Embed Size (px)
Citation preview

UNCORRECTED PROOF
Ontogenetic change in novel functions: waterfall climbingin adult Hawaiian gobiid fishes
R. W. Blob1, K. M. Wright1, M. Becker2, T. Maie1, T. J. Iverson2, M. L. Julius2 & H. L. Schoenfuss2
1 Department of Biological Sciences, Clemson University, Clemson, SC, USA
2 Department of Biological Sciences, St. Cloud State University, Math and Science Center, St Cloud, MN, USA
Keywords
locomotion; performance; kinematics;
ontogeny; climbing; biomechanics; fish;
muscle.
Correspondence
Richard W. Blob, Department of Biological
Sciences, Clemson University, 132 Long
Hall, Clemson, SC, 29634, USA.
Tel: + 1 864 656 3602;
Fax: + 1 864 656 0435
Email: [email protected]
Received 31 October 2006; accepted
19 January 2007
doi:10.1111/j.1469-7998.2007.00315.x
Abstract
Juveniles from three species of Hawaiian gobiid fish climb waterfalls as part of an
amphidromous life cycle, allowing them to re-penetrate adult upstream habitats
after being swept out to the ocean upon hatching. The importance of climbing for
juvenile stream gobies is well established, but adult fish in upstream island habitats
also face potential downstream displacement by periodic disturbances. Thus,
retention of climbing ability could be advantageous for adult stream gobies.
Climbing performance might be expected to decline among adults, however, due
to the tendency for mass-specific muscular power production to decrease with
body size, and a lack of positively allometric growth among structures like the
pelvic sucker that support body weight against gravity. To evaluate changes in
waterfall-climbing ability with body size in Hawaiian stream gobies, we compared
the climbing performance and kinematics between adults and juveniles from three
species Awaous guamensis, Sicyopterus stimpsoni and Lentipes concolor. For
species in which juveniles climbed using ‘powerbursts’ of axial undulation, adult
performance and kinematics showed marked changes: adult A. guamensis failed to
climb, and adult L. concolor used multiple pectoral fin adductions to crutch up
surfaces at slow speeds, rather than rapid powerbursts. Adult S. stimpsoni, like
juveniles, still used oral and pelvic suckers to ‘inch’ up surfaces and climbed
at speeds comparable to those of juveniles. However, unlike juveniles, adult
S. stimpsoni also add pectoral fin crutching to every climbing cycle. Thus, although
powerburst species appear to be particularly susceptible to size-related declines in
waterfall-climbing performance, the addition of compensatory mechanisms pre-
vents the loss of this novel function in some species.
Introduction
Physical forces of the external environment place many
functional demands on animals. As animals grow, these
forces may change as a function of an animal’s increase in
size. To accommodate growth-related changes in external
forces, animals often exhibit compensatory mechanisms
such as allometric growth of support or propulsive struc-
tures (Carrier, 1996; Toro et al., 2003), or changes in
behavior (Higham et al., 2005), that help maintain func-
tional performance at a comparable level throughout
growth. Without such mechanisms, the performance of
some functions could decrease as juveniles mature into
adults, even if those functions continued to be advantageous
among larger individuals (Maie, Schoenfuss & Blob, in
review).
The freshwater fauna of small volcanic islands encounter
particularly challenging environments. Streams on these
islands are commonly subject to flash floods, high-velocity
flows and waterfalls that obstruct access to upstream
reaches (Fitzsimons & Nishimoto, 1995; Fig. 1a). Although
the physical forces exerted by these hazards can be rigorous
for stream-dwelling animals, many species have evolved
novel functional capacities that help meet such demands.
For example, in the Hawaiian Islands, the five native species
of stream fishes (four gobies and one eleotrid) hatch in
freshwater but are swept by currents to the ocean, where
they share an oceanic larval phase for several months before
returning to freshwater as juveniles (Ego, 1956; Julius, Blob
& Schoenfuss, 2005). However, three of these species face
further challenges because their adult habitats are in up-
stream reaches that can be obstructed by waterfalls up to
thousands of body lengths tall (Fitzsimons & Nishimoto,
1995). Juveniles of these species use two different mechan-
isms to climb waterfalls (Nishimoto & Kuamo’o, 1992;
Schoenfuss & Blob, 2003). Juvenile Awaous guamensis (Fig.
1b) and Lentipes concolor (Fig. 1c) climb using rapid ‘power-
bursts’ of axial undulation initiated by pectoral fin adduc-
tion. These are followed by rest periods during which fish
adhere to the substrate using a ventral sucking disk formed
from fused pelvic fins. In contrast, the third species, Sicyop-
terus stimpsoni (Fig. 1d), slowly inches up waterfalls using
J Z O 3 1 5 B Dispatch: 5.3.07 Journal: JZO CE: Chandrika
Journal Name Manuscript No. Author Received: No. of pages: 10 TE: Premalatha Op: gsravi
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London 1
Journal of Zoology. Print ISSN 0952-8369
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)

UNCORRECTED PROOFthe pelvic disk and a second, oral disk with little or no fin
motion or axial undulation. The use of these mechanisms
allows repopulation of upstream habitats by oceanic larvae
after disturbances such as floods or hurricanes (Fitzsimons
& Nishimoto, 1995).
Although the importance of climbing to juvenile
A. guamensis, L. concolor and S. stimpsoni has been estab-
lished, adult fish in upstream Hawaiian habitats face many
similar risks, including displacement by catastrophic distur-
bances (Fitzsimons & Nishimoto, 1995). As a result, the
retention of climbing ability could be advantageous for
adult fish. However, as gobies increase in size, their ability
to climb waterfalls might decline for several reasons. First,
because mass-specific power production tends to decrease
with body size (Irschick et al., 2003), size-related limits to
power production might restrict the maximum body size at
which ‘powerbursting’ is a viable climbing mechanism (Blob
et al., 2006). This might limit climbing ability in adult
A. guamensis and L. concolor, or force these species to adopt
different climbing mechanics. Power demands might not
limit climbing by S. stimpsoni because of the slow cycle
frequencies of inching (Schoenfuss & Blob, 2003). However,
studies of ontogenetic scaling of body proportions in gobies
(including S. stimpsoni) have shown that the area of the
pelvic sucking disk grows isometrically with body length (i.e.
scales / BL2) and, thus, does not keep pace with increases
in the mass of the body (/ BL3) that it must keep attached
to the substrate (Maie et al., in review). As a result, even
inching climbers like S. stimpsoni might show reduced
climbing performance as they grow larger.
In this study, we test for size-related changes in climbing
performance and behavior by evaluating the climbing
of adult stream gobies (A. guamensis, L. concolor and
S. stimpsoni) and comparing their performance and kine-
matics with previous data (Schoenfuss & Blob, 2003; Blob
et al., 2006) from juveniles. Because of possible limits to
muscular power production and isometric scaling of the
adhesive sucker, we expect adult climbing performance to
decrease relative to that of conspecific juveniles. These
changes might be especially dramatic for powerburst species
because their high locomotor frequencies (Schoenfuss &
Blob, 2003) are disadvantageous for the high power produc-
tion required for climbing by larger fishes. Thus, species that
use powerburst climbing as juveniles might require major
changes in locomotor behavior in order to climb as adults.
