Embed Size (px)
Citation preview

Journal of Controlled Release 129 (2008) 26–32
Contents lists available at ScienceDirect
Journal of Controlled Release
j ourna l homepage: www.e lsev ie r.com/ locate / jconre l
Oligomannose-coated liposomes as a therapeutic antigen-delivery and an adjuvantvehicle for induction of in vivo tumor immunity
Naoya Kojima a,⁎, Le Biao a, Tomoko Nakayama a, Mariko Ishii a, Yuzuru Ikehara b, Kunio Tsujimura c
a Department of Applied Biochemistry, Tokai University, Hiratsuka, Kanagawa 259-1292, Japanb Molecular Medicine Team of Research Center for Medical Glycoscience, National Institute of Advanced Industrial Science and Technology, Tsukuba 305-8568, Japanc Department of Infectious Diseases, Hamamatsu University School of Medicine, Hamamatsu 431-3192, Japan
⁎ Corresponding author. Tel.: +81 463 58 1211; fax: +E-mail address: [email protected] (N. K
0168-3659/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.jconrel.2008.03.023
A B S T R A C T
A R T I C L E I N F OArticle history:
In the present study, the adj Received 5 November 2007Accepted 29 March 2008Available online 7 April 2008Keywords:Antigen deliveryCancer immunotherapyCytotoxic T cellsOligomannose-coated liposomeVaccine
uvant capacity of oligomannose-coated liposomes (OMLs) was evaluated in mice,and an OML-based vaccine was shown to induce effective anti-tumor immunity. C57BL/6 mice wereimmunized subcutaneously with OML-encased ovalbumin (OVA) and challenged with OVA-expressing E.G7-OVA tumor cells. All mice that received OVA in OMLs completely rejected the E.G7-OVA tumor. Spleen cellsfrom the immunized mice showed strong cytotoxic activity against E.G7-OVA cells, but not against theparental EL4 cells. The therapeutic efficacy of OML-encased OVA against established E.G7-OVA tumors wasthen investigated. When the tumor mass became palpable (8–10 mm in length), the mice were treated with asingle injection of 1 μg of OML-encased OVA. Tumor growth was reduced significantly in mice treated withOML-encased OVA and tumors were completely eliminated in about 40% of these mice. Similar results wereobtained using EL4 tumors, with the EL4 cell lysate used as an antigen. These results indicate that an OML-based vaccine with an encased tumor antigen might be useful clinically to raise an effective immuneresponse against a tumor.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Cytotoxic T lymphocytes (CTLs) have a critical role in tumorimmunity. Activation of antigen-presenting cells (APCs) such asdendritic cells and macrophages is required to induce CTLs, and thiscan be achieved using various adjuvants, including microbialcomponents such as lipoarabinomannan, lipopolysaccharide, lipotei-choic acid, and bacterial DNA [1–5]. These bacterial components arereferred to as pathogen-associated molecular patterns (PAMPs) andare recognized by pattern recognition receptors expressed on APCs[5,6]. To induce tumor-specific CTLs, it is also important that APCspresent antigenic peptides derived from tumor-associated antigens(TAAs) on major histocompatibility complex (MHC) class I molecules.Thus, strategies aimed at delivery of TAAs in the MHC class I and classII pathways of APCs require more than just activation of APCs [7].Liposomes (vesicles formed from phospholipid bilayers) can encap-sulate antigen and immunomodulatory factors, thus serving as potentdelivery vehicles [8,9], and delivery of antigen-containing liposomesto immune cells can be facilitated by agents that bind selectively tomolecules on the surface of target cells. Hence, antibodies andrecombinant proteins, such as soluble forms of cell surface receptors
81 463 50 2012.ojima).
l rights reserved.
or their ligands, can be used to target liposomes to receptors on cells[10–13]. The lectin-like receptors expressed on APCs, including themannose receptors thought to be involved in phagocytosis ofpathogens, can facilitate binding and endocytosis of ligands thathave a terminal sugar such as mannose, and hence could be utilized todirect delivery of liposome-encapsulated antigens to APCs [11,14,15].
We have recently demonstrated that liposomes coated withneoglycolipids constructed from mannotriose and dipalmitoylpho-sphatidylethanolamine (Man3-DPPE) (oligomannose-coated lipo-somes, OMLs) are preferentially and rapidly taken up by peritonealmacrophages (PEMs) and that the liposome-containing PEMs accu-mulate in extra-nodal lymphoid tissues following administration ofOMLs into the peritoneal cavity [16,17]. More recently, we have shownthat OML treatment of PEMs leads to upregulation of co-stimulatorymolecules and MHC class II molecules, that the OML-stimulatedmacrophages preferentially produce IL-12, which is essential forinduction of a Th1 immune response, and that PEMs containingOMLs effectively present encased antigen-derived peptides via MHCclass I and II molecules and activate both CD4-positive and CD8-positive T cells [18,19]. Furthermore, intraperitoneal (i.p.) immuniza-tion of an OML-entrapped soluble leishmanial antigen induces anantigen-specific Th1 immune response and protects against subse-quent infection by Leishmania major in BALB/c mice [20,21]. Inaddition, i.p. administration of OMLs caused complete rejection of E.G7-OVA tumors challenged after immunization [19]. These results

27N. Kojima et al. / Journal of Controlled Release 129 (2008) 26–32
indicate that OMLs can be used both as an antigen-delivery vehicle toAPCs and as an APC-activating adjuvant to induce strong cellular andanti-tumor immunity following i.p. administration.
However, the i.p. route is not viable for vaccination in humans, andapplication of OMLs in cancer immunotherapy requires a differentvaccination route. The aim of this study is thus to clarify whethersubcutaneous (s.c.) administration of OMLs gives sufficient anti-tumorimmunity to protect against tumor inoculation, as seen with i.p.administration. In addition, we evaluated the therapeutic efficacy of s.c.administration of OML-entrapped tumor-associated antigen againstestablished tumors. Our results indicate that OMLs containing a tumorantigen can be used as a highly efficient vaccine to enhance induction ofin vivo tumor immunity.
