-
8/2/2019 Paper 1 10
1/30
The Microclimates and Characteristics of Roost Buildings of Bats
(Chiroptera) In Urban
Area of West Coast Of Sabah
Ehsan Hadi1, Nurul Najmaini1, Abdul Hamid Ahmad1 and Azniza
Mahyudin1
1Institute for Tropical Biology and Conservation, Universiti
Malaysia Sabah.
Abstract
This study was design to identify species of bat that roost in
building and to collect information
about the characteristics of the roosts in urban area of West
Coast of Sabah. Bats were found
opportunistically and examined in six buildings with a total of
11 roosting sites. Three species of
bats were identified in the buildings, namely: 1) Taphozous
melanopogon; 2) Taphozous
saccolaimus and 3) Scotophilus kuhlii. Among these three,
Taphozous melanopogon is the most
abundant species. Bats were found roost in four types of
buildings; commercial, mosque, tower
and residential house. They roost on walls, attics, crevice and
ceiling. Generally, rooting sites are
situated high from ground and relatively covered from weather.
Roosting sites also found located
within radius 1 km distance from water resource and forest
remnant. At the temperatures scale,
roosting sites are characterized by warm temperature. However,
the temperatures variables inside
roosting sites were varied suggest that structure of buildings
influenced the temperature. The
roosting sites ofTaphozous melanopogon and Taphozous saccolaimus
were significantly warmer
than Scotophilus kuhlii. Despite that, the temperatures occurred
in Taphozous melanopogons and
Taphozous saccolaimusroosting siteswere relatively similar.
Keywords: Bats; Urban; Roost building; Roost characteristics;
Roost temperature
1
-
8/2/2019 Paper 1 10
2/30
1. Introduction
Bats utilize a wide variety of structures as roost. Naturally
they use trees, caves and overhangs.
However, in urban environment where the natural roosts are
scarce, bats might opt for many
different types of roost including man-made structures such as
bridge, tower, attic and even the
hollow floor spaces of house as substitutes for tree cavities or
caves (Altringham, 1996). Among
those structures, the used of building as roost by bats is the
most well-documented (Riskin and
Pybus, 1998, Jenkins et. al, 1998, Kunz, 1982). During the past
40 years, evidences that certain
species of bats is well adapted to building were recorded in
increasing amount of literatures. Many
species such as Taphozous melanopogon, Eptesicus fuscus and
Pipistrellus pipstrellus whose
originally roost in the caves has been link almost exclusively
to building (Kunz, 1982).
The roost selection in bats involves many mutually important
factors. Generally, bats choose area
that has lower predation risk, reduced ectoparasite infection,
higher from ground (Psyllakis and
Brigham, 2006), and relatively close to water resource (Evelyn
et al., 2004) and foraging area
(Kunz, 1982). Thermal stability also one of the most important
factor in bats roost selection.
Lausen and Barclay (2006) explained the main reason for bats
occupied human settlements is
because of thermal advantages where warm temperatures can reduce
energy expenditure and also
offer benefit in term of productivity.
Despite the consistency of published studies, there is still
paucity of roosting characteristics for
many species especially around human settlement (Evelyn et al.,
2004). Besides, far too little
attention has been paid to the roosting information of bats in
tropics region (Rodriguez-Duran and
Soto-Centeno, 2003). Many studies have been conducted but mainly
in temperate countries. Recent
2
-
8/2/2019 Paper 1 10
3/30
research has identified roost as the most crucial element in
bats conservation. Therefore, such
information is urgently needed as the bats remain unprotected in
many parts in the world.
This research was conducted to identify the species of bats that
occupy the buildings in
urban area of Kota Kinabalu, West Coast of Sabah. This study
also examines the
characteristics and microclimate of the roost. More
understanding about the animals needs and
their behaviour may be able to provide solution for human-bats
conflicts since bats always
regarded as nuisance and frequently persecuted.
2. Material and Methods
Random surveys were conducted from April 2008 to June 2008 to
locate possible bats roosting site
in Kota Kinabalu. Residents, or workers of buildings were
interviewed to get the information about
bats occurrence. Positive feedback was then followed by
examination of the roosting sites to
confirm the existence of bats in the buildings.
2.1 Buildings Characteristics
Seven variables were used to characterize roost site:
temperature (C); building types, i.e house,
factory, commercial/ species (if trees); Wall material (wooden,
bricks etc), canopy cover (%);
distance from brackish/ fresh water source (m); distance from
forest remnant (m); roost height
from the ground (m).
3
-
8/2/2019 Paper 1 10
4/30
2.2 Microclimate Variables
The microclimate of the sites was described with temperature
used as a measurement.
Microclimate was recorded using data loggers (Onset HOBO H8 Pro
series, temperature range
-30C to +50C). To reduce disturbance to roost, the data loggers
were positioned 1 to 3 m away
from roost.
Data were averaged per hour and categorised into hourly
intervals over each period of 24-h period.
To calculate variables that would show variation and pattern of
24-h temperatures, the hourly data
were used to calculate: average temperature, temperature
fluctuation (hourly); maximum;
minimum; range of day; time of day maximum temperature occurred;
and length of time maximum
temperature of a day was maintained. The readings for each
roosting sites were then compared.
