Embed Size (px)
Citation preview

J. Cell Sci. 42, 23-32 (1980) 2 , ,Printed in Great Britain © Company of Biologists Limited 1980
PARACRYSTALLINE ARRAYS IN ATYPICAL
CRISTAE AND MITOCHONDRIAL DIVISION
DORIS R. NATHANIELDepartment of Anatomy, Faculty of Medicine, University of Manitoba,Winnipeg, Manitoba, Canada, R3E 0W3
SUMMARY
Paracrystalline arrays in atypical cristae were observed in a few large mitochondria in theprocess of replication, in the follicular cell of the normal and gonadectomized rabbit thyroidgland. They were about 30 nra in diameter and spaced approximately 35-40 nm apart, andoblique sections showed parallel arrays which had a pitch of 25 nm. The number of inclusionsvaried in number and were similar to those reported in the normal follicular cell. The site ofreplication was often at the base of the mitochondrion and in some instances the continuity ofthe outer membrane was present. The helical nature of the inclusions suggests that they may beassociated with the complex proteins of the inner mitochondrial membrane and space whichhave the capacity of helix formation. The cristae with the paracrystalline arrays appear to be inthe orthodox configuration, a non-energized stage of the energy cycle. The mitochondriaprobably utilizes its unique ability to control respiration according to its functional requirementduring replication.
INTRODUCTION
Reports of paracrystalline arrays or inclusions within mitochondria are increasing.Several forms have been reported in animal and plant tissues. They also occur inpathological conditions. The diverse forms and dimensions of these paracrystalloidsincrease the problem of correlating function and morphology. The literature shows aprevailing uncertainty in interpretation of their role in mitochondria. Inclusions inmitochondria have been dealt with in a comprehensive manner by Tandler & Hoppel(1972) and Munn (1974). A few examples of the inclusions are, the prismatic cristae inthe astrocytes of the hamster brain (Blinzinger, Rewcastle & Hager, 1965), patternslike the star of David in the mitochondria of the kidney of the ammocoetes (Youson,1971) and intramitochondrial yolk crystals in frog oocytes (Massover, 1971). Someparacrystalloids are involved with cristal membranes and the intracristal space.Suzuki & Mostofi (1967), provided a classification which denned them as those thatoccur within the matrix and those that are present with the cristal space. The inclusionsthat are associated with the cristal membrane and space vary in configuration, and areoften helical in nature, such as the ones observed in the mitochondria of the bean root(Newcombe, Steer, Hepler & Wergin, 1968), the long helical structures within theastrocytes of the corpus striatum (Mugnaini, 1964), in the ameloblasts of the enamelorgan (Jessen, 1968), inclusions in the mitochondria of beef heart (Hall & Crane, 1970),in the follicular cell of the thyroid gland of the rabbit (Nathaniel, 1976, 1978),

24 D. R. Nathaniel
and in the thyroid gland of the bat (Nunez, Grief & Gershon, 1975). Enlarged mito-chondria with paracrystalloids occur in several pathological conditions. The studies ofliver biopsies of women with trophoblastic tumour showed the presence of crystalloidinclusions in enlarged mitochondria (Gonzalez-Angula et al. 1970). Martinez-Manautou, Aznar-Ramos, Bautista-O'Farril & Gonzalez-Augulo (1970), observedparacrystalloids within mitochondria of liver of women under contraceptive therapy.The morphology of the mitochondria reverted to normal after cessation of the drug.The administration of Clofibrate brought about ultrastructural changes in mito-chondria presenting multiple spiral structures, some of which appeared to have origi-nated from cristae. The changes occurred in enlarged mitochondria with cristae run-ning in the longitudinal axis of mitochondria (Lee, Kim & Lee, 1974). Crystalloidswere also observed in the dense matrix of giant or mega mitochondria in the liver ofjaundiced patients (Takasan, Yamada, Ozawa & Honjo, 1968), and Willis & Walton(1978) reported that in some cases of halothane hepatitis the mitochondria possessedcrystalloids in the matrix.