2. Materials and methods
2.1. Chemicals
Cholesterol, ovalbumin (OVA), dipalmitoylphosphatidylethanola-mine (DPPE), and dipalmitoylphosphatidylcholine (DPPC) werepurchased from Sigma (St Louis, MO). Mannotriose (Man3) withthe structure Manα1-6(Manα1-3)Man was purchased from Funa-koshi (Tokyo, Japan). Man3-DPPE was prepared in our laboratory byconjugation of these oligosaccharides with DPPE, as describedpreviously [16]. The structure and purity of Man3-DPPE wereconfirmed by matrix-assisted laser desorption/ionization-time-of-flight mass spectrometry (Auto FLEX, Bruker Daltonics, Bremen,Germany) and high-performance thin-layer chromatography,respectively. The purity of Man3-DPPE used in the study was atleast 98%.
2.2. Cell lines and animals
A C57BL/6-derived thymoma cell line of the H-2b haplotype, EL4,was obtained from the American Type Culture Collection (Mana-ssas, VA) and grown in RPMI 1640 medium (GIBCO) supplementedwith 10% FCS, 100 U/ml penicillin, 100 µg/ml streptomycin, and2 mM L-glutamine at 37°C in 5% CO2 (complete RPMI medium). Toprepare EL4 tumor-cell lysate (EL4-TCL), EL4 tumor cells wereharvested, washed twice with phosphate-buffered saline (PBS), andsuspended in PBS at a concentration of 108 108 cells/ml. The cellswere then subjected to seven freeze–thaw cycles. The suspensionwas centrifuged (8000 ×g) and the supernatant was sonicated at4 °C with twenty one-second pulse cycles using an ultrasonicdisruptor (UD-201; TOMY, Tokyo, Japan). The sonicated supernatantwas centrifuged at 100,000 ×g and then passed through a 0.2-μmfilter to give EL4-TCL. The E.G7-OVA cell line, an EL4 cell line that isstably transfected with cDNA of OVA to allow endogenousproduction of OVA with an H-2Kb-restricted CTL epitope [22], wasobtained from the American Type Culture Collection and main-tained in complete RPMI medium supplemented with 400 µg/mlG418 (Wako, Osaka, Japan) at 37 °C in 5% CO2. Six- to 8-week-oldfemale C57BL/6 mice were purchased from Japan SLC (Hamamatsu,Japan) and used 1 week after arrival at our animal facility. Allanimal experiments were conducted in accordance with a protocolapproved by Tokai University.
2.3. Preparation of liposomes
Liposomes were prepared as described previously [16,18]. Briefly, achloroform:methanol (2:1, v/v) solution containing 1.5 μmol of DPPC,1.5 μmol of cholesterol, and different amounts (0.15–0.0015 μmol) ofMan3-DPPE was added to the flask and evaporated to prepare a lipidfilm. PBS containing 5 mg/ml of OVA or 2.5 mg/ml of EL4-TCL wasadded to the dried lipid film andmultilamellar vesicles were preparedby intense vortex dispersion. The vesicles were extruded 10 times
through a 1-μm pore polycarbonate membrane (Nucleopore, Plea-santon, CA). The amount of entrapped protein was measured using amodified Lowry protein assay kit (Pierce, Rockford, IL) in the presenceof 0.3% (w/v) sodium dodecyl sulfate, using BSA as the standard. Molarratios of the lipid components of the liposomes were determinedusing HPLC. Particle sizes of liposomes were determined using adynamic light scattering particle size analyzer (LB-550, Horiba, Kyoto,Japan).
2.4. Immunization of mice and tumor inoculation
C57BL6micewere immunized s.c. in the right dorsal areawith 1 μgOVA or EL4-TCL encased in Man3-DPPE-coated liposomes (OML/OVAor OML/EL4), 1 μg OVA or EL4-TCL encased in uncoated liposomes(Bare/OVA or Bare/EL4), 1 μg OVA or EL4-TCL in 100 μl PBS, or PBS(100 μl each). This vaccination procedure was repeated 3 times at one-week intervals. One week after the last immunization, 5 mice in eachgroup were challenged with dorsal s.c. injection of 1×106 E.G7-OVA orEL4 cells. Tumor growth was assessed using dial gauge calipers andexpressed as tumor volume using the following formula: tumorvolume (mm3)=[length/2 (mm)]×[width/2 (mm)]2×π.
2.5. In vitro stimulation of spleen cells and CTL assay
The remaining 5 mice from each group were sacrificed one weekafter the third immunization and single cell suspensions from thespleens of individual mice were prepared in Hanks' balanced saltsolution containing 2% FCS. The spleen cells were washed with RPMI1640 medium containing 10% FCS, suspended at a density of 5×106
106 cells/ml, dispensed in triplicate onto 24-well plates, and culturedin 1 ml of RPMI 1640 medium containing 10% FCS in the presence orabsence of 10 µg/ml OVA. After incubation for 72 h at 37 °C in 5% CO2,CD8-positive cells were isolated by negative selection using a MACSsystem (Miltenyi Biotec, Bergisch Gladbach, Germany). The super-natants of the spleen cell cultures were collected and kept at −80°Cuntil assayed for cytokines. The effector cells (spleen cells or CD8-positive cells) thus prepared were co-cultured with target cells (E.G7-OVA or EL4) at various effector/target ratios for 8 h at 37 °C andcytotoxicity was measured with a CytoTox96 Non-RadioactiveCytotoxicity assay kit (Promega, Madison, WI). Levels of IFN-γ andIL-4 were quantified by enzyme-linked immunosorbent assay (ELISA)using a commercial ELISA kit (Pierce, Rockford, IL).
2.6. Therapeutic immunization using OML-encased antigen
The therapeutic efficacy of an OML-based vaccine against estab-lished E.G7-OVA or EL4 tumors was investigated. E.G7-OVA tumorcells (1×106) were injected s.c. into the left dorsal area of C57BL/6mice on day 0. When the tumor mass became palpable (8–10 mm inlength, around day 10), the mice were immunized s.c. in the rightdorsal area with 1 µg OVA encased in OMLs (OML/OVA), 1 µg OVAencased in uncoated liposomes (Bare/OVA), 1 µg OVA in 100 µl PBS, orPBS (100 µl each). The anti-tumor activity mediated by the transferredcells was determined by measuring perpendicular tumor diameters.The same experiments were performed using EL4 tumors with EL4-TCL as antigen. In this case, the vaccination procedure was repeated 3times at one-week intervals.
