Embed Size (px)
Citation preview

Universidad Andrés Bello
Facultad de Ingeniería
Geología
Paleontología sistemática y estratigrafía de la Facies (a) de
Formación Toqui (Titoniano-Valanginiano) y sus implicancias
paleoambientales en las cercanías de Puerto Guadal, región
de Aysén
MEMORIA PARA OPTAR AL TÍTULO DE GEÓLOGO
Vicente Mimica Davet
Profesor Guía:
Manuel Suárez Dittus
Miembros de la Comisión:
Jean-Baptiste Gressier
Pierre-Yves Descote

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
1 Índices
Índice de contenido
1 Introducción 4
1.1 Formulación del problema y área de estudio 4
1.2 Marco Geográfico 5
1.3 Objetivos
1.3.1 Objetivo general
1.3.2 Objetivos específicos
6
6
6
1.4 Metodología
1.4.1 Trabajo pre-terreno
1.4.2 Trabajo en terreno
1.4.3 Trabajo post-terreno
7
7
7
8
2 Marco Geológico 10
2.1 Geología regional 10
2.2 Geología local y unidades estratigráficas
2.2.1 Formación Ibáñez
2.2.2 Formación Toqui
2.2.3 Formación Ligorio Márquez
2.2.5 Formación San José
2.2.5 Formación Guadal
13
13
14
17
17
17
3 Marco Teórico 18
3.1 Estromatolitos algales 19
3.2 Troncos fosilizados 26
3.3 Microfósiles 30
4 Resultados
4.1 Estratigrafía
33
33
4.2 Troncos fosilizados 46
4.3 Microfósiles 48
4.4 Paleontología Sistemática 50
4.5 Estromatolitos 55
5 Discusión 58
6 Conclusión 66
7 Agradecimientos 68
8 Bibliografía 69

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
2 Índices
Índice de Imágenes
Figura 1.1 Imagen de ubicación de la zona. 6
Figura 2.1 Figura esquemática que muestra la Cuenca Austral e indica las subcuencas
de Aysén, Magallanes y Rocas verdes (Salgado et al. 2008).
11
Figura 2.2 Estratigrafía Generalizada del Mesozoico de la parte este de la Cordillera
Patagónica Central, Chile (Salgado et al. 2008).
11
Figura 2.3 Estratigrafía Esquemática del Mesozoico (Suáres et al. 2015). 13
Figura 2.4 Columna Estratigráfica de las capas con fósiles de la formación Toqui
(Suáres et al. 2015).
15
Figura 2.5 Esquema geológico (Modificado de De La Cruz et al. 2006). 16
Figura 3.1 Clasificación de estructuras de estromatolitos de tipo LLH, SH y SS de
Logan et al. 1968.
20
Figura 3.2 Estructura de estromatolito compuesto, (Logan et al. 1964) 21
Figura 3.3 Morfologías externas de estromatolitos basadas en observaciones de
campo. Su crecimiento interno, forma de lámina, tipo de márgenes de
lámina, Figura por Martin-Bello (2019) y definidos por Kennard & Burne
(1989).
25
Figura 3.4 Estructura de la madera mostrando sus tres planos: transversal (Tr),
tangencial (Ta) y radial (R), así como anillos de crecimiento y el eje
longitudinal. Modificado de Franke y Quenneville (2011).
26
Figura 3.5 Cortes modelo de conífera, de Guía para cortes de la madera (2018). 27
Figura 3.6 Diagrama de la estructura típica de una dasycladecean adaptado por
Scholle & Ulmer-Scholle (2003) de Wray (1977).
30
Figura 3.7 Morfologías comunes observables en los foraminíferos adaptada de Moore et al. (1952), Loeblich and Tappan (1964) y Culver (1987) por Scholle & Ulmer-Scholle (2003).
31
Figura 3.8 Figura de Scholle & Ulmer-Scholle (2003) redibujada a partir de Moore et al. 32

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
3 Índices
(1952). Muestra la estructura de bola y encaje de los equinodermos
Figura 4.1 Imagen Satelital que indica las transectas por las que las Columnas 1 - 4 34
Figura 4.2 Columna Estratigráfica 1 35
Figura 4.3 contacto a nivel microscópico de la muestra entre los estromatolitos algales
y la arenisca de grano medio que presentaba fósiles calcáreos.
36
Figura 4.4 Columna Estratigráfica 2 38
Figura 4.5 Columna Estratigráfica 3 41
Figura 4.6 Columna Estratigráfica 4 43
Figura 4.7 Columna Esquemática que representa la Facies (a) de la Formación Toqui 45
Figura 4.8 cortes 2A y 2B de tronco T2 46
Figura 4.9 Muestra los cortes 3ª, y 3C de la muestra de tronco T3 47
Figura 4.10 recuperación del material antes del tamizado 49
Figura 4.11 estructura de origen orgánico 50
Figura 4.12 Cymopolia eochoristosporica 51
Figura 4.13 Quinqueloculina sp 52
Figura 4.14 Foraminíferos Glovigerínidos 53
Figura 4.15 Muestra una espina de equinodermo 54
Figura 4.16 Fotrografía de un estromatolito algal observado frente al campamento 55
Figura 4.17 Fotografía de un estromatolito algal, observado en transecta de la Columna
3
55
Figura 4.18 Fotografía de un estromatolito algal observado en la transecta de la
Columna 1
56
Figura 4.19 Muestra pellets y peloides en la zona del corte que incluye los
estromatolitos.
57
Figura 4.20 Corresponde a una imagen de la muestra AYE82 que muestra la presencia
de siliciclastos y recristalización de calcedonia en los estromatolitos.
57
Figura 4.21 Figura 4.20 Modificado de Flüguel (2004), muestra los ambientes
depositacionales de oncoides en el Jurásico Tardío.
64

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
4 Introducción
1 Introducción
1.1 Formulación del problema y área de estudio
En la zona sur del Lago General Carrera, cercano a Puerto Guadal, esta área,
geológicamente, está compuesta por la Formación Ibáñez de edad que va desde el
Jurásico superior al Cretácico inferior bajo; corresponde a una sucesión de rocas
volcánicas y volcanoclásticas de composición principalmente silícea (De La Cruz et al.
2006) Y por la Formación Toqui datada con edad Titoniano al Cretácica inferior, la cual
es una unidad sedimentaria y piroclástica. Ambas Formaciones están en contacto por
estructuras como la Falla Visera (De la Cruz et al. 2006) y a su vez se encuentran
interdigitados los niveles superiores de Formación Ibañez con los niveles inferiores del
Grupo Coyhaique (Suárez et al. 2009).
Los depósitos provenientes de la sucesión marina del Jurásico superior a Cretácico
inferior, acumulados en la parte norte de la cuenca Austral (43º-49ºS), conforman el
Grupo Coyhaique (De La Cruz et al. 2003).
La Formación Ligorio Márquez, correspondiendo a Paleoceno superior, definida como
una unidad fluvial de areniscas de cuarzo con intercalaciones de lutitas con algunos
niveles carbonosos (De La Cruz et al. 2006). Por la Formación San José de edad
Eoceno inferior, la cual es definida como una unidad formada por una sección basal de
conglomerados clastosoportados de base erosiva y areniscas de grano grueso (De La
Cruz et al. 2006). Y por último la Formación Guadal del Oligoceno superior al Mioceno
inferior, descrita como una sucesión sedimentaria marina fosilífera (De La Cruz et al.
2006).
El Grupo Coyhaique está integrado en orden ascendente por las Formaciones Toqui,
Katterfeld y Apeleg que representan una sucesión transgresiva-regresiva (De La Cruz
et al. 2003). La Formación Toqui está compuesta por calizas fosilíferas, areniscas y
tufitas, representado depósitos marinos someros, con fósiles del Titoniano-Berriasiano
(De La Cruz et al. 1996, 2003).

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
5 Introducción
En particular, la Formación Toqui, no ha sido ampliamente estudiada como otras
Formaciones del país. Pese a que en zonas aledañas, aconteció el descubrimiento del
Chilesaurus diegosuarezi.
Debido a esto, y a que no hay publicaciones de estudios de reconstrucciones
paleoambientales en base a microfósiles, se reconoce como una oportunidad de
estudio, donde se halla una zona (18G 0696710E 4807647S) en la que es posible
observar estromatolitos, además, teniendo en consideración el tipo de depósito de la
zona, es posible inferir que habrá presencia de otros tipos de microfósiles, los que
entregarán mayor exactitud del tipo de ambiente, edad y temperatura.
De los microfósiles esperables a ser encontrados en las muestras de rocas
sedimentarias extraídas de la zona de los estromatolitos, corresponden los de tipo
planctónicos y bentónicos, debido a su amplia distribución geográfica, su elevado
número de especies, su habilidad para colonizar diferentes medios y su capacidad de
respuesta a cambios en su entorno. Son una valiosa herramienta para realizar
inferencias paleoclimáticas debido a su sensibilidad a variables indicadoras de
cambios ambientales, como por ejemplo profundidad del agua, temperatura, salinidad
y pH (Round et al. 1990).
Este trabajo forma parte del Proyecto Puesta en valor de geología y conocimiento de
dinosaurios FIC 40000501 a cargo de Geólogos asociados a la Universidad Andrés
Bello, los doctores Manuel Suárez y Jean-Baptiste Gressier además de contar con la
participación del doctor Pierre-Yves Descote asociado a la misma universidad.
1.2 Marco Geográfico
La zona cubierta por esta campaña geológica (Figura 1.1) se encuentra en las
cercanías de Puerto Guadal, a 6,2Km al sureste del sector Las Horquetas, siguiendo
los caminos rurales hechos por los propietarios de la zona hasta la cota de los 1180m
que corresponde al límite arbóreo, donde se ubica el campamento base.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
6 Introducción
La forma de acceso al sector, puede ser realizada en vehículos 4x4 o a cabalgata, ya
que el camino puede encontrarse en mal estado y es arreglado cuando se le da uso a
los aserraderos que existen entre Las Horquetas y el campamento base.
Figura 1.1 Imagen de ubicación de la zona. Figura de color verde corresponde al sitio del campamento
base, ubicado en 46°51’15’’S y 72°25’27’’O al sur del lago General Carrera, donde se muestra en la
derecha la zona cubierta por este estudio que consta principalmente en el área sobre la línea arbórea.
1.3 Objetivos
1.3.1 Objetivo general
Reconstruir el paleoambiente de la Formación Toqui (Cretácico) en la región de Aysén,
a partir de evidencias micropaleontológicas.
1.3.2 Objetivos específicos
● Establecer la estratigrafía de la zona de estudio dentro de la Formación
Toqui y caracterizar las distintas facies.
● Componer una base de muestras micropaleontológicas de la Formación
Toqui, mediante la recuperación microestructuras.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
7 Introducción
● Analizar cortes petrográficos de muestras de estromatolitos y facies
asociadas, y contrastar con muestras macro de fósiles de la zona.
● Determinar el paleoambiente de la zona, en base a las evidencias
obtenidas.
1.4 Metodología
1.4.1 Trabajo pre-terreno
● Se estudia la zona satelitalmente, así como la carta geológica del área de
Puerto Guadal - Puerto Sánchez (De la Cruz et al. 2006). Se prepara
campaña geológica a cargo de los doctores Manuel Suárez y Jean-
Baptiste Gressier.
1.4.2 Trabajo en terreno
● 11 al 18 de Enero: Se levantan columnas estratigráficas de la Formación
Toqui, específicamente de secciones cuya base se encuentra definida por
la presencia de estromatolitos, y se recuperan muestras de las distintas
facies descritas.
● 12 de Enero: Se recuperan rodados de fósiles de vertebrados de la
Formación Toqui y se estudia el contacto de esta con la Formación
Ibañez.
● 19 de Enero: Se recuperan fósiles de troncos de la Formación Toqui y se
continúa sección estratigráfica, y toma de muestras.
● 20 y 21 de Enero: Se complementa la estratigrafía de la Formación Toqui
y se toman datos estructurales a lo largo de esta, así como más muestras
y fotografías.
Además de lo anteriormente mencionado, cabe destacar que al terreno se accedió en
camioneta, habilitando los caminos hasta llegar al campamento base los que se
encontraban bloqueados por árboles caídos. Se trabajó entre los 1200 y 1400 metros
de altura, recolectando cerca de 400 kilos de muestras geológicas a caminata o