Materials and methods
During three field seasons (2003–2005), adult climbing
gobies were collected with ‘opae nets’ (bowl-shaped mesh
nets with a narrow mouth) from habitats above waterfalls
in mid- (A. guamensis and S. stimpsoni) or upstream
(L. concolor) reaches of Hakalau Stream on the Island of
Hawai’i. Within 2 h of capture, animals were transferred in
stream water to a facility provided by the Hawai’i Depart-
ment of Land and Natural Resources, Division of Aquatic
Resources (DAR). Animals were separated by species into
groups of three to five individuals and acclimated together
for several hours at ambient stream temperature (19 1C).
Trials were conducted on individuals that overlapped in
body size (Table 1).
Our climbing arena (Fig. 2) was a chute that extended up
from a catch basin (Blob et al., 2006). The basin was 60 cm
long� 45 cm wide� 15 cm deep, half filled with Hakalau
stream water, and contained a rock to provide cover for the
fish. Chutes extended from the basin at 571 from horizontal,
and consisted of 1.5m long sections of a 4 in. diameter PVC
pipe (cut in half longitudinally) with fine-grained sand
Figure 1 (a) Falls of Nanue Stream on the Hamakua Coast of the island of Hawai’i, a waterfall climbed by juveniles of all three species of climbing
Hawaiian stream gobies: powerburst climbing Awaous guamensis and Lentipes concolor, and the inching climber Sicyopterus stimpsoni. (b)
Photo of adult A. guamensis. (c) Photo of adult male L. concolor. (d) Photo of adult male S. stimpsoni. Photos in parts b–d were provided courtesy
of Mike Yamamoto; all fish were free-swimming wild adults.
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London2
Ontogeny of waterfall climbing in gobiid fishes R. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)
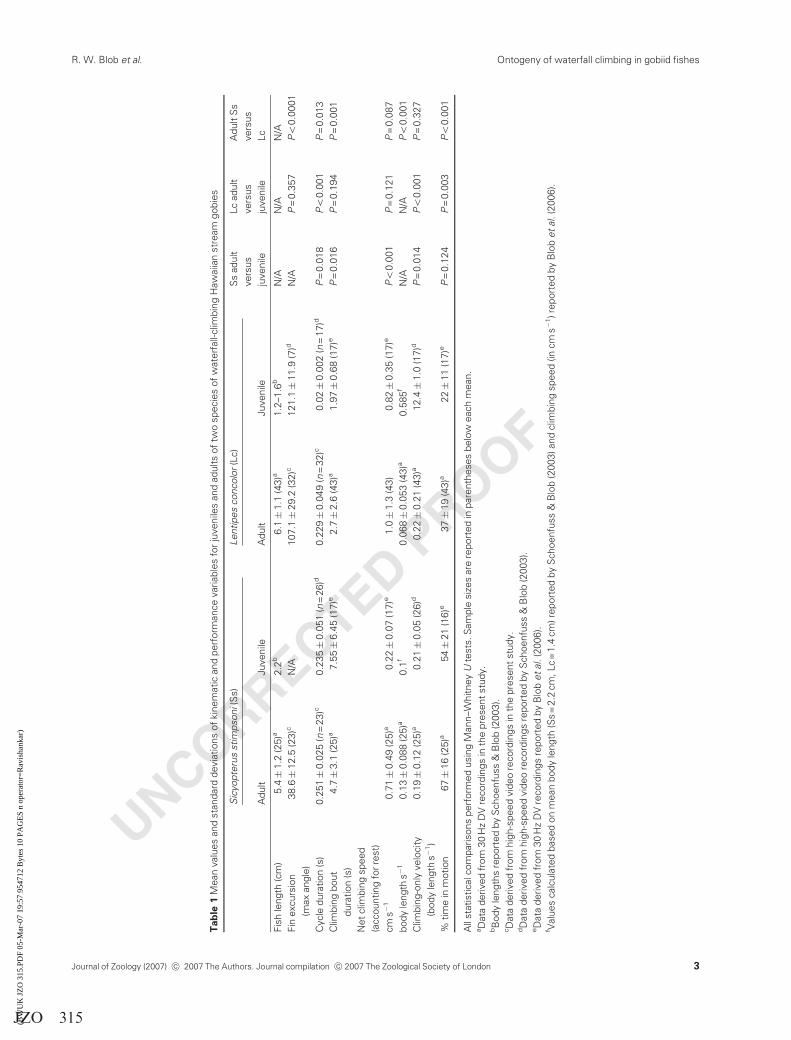
UNCORRECTED PROOF
Tab
le1
Mean
valu
es
and
sta
ndard
devia
tions
of
kin
em
atic
and
perf
orm
ance
variable
sfo
rju
venile
sand
adults
of
two
specie
sof
wate
rfall-
clim
bin
gH
aw
aiia
nstr
eam
gobie
s
Sic
yopte
rus
stim
psoni(S
s)
Lentipes
concolo
r(L
c)
Ss
adult
vers
us
juvenile
Lc
adult
vers
us
juvenile
Adult
Ss
vers
us
Lc
Adult
Juvenile
Adult
Juvenile
Fis
hle
ngth
(cm
)5.4�
1.2
(25)a
2.2
b6.1�
1.1
(43)a
1.2
–1.6
bN
/AN
/AN
/A
Fin
excurs
ion
(max
angle
)
38.6�
12.5
(23)c
N/A
107.1�
29.2
(32)c
121.1�
11.9
(7)d
N/A
P=
0.3
57
Po
0.0
001
Cycle
dura
tion
(s)
0.2
51�
0.0
25
(n=
23)c
0.2
35�
0.0
51
(n=
26)d
0.2
29�
0.0
49
(n=
32)c
0.0
2�
0.0
02
(n=
17)d
P=
0.0
18
Po
0.0
01
P=
0.0
13
Clim
bin
gbout
dura
tion
(s)
4.7�
3.1
(25)a
7.5
5�
6.4
5(1
7)e
2.7�
2.6
(43)a
1.9
7�
0.6
8(1
7)e
P=
0.0
16
P=
0.1
94
P=
0.0
01
Net
clim
bin
gspeed
(accounting
for
rest)
cm
s�
10.7
1�
0.4
9(2
5)a
0.2
2�
0.0
7(1
7)e
1.0�
1.3
(43)
0.8
2�
0.3
5(1
7)e
Po
0.0
01
P=
0.1
21
P=
0.0
87
body
length
s�
10.1
3�
0.0
88
(25)a
0.1
f0.0
68�
0.0
53
(43)a
0.5
85
fN
/AN
/APo
0.0
01
Clim
bin
g-o
nly
velo
city
(body
length
s�
1)
0.1
9�
0.1
2(2
5)a
0.2
1�
0.0
5(2
6)d
0.2
2�
0.2
1(4
3)a
12.4�
1.0
(17)d
P=
0.0
14
Po
0.0
01
P=
0.3
27
%tim
ein
motion
67�
16
(25)a
54�
21
(16)e
37�
19
(43)a
22�
11
(17)e
P=
0.1
24
P=
0.0
03
Po
0.0
01
All
sta
tisticalcom
parisons
perf
orm
ed
usin
gM
ann–W
hitney
Ute
sts
.S
am
ple
siz
es
are
report
ed
inpare
nth
eses
belo
weach
mean.
aD
ata
derived
from
30
Hz
DV
record
ings
inth
epre
sent
stu
dy.
bB
ody
length
sre
port
ed
by
Schoenfu
ss
&B
lob
(2003).
cD
ata
derived
from
hig
h-s
peed
vid
eo
record
ings
inth
epre
sent
stu
dy.
dD
ata
derived
from
hig
h-s
peed
vid
eo
record
ings
report
ed
by
Schoenfu
ss
&B
lob
(2003).
eD
ata
derived
from
30
Hz
DV
record
ings
report
ed
by
Blo
bet
al.