This paper reports the presence of paracrystalloids associated with cristal mem-branes and space in mitochondria presumably in the state of division. They were veryfew in number and were present in the normal as well as in the thyroid gland of themale rabbit following gonadectomy. They were encountered less frequently in gona-dectomized rabbits. Such an observation has not been reported and may lend someinsight to a possible role of these complex structures in respiration and functional stateof the mitochondria.
MATERIALS AND METHODS
Adult male rabbits of the New Zealand strain weighing about 2-3 kg were used to study theeffect of gonadectomy on the thyroid gland. The rabbits were maintained on normal rabbit chowad libitum and allowed to get acclimatized to their environment for a week. The controls andthe experimental animals were anaesthetized with intravenous pentobarbital (Nembutal), andfixed by intravascular perfusion via the left ventricle with buffered paraformaldehyde. Theright atrium was opened to enhance adequate flow of the fixative. The thyroid gland was dis-sected out of the capsule and cut into cubes of 1-2 mm and fixed for a further period of 30 minin paraformaldehyde. After sufficient buffer wash, tissues were fixed in 1 % osmium tetroxidefor a period of 1 h and then embedded in Araldite. One-half micron sections were cut routinelyfor evaluation of the tissue at the light-microscope level. Thin sections cut from representativeblocks were stained with uranyl acetate and lead citrate for electron-microscopic examinationwith Philips EM 200 and 300.
RESULTS
The inclusions in the atypical cristae of the mitochondria in the rabbit thyroid glandshow a unique method of formation. They also display parallel arrays in obliquesections and a tight-junction configuration in the vertical plane (Nathaniel, 1976,1978). While searching for mitochondria with the paracrystalline arrays in the con-trols and in the experimental group dealing with gonadectomy and its effect on thethyroid gland, a few large mitochondria that appeared to be replicating showed thepresence of inclusions in atypical cristae. Several stages of the process of replicationcan be seen in the micrographs (Figs. 1-7). In Figs. 1-4, a constriction connects the
Mitochondrial inclusions and division
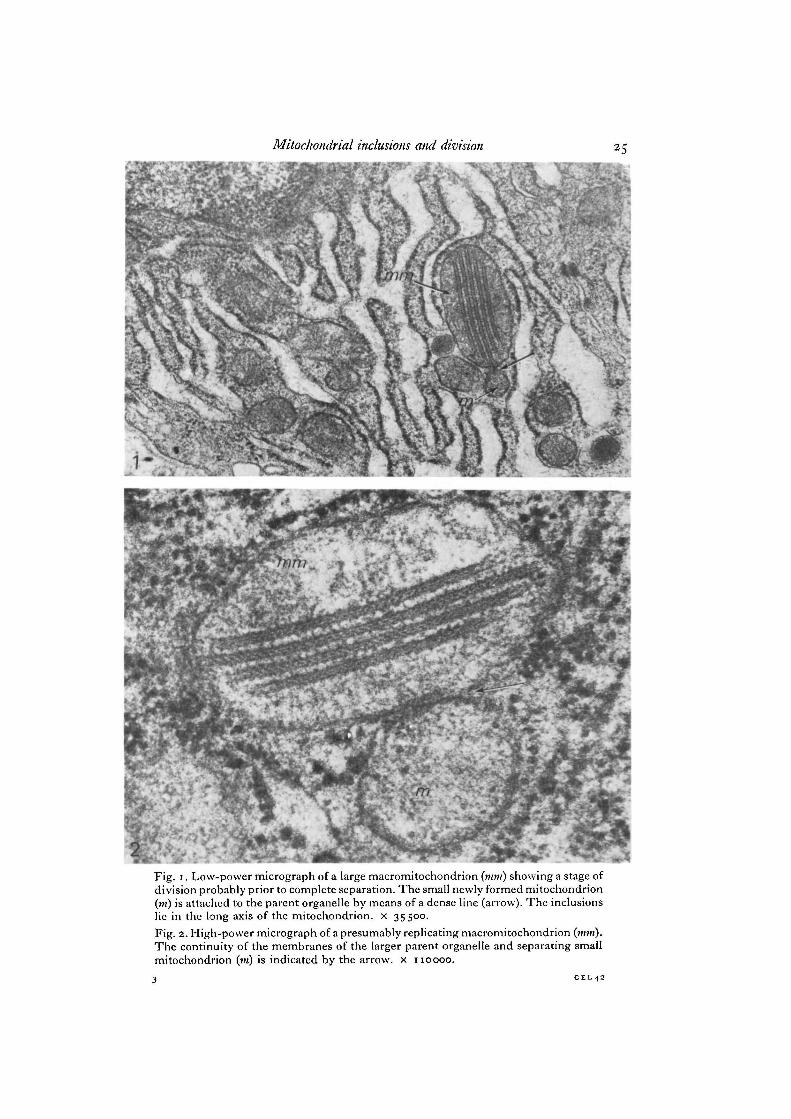
Fig. i. Low-power micrograph of a large macromitochondrion (mm) showing a stage ofdivision probably prior to complete separation. The small newly formed mitochondrion(m) is attached to the parent organelle by means of a dense line (arrow). The inclusionslie in the long axis of the mitochondrion, x 35500.Fig. 2. High-power micrograph of a presumably replicating macromitochondrion (mm).The continuity of the membranes of the larger parent organelle and separating smallmitochondrion (in) is indicated by the arrow, x 110000.