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
8 Introducción
cabalgata. Se contó con buenas condiciones climáticas hasta que una tormenta de
nieve dio por terminada la campaña prematuramente.
1.4.3 Trabajo post-terreno
● Orden de muestras: Se almacenan las muestras de forma temporal en la
oficina del doctor Jean-Baptiste Gressier, en el edificio de geología de la
Universidad Andres Bello, sede república. Estas se encuentran rotuladas
con sus códigos otorgados en la campaña.
● Digitalización de información recolectada: Se realizan tablas con la
información de posición, fecha y descripción de puntos importantes de la
campaña geológica y tablas del origen de las muestras obtenidas. Todo
esto como parte del informe de práctica profesional, la cual corresponde a
la participación en la campaña mencionada.
● Selección de muestras para corte: Se realizan listas preliminares de
muestras de las que se requerirá un corte petrográfico
● Realización de columnas estratigráficas: Se realizan columnas
estratigráficas a partir de los datos obtenidos en terreno.
● Trabajo preliminar en mapa: Se avanza en el trabajo de mapeo
utilizando el programa ArcMap, georreferenciando fotografías, realizando
polígonos, polilíneas y
● Reuniones: Se realizan reuniones semanales con los integrantes de la
campaña, para organizar el avance de los informes, y trabajos que se
buscan como resultados de esta.
● Toma de contacto con especialistas: Se hace contacto con Teresa
Torres, doctora de la Universidad de Chile, especialista en Xilología,
quien personalmente ubica estructuras en muestras de madera fósil para
realizar la preparación para el estudio de estas.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
9 Introducción
● Redacción del informe de Título 1: Se redacta el informe de título 1,
como una primera parte a una memoria de carácter científico, la cuál es
resultado directo de la información obtenida en la campaña geológica ya
mencionada.
● Realización de corte de Muestras: Se mandan las rocas y troncos
seleccionados para realizar los cortes petrográficos necesarios para su
posterior análisis.
● Trabajo en laboratorio: Se aplica protocolo para la obtención de
microfósiles mediante la disgregación de rocas, esto se explica en el
capítulo de Micropaleontología.
● Trabajo estratigráfico: Se realiza una descripción estratigráfica y se
elaboran 4 columnas según el área seleccionada para este estudio.
● Análisis de los cortes: Se identifican las rocas en los cortes y los
elementos micropaleontológicos observables en estos con aumentos
x40,x100,x200 y x400.
● Recopilación bibliográfica: Se realiza una segunda ronda bibliográfica
para poder describir los elementos no previstos en un principio, sus
implicancias y la realización de la paleontología sistemática.
● Descripción, Sistemática e Interpretación Ambiental: Se lleva a cabo
la descripción de las muestras obtenidas, se realiza paleontología
sistemática en los microfósiles hallados y se realiza una interpretación
paleoambiental en base a las evidencias.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
10 Marco Geológico
2 Marco Geológico
Este capítulo recoge la caracterización geológica de la región de Aysén mencionando
la geología regional y las Formaciones que aparecen al sur del lago General Carrera, lo
que será necesario para poder determinar el paleoambiente y establecer la estratigrafía
de la zona estudiada más adelante.
2.1 Geología regional:
El Basamento está definido como el complejo metamórfico andino oriental, del
devónico al carbonífero (Hervé, 1993; Hervé et al. 1998). Consta de rocas
metamórficas pre Jurásico Tardío, expuestas en la zona centrooriental de la cordillera
patagónica. El complejo subyace en discordancia angular a la Formación Ibáñez y la
Formación Ligorio Márquez, y constituye cerca de un 45% de las rocas aflorantes en el
área de Puerto Guadal-Puerto Sánchez (De La Cruz et al. 2006).
La configuración geológica del mesozoico en la región de Aysén (44-47°S) incluye tres
episodios volcánicos principales, siendo el primero de estos en el Jurásico Tardío
rellenando una cuenca marina de intra-arco y tras-arco durante el Titoniano al Aptiano
conocida como la cuenca de Aysén (Figura 2.1, Suárez et al. 2015). Siendo entonces
la geología del Jurásico Tardío al Cretácico Temprano de la parte sur de Sudamérica
dominada por un arco magmático a lo largo de la cordillera Andina y una cuenca de
trasarco (Suárez et al. 2009).
Este primer episodio volcánico es representado por la formación Ibáñez siendo
principalmente una unidad volcánica silícea subaerea del Oxfordiano al Valanginiano
(Suárez et al. 2009).
El segundo episodio volcánico es representado por el grupo Divisadero del Aptiano al
Cenomaniano temprano, formado principalmente por rocas volcánicas intermedias
subaereas. Mientras que el tercer episodio es representado por rocas de la Formación
El Toro, del santoniano al Daniano, representada por rocas que incluyen andesitas,
domos riolíticos y basaltos, sobreyace inconformemente al grupo Divisadero (De La
Cruz et al. 2003; Demant et al. 2007)

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
11 Marco Geológico
Mientras que las sucesiones sedimentarias mesozoicas son representadas por las
formaciones Toqui, Katterfeld y Apeleg, las
cuales forman el grupo de Coyhaique,
depositadas en la cuenca de Aysén. (De La
Cruz et al. 2003).
El grupo Coyhaique representa los
sedimentos marinos acumulados en él y
estos son cubiertos por el grupo Divisadero
como se muestran en la Figura 2.2.
Figura 2.1. Figura esquemática que muestra la
Cuenca Austral e indica las subcuencas de
Aysén, Magallanes y Rocas verdes (Salgado et
al. 2008). 1 muestra la cuenca de Aysén y 2 la
cuenca Austral.
Figura 2.2. Estratigrafía Generalizada del Mesozoico de la parte este de la Cordillera
Patagónica Central, Chile (Salgado et al. 2008).

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
12 Marco Geológico
El grupo Coyhaique, está conformado desde su base por la Formación Toqui, del
Titoniano al Hauteriviano temprano, compuesta de areniscas marinas someras, lechos
de ostras y rocas volcanoclásticas, la cual se interdigita con la parte superior de la
Formación Ibáñez (De La Cruz et al. 2003, 2006; Suárez et al 2009). Y al norte del lago
General Carrera, la Formación Toqui subyace a las lutitas negras de la Formación
Katterfeld, que a su vez subyace principalmente a areniscas marinas de facies
mareales de la Formación Apeleg (Bell et al 1997).
Entre el Paleoceno al Mioceno se tiene a la Formación Ligorio Márquez, que pertenece
al Paleoceno Superior (Suárez et al 2000), definida como una unidad de areniscas de
cuarzo con intercalaciones de lutitas y algunos niveles de lutitas carbonosas y de
carbón (De La Cruz et al. 2006). La Formación San José que pertenece al Eoceno
inferior (Flint et al. 1994) definida como una unidad formada por una sección basal de
conglomerados clasto soportados de base erosiva y areniscas de grano grueso que
pasan estratigráficamente hacia arriba a una alternancia de limolitas rojas y púrpuras y
bancos de areniscas (De La Cruz et al. 2006). Y la Formación Guadal del Oligoceno
Superior al Mioceno inferior (Niemeyer, 1975) la cuál está descrita como una sucesión
sedimentaria marina fosilífera y sobreyace en concordancia aparente a la Formación
San José (De La Cruz et al. 2006).
Por último, desde el Mioceno superior la región ha estado sujeta a glaciaciones
intermitentes lo que ha labrado al actual lago General Carrera en su extremo oriental,
dejando una serie de morrenas frontales diferentes (Mörner et al 1989). Diferentes
terrazas lacustres y deltaicas se identifican en los márgenes del lago General Carrera,
lo que podría reflejar distintas líneas de costa de distintos lagos interglaciares
(Lagabrielle et al. 2004).

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
13 Marco Geológico
2.2 Geología local y unidades estratigráficas:
2.2.1 Formación Ibáñez (Jurásico Superior-Cretácico Inferior bajo) (Niemeyer, 1975)
El episodio volcánico que dio lugar a la formación Ibañez, corresponde al
emplazamiento de volcanismo andesítico y félsico, siendo este último representado
principalmente por ignimbritas que son la parte más joven de una gigantesca provincia
ígnea silicia cuyos productos cubren gran parte de la Patagonia, toda el área hasta el
océano Atlántico (Pankhurst and Rapela, 1995; Pankhurst et al., 1998; Féraud et al.,
1999; Pankhurst et al., 2000). Los niveles superiores de la formación Ibáñez
sobreyacen e interdigitan las rocas sedimentarias del grupo Coyhaique (Suárez et al.,
2009), como se muestra en la Figura 2.3, esto se indica por las edades radiométricas
de la Formación Ibáñez y el registro fósil de ammonites berriasianos en la Formación
Toqui (Suárez et al., 2007).
Figura 2.3: Estratigrafía Esquemática del Mesozoico (Suáres et al. 2015)
La Formación Ibañez posee entre 300 a 1300 metros de espesor. Se dispone en
discordancia angular y de erosión sobre el Complejo Metamórfico Andino Oriental y
subyace a la formación Toqui. En sus niveles inferiores, posee una asociación de

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
14 Marco Geológico
brechas caóticas basales, que representan una unidad discontinua de espesor variable
inferior. A estas sobreyacen una asociación de intercalaciones volcanosedimentarias
bien estratificadas (De la Cruz et al. 2006). La Formación se correlaciona
litoestratigráficamente en la región de Magallanes, con la Formación Tobífera
(modificado de Thomas, 1949 en Suárez, 1978) y en la Patagonia Argentina, con otras
unidades de la Formación Lago La Plata (Haller y Lapido, 1980) y el Complejo El
Quemado (Riccardi, 1971).
Mientras que en la base del grupo de Coyhaique (Figura 2.3) se encuentra la
formación Toqui, donde su facies calcárea es equivalente a la Formación Cotidiano
(Ramos, 1976, 1981) y su facies arenosa es equivalente a Las Tres Lagunas
(Ploszkiewicz y Ramos, 1977) en Argentina. La cuál es producto de una transgresión
marina sobre rocas volcánicas subaéreas del Jurásico Superior al Cretácico Inferior del
grupo Ibañez. Consiste de sedimentos clásticos y calcáreos depositados en costas de
alta energía cercanas a volcanos andesíticos (Suárez y De la Cruz, 1994).
2.2.2 Formación Toqui (Titoniano-Valanginiano) (Suárez y De la Cruz, 1994)
La columna estratigráfica de la Figura 2.4 corresponde parcialmente al nivel superior
de la Formación Toqui, puesto que la subyace una sucesión de estratos de areniscas
con y sin estructuras de laminaciones, estructuras de escape de agua, bioturbaciones,
fósiles calcáreos, trazas fósiles y estromatolitos de laminación algal, donde estos
últimos representan arrecifes de algas. Además, que la columna cuenta con tan solo 70
metros de los 300 metros mínimos de espesor que De la Cruz et al. 2006 describe.
Mientras que los troncos fósiles hallados en el nivel inferior con fósiles de vertebrados
de la Figura 2.4 de Podocarpoxylon y esporas trilete (de helechos) que indicarían
condiciones húmedas (Salgado et al. 2008), así como la evidencia de huesos de
cocodrilos (Lio et al. 2011) podrían indicar un ambiente más cercano a uno lacustre.
Finalmente la identificación de glauconita en algunas areniscas verdosas y la
ocurrencia de dos areniscas tuffaceas con trazas fósiles de Thalassinoides y
probablemente skolithos indicarían una sedimentación marina para esas capas (ver
Frey, 1975; Ekdale et al., 1984; Goldring, 1991; Pemberton et al., 1992), pero análisis

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
15 Marco Geológico
posteriores llevados a cabo en otras areniscas verdosas no muestran señales de
glauconita sino de chamosita y celadonita (Suárez et al. 2015).
Figura 2.4: Columna Estratigráfica de las
capas con fósiles de Formación Toqui
(Suárez et al. 2015).
La Formación Toqui se subdivide en cuatro facies principales. La primera (a) es una
asociación calcáreo-arenosa de 30 a 50 metros de espesor que sobreyacen a la
Formación Ibañez y subyace de la segunda facies (b), que es una asociación de

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
16 Marco Geológico
areniscas, brechas y tobas rojas, con fósiles de vertebrados. La tercera (c) facies
posee 150 metros de espesor mínimo y es una asociación de areniscas, brechas y
tobas verdes, la que sobreyace a la anterior y no presenta el techo expuesto. Por
último, la facies (d) de 40 metros de espesor mínimo es una asociación volcanoclástica
con fósiles de invertebrados de color verdoso. La que se apoya sobre ignimbritas
dacíticas de la Formación Ibáñez y subyace en leve discordancia angular a la
Formación Ligorio Márquez. (De la Cruz et al. 2006)
Figura 2.5: Esquema geológico (Modificado de De La Cruz et al. 2006).