2.7. Immunohistochemical analysis
For immunohistochemical detection of FITC-conjugated BSAcarried by macrophages to the draining lymph nodes, sections of thepopliteal lymph nodes were incubated with peroxidase-labeledpolyclonal anti-FITC antibody (Roche Diagnostic Co., Tokyo, Japan)overnight at 4 °C. The antibody reaction was visualized with adiaminobenzidine (DAB)/H2O2 solution.

Fig. 1. Accumulation of OMLs in draining lymph nodes. Mice were treated in the lefthind footpad with OMLs (A) or uncoated liposomes (bare liposomes) (B) includingentrapped FITC-BSA. The popliteal lymph nodes were isolated 24 h after administrationand FITC signals were detected using an anti-FITC antibody.
28 N. Kojima et al. / Journal of Controlled Release 129 (2008) 26–32
2.8. Statistical analysis
Difference between means for experimental groups was analyzedusing Student t-tests, and was considered significant at Pb0.05.
3. Results
3.1. OMLs accumulate in the draining lymph nodes followingsubcutaneous administration
Multilamellar liposomes prepared from DPPC, cholesterol, andMan3-DPPE at the molar ratio of 10:10:1 (oligomannose-coatedliposomes, OMLs) and extruded through a membrane with 1-μmpores were used in all experiments unless otherwise indicated. Thecompositions of the OMLs and uncoated liposomes without Man3-DPPE (Bare) used in this study were summarized in Table 1.
FITC-labeled BSA-containing OMLs (OML/FITC) or FITC-labeledBSA-containing uncoated liposomes (Bare/FITC) (30 µg of choles-terol each) were administered s.c. in the left hind footpad, and thepopliteal lymph nodes were isolated 24 h after administration. Thesignals from liposomes were confirmed immunohistochemicallyusing a specific antibody against FITC. As shown in Fig. 1, a strongsignal for FITC was seen in the draining lymph nodes following OML/FITC administration, while no apparent accumulation was seen inmice that received Bare/FITC. This result indicates that the OMLs canbe used for delivery of antigens to the draining lymphoid tissues bys.c. administration.
3.2. Subcutaneous vaccination using OMLs induces a strong anti-tumoreffect against E.G7-OVA cells
Subcutaneous vaccination using OMLs was first evaluated forprotection of mice against E.G7-OVA cell challenge. E.G7-OVA tumorcells present OVA peptide (OVA257–264) on MHC class I molecules as amodel tumor-associated antigen. Each group of 5 mice underwent s.c.injection of 1 µg OVA encased in OMLs (OML/OVA), 1 µg OVA inuncoated liposomes (Bare/OVA),1 µg OVA alone, or PBS 3 times at one-week intervals. One week after the last immunization, mice werechallenged with E.G7-OVA cells (1×106) and tumor growth wasdetermined in each mouse. All mice that received OML/OVA (3injections) completely rejected the E.G7-OVA tumor and survived forover 80 days after tumor challenge (Fig. 2A). A single s.c. injection ofOML/OVA also induced sufficient anti-tumor immunity to reject the E.G7-OVA tumor (data not shown). In contrast, tumor rejection was notobserved inmice immunizedwith OVA alone or Bare/OVA and all micedied about 50 days after the tumor challenge.
The levels of IFN-γ and IL-4 were quantified for assessment of Th1and Th2 immune responses, since IFN-γ and IL-4 are produced by Th1and Th2 cells, respectively. Upon stimulationwith OVA in vitro, spleencells from mice immunized with OML/OVA produced significantamounts of IFN-γ, whereas cytokines were not produced by spleencells from mice that received Bare/OVA, OVA or PBS (Fig. 2B). On theother hand, only small amounts of IL-4 were produced from cellstreated with OML/OVA, indicating that s.c. vaccination with OML/OVA
Table 1Composition of liposomes used in this study
Liposome DPPC Cholesterol Man3-DPPE Protein Particle size
mol/mol µg/mg cholesterol nm
OML/OVA 1.00 1.02±0.08 0.089±0.006 24.7±3.3 1053±98Bare/OVA 1.00 0.98±0.11 0 27.8±4.1 1078±104OML/EL4 1.00 1.08±0.09 0.095±0.012 19.3±3.8 1127±121Bare/EL4 1.00 1.04±0.13 0 18.4±4.3 1108±109
Values are represented as mean±SD from five independent preparations.
induces an antigen-specific Th1-type immune response. Next, CTLlevels in spleen cells were determined to examine the presence ofOVA-specific T cells. Only spleen cells from mice immunized withOML/OVA showed cytotoxicity against E.G7-OVA (Fig. 2C). CD8+ cellsisolated from spleen cells of mice immunized with OML/OVA showedcytotoxicity against E.G7-OVA tumor cells, but not against parentalEL4 tumor cells, which do not present OVA peptide as a tumor antigen(Fig. 3A). Furthermore, mice immunized with OML/OVA were unableto reject an EL4 tumor (Fig. 3B). Tumor-associated antigens have notbeen identified for the EL4 tumor, and therefore EL4 tumor-cell lysate(EL4-TCL), which includes unidentified tumor-associated antigens,was used as the antigen source. With co-administration of EL4-TCL, allmice that received OML/EL4 completely rejected the EL4 tumor,whereas EL4 tumor growth was seen in mice that received Bare/EL4 orEL4-TCL alone (Fig. 3C). These results indicate that OML-mediatedimmunization induces a systemic immune response that protectsmice from tumor challenge in an antigen-specific manner.
3.3. Preferential OML composition for tumor rejection
To assess the required amount of antigen for induction of anti-tumor immunity, different amounts of OVA encased in OMLs wereadministered to mice. The minimal amount of OML-encased OVArequired to prime mice for complete rejection of an E.G7-OVA tumorwas 0.1 μg/mouse (Fig. 4A). A statistically significant suppression oftumor growth was seen in mice immunized with 0.05 μg or 0.01 μg ofOVA encased in OMLs (Pb0.005), but complete tumor rejection wasnot observed at this dose (Data not shown). Immunization with OMLswithout OVA (empty OML) or with a mixture of soluble OVA andempty OMLs did not suppress tumor growth (PN0.1). Spleen cellsfrom mice immunized with 1 μg of OVA encased in OMLs showedcytotoxicity against E.G7-OVA tumor cells that was similar to thatdetected in cells from mice immunized with 10 μg of OVA encased inOMLs, suggesting that 1 μg of OVA per mouse is sufficient to inducemaximal cytotoxic activity. The cytotoxicity of the spleen cellsdecreased in a dose-dependent manner (Fig. 4B).
