Embed Size (px)
Citation preview

Molecular and Cellular Endocrinology, 74 (1990) R7-RlO
Elsevier Scientific Publishers Ireland, Ltd.
R7
MOLCEL 02421
Rapid Paper
Ontogeny of 3P-hydroxysteroid dehydrogenase/A5-A4 isomerase (3P-HSD) in human adrenal gland performed by immunocytochemistry
E. DuPont, V. Luu-The, F. Labrie and G. Pelletier MRC Group in Molecular Endocrinology, CHUL Research Center and Lava1 University, Quebec GI V 4G2, Canada
(Received 15 October 1990; accepted 16 October 1990)
Key words: 3B-Hydroxysteroid dehydrogenase/A’-A4 isomerase; Adrenal; Ontogeny; Immunocytochemistry; (Human)
Summary
Human foetal adrenal plays a major role in the synthesis and secretion of steroids which are essential to several physiological events during foetal development. Since 3P-hydroxysteroid dehydrogenase 5-ene-4-ene isomerase (3b-HSD) is involved in the biosynthesis of all classes of hormonal steroids, namely pro-
gesterone, glucocorticoids, mineralocorticoids, androgens and estrogens, knowledge of the ontogeny of 3fi-HSD should lead to a better understanding of steroidogenesis in the foetal adrenal. Polyclonal
antibodies raised against purified human placental 3/?-HSD were used to localize the enzyme in 22-week, 28-week, and term foetal adrenals as well as in the adrenals of 2-month-, g-month- and 2-year-old infants.
3/3-HSD has been identified in the outer cortex, neocortex and zona glomerulosa zone of foetal adrenal, respectively. In 2-month- and 8-month-old infant adrenals, the enzyme was localized in the zona glomerulosa and zone fasciculata while in 2-year-old infants, the immunoreaction was observed also in the zona reticularis, the medulla being devoid of any reaction.
Introduction
The enzyme 3&hydroxy-5-ene-steroid dehydro- genase/A5-A4 isomerase (3/3-HSD) is responsible for the conversion of all the A5-3/%hydroxysteroids
into A4-3-ketosteroids. This enzymatic step is thus essential for the biosynthesis of all biologically
active steroid hormones, namely glucocorticoids, mineralocorticoids, progesterone, estrogens and androgens (Lipsett, 1986).
The 3/3-HSD enzymatic system is present not only in the classical steroidogenic tissues, namely the adrenals, testes, ovaries and placenta (Ishii- Ohba et al., 1987; Luu-The et al., 1989; Zhao et
al., 1989) but also in many peripheral tissues in- eluding the prostate, breast, brain, liver and skin
(Lax and Schriefers, 1981; Lacoste et al., 1990). The human foetal adrenal cortex is morphologi-
tally and functionally differentiated into two
zones, namely the neocortex and the foetal zone. While the foetal zone primarily secretes A5-steroid 3-sulfates (Seron-Ferre et al., 1978; Simonian and Gill, 1981), the neo-cortex secretes higher amounts of A4-3-ketosteroids and is the primary site of cortisol production in the foetus (Seron-Ferre et al., 1978). Circulating levels of free and con-
Address for correspondence: Dr. Georges Ph.D., MRC Group in Molecular Endocrinol
search Center, 2705, Laurier Boulevard, Ste-FI _ _
4G2, Canada. jugated A5-3/3-hydroxysteroids such as dehydro-
_ Pelletier, M.D.,
ogy, CHUL Re-
DY, Quebec GlV
0303-7207/90/$03.50 0 1990 Elsevier Scientific Publishers Ireland, Ltd

R8
epiandrosterone (DHEA), pregnenolone (PREG) and 17-hydroxypregnolone (17-OH-PREG) un- dergo dramatic changes with a rapid decline after birth, a gradual rise during late childhood and puberty, and a smaller decline in old age (De Peretti and Forest, 1978; De Peretti and Mappas, 1983; Orentreich et al., 1984). Analyses of in- traadrenal (Dickerman et al., 1984) circulating and urinary (Kelner and Brook, 1983) A4- to A5-
steroid ratios during development have suggested that these phenomena reflect, at least in part, changes in the activity of the adrenal microsomal
3P-HSD complex. In order to obtain more infor- mation about the age-specific expression of 3p- HSD in the human adrenal, we have localized this
enzyme by immunocytochemistry at the light mi- croscopic level during foetal and postnatal periods of development in the human adrenal.