(2006).
f Valu
es
calc
ula
ted
based
on
mean
body
length
(Ss
=2.2
cm
,Lc
=1.4
cm
)re
port
ed
by
Schoenfu
ss
&B
lob
(2003)and
clim
bin
gspeed
(in
cm
s�
1)re
port
ed
by
Blo
bet
al.
(2006).
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London 3
Ontogeny of waterfall climbing in gobiid fishesR. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)

UNCORRECTED PROOF
(grains mean� SD=392� 99 mm, n=50) glued to the pipe.
Stream water was run over chutes for several days before
experiments to remove chemical residue, and a distance
scale was marked on each chute. To generate flow over the
climbing surface during trials, a siphon directed Hakalau
stream water down the chute from a 20L bucket at
200mLmin�1 (within natural flow rates: H. L. Schoenfuss,
pers. obs.). We began each trial by introducing three to five
fish (from a single species) into the basin with water flowing
down the chute. If climbing did not commence within
20min, the fish were removed and replaced with a new
group.
Two types of video records were collected. First, to
measure the climbing performance over a distance of several
body lengths, a SonyQ1 DTV 1020 digital camcorder (30Hz
framing rate) filmed animals in dorsal view as they ascended
the chute. Camera field of view covered 20 cm of pipe
c. 10 cm above the water level. Each fish was used in a single
trial and then returned to the stream. Trials were only
analyzed if the fish climbed the entire 20 cm. DV tape
footage was transferred to a computer (Macintosh G4)
using iMovie software. One of us (M. B.) tracked individual
animals frame by frame. The beginning and end of each
climbing bout were recorded, as were rest periods between
bouts and instances of pectoral fin and tail use during
climbing.
Second, to evaluate detailed climbing kinematics, a high-
speed digital video was collected for a subset of trials using
either a PhantomQ2 V4.1 (200Hz) or Redlake MotionscopeQ3 (250Hz) camera. Fish were filmed in dorsal view as they
climbed the chute and trials were saved as AVI files. The
positions of anatomical landmarks on the fishes were
digitized for each analyzed video frame using a modification
of the public domain NIH Image program for Macintosh,
developed at the US National Institutes of Health (the
modification, QuickImage, was developed by J. Walker and
is available at http://www.usm.maine.edu/�walker/software.html). For the inching climber S. stimpsoni, points at the
base and tip of both pectoral fins were digitized along with
four points on the head representing the maximal anterior,
left and right margins of the oral sucker, as well as the
midpoint between the eyes. At least nine additional points
along the body midline were also digitized, with the most
posterior point at the base of the caudal peduncle. For
species without an oral sucker, the same points were
digitized, except for those on head, for which the three
sucker landmarks were replaced with points at the anterior
midline of the face and the left and right eyes.
Custom programs (Matlab 5.0; Mathworks, Inc.; Natick,
MA, USA) calculated kinematic variables from digitized
coordinates. For all trials, fin movements and the displace-
ment of the front lip through each cycle were evaluated. The
angle of each fin to the direction of travel (determined by the
two midline head points) was calculated for each frame,
allowing us to calculate profiles of fin movements through-
out locomotor cycles and fin excursion angles. In addition,
for S. stimpsoni, an index of the instantaneous area of the
oral sucker was calculated as
Mouth area ¼ ðp=4ÞðDLRÞðDAPÞ ð1Þ
where DLR is the distance between the left and right edges of
the mouth and DAP is the distance from the point between
the eyes to the anterior edge of the mouth. This index
simplifies mouth shape, but allows evaluation of whether
the oral disk is increasing or decreasing in area (Schoenfuss
& Blob, 2003). After evaluating these parameters, we used
QuickSAND (Walker, 1998; available at http://www.usm.
maine.edu/�walker/software.html) to fit a quintic spline to
the kinematic calculations for each trial, smooth the data
and normalize all trials to the same duration to calculate the
mean kinematic profiles for each variable.
Statistical analyses were performed using GraphPad
Prism 4.0c (San Diego, CA, USA) or Statview 5.0 (SAS
Institute Inc, Cary, NC, USA) for Macintosh. The mean
locomotor cycle duration for each species was calculated
from a high-speed video, whereas the mean durations of
climbing and rest periods for entire climbing bouts (over the
20 cm distance) were calculated from standard video. Net
climbing speed (accounting for rest periods) was calculated
by dividing the 20 cm distance by the sum of climbing and
rest times. In addition, the percentage of time that each
individual spent moving was calculated as time in motion
divided by the sum of motion and rest times. Differences in
kinematic and performance variables between species and
between conspecific juveniles and adults were evaluated
using non-parametric Mann–Whitney U tests.
Results
Climbing by adult gobies: comparisons withjuveniles
Awaous guamensis
In contrast to powerbursting juveniles (Blob et al., 2006),
none of the 10 adult A. guamensis successfully climbed (or
Mini-DVcamera
High-speedcamera
Climbingchute
Catch basin
Siphon
Goby
Source bucketfor water
Figure 2 Illustration of experimental arena used for climbing trials with
adult Hawaiian stream gobies.
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London4
Ontogeny of waterfall climbing in gobiid fishes R. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)
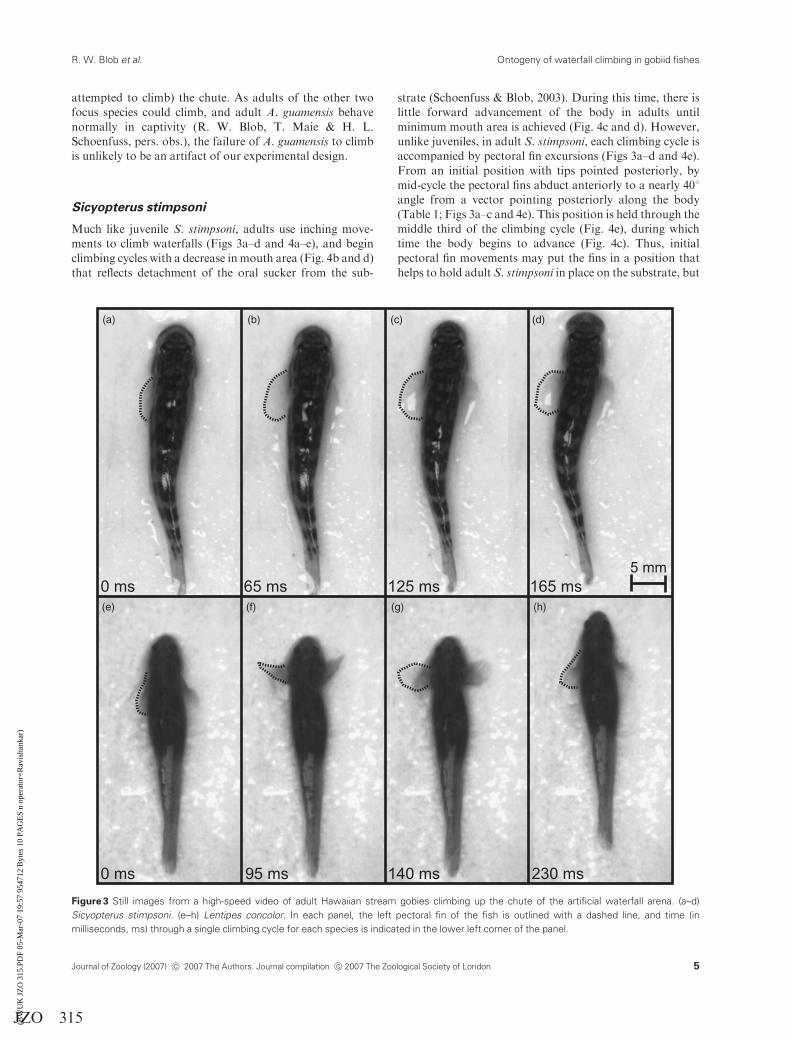
UNCORRECTED PROOF
attempted to climb) the chute. As adults of the other two
focus species could climb, and adult A. guamensis behave
normally in captivity (R. W. Blob, T. Maie & H. L.