To assess the most effective density of oligomannose on theliposome surface, OMLs were prepared from equimolar amounts ofDPPC and cholesterol and different amounts of Man3-DPPE. Miceimmunized with 1 μg of OVA encased in these OMLs were challengedwith E.G7-OVA tumor cells. Tumor growth in mice that received OMLsconsisting of DPPC:cholesterol:Man3-DPPE at ratios of 10:10:0.01
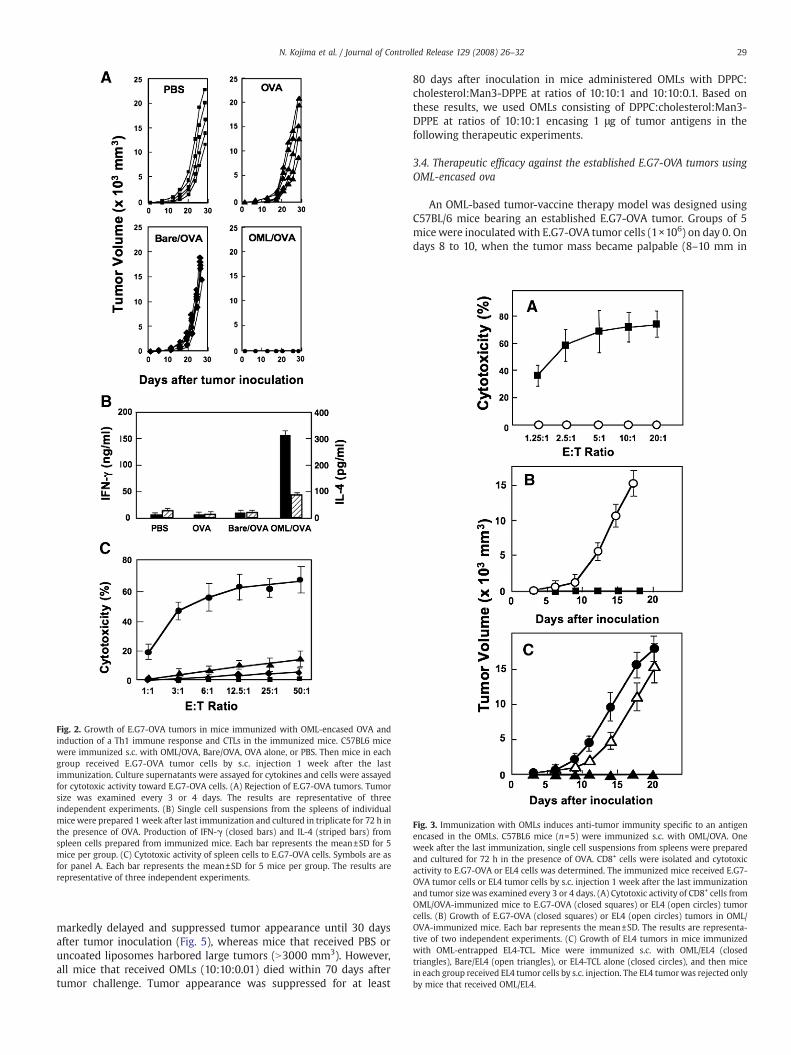
Fig. 2. Growth of E.G7-OVA tumors in mice immunized with OML-encased OVA andinduction of a Th1 immune response and CTLs in the immunized mice. C57BL6 micewere immunized s.c. with OML/OVA, Bare/OVA, OVA alone, or PBS. Then mice in eachgroup received E.G7-OVA tumor cells by s.c. injection 1 week after the lastimmunization. Culture supernatants were assayed for cytokines and cells were assayedfor cytotoxic activity toward E.G7-OVA cells. (A) Rejection of E.G7-OVA tumors. Tumorsize was examined every 3 or 4 days. The results are representative of threeindependent experiments. (B) Single cell suspensions from the spleens of individualmice were prepared 1 week after last immunization and cultured in triplicate for 72 h inthe presence of OVA. Production of IFN-γ (closed bars) and IL-4 (striped bars) fromspleen cells prepared from immunized mice. Each bar represents the mean±SD for 5mice per group. (C) Cytotoxic activity of spleen cells to E.G7-OVA cells. Symbols are asfor panel A. Each bar represents the mean±SD for 5 mice per group. The results arerepresentative of three independent experiments.
Fig. 3. Immunization with OMLs induces anti-tumor immunity specific to an antigenencased in the OMLs. C57BL6 mice (n=5) were immunized s.c. with OML/OVA. Oneweek after the last immunization, single cell suspensions from spleens were preparedand cultured for 72 h in the presence of OVA. CD8+ cells were isolated and cytotoxicactivity to E.G7-OVA or EL4 cells was determined. The immunized mice received E.G7-OVA tumor cells or EL4 tumor cells by s.c. injection 1 week after the last immunizationand tumor size was examined every 3 or 4 days. (A) Cytotoxic activity of CD8+ cells fromOML/OVA-immunized mice to E.G7-OVA (closed squares) or EL4 (open circles) tumorcells. (B) Growth of E.G7-OVA (closed squares) or EL4 (open circles) tumors in OML/OVA-immunized mice. Each bar represents the mean±SD. The results are representa-tive of two independent experiments. (C) Growth of EL4 tumors in mice immunizedwith OML-entrapped EL4-TCL. Mice were immunized s.c. with OML/EL4 (closedtriangles), Bare/EL4 (open triangles), or EL4-TCL alone (closed circles), and then micein each group received EL4 tumor cells by s.c. injection. The EL4 tumor was rejected onlyby mice that received OML/EL4.
29N. Kojima et al. / Journal of Controlled Release 129 (2008) 26–32
markedly delayed and suppressed tumor appearance until 30 daysafter tumor inoculation (Fig. 5), whereas mice that received PBS oruncoated liposomes harbored large tumors (N3000 mm3). However,all mice that received OMLs (10:10:0.01) died within 70 days aftertumor challenge. Tumor appearance was suppressed for at least
80 days after inoculation in mice administered OMLs with DPPC:cholesterol:Man3-DPPE at ratios of 10:10:1 and 10:10:0.1. Based onthese results, we used OMLs consisting of DPPC:cholesterol:Man3-DPPE at ratios of 10:10:1 encasing 1 μg of tumor antigens in thefollowing therapeutic experiments.