2.3 Data Analysis
Each variable was tested for normality using Kolmogorov-Smirnov
test and most of variables were
not normally distributed and could not be transformed
successfully. Thus non-parametric tests
were used in this study.
To test differentiation of temperatures between roosting sites,
Kruskal-Wallis One Way Analysis
of Variance test was performed on five temperatures variables:
average temperature, fluctuation,
range of temperature, time of maximum temperature occurs and
length of time maximum
temperature was maintained.
4
-
8/2/2019 Paper 1 10
5/30
Each of temperatures variables had expected to be different
between roosting sites (at least one).
The differences were suspected to be influenced by the buildings
construction materials.
Therefore, a comparison using Mann Whitney U-Test (p
-
8/2/2019 Paper 1 10
6/30
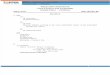
Figure 1: Roost buildings in urban area of Kota Kinabalu, West
Coast of Sabah
3.1 Roost Buildings
All roost buildings are older than 10 years. WLLT, WM and WK are
classified as commercial
building, CM and SM as place for worship, while THC is a
residential house. A higher proportion
of the 11 specific roosts were found in wall (45.5%), others in
attic (36.4%), crevices (9%) and
ceiling (9%). All roost was communal and no solitary roost was
found. Roosts are high from
ground (med=16.90), highly covered (med=100.00) and located
within 1 km from water
(med=116.9) and forest remnants (med=268.89). Interestingly, all
roost is surrounded by open
space. Table 1 described the physical characteristics of the
buildings.
Table 1. The characteristics of the roost buildings found in
Kota Kinabalu.
6
-
8/2/2019 Paper 1 10
7/30
Roost sites Roost
type
Species Roost
height
Entrance
direction
Canopy
cover
Distance
from water
Distance
from forest
CM1 Wall T. melanopogon 5.6 SE 100 26.35 556.58
CM2 Wall T. melanopogon 9.3 SE 100 30.35 556.58
CM3 Ceiling T. melanopogon 45.4 SE 100 48.64 575.38
CM4 Wall T. melanopogon 20 SE 100 47.01 592.18
WK1 Wall T. melanopogon 21.6 SE 100 343.95 76.48
WK2 Wall T. melanopogon 16.9 SE 100 343.95 72.24
WM Crevice T. melanopogon 4.5 SE 40 872.91 268.89
SM Attic T. melanopogon 67.6 SE 80 297.46 574.74
WLLT AtticT. melanopogon
18 SE 100 30.17 260.75THC1 Attic T. saccolaimus 6.3 NW 100 219
33
THC2 Attic S. kuhlii 7.1 SW 100 222 33
SE=southeast, NW=Northwest and SW=southwest.
3.2 Temperatures
Overall, most site recorded temperature above 27C. Highest
temperature was 51.04C while the
lowest was 23.24. Kruskal-Wallis Analysis showed that there were
some variations of
temperatures variables between roosting sites. Among all roost,
CM1 was regarded as most stable
site as it have lowest range (med=1.98) and maintained highest
temperature longer (med=7:00)
(Table 2).
3.3 Concrete Buildings versus Wood
The temperature inside concrete roost buildings were more stable
than wood building. The
temperature in concrete were fluctuated slower than wood. The
range of temperature occurs in
concrete buildings were significantly lower and minimum
temperature during night were
7
-
8/2/2019 Paper 1 10
8/30
significantly warmer than wood building. Concrete roost
buildings also reached maximum
temperature earlier and it can hold up maximum temperature much
longer compare to wood
structures. Although, mean rank of temperature and maximum
temperature during daylight
(between these two sites) was not significantly differed (Table
3).
Table 2. Comparison of Temperatures Variable Between Roosting
Sites.
Temperature Fluctuation Range LMT TMO
CM1 27.91 (0.79) 0.30 (0.41) 1.98 (0.78) 7:00 (10:00) 11:00
(3:00)
CM2 27.91 (1.43) 0.32 (0.44) 2.36 (2.02) 5:00 (4:15) 11:00
(2:00)
CM3 27.5 (6.81) 1.18 (1.36) 23.17 (6.44) 1:00 (1:00) 10:00
(0:15)
CM4 27.91 (1.12) 0.25 (0.32) 2.36 (0.41) 5:00 (5:15) 12:00
(2:00)
WK1 28.31 (2.58) 0.75 (0.88) 4.80 (1.41) 3:30 (2:15) 12:00
(1:15)
WK2 26.67 (3.75) 0.72 (1.00) 6.73 (1.56) 2:00 (3:00) 12:00
(2:00)
THC1 27.85 (4.62) 0.65 (0.94) 8.24 (2.58) 2:00 (2:00) 11:00
(3:15)
THC2 27.39 (4.8) 0.72 (0.98) 8.47 (3.43) 2:00 (1:15) 13:00
(3:00)
WM 27.33 (4.29) 0.83 (1.42) 7.91 (2.11) 3:00 (3:00) 10:00
(3:15)
SM 27.56 (2.1) 0.70 (0.72) 3.57 (1.58) 4:00 (4:15) 13:00
(3:15)
WLLT 29.5 (1.61) 0.53 (0.70) 2.40 (1.19) 6:00 (4:15) 13:00
(3:00)
H 728.41 50.49 268.40 110.43 83.29
df 10 10 10 10 10
P value p
-
8/2/2019 Paper 1 10
9/30
3.5 Species Roost Temperature
Overall, roost temperature ofS. kuhlii is significantly lower
than the other two species. Despite
that, the roost temperature ofT. melanopogon and T. saccolaimus
was not significantly differed
(Table 3).
