Embed Size (px)
Citation preview

�������� ����� ��
Ontogeny of the digestive enzyme activities in hatchery produced beluga(Huso huso)
Reza Asgari, Gholamreza Rafiee, Soheil Eagdei, Farzaneh Noori, NaserAgh, Hadi Pourbagher, Enric Gisbert
PII: S0044-8486(13)00412-2DOI: doi: 10.1016/j.aquaculture.2013.08.014Reference: AQUA 630792
To appear in: Aquaculture
Received date: 18 June 2013Revised date: 6 August 2013Accepted date: 15 August 2013
Please cite this article as: Asgari, Reza, Rafiee, Gholamreza, Eagdei, Soheil, Noori,Farzaneh, Agh, Naser, Pourbagher, Hadi, Gisbert, Enric, Ontogeny of the digestiveenzyme activities in hatchery produced beluga (Huso huso), Aquaculture (2013), doi:10.1016/j.aquaculture.2013.08.014
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Ontogeny of the digestive enzyme activities in hatchery
produced beluga (Huso huso)
Reza Asgari1, Gholamreza Rafiee
1, Soheil Eagdei
1, Farzaneh Noori
2, Naser
Agh2, Hadi Pourbagher
1, Enric Gisbert
3
1 Department of Fisheries, Faculty of Natural Resources, University of Tehran, 31585-4314
Karaj, Iran; 2 Artemia & Aquatic Animals Research Institute, Urmia University, Urmia, Iran;
3 Institut de Recerca i Tecnologia Agroalimentaries (IRTA),Centre de Sant Carles de la
Ràpita, Crta. Poble Nou del Delta km 5.5, 43540 - Sant Carles de la Ràpita, Spain
Correspondence: Reza Asgari, Department of Fisheries, Faculty of Natural Resources,
University of Tehran, 31585-4314 Karaj, Iran, Tel: 0098- 32223044. E-mail:

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Abstract
The development of digestive enzymes of the stomach (pepsin), pancreas (trypsin,
chymotrypsin, amylase, and lipase) and intestine (alkaline phosphatase) were studied in
beluga (Huso huso) from hatching time to 50th day post hatching. Sampling was carried out
immediately after hatching, 111.3, 235.2, 306, 400.2, 490, 612.5, 756 and 906.5 degree day
post hatching, ddph (0, 7, 14, 18, 23, 28, 35, 42 and 50 days post hatching, dph, respectively)
for the digestive enzymes activity. Beluga larvae had near 70% mortality during rearing
procedure. The highest mortality rate was accrued in two stages; firstly, during endogenous
feeding (2-9 dph; 30.4 - 144.9 ddph) and then from mixed feeding stage (12 dph; 253.5 ddph)
until starting time of weaning (28 dph; 490 ddph), with values ranging from 50 to 40% of
total mortality, respectively. Furthermore, the results showed that at the onset of exogenous
feeding gastric glands were already functional as indicated by the steady increase in pepsin
specific activity. In contrast, trypsin and chymotrypsin significantly decreased after the onset
of exogenous feeding, showing the importance of these types of enzymes in the cleavage of
yolk proteins during the endogenous feeding phase. Moreover, amylolytic activity fluctuated
considerably over the study period; after onset of exogenous feeding alpha-amylase activity
increased (P < 0.05), whereas it reached a maximum value at the end of the study (P<0.05).
Lipolytic activity was low in the first week (P < 0.05), whereas it increased until onset of
exogenous feeding and then, until 490 ddph (28 dph) went down again and reached the same
amount in the first week. At the end of the study, lipolytic activity reached 4 times relative to
the activity levels recorded during the first 4 weeks of exogenous feeding. Furthermore,
changes in enzyme activities from the stomach and pancreas were coupled with those in the
intestine (brush border membrane), whereas the increase in alkaline phosphatase from
hatching until 111.3 ddph (7 dph) suggested that the beluga larvae had a developed intestine
at the onset of exogenous feeding. The results obtained from this study proposed that more

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
beluga weaned earlier, more larvae lived and hence weaning should begin immediately after
commencement of exogenous feeding.
Keywords: beluga (Huso huso), digestive enzymes, weaning, mixed feeding, larvae
1. Introduction
Sturgeon fish are commercially cultured for caviar production, as a result of the sharp
decrease in fisheries of this group of fishes. Sturgeons are a group of slow-growing fish that
mature very late in life, and consequently, they are particularly vulnerable to overfishing and
to other threats, like the loss of their natural habitat, river damming, and deterioration of
water quality (May et al., 1997). As a consequence, wild stocks of sturgeons have been
decreasing dramatically during these last decades (Billard and Lecointre, 2001) and sturgeons
are considered as one of the most threatened group of animals on the IUCN Red List of
Threatened Species™ (IUCN, 2013).
The beluga, Huso huso, is an important commercial species in the Caspian Sea and a
good candidate for aquaculture because of its fast growth and easy propagation in captivity.
In addition, this is the most appreciated species in terms of caviar quality. Consequently, this
species has been cultured for commercial and restocking purposes in Iran since 1991 (IFO,
2002). Based on fishery data and number of recorded spawning individuals, it has been
estimated that the wild native populations of this species have declined over 90% in the past
three generations (ca. 60 years), and overfishing for meat and caviar will soon cause global
extinction of the remaining natural wild populations if conservation measures are not taken in
its area of distribution (Gessner et al., 2010). Independently of the final purpose of this