Schoenfuss, pers. obs.), the failure of A. guamensis to climb
is unlikely to be an artifact of our experimental design.
Sicyopterus stimpsoni
Much like juvenile S. stimpsoni, adults use inching move-
ments to climb waterfalls (Figs 3a–d and 4a–e), and begin
climbing cycles with a decrease in mouth area (Fig. 4b and d)
that reflects detachment of the oral sucker from the sub-
strate (Schoenfuss & Blob, 2003). During this time, there is
little forward advancement of the body in adults until
minimum mouth area is achieved (Fig. 4c and d). However,
unlike juveniles, in adult S. stimpsoni, each climbing cycle is
accompanied by pectoral fin excursions (Figs 3a–d and 4e).
From an initial position with tips pointed posteriorly, by
mid-cycle the pectoral fins abduct anteriorly to a nearly 401
angle from a vector pointing posteriorly along the body
(Table 1; Figs 3a–c and 4e). This position is held through the
middle third of the climbing cycle (Fig. 4e), during which
time the body begins to advance (Fig. 4c). Thus, initial
pectoral fin movements may put the fins in a position that
helps to hold adult S. stimpsoni in place on the substrate, but
(a) (b) (c) (d)
(h)(g)(f)(e)
Figure 3 Still images from a high-speed video of adult Hawaiian stream gobies climbing up the chute of the artificial waterfall arena. (a–d)
Sicyopterus stimpsoni. (e–h) Lentipes concolor. In each panel, the left pectoral fin of the fish is outlined with a dashed line, and time (in
milliseconds, ms) through a single climbing cycle for each species is indicated in the lower left corner of the panel.
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London 5
Ontogeny of waterfall climbing in gobiid fishesR. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)
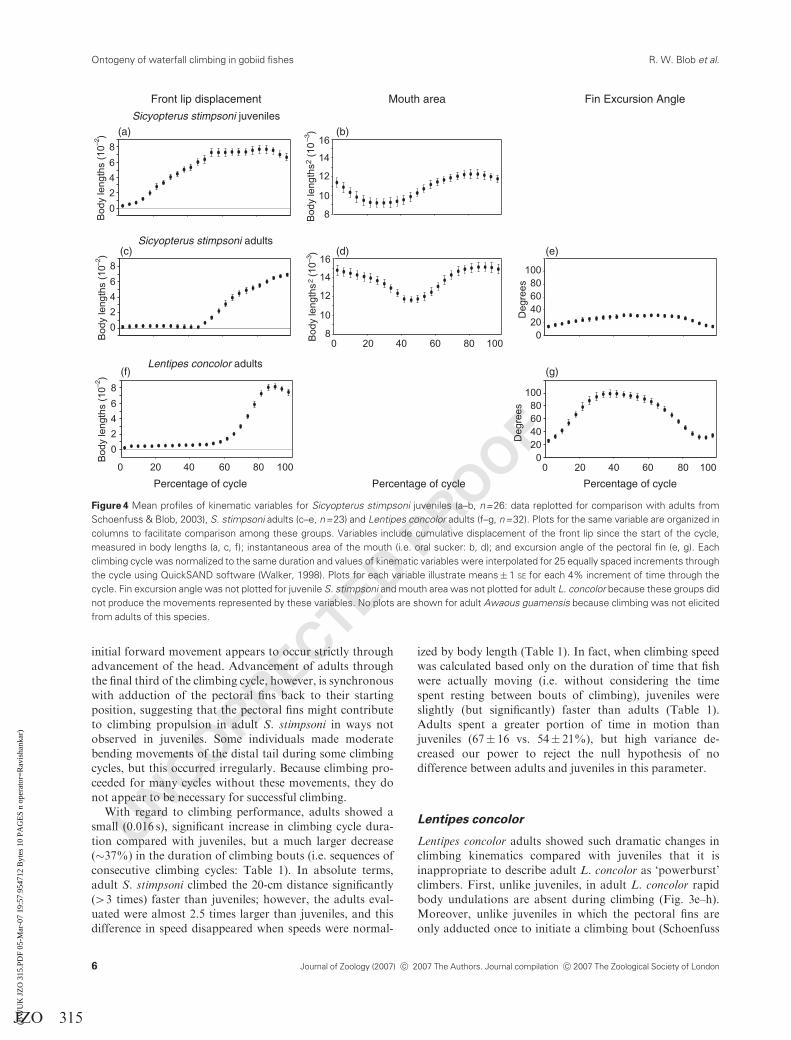
UNCORRECTED PROOF
initial forward movement appears to occur strictly through
advancement of the head. Advancement of adults through
the final third of the climbing cycle, however, is synchronous
with adduction of the pectoral fins back to their starting
position, suggesting that the pectoral fins might contribute
to climbing propulsion in adult S. stimpsoni in ways not
observed in juveniles. Some individuals made moderate
bending movements of the distal tail during some climbing
cycles, but this occurred irregularly. Because climbing pro-
ceeded for many cycles without these movements, they do
not appear to be necessary for successful climbing.
With regard to climbing performance, adults showed a
small (0.016 s), significant increase in climbing cycle dura-
tion compared with juveniles, but a much larger decrease
(�37%) in the duration of climbing bouts (i.e. sequences of
consecutive climbing cycles: Table 1). In absolute terms,
adult S. stimpsoni climbed the 20-cm distance significantly
(43 times) faster than juveniles; however, the adults eval-
uated were almost 2.5 times larger than juveniles, and this
difference in speed disappeared when speeds were normal-
ized by body length (Table 1). In fact, when climbing speed
was calculated based only on the duration of time that fish
were actually moving (i.e. without considering the time
spent resting between bouts of climbing), juveniles were
slightly (but significantly) faster than adults (Table 1).
Adults spent a greater portion of time in motion than
juveniles (67� 16 vs. 54� 21%), but high variance de-
creased our power to reject the null hypothesis of no
difference between adults and juveniles in this parameter.
Lentipes concolor
Lentipes concolor adults showed such dramatic changes in
climbing kinematics compared with juveniles that it is
inappropriate to describe adult L. concolor as ‘powerburst’
climbers. First, unlike juveniles, in adult L. concolor rapid
body undulations are absent during climbing (Fig. 3e–h).