3.4. Therapeutic efficacy against the established E.G7-OVA tumors usingOML-encased ova
An OML-based tumor-vaccine therapy model was designed usingC57BL/6 mice bearing an established E.G7-OVA tumor. Groups of 5micewere inoculated with E.G7-OVA tumor cells (1×106) on day 0. Ondays 8 to 10, when the tumor mass became palpable (8–10 mm in

Fig. 6. Therapeutic efficacy of OML/OVA vaccination against an established E.G7-OVAtumor. Mice (n=5) were challenged by s.c. injection of E.G7-OVA tumor cells in the rightdorsal area on day 0. When the tumor mass became palpable (around day 10), OML/OVA, Bare/OVA, or PBS was injected into the side where the tumor had grown.(A) Suppression of tumor growth, with tumor size examined every 3 days. †, mouse diedat the indicated time point. (B) Surviving mice, expressed as a percentage of the totalnumber of mice in each group. Results are representative of five independentexperiments.
Fig. 4. Effect of the dose of OML/OVA on tumor growth and CTL induction. C57BL6 mice(n=10) were immunized s.c. with 10 µg (closed squares), 1 µg (closed circles), 0.1 µg(open circles), 0.05 µg (open triangles), or 0.01 µg (open diamonds) of OML-encasedOVA at one-week intervals. As a negative control, mice received OMLs without OVA(empty OML, open squares) or PBS (closed triangles). Alternatively, mice received amixture of empty OML and OVA (closed diamonds). E.G7-OVA tumor cells wereadministered by s.c. injection in the right dorsal area 1 week after the lastimmunization. Tumor growth in the mice in each group (A) and cytotoxic activity ofspleen cells (B) were determined as described in the legend to Fig. 2. Note that tumorgrowth was not detected in the mice that received over 0.1 µg of antigen in OMLs, andthus, the groups of mice that received 1 µg and 10 µg of antigens were not displayed inPanel A. In addition, cytotoxic activity was not observed in mice that received a mixtureof empty OML and OVA, empty OML, and PBS. Each bar represents the mean±SD.⁎Statistically significant difference (Pb0.005) compared to PBS.
30 N. Kojima et al. / Journal of Controlled Release 129 (2008) 26–32
length), tumor-bearing mice were immunized with a single s.c.injection of 1 µg OVA encased in OMLs (OML/OVA), 1 µg OVA inuncoated liposomes (Bare/OVA), or PBS. Significant suppression oftumor growth and reduction of tumor mass were observed in all mice
Fig. 5. Effect on tumor growth of the concentration of Man3-DPPE in liposomes. Mice(n=5) were immunized s.c. with 1 µg of OVA encased in OMLs consisting of DPPC:cholesterol:Man3-DPPE at ratios of 10:10:1 (closed circles), 10:10:0.1 (open squares) or10:10:0.01 (open triangles) at one-week intervals. In addition, mice were treated with1 µg of OVA encased in uncoated liposomes (closed squares) or with PBS (open circles).E.G7-OVA tumor cells were administered by s.c. injection 1 week after the lastimmunization and tumor growth was determined until the tumor volume reached3000 mm3 (†).
treated with OML/OVA (Fig. 6A), whereas mice treated with Bare/OVAshowed no marked suppressive effect on E.G7-OVA tumor growth. Allmice treated with OML/OVA survived until day 53, by which time allmice that received Bare/OVA and PBS had died. In addition, two of thefive OML/OVA-treatedmice showed complete elimination of the E.G7-OVA tumor (Fig. 6B). These results are representative of fourindependent sets of experiments, and 11 of 25 tumor-bearing micethat received OML/OVA showed elimination of the E.G7-OVA tumor(data not shown). Thus, a single injection of an OML-based tumorvaccine induced effective anti-tumor activity in model E.G7-OVAtumor-bearing mice.
3.5. Therapeutic efficacy against established EL4 tumors using OML-encased EL4 tumor-cell lysate
We next examined therapeutic efficacy against an established EL4tumor. Groups of 5 micewere inoculated with EL4 tumor cells (1×106)on day 0. On days 5 to 7, when the tumor mass became palpable (8–10 mm in length), tumor-bearing mice were immunized with a singles.c. injection of 1 µg EL4-TCL encased in OMLs (OML/EL4), 1 µg EL4-TCLin uncoated liposomes (Bare/EL4), 1 µg EL4-TCL or PBS (control). Thevaccination procedures were repeated three times at one-weekintervals. Tumor growth was significantly reduced in all mice thatreceived OML/EL4 compared with mice that treated with Bare/EL4 orEL4-TCL (Fig. 7A). Mice that received Bare/EL4 or EL4-TCL died on days20 to 25, whereas all OML/EL4-treated mice survived to that point(Fig. 7B). The mean survival times for control mice, and mice receivingEL4-TCL, Bare/EL4, and OML/EL4 were 18.6±1.5, 22.8±3.0, 22.0±2.8,and 46.8±10.9 days, respectively; Therefore, mice vaccinated with
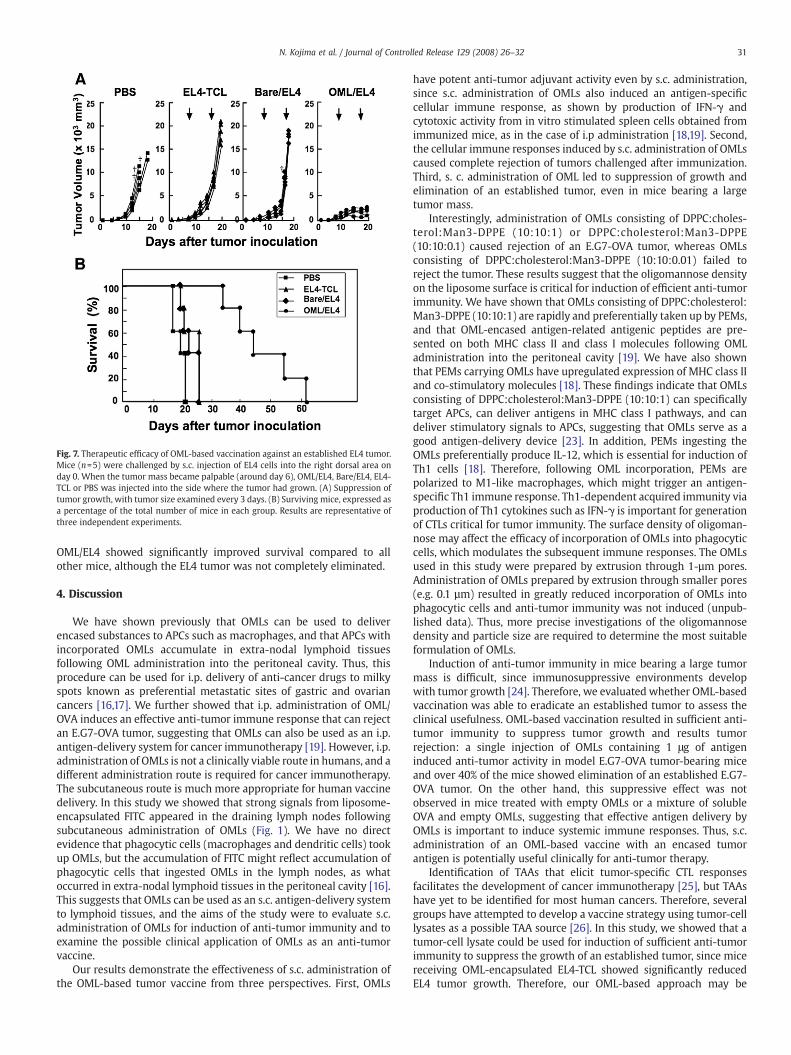