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
17 Marco Geológico
2.2.3 Formación Ligorio Márquez (Paleoceno Superior) (Suárez et al. 2000)
Por último, la Formación del Paleoceno Temprano al Eoceno Temprano, Ligorio
Márquez, sobreyace al Mesozoico inconformemente recubriéndolo con depósitos
fluviales (De La Cruz et al. 2006), lo que se observa en el “Cerro Negro” dentro de la
zona de estudio y se indica en la Figura 2.5 al igual que las relaciones de contacto
entre las formaciones Ibañez y Toqui según describe De La Cruz et al. 2006.
La Formación tiene un espesor mínimo de 200 metros y se le asigna una sucesión de
color blanquecino compuesta por areniscas y conglomerados de cuarzo con
estratificación cruzada que incluyen ocasionales intercalaciones de areniscas de grano
fino a muy fino y limolitas, que describe un ambiente fluvial (De La Cruz et al. 2006).
2.2.5 Formación San José (Eoceno inferior) (Flint et al., 1994)
Esta Formación fue definida como una unidad conformada por una sección basal de
conglomerados clastosoportados de base erosiva y areniscas de grano grueso, posee
espesores variables entre 80 y 200 metros y es una unidad continental caracterizada
por la presencia de paleocanales. La parte inferior de es de un color amarillento a
verdoso y la sección superior es en general de un color rojizo a amarillento (De La Cruz
et al. 2006).
2.2.5 Formación Guadal (Oligoceno Superior-Mioceno Inferior) (Niemeyer, 1975)
Si bien ningún material recuperado por esta campaña geológica pertenece a la
Formación Guadal, su cercanía con el área de estudio y su importancia en la
Paleontología de la región de Aysén, hacen que se haga indispensable mencionarla.
Es una Formación de aproximadamente 110 metros de potencia y color verde-pardusco
que sobreyace discordantemente a la Formación San José. (De La Cruz et al. 2006).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
18 Marco Teórico
3 Marco Teórico
Con motivo de concretar los objetivos propuestos, se analizarán evidencias
paleontológicas en base a muestras de estromatolitos, restos de troncos y microfósiles,
obtenidos en trabajo de terreno, además de trabajo de laboratorio de este material.
Posteriormente, se realizará su clasificación taxonómica en base a caracteres
morfológicos y anatómicos en el capítulo de paleontología sistemática, para luego
compararlos con ejemplares homólogos, ya sea en el presente o con otras evidencias
de Formaciones cretácicas. Para así determinar gracias a estas evidencias el
paleoambiente de la zona de estudio.
Respecto a las muestras de estromatolitos, se cuenta con material fotográfico,
muestras de mano y se realizarán cortes petrográficos. Dos muestras de troncos
fosilizados las que se trabajarán en base a tres tipos de cortes: transversales, radiales
y tangenciales de cada muestra. Por último, muestras de rocas sedimentarias dónde se
hallan los estromatolitos en forma de intercalaciones estromatolitos-areniscas, se
trabajarán estas areniscas para buscar otros microfósiles propios de una columna de
agua, en base a evidencia bibliográfica.
Los resultados del trabajo sobre las muestras descritas anteriormente se incluirán en
un capítulo posterior, mientras que en este, se discutirá la teoría detrás del trabajo que
se desarrollará para dicho análisis y las razones de por qué servirá para cumplir el
objetivo general.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
19 Marco Teórico
3.1 Estromatolitos algales:
El uso y la definición del termino estromatolito ha pasado por una revisión gradual
(Riding 1977; Krumbein 1983) y osilan entre una definición descriptiva o genética
(Monty 1977; Semikhatov et al. 1979; Awramik 1984, 1990). Flügel (2004) recomienda
la definición propuesta por Riding (1999) quien los define como: Los estromatolitos son
depósitos microbiales bentónicos laminares.
En el trabajo ‘Classification and environmental significance of algal stromatolites’
(Logan et al. 1968) se realiza una clasificación presentada en la Figura 3.2. Para
estromatolitos de tipo algal, la cual se utilizará para las descripciones de las muestras
de estromatolitos y sus fragmentos, tanto en fotografías, muestras de mano como
cortes petrográficos.
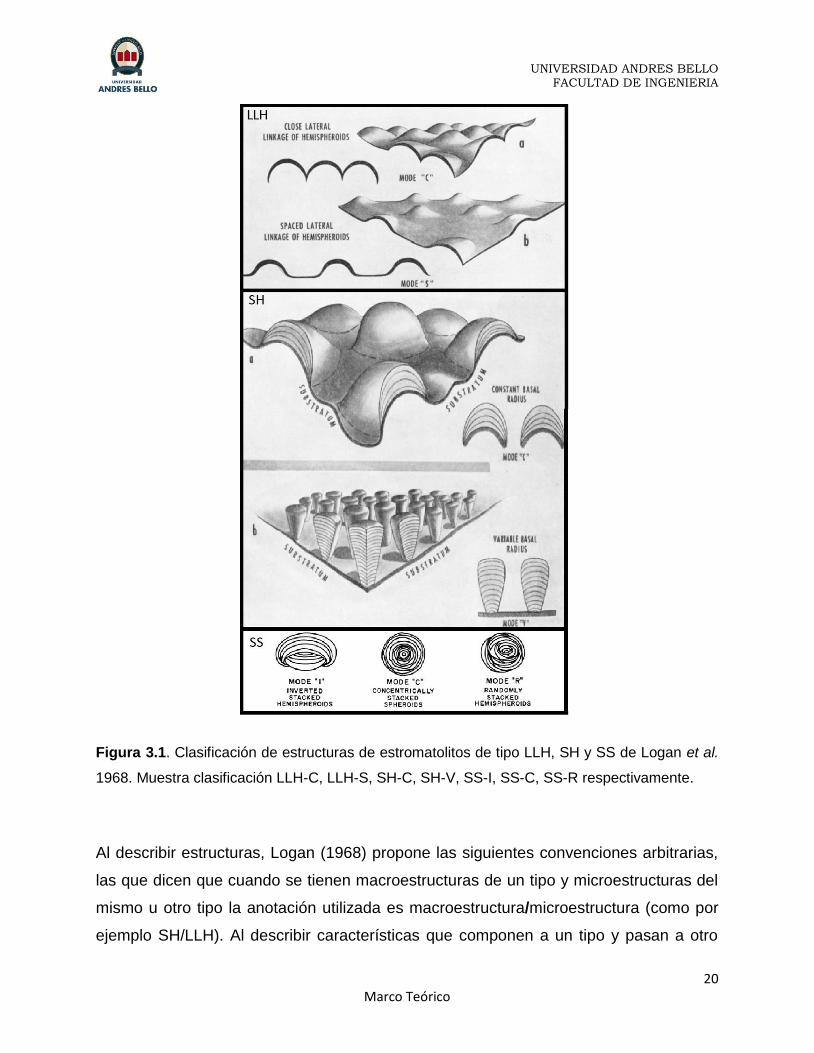
La fórmula estructural que desarrolla Logan (1968) es un sistema que permite expresar
cualquier arreglo de estructuras, las cuales se componen por hemisferoides
lateralmente ligados (LLH), hemisferoides apilados (SH) y estructuras esferoidales
(SS). Siendo útil tanto para describir estromatolitos bien preservados y expuestos como
para fragmentos de estos. (Figura 3.1)
UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
20 Marco Teórico
Figura 3.1. Clasificación de estructuras de estromatolitos de tipo LLH, SH y SS de Logan et al.
1968. Muestra clasificación LLH-C, LLH-S, SH-C, SH-V, SS-I, SS-C, SS-R respectivamente.
Al describir estructuras, Logan (1968) propone las siguientes convenciones arbitrarias,
las que dicen que cuando se tienen macroestructuras de un tipo y microestructuras del
mismo u otro tipo la anotación utilizada es macroestructura/microestructura (como por
ejemplo SH/LLH). Al describir características que componen a un tipo y pasan a otro

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
21 Marco Teórico
tipo se utilizará una flecha indicando el cambio a la nueva estructura (un ejemplo sería
LLH→SH), describiendo siempre las estructuras de base a techo ya que el crecimiento
de los estromatolitos es hacia arriba. Esta convención se encuentra indicada en la
Figura 3.2 y se pueden tomar las secciones verticales mostradas como ejemplos para
las fórmulas.
Figura 3.2. Estructura de estromatolito compuesto, (Logan et al. 1964)

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
22 Marco Teórico
La significancia ambiental de la formación de estructuras de estromatolitos de tipo algal
y los posibles cambios de estas estructuras son resultado de una sucesión de eventos
en el ambiente y se cree que las bases para la reconstrucción paleo-ecológica se
encuentra en los límites del conocimiento de los estromatolitos algales modernos
(Logan et al. 1968).
Las estructuras de tipo LLH se desarrolla característicamente en esteras (mats) y
sedimentos ligados a algas de ambiente marino intermareal lodoso, principalmente en
sectores protegidos como tras islas y crestas que hacen de barrera y la acción de las
olas es leve. (Black, 1933; Ginsburg et al., 1954; and Logan, 1961). Aunque la mayoría
de las esteras de algas (algal mats) están descritos desde ambientes intermareales,
hay ocurrencia en lagos como el lago Cowan del Oeste de Australia (Clarke and
Teichert, 1946).
La formación de domos de las articulaciones laterales puede deberse a cuatro factores
ya sea de forma independiente o en combinación, como la expansión lateral de las
esteras, crecimiento sobre irregularidades preexistentes (Ginsburg et al., 1954),
diferencias en características del relieve donde el sedimento será más activo en las
elevaciones y las esteras algales en las depresiones (Ginsburg et al., 1954; Logan,
1961) y por evolución de gases bajo las esteras (Logan et al. 1968).
Logan (1968) describe 8 factores que cambian las estructuras LLH a domos
individuales y formas compuestas de estromatolitos de esteras algales en los terrenos
protegidos en los ambientes litorales marinos y en lagos continentales los cuales, estos
son:
1) Mojado y secado por la fluctuación mareal.
2) Inundaciones por olas altas de tormenta en el litoral marino y afluencias de
escorrentía aperiódicas en lagos
3) Mojado por salpicaduras de olas
4) Prolongados periodos de desecación durante la marea baja y por evaporación de
aguas poco profundas en lagos.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
23 Marco Teórico
5) Daños y fragmentación mecánica por olas de tormenta
6) Roce y canalización por escurrimiento de aguas mareales
7) Enterramiento por afluencia de sedimentos al terreno con las esteras
8) Actividad biótica de animales.
La formación y mantención de estructuras verticales SH dependen de la inhabilidad de
los materiales algales activos en los domos para atravesar las áreas separándolos o
por la destrucción de las esteras y laminas en las interáreas, y Logan (1968) explica
que hay 6 factores que interactúan en la mantención o formación de domos discretos
SH:
1) El relieve, cuando el relieve entre elevaciones y depresiones es muy alto y las
esteras solo ocupan el espacio en las elevaciones y no logran conectarse por las
depresiones.
2) El endurecimiento y mantención de domos de alto relieve compuestos de algas
ligadas a granos de sedimentos como los descritos en Shark Bay (Logan, 1961),
dependen en el endurecimiento del sedimento carbonático, particularmente en las
porciones inferiores de las colonias, siendo este rápido endurecimiento una
característica del ambiente litoral (Rodgers, 1957).
3) Mojado prolongado por el efecto del agua mareal y sus salpicaduras que tienden a
acumular depresiones que inhiben el crecimiento de las algas (Logan, 1961).
4) Factores mecánicos como el escape de aguas mariales y la moción transitoria de las
olas concentradas en depresiones en la superficie del litoral inhiben el crecimiento de
las algas. El efecto es de desarrollar domos elongados aproximadamente paralelos a la
dirección de la corriente (Logan, 1961).
5) Desecación y agrietamiento de las esteras algales y sedimentos en las periferias de
los domos como los descritos por Black (1933) como el mecanismo para la formación
de SH-C en las Bahamas.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
24 Marco Teórico
6) Altas tazas de sedimentación inundando el interárea entre domos con sedimento rico
en cuarzo resulta en la destrucción de las esteras en el interárea. Aunque esta puede
ser una condición temporal y la estera puede reestablecerse si los otros factores no
intervienen.
Se conocen estructuras SH recientes en lagoons hipersalinos como el de Shark Bay al
Oeste de Australia (Logan, 1961). Y Domos discretos SH-V son comunes en arrecifes
donde las olas son moderadas. Y en cualquier estera algal LLH afectada por los
factores anteriores (Logan et al. 1968).
Finalmente, las estructuras de tipo SS (o de tipo oncolitos) a diferencia de los
estromatolitos SH y LLH no requiere de períodos de desecación, sino que
generalmente indica un sumergimiento permanente (Ginsburg, 1960, p. 29). Las
estructuras SS-I son estructuras que hoy se observan en ambientes poco profundos de
zonas intermareales, las que pueden formarse por desprendimiento de estromatolitos
SH. Los de tipo SS-R representan frecuente agitación separada por largos períodos lo
que resulta en organizaciones aleatorias quedando en diferente posición con cada
movimiento. Y por último las estructuras SS-C indican un movimiento continuo del
agua, restringido a áreas continuamente bajo el agua y suficientemente agitadas como
para permitir la formación continua del esferoide (Logan et al. 1968).
Flüguel (2004), explica que la clasificación de estromatolitos de Logan et al. (1968) Fue
desarrollada para describir formas de crecimiento de estromatolitos intermareales y
submareales de baja profundidad, los cuales se creían ser predominantemente
controlados por el grado de turbulencia del agua. Expone que esta interpretación puede
caber para cuerpos grandes de estromatolitos, pero que en el caso de que sean
pequeños, su composición estructural depende fuertemente de la biota asociada. Por
último explica que se deben utilizar métodos cuantitativos para el estudio de la fábrica
de los estromatolitos, los que facilitarían el reconocimiento de cambios
microambientales.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
25 Marco Teórico
La descripción de los estromatolitos en distintos puntos de la columna sedimentaria
estudiada, se describirán según la Figura 3.3 y su morfología externa observada en
terreno basándose en los términos definidos en Kennard & Burne (1989).
Figura 3.3. Morfologías externas de estromatolitos basadas en observaciones de campo. Su
crecimiento interno, forma de lámina, tipo de márgenes de lámina, Figura por Martin-Bello
(2019) y definidos por Kennard & Burne (1989).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
26 Marco Teórico
3.2 Troncos fosilizados:
Respecto a los troncos fosilizados, (Salgado et al. 2008), clasificó como taxón
Podocarpoxylon (coníferas) una muestra de tronco fósil obtenida de la zona de estudio.
Debido a esto, las muestras con las que se cuenta serán estudiadas para determinar si
se trata de un nuevo taxón o si se puede mejorar la clasificación ya descrita realizando
un análisis xilotómico a partir de las descripciones sistemáticas.
Las descripciones sistemáticas se realizarán a partir de tres cortes petrográficos de
cada muestra que coincidan con los planos transversales, radiales y tangenciales
(Figura 3.4 y Figura 3.5) rigiéndose por el código de la ‘International Association of
Wood Anatomists’ basándose en el método utilizado por Torres et al. 2002.
Figura 3.4: Estructura de la madera mostrando sus tres planos: transversal (Tr), tangencial (Ta)
y radial (R), así como anillos de crecimiento y el eje longitudinal. Modificado de Franke y
Quenneville (2011).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
27 Marco Teórico
Figura 3.5: Cortes modelo de conífera, de Guía para cortes de la madera (2018).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
28 Marco Teórico
Para clasificar la muestra con su homólogo se utilizará la revisión de Bamford &
Philippe (2001) quienes generaron una base de datos con 348 troncos fósiles del
jurásico al cretácico temprano donde se incluyen 20 casos distintos de Podocarpoxylon.
La anatomía vegetal como rama de la botánica se encarga del estudio de la morfología
de los tejidos en las plantas y de la formación de anillos anuales en el leño por efecto
del crecimiento secundario (Evert 2006). Paralelamente, la dendrocronología es una
disciplina científica fuertemente arraigada en la anatomía de la madera y se basa en la
comparación de secuencias de anchos de anillos en especies leñosas (Fritts 1976,
Schweingruber 1996).
En coníferas, los anillos de crecimiento están formados por células con paredes
delgadas y lúmenes amplios al inicio de la temporada de crecimiento representado por
una tonalidad de la madera más clara (leño temprano); luego se forman gradualmente
paredes celulares más gruesas y lúmenes más estrechos con apariencia más oscura al
final de la temporada de crecimiento (leño tardío) (Tortorelli 1956, Roig 1992, Díaz-Vaz
2003). Adicionalmente, existen eventos ambientales que ocurren dentro de la estación
de crecimiento y formación de anillos (e.g. sequías, bajas temperaturas, cambios
bruscos y/o extremos de temperatura y precipitación), que se registran como anomalías
que dificultan la correcta identificación de los límites de cada temporada. Las anomalías
en los anillos de crecimiento son estructuras anatómicas generadas a partir de un daño
celular o formación anormal del tejido xilemático (Schweingruber et al. 1990).
La madera petrificada, es un fósil común y mejor distribuido que restos foliares u
órganos reproductivos entre los sedimentos, menos retrabajados que los palinomorfos
y representando el 80% de cualquier biomasa del ecosistema terrestre. En la
etimología de la anatomía de maderas, esta es utilizada en dos niveles, existe es de
morfotaxa y otro para términos descriptivos. (Philippe y Bamford 2008). Por ejemplo, el
morfogenero de Podocarpoxylon, se considera anatómicamente análogo con el actual
género de Podocarpus o con características anatómicas de la familia Podocarpaceae.
Es por estos motivos que la madera fosilizada se utiliza de forma análoga a la madera
actual. En el proceso de fosilización o también llamado, petrificación, los minerales