4. Discussion
9
Table 3. Comparison of temperatures variables of construction
materials and species
Materials Concrete Wood U p value Effect size (R )
Day (Max) 29.10 (2.40) 31.50 (4.47) 4494196 ns -0.014
Night (min) 27.12 (1.57) 26.34 (1.56) 4400301 p
-
8/2/2019 Paper 1 10
10/30
In this study, roosts were found exclusively in buildings that
shared common characteristics. All
roost building aged more than 10 years and historically, showed
no sign of major modification.
The walls were not smooth with some cracks; allowing bats to
cling. Bats accessed the interior
buildings through windows, doors or attic which were opened all
the time. Moreover, the existence
of ruined rough bricks or concrete which comparable to crevices;
create a good roosting habitat for
bats.
4.1 Species distribution
Only three species of bats were found in this study. The low
number of species is expected as
species richness, abundance and activity of bats were reduced in
disturbed habitats (Hourigan et
al., 2006; Lumsden and Bennett, 2002; Kirsten and Klomp, 1998).
The species in urban ecosystem
are also dominated by one or a few abundant species
(Furmankiewicz and Grniak, 2002). This is
because not many species of bats have the ability to survive in
open and edge habitat. Therefore,
the community structure of bats may have changed; particularly
for microchiropteran which have
narrower and specialize habitat requirements. This may also
explain why the number of species
found in buildings was low (See Hourigan et al., 2006; Kurta and
Teramino, 1992).
T. melanopogon was the most abundant species found in this
research (>800). Discovered roost
buildings also dominated by this species revealed that T.
melanopogon has successfully adapted to
the current environment in this city. Roost buildings which were
occupied by T. melanopogon
were found distributed along coastal area. This record was
different from study of Bates and
Harrison (1997) in India, which found T. melanopogon occur near
fresh water areas such lakes,
rivers and ponds.
10
-
8/2/2019 Paper 1 10
11/30
Conversely, T. saccolaimus and S. kuhlii showed a restrictive
distribution in this study. This
finding is in contrast to what was found by previous studies.
Both species are intimately associated
with modified habitats especially S. kuhlii which were known to
occur in all type of mosaic
particularly urban or suburban parklands or rural areas (Pottie
et al., 2005). A possible explanation
for this might be that both species prefer to roost in rural
area rather than sub urban or urban
mosaic. This result also fit with population status of this
species which is known to be in low
density in Sabah (Yasuma et al., 2003).
4.2 Roost Characteristics
The differences in roost distribution revealed that there are
might be some flexibility and
preferences behaviour among these species. However, the roost
characteristics might be
underestimated forT. saccolaimus and S. kuhlii because of the
small sample size. Nevertheless, in
general rooting sites are located high from ground and
relatively covered from weather. The
temperatures in all roosting sites were also warm. Roosting
sites also found located within radius 1
km distance from water resource and forest remnant.
4.3 Temperature
Overall, all species found in this study roost in warm roosting
site. Preference of bats toward
warmer temperature has been observed in several studies in other
tropical area (Table 4).
However, the factors guiding bats in this study to roost in warm
site are poorly known. To date, for
other species high roost temperatures offer several advantages
to bats particularly reproductive
females. Roosting in warmer roost enables reproductive females
to save energy maintaining their
body temperature and use them for reproduction purposes. It has
been hypothesized that warm
11
-
8/2/2019 Paper 1 10
12/30
roosts accelerate embryonic development (which resulted in early
birth) and increase milk
production.
Table 4. Research investigating roost temperature in tropical
bats.
Bat species Type Temperature (C) Country Source
T. melanopogon Building 28.3 2.7 Malaysia This study
T. saccolaimus Attic 28.7 2.9 Malaysia This study
S. kuhlii Attic 28.3 3.1 Malaysia This studyP. quadridens Cave
30.2 4.6 West Indies Rodriguez-Duran & Soto
Centeno (2003)E. sezerkoni Cave 28.2 2.3 West Indies
Rodriguez-Duran & Soto
Centeno (2003)L. silvicolum Termite nest 27.9 1.0 Panama
Dechmann et al., (2004)
C. brevicauda Cave 19.2 2.7 Mexico Avila Flores &
Medellin
12
-
8/2/2019 Paper 1 10
13/30
(2004)
B. plicata Cave 26.7 3.1 Mexico Avila Flores & Medellin
(2004)
A. jamaicensis Cave 21.9 2.6 Mexico Avila Flores &
Medellin
(2004)
E. alba Tents 23.3 0.7 Costa Rica Rodriguez-Herera et al.,
(2008)Temperatures data presented in mean (SD).