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
activity, conservation and restocking programs or commercial purposes, a reliable larval
rearing technique must be developed to ensure consistent production of good quality fry.
However, the high mortality rate during beluga larval rearing, up to 80% in some cases, has
been identified as one of the most serious bottlenecks for the culture of this species.
Therefore, the success and development of aquaculture activity of this critically endangered
species still requires some improvements in rearing techniques, especially those affecting
larval stages such as the partial or complete replacement of live prey with a compound diet
and the formulation of species-specific weaning protocols and diets. Traditionally, hatchery-
produced sturgeon larvae were raised on live prey, e.g., oligochaetes (Enchytraeus sp. and
Tubifex sp.), cladocerans (Daphnia sp. and Moina sp.) or Artemia sp. (see review in Gisbert
and Williot, 2002). However, the production of live food is a labor-intensive and expensive
process, since their production and enrichment require of considerable space, manpower and
labor. Moreover, the nutritional supplies of live food are often inadequate to complete the
growth-out phase and variable along the production cycle. From a nutritional point of view,
newly hatched larvae remain absolutely dependent on yolk sac reserves. When yolk is
depleted, larvae must begin to ingest external food to cope with increasing energy demands
for growth. In this context, larvae of different Acipenserid species possess an anatomically
complete and functional digestive system with a high degree of morphological organization
and functionality at the onset of exogenous feeding (Buddington and Doroshov, 1986;
Dettlaff et al., 1993; Gawlicka et al., 1995). During the last twenty years, several studies have
demonstrated that compound larval diets can be successfully used for intensive commercial
culture of several sturgeon species from the onset of exogenous feeding (e.g., Acipenser
baerii, A. transmontanus, A. oxyrinchus and A. medirostris) (Bardi et al., 1998; Buddington
and Doroshov, 1984; Dabrowski et al., 1985; Gisbert and Doroshov, 2006).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
The efficiency of food depends on the physiological capacities in fish to digest and
transform ingested nutrients (Furné et al., 2008). The analysis of digestive enzyme activities
is an easy and reliable biochemical method that can provide insight into the digestive
physiology of fish larvae (Rønnestad et al., 2013), their nutritional condition (Bolasina et al.,
2006) and assist in defining their nutritional requirements (Twining et al., 1983). In addition,
a comprehensive analysis of the ontogenic changes during the early life stages of fish is
essential for the proper design of feeding strategies and formulation of compound diets
(Verreth and Segner, 1995). Thus, in order to enhance the success of larval rearing of this
sturgeon species and to facilitate overcoming one of the major bottlenecks of beluga
hatcheries, the description of the ontogeny of the larval digestive enzymes is a necessary tool.
Although the ontogenetic development of the digestive enzymes has been described in a large
number of marine and freshwater fish species (see reviews in Lazo et al., 2011; Zambonino-
Infante et al., 2009), there is very limited information on this group of primitive fish species
with fragmentary information from A. transmontanus (Buddington and Doroshov, 1986;
Gawlicka et al., 1995), A. baerii (Gisbert et al., 1999; Zółtowska et al., 1999), A. nacarii
(Furné et al., 2005; Sanz et al., 2011) and the hybrid bester (Timeiko and Bondarenko, 1988).
No data are available on the ontogenetic development of digestive enzymes in beluga.
The aim of this study was to characterize the development of the main digestive
enzymes from the stomach (pepsin), pancreas (trypsin, chymotrypsin, α-amylase and lipase)
and intestine (alkaline phosphatase) in beluga in order to provide insight into the digestive
physiology of this species. This information will be useful for the development of a
formulated compound diet and establishing the most convenient weaning time for this
sturgeon species, and synchronize the state of development of the organism, with regards to
its digestive capacity, with the larval rearing techniques.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
2. Materials and Methods
2.1. Larval supply and rearing protocol
Beluga specimens used in the present study were obtained by the hormonally-induced
(LHRHa2) spawning of 2 females that were fertilized with the milt of 3 males at the Shahid
Marjani Sturgeon Center (Gorgan, Iran). Eggs were incubated in Yushchenko incubators at
11-12ºC in a closed freshwater recirculation system. After eight days of incubation, hatching
took place (hatching rate: 68%) and 1800 newly hatched larvae (body weight, BW: 15.6 ± 0.7
mg; total length, TL: 11.2 ± 0.4 mm) were placed in three 500 liter circular fiberglass tanks
connected to a flow-through freshwater system. Malformation rate at hatching (11.2 mm TL)
was 6.1%. During the larval rearing period, water temperature, dissolved oxygen, pH and
flow rate were 16.5 ± 0.2ºC, 10.7 ± 0.3 mg/l, 7.8 ± 0.1 and 5.7 ± 0.4 l/min, respectively.
Larvae were reared under natural photoperiod (37º4'30.62'' N, 54º40'10.45'' E). All tank
surfaces were scrubbed and the bottom of the tank siphoned to remove uneaten feed and feces
3 times a day.
The feeding protocol used for H. huso larval rearing from hatching to the juvenile
stage is summarized in Figure 1. In brief, larvae were fed by mixed of Artemia nauplii (EG,
INVE, Belgium) and Daphnia sp. from 12 to 25 dph (199.2- 432.5 degree days post hatch,
ddph) (500 - 800 nauplii/larvae/day). A short co-feeding phase based on Cladocerans and a
compound diet (Biomar, Denmark; D1- particle size = 0.5 mm) was conducted from 25 to 30
dph (432.5- 534 ddph), whereas from this age onward, fish were fed only with the inert feed
(Biomar, Denmark; D2- particle size = 0.8 mm) at a feeding rate of 30 and 20% of stocked
fish biomass respectively. The compound diet was given to larvae between 4 to 6 times per
day and feed ration and particle size were progressively adjusted for fish size from 25 to 50
dph (432.5- 906.5 ddph).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Proximate composition (dry matter) of live prey (Artemia nauplii and Daphnia sp.)
was determined in 3 samples per type of live prey. Crude protein was determined by the
Kjeldahl method (N × 6.25) using an automatic Kjeldahl system (Behrotest WD 40,
Germany) and crude fat content was determined by Soxhlet method (Folch et al., 1957). Total
protein and lipid content in Artemia were 65.6 ± 1.4% and 15.1 ± 0.2%, respectively, whereas
Daphnia sp. contained 52.6 ± 1.9% of proteins and 11.1 ± 0.1% of lipids. The proximate
composition of compound diets (D1 and D2) was 63 and 58% of total protein, 11 and 15% of
total lipids, and 11.6 and 11.5% total carbohydrate respectively (data provided by feed
manufacturer).
2.2. Fish growth measurements and sampling schedule
In order to determine larval growth in weight (BW) and total length (TL), fish (n =
40) were daily sampled randomly prior to feeding in the morning using a pipette with a large
opening from hatching to 18 dph (306 ddph), whereas samples were collected every 2 days
from 18 to 50 dph (306 - 906.5 ddph). Sampled larvae were sacrificed with an overdose of
tricaine methanosulphonate (MS-222, Sigma-Aldrich, Munich, Germany), rinsed in distilled
water, and their TL and BW measured to the nearest 0.01 mm and 0.1 g, respectively. Fish
TL was measured by means of a stereomicroscope equipped with a drawing tube and
micrometer (Zeizz, Oberkochen, Germany) and BW was determined with an analytical
microbalance (Mettler Toledo, XS 205, Switzerland). Furthermore, dead larvae were
removed and counted 2–3 times per day. From this data, cumulative mortality was calculated.
Larval malformation rate at hatching was 6.1%.
Samples (n = 100) for analysis of the digestive enzymes activities were taken at
hatching, 7, 14, 18, 23, 28, 35, 42 and 50 dph (0, 111.3, 235.2, 306, 400.2, 490, 612.5, 756

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
and 906.5 ddph respectively). No food was added to the rearing tank the night prior to
sampling, and larvae were sampled in the early morning to minimize the effects of exogenous
enzymes from live food in fish guts (Kolkovski, 2001) using a pipette with a large opening.
Sampled fish were washed in distilled water, flash-frozen in liquid nitrogen and stored at
−80°C until further analyses.
2.3. Sample preparation and enzymes determination
From hatching till 18 dph (306 ddph) the whole body of each larva and from this age
onward, complete digestive system with the exception of intestine for pancreatic and gastric
enzymes and separated intestine for brush border enzyme were dissected over a cold (0- 4ºC)
glass plate according to Cahu and Zambonino-Infante (1994). Then, all samples except for
intestine were homogenized (1: 9, w: v) in a buffer (100 mM Tris–HCl, 0.1 mM EDTA, 0.1%
triton X-100, pH= 7.8) with an electric homogenizer (Heidolph Instruments, Germany).
Tissue homogenates were centrifuged at 30,000 g for 30 min at 4oC (Centrikon H- 401
centrifuge, Kontron) and the supernatant was collected and frozen at -80oC until its analysis.
In addition, enterocytes’ brush border extracts were prepared as described by Crane et al.
(1979). In brief, the intestinal regions of the digestive tract were homogenized in 30 v/w
fractions of Tris (2 mM)–mannitol (50 mM), pH 7.0 for 30 s with an electric homogenizer
(Heidolph Instruments, Germany). Then, tissue homogenates were centrifuged at 9000 g for
10 min after the addition of 0.1 M CaCl2. The supernatants were transferred to new vials and
stored frozen (−80 °C) until analysis of enzyme activity or protein content.
Pepsin (E.C.3.4.23.1) activity quantification was conducted according to
Rungruangsak and Utne (1981) using casein as substrate. In brief, 200 µl of the crude enzyme