Fig. 7. Therapeutic efficacy of OML-based vaccination against an established EL4 tumor.Mice (n=5) were challenged by s.c. injection of EL4 cells into the right dorsal area onday 0. When the tumor mass became palpable (around day 6), OML/EL4, Bare/EL4, EL4-TCL or PBS was injected into the side where the tumor had grown. (A) Suppression oftumor growth, with tumor size examined every 3 days. (B) Surviving mice, expressed asa percentage of the total number of mice in each group. Results are representative ofthree independent experiments.
31N. Kojima et al. / Journal of Controlled Release 129 (2008) 26–32
OML/EL4 showed significantly improved survival compared to allother mice, although the EL4 tumor was not completely eliminated.
4. Discussion
We have shown previously that OMLs can be used to deliverencased substances to APCs such as macrophages, and that APCs withincorporated OMLs accumulate in extra-nodal lymphoid tissuesfollowing OML administration into the peritoneal cavity. Thus, thisprocedure can be used for i.p. delivery of anti-cancer drugs to milkyspots known as preferential metastatic sites of gastric and ovariancancers [16,17]. We further showed that i.p. administration of OML/OVA induces an effective anti-tumor immune response that can rejectan E.G7-OVA tumor, suggesting that OMLs can also be used as an i.p.antigen-delivery system for cancer immunotherapy [19]. However, i.p.administration of OMLs is not a clinically viable route in humans, and adifferent administration route is required for cancer immunotherapy.The subcutaneous route is much more appropriate for human vaccinedelivery. In this study we showed that strong signals from liposome-encapsulated FITC appeared in the draining lymph nodes followingsubcutaneous administration of OMLs (Fig. 1). We have no directevidence that phagocytic cells (macrophages and dendritic cells) tookup OMLs, but the accumulation of FITC might reflect accumulation ofphagocytic cells that ingested OMLs in the lymph nodes, as whatoccurred in extra-nodal lymphoid tissues in the peritoneal cavity [16].This suggests that OMLs can be used as an s.c. antigen-delivery systemto lymphoid tissues, and the aims of the study were to evaluate s.c.administration of OMLs for induction of anti-tumor immunity and toexamine the possible clinical application of OMLs as an anti-tumorvaccine.
Our results demonstrate the effectiveness of s.c. administration ofthe OML-based tumor vaccine from three perspectives. First, OMLs
have potent anti-tumor adjuvant activity even by s.c. administration,since s.c. administration of OMLs also induced an antigen-specificcellular immune response, as shown by production of IFN-γ andcytotoxic activity from in vitro stimulated spleen cells obtained fromimmunized mice, as in the case of i.p administration [18,19]. Second,the cellular immune responses induced by s.c. administration of OMLscaused complete rejection of tumors challenged after immunization.Third, s. c. administration of OML led to suppression of growth andelimination of an established tumor, even in mice bearing a largetumor mass.
Interestingly, administration of OMLs consisting of DPPC:choles-terol:Man3-DPPE (10:10:1) or DPPC:cholesterol:Man3-DPPE(10:10:0.1) caused rejection of an E.G7-OVA tumor, whereas OMLsconsisting of DPPC:cholesterol:Man3-DPPE (10:10:0.01) failed toreject the tumor. These results suggest that the oligomannose densityon the liposome surface is critical for induction of efficient anti-tumorimmunity. We have shown that OMLs consisting of DPPC:cholesterol:Man3-DPPE (10:10:1) are rapidly and preferentially taken up by PEMs,and that OML-encased antigen-related antigenic peptides are pre-sented on both MHC class II and class I molecules following OMLadministration into the peritoneal cavity [19]. We have also shownthat PEMs carrying OMLs have upregulated expression of MHC class IIand co-stimulatory molecules [18]. These findings indicate that OMLsconsisting of DPPC:cholesterol:Man3-DPPE (10:10:1) can specificallytarget APCs, can deliver antigens in MHC class I pathways, and candeliver stimulatory signals to APCs, suggesting that OMLs serve as agood antigen-delivery device [23]. In addition, PEMs ingesting theOMLs preferentially produce IL-12, which is essential for induction ofTh1 cells [18]. Therefore, following OML incorporation, PEMs arepolarized to M1-like macrophages, which might trigger an antigen-specific Th1 immune response. Th1-dependent acquired immunity viaproduction of Th1 cytokines such as IFN-γ is important for generationof CTLs critical for tumor immunity. The surface density of oligoman-nose may affect the efficacy of incorporation of OMLs into phagocyticcells, which modulates the subsequent immune responses. The OMLsused in this study were prepared by extrusion through 1-µm pores.Administration of OMLs prepared by extrusion through smaller pores(e.g. 0.1 µm) resulted in greatly reduced incorporation of OMLs intophagocytic cells and anti-tumor immunity was not induced (unpub-lished data). Thus, more precise investigations of the oligomannosedensity and particle size are required to determine the most suitableformulation of OMLs.