The high temperature also benefits young bats which had low
thermoregulating ability. During
early birth, when their mother was away warm roosts help them
maintain their body temperature.
When they are able to regulate their body temperature, warm
temperature lower the
thermoregulatory cost and will increase their growth rates
(Lausen and Barclay, 2006; Kerth et al.,
2001). In temperate, this strategy was used to increase winter
survival (Kunz, 1982).
4.3.1 Thermal Variation among Roost
The variation in microclimate found among roost buildings (Table
2) indicated that temperature
was influenced by the diverse microstructures or substrates of
buildings; such as crevices, walls,
attics, wood, bricks or other features of the buildings
architecture. Some structures may expose
more to solar radiation or can retain heat longer than others.
Besides, the ventilation system also
contributed to temperature fluctuation in the buildings. The
existence of vent-holes and peripherals
such as ventilator could avoid heat entrapment.
The bats also were most common roost in concrete buildings. One
reason bats dwelled in this type
of buildings might be because of the compacted stable
temperature and ability to retain heat longer.
13
-
8/2/2019 Paper 1 10
14/30
Similarly, the bats preferences for bricks or concrete
structures have been seen in several past
studies (Lausen and Barclay, 2006; Lance et al., 2001). In
Sweden during winters, bats occupied
concrete bat boxes more common than wooden. The research of
exposed roost in Alberta also
found that most bats roost on bricks which suggest bricks or
concrete may have higher capacity of
trapping heat (Riskin and Pybus, 1998).
4.3.2 Temperatures Difference among Species
Microclimate variation also can encourage colonization by
several species (Hill and Smith, 1985)
and may influence pattern association of bats in certain roosts
(Rodriguez and Soto-Centeno,
2003). Despite wide range of temperatures recorded in many roost
buildings, only T. saccolaimus
and Scotophlus kuhlii were found sharing roost. The reason
anyhow might be related to the
condition site of study which concentrated around urban
environment that known to have limited
number of species and the limited number of available
roosts.
Roost selection is a complex processes. It influenced by many
factors that mutually dependent
between each other. Rodriguez-Duran (1995) suggested the
occurrence of certain species in a
given cave was determined by physiology of the bats which
influenced by body mass, diet and
microclimate condition. For example, Baudinette (2000) found
microchiropterans species in a cave
in Australia preferred different temperatures and humidities.
Similarly, the study of bats in West
Indies also revealed that species ofPteronotus quadridens which
is smaller species, roost in
warmer site compare to Erophylla sezekorni although they roost
in the same cave (Rodriguez-
Duran and Soto-Centeno, 2003).
14
-
8/2/2019 Paper 1 10
15/30
Suprisingly, the result of this study was inconsistent with the
previous studies. S. kuhlii which is
smaller, roost in cooler site compare to T. saccolaimus and T.
melanopogon. The inconsistency
maybe due to morphological features ofS. kuhlii. This species
longer tail is probably significant in
conserving heat as seen in Lasiurus borealis (Davis and Reite,
1967). The comparison of other
temperatures variables also showed that T. saccolaimus is
closely related to S. kuhlii. This
similarity maybe explained by the fact that they shared common
preference for roost type; that is in
attic and hollow trees (Yasuma et al., 2003).
4.4 Canopy Cover and Surrounding Areas
The finding of this study indicated most roosting sites are
situated in highly covered area (average
92.7 %). Most bats roost under roof except WM which is located
in the narrow slit between two
buildings. In natural roosting site, several other bat species
also have been shown to roost
preferentially in highly covered roost (Table 5).
Table 5. Research investigating roost canopy cover in tropical
bats and other part of the world.
Bat species Canopy cover (%) Country Source
T. melanopogon 91.1 (n=9) Malaysia This study
T. saccolaimus 100 (n=1) Malaysia This study
S. kuhlii 100 (n=1) Malaysia This study
E. alba 83.1 (n=50) Costa Rica Rodriguez-Herera et al.,
(2008)
M. molossus 81 (n=5) Australia Aguirre et al., (2003)D. rotundus
62.7 (n=7) Bolivia Aguirre et al., (2003)
N. albiventris 95 (n=1) Bolivia Aguirre et al., (2003)
T. georgianus 53.5 (n=16) Australia Milne et al., (2005)
H. diadema 48.5 (n=1) Australia Milne et al., (2005)
H. ater 48.5 (n=6) Australia Milne et al., (2005)Data presented
as mean, (n=number of roost).
15
-
8/2/2019 Paper 1 10
16/30
A highly covered roost can protect bats from extreme weathers
and avian predators. It also
provides bats with more stable temperature (Kunz, 1982). The
finding of this study seems to be
consistent with the theory. The temperatures fluctuations of
most roosting site were very low with
an average 0.51 C. Only CM3 were considered as less stable with
high range of temperature
(Table 2).