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
extract were mixed with 200 µl of substrate (1% cazein solution in 60 mM HCl) and
incubated for 10 min at 37°C. The reaction was stopped with 1 ml of 5% trichloroacetic acid,
and after 30 min, tubes were centrifuged at 5,000 g for 20 min at 4°C. Afterwards, 0.5 µl of
the supernatant were incubated with 0.5 M NaOH (1 ml) and Folin-Ciocalteu reagent (0.3 ml)
for 10 min at 25°C and the absorbance of the supernatant measured at 720 nm. The range of
tyrosine standard curve was 0-50 µg tyrosine ml-1
. One unit of pepsin was defined as the μg
of tyrosine released per minute and ml at 37°C.
Trypsin (E.C.3.4.21.4) activity was measured with N-α-benzoyldlarginine-p-
nitroanilide (BAPNA) as substrate. BAPNA (1 mM in 50 mM Tris–HCl, 20 mM CaCl2, pH
7.5) was incubated with the enzyme extract at 37°C. Changes in absorbance were read at 410
nm for 10 min (Erlanger et al., 1961). Chymotrypsin (EC. 3.4.21.1) activity was measured by
using 0.1 mM Suc-Ala-Ala-Pro-Phe-p-nitroanilide (SAPNA) in 50 mM Tris–HCl and 20 mM
CaCl2 (pH = 7.5). The crude enzyme extract was incubated with substrate at 37°C and
changes in absorbance (410 nm) recorded for 3 min (Erlanger et al., 1961). Trypsin and
chymotrypsin activity units were expressed as change in absorbance per minute per mg
soluble protein and calculated by the following equation:
Alpha-amylase (E.C.3.2.1.1) activity was determined according to Worthington
(1991) using starch as substrate. In brief, starch (1%) was diluted in a 0.02 M Na2HPO4 and
0.006 M NaCl buffer (pH= 6.9) and incubated with the enzyme crude extract for 4 min at 25
°C. Then, 0.5 ml of 1% dinitrosalicylic acid (DNS) solution was added and boiled for 5 min.
After boiling, 5 ml of distilled water was added to the mixture and the absorbance of the
solution measured at 540 nm. Blanks were similarly prepared, but without the crude enzyme

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
extracts. Maltose (0.3–5 µmol ml -1
) was used for the preparation of the standard curve. The
α-amylase specific activity was defined by the μmol of maltose produced per min per mg
protein at 25ºC.
Lipase (E.C.3.1.1) activity was determined using nitrophenyl myristate as substrate.
Each assay (0.5 ml) contained 0.53 mM p-nitrophenyl myristate, 0.25 mM 2-methoxyethanol,
5 mM sodium cholate and 0.25 M Tris–HCl (pH= 9.0). Samples were incubated for 15 min at
30°C, and the reaction was stopped by the addition of 0.7 ml of acetone/n-heptane (5:2, v/v).
The reaction mixture was vigorously mixed and centrifuged at 6,080 g for 2 min and the
absorbance of the aqueous solution read at 405 nm. One unit of lipase was defined as 1 μmol
of n-nitrophenol released per min (Iijima et al., 1998).
Alkaline phosphatase (E.C.3.1.3.1) from purified enterocytes’ brush borders was
quantified at 37°C using 4-nitrophenyl phosphate (PNPP) as substrate in 30 mM Na2CO3
buffer (pH= 9.8). One unit (U) was defined as 1 μg BTEE released per min per ml of brush
border homogenate at 407 nm (Bessey et al., 1946).
Total soluble protein was measured by the Bradford (1976) method using bovine
serum albumin as a standard. Enzyme activities were expressed as specific activity (U
mg/protein). All samples were analysed in triplicate (biological replicates) and each of them
in triplicate (methodological replicate).
2.4. Statistical analysis
Data were checked for normality (Kolmogorov– Smirnov test) and homogeneity of
variances (Bartlett's test) prior to their comparison. All data are expressed as the mean ± SD