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
29 Marco Teórico
reemplazan lentamente la estructura celular de los troncos. Es por ello, que en
paleoxilología se utilizan los tres cortes previamente descritos, los cuales también son
utilizados para maderas actuales.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
30 Marco Teórico
3.3 Microfósiles:
En base a los resultados obtenidos, que son revisados en el siguiente capítulo, se hace
necesario mencionar las características principales con las que algunos Filos son
identificados en cortes petrográficos. Esto se resumirá a partir del trabajo de Scholle &
Ulmer-Scholle (2003) en su libro “A Color Guide to the Petrography of Carbonate
Rocks: Grains, textures, porositty, diagénesis:
1) Algas marinas verdes:
De la División Chlorophyta, Familia Codiaceae (Ordovícico-Reciente) y la Familia
Dasycladaceae (Cámbrico-Reciente), son fotosintéticos y por lo tanto requieren luz.
Son comúnes en profundidades entre los 2 a 30 metros, aunque algunas algas
modernas fuertemente calcificadas son más abundantes en profundidades entre los 50
y 100 metros. La mayoría de las formas calcificadas crecen principalmente en áreas de
clima templado a tropical en niveles normales de salinidad, aunque tienen un alto rango
de tolerancia a esta. En la Figura 3.6 se observa la estructura típica de un alga verde
dasycladecean.
Figura 3.6. Diagrama de la estructura típica de una Dasycladecean adaptado por Scholle & Ulmer-Scholle (2003) de Wray (1977). Son pequeñas estructuras rectas el talo, posee en un arreglo radial sobre un eje central. Además, pueden desarticularse en segmentos individuales.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
31 Marco Teórico
2) Foraminíferos:
Del Orden Foraminiferida, existen los foraminíferos bentónicos, desde el Cámbrico al
presente, los foraminíferos bentónicos calcáreos, desde el Ordovícico a la actualidad y
los foraminíferos planctónicos, desde el Jurásico Medio al presente.
La mayoría de los foraminíferos son organismos bentónicos, existiendo cercanamente
a 4000 especies modernas, tan solo 40 son plantónicas. Por razones principalmente de
alimentación, los foraminíferos viven en los primeros 300 metros de la columna de
agua, pero después de morir, sus restos pueden caer al fondo marino.
Los foraminíferos pueden ser un grupo principal de los elementos formadores de roca
en depósitos marinos profundos o en plataformas abiertas o restringidas. Y en algunos
casos su abundancia llega a decenas de miles de individuos por metro cuadrado de
sedimento. Las morfologías comunes de los foraminíferos se presentan en la Figura
3.7. Y típicamente tienen un rango de tamaño de entre 0.1 milímetro a 1 milímetro,
aunque existen fósiles que pueden casi alcanzar hasta los 20 centímetros en longitud.
Figura 3.7. adaptada de Moore et al.(1952), Loeblich and Tappan (1964) y Culver (1987) por Scholle & Ulmer-Scholle (2003). Muestra las morfologías comunes observables en los foraminíferos.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
32 Marco Teórico
3) Equinodermos:
Filo Echinodermata, Clase Echinoidea (Ordovícico Tardío - Reciente), viven en
condiciones marinas normales, ya que tienen un rango limitado de tolerancia a la
salinidad. Aparecen principalmente como herbívoros o excavadores en áreas arenosas
de plataforma o a lo largo de costas rocosas, también en aguas más profundas
extendiéndose hasta profundidades abisales. Pese a esto, los fósiles de equinodermos
son mucho más comunes en depósitos plataformales en ambientes marinos normales.
Muestran simetría pentameral cuando son adultos, tienen un exoesqueleto fuertemente
calcificado, desde globular a discoidal y vacío que se componen de placas suturadas o
imbricadas calcáreas. En vida, sus placas están cubiertas de espinas elongadas, muy
cortas en algunas especies pero en otras llegan hasta los 8cm de longitud, y
normalmente se desacoplan una vez muerto, lo que las hace un contribuidor
significante de sedimento. Sus espinas están unidas al cuerpo del organismo por un
tipo de estructura de bola y encajecomo se muestra en la Figura 3.8.
Figura 3.8. Figura de Scholle & Ulmer-Scholle (2003) redibujada a partir de Moore et al. (1952). Muestra la estructura de bola y encaje de los equinodermos, que en vida se mantiene unida por tejido orgánico, lo que implica que pierdan las espinas debido a la descomposición y factores mecánicos o biológicos en el retrabajo del sedimento.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
33 Resultados
4. Resultados
En este capítulo se presentan los resultados obtenidos en base a los objetivos
propuestos. Así como el resultado estratigráfico obtenido a partir del trabajo en terreno,
las conclusiones obtenidas en base al trabajo en laboratorio de las muestras de
estromatolitos y la clasificación taxonómica en base a los caracteres morfológicos
observados en los cortes de rocas y troncos de los fósiles. Determinando, en base a los
datos obtenidos, la interpretación paleoambiental de la zona de estudio.
Esto se lleva a cabo con la realización de cuatro columnas estratigráficas, el análisis de
seis cortes de troncos fósiles, el trabajo en laboratorio en la separación de microfósiles
de los estromatolitos y en la observación de seis cortes petrográficos.
Estratigrafía:
Como se mencionó anteriormente, se realizaron cuatro columnas estratigráficas, todas
en el sector de la Formación Toqui, debido a su coloración y contenido correspondería
a la facies (a) Calcáreo arenosa estando ubicada en la parte basal de esta Formación,
esta, en particular, contiene estromatolitos hasta llegar a los niveles de ignimbritas
verdes y rojas. La posición de las transectas estudiadas se indican en la Figura 4.1.
Las columnas serán referidas en este texto como Columna 1 (Figura 4.2), Columna 2
(Figura 4.4), Columna 3 (Figura 4.5) y Columna 4 (Figura 4.6) y la simbología utilizada
en estas columnas se muestra dentro de las mismas figuras. Entregando la totalidad de
la estratigrafía que se puede observar de la base de la unidad Toqui en la zona de
estudio.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
34 Resultados
Figura 4.1. Imagen Satelital que indica las transectas por las que las Columnas 1, 2, 3 y 4
fueron realizadas, el punto de color café muestra el lugar en donde se encontraron los Troncos
Fósiles y las líneas segmentadas la separación entre Ligorio Márquez y Toqui, así como las
facies a y b de Toqui dónde se indica.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
35 Resultados
Figura 4.2. Columna Estratigráfica 1, de la transecta que se observa en la Figura 4.1

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
36 Resultados
La Columna 1, representa 108 metros de la facies (a) de areniscas calcáreo arenosas
de la Formación Toqui, estos se ven representados en 3 afloramientos; el primero de
estos, mantea 20° al sur con un rumbo de N42E, el segundo, mantea en su base igual
que el primero, pero a partir de los 5 metros comienza a mantear con 13° en la misma
dirección de rumbo, y el tercer afloramiento mantiene el rumbo y manteo de la parte
superior del segundo.
Respecto al primero de estos afloramientos (basal) consta de 5,4 metros con una base
de estromatolitos algales de 1,1 metros seguido por una sucesión de areniscas de
grano fino de tan solo 30 centímetros de espesor, en la que se encontraron fragmentos
de fósiles calcáreos, cuyo contacto entre los estromatolitos y las areniscas finas, se
observa en la Figura 4.3, luego en los últimos 4 metros del afloramiento se observa
una intercalación de areniscas de grano medio con grano fino los cuales presentan
estructuras de escape de fluidos y laminaciones y bioturbaciones.
Figura 4.3. Muestra parcialmente el contacto a nivel microscópico de la muestra entre los
estromatolitos algales y la arenisca de grano fino que presentaba fósiles calcáreos. Parte del
corte AYB-01 de la muestra obtenida en la realización de la Columna 1.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
37 Resultados
El segundo de los afloramientos posee de 40 metros de altura, donde continúa la
intercalación de areniscas de grano fino y medio por los primeros 5 metros donde se
siguen observando laminaciones horizontales en las areniscas finas y se estructuras de
escape de fluidos. A esto le siguen 35 metros de tobas de ceniza donde aparecen de
forma intermitente pómez y donde además se observaron laminaciones horizontales y
cruzadas, slumps y restos de carbón.
Finalmente, el tercer afloramiento consta de 35 metros de tobas de ceniza con las
mismas características generales, aunque no se observaron laminaciones, presenta
pómez, restos de carbón y en la parte superior, bioturbaciones.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
38 Resultados
Figura 4.4. Columna 2 de la transecta indicada en la Figura 4.1