Moreover, unlike juveniles in which the pectoral fins are
only adducted once to initiate a climbing bout (Schoenfuss
(a) (b)
(d) (e)
(g)(f)
(c)
Front lip displacement Mouth area Fin Excursion Angle
Sicyopterus stimpsoni juveniles
Sicyopterus stimpsoni adults
Lentipes concolor adults
Figure 4 Mean profiles of kinematic variables for Sicyopterus stimpsoni juveniles (a–b, n=26: data replotted for comparison with adults from
Schoenfuss & Blob, 2003), S. stimpsoni adults (c–e, n=23) and Lentipes concolor adults (f–g, n=32). Plots for the same variable are organized in
columns to facilitate comparison among these groups. Variables include cumulative displacement of the front lip since the start of the cycle,
measured in body lengths (a, c, f); instantaneous area of the mouth (i.e. oral sucker: b, d); and excursion angle of the pectoral fin (e, g). Each
climbing cycle was normalized to the same duration and values of kinematic variables were interpolated for 25 equally spaced increments through
the cycle using QuickSAND software (Walker, 1998). Plots for each variable illustrate means� 1 SE for each 4% increment of time through the
cycle. Fin excursion angle was not plotted for juvenile S. stimpsoni and mouth area was not plotted for adult L. concolor because these groups did
not produce the movements represented by these variables. No plots are shown for adult Awaous guamensis because climbing was not elicited
from adults of this species.
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London6
Ontogeny of waterfall climbing in gobiid fishes R. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)
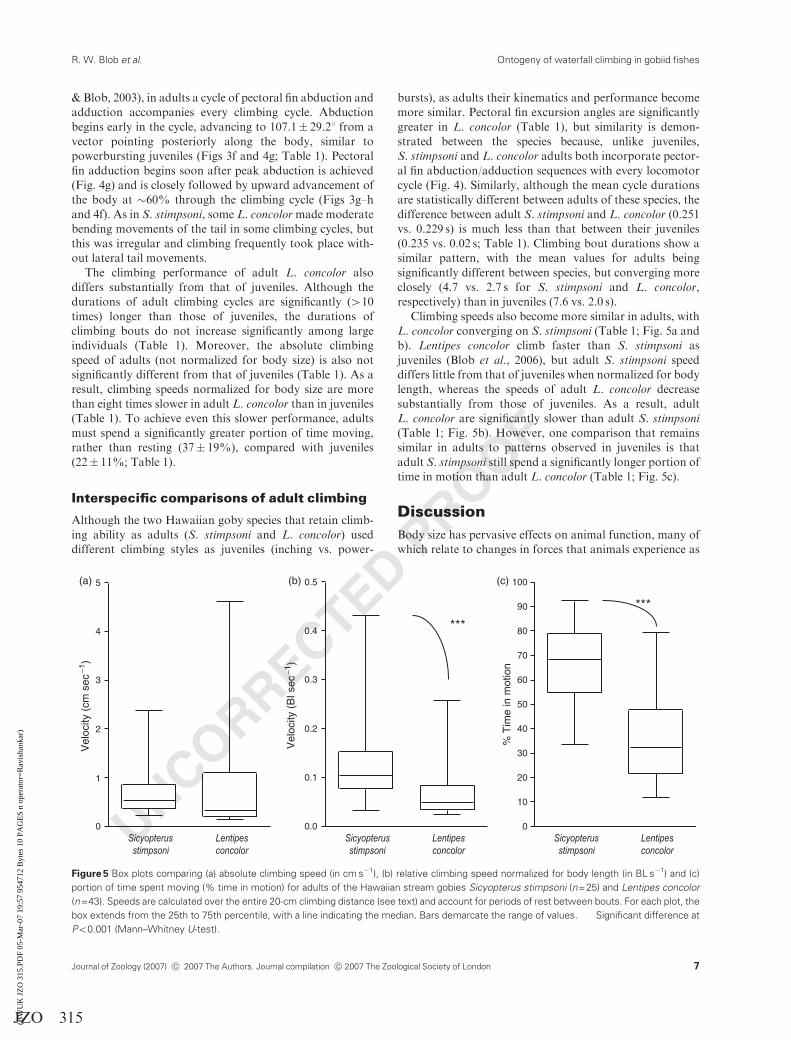
UNCORRECTED PROOF
&Blob, 2003), in adults a cycle of pectoral fin abduction and
adduction accompanies every climbing cycle. Abduction
begins early in the cycle, advancing to 107.1� 29.21 from a
vector pointing posteriorly along the body, similar to
powerbursting juveniles (Figs 3f and 4g; Table 1). Pectoral
fin adduction begins soon after peak abduction is achieved
(Fig. 4g) and is closely followed by upward advancement of
the body at �60% through the climbing cycle (Figs 3g–h
and 4f). As in S. stimpsoni, some L. concolormade moderate
bending movements of the tail in some climbing cycles, but
this was irregular and climbing frequently took place with-
out lateral tail movements.
The climbing performance of adult L. concolor also
differs substantially from that of juveniles. Although the
durations of adult climbing cycles are significantly (410
times) longer than those of juveniles, the durations of
climbing bouts do not increase significantly among large
individuals (Table 1). Moreover, the absolute climbing
speed of adults (not normalized for body size) is also not
significantly different from that of juveniles (Table 1). As a
result, climbing speeds normalized for body size are more
than eight times slower in adult L. concolor than in juveniles
(Table 1). To achieve even this slower performance, adults
must spend a significantly greater portion of time moving,
rather than resting (37� 19%), compared with juveniles
(22� 11%; Table 1).
Interspecific comparisons of adult climbing
Although the two Hawaiian goby species that retain climb-
ing ability as adults (S. stimpsoni and L. concolor) used
different climbing styles as juveniles (inching vs. power-
bursts), as adults their kinematics and performance become
more similar. Pectoral fin excursion angles are significantly
greater in L. concolor (Table 1), but similarity is demon-
strated between the species because, unlike juveniles,
S. stimpsoni and L. concolor adults both incorporate pector-
al fin abduction/adduction sequences with every locomotor
cycle (Fig. 4). Similarly, although the mean cycle durations
are statistically different between adults of these species, the
difference between adult S. stimpsoni and L. concolor (0.251
vs. 0.229 s) is much less than that between their juveniles
(0.235 vs. 0.02 s; Table 1). Climbing bout durations show a
similar pattern, with the mean values for adults being
significantly different between species, but converging more
closely (4.7 vs. 2.7 s for S. stimpsoni and L. concolor,
respectively) than in juveniles (7.6 vs. 2.0 s).
Climbing speeds also become more similar in adults, with
L. concolor converging on S. stimpsoni (Table 1; Fig. 5a and
b). Lentipes concolor climb faster than S. stimpsoni as
juveniles (Blob et al., 2006), but adult S. stimpsoni speed
differs little from that of juveniles when normalized for body
length, whereas the speeds of adult L. concolor decrease
substantially from those of juveniles. As a result, adult
L. concolor are significantly slower than adult S. stimpsoni
(Table 1; Fig. 5b). However, one comparison that remains
similar in adults to patterns observed in juveniles is that
adult S. stimpsoni still spend a significantly longer portion of
time in motion than adult L. concolor (Table 1; Fig. 5c).