On the other hand, the surrounding roosts are open areas. The
reasons of this preference are not
fully understood. Studies for other species such as Chalinolobus
tuberculatus, Eptesicus fuscus and
Myotis bechsteinii suggest the main reasons why bats prefer open
space were due to their
ecomorphology, predator avoidance, ease at roost relocation and
thermal benefits (Vonhof and
Gwilliam, 2007; Psyllakis and Brigham, 2006; Sedgeley and
ODonnell, 1999).
Species that was found in this study is consistent with
ecomorphological theory. All three species
in this study have been described as having high wing loadings
(Yasuma et al., 2003). The
morphology allows these species to fly fast in open areas, above
treetops and along forest gaps.
However, this feature reduced the flight maneuverability and may
limit bats from accessing
cluttered habitats (Wei et al., 2008). Therefore, roosts which
are far from ground in open spaces
would facilitate bats that lack maneuverability (Sedgeley and
ODonnell, 1999).
Open spaces also allow bats to detect predators and this area
most likely receive more direct
sunlight compared to dense area. Exposure to solar radiation
produced warmer temperatures which
16
-
8/2/2019 Paper 1 10
17/30
could benefit bats in many ways (Perry et al., 2007). The
surrounding habitats might be the cause
dictating the temperatures in the roosting sites.
4.5 Higher from ground
Another important finding is those roosts located high from the
ground (at least 4.5 m above the
ground). This characteristic was reviewed as one of main
important variables in roost selection
(Agosta, 2002). High roost is commonly selected by bats because
it can avoid terrestrial predators
such as snake, rats and etc. However, in urban area, cats and
dogs are more abundant.
Domestic animals were known to prey bats in many studies
(Sedgeley and ODonnell, 2004;
Riskin and Pybus, 1998). In this study, these animals were found
abundant in the study sites. Cats
were also seen tried to catch bats when they forage close to
ground and during bats-released from
mist nets in THC. Taller roost also accommodates bats to gain
velocity during early flight. This
roosts characteristic is particularly important for bats that
are in less maneuverable conditions
such as pregnant adults, adults moving non-volant young and
young bats during their first flight
(Norberg and Rayner, 1987).
Roosts that have greater height from ground are also more
exposed to solar radiations which offer
warmer condition to bats (Russo et al., 2004). The preference of
bats for high roost to gain
microclimate advantage by choosing higher roost was confirmed in
this study which was observed
in T. melanopogon in CM3. Many other research finding also has
showed preference of bats to
roost in high roosting site. However, it is interesting to note
that roost height in this study was
unusual. Mean roost height found is 20.2 m and the maximum roost
height recorded is 67.6 m;
both exceeds those all recorded in other published studies
(Table 6).
17
-
8/2/2019 Paper 1 10
18/30
Table 6. Research investigating roost height in tropical bats
and other part of the world.
Bat species Type Roost height (m) Country Source
T. melanopogon Building 23.2 (n=9) Malaysia This study
T. saccolaimus Attic 6.3 (n=1) Malaysia This study
T. saccolaimus Tree 7 (n=1) Australia Schulz & Thomson
(2007)
S. kuhlii Attic 7.1 (n=1) Malaysia This study
S. kuhlii Building 12 (n=1) Philippines Rickert et al.,
(1989)
E. alba Tree 1.3 - 2.6 (n=26) Costa Rica Timm & Mortimer
(1976)
S. lilium Tree 2.1 to 7.9 (n=8) Belize Fenton et al., (2000)
E. furinalis Tree 9.5 (n=2) Bolivia Aguirre et al., (2003)
N. albiventris Tree 8.4 (n=1) Bolivia Aguirre et al., (2003)
N. noctula Building 7.5 (n=131) Hungary Bihari (2004)
A. lituratus Tree 8.1 (n=15) Venezuela Munoz-romo et al.,
(2007)
Data presented as mean, (n=number of roost).
4.6 Distance from Water Source
Being close to permanent water resource is always associated
with roost selection by many species
of bats (Rabe et al., 1998; Entwistle et al., 1997). The finding
of this study seems to be consistent
with other research. Compare to other studies, roosting sites in
this study are located relatively
close to water (
-
8/2/2019 Paper 1 10
19/30
T. melanopogon 0.23 (n=9) Malaysia This study
T. saccolaimus 0.22 (n=1) Malaysia This study
S. kuhlii 0.22 (n=1) Malaysia This study
Myotis spp 0.48 (n=22) Canada Jung et al., (2004)
B. barbastellus 1.79 (n=33) Italy Russo et al., (2004)
M. yumanensis 0.13 (n=20) USA Evelyn et al., (2004)T. georgianus
2.7 (n=16) Australia Milne et al., (2005)
T. kapalgensis 6.6 (n=4) Australia Milne et al., (2005)
H. diadema 1.6 (n=1) Australia Milne et al., (2005)
H. ater 1.6 (n=1) Australia Milne et al., (2005)Data presented
as mean, (n=number of roost).
Roost near water body offers bats benefit in several ways.
First, the insect availability is usually
abundant of near water. Hence, by roosting near water, bats are
close to prospective foraging
location (see Evelyn et al., 2004). Second, waterway provides
bats open a flight passage which is
benefit most fast flier bat. However, water bodies that lack of
vegetation are often avoided by bats.