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
39 Resultados
La Columna 2 (Figura 4.4) se sigue la transecta a través de una quebrada indicada en
Figura 4.1 cuya base, tal como se observa en terreno y se indica en la Columna C2,
correspondería a una parte de la Formación Ligorio Márquez, con sus característicos
conglomerados cuarcíferos de clastos rojos. En el contacto entre las Formaciones
Ligorio Márquez y Toqui no se observa de manera definida, puesto que aparece un
cuerpo magmático de un basalto con amígdalas y vetillas de Cuarzo en este. A su vez
la primera litología que se observa en la base de la Formación Toqui, son calizas
plegadas y deformadas por estructuras de tipo slump y cuentan con presencia de
estromatolitos. Por esto y la diferencia de edad de ambas formaciones es que se
interpreta como la falla inversa de tipo cabalgamiento descrita en De La Cruz (2006), la
cual habría utilizado las calizas como plano de despegue para que la formación Toqui
se alzara sobre la formación Ligorio Márquez, además que existe la posibilidad de que
el basalto utilizara esta estructura de falla, pudiendo determinar una edad mínima para
la ocurrencia de la génesis de la estructura en caso de datarse.
Por lo tanto la parte que corresponde a la Formación Toqui de esta columna representa
los 85,7 metros superiores. La base corresponde a calizas plegadas de las que se
aprecian 7 metros. A estas lo siguen 18 metros de areniscas de grano fino de
coloración verdosas las que presentan vetillas de calcita. Posteriormente siguen 5
metros, nuevamente de calizas plegadas y deformadas con estructuras de tipo slump
en las que también se observaron vetillas de calcita. Sucesivamente siguen 56 metros
de areniscas donde se intercalan de grano medio con areniscas de grano fino, en los
primeros 25 metros, además, sólo existe presencia de areniscas de grano fino en la
parte superior. En estas areniscas se observan repetidamente estromatolitos. En la
base (primeros 25 metros) hacen aparición ondulitas, las cuales no se vuelven a
observar en los niveles superiores. A los 25 metros aparece un estrato plegado que se
define como de calizas arenosas. Finalmente en la parte superior, dominan estructuras
de laminación plana, y aparecen a su vez fragmentos de carbón y trazas fósiles. Los
troncos fósiles T2 y T3 como se indican en la Columna, no fueron encontrados en la
transecta indicada en la Figura 4.1 sino que se encontraron en el nivel indicado de la

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
40 Resultados
columna a pocos metros de esta transecta (Estos se describen más adelante en este
trabajo).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
41 Resultados
Figura 4.5. Presenta Columna 3 de la transecta indicada en la Figura 4.1

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
42 Resultados
La Columna 3 (Figura 4.5) tiene un manteo de 18° al Sur con un Rumbo S88E,
representa 24,5 metros y corresponde a 2 afloramientos, el primero de 6.5 metros y un
segundo de 8,1 metros.
El primero de estos afloramientos tiene como base 80 centímetros de tobas blancas de
ceniza, seguido de 5,7 metros de areniscas de grano medio con trazas fósiles y
laminación. El segundo de estos afloramientos tiene como base nuevamente
ignimbritas blancas que presentan trazas fósiles y pómez, seguido de una estructura
con un tamaño de 1 metro correspondiente a estromatolitos, seguido por 3 metros de
areniscas de grano medio (de los cuales solo los primeros 2 metros reaccionan al ácido
clorhídrico), sucedido por nuevamente tobas blancas de ceniza, las que esta vez
presentan pómez de coloración verdosa y donde se observaron estrías de falla (223/80
como plano de estría y 310° como dirección de la estría), y finalmente termina el
afloramiento con 20cm de areniscas grises de grano medio.
UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
43 Resultados
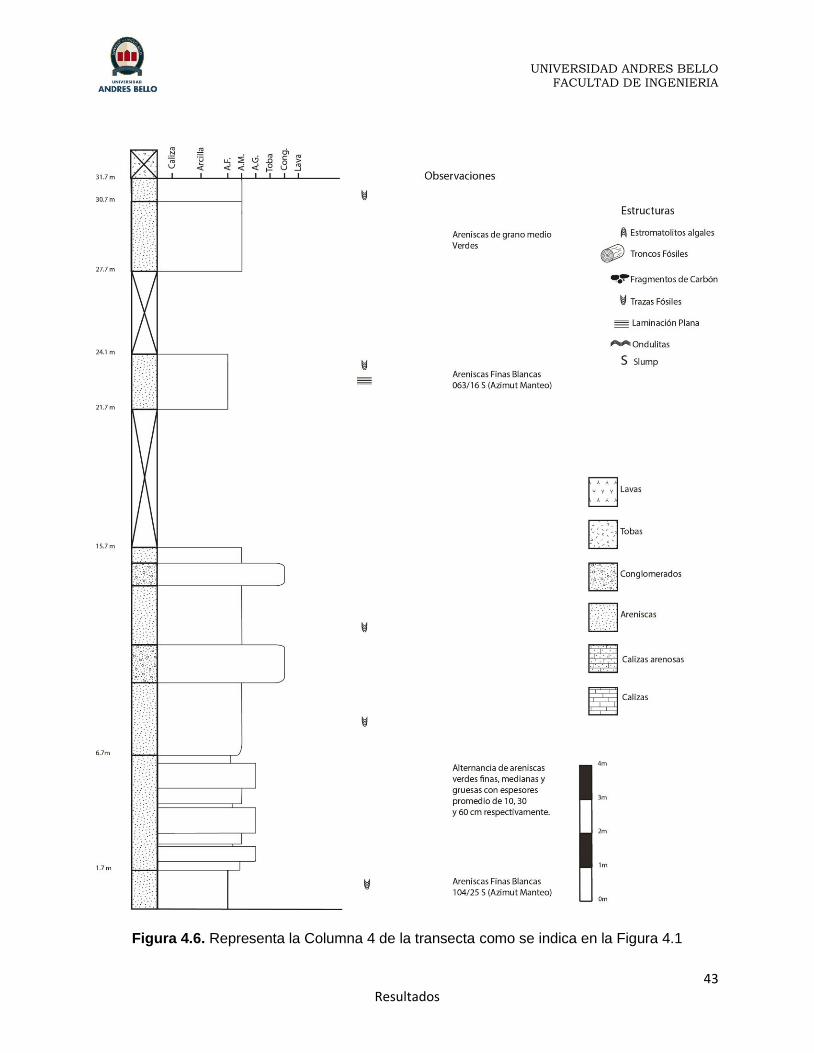
Figura 4.6. Representa la Columna 4 de la transecta como se indica en la Figura 4.1

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
44 Resultados
La Columna 4 (Figura 4.6) posee un rumbo y manteo de S80E/25S y corresponde a
31.7 metros definidos por 3 afloramientos, el primero de 15.7 metros, el segundo de 2.4
metros y un último afloramiento de 4 metros, llegando a los niveles de ignimbritas rojas
en el techo, que corresponderían a facies de la Formación Toqui, diferente a las
areniscas calcáreo arenosas estudiada en este trabajo, y así como la Columna 2
definía la base de estas facies, esta columna define el techo de la misma.
El primer afloramiento de 15.7 metros, presenta en sus primeros 1.7 metros areniscas
finas blancas con trazas fósiles, seguido por 5 metros de una intercalación de areniscas
gruesas, medias y finas donde estas últimas tienen una coloración verdosa, y por
último una intercalación de areniscas de grano medio, las que poseen trazas fósiles,
con conglomerados matriz soportados de matriz fina en coloración café.
El segundo afloramiento, de tan solo 2,4 metros es de areniscas de grano fino
finamente laminadas de coloración blanca que presentan trazas fósiles en las que el
manteo cambia ligeramente a 16° y su rumbo a N63E. En los metros entre el primer
afloramiento y este se observaron rodados de ignimbritas con pomes rojas, ignimbritas
verdes y blancas y se dejan de encontrar rodados de estromatolitos, lo que era común
hasta este punto.
El último de los afloramientos, de 4 metros, corresponde a areniscas de grano medio
con coloración verde y trazas fósiles. Luego comienza un nivel de tobas rojas
intercaladas con ignimbritas de coloración verde de espesor similar (80 centímetros)
muy erosionadas. Se toman estas ignimbritas como el inicio de otra facies de la
Formación Toqui de acuerdo a la descripción de estas. Los contactos se encuentran
con un rumbo y manteo N75E/14°S.
Se realizó una columna estratigráfica esquemática entre estas en la Figura 4.7, para
representar la facies (a) de la Formación Toqui. Mientras que las diferencias entre las
columnas 1 y 3 se explicarían cómo variaciones laterales en la deposición de estas.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
45 Resultados
Figura 4.7. Muestra Columna Esquemática que representa la Facies (a) de la Formación Toqui,
a partir del análisis de las columnas 1,2,3 y 4 presentadas en este trabajo.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
46 Resultados
Troncos fosilizados:
Los troncos fosilizados encontrados T1, T2 y T3 fueron hallados como rodados en el
sector indicado en la Figura 4.1 debido a sus características se logra hacer una
correlación litológica con la Columna 2 (Figura 4.4), la zona se caracterizaba por altura
y poseer baja pendiente, además, a pocos metros (hacia arriba) comienzan los estratos
de ignimbritas verdes y rojas de la Formación Toqui, con características diferentes.
Figura 4.8. Se observan los cortes 2A (izquierda) y 2B (derecha) utilizando el lente de aumento
x10 del microscopio óptico. Estos cortes tienen componentes transversal y tangencial-radial
respectivamente.
Se realizaron cortes 6 cortes de los troncos, 3 de la muestra T2 y 3 cortes de la
muestra T3. Los cortes de T2 (Figura 4.8) corresponden a 1 corte transversal en 2A y
otro, 2B, que salió con componentes tangenciales y radiales, mientras que el tercer
corte observa las mismas componentes que el anterior. Para el segundo fósil, los
cortes de T3 (Figura 4.9) muestran, un corte transversal para 3A, un corte tangencial
para 3B y un corte con componentes radial y tangencial para 3C.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
47 Resultados
Figura 4.9. Muestra los cortes 3A (izquierda) 3B (centro) y 3C (derecha) de la muestra de
tronco T3, con componentes transversal, tangencial y radial-tangencial respectivamente. La
barra de escala mide 0.1mm.
En el caso de T2, no se obtuvieron cortes correctos, debido a la presencia de un nudo,
observable en el corte de la Figura 4.8A, lo que cambia la dirección de las estructuras
de la madera. A su vez en el corte 2A se observaron 54 anillos de crecimiento en 3
centímetros y al menos 4 anillos falsos.
En cuanto al corte 2B, los tipos celulares observados, fueron de compleja identificación,
ya que se presentan deformadas como se observa en la Figura 4.8B. En cuanto al
corte 3B, lo único distinto observado a 2B fue al menos una estructura de punteaduras.
Para el tronco T3, se obtuvieron mejores resultados con los cortes. Y si bien el corte
transversal 3A (Figura 4.9A) no muestra los anillos de crecimiento o estos son poco
notorios, aparecen estructuras de radios uniseriados, y células rellenas de calcita. En el
corte 3B, de componente tangencial (Figura 4.9B) se observan bien los radios leñosos
que van desde 2 células hasta 16 con la mediana en 4 células, además mucha resina,
la cual podría estar rellenando algunos conductos. Y finalmente en el corte 3C (Figura
4.9C) de componente radial y tangencial, se observan punteaduras uniseriadas y
células deformadas por mineralización.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
48 Resultados
Microfósiles:
En la búsqueda de microfósiles, se trabajó con el procedimiento propuesto por Hinchey
1994, por lo que la primera experiencia realizada fue la de asegurar si este
procedimiento era aplicable para las rocas disponibles. Se utilizó la muestra AYE57, la
cual corresponde a un estromatolito recuperado en el nivel indicado en la Columna 2
(Figura 4.4).
Bajo este procedimiento, primero se realizó un quiebre mecánico de la muestra para
reducir el tamaño de esta a los recomendados 10 milímetros de diámetro. Luego se
tomó un volumen de alrededor de 20 mililitros de la muestra para pasar por ácido
clorhídrico al 15% con el objetivo de remover los carbonatos de esta, ya que estos
forman parte del cemento de la roca y la obtención de los microfósiles requería su
disgregación, se neutralizó la muestra con agua destilada y se calentó sobre papel filtro
levemente hasta que se encontrara seca. Luego se le aplicó 20 mililitros de peróxido de
hidrógeno al 30% para eliminar la materia orgánica, este dejó de reaccionar 4 horas
posterior a su aplicación, luego se neutralizó la muestra y se procedió al secado.
Finalmente el resultado se puede observar en la Figura 4.10, dónde es posible apreciar
clastos de gran tamaño.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
49 Resultados
Figura 4.10. Muestra cómo se veía la recuperación del material antes del tamizado, los clastos
de gran tamaño impedían el uso de más aumento en este punto del trabajo, por lo que
reconocer microfósiles a esta altura del trabajo se hizo imposible.
Se pretendía concentrar los restos que parecieran fosilíferos y pasarlos por el
microscopio electrónico de barrido para su posterior identificación, pero para esto sería
necesario repetir el experimento, cambiando algunas de las etapas realizadas, como
por ejemplo, tratar de trabajar las muestras en líquido, y realizar las separaciones por
diferencia de densidad en vez de utilizar un mecanismo mecánico como el tamizaje.
A pesar de esto, se obtuvieron buenos resultados a partir de la identificación de
microfósiles en los cortes de las muestras de estromatolitos obtenidos en terreno.
Donde se pueden observar tanto foraminíferos, restos de moluscos y equinodermos,
otros bioclastos, así como evidencias de algas y granos no esqueléticos como pellets
acompañados de granos terrígenos. Los que se identifican a continuación en el capítulo
de paleontología sistemática.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
50 Resultados
Paleontología Sistemática:
División Chlorophyta Engler, 1903.
Clase Ulvophyceae Stewart KD, Mattox KR (1978).
Orden Dasycladales Pascher, 1931.
Familia Dasycladaceae Kützing, 1843.
Género Cymopolia Lamouroux, 1816.
Especie Cymopolia eochoristosporica Elliott, 1968.
El corte AYE82 (Figura 4.12) fue comparado con diversa bibliografía para determinar
su sistemática.
Figura 4.11. A la izquierda se muestra la estructura de origen orgánico (arriba a NP y abajo a
NX), a la derecha se muestra figura de Horowitz & Potter (2012) de un estromatolito mostrando
capas de limo y arcilla entre estructuras de calcita (Limestone, Middle Ordovician, Garrard
County, Kentucky, U.S.A. IU 8099-910, X 20).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
51 Resultados
Figura 4.12 Muestra parte de la evidencia hallada en AYE82 (arriba), se observa parte del talo
en la parte inferior y las ampollas en el centro. Abajo un corte de Rashidi, Schlagintweit (2017)
tangencial-oblícuo de una Cymopolia eochoristosporica Elliott, 1968