Materials and methods
Adrenal gland. Foetal adrenal tissues were ob- tained at autopsy following premature termination of pregnancy during the second (22 weeks) and third (28 weeks) trimester of gestation as well as at term-gestation (35 weeks). Postnatal human
adrenal tissue was obtained at autopsy from pa- tients (aged of 2 and 8 months and 2 and 20 years) deceased from non-endocrine causes. For each time interval, at least three specimens were
analyzed. Histological techniques. The adrenals were
fixed by immersion in Bouin’s fluid for 3 days. The tissues were then dehydrated in ethanol and embedded in paraffin. 6 urn sections were cut and mounted onto glass slides.
Immunohistochemistry. The sections were in- cubated overnight at 4°C with the 3/3-HSD anti- serum (Luu-The et al., 1989) raised in rabbits against purified human placenta 3&HSD (Luu- The et al., 1989). The antiserum was diluted 1 : 500 in Tris-saline (pH 7.6). The sections were then washed in the same buffer and incubated at room temperature for 4 h with peroxidase-labeled goat antirabbit y-globulins (Hyclone, Logan, Utah, U.S.A.) diluted 1 : 500. The same antibody has recently been used to screen a hgtll cDNA library which led to first elucidation of the nucleotide sequence of human 3b-HSD cDNA and the pre-
dieted amino acid sequence of the human enzyme (Luu-The et al., 1989). The antiserum has also been successfully used for the localization of 3/% HSD in the human placenta as well as in guinea pig and rat gonads and adrenals (DuPont et al., 1990a, b). Control experiments were performed on adjacent sections by substituting preimmunized rabbit serum or the primary antiserum immuno-
absorbed with excess (lop6 M) purified human placental 3P-HSD (Luu-The et al., 1989).
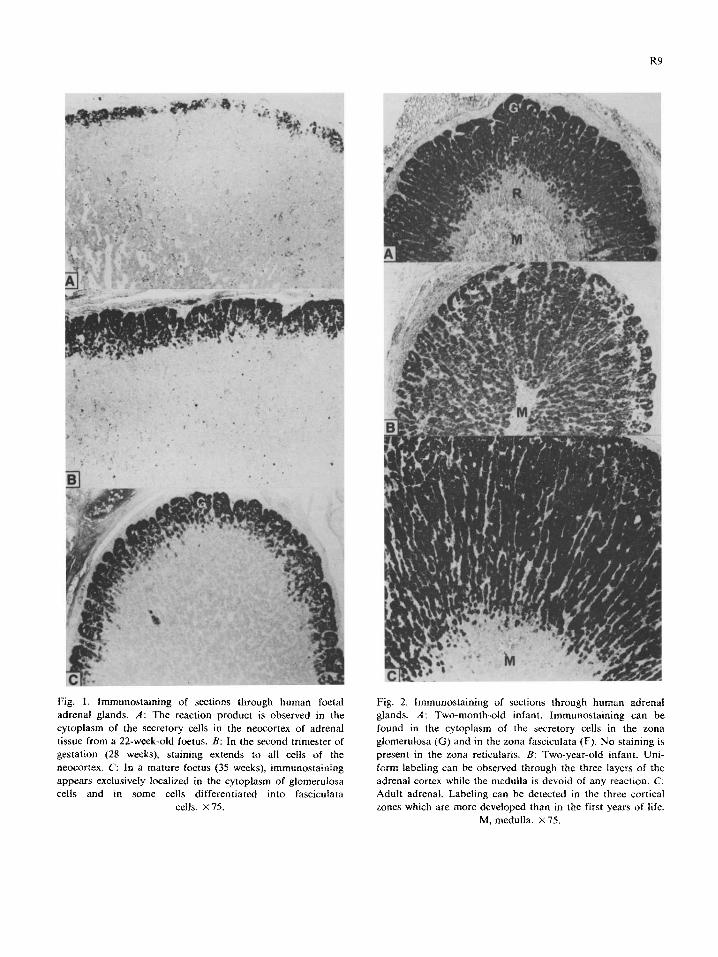
In the foetal adrenal from mid-gestation (22 weeks), immunostaining was observed exclusively in the cytoplasm of cells of the outer cortex (Fig. 1A). Six weeks later, at the beginning of the third trimester (28 weeks) (Fig. lB), immunolabeling was more widely distributed and extended to all cells of the neocortex. In term foetal adrenals, on the other hand, strong staining was observed in the presumptive zona glomerulosa (Fig. 1C).