3 CKL42

26 D. R. Nathaniel

Mitochondrial inclusions and division 27
major macromitochondrion with the smaller newly forming one. The matrix of thelarger parent mitochondrion appears to be continuous with that of the new one at thesite of the constriction in Fig. 3. But in Figs. 1 and 4 an almost complete division isobserved, although a small attachment still persists as a dense line. The separating neworganelle is attached to the large mitochondrion by means of the continuity of theouter limiting membrane in Figs. 5 and 6, whereas in Figs. 2 and 7, the outer mem-branes are adherent just at one point indicated by the arrow. In all instances theinclusions were present only in the larger parent mitochondrion.
The morphological features of the inclusions are similar to those reported in thethyroid gland of the rabbit (Nathaniel, 1976, 1978). The paracrystalline arrays lie inthe longitudinal axis of the mitochondrion (Figs. 1-7). The tight junction configurationin a vertical plane is often observed (Figs. 1,2,4, 5). In the vertical sections the electron-dense outer layers enclose an electron-lucent space which has a centrally located denseline with a beaded appearance displaying periodicity of 25 nm (Figs. 2, 4). The obliquesections show parallel arrays revealing the filamentous helical nature of the inclusion(Figs. 5, 6). The obliquely disposed parallel arrays have a pitch of 25 nm and the fibresmake an angle of 450 with the long axis of the inclusion. Infrequently a honeycombpattern was present, probably due to a tangential section (Figs. 3, 7). In Fig. 5, themitochondrion shows the inclusions sectioned in 2 different planes. The lower 4 aresectioned vertically whereas the uppermost inclusion is sectioned in an oblique planeshowing the filamentous nature effectively. Large dense bodies are present in themitochondrion in Fig. 6, one of them being closely related to the inclusion and mayrepresent binding sites for ions particularly cations such as Ca2+ in the matrix.
DISCUSSION
The autonomous nature of mitochondria has puzzled scientists for many years.The labelling studies of Luck (1963), and Parsons & Rustard (1968) lend support to the
Fig. 3. Micrograph of macromitochondrion (mm) with a constriction (arrow), wherethe matrix is continuous with the smaller newly forming mitochondrion (in). The in-clusion within the large mitochondrion presents a honeycomb pattern as a result of atangential section, x 87000.