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
52 Resultados
Filo Foraminifera D’Orbigny, 1826.
Clase Tubothalamea Pawlowski, Holzmann & Tyszka, 2013.
Orden Miliolida Delage & Hérquard, 1896.
Familia Quinqueloculinidae Cushman, 1917.
Género Quinqueloculina D’Orbigny, 1826.
En base a la evidencia encontrada en un corte de la muestra AYE92, la cual
corresponde a un estromatolito algal recuperado en 697433E 4807554S a pocos
metros al Este de la transecta en base de la Columna 3, se observa un foraminífero de
morfología milloide
Figura 4.13. A la izquierda se observa un foraminífero bentónico Quinqueloculina sp. Presente
en la muestra AYE92 de un estromatolito algal. A la derecha una imagen de referencia de un
fragmento de una alga dasycladal rellena de arcilla con un foraminífero bentónico de
quinqueloculina (Lower Cretaceous, Coupe de St. Chamas, Bouches du Rhone, France. IU
8099-777, X 20).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
53 Resultados
Filo Foraminifera D’Orbigny, 1826.
Clase Globothalamea D’Orbigny, 1826.
Orden Rotaliida Delage & Hérquard, 1896.
Familia Globigerinidae Carpenter et al., 1862.
Género Globigerina D’Orbigny, 1826.
La Muestra AYE92 también presenta foraminíferos con estructura planispiral (Figura
4.14) típica de los Globigerinida,
Figura 4.14. Foraminíferos Glovigerínidos presentes en la muestra AYE92 de un estromatolito
algal.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
54 Resultados
Filo Echinodermata Bruguière, 1791.
En la muestra AYE82, se encontró una espina de equinodermo (Figura 4.15)
Figura 4.15. Muestra una espina de equinodermo, presente en el corte AYE82, con su encaje
bulboso en el lado inferior derecho y la elongada espina desde el bulbo hasta el lado superior
izquierdo.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
55 Resultados
Estromatolitos:
Figura 4.16. Fotrografía de un estromatolito algal observado frente al campamento de la zona
de estudio, estratiforme, planarmente laminado.
Figura 4.17. Fotografía de un estromatolito algal, observado en transecta de la Columna 3
(Figura 4.1) estratiforme, planarmente laminado.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
56 Resultados
Figura 4.18. Fotografía de un estromatolito algal observado en la transecta de la Columna 1
(Figura 4.1) estratiforme, planarmente laminado.
En las Figuras 4.16, 4.17 y 4.18 se puede acotar que los estromatolitos observados en
terreno, corresponden a la clasificación LLH por Logan et al. 1964 y no se presentan
estructuras de tipo SH (domos). Además, la caracterización de estos en base a los
definidos por Kennard & Burne (1989) se describen como estromatolitos estratiformes
con formas internas laminadas planas, con sus láminas onduladas y de márgenes no
envolventes.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
57 Resultados
Figura 4.19. Corresponde a una sección del corte de la muestra AYB01 de la Columna1
(Figura 4.2). Muestra pellets y peloides en la zona del corte que incluye los estromatolitos.
La gran abundancia de pellets y peloides hallados en los cortes de Estromatolitos
AYB01, AYE82 y AYE83 (Figura 4.19). Parte de estos peloides corresponden a pellets
fecales por su geometría de bastón, ovalada y elongada.
Figura 4.20. Corresponde a una imagen de la muestra AYE82 que muestra la presencia de
siliciclastos y recristalización de calcedonia en los estromatolitos. (la barra roja mide 0.1mm).
La presencia de siliciclastos en los estromatolitos, como se observa en la Figura 4.20,
es un indicador de un ambiente de marismas temperadas (Cameron et al. 1985;
Gerdes and Krumbein 1986; Noffke 1998).
.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
58 Discusión
5. Discusión
Respecto a las muestras de troncos, T1, T2 y T3 fueron hallados como rodados, pero
tanto la coloración, la evidencia microscópica y la roca que acompaña a estos, permite
hacer una correlación litológica con la Columna 2, así también como factores
geográficos, tanto como la altura y la baja pendiente, lo que dificultaría la posibilidad de
que estos provengan de un afloramiento que se encontrara por sobre el indicado.
Finalmente, el hecho de que a pocos metros (hacia arriba) comienzan los estratos de
ignimbritas verdes y rojas de la Formación Toqui, y no comparten coloración ni con el
tronco ni los restos de roca que lo acompaña.
En cuanto al estudio, en el caso de la muestra T2, lo que dificultó que se obtuviesen
cortes correctos, fue la presencia de un nudo, observable en el corte de la Figura 4.8A,
esto provoca que se cambie la dirección de las estructuras de la madera. A su vez en el
corte 2A se observaron 54 anillos de crecimiento en solo 3 centímetros y al menos 4
anillos falsos. De este modo al tener tal número de anillos de crecimiento en el área
previamente nombrada y teniendo como referencia a las podocarpacéas actuales y
fósiles ya que son las más abundantes en la zona de hallazgo, se lograr inferir que el
tronco fósil, proviene de un individuo con hábito arbustivo, por lo que su crecimiento no
sería mayor a tres metros de longitud. También cabe señalar que es probable que la
muestra provenga de una rama lateral y no del eje principal del individuo.
En cuanto al corte 2B, en un principio se pensó ver células del tejido parenquimático,
pero estas estructuras bien pueden explicarse por los crecimientos mineralógicos que
pudieron deformar a las células como se observa en la Figura 4.8B.
A falta de estructuras observables y la baja calidad obtenida en los cortes debido al
nudo, se dificulta realizar una clasificación taxonómica del tronco. No obstante, la
utilidad del corte transversal 2A se mantiene y podría servir para estudios
dendrocronológicos que podrían explicar eventos que ocurrieron en los 54 años de vida
de este espécimen que han quedado registrados.
Respecto a la muestra T3, en el corte 3B, de componente tangencial, se logran
observar bien los radios leñosos que van desde 2 células hasta 16 con la mediana en 4

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
59 Discusión
células. Esto es un dato relevante, ya que el número celular sirve para realizar la
primera gran diferenciación entre grupos de plantas. Variados autores señalan que el
número de células observadas en un corte tangencial, sirve para la diferenciación entre
gimnospermas y angiospermas, en el período geológico del cuál corresponden estas
muestras, ya se tienen registros de las primeras angiospermas, pero, estás poseen un
número mucho mayor de células entre los radios leñosos, así es posible determinar
que la muestra corresponde a una gimnosperma, lo que es concluyente con los datos
previamente obtenidos.
Debido a lo expuesto previamente y cómo no fue posible observar estructuras que
pudieran guiar la descripción a otro tipo de flora, se clasifica T3 como miembro de la
Familia Podocarpaceae de manera preliminar; de todos modos se requiere
complementar la clasificación en base a la una revisión más completa de los cortes de
T3. T2 en cambio, no puede ser clasificado por la falta de cortes en las direcciones
necesarias para realizarlo de forma correcta, pero como se mencionó anteriormente,
tiene uso si se llegan a hacer estudios dendrocronológicos de la zona, aunque en estos
se podrían necesitar nuevos cortes de la muestra.
Es necesario recalcar que también ha sido posible identificar algunas características
que narran la historia de vida de los árboles de los cuales los troncos fosilizados
corresponden.
Respecto a los anillos de crecimiento, la disposición, número de células y espesor son
utilizados como indicadores ambientales.
Dentro de esto, cabe señalar que se observaron anillos falsos, estos, ‘’se reconocen
como bandas delgadas de traqueidas con un tamaño menor al normal incompletas
alrededor del eje del tronco’’. (Brea, M. et al. 2005) Y no se deben considerar para
realizar una reconstrucción paleoambiental o paleoclimatológica.
El indicador tomado a consideración, correspondió al del espesor de los anillos, ya que,
‘’a nivel madera, se observa que los anillos de crecimiento
correspondientes a años de sequías registran menor ancho en comparación con los de
años más húmedos’’ (Lewis, 1993). En las muestras estudiadas se observa de manera

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
60 Discusión
clara este factor de estrés hídrico, ya que es posible observar una alternancia entre dos
anillos gruesos y tres que corresponden a un tercio del tamaño de los anillos normales.
Pese a que no se obtuvo una lámina de rodela del tronco, es posible determinar que los
anillos estudiados no eran parte de la zona central del tronco (zona en la cual los
anillos tienden a ser de menor tamaño), esto se refleja en el grosor de los anillos
observados y la alternancia con anillos de menor tamaño.
Teniendo en consideración estos antecedentes, se puede inferir que el árbol original de
la Familia Podocarpaceae, sufrió en vida un estrés hídrico, el cual fue provocado por
menor cantidad de agua en el ambiente (sequías) o también, etapas con temperaturas
muy bajas, ya que el congelamiento se expresa en los anillos de crecimiento de
manera muy similar a una sequía; pese a ser eventos muy diferentes, generan un
estrés a nivel celular, lo cual es observable y medible tangiblemente, esto se debe a
que su capacidad de transporte de agua a través del tejido conductor se ve disminuido,
por tanto provoca que el espesor del anillo sea menor al normal en condiciones
óptimas.
Respecto al crecimiento, durante la estación favorable es lento, lo que genera anillos
de mayor espesor, además, lo observado en las muestras, corresponden a anillos
incompletos e irregulares alrededor de la circunferencia del tronco. Estos tipos de
anillos comúnmente se observan en árboles que viven bajo condiciones de crecimiento
inestable, produciéndose en períodos con bajas temperaturas, congelamiento, sequías
o por severos ataques de insectos. (Fritts, 1976; Parrish y Spicer, 1988).
La diferencia de grosor en anillos, a pesar de su relación con la edad de los árboles, no
tiene directa relación con un año regular, ya que lo que se registra a nivel celular, es
más bien, los cambios estacionales. Las células tienden a acumularse en las zonas
cercanas a los anillos, demarcando así el cambio de estación no favorable a una más
bien favorable, en el caso que se produce un estrés hídrico, el árbol no tiene tiempo de
generar células que aporten al transporte de nutrientes y agua, de manera normal para
el espécimen.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
61 Discusión
En relación a los anillos que se pueden interpretar como anillos de congelamiento, se
suelen caracterizar por la presencia de una zona interna de células muertas y una zona
externa de células xilemáticas anormales, principalmente de parénquima axial,
producidas después del brusco enfriamiento (Glerum y Farrar, 1966).
En el caso de los anillos analizados, no se presenciaron dichas estructuras, de manera
que se puede interpretar que el espécimen en cuestión estuvo expuesto a períodos con
menor cantidad de agua en el ambiente y no por niveles de congelamiento, es por ello
que presenta anillos más pequeños entre anillos de mayor espesor, ya que tuvo
periodos de estrés hídrico provocado por una ausencia de recurso de agua.
Otro factor muy relevante de este estudio, fue el análisis de microfósiles y su
importancia para el aporte en una reconstrucción paleoambiental.
En relación al procedimiento de estudio de muestras, a través del trabajo en
laboratorio, con una muestra en particular, la cual es posible observar en la Figura 4.10
no fue posible la identificación de microfósiles, puesto que el aumento disponible era
100x, ya que clastos que no se disgregaron luego del procedimiento chocarían con
otros lentes. Entonces para buscar una solución a este problema, se tamizó la muestra,
lo que dio como resultado la destrucción de los posibles microfósiles como los que se
observaban en la Figura, dando por terminado el experimento sin resultados positivos.
Pese a este inconveniente se lograron recuperar microfósiles los que corresponden a
estromatolitos algales y microfauna marina. Tanto las algas, como la microfauna,
coexisten en condiciones específicas de profundidad, luz solar y temperatura y allí
radica su importancia para realizar una reconstrucción paleoambiental.
De las muestras de estromatolitos fue posible obtener otros microfósiles,
correspondientes a microfauna como foraminíferos, restos de moluscos y
equinodermos, además de otros bioclastos, así como evidencias de algas y granos no
esqueléticos como pellets acompañados de granos terrígenos.
En relación a la paleontología sistemática, algunos especímenes presentaron mayor
grado de dificultad de lograr categorías taxonómicas más bajas, quedando sólo en la
categoría de Filo, a continuación se detallan por especímenes.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
62 Discusión
Los estromatolitos corresponden a una agrupación entre células algales y sedimentos,
las algas se lograron identificar como parte de la Especie de Cymopolia
eochoristosporica. Esta fue posible lograr su identificación basando las características
morfológicas observadas con la bibliografía sobre estructuras similares. Por ejemplo la
evidencia que Rashidi, Schlagintweit (2017) muestra, se hace una comparación con la
evidencia obtenida en el corte AYE82 (Figura 4.12). Además Horowitz & Potter (2012)
incluye en una de sus imágenes, una estructura de un estromatolito (Figura 4.11) de
origen orgánico en estromatolitos algales. Esto permite identificar el alga verde que
presenta la muestra ya mencionada, limitando el área de la formación de nuestro
estromatolito entre los 2 a 30 metros, según lo que explica Scholle & Ulmer-Scholle
(2003). Cabe recalcarla que son parte de la División Chlorophyta, que corresponden a
las llamadas algas verdes; estas requieren de luz solar directa, ya que contienen
clorofila A y B, y no poseen pigmentos accesorios, por lo que no pueden hallarse a una
profundidad mayor a los 7 metros, ya que los rayos de luz solar no penetran
directamente a profundidades mayores. Además se debe tener en consideración que
presentan estructuras de tipo LLH las cuales se desarrollan característicamente en
esteras (mats) y sedimentos ligados a algas de ambiente marino intermareal lodoso,
principalmente en sectores protegidos como tras islas y crestas que hacen de barrera y
la acción de las olas es leve. (Black, 1933; Ginsburg et al., 1954; and Logan, 1961).
Esta información además indica que la zona era de baja profundidad y de
características de un medio acuoso sin oleaje y protegidos.
Respecto a la microfauna, fue posible encontrar foraminíferos, de los que se logró
llegar taxonómicamente hasta Género, el cual corresponde a Quinqueloculina. Esto se
debe a que posee morfología milloide, se realíza su clasificación hasta el Orden
Miliolida. Luego, comparando la morfología observada con la evidencia presentada por
Horowitz & Potter (2012) se realiza la clasificación del género. Respecto a otra muestra
de foraminífero, fue posible también lograr la clasificación taxonómica hasta Género,
correspondiendo a Globigerina de estructura planispiral, típica de los Globigerinida,
clasificándolo en dicho orden en base a la descripción y comparación de imágenes con
Scholle& Ulmer-Scholle (2003).