In 2-month-old infant adrenals (Fig. 2A), im-
munostaining was seen in both the zona glomer- ulosa and the zona fasciculata. No staining could be detected in the zona reticularis which could be easily identified. Similar 3/?-HSD localization was
seen in the &month adrenals. In 2-year-old infants and adults (Fig. 2B and C), on the other hand, immunostaining was observed in the three layers of the cortex, the medulla being devoid of any reaction. Moreover, the intensity of staining was similar in the zona reticularis, fasciculata and glomerulosa. At the subcellular level, the labeling was exclusively observed in the cytoplasm.
Discussion
The foetal adrenal gland derives from the dor- sal coelomic epithelium. At 8 weeks of gestation, the human adrenal comprises chromaffin cells which will later form the adrenal medulla as well as the cortex which is divided into two zones, namely the outer or definitive zone and the inner or foetal zone. The outer zone further differenti- ates into zona glomerulosa, fasciculata and reti- cularis during the period of infancy and early
R9
Fig. 1. Jmmunostain~ng of sections through human foetal
adrenal glands. A: The reaction product is observed in the
cytoplasm of the secretory cells in the neocortex of adrenal tissue from a 22-week-old foetus. B: In the second trimester of
gestation (28 weeks), staining extends to all cells of the
neocortex. C: In a mature foetus (35 weeks), ~mmunostaining
appears exclusively localized in the cytoplasm of glomerulosa cells and in some cells differentiated into fasciculata
cells. X 75.
Fig. 2. lmmunostaining of sections through human adrenal glands. A: Two-month-old infant. Immunostaining can be
found in the cytoplasm of the secretory cells in the zona
glomerulosa (G) and in the zona fasciculata (F). No staining is
present in the zona reticularis. B: Two-year-old infant. Uni-
form labeling can be observed through the three layers of the
adrenal cortex while the medulla is devoid of any reaction. C:
Adult adrenal. Labeling can be detected in the three cortical
zones which are more developed than in the first years of life.
M. medulla. x 75.

R10
childhood, while the inner zone which occupies approximately 80% of the adrenal volume during foetal life regresses following birth.
Contrary to the observation of Murakoshi et al. (1983) who localized 3P-HSD in the inner zone of the human 20-week foetal adrenal by histochem- istry, our data clearly identify 3P-HSD exclusively
in the outer zone at this period of foetal life. During early to mid-gestation, it is generally thought that 3/3-HSD activity in foetal adrenal
tissue is minimal (1983) and must use placental progesterone as a precursor for the synthesis of cortisol. At mid-gestation and approaching term, the present data indicate an increase in 3/?-HSD levels (Fig. 1B and C), a finding which could well
explain the increase in circulating cortisol at mid- gestation (Migeon et al., 1957). Thus, the increase in 3P-HSD level in the outer zone of the adrenal cortex could provide the basis for increased corti- sol secretion by the foetal adrenal independently from placental transfer of cortisol (Beitins et al., 1973; Murphy, 1982). Such data also suggest that the foetal adrenal is actively involved in pro-
gesterone synthesis contrary to the generally accepted view that progesterone is synthesized from precursor pregnenolone by the placental 3/L
HSD. In fact, 3P-HSD activity is only 3-fold higher in the outer adult-type definitive adrenal tissue compared with that in the foetal cortex (Simson and Capp, 1984). Moreover, removal of the foetus in baboons resulted in a 80% decline in serum estradiol concentration and a 70% decrease in
serum progesterone (Albrecht et al., 1980). The decrease in circulating estrogen presumably re- flects removal of estrogen precursors secreted by the foetus while the loss in circulating pro- gesterone might be due, at least in part, to the removal of foetal 3P-HSD or to factors of foetal
origin which regulate placental progesterone pro- duction (Hagemenas et al., 1975).