It is believed that loss of buffer zone resulted in loss of
invertebrate diversity and abundance (see
Russ and Montgomery, 2002).
In this study, all three species are described as fast fliers
with high wing loadings and high aspect
ratios. It also means these species lack maneuverability.
Therefore, the water bodies might be
useful as foraging site for these species as they prefer to
forage in open space. However,
exploitation of water body as foraging site only has been seen
in T. melanopogon near CM and
WLLT.
4.7 Distance from forest
To date, distance from forest plays a major role in roost
selection. In general closer to forest offer
several benefits to roost dwellers. With more complex structure
of vegetation, forest remnants offer
19
-
8/2/2019 Paper 1 10
20/30
bats variation and abundant of insects. Even small or poor
quality remnants harbour resource for
bats particularly where large forests are rare (see Law et al.,
1999). Therefore, by being close to
forest bats could reduce energetic consumption during flight to
the foraging sites (Jenkins et al.,
1998).
Roosting near forest also may protect bats from avian predators.
The presences of trees in forest
allow bats to emerge with low detection of predators (Russo et
al., 2007). There is several other bat
species have been shown to roost preferentially near forest
remnant (Table 8).
Table 8. Research investigating distance of roost from forest
resource in other studies.
Bat species Distance from forest (m) Country Source
T. melanopogon 392.6 (n=9) Malaysia This study
T. saccolaimus 33 (n=1) Malaysia This study
S. kuhlii 33 (n=1) Malaysia This study
C. tuberculatus 15 (n=1) New Zealand Sedgeley & O' Donnell
(1999)Mystacinidae sp 220 to 530 (n=8) New Zealand O' Donnell et
al., (1999)
M. sodalis 14.2 (n=47) USA Carter & Feldhamer (2005)
Data presented as mean, (n=number of roost).
During investigating period, T. melanopogon were seen forage in
open space near forest edge in
two places: WK and WLLT while S. kuhlii also use the same space
in THC. Unfortunately, this
20
-
8/2/2019 Paper 1 10
21/30
study was unable to trace foraging site of T. saccolaimus. T.
melanopogon and S. kuhlii which
known to forage in open habitats also seems cant utilized
benefits of forest remnants to its fullest.
These two species might be gain advantage of food supply at the
corridor but, they are most likely
not covered from predators because they are open space flier.
The protection might just apply to
low wing loadings bats that have slow flight but better
maneuverability (e.g, Russo et al., 2007).
5. Conclusion
Taken together, these results suggest that bats choose roost
based on warmer temperature, high
level from ground and high percentage of canopy cover. An
implication of this is the possibility
that any renovations done on the buildings should not change the
microclimate. Human activities
near roost area also at least must not disturb the animals. The
current findings add substantially to
our understanding that bats need certain type of roost which
influences most aspect of their life.
This research will serve as a base for future studies and more
investigations are needed.
Knowledge of bats metabolic rates relative to warmer roost will
help to answer the questions that
unsolved in this study. Further research using radiotelemetry
also could help tracking the bats and
consolidate the results of this study.
Acknowledgement
We gratefully acknowledge and thank Mr. Hairul Hafiz Mahsol for
his statistics advice. We thank
Kota Kinabalu Health Office and manager of Wisma Merdeka for
giving us permission to conduct
21
-
8/2/2019 Paper 1 10
22/30
this research in their buildings. We also thank staff of City
Mosque especially Hj. Gustin, Hj. Asli,
Abdullah, Mr. Mudim and Mr. Rosli for their incredible assistant
all through the studies that we
did there. Further thank people who helped us during the
fieldwork in Telipok Health Centre and
Wisma Kosan.
References
Altringham J D. 1996.Bats biology and behaviour. Oxford
University Press, United State.
Agosta S J. 2002. Habitat use, diet and roost preferences of the
big brown bats (Eptesicus fuscus)
in North America: a case of conserving an abundant
species.Mammal Review 32: 179-198.
Aguirre L F, Lens L, Matthysen E. 2003. Patterns of roost use by
bats in a neotropical savanna:
implications for conservation.Bulletin of the American Museum of
Natural History111: 435-443.
Avila Flores R, Medellin R A. 2004. Ecological, taxonomic and
physiological correlates of cave
use by the mexican bats.Journal of Mammalogy85(4): 675-687.
22
-
8/2/2019 Paper 1 10
23/30
Barclay R M R. 1991. Population structure of temperature zone
insectivorous bats in relation to
foraging behaviour and energy demand.Journal of Animal Ecology
60: 165-78.
Bates P J J, Harrison D L. 1997. Bats of the Indian
Subcontinent. Harrison Zoological Museum
Publications, 258.
Baudinette RV, Churchill S K, Christian K A, Nelson J E, Hudson
P J. 2000. Energy, water
balance and the roost microenvironment in three australian
cave-dwelling bats (microchiroptera).
Journal of Comparative PhysiolologyB 170: 439446.
Bihari Z. 2004. The roost preference ofNyctalus noctula
(Chiroptera, Vespertilionidae) in summer
and the ecological background of their
urbanization.Mammalia68(4): 329-336.