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
63 Discusión
También se halló un fragmento, el cual pudo ser clasificado como parte del Filo
Echinodermata, de este espécimen sólo se halló una espina en sección longitudinal,
con la estructura de encaje de bulbosa. Esto entrega evidencias de que las condiciones
marinas eran normales, por la limitada tolerancia a la salinidad que poseen estos seres
bentónicos.
Con la evidencia micropaleontológica que entregaron los estromatolitos es suficiente
para acotar el ambiente en el que estos se formaron corresponde a marino, así
también, la evidencia de las estructuras sedimentarias descritas en el capítulo de
estratigrafía junto a la composición biótica y abiótica de los estromatolitos, permiten
interpretar en base al estudio de Flüguel 2010 el paleoambiente en el que el que estos
se formaron.
La presencia de clastos siliceos en los estromatolitos, sumado a la evidencia
morfológica de tipo LLH, permite definir el ambiente como uno templado y protegido del
oleaje constante, donde afloran los estromatolitos de estructura planar.
La gran abundancia de pellets y peloides hallados en los cortes de Estromatolitos
AYB01, AYE82 y AYE83 (Figura 4.19) permiten inferir la presencia de agregados de
arcillas carbonáticas y/o precipitados de carbonato de calcio, lo que indica que su
composición original fue aragonito o calcita o una mezcla de ambos según lo que
explica Scholle & Ulmer-Scholle (2003). Además este autor señala que la forma
ovalada y/o alargada son características comunes de pellets fecales. Lo que enriquece
las características del estromatolito en cuanto a biodiversidad se refiere, aunque no se
logran identificar especies a partir de este.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
64 Discusión
Figura 4.21 Modificado de Flüguel (2004), muestra los ambientes depositacionales de oncoides en el Jurásico Tardío. Es una simplificación de la figura original, donde se encierra en rojo la zona ambiental de depositación que cumple con la evidencia observada por este trabajo.
Flüguel (2004) describe los ambientes depositacionales de los oncoides y sus
características en el Jurásico Tardío, y como parte de esto describe los fósiles
asociados en los distintos ambientes como se observa en la Figura 4.21, a partir de
esto, a la aparición de un resto fósil de un equinoideo, la presencia de ondulitas en un
nivel de la Columna 2, de restos de fósiles calcáreos (conchas) en la columna 1 y de
estratificación cruzada en otro nivel y a la intermitencia de los estromatolitos en
diferentes niveles de las columnas, se propone la variabilidad del nivel del mar
provocando el cambio de ambiente entre uno de marisma o playa de barrera, uno
intermareal hasta uno plataformal.
Con dichas evidencias y agregando los antecedentes de la microfauna, es posible
agregar que el ambiente de formación era de una zona de baja profundidad.
De acuerdo a Flügel (2004) la ocurrencia de los peloides es común en ambientes
marinos de baja profundidad, en plataformas y arrecifes o montículos de barro, siendo
también abundantes en carbonatos de aguas profundas. Pero en contraste a su
abundancia en climas tropicales, son raros y/o ausentes en carbonatos de aguas
heladas no tropicales.

UNIVERSIDAD ANDRES BELLO
FACULTAD DE INGENIERIA
65 Discusión
Además Flügel (2004) menciona que si bien los pellets fecales son de origen animal, ya
sean herbívoros, consumidores de plancton y consumidores de detritos, incluyendo
artrópodos, moluscos, gusanos, equinodermos y peces. La asignación de estos pellets
fecales a un productor de este en específico es difícil, ya que la mayor parte de las
fecas producidas por los invertebrados carece de un criterio morfológico.
Por lo que la identificación y abundancia de estos pellets fecales en los cortes, nos
entrega información del clima, el cual se puede suponer dado su abundancia, como
uno de aguas templadas.
Finalmente, respecto a la estratigrafía observada en las columnas 1,2,3 y 4, estas
concuerdan con el ambiente de formación ya identificado en base a la evidencia
paleontológica (entre uno de marisma o playa de barrera a uno intermareal).
De acuerdo a las estructuras observadas en la Figura 4.7 (Columna Esquemática de la
Facies (a) de la Formación Toqui), la interpretación de una playa de barrera sobre un
ambiente de marisma se hace más evidente, puesto que el primero permitiría un
cambio alternante entre sistemas de corriente y sistemas protegidos de la corriente,
debido a la migración de la barrera, la destrucción de esta o su reaparición, así como
los cambios morfológicos que pudo haber sufrido el sistema de playa; permiten explicar
la migración de los estromatolitos en respuesta a estos cambios, explicándose así su
alternancia de aparición en la columna.
El techo de esta columna, está conformado por Ignimbritas blancas con presencia de
pómez, las que se pueden explicar en un ambiente marino si este es somero, como ha
indicado la abundante evidencia ya presentada.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
66 Conclusión
6 Conclusión
La zona de estudio, referida como la facies (a) de la Formación Toqui, en cercanías a
Puerto Guadal, al sur de Mallín Grande, está constituida por una asociación calcáreo
arenosa con estromatolitos algales, que sobreyace a la Formación Ibáñez y se
encuentra cabalgada sobre la Formación Ligorio Márquez. Varía entre los 30 y 108
metros de espesor mínimo. La parte inferior de esta está compuesta principalmente de
areniscas calcáreas y calizas con presencia de estromatolitos algales, trazas fósiles y
laminaciones, mientras que la parte superior aparecen rocas volcanoclásticas de
tamaño ceniza subaéreas con evidencias de actividad biológica relacionada a los
cuerpos de agua. Se encuentra manteando entre los 18° y 25° al Sur cuando no es
afectada por estructuras o fallas.
El ambiente de estas facies, de la Formación Toqui, varía entre uno de marisma o
playa de barrera y uno intermareal, logrando así la deposición de diversos organismos
como partes formadoras del sustrato.
Los estromatolitos algales estudiados, son un buen referente para ubicar las partes que
corresponden al ambiente de marisma o playa de barrera cuando se visita el lugar,
pues su morfología y biogenética así lo indica, ya que son de estructura planar y en
ellos, además, se observaron pellets fecales y seres planctónicos y bentónicos.
La intermitencia de estos estromatolitos podría indicar que el ambiente probable en el
que aparecen, es de playa de barrera, explicando en los cambios morfológicos, de la
playa y su barrera a travez del tiempo, su reaparición a lo largo de la columna.
Finalmente, se identificaron equinoideos, foraminíferos bentónicos (Quinqueloculina sp)
y planctónicos (Globigerinidos), además de algas verdes fotosintéticas (Cymopolia
eochoristosporica), de estos, las algas resultaron ser un importante aporte directo al
sustrato.
Con los antecedentes obtenidos se logró determinar el tipo de ambiente en el que se
formaron las rocas y fósiles estudiados.
Con la información de los Estromatolitos, y al entender el tipo de alga al cual
corresponden esto ya delimitaba la zona a un ambiente poco profundo, la presencia de