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
(n= 3). Digestive enzyme activities among different sampling times (ddph) were compared by
means of a one-way ANOVA, and the mean comparison was performed with a Duncan's test
at a reliability level of 5% (SPSS version 15; IBM, Somers, New York, USA).
3. Results
3.1. Fish growth and mortality rate
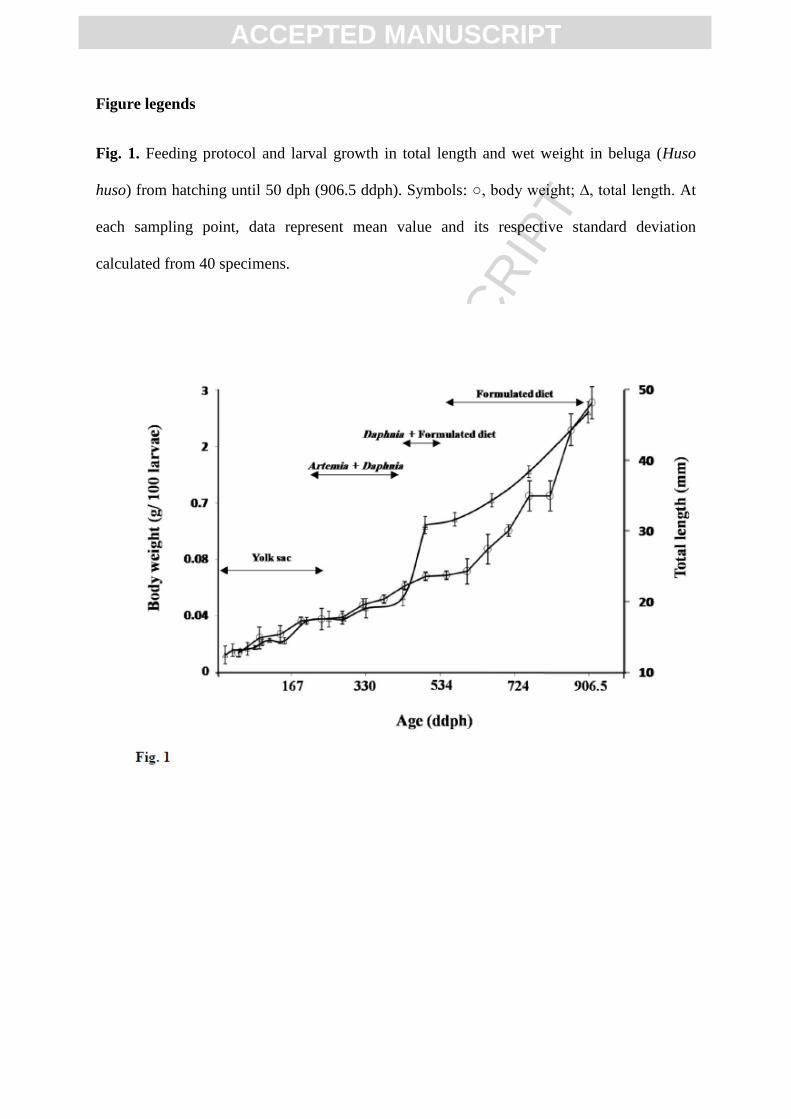
Total length and wet weight of beluga larvae averaged 12.1 ±1.01 mm and 11.0 ± 0.5
mg , respectively at hatching, and increased to 47.5 ± 6.5 mm and 2.5 ± 0.8 g at 906.5 ddph
(50 dph) (Fig. 1). Furthermore, at the end of the study, cumulative mortality was 69.9 ± 1.2
%. From 30.4 ddph (2 dph) to 144.9 ddph (9 dph), mortality sharply increased (50% of total
mortality), whereas it remained stable until the onset of the mixed feeding stage (12 dph;
199.2 ddph). From this stage until the onset of weaning (28 dph; 490 ddph), mortality
progressively increased (40% of total mortality), whereas after weaning almost no mortality
was observed in the rearing tanks (Fig. 2).
3.2. Digestive enzymes activities
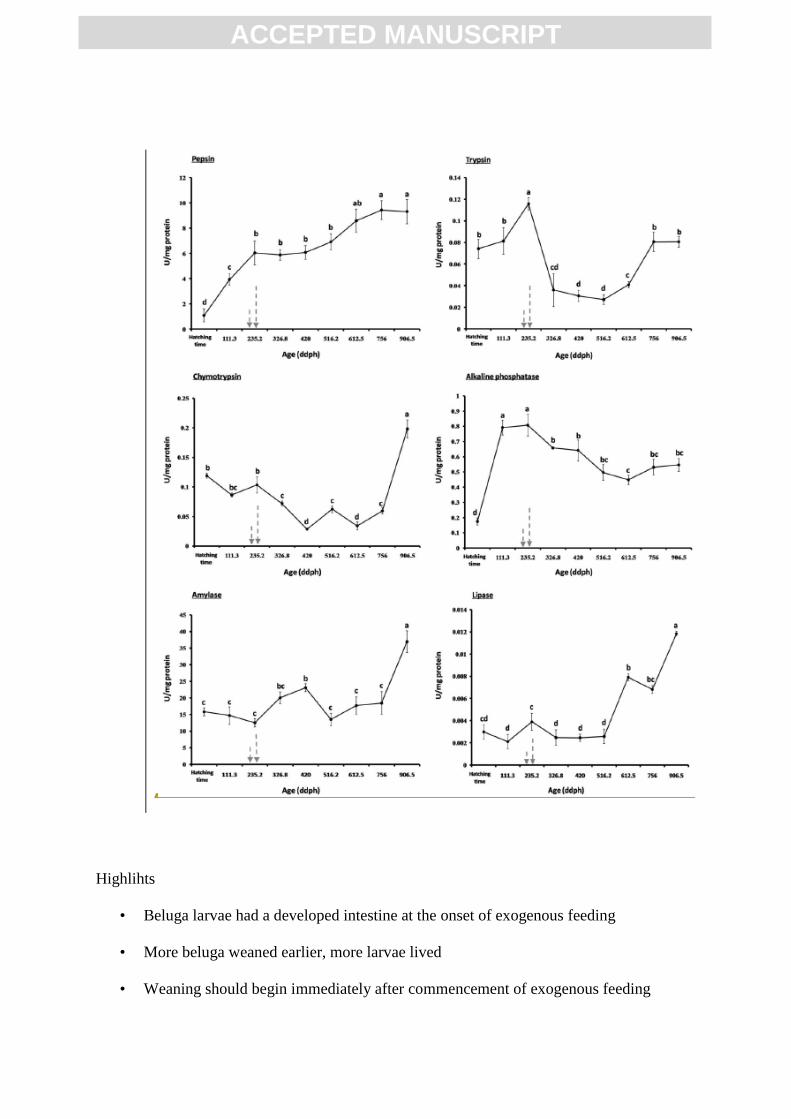
At hatching, pepsin activity was already detected in beluga larvae (1.08 ± 0.52 U mg−1
protein). From this stage until the onset of exogenous feeding at 14 dph (235.2 ddph), pepsin
specific activity increased six times (6.03 ± 0.94 U mg−1
protein) (P < 0.05) and become
stable until 28 dph (490 ddph) (P > 0.05). From 28 to 42 dph (490 to 756 ddph), pepsin

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
specific activity increased again and reached the maximal activity values (9.42 ± 0.75 U mg−1
protein) and then remained stable until the end of the study at 50 dph (906.5 ddph) (Fig. 3a).
Alkaline proteases, trypsin and chymotrypsin, were detected in newly hatched beluga
larvae. The highest values of trypsin specific activity were reached at the transition to
exogenous feeding at 14 dph (235.2 ddph) (0.12 ± 0.01 U mg−1
protein) (P < 0.05). Then,
trypsin activity decreased abruptly and remained constant until 35 dph (612.5 ddph).
Afterwards, trypsin specific activity increased until 42 dph (756 ddph) when it became
constant until the end of study (Fig. 3b). Chymotrypsin specific activity remained constant
from hatching until the onset of exogenous feeding, whereas after then, it progressively
decreased until 23 dph (400.2 ddph). From 23 to 50 dph (906.5 ddph), chymotrypsin specific
activity increased, reaching its maximum values (0.20 ± 0.02 U mg−1
protein) (P < 0.05) at
the end of the study (Fig. 3c).
Alpha-amylase specific activity remained constant (15.81 ± 1.22 U mg−1
protein)
from hatching to first feeding (P > 0.05), peaked between 18 and 23 dph (306 and 400.2
ddph) (P < 0.05) and decreased again from 28 to 42 dph (490 - 756 ddph) when it reached
similar values than in newly hatched larvae (P > 0.05). From this age α-amylase specific
activity doubled (36.93 ± 3.28 U mg−1
protein) and reached maximum values in 50 dph
(906.5 ddph) larvae (P < 0.05) (Fig. 3e).
Lipolytic activity was low in the first week (P < 0.05), whereas it increased until onset
of exogenous feeding and then, until 490 ddph (28 dph) went down again and reached the
same amount in the first week. After then, lipase activity sharply increased until the end of
the study when it reached maximal activity values (0.01 ± 0.00 U mg−1
protein) (P < 0.05).
Lipase specific activity recorded at the end of the study was four times than measured in
newly hatched and first feeding beluga larvae (Fig. 3f).