Fig. 4. This micrograph is an enlargement of mitochondrion in Fig. 1 showing a stageprobably prior to complete separation of the small mitochondrion (m) from the mainmacromitochondrion (vim). The connexion with the main organelle is indicated by adense line (arrow), which appears to be continuous with the outer membrane of thelarge mitochondrion. The inclusions are 30 nm in width enclosing an electron-lucentspace of about 15 nm. The dense line in the middle of the electron-lucent spacepresents a beaded appearance which has a point-to-point spacing of 25 nm, x 93400.Fig. 5. Electron micrograph of a large mitochondrion (mm) in process of replication.The newly formed organelle (m) or the small mitochondrion is attached to the parentmitochondrion by connecting membranes (arrows). The inclusions are in the longaxis of the large mitochondrion and present the longitudinal and oblique profile. Thehelical and filamentous nature of the paracrystalline arrays is effectively displayed inthe uppermost inclusion sectioned obliquely (paired arrows). The parallel arrays make anangle of 45° with the vertical axis of the inclusion and the pitch is 25 nm. x 82800.
3-2

D. R. Nathaniel
it*.

Mitochondrial inclusions and division 29
hypothesis that mitochondria arise by steady growth and division of pre-existingmitochondria. The evidence that mitochondria contain DNA provided by Nass &Nass (1963), Kislev, Swift & Bogorad (1965) and Leduc, Bernard & Tournier (1966),explains their ability to divide and separate as independent entities. Tandler,Erlandson, Smith & Wunder (1969) reported that the mitochondria of the liver inriboflavin-deficient mice attained enormous size and when the deficiency was cor-rected, numerous partitioned mitochondria with protrusions often at the base werepresent. The protrusions eventually separated from the main mass. The partitionconsisted of 2 parallel membranes which were joined to the internal limiting mem-brane. Since there was a corresponding increase in mitochondrial number, Tandleret al. (1969) presumed that the partitioned mitochondria were in process of replication.In the skipper butterly, 22-34 n a^ter t n e emergence of the adult, the mitochondriaof the metamorphosing fat body increase 700% as determined morphometrically.During the period of increase, numerous mitochondria with multiple partitions andconstrictions were observed (Larson, 1970). Partitioned mitochondria were also foundin the cardiac muscle of the mouse (Tandler & Hoppel, 1970) and in oncocytoma of theparotid gland (Tandler & Hoppel, 1972). These observations add strength to theassumption that mitochondria grow and divide by partition. In the present study en-larged elongated mitochondria show constrictions and stages of separation. The uniquefeature in the present report is the presence of helical inclusions in the mitochondriathat are in process of division.
There is growing awareness that mitochondria exist in a variety of morphologicalstates and profound changes take place in response to metabolic alterations. Experi-mental evidence show that rapid changes in configurational state of the cristae accom-pany changes in the energy state of the mitochondria (Williams et al. 1970; Green &Sungchul, 1972). The transition from an energized to non-energized configurationis rapid or instantaneous (Green & Baum, 1970).
The paracrystalline arrays in the mitochondria of the present study bear a closeresemblance to the helical inclusions reported in the Mung bean root (Newcombe et al.1968) and those in the guinea-pig liver (Valdivia, 1970), which according to Korman,Addink, Wakabayashi & Green (1970a), are in the orthodox configuration of theenergy cycle, where the intracristal space is minimal when compared to the matrix(Korman et al. 1970/1). Korman et al. (1970a) suggested that the paracrystalline arrayscan arise from any configuration of the energy cycle, the essential feature being the
Fig. 6. Micrograph showing a dividing macromitochondrion (mm). The larger mito-chondrion (mm) is connected to the smaller budding mitochondrion (m) by means ofa continuous outer membrane (arrow). This, probably, is a stage prior to completeseparation. The inclusions sectioned in an oblique plane show parallel arrays of ahelical nature (paired arrows). Two large dense bodies (d) are present in the matrix, oneof which lies in close apposition to an inclusion, x 93500.Fig. 7. A large mitochondrion (mm) in the process of budding. The only point ofattachment to the small mitochondrion (m) is indicated by an arrow where there is adensity at the site of the attachment of membranes. The inclusions are tangentially sec-tioned and therefore present a honeycomb pattern, x 82700.