References
Albrecht, E.D., Haskins, A.L. and Pepe, G.J. (1980) Endo-
crinology 107, 766-770.
Beitins, I.Z., Bayard, F., Antes, I.G., Kowanski, A. and Mi-
geon, C.J. (1973) Pediatr. Res. 7, 509.
De Peretti, E. and Forest, M.G. (1978) J. Clin. Endocrinol.
Metab. 47, 572-571.
De Peretti, E. and Mappas, E. (1983) J. Clin. Endocrinol.
Metab. 51, 550-556.
Dickerman, Z., Grant, D.R., Faiman, C. and Winter, J.S.D.
(1984) J. Clin. Endocrinol. Metab. 59, 1031-1036.
DuPont, E., Luu-The, V., Labrie, F. and Pelletier, G. (1990a)
Endocrinology 126, 2906-1909.
DuPont, E., Zhao, H.F., Rheaume, E., Simard, J., Luu-The, V.,
Labrie, F. and Pelletier, G. (1990b) Endocrinology 127,
1394-1403.
Hagemenas, F.L., Baughman, W.L. and Kittinger, G.W. (1975)
Endocrinology 96, 1059-1062.
Ishii-Ohba, H., Inano, H. and Tamaoki, B.I. (1987) J. Steroid
Biochem. 21, 7155779.
Kelnar, C.J.H. and Brook. C.G.D. (1983) Clin. Endocrinol.
(Oxford) 19, 117-129.
Lacoste, D., Belanger, A. and Labrie, F. (1990) in Steroid
Formation, Degradation and Action in Peripheral Tissues
(Bradlow, H., Castagnatta, L., d’Aquino, S. and Labrie, F.,
eds.), Ann. N.Y. Acad Sci. 595, 389-391.
Lax, E.R. and Schriefers, H. (1981) Acta Endocrinol. 98,
261-266.
Lipsett, M.B. (1986) in Reproductive Endocrinology (Yen,
S.S.C. and Jaffe, R.B., eds.), pp. 140-153, Saunders, Phila-
delphia, PA.
Luu-The, V., Lachance, Y., Labrie, C., Leblanc, G., Thomas,
J.L., Strickler, R.C. and Labrie, C. (1989) Mol. Endocrinol.
3, 1310-1312.
Migeon, C.J., Bertrand, J. and Wale, P.E. (1957) J. Clin. Invest.
36, 1350-1362.
Murakoshi, M., Osamura, Y., Watanabe, K. and Kuroshima,
V. (1983) Tokai J. Exp. Clin. Med. 8, 89-96.
Murphy, B.E.P. (1982) Am. J. Obstet. Gynecol. 144, 276-282.
Orentreich, N., Brind, J.L., Rizer, R.L. and Vogelman, J.H.
(1984) J. Clin. Endocrinol. Metab. 59, 551-555.
Seron-Ferre, M., Lawrence, CC., Siiteri, P.K. and Jaffe, R.B.
(1978) J. Clin. Endocrinol. Metab. 47, 603-609.
Simonian, M.H. and Gill, G.N. (1981) Endocrinology 108,
1769-1779.
Simson, M.H. and Capp, M.W. (1984) J. Clin. Endocrinol.
Metab. 59, 643-651.
Zhao, H.F., Simard, J., Labrie, C., Breton, N., Rheaume. E.,
Luu-The, V. and Labrie, F. (1989) FEBS Lett. 259.153-157.





