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Alkaline phosphatase specific activity sharply increased from hatching to 7 dph
(111.3 ddph) (P < 0.05), reaching a maximum value (0.81 ± 0.07 U mg−1
protein) between 7
and 14 dph (111.3 – 235.2 ddph). After the onset of exogenous feeding, alkaline phosphatase
specific activity progressively decreased until 35 dph (612.5 ddph), when it become stable
until the end of the study (P > 0.05) (Fig. 3d).
4. Discussion
Physiological studies during the early stages of fish development, as well as
ontogenetic changes of the digestive enzyme activities, are valuable tools for better
understanding the nutritional capabilities of young larvae (Diaz et al., 1997) and optimizing
feeding protocols (Cahu and Zambonino-Infante, 1997; Lazo et al., 2000). Therefore, a better
understanding of the enzymes involved in digestion and their functionality during larval
ontogeny, in combination with the knowledge of the natural diet of fish will aid in optimizing
the nutritional composition of formulated microdiets to be compatible with the digestive
physiology of the developing organism (Lazo et al., 2011). In this sense, it is very important
to synchronize the physiological status of the larva in terms of its digestive capabilities with
the feeding protocol and weaning process, because the success of larval rearing is highly
dependent upon it.
In the present study, most of the mortality during H. huso larval rearing was observed
in two different stages. A first one comprised between hatching and the onset of exogenous
feeding, when larvae underwent an intense process of organogenesis (Dettlaff et al., 1993),
and a second one linked to larval weaning. Mortality rates occurring during the endogenous
feeding phase of beluga were higher (50% vs 2.5%) than those found in A. baerii (Gisbert et

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
al., 2002), which may be attributed to differences in egg quality (Dettlaff et al., 1993;
Kamler, 2008). Thus, it seems advisable to pay more attention into beluga broodstock
management from a reproductive and nutritional point of view (see reviews in Dettlaff and
Goncharov, 2002; Migaud et al., 2013). In addition, the sharp increase in mortality observed
in beluga larvae during the transition from live prey to a compound feed clearly indicated that
actual feeding practices and diets should be refined and tuned-up to match the digestive
physiology and nutritional requirements of this species.
In the present study, all the assayed digestive enzymes were already detected in
beluga hatchlings, which confirmed that the process of enzyme production is triggered by the
underlying genetic mechanisms (Buddington and Diamond, 1989) rather than dietary induced
(Cahu and Zambonino-Infante, 1994; Lazo et al., 2000). This early detection of enzymes has
been reported in many marine and freshwater fish species (Rønnestad and Morais, 2007;
Zambonino-Infante et al., 2009), and it would allow larvae to start digesting food items as
soon as the onset of exogenous feeding takes place.
During larval stages, protein digestion occurs mainly by the action of alkaline
proteases such as trypsin and chymotrypsin in combination with intestinal cytosolic
peptidases (Zambonino-Infante and Cahu, 2001). These enzymes have limited capacity for
digesting macromolecules that are absorbed by the pinocytotic activity of the enterocytes in
the posterior intestine where they are intracellularly digested. Similarly to other acipenserids
(Babaei et al., 2011) and other marine and freshwater teleosts these two alkaline proteaseas
are detected at hatching (Darias et al., 2007; Srivastava et al., 2002; Pradhan et al., 2013;
Uscanga-Martínez et al., 2011, among others) The activity of trypsin and chymotrypsin in the
newly hatched H. huso larvae reveals the importance of these alkaline pancreatic enzymes
during organogenesis and early larval development because both types of proteases might be
involved in the cleavage of proteins contained in the yolk and food items once exogenous

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
feeding takes place (Gisbert et al., 2009), as well as during the hatching process in which the
hatching gland participates in the digestion and breakage of the egg chorion (Dettlaff et al.,
1993). Thus, the high activity of these two alkaline pancreatic enzymes during early larval
development confirmed their important digestive role in chondrosteans, as well as previously
described in modern teleosts (Zambonino-Infante and Cahu, 2001). In addition, the decline in
specific activity of both enzymes along larval development might be mainly explained by the
progressive transformation of the digestion mode from an alkaline larval digestion, mainly
characterized by pancreatic proteases like trypsin and chymotrypsin, to a juvenile acid
digestion mode.
The development of the stomach generally involves an acid digestion and
consequently a more efficient extracellular digestion of proteins (Segner et al., 1994). In the
present study, the increase in pepsin activity was correlated to an abrupt decrease in markedly
decreasing in trypsin and chymotrypsin activities after the exhaustion of yolk sac and onset of
exogenous feeding. Although the histological development of gastric glands was not
observed in H. huso until 3 dph (49.5 ddph) (Asgari et al., 2013), pepsin activity was already
detected in newly hatched larvae. These results are similar to those reported in A.
transmontanus, A. fulvescens, A. gueldenstaedti, A. stellatus and H. huso embryos
(Buddington and Doroshov, 1986; Kopylenko et al., 1984). In this sense, this activity is most
likely not stomach pepsin, but some other acid proteases such as lysosomal cathepsins that
might be involved in the digestion of yolk proteins (Carnevali et al., 2001; Sire et al., 1994).
However, the proteolytic activity found in yolk-sac H. huso larvae needs to be characterized
by means of specific inhibitor assays and substrate-SDS-PAGE procedures.
Similar to previous studies of embryos and sturgeon larvae (Kuzmina and Gelman,
1998), trace levels of α-amylase activity were detected in H. huso hatchlings, although they
sharply increased after the transition to exogenous feeding. These results were similar to

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
those reported in A. persicus (Babaei et al., 2011) and A. baerii (Zółtowska et al., 1999), as
well as in most of the teleost species described so far (Lazo et al., 2007; Ma et al., 2005;
Pradhan et al., 2013; Zambonino-Infante and Cahu, 2001, among others). This increase in α-
amylase activity after onset of exogenous feeding reflected the relatively high carbohydrate
content (6–10%) in live zooplanktonic prey (Ma et al., 2005) used for feeding larvae, which
might have differentially stimulated the synthesis and secretion of α-amylase during beluga
larval development. The detection of α-amylase specific activity from hatching in sturgeon
species similar to teleost larvae supported the idea of the existence of a hereditary component
in this process and also indicates that the predisposition to synthesize this enzyme before the
commencement of the exogenous feeding phase is independent of external diet (Lazo et al.,
2011). As a rule of thumb, α-amylase specific activity has been shown to be high during
larval stages and generally decreases during development when the juvenile stage is reached,
which is considered as an indicator of pancreas maturation in carnivorous fish species (Cahu
et al., 2004; Cahu and Zambonino-Infante, 1994; Rønnestad et al., 2013). In this study, the
decrease in α-amylase observed in H. huso from 400.2 to 490 ddph (23- 28 dph) might be
attributed to the acquisition of a juvenile-like digestion mode, rather than a dietary induced
change in α-amylase production and secretion, because there were not any significant
differences between carbohydrate content of live zooplanktonic preys and the inert diet (6-10
% and 11.6 %, respectively). Furthermore, increase in amylase activity from 28 dph (490
ddph) onwards in this study would enable fish to more adequately take advantage of nutrients
through the hydrolysis of glycogen from ingested food (Uscanga-Martínez et al., 2011).
Despite of the major advances in the understanding lipid digestion in juvenile and
adult fish, the knowledge of lipolytic enzymes in larvae is still scarce, particularly in terms of
luminal digestion of dietary lipids (Rønnestad and Morais, 2007). In this study, , lipase
specific activity was detected at hatching and during the endogenous feeding stage, which