Carter T C, Feldhamer G A. 2005. Roost tree use by maternity
colonies of indiana bats and
northern long-eared bats in southern Illinois.Forest Ecology and
Management219: 259-268.
Chruszcz, B. J., and Barclay, R. M. R., 2003. Prolonged foraging
bouts of a solitary gleaning /
hawking bat, Myotis evotis. Convergence, 826, 823-826.
Davis W H, Reite O L. 1967. Responses of bats from temperate
regions to changes in ambient
temperature.Biological Bulletin 132: 320328.
23
-
8/2/2019 Paper 1 10
24/30
Dechmann D K N, Kalko E K V, Kerth G. 2004. Ecology of an
exceptional roost: energetic
benefits could explain why the bat Lophostoma silvicolum roosts
in active termite nests.
Evolutionary Ecology: 1037-1050.
Entwistle A C, Racey P A, Speakman J R. 1997. Roost selection by
the brown long-eared bat
Plecotus auritus.Journal of Applied Ecology Vol. 34 (2):
399-408
Evelyn M. 2004. Conservation of bats in suburban landscapes:
roost selection by Myotis
yumanensis in a residential area in California.Biological
Conservation115(3): 463-473
Fenton M B, Vonhof M J, Bouchard S, Gill S A, York N, Johnston D
S, Reid F A, Riskin D K,
Standing K L, Taylor J R, Wagner R. 2000. Roosts used by
Sturnira lilium
(Chiroptera:Phyllostomidae).Biotropica32: 729-733.
Furmankiewicz J, Grniak J. 2002. Seasonal changes in number and
diversity of bat species
(Chiroptera) in the Stolec mine (SW Poland).Przyroda Sudetw
Zachodnich Suplement 2: 49-70.
Hill J E, Smith J D. 1985.Bats, A Natural History. British
Museum (Natural History), England, 53.
Hourigan C L, Johnson C, Robson S K A. 2006. The structure of a
micro-bat community in
relation to gradients of environmental variation in a tropical
urban area. Urban Ecosystems9(2):
67-82.
24
-
8/2/2019 Paper 1 10
25/30
Hutson A M, Mickleburgh S P, Racey P A. 2001. Global Status
Survey and Conservation Action
Plan Microchiropteran Bats. IUCN, Gland, Switzerland and
Cambridge, United Kingdom, 35-51.
Jenkins E V, Laine T, Morgan S E, Cole K R, Speakman J R. 1998.
Roost selection in the
pipistrelle bat,Pipistrellus pipistrellus
(Chiroptera:Vespertilionidae) in northeast Scotland.Animal
Behaviour56: 909917.
Jung T, Thompson I, Titman R. 2004. Roost site selection by
forest-dwelling male in central
Ontario, Canada.Forest Ecology and Management202(1-3):
325-335.
Kerth G, Weissmann K, Konig B. 2001. Day roost selection in
female Bechstein s bats
(Myotis:bechsteinii): a field experiment to determine the
influence of roost temperature.
Oecologia, 126(1): 1-9.
Kirsten I, Klomp N. 1998. Microchiroptera in urban, rural and
forest areas of southern NSW.
Australasian Bat
Kunz T H. 1982. Roosting Ecology of Bats. in Kunz, T. H.
(ed.)Ecology of bats. 2-3.
Kurta A, Teramino J A. 1992. Bat community structure in an urban
park.Ecography 15: 257-261.
25
-
8/2/2019 Paper 1 10
26/30
Lance R F, Hardcastle B T, Talley A, Leberg P L. 2001. Day-roost
selection by Rafinesques big-
eared bats (Corynorhinus Rafinesquii) in Louisiana Forests.
Journal of Mammalogy, 82(1): 166
172.
Lausen C L, Barclay R M R. 2006. Benefits of living in a
building: the big brown bats (Eptesicus
fuscus) in rocks versus buildings.Journal of Mammalogy87(2):
362-370.
Law B S, Anderson J, Chidel M. 1999. Bat communities in a
fragmented forest landscape on the
south-west slopes of New South Wales, Australia.Biological
Conservation: 88.
Lumsden L, Bennett A. 2005. Scattered trees in rural landscapes:
foraging habitat for insectivorous
bats in south-eastern Australia.Biological Conservation122(2):
205-222.
Milne D J, Armstrong M, Fisher A, Flores T, Pavey C R. 2005.
Structure and environmental
relationships of insectivorous bat assemblages in Tropical
Australian Savannas. Austral Ecology
30: 906-919.
Munoz-Romo M, Herrera E A, Kunz T H. 2007. Roosting behavior and
group stability of the big
fruit-eating bat Artibeus lituratus (Chiroptera:Phyllostomidae).
Mammalian Biology 73(3): 214-
221.
26
-
8/2/2019 Paper 1 10
27/30
Norberg U, Rayner J M V. 1987. Ecological morphology and flight
in bats (Mammalia:
Chiroptera): wing adaptations, flight performance, foraging
strategy and echolocation.
Philosophical Transactions of the Royal Society of London, B
316: 335427.