Induction of anti-tumor immunity in mice bearing a large tumormass is difficult, since immunosuppressive environments developwith tumor growth [24]. Therefore, we evaluated whether OML-basedvaccination was able to eradicate an established tumor to assess theclinical usefulness. OML-based vaccination resulted in sufficient anti-tumor immunity to suppress tumor growth and results tumorrejection: a single injection of OMLs containing 1 μg of antigeninduced anti-tumor activity in model E.G7-OVA tumor-bearing miceand over 40% of the mice showed elimination of an established E.G7-OVA tumor. On the other hand, this suppressive effect was notobserved in mice treated with empty OMLs or a mixture of solubleOVA and empty OMLs, suggesting that effective antigen delivery byOMLs is important to induce systemic immune responses. Thus, s.c.administration of an OML-based vaccine with an encased tumorantigen is potentially useful clinically for anti-tumor therapy.
Identification of TAAs that elicit tumor-specific CTL responsesfacilitates the development of cancer immunotherapy [25], but TAAshave yet to be identified for most human cancers. Therefore, severalgroups have attempted to develop a vaccine strategy using tumor-celllysates as a possible TAA source [26]. In this study, we showed that atumor-cell lysate could be used for induction of sufficient anti-tumorimmunity to suppress the growth of an established tumor, since micereceiving OML-encapsulated EL4-TCL showed significantly reducedEL4 tumor growth. Therefore, our OML-based approach may be

32 N. Kojima et al. / Journal of Controlled Release 129 (2008) 26–32
promising in TAA-based cancer immunotherapy for control of tumorsthat express tumor-associated antigens.
Microbial components such as lipoarabinomannan, LPS and theouter membrane protein of gram-negative bacteria, bacterial holotox-ins such as cholera toxin, and bacterial DNA have been used asadjuvants in therapeutic immunization against infectious and non-infectious diseases [1–5]. In particular, unmethylated bacterial CpG-ODNs, which is recognized by TLR-9 expressed on APCs, promotes aTh1 immune response via APC maturation [27,28]. Although admin-istration of CpG-ODNs induces strong anti-tumor immunity [29],repeated or intravenous injection of CpG-ODNs has been reported tohave unfavorable immunosuppressive effects in mice [30]. We havedemonstrated that OMLs activate peritoneal macrophages, leading toupregulation of co-stimulatory molecules and MHC class II molecules,and secretion of IL-12 [18]. Furthermore, a comparative study ofcytokine production from OML- and CpG-activated peritoneal macro-phages suggested that the signaling pathway for activation of the cellby OMLs is distinct from the CpG-TLR9 signaling pathway [18].Therefore, OML-based vaccination may overcome the immunosup-pressive effects of CpG-ODNs. Successful development of newvaccines requires an effective tumor–antigen delivery system and anadjuvant that induces an effective anti-tumor immune response.Based on our previous data [16–21] and the current results, weconclude that OMLs have excellent properties as novel antigen-delivery vehicles and have a strong adjuvant effect even via s.c.administration, suggesting that they have potential for clinical use inanti-tumor vaccination.
Acknowledgements
This work was supported by the Program for Promotion of BasicResearch Activities for Innovative Biosciences (PROBRAIN), in part by agrant for Hi-Tech Research from Tokai University, and by the IndustrialTechnology Research Grant Program of the New Energy and IndustrialTechnology Development Organization (NEDO) of Japan.
References
[1] R. Dziarski, Q. Wang, K. Miyake, C.J. Kirschning, D. Gupta, MD-2 enables Toll-likereceptor 2 (TLR2)-mediated responses to lipopolysaccharide and enhances TLR2-mediated responses to Gram-positive and Gram-negative bacteria and their cellwall components, J. Immunol. 166 (3) (2001) 1938–1944.
[2] K. Matsui, R. Motohashi, A. Nishikawa, Cell wall components of Staphylococcusaureus induce interleukin-5 production in patients with atopic dermatitis, J.Interferon Cytokine Res. 20 (3) (2000) 321–324.
[3] D.N. Dao, L. Kremer, Y. Guerardel, A. Molano, W.R. Jacobs Jr, S.A. Porcelli, V. Briken,Mycobacterium tuberculosis lipomannan induces apoptosis and interleukin-12production in macrophages, Infect. Immun. 72 (4) (2004) 2067–2074.
[4] F. Zhou, L. Huang, Monophosphoryl lipid A enhances specific CTL induction by asoluble protein antigen entrapped in liposomes, Vaccine 11 (11) (1993) 1139–1144.
[5] O. Takeuchi, S. Sato, T. Horiuchi, K. Hoshino, K. Takeda, Z. Dong, R.L. Modlin, S. Akira,Cutting edge: role of Toll-like receptor 1 in mediating immune response tomicrobial lipoproteins, J. Immunol. 169 (1) (2002) 10–14.
[6] T. Kawai, S. Akira, Pathogen recognition with Toll-like receptors, Curr. Opin.Immunol. 17 (4) (2005) 338–344.
[7] P.J. Tacken, R. Torensma, C.G. Figdor, Targeting antigens to dendritic cells in vivo,Immunobiology 211 (6–8) (2006) 599–608.
[8] J.G. Altin, C.R. Parish, Liposomal vaccines—targeting the delivery of antigen,Methods 40 (1) (2006) 39–52.
[9] J.G. Altin, C.L. van Broekhoven, C.R. Parish, Targeting dendritic cells with antigen-containing liposomes: antitumour immunity, Expert Opin. Biol. Ther. 4 (11) (2004)1735–1747.
[10] J.W. Park, B.D. Kirpotin, K. Hong, R. Shalaby, Y. Shao, U.B. Nielsen, J.D. Marks, D.Papahadjopolous, C.C. Benz, Tumor targeting using anti-her2 immunoliposomes, J.Control. Release 74 (1–3) (2001) 95–113.
[11] R.K. Gieseler, G. Marquitan, M.J. Hahn, L.A. Perdon, W.H. Driessen, S.M. Sullivan, M.J.Scolaro, DC-SIGN-specific liposomal targeting and selective intracellular compounddelivery to human myeloid dendritic cells: implications for HIV disease, Scand. J.Immunol. 59 (5) (2004) 415–424.