Discussion
Body size has pervasive effects on animal function, many of
which relate to changes in forces that animals experience as
1000.55
4
3
2
1
0
0.4
0.3
0.2
0.1
0.0
% T
ime
in m
otio
n
Vel
ocity
(B
I sec
−1)
Vel
ocity
(cm
sec
−1)
90
80
70
60
50
40
30
20
10
0
(a) (b) (c)
***
***
Figure 5 Box plots comparing (a) absolute climbing speed (in cm s�1), (b) relative climbing speed normalized for body length (in BL s�1) and (c)
portion of time spent moving (% time in motion) for adults of the Hawaiian stream gobies Sicyopterus stimpsoni (n=25) and Lentipes concolor
(n=43). Speeds are calculated over the entire 20-cm climbing distance (see text) and account for periods of rest between bouts. For each plot, the
box extends from the 25th to 75th percentile, with a line indicating the median. Bars demarcate the range of values. ���Significant difference at
Po0.001 (Mann–Whitney U-test).
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London 7
Ontogeny of waterfall climbing in gobiid fishesR. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)

UNCORRECTED PROOF
they grow (Schmidt-Nielsen, 1984; McHenry & Lauder,
2005). Unless growing animals adopt mechanisms to accom-
modate these changing forces, the performance of many
functions might be impaired or precluded in larger indivi-
duals (Maie et al., in review). The three species of Hawaiian
gobies that climb waterfalls as juveniles show differing
functional responses as they increase in size. In A. guamen-
sis, climbing ability appears to have been lost in adults.
However, in the other two species of climbing Hawaiian
gobies, changes in locomotor kinematics preserve at least
some climbing ability among adults, allowing continued
opportunity for this novel behavior to meet the functional
requirements of a habitat with extreme physical demands.
Ontogenetic changes in waterfall climbing:mechanisms for accommodating larger bodysize
Three general mechanisms might enable animals to accom-
modate size-related increases in locomotor forces that they
experience (or must produce) due to growth: allometric
increases in morphological features (e.g. fin breadth); allo-
metric changes in physiological parameters (e.g. muscle
force generation); or changes in locomotor behavior.
Although morphological and physiological scaling have
not been completely surveyed in climbing gobies, at least
one feature integral to climbing, the area of the pelvic
sucker, fails to exhibit allometric growth that could help
accommodate the greater gravitational forces that must be
resisted by larger individuals as they climb (Maie et al., in
review). This led us to predict that if adult gobies could
climb, they would be expected to show behavioral adjust-
ments relative to juveniles that would facilitate maintenance
of climbing performance. Such adjustments appeared in
both species that continued to climb as adults. In adult S.
stimpsoni and L. concolor, pectoral fin movements were
added to every climbing cycle, contrasting with juveniles in
which the pectoral fins were used once (L. concolor) or not at
all (S. stimpsoni) during climbing bouts. In S. stimpsoni,
pectoral fin movements are less extensive than in L. con-
color, and fin adduction is delayed until after forward
advancement has commenced (Fig. 4c and e). Thus, exten-
sion of the fins may initially stabilize the body of adult
S. stimpsoni against the substrate during extension of the
head, only aiding forward propulsion toward the end of
each cycle. In contrast, the sweeping pectoral adductions of
adult L. concolor synchronize closely with body advance-
ment, and appear to provide the primary upward thrust in
this species in which axial motions are reduced compared
with powerbursting juveniles.
Size-related changes in climbing performance and kine-
matics among species using powerbursts as juveniles were
more drastic than those in the inching climber S. stimpsoni:
one powerburst species (A. guamensis) did not climb as
adults, and the other (L. concolor) shifted from using the
body axis as its main propulsor to primary reliance on the
pectoral fins. This divergence in climbing performance and
mechanics between juveniles and adults suggests that power-
bursts may not be a viable climbing mechanism for larger
individuals. Adult A. guamensis and L. concolor still appear
capable of performing body movements associated with
powerburst climbing: body axis movements seen in juvenile
climbers (Schoenfuss & Blob, 2003) are seen in swimming
adults (R. W. Blob, H. L. Schoenfuss, pers. obs.), and
pectoral fin movements used by climbing adult L. concolor
simply represent more frequent use of a behavior exhibited
by juveniles (Table 1). However, adult powerburst climbing
might be precluded because of its high demand for muscular
power (Blob et al., 2006). Ascension of vertical surfaces has
high power requirements (Irschick et al., 2003), but muscles
contracting at the high frequencies used during powerburst
climbing tend to have reduced power-producing capacities
(Rome, 1998). Because mass-specific power production also
tends to decrease with body size (Irschick et al., 2003), it
might not be possible to climb using rapid movements at
larger body sizes, leaving slower movements like pectoral fin
crutching and the inching of S. stimpsoni as the only viable
climbing mechanisms for adult gobies. Such size-related
limitations could also help explain the lack of powerburst
climbing in juvenile S. stimpsoni, which can be three times
heavier than juveniles from powerburst species and possibly
already restricted in climbing behaviors (Schoenfuss, 1997).
Physiological implications of climbingperformance in adult Hawaiian streamgobies
Like juveniles, both species that successfully climb waterfalls
as adults use intermittent bouts of locomotion. For other
species moving over terrestrial substrates, the use of inter-
mittent locomotion significantly increases the distance tra-
veled before fatigue (Weinstein & Full, 1998, 1999). This
benefit results if pauses are of appropriate duration (abso-
lute and relative to time spent moving) to allow removal of
fatigue-inducing products and restoration of metabolic fuels
(Allen & Westerblad, 2001; Kramer & McLoughlin, 2001).
Intermittent inching by S. stimpsoni juveniles did not meet
these criteria (Blob et al., 2006), as activity durations ranged
up to twice as long as rest periods, a ratio that does not
typically improve performance (Weinstein & Full, 1998,
1999). Adult S. stimpsoni did not differ significantly from
juveniles in the ratio of time spent climbing versus resting
(Table 1); thus, they also seem unlikely to increase the
distance traveled before fatigue through intermittent climb-
ing. In contrast, juvenile powerburst climbers rest between
three and five times longer than they move (Blob et al.,
2006). With the short durations of their activity periods
(o2 s), the motion-to-rest ratio of juvenile powerburst
climbers is appropriate to increase their pre-fatigue travel
distance (Weinstein & Full, 1998, 1999) at the cold tempera-
tures typical of their stream habitats (18–19 1C: Schoenfuss,
Julius & Blob, 2004). Although adult L. concolor rested for a
significantly shorter portion of time than juveniles, they still
rested for over 1.7 times longer than their activity periods
(i.e. motion-to-rest ratio=0.59); in addition, adult locomo-
tor bouts did not last significantly longer than those of
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London8
Ontogeny of waterfall climbing in gobiid fishes R. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)

UNCORRECTED PROOF
juveniles (Table 1). This ratio of activity to rest is close to
0.5, which the increases distance capacity in species ranging
from crabs to geckos (Weinstein & Full, 1998, 1999). Thus,
although adult L. concolor use climbing mechanisms differ-
ent from juveniles, they may still achieve some of the same
performance benefits.