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
coincides with previous data on A. baerii (Gisbert et al., 1999) that suggested the
participation of this enzyme in the digestion of lipids contained in yolk. In this study, lipase
activity remained stable during all the phase of larval rearing during which larvae were fed
with live prey (Artemia nauplli and cladocerans), whereas it sharply increased as a result of
weaning. Changes in lipase activity might be attributed to changes in food quality and
quantity rather than lipolytic digestive capabilities of fish (Morais et al., 2004; Rønnestad et
al., 2013; Rønnestad and Morais, 2007). For instance, some studies have correlated a
decrease in lipase specific activity to a lower lipid content of cladocerans with regards to
Artemia nauplii (Øie et al., 2011), as well as higher proportion of sterols and phospholipids
(Bychek et al., 2005). However, present results seemed to indicate that the slight increase in
dietary lipid content (11% vs 15%) during weaning, as well as the different lipid class
composition of live prey versus compound diets might explain the observed increase in
lipase production in H. huso. These results differed from those published in other sturgeon
species such as A. ruthenus (Napora-Rutkowski et al., 2009) and A. transmontanus
(Buddington and Doroshov, 1986) where the lipid content of the diet did not significantly
affect lipase activity. However, considering the presence of lipase activity in several tissues
that have nothing to do with lipid digestion, added to the fact that the relationship between
lipolytic activity in the digestive organs and lipolytic activity in non-digestive organs varies
during development (see review in Rønnestad et al., 2013), further studies using protein or
gene expression analyses are needed to clarify whether the increase in lipase activity in H.
huso larvae was due to a maturation of their digestive function or dietary induced. Enzymes
associated with the enterocytes also contribute to the final digestion of proteins and peptides
(Zambonino-Infante and Cahu, 2001). The development of a functional intestine implies
different maturational and morphological events that are very well preserved among
vertebrates (Henning, 1987). From an enzymatic point of view, the appearance of functional

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
microvilli in enterocytes constitutes a crucial step during larval development of fish for the
acquisition of an adult mode of digestion (see review in Zambonino-Infante et al., 2009). In
this sense, it has been suggested that the ontogenetic pattern of trypsin, membranous and
cytosolic peptidase activities during larval development reflect changes in the mode of larval
digestion, which is believed to become increasingly dependent on luminal digestion and less
so on intracellular digestion (Cahu and Zambonino-Infante, 2001). According to the former
authors, this change has been reported to occur around the 3rd and 4th week after hatching in
temperate marine teleost fish species, whereas it takes place at 420 ddph in other Acipenserid
species like A. persicus (Babaei et al., 2011). In contrast, present results revealed that alkaline
phosphatase from enteroctyes’ microvilli sharply increased after first feeding, whereas it
progressively decreased afterwards. The above-mentioned decline in specific enzyme activity
of alkaline phosphatase can be mainly explained by the normal increase of tissue proteins in
growing larvae, which reflects anatomical and physiological changes in fish intestine (Asgari
et al., 2013), and does not correspond to a lowering in the amount of digestive enzymes or
dietary shifts (Zambonino Infante and Cahu, 2001). Although in the present study the activity
of intestinal cytosolic enzymes was not analyzed, it seems plausible that the observed
increase in alkaline phosphatase activity was coupled with a progressive decline in cytosolic
enzyme activities, as this process characterizes the normal development of the intestine and
maturation of the enterocytes in vertebrates (Zambonino-Infante et al., 2009). These changes
in enzyme activity might be in agreement with the increase in the level of histological
organization and complexity reported in the intestine of H. huso (Asgari et al., 2013).
5. Conclusion

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
According to the results from the present study and those previously reported on the
histological organization of the digestive tract in H. huso (Asgari et al., 2013), larvae could
be co-fed with a combination of Artemia nauplii and commercial feed from the onset of
exogenous feeding, although the complete substitution of live prey by inert feed is not
recommended until 235 ddph (14 dph), when larvae exhausted yolk reserves and gastric
stomach was completely functional. Further trials are needed in order to optimize larval
rearing practices under co-feeding regimes with regards to maximal larval survival, growth
performance and quality.
Acknowledgments
We would like to thank the Shahid Marjani Sturgeon rearing center for the assistance
in obtaining beluga broodstocks for spawning and Hamid Eshaghzadeh, Reza Jalili and Dr.
Ahmad Imani for technical assistances during larval rearing and enzyme assay.
References
Asgari, R., Rafiee, Gh., Eagdei, S., Shahrooz, R., Pourbagher, H., Agh, N., Gisbert, E., 2013.
Ontogeny of the digestive system in hatchery produced Beluga (Huso huso Linnaeus,
1758); a comparative study between Beluga and genus Acipenser. Aquacult. Nutr. In
press.
Babaei, S.S., Abedian Kenari, A., Rajabmohammad Nazari, R.M., Gisbert, E., 2011.
Developmental changes of digestive enzymes in Persian sturgeon (Acipenser persicus)
during larval ontogeny. Aquaculture 318, 138–144.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Bardi, R.W., Chapman, F.A., Barrows, F.T., 1998. Feeding trials with hatchery-produced
Gulf of Mexico sturgeon larvae. Prog. fish-Cult. 60, 25–31.
Billard, R., Lecointre, G., 2001. Biology and conservation of sturgeon and paddlefish. Rev.
Fish Biol. Fisher. 10, 355-392.
Bolasina, S., Perez, A., Yamashita, Y., 2006. Digestive enzymes activity during ontogenetic
development and effect of starvation in Japanese flounder, Paralichthys olivaceus.
Aquaculture 252, 503–515.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 72,
248–254.
Buddington, R.K., Diamond, J.M., 1989. Ontogenic development of intestinal nutrient
transporters. Annu. Rev. Physiol. 51, 601-619.
Buddington, R.K., Doroshov, S.I., 1984. Feeding trials with hatchery produced white
sturgeon juveniles (Acipenser transmontanus). Aquaculture 36, 237–243.
Buddington, R.K., Doroshov, S.I., 1986. Structural and functional relations of white sturgeon
alimentary canal (Acipenser transmontaneus). J. Morphol. 190, 201-213.
Bychek, E.A., Dobson, G.A., Harwood, G.L., Guschina, I.A., 2005. Daphnia magna can
tolerate short-term starvation without major changes in lipid metabolism. Lipids 40,
599-608.
Cahu, C., Rønnestad, I., Grangier, V., Zambonino-Infante, J.L., 2004. Expression and
activities of pancreatic enzymes in developing sea bass larvae (Dicentrarchus labrax)