O Donnell F J, Christie J, Corben C, Sedgeley J A, Simpson W.
1999. Rediscovery of short-tailed
bats (mystacina sp.) In Fiordland, New Zealand : preliminary
observations of taxonomy,
echolocation calls, population size, home range and habitat use.
Society23: 21-30.
Perry R W, Thill R E. 2007. Roost selection by male and female
northern long-eared bats in a pine
dominated landscape.Forest Ecology and Management 247:
220-226.
Pottie S A, Lane D J W, Kingston T, Lee Y H. 2005. The
microchiropteran bat fauna of Singapore.
Acta Chiropterologica7(2): 237-247.
Psyllakis J, Brigham R. 2006. Characteristics of diurnal roosts
used by female Myotis bats in sub-
boreal forests.Forest Ecology and Management223(1-3):
93-102.
Rabe M J, Morrell T E, Green H, deVos J C, Miller C R. 1998.
Characteristics of ponderosa pine
snag roosts used by reproductive bats in Northern Arizona.
Journal Wildlife Management62: 612
621.
Rancourt S, Rule M, O Connell M. 2007. Maternity roost site
selection of big brown bats in
ponderosa pine forests of the Channeled Scablands of
Northeastern Washington State, USA.
Forest Ecology and Management248(3): 183-192.
27
-
8/2/2019 Paper 1 10
28/30
Rickert E A, Heideman P D, Utzurrum R C B. 1989. Tents roosting
by Scotophilus kuhlii in the
Philippines.Journal of Tropical Ecology 5: 433-436.
Riskin D K, Pybus M J. 1998. The Use of Exposed Diurnal Roosts
in Alberta by The Little Brown
Bat,Myotis lucifugus. Canadian Journal of Zoology 76,
767-772.
Rodrguez-Durn A. 1995. Metabolic rates and thermal conductance
in four species of neotropical
bats roosting in hot caves. Comparative biochemistry and
physiology. Part A, Physiology110(4):
347-55.
Rodriguez-Duran A, Soto-Centeno J A. 2003. Temperature selection
by tropical bats roosting in
caves.Journal of Thermal Biology28(6-7): 465-468.
Rodrguez-Herrera B, Medelln R A, Gamba-Rios M. 2008. Roosting
requirements of white tent-
making batEctophylla alba (Chiroptera:Phyllostomidae).Acta
Chiropterologica10(1): 89-95.
Russ J M, Montgomery W I. 2002. Habitat associations of bats in
Northern Ireland : implications
for conservation.Journal of Wildlife Management108: 49-58.
Russo D, Jones G, Migliozzi A. 2002. Habitat selection by the
Mediterranean horseshoe bat,
Rhinolophus euryale (Chiroptera: Rhinolophidae) in rural area of
Southern Italy and implication
for conservation.Biological Conservation107: 71-81.
28
-
8/2/2019 Paper 1 10
29/30
Russo D, Cistrone L, Jones G, Mazzoleni S. 2004. Roost selection
by Barbastelle Bats
(Barbastella barbastellus, Chiroptera:Vespertilionidae) in Beech
Woodlands Of Central Italy:
Consequences For Conservation.Biological Conservation117(1):
73-81.
Russo D, Cistrone L, Jones G. 2007. Emergence time in forest
bats: the influence of canopy
closure.Acta Oecologica31(1): 119-126.
Schulz M, Thomson B. 2007. National recovery plan for the
bare-rumped sheathtail bat
Saccolaimus saccolaimus nudicluniatus. Report of Department of
the Environment and Water
Resources, Canberra. Queensland Parks and Wildlife Service,
Brisbane.
Sedgeley J A, O Donnell C F J. 1999. Roost selection by the
long-tailed bat, Chalinolobus
tuberculatus, in temperate New Zealand rainforest and its
implications for the conservation of bats
in managed forests.Biological Conservation88 (2): 261-276.
Sedgeley J A, O Donnell C F J. 2004. Roost use by long-tailed
bats in South Canterbury:
examining predictions of roost-site selection in a highly
fragmented landscape. Society Newsletter
11: 28-30.
Timm R M, Mortimer J. 1976. Selection Of roost sites by honduran
white bats, Ectophylla alba
(Chiroptera: Phyllostomatidae).Ecology 57: 385389.
29
-
8/2/2019 Paper 1 10
30/30
Vonhof M J, Gwilliam J C. 2007. Intra- and interspecific
patterns of day roost selection by three
species of forest-dwelling bats in Southern British Columbia.
Forest Ecology and Management,
252(1-3): 165-175.
Wei L, Han N, Zhang L, Helgen K M, Parsons S, Zhou S, Zhang S.
2008. Wing morphology,
echolocation calls, diet and emergence time of black-bearded
tomb bats (Taphozous melanopogon,
Emballonuridae) from Southwest China.Acta Chiropterologica
Vol.10 (1): 51-59.
Yasuma S, Azniza M, Bernard H, 2005. The Bornean Mammals Vol. 2
& 3. Research and
Education Component, BBEC Programme c/o Institute for Tropical
Biology and Conservation,
Kota Kinabalu.
30