Slow climbing by adult gobies may also help prevent
fatigue. Climbing cycle frequencies for adult S. stimpsoni
and L. concolor (calculated from cycle durations, Table 1)
average 4.0 and 4.4Hz, respectively, and climbing speeds
(considering motion periods only) are o0.25BL s�1 (Table
1). Motion at such low speeds and frequencies in fish is
typically powered by slow oxidative (red) muscle without
activity in other fiber types (Jayne & Lauder, 1994; Gillis,
1998). If adult gobies used only red muscle during climbing,
their potential for fatigue could be reduced because red
muscle activity depends on aerobic metabolism that is suited
for endurance behaviors (Sanger & Stoiber, 2001). Extensive
fast glycolytic (white) fibers would still be expected in adults,
however, as these would facilitate swimming in rapid cur-
rents, likely their most common locomotor behavior.
Ecological implications of climbingperformance in adult Hawaiian streamgobies
The climbing performance exhibited by adult stream gobies
may relate to several aspects of their ecology. First, because
A. guamensis has the poorest climbing juveniles among the
three climbing species (Blob et al., 2006), it is not surprising
that adults of this species have lost climbing ability that was
retained by the other species. Moreover, the loss of climbing
among larger A. guamensis would help explain why this
species showed the greatest delay in the upstream appear-
ance of reproductive adults in streams on Kaua’i that were
devastated by Hurricane Iniki in 1992 (Fitzsimons & Nishi-
moto, 1995), as repopulation may have been achieved solely
by small individuals and juveniles migrating in from the
ocean.
One of the most striking features of the climbing exhib-
ited by juvenile Hawaiian gobies is the diversity of mechan-
isms and levels of performance that they use. Not only do
juveniles use two dramatically different climbing styles
(inching vs. powerbursts), but different species of power-
burst climbers exhibit different performance. This diversity
resembles the ‘many-to-one’ mapping of morphology to
function documented in fish feeding studies (Alfaro, Bolnick
&Wainwright, 2005; Wainwright et al., 2005), and indicates
that a wide range of performance can be maintained even
under the selective pressures of extreme environmental
conditions (Blob et al., 2006). However, functional diversity
is diminished among adult climbers, suggesting a ‘fewer-to-
one’ mapping of performance to functional demand.
Although there are statistically significant differences in
several performance variables between adult S. stimpsoni
and L. concolor, the magnitude of those differences is
smaller between adults than between juveniles (Table 1).
Moreover, adults of both species incorporate the same
additional propulsive mechanism (pectoral fin adduction)
beyond those seen in juveniles (Table 1). Thus, adult
S. stimpsoni and L. concolor have converged considerably
in their strategies and capacity for climbing waterfalls.
Although functional diversity might be maintained among
juveniles of these species despite strong selection for success-
ful waterfall climbing, the additional pressures of increasing
body size may finally limit the range of mechanics and
performance that can execute this novel behavior.
Acknowledgements
Funding was provided by the Hawai’i DAR (Sport Fish
Restoration Project F-14-R-28) to H.L.S., M.L.J. and
R.W.B.; the Clemson Department of Biological Sciences; a
Raney Award from the American Society of Ichthyologists
and Herpetologists (T.M.); a St Cloud State Faculty Re-
search Grant (H.L.S.); and Stearns Manufacturing Inc. We
thank Bob Nishimoto, Darrell Kuamo’o, Mike Fitzsimons
and Nora Espinoza for suggestions, Christopher Mims for
data analysis and Mike Yamamoto for the photos used in
Fig. 1b and d. We also thank Lance Nishiura, Tim Shindo,
Troy Sakihara, Troy Shimoda and Bruce Kaya for logistical
support at the Hilo DAR station.
References
Alfaro, M.E., Bolnick, D.I. & Wainwright, P.C. (2005).
Evolutionary consequences of many-to-one mapping of
jaw morphology to mechanics in labrid fishes. Am. Nat.
165, E140–E154.
Allen, D.G. & Westerblad, H. (2001). Role of phosphate and
calcium stores in muscle fatigue. J. Physiol. 536, 657–665.
Blob, R.W., Rai, R., Julius, M.L. & Schoenfuss, H.L. (2006).
Functional diversity in extreme environments: effects of
locomotor style and substrate texture on the waterfall
climbing performance of Hawaiian gobiid fishes. J. Zool.
(Lond.) 268, 315–324.
Carrier, D.R. (1996). Ontogenetic limits on locomotor per-
formance. Physiol. Zool. 69, 467–488.
Ego, K. (1956). Life history of fresh water gobies. Project No.
4-4-R. Fresh water game fish management research, Hon-
olulu, Territory of Hawai’i.
Fitzsimons, J.M. & Nishimoto, R.T. (1995). Use of fish
behavior in assessing the effects of Hurricane Iniki on the
Hawaiian island of Kaua’i. Environ. Biol. Fish. 43, 39–50.
Gillis, G.B. (1998). Neuromuscular control of anguilliform
locomotion: patterns of red and white muscle activity
during swimming in the American eel, Anguilla rostrata.
J. Exp. Biol. 201, 3245–3256.
Higham, T.E., Malas, B., Jayne, B.C. & Lauder, G.V. (2005).
Constraints on starting and stopping: behavior compen-
sates for reduced pectoral fin area during breaking of the
bluegill sunfish Lepomis macrochirus. J. Exp. Biol. 208,
4735–4746.
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London 9
Ontogeny of waterfall climbing in gobiid fishesR. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)

UNCORRECTED PROOF
Irschick, D.J., Vanhooydonck, B., Herrel, A. & Andronescu,
A. (2003). Effects of loading and size on maximum power
output and gait characteristics in geckos. J. Exp. Biol. 206,
3923–3934.
Jayne, B.C. & Lauder, G.V. (1994). How fish use slow and fast
muscle fibers: implications for models of vertebrate muscle
recruitment. J. Comp. Physiol. A 175, 123–131.
Julius, M.L., Blob, R.W. & Schoenfuss, H.L. (2005). The
survival of Sicyopterus stimpsoni, an endemic amphidro-
mous Hawaiian gobiid fish, relies on the hydrological
cycles of streams: evidence from changes in algal composi-
tion of diet through growth stages of fish. Aquat. Ecol. 39,
473–484.
Kramer, D.L. & McLoughlin, R.L. (2001). The behavioral
ecology of intermittent locomotion. Am. Zool. 41,
137–153.
Maie, T., Schoenfuss, H.L. & Blob, R.W. (In review). Onto-
genetic scaling of body proportions in waterfall-climbing
gobiid fishes from Hawai’i and Dominica: implications for
locomotor functionQ4 .
McHenry, M.J. & Lauder, G.V. (2005). Ontogeny of form
and function: locomotor morphology and drag in zebrafish
(Danio rerio). J. Morphol. 267, 1099–1109.
Nishimoto, R.T. & Kuamo’o, D.G.K. (1992). Waterfall
climbing behavior in the postlarvae of native Hawaiian
stream gobies. Technical paper, Division of Aquatic
Resources, State of Hawai’i.
Rome, L.C. (1998). Some advances in integrative muscle
physiology. Comp. Biochem. Physiol. B 120, 51–72.
Sanger, A.M. & Stoiber, W. (2001). Muscle fiber diversity and
plasticity. In Fish physiology, Vol. 18: 187–250. Johnston,
I.A. (Ed.). San Diego: Academic Press.
Schmidt-Nielsen, K. (1984). Scaling: why is animal size so
important? Cambridge: Cambridge University Press.