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
in relation to intact and hydrolyzed dietary protein; involvement of cholecystokinin.
Aquaculture 238, 295–308.
Cahu, C.L., Zambonino-Infante, J.L., 1994. Early weaning of sea bass (Dicentrarchus labrax)
larvae with a compound diet: effect on digestive enzyme. Comp. Biochem. Physiol.
109A, 213–222.
Cahu, C., Zambonino Infante, J.L., 1997. Is the digestive capacity of marine fish larvae
sufficient for a compound diet feeding? Aquac. Int. 5, 151-160.
Carnevali, O., Mosconi, G., Cambi, A., Ridolfi, S., Zanuy, S., Polzonetti-Magni, A.M., 2001.
Changes of lysosomal enzyme activities in sea bass (Dicentrarchus labrax) eggs and
developing embryos. Aquaculture 202, 249–256.
Crane, R.K., Boge, G., Rigal, A., 1979. Isolation of brush border membranes in vesicular
form from the intestinal spiral valve of the small dogfish Scyliorhinus canicula.
Biochim. Biophys. Acta. 554, 264–267.
Dabrowski, K., Kaushik, S.J., Fauconneau, B., 1985. Rearing of sturgeon (Acipenser baeri
Brandt) I. Feeding trial. Aquaculture 47, 185–192.
Darias, M.J., Murray, H.M., Gallant, J.W., Douglas, S.E., Yúfera, M., Martínez-Rodríguez,
G., 2007. Ontogeny of pepsinogen and gastric proton pump expression in red porgy
(Pagrus pagrus): determination of stomach functionality. Aquaculture 270, 369–378.
Dettlaff, T.A., Ginsburg, A.S., Schmalhausen, O.I., 1993. Development of prelarvae.
Sturgeon Fishes. Developmental Biology and Aquaculture. Springer-Verlag Ed, Berlin,
Germany, pp. 155–221.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Dettlaff, T.A., Goncharov, B.F., 2002. Contribution of developmental biology to artificial
propagation of sturgeon in Russia, J. Appl. Ichthyol. 18, 266–270.
Diaz, M., Moyano, F.J., Garcia-Carreño, L.F., Alarcon, F.J., Sarasquete, M.C., 1997.
Substrate-SDS-PAGE determination of protease activity through larval development in
sea bream. Aquac. Int. 5, 461–471.
Erlanger, B., Kokowsky, N., Cohen, W., 1961. The preparation and properties of two new
chromogenic substrates of trypsin. Arch. Biochem. Biophys. 95, 271–278.
Folch, J., Lees, G., Sloane-Stanely, H., 1957. A simple method for the isolation and
purification of total lipids from animal tissues. J. Biol. Chem. 226, 497–507.
Furne, M., García-Gallego, M., Hidalgo, M.C., Morales, A.E., Domezain, A., Domezain, J.,
Sanz, A., 2008. Effect of starvation and refeeding on digestive enzyme activities in
sturgeon (Acipenser naccarii) and trout (Oncorhynchus mykiss). Comp. Biochem.
Physiol. 149, 420–425.
Furne, M., Hidalgo, M.C., Lopez, A., Garcia-Gallego, M., Morales, A.E., Domezain, A.,
Domezaine, J., Sanz, A., 2005. Digestive enzyme activities in Adriatic sturgeon,
Acipenser naccarii, and rainbow trout, Oncorhynchus mykiss; a comparative study.
Aquaculture 50, 391-398.
Gawlicka, A., The, S.J., Hung, S.S.O., Hinton, D.E., de la Noüe, J., 1995. Histological and
histochemical changes in the digestive tract of white sturgeon larvae during ontogeny.
Fish Physiol. Biochem. 14, 357-371.
Gessner, J., Chebanov, M., Freyhof, J., 2010. Huso huso. In: IUCN 2012. IUCN Red List of
Threatened Species. Version 2012.2. http://www.iucnredlist.org. Last accessed on 14th
February 2013.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Gisbert, E., Doroshov, S.I., 2006. Allometric growth in green sturgeon larvae. J. Appl.
Ichthyol. 22, 202–207.
Gisbert, E., Gimenez, G., Fernandez, I., Kotzamanis, Y., Estevez, A., 2009. Development of
digestive enzymes in common dentex, Dentex dentex, during early ontogeny.
Aquaculture 287, 381–387.
Gisbert, E., Ortiz-Delgado, J.B., Sarasquete, C., 2008. Nutritional cellular biomarkers in early
life stages of fish. Histol. Histopathol. 23, 1525–1539.
Gisbert, E., Sarasquete, M.C., Williot, P., Castello-Orvay, F., 1999. Histochemistry of the
development of the digestive system of Siberian sturgeon (Acipenser baeri, Brandt)
during early ontogeny. J. Fish Biol. 55, 596–616.
Gisbert, E., Williot, P., 2002. Advances in the larval rearing of Siberian Sturgeon. J. Fish
Biol. 60, 1071-1077.
Henning, S.J., 1987. Functional development of the gastrointestinal tract, In: Johnson, L.R.
(Ed.), Physiology of the Gastrointestinal Tract, 2nd edition. Raven Press, New York,
USA, pp. 285–300.
IFO, 2002. The Statistical Yearbook of Iranian Fisheries Organization. pp. 30- 31. In Farsi.
Iijima, N., Tanaka, S., Ota, Y., 1998. Purification and characterization of bile salt activated
lipase from the hepatopancreas of red sea bream, Pagrus major. Fish Physiol. Biochem.
18, 59–69.
IUCN, 2013. The IUCN Red List of Threatened SpeciesTM. http://www.iucnredlist.org/. Last
accessed on 18th February 2013.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Kamler, E., 2008. Resource allocation in yolk-feeding fish. Rev. Fish Biol. Fisher. 18, 143–
200.
Kolkovski, S., 2001. Digestive enzymes in fish larvae and juveniles—implications and
applications to formulated diets. Aquaculture 200, 181–201.
Kopylenko, L.R., Mitskevich, L.G., Vaitman, G.A., Mosolov, V.V., 1984. Proteinases in the
spawn of sturgeon. Prikl. Biokhim. Mikrobiol. 20, 373–377.
Kuzmina, V.V., Gelman, A.G., 1998. Traits in the development of the digestive function in
fish. J. Ichthyol. 39, 106–115.
Lazo, J.P., Darias, M.J., Gisbert, E., 2011. Ontogeny of the Digestive Tract. in: Holt, G.J.
(Ed.), Larval Fish Nutrition. Wiley-Blackwell, West Sussex, pp. 3-46.
Lazo, J.P., Holt, G.J., Arnold, C.R., 2000. Ontogeny of pancreatic enzymes in larval red drum
Sciaenops ocellatus. Aquacult. Nutr. 6, 183–192.
Lazo, J.P., Mendoza, R., Holt, G.J., Aguilera, C., Arnold, C.R., 2007. Characterization of
digestive enzymes during larval development of red drum (Sciaenops ocellatus).
Aquaculture 265, 194–205.
Ma, H., Cahu, C., Zambonino-Infante, J.L., Yu, H., Duan, Q., Le Gall, M.M., Mai, K., 2005.
Activities of selected digestive enzymes during larval development of large yellow
croaker (Pseudosciaena crocea). Aquaculture 245, 239–248.
May, B., Krueger, C.C. & Kincaid, H.L. 1997. Genetic variation at microsatellite loci in