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
67 Conclusión
un equinodermo, reforzó la idea de un ambiente marino de salinidad normal, y el tipo de
microfauna hallada apoya la idea de ambiente propuesto.
Además, con el estudio de los troncos fosilizados se logró determinar la Familia de
procedencia (Podocarpaceae) y el hábito (arbustivo), dando una idea de cómo era
también el ambiente en tierra, dada el tipo de flora descrita.
Junto a la evidencia de los peloides, pellets fecales y la Familia identificada con los
troncos fósiles, se logra concluir que el paleoclima de la zona estudiada correspondería
a uno húmedo y de aguas templadas, cercano a un clima mediterráneo.
A pesar de que no se lograron recuperar microfósiles a partir de la disgregación de las
rocas asociadas a los estromatolitos, este trabajo cumple con el objetivo de haber
identificado algunos de estos fósiles mediante cortes petrográficos y se abre la
oportunidad de asegurar que estudios posteriores sobre estas rocas puedan arrojar
algunos que permitan una interpretación paleoambiental más exacta que la que aquí se
ofrece.
En cuanto a los objetivos del trabajo, se lograron, ofreciendo una explicación del
paleoambiente en base a evidencias obtenidas. Se logra realizando un análisis de los
cortes petrográficos de los estromatolitos y facies asociadas. A pesar que no se
compone una base de muestras micropaleontológicas mediante la recuperación de
microestructuras por medio de la disgregación de las muestras de roca, se obtienen
ejemplares a partir de los cortes realizados. Y se establece la estratigrafía de la zona
estudiada.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
68 Agradecimientos
7. Agradecimientos
Al autor le gustaría agradecer al proyecto FIC 40000501 Puesta en valor de geología y
conocimiento de dinosaurios. A los geólogos Manuel Suárez, Jean-Baptiste Gressier y
Pierre-Yves Descote por el apoyo durante la realización de este trabajo. A los
compañeros Daniel Riveros, Benjamín Aldridge, Tomás Piñeiro y Camila Oportus. A la
paleoxilóloga Teresa Torres y geóloga Samantha Leiva. Y por último, a su compañera
de vida, Andréa Yévenes quién le acompañó a lo largo de todo el proceso para
formarse como geólogo.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
69 Bibliografía
8 Bibliografía
Aguirre-Urreta, B.; Suárez, M.; De La Cruz, R.; Ramos, V. 2007. Ammonoids
(Crioceratitinae, Hauterivian) from the Austral Basin, Chile. Revista de la Asociación
Geológica Argentina, Vol.44, p. 387-397.
Bamford, M.; Philippe, M. 2001. Gondwanan Jurassic - Early Cretaceous homoxylous
woods: a nomenclatural revision of the genera with taxonomical notes. Review of
Paleobotany and Palynology, Vol. 113, No. 4, p. 287- 297.
Bell CM, Suárez M (1997) The lower cretaceous Apeleg formation of the Aisén basin,
southern Chile. Tidal sandbar deposits of an epicontinental sea. Revista Geológica de
Chile 24:203–226.
Brea, M. et al. 2005. Análisis de los anillos de crecimiento del bosque fósil de Víctor
Szlápelis, Terciario inferior del Chubut, Argentina. Ameghiniana (Rev. Asoc. Paleontol.
Argent.) 42 (2): 407-418. pp.
Caballero, N., Nass, Y., & Gutièrrez Téllez, B. Diatomeas como indicadoras
paleoambientales en la Formación Río Negro, Provincia de Río Negro, Argentina.
Canizal, A. 2009. Catálogo ilustrado de diatomeas dulceacuícolas mexicanas. Facultad
de Ciencias. Universidad Nacional Autónoma de México.
CLARKE, E. DE D., and TEICHERT, CURT, 1946, Algal structures in a West Australian
salt lake: Am. Jour. Sci., v.224, n.4, p.271-276.
De La Cruz, R.; Suárez, M.; Covacevich, V.; Quiroz, D. 1996. Estratigrafía de la zona
de Palena y Futaleufú (43°15– 43°45′ Latitud S), X Región, Chile. XIII Congreso
Geológico Argentino y III Congreso de Exploración de Hidrocarburos, Actas 1, p. 417–
424.
De La Cruz, R.; Suárez, M.; Belmar, M.; Quiroz, D.; Bell, M. 2003. Geología del área
de Coihaique-Balmaceda, Región Aysén del General Carlos Ibáñez del Campo.
Servicio Nacional de Geología y Minería, Carta Geológica de Chile, Serie Geológica
básica No. 80, p. 5-30.
De La Cruz, R.; Suárez, M. 2006. Geología del área de Puerto Guadal - Puerto
Sánchez, Región de Aisén del General Carlos Ibáñez del Campo. Servicio Nacional de
Geología y Minería, Carta Geológica de Chile, Serie Geología Básica No. 112, p. 5-42.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
70 Bibliografía
Demant A, Suárez M, De la Cruz R (2007) Geochronology and petrochemistry of Late
Cretaceous—(?) Paleogene volcanic sequences from the eastern central Patagonian
Cordillera (45o–45o40’S). Revista Geológica de Chile 34:3–21.
DÍAZ-VAZ, J.E. 2003. Anatomía de maderas. Universidad Austral de Chile, Editorial
Marisa Cuneo, Valdivia. 151pp.
EVERT, R.F.2006. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant
Body: Their Structure, Function, and Development (3rd Edition). John Wiley & Sons,
Inc., New Jersey. 642pp.
Fayó, R & Espinosa, M. 2014. Reconstrucción paleoambiental de la planicie costera
de Mar Chiquita (Provincia de Buenos Aires, Argentina) durante el Holoceno, basada
en diatomeas. Ameghiniana.
Feijoo, C., Ramón, D., Pucha, D., (2018). Guía para cortes anatómicos de la madera.
Universidad Nacional de Loja. Facultad Agropecuaria y de Recursos Naturales
Renovables Carrera de Ingeniería Forestal. Laboratorio de Anatomía de Maderas
Tropicales.
Feraud, G., Alric, V., Fornari, M., Bertrand, H. & Haller, M. 1999. 40Ar-39Ar dating of
the Jurassic volcanic province of Patagonia: migrating magmatism related to Gondwana
break-up and subduction. Earth & Planetary Science Letters. Vol. 172. p. 83-96.
Flint, S.S., Prior, D.J., Agar, S.M. and Turner, P. 1994. Stratigraphic and structural
evolution of the Tertiary Cosmelli Basin and its relationship to the Chile triple junction.
Journal of the Geological Society. London 151. P. 251-68.
Flügel, E. (2004) Microfacies of Carbonate Rocks, Analysis, Interpretation and
Application. Springer, Berlin, 976 p.
Fritts, H.C. 1976. Tree-rings and Climate. Academic Press. Nueva York, 567 pp.
Ginsburg. N., 1955, Recent stromatolitic sediments from South Florida (abs.): Jour.
Paleontology, v.29, p.723.
Ginsburg, 1960, Ancient analogues of recent stromatolites: XXI International Geol.
Cong., pt.22, p.26-35, figs.1-15.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
71 Bibliografía
Glerum, C. y Farrar, J.L. 1966. Frost ring formation in the stems of some coniferous
species. Canadian Journal of Botany 44: 879- 886
Guiry, M.D. & Guiry, G. M. 2009. Algaebase. National University of Ireland. Avaliable
at http://www.algaebase.org; accessed on 07/12/2015.
Gutiérrez Téllez, B. 2007. Reconstrucción paleoambiental del cuaternario de la
cuenca del arroyo Napostá Grande, provincia de Buenos Aires. Universidad Nacional
del Sur, Ph.D. Thesis, 165 p.
Haller, M.; Lapido, O. 1980. El Mesozoico de la Cordillera Patagónica Central.
Revista de la Asociación Geológica Argentina 35, p.230-247.
Hervé, F. 1993. Paleozoic metamorphic complexes in the Andes of Aysén, Southern
Chile (West of Occidentalia). In First Circum-Pacific and Circum-Atlantic Terrane
Conference, Proceedings, p.64-65. Mexico.
Hervé, F.; Aguirre, L.; Godoy, E.; Massone, H.; Morata, D.; Pankhurst, R.; Ramírez,
E.; Sepúlveda, V.; Willner, A. 1998. Nuevos antecedentes acerca de la edad y las
condiciones P-T de los Complejos Metamórficos en Aysén, Chile. X Congreso
Latinoamericano de Geología, Buenos Aires, Vol. II, 134-137.
Hinchey, J. V., & Green, O. R. (1994). A Guide to the Extraction of Fossil Diatoms from
Lithified or Partially Consolidated Sediments. Micropaleontology, 40(4), 368.
Horowitz, A. S., Ringer, G. R., & Potter, P. E. (2012). Introductory Petrography of
Fossils. Berlin, Germany: Springer Berlin Heidelberg.
International Association of Wood Anatomists. Committee on Nomenclature.1964.
Multilingual glossary used in wood anatomy. Verlag Sannstalt Buchdruckerei Konkodie
Winterthur, p. 3-186.
J. V. Hinchey, O. R. Green 1994. A guide to the extraction of fossil diatoms from
lithified or partially consolidated sediments. Micropaleontology.
Kennard, J.M. and Burne, R.V. (1989) Stromatolite Newsletter Number 14. Bureau of
Mineral Resources, Geology and Geophisics, Camberra, 179 pp.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
72 Bibliografía
Lagabrielle, Y.; Suárez, M.; Rosselló, E.A.; Hérail, G.; Martinod, J.; Régnier, M.; De
la Cruz, R. 2004. Neogene to Quaternary tectonic evolution of the Patagonian Andes at
the latitude of the Chile triple junction. Tectonophysics 385: 211-241.
Lewis, T., 1993. Tree Rings as Indicators of Ecosystem Health. Penn State
University. United States of America.
Lio G, Novas F, Salgado L, Suárez M, De La Cruz R (2011) First record of a non-
marine Crocodylomorph (Archosauria) from the Upper Jurassic of Chile. IV Congreso
Latinoamericano de Paleontología de Vertebrados, San Juan, Argentina.
Logan, Brian W., 1961, Cryptozoon and associate stromatolites from the Recent of
Shark Bay. Western Australia: Jour. Geol., v.69, n.5, p.517-533, figs. 1-3, pls. 1, 2.
Logan, B.W., Rezak, R. and Ginsburg, R.N. (1964) Classification and environmental
significance of algal stromatolites. J. Geol., 72, 68–83.
Martin‐Bello, L., Arenas, C. and Jones, B. (2019), Lacustrine stromatolites: Useful
structures for environmental interpretation – an example from the Miocene Ebro Basin.
Sedimentology.
Mörner, N.A., Sylwan, C., 1989. Magnetostratigraphy of the Patagonian moraine
sequence at Lago Buenos Aires. Journal of South American Earth Sciences 2, 385–
389.
Niemeyer, H. 1975. Geología regional del territorio comprendido entre el lago General
Carrera y el río Chacabuco, Provincia de Aysén, Chile. Memoria de grado.
Departamento de Geología, Universidad de Chile, Santiago.
Pankhurst, R.J. & Rapela, C.W. 1995. Production of Jurassic rhyolite by anatexis in
the lower crust of Patagonia. Earth and Planetary Science Letters. Vol. 134. p. 23-36.
Pankhurst, R.J., Leat, P.T., Sruoga, P, Rapela, C.W., Márquez, M., Storey, B.C. &
Riley, T.R. 1998. The Chon-Aike silicic igneous province of Patagonia and related rocks
in West Antarctica: a silicic LIP. Journal of Volcanology and Geothermal Research. Vol.
81. p. 113-136.
Pankhurst, R.J., Riley, T.R., Fanning, C.M. & Kelley, S.P. 2000. Episodic silicic
volcanism in Patagonia and the Antarctic Peninsula: chronology of magmatism
associated with the break-up of Gondwana. Journal of Petrology. Vol. 41. p. 605-625.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
73 Bibliografía
Parrish, J.T. y Spicer, R.A. 1988. Middle Cretaceous wood from the Nanushuk Group,
Central North Slope, Alaska. Palaeontology 31: 19-34.
Philippe, M. y Bamford M. K. 2008. A key to morphogenera used for Mesozoic conifer-
like woods. Review of Palaeobotany and Palynology148: 184-207.
Riccardi, A., 1971. Estratigrafía en el oriente de la Bahía La Lan-cha, Lago San Martín,
Santa Cruz, Argentina. Revista Museo de la Plata (nueva serie). Tomo 7, 61.
ROIG, F. 1992. Comparative wood anatomy of Southern South American
Cupressaceae. IAWA Bulletin13:151-162.
Round, F.E., Crawford, R.M. and Mann, D.G. (1990) The Diatoms. Biology and
Morphology of the Genera. Cambridge University Press, Cambridge, 747 pp.
Salgado, L., De La Cruz, R., Suárez, M., Fernández, M., Gasparini, Z., Palma-Heldt,
S., & Fanning, M. (2008). First Late Jurassic dinosaur bones from Chile. Journal of
Vertebrate Paleontology.
Scherer, R., Gladenkov, A., & Barron, J. (2007). Methods and Applications of
Cenozoic Marine Diatom Biostratigraphy. The Paleontological Society Papers, 13, 61-
83.
Scholle, P. A., & Ulmer-Scholle, D. S. (2003). A Color Guide to the Petrography of
Carbonate Rocks: Grains, Textures, Porosity, Diagenesis, AAPG Memoir 77 (Vol. 77).
SCHWEINGRUBER, F.H., ECKSTEIN, D., SERRE-BACHET, F., BRÄKER, O.U. 1990.
Identification, presentation and interpretation of event years and pointer years in
dendrochronology. Dendrochronologia 8:9-39.
SCHWEINGRUBER, F.H. 1996. Tree Rings and environment, Dendroecology. Swiss
Federal Institute for Forest, Snow, and Landscape Research, Birmensdorf. 609pp.
Suárez, M. 1978. Carta Geológica de Chile Nº 36. Región al Sur del Canal Beagle.
Región de Magallanes y Antártica Chilena, escala 1 : 500.000. Santiago. 48 pp.
Suárez, M.; De La Cruz, R. 1994. Estratigrafía y paleogeografía Mesozoica de
Aysén Nororiental (45°- 46°S), Chile. In Congreso Geológico Chileno, No.
7, p. 533-537. Concepción.

UNIVERSIDAD ANDRES BELLO FACULTAD DE INGENIERIA
74 Bibliografía
Suárez, M., De La Cruz, R., Troncoso, A. 2000 Tropical/subtropical upper paleocene-
lower eocene fluvial deposits in Eastern Central Patagonia, Chile (46°45’S). Journal
South American Earth Sciences, 13, 527-536.
Suárez, M., De La Cruz, R. and Bell, M. 2007. Geología del área Ñireguao-Baño
Nuevo, Región de Aisén del General Carlos Ibáñez del Campo, escala 1:100.000. Carta
Geológica de Chile, Serie Geología Básica , Nº 108. Servicio Nacional de Geología y
Minería, Santiago, Chile.
Suárez, M.; De La Cruz, R.; Aguirre-Urreta, B.; Fanning, M. 2009. Relationship
between volcanism and marine sedimentation in Northern Austral (Aysén) Basin, central
Patagonia: Stratigraphic, U-Pb SHRIMP and paleontological evidence. In Journal of
South American Sciences, Vol. 27, p. 309-325.
Suárez, M., Cruz, R. D., Fanning, M., Novas, F., & Salgado, L. (2015). Tithonian age
of dinosaur fossils in central Patagonian, Chile: U–Pb SHRIMP geochronology.
International Journal of Earth Sciences, 105(8), 2273-2284. doi:10.1007/s00531-015-
1287-7
Torres, Teresa, & Philippe, Marc. (2002). Nuevas especies de Agathoxylon y
Baieroxylon del Lías de La Ligua (Chile) con una evaluación del registro paleoxilológico
en el Jurásico de Sudamérica. Revista geológica de Chile, 29(2), 151-165.
Tortorelli, L.A.1956. Maderas y Bosques Argentinos. Editorial ACME, Buenos Aires.
910pp.
Rashidi, K., Schlagintweit, F., Cymopolia eochoristosporica Elliott, 1968 (green alga,
Dasycladale) from the upper Maastrichtian of the Tarbur Formation (SW Iran),
Cretaceous Research (2017).
Wehr, J.D. and Sheath R.G., 2003: Freshwater Algae of North America: Ecology and
Clasification. Academic Press.





