Schoenfuss, H.L. (1997). Metamorphosis of the Hawaiian
stream goby Sicyopterus stimpsoni: a structural, functional,
and behavioral analysis. PhD thesis, Louisiana State Uni-
versity.
Schoenfuss, H.L. & Blob, R.W. (2003). Kinematics of water-
fall climbing in Hawaiian freshwater fishes (Gobiidae):
vertical propulsion at the aquatic–terrestrial interface.
J. Zool. (Lond.) 261, 191–205.
Schoenfuss, H.L., Julius, M.L. & Blob, R.W. (2004). Coloni-
zation of a recent, volcanically formed freshwater habitat:
an example of primary succession. Ichthyol. Explor. Fresh-
waters 15, 83–90.
Toro, E., Herrel, A., Vanhooydonck, B. & Irschick, D.J.
(2003). A biomechanical analysis of intra- and interspecific
scaling of jumping and morphology in Caribbean Anolis
lizards. J. Exp. Biol. 206, 2641–2652.
Wainwright, P.C., Alfaro,M.E., Bolnick, D.I. &Hulsey, C.D.
(2005). Many-to-one mapping of form to function: a
general principle in organismal design? Int. Comp. Biol. 45,
256–262.
Walker, J.A. (1998). Estimating velocities and accelerations of
animal locomotion: a simulation experiment comparing
numerically different alogorithms. J. Exp. Biol. 201, 981–995.
Weinstein, R.B. & Full, R.J. (1998). Performance limits of
low-temperature, continuous locomotion are exceeded
when locomotion is intermittent in the ghost crab. Physiol.
Zool. 71, 274–284.
Weinstein, R.B. & Full, R.J. (1999). Intermittent locomotion
increases endurance in a gecko. Physiol. Biochem. Zool. 72,
732–739.
Journal of Zoology (2007) c� 2007 The Authors. Journal compilation c� 2007 The Zoological Society of London10
Ontogeny of waterfall climbing in gobiid fishes R. W. Blob et al.
JZO 315(BW
UK
JZ
O 3
15.P
DF
05-M
ar-0
7 19
:57
9547
12 B
ytes
10
PAG
ES
n op
erat
or=
Rav
isha
nkar
)
Author Query Form
_______________________________________________________
_______________________________________________________
Dear Author,
During the copy-editing of your paper, the following queries arose. Please respond to these by marking up your proofs with the necessary changes/additions. Please write your answers clearly on the query sheet if there is insufficient space on the page proofs. If returning the proof by fax do not write too close to the paper's edge. Please remember that illegible mark-ups may delay publication.
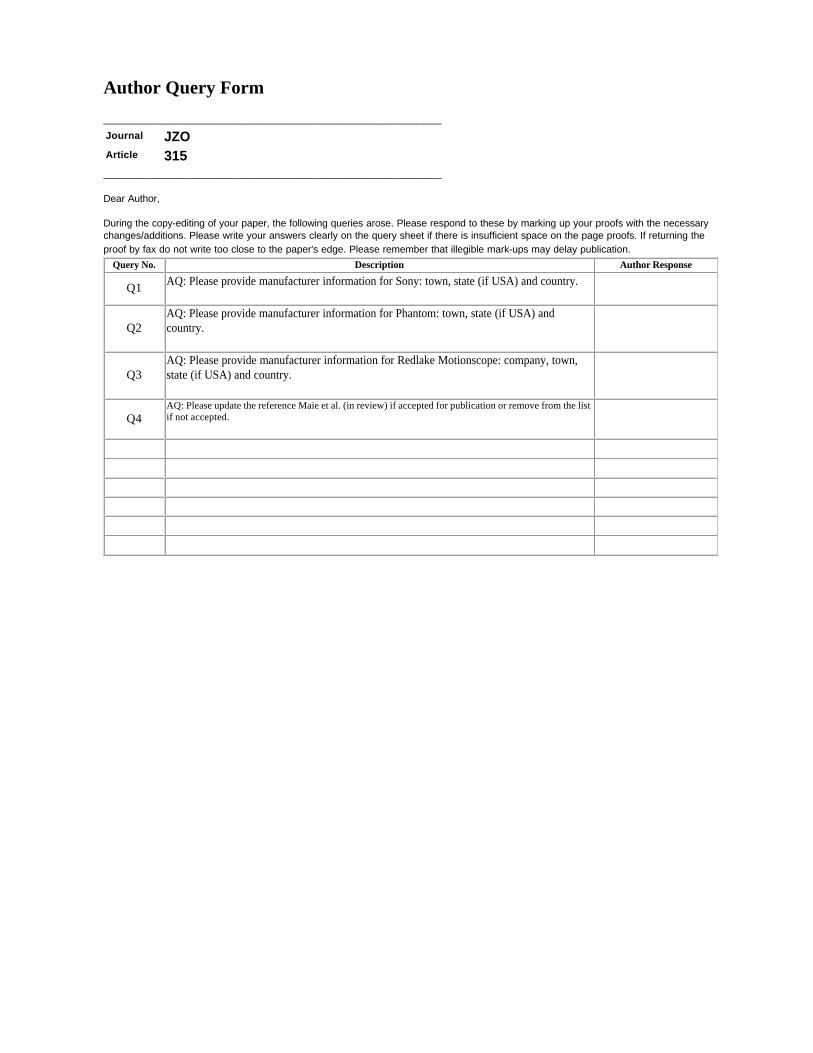
Journal JZOArticle 315
Query No. Description Author Response
Q1AQ: Please provide manufacturer information for Sony: town, state (if USA) and country.
Q2AQ: Please provide manufacturer information for Phantom: town, state (if USA) and country.
Q3AQ: Please provide manufacturer information for Redlake Motionscope: company, town, state (if USA) and country.
Q4AQ: Please update the reference Maie et al. (in review) if accepted for publication or remove from the list if not accepted.
MARKED PROOF
Please correct and return this set
Instruction to printer
Leave unchanged under matter to remain
through single character, rule or underline
New matter followed byor
or
or
or
or
or
or
or
or
and/or
and/or
e.g.
e.g.
under character
over character
new character new characters
through all characters to be deleted
through letter orthrough characters
under matter to be changedunder matter to be changedunder matter to be changedunder matter to be changedunder matter to be changed
Encircle matter to be changed
(As above)
(As above)
(As above)
(As above)
(As above)
(As above)
(As above)
(As above)
linking characters
through character orwhere required
between characters orwords affected
through character orwhere required
or
indicated in the marginDelete
Substitute character orsubstitute part of one ormore word(s)
Change to italicsChange to capitalsChange to small capitalsChange to bold typeChange to bold italicChange to lower case
Change italic to upright type
Change bold to non-bold type
Insert ‘superior’ character
Insert ‘inferior’ character
Insert full stop
Insert comma
Insert single quotation marks
Insert double quotation marks
Insert hyphenStart new paragraph
No new paragraph
Transpose
Close up
Insert or substitute spacebetween characters or words
Reduce space betweencharacters or words
Insert in text the matter
Textual mark Marginal mark
Please use the proof correction marks shown below for all alterations and corrections. If you
in dark ink and are made well within the page margins.wish to return your proof by fax you should ensure that all amendments are written clearly





