sturgeon: primer sequence homology in Acipenser and Scaphirhynchus. Can. J.
Fisher. Aquat. Sci. 54, 1542-1547.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Migaud, H., Bell, G., Cabrita, E., McAndrew, B., Davie, A., Bobe, J., Herráez, M.P., Carrillo,
M., 2013. Gamete quality and broodstock management in temperate fish. Rev.
Aquacult. 5, 194–S223.
Morais, S., Cahu, C., Zambonino-Infante, J.L., Robin, J., Rnnestad, I., Dinis, M.T.,
Conceicao, L.E.C., 2004. Dietary TAG source and level affect performance and lipase
expression in larval sea bass (Dicentrarchus labrax). Lipids 39, 449–458.
Napora-Rutkowski, L., Kamaszewski, M., Bielawski, W., Ostaszewska1, T., Wegner., A.,
2009. Effects of Starter Diets on Pancreatic Enzyme Activity in Juvenile Sterlet
(Acipenser ruthenus). Isr. J. Aquacult. – Bamid. 61, 143-150.
Øie, G., Reitan, K. I., Evjemo, J, O., Støttrup, J. S., Olsen, Y., 2011. Ontogeny of the
Digestive Tract. in: Holt, G.J. (Ed.), Larval Fish Nutrition. Wiley-Blackwell, West
Sussex, pp. 307- 334.
Pradhan, P. K., Jena, J., Mitra, G., Sood, N., Gisbert, Enric., 2013. Ontogeny of the digestive
enzymes in butter catfish, Ompok bimaculatus, (Bloch) larvae. Aquaculture 372-375,
62–69.
Rønnestad, I., Morais, S., 2007. Digestion. In: Fin, R.N., Kapoor, B.G. (Eds.), Fish Larval
Physiology. Enfield, Science Publishers, pp. 201–262.
Rønnestad, I., Yúfera, M., Ueberschär, B., Ribeiro, L., Sæle, Ø., Boglione, C., 2013. Feeding
behaviour and digestive physiology in larval fish: current knowledge and gaps and
bottlenecks in research. Rev. Aquacult. 5, 559-598.
Rungruangsak, K., Utne, F., 1981. Effect of different acidified wet feeds on protease
activities in the digestive tract and on growth rate of rainbow trout (Salmo
gairdneri Richardson). Aquaculture 22, 67–79.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Sanz, A., Llorente, J.I., Furné, M., Ostos-Garrido, M.V., Carmona, R., Domezain, A.,
Hidalgo, M.C., 2011. Digestive enzymes during ontogeny of the sturgeon Acipenser
naccarii: intestine and pancreas development. J. Appl. Ichthyol. 27, 1139–1146.
Segner, H., Storch, V., Reinecke, M., Kloas, W., Hanke, W., 1994. The development of
functional digestive and metabolic organs in turbot, Scophthalmus maximus. Mar. Biol.
119, 471–486.
Sire, M.F., Babin, P.J., Vernier, J.M., 1994. Involvement of the lysosomal system in yolk
protein deposit and degradation during vitellogenesis and embryonic development in
trout. J. Exp. Zool. 269, 69-83.
Srivastava, A.S., Kurokawa, T., Suzuki, T., 2002. mRNA expression of pancreatic enzyme
precursors and estimation of protein digestibility in first feeding larvae of the Japanese
flounder, Paralichthys olivaceus. Comp. Biochem. Physiol. 132A, 629–635.
Timeiko, V.N., Bondarenko, L.G., 1988. A study of digestive enzymes in bester (giant
sturgeon sterlet hybrid) during postembryonic period. Vop. Ikhtiol. 1, 117–123.
Twining, S.S., Alexander, P.A., Huibregste, K., Glick, D.M., 1983. A pepsinogen from
rainbow trout. Comp. Biochem. Physiol. 75, 109–112.
Uscanga-Martínez, A., Perales-García, N., Alvarez-González, C.A., Moyano, F.J., Tovar-
Ramírez, D.G., Gisbert, E., Márquez-Couturier, G., Contreras-Sánchez, W.M., Arias-
Rodríguez, L., Indy, J.R., 2011. Changes in digestive enzyme activity during initial
ontogeny of bay snook, Petenia splendida. Fish Physiol. Biochem. 37, 667-680.
Verreth, J., Segner, H., 1995. The impact of development on larval nutrition. In: Lavens, P.,
Jasper, E., Roelants, I. (Eds.), Larvi' 95, Fish and Shellfish Larviculture Symposium,
Europe. Aquaculture Society, Special publication, 24, Ghent, Belgium.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Worthington, C.C., 1991. Worthigton Enzyme Manual Related Biochemical, 3th Edition.
Freehold, New Jersey. USA.
Zambonino-Infante, J.L., Cahu, C., 2001.Ontogeny of the gastrointestinal tract of marine fish
larvae. Comp. Biochem. Physiol. 130, 477–487.
Zambonino-Infante, J., Gisbert, E., Sarasquete, C., Navarro, I., Gutiérrez, J., Cahu, C.L.,
2009. Ontogeny and physiology of the digestive system of marine fish larvae. In:
Cyrino, J.E.O., Bureau, D., Kapoor, B.G. (Eds.), Feeding and Digestive Functions of
Fish. Science Publishers, Inc, Enfield, USA, pp. 277–344.
Zółtowska, K., Kolman, R., Lopienska, E., Kolman, H., 1999. Activity of digestive enzymes
in Siberian sturgeon juveniles (Acipenser baeri Brandt), a preliminary study. Arch. Pol.
Fish. 7, 201–211.
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Figure legends
Fig. 1. Feeding protocol and larval growth in total length and wet weight in beluga (Huso
huso) from hatching until 50 dph (906.5 ddph). Symbols: ○, body weight; ∆, total length. At
each sampling point, data represent mean value and its respective standard deviation
calculated from 40 specimens.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Fig. 2. Cumulative mortality and summary of the major histological events data of digestive
system (Asgari et al., 2013) of Beluga (Huso huso) during larval rearing. Different dash lines
indicate the exact time of each histological events.

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Fig. 3. Specific activity (U mg−1
protein) of different digestive enzymes in beluga (Huso
huso) from hatching to 906.5 ddph (50 dph). a: pepsin (gastric enzyme). b: trypsin. c:
chymotrypsin. e: α-amylase. f: lipase (pancreatic enzymes) and d: alkaline phosphatase
(intestinal brush border enzyme). Different values of enzyme activity (mean ± SD, n = 6)
with different superscript letters are statistically significant (P < 0.05). The arrows indicate
the commencement of mixed and exogenous feeding, respectively.
ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
Highlihts
• Beluga larvae had a developed intestine at the onset of exogenous feeding
• More beluga weaned earlier, more larvae lived
• Weaning should begin immediately after commencement of exogenous feeding

ACC
EPTE
D M
ANU
SCR
IPT
ACCEPTED MANUSCRIPT
• Complete substitution of live prey by inert feed is not recommended until 235 ddph





