[12] C.L. van Broekhoven, C.R. Parish, C. Demangel, W.J. Britton, J.G. Altin, Targetingdendritic cells with antigen-containing liposomes: a highly effective procedure forinduction of antitumor immunity and for tumor immunotherapy, Cancer Res. 64(12) (2004) 4357–4365.
[13] K. Kawamura, N. Kadowaki, R. Suzuki, S. Udagawa, S. Kasaoka, N. Utoguchi, T.Kitawaki, N. Sugimoto, N. Okada, K. Maruyama, T. Uchiyama, Dendritic cells thatendocytosed antigen-containing IgG-liposomes elicit effective antitumor immu-nity, J. Immunother. (1997) 29 (2) (2006) 165–174.
[14] K.L. White, T. Rades, R.H. Furneaux, P.C. Tyler, S. Hook, Mannosylated liposomes asantigen delivery vehicles for targeting to dendritic cells, J. Pharm. Pharmacol. 58(6) (2006) 729–737.
[15] M.J. Copland,M.A.Baird, T. Rades, J.L.McKenzie, B. Becker, F. Reck,P.C. Tyler,N.M.Davies,Liposomal delivery of antigen to human dendritic cells, Vaccine 21 (9–10) (2003)883–890.
[16] Y. Ikehara, T. Niwa, L. Biao, S. Ikehara, N. Ohashi, T. Kobayashi, Y. Shimizu, N. Kojima,H. Nakanishi, A carbohydrate recognition based drug delivery and controlledrelease system using intraperitoneal macrophages as a cellular vehicle, Cancer Res.66 (17) (2006) 8740–8748.
[17] Y. Ikehara, N. Kojima, Development of a novel oligomannose-coated liposome-based anticancer drug delivery system for intraperitoneal cancer, Curr. Opin. Mol.Ther. 9 (1) (2007) 53–61.
[18] H. Takagi, N. Furuya, N. Kojima, Preferential production of IL-12 by peritonealmacrophages activated by liposomes prepared from neoglycolipids containingoligomannose residues, Cytokine 40 (3) (2007) 241–250.
[19] Y. Ikehara, N. Shiuchi, S. Kabata-Ikehara, H. Nakanishi, N. Yokoyama, H. Takagi,T. Nagata, Y. Koide, K. Kuzushima, T. Takahashi, K. Tsujimura, N. Kojima,Effective induction of anti-tumor immune responses with oligomannose-coated liposome targeting to intraperitoneal macrophage, Cancer Lett. 260(1–2) (2008) 137–145.
[20] Y. Shimizu, K. Yamakami, T. Gomi, M. Nakata, H. Asanuma, T. Tadakuma, N. Kojima,Protection against Leishmania major infection by oligomannose-coated liposomes,Bioorg. Med. Chem. 11 (7) (2003) 1191–1195.
[21] Y. Shimizu,H. Takagi, T. Nakayama, K. Yamakami, T. Tadakuma,N. Yokoyama,N.Kojima,Intraperitoneal immunizationwith oligomannose-coated liposome-entrapped solubleleishmanial antigen induces antigen-specific T-helper type 1 immune response inBalb/c mice through uptake by peritoneal macrophages, Parasite Immunol. 29 (5)(2007) 229–239.
[22] M.W. Moore, F.R. Carbone, M.J. Bevan, Introduction of soluble protein into the classI pathway of antigen processing and presentation, Cell 54 (6) (1988) 777–785.
[23] S.T. Reddy, M.A. Swart, J.A. Hubbell, Targeting dendritic cells with biomaterials:developing the next generation of vaccines, Trends Immunol. 27 (12) (2006)573–579.
[24] T.J. Curiel, G. Coukos, L. Zou, X. Alvarez, P. Cheng, P. Mottram,M. Evdemon-Hogan, J.R.Conejo-Garcia, L. Zhang, M. Burow, Y. Zhu, S. Wei, I. Kryczek, B. Daniel, A. Gordon, L.Myers, A. Lackner, M.L. Disis, K.L. Knutson, L. Chen, W. Zou, Specific recruitment ofregulatory T cells in ovarian carcinoma fosters immune privilege and predictsreduced survival, Nat. Med. 10 (9) (2004) 942–949.
[25] C.N. Baxevanis, N.N. Sotiriadou, A.D. Gritzapis, P.A. Sotiropoulou, S.A. Perez, N.T.Cacoullos, M. Papamichail, Immunogenic HER-2/neu peptides as tumor vaccines,Cancer Immunol. Immunother. 55 (1) (2006) 85–95.
[26] A.E. Chang, B.G. Redman, J.R. Whitfield, B.J. Nickoloff, T.M. Braun, P.P. Lee, J.D.Geiger, J.J. Mule, A phase I trial of tumor lysate-pulsed dendritic cells in thetreatment of advanced cancer, Clin. Cancer Res. 8 (4) (2002) 1021–1032.
[27] A.M. Krieg, CpG motifs in bacterial DNA and their immune effects, Annu. Rev.Immunol. 20 (2002) 709–760.
[28] B. Jahrsdorfer, G.J. Weiner, CpG oligodeoxynucleotides for immune stimulation incancer immunotherapy, Curr. Opin. Investig. Drugs 4 (6) (2003) 686–690.
[29] D. van Duin, R. Medzhitov, A.C. Shaw, Triggering TLR signaling in vaccination,Trends Immunol. 27 (1) (2006) 49–55.
[30] M. Heikenwalder, M. Polymenidou, T. Junt, C. Sigurdson, H. Wagner, S. Akira, R.Zinkernagel, A. Aguzzi, Lymphoid follicle destruction and immunosuppressionafter repeated CpG oligodeoxynucleotide administration, Nat. Med. 10 (2) (2004)187–192.









![Three decades of messenger RNA vaccine development · information to antigen-presenting cells (APCs) [3, 4]. As pioneers, Martinon et al. showed that liposomes containing mRNA encoding](https://img.pdfslide.net/doc/110x75/60b8282dbb75f712773ecb5d/three-decades-of-messenger-rna-vaccine-development-information-to-antigen-presenting.jpg)



