30 D. R. Nathaniel
possibility for extensive apposition of base piece to base piece. They mentioned thatthe elongation of the mitochondrion enhances the apposition of cristal membranes.Elongation of mitochondria of the guinea-pig liver could be achieved by subjectingthe animal to cooling or low oxygen tension for an extended period of time, and the num-ber of paracrystalline arrays within the mitochondria were simultaneously increased(Valdivia, 1970). The treatment with dinitrophenol increased the number of para-crystalline arrays within mitochondria of the guinea-pig liver (Korman et al. 1970a).Dinitrophenol is one of the prime uncouplers and is known to discharge the energizedconfiguration to a non-energized configuration, one of which is the orthodox form(Williams et al. 1970). It has been shown that, calcium and low osmotic pressure caninduce the orthodox configuration of the energy cycle (Allman, Wakabayashi, Korman& Green, 1970). Large dense granule which is probably the binding site for cationssuch as Ca2+, is present in close association with the paracrystalline arrays in the mito-chondrion in Fig. 6. But the true nature of the granule cannot be ascertained morpho-logically.
Recently, tubulofilamentous components were observed in cristae of mitochondriaof sperm (Fucus). They were not present in younger, less differentiated sperms andtheir presence in mitochondria of older cells may fulfil a functional requirement of freeswimming sperms (Pollock & Cassell, 1977). A functional role of the paracrystallinearrays appears to be forthcoming. The intermembranous inclusions or paracrys-talline arrays found in mitochondria from ischaemic muscle were protein in natureas observed from staining properties and enzyme extraction procedures (Hanzlikova& Schiaffino, 1977). In the present study the inclusions sectioned obliquely show ahelical structure, filamentous in nature. The catalytic as well as the non-catalyticproteins of the inner mitochondrial membrane have high capability of helix formation(Green & Sungchul, 1972). One can only hypothesize that the helical components areassociated with the complex proteins of the inner mitochondrial membrane and spaceand are formed during apposition of cristal membranes in the orthodox configurationof the energy cycle during division. It is perhaps a unique ability of the mitochondriato use a control mechanism to utilize energy according to the functional requirementof its environment. This adaptive phenomenon of the mitochondria could play aprominent role in recovery as well as in the normal function of the cell.
The technical assistance of Paul Perumal is gratefully acknowledged. Thanks are due toRoberta Van Aertselaer and Cheryl Wagner, Department of Anatomy, University of Manitobafor typing the manuscript.
This work was supported in partby the University of Manitoba Grant-in-Aid (431-3113-01),Multiple Sclerosis Society of Canada and the Technology and Research Foundation of theParalysed Veterans of America.
REFERENCES
ALLMAN, D. W., WAKABAYASHI, T., KORMAN, E. F. & GREEN, D. E. (1970). Studies on thetransition of the cristal membrane from the orthodox to the aggregated configuration. Top-ology of bovine adrenal cortex mitochondria in the orthodox configuration. Bioenergetics 1,73-86.
BLINZINGER, K., REWCASTLE, N. B. & HAGER, H. (1965). Observations on prismatic type mito-chondria within astrocytes of the Syrian hamster brain. J. Cell Biol. 25, 293-303.

Mitochondrial inclusions and division 31
GONZALEZ-ANCULA, A., AZNAR-RAMOS, R., MARQUEZ-MONTER, H., BIERZWINSKY, G. &
MARTINEZ-MANAUTOU, J. (1970). The ultrastructure of liver cell in women under steroidtherapy. Acta endocr., Copnh. 65, 193-206.
GREEN, D. E. &BAUM, H. (1970). Energy and the Mitochondrion. New York, London: AcademicPress.
GREEN, D. E. & SUNCCHUL, J. (1972). The electronmechano-chemical model of mitochondrialstructure and function. Bioenergetics 3, 159-202.
HALL, J. D. & CRANE, F. L. (1970). An intracristal structure in beef heart mitochondria. ExplCell Res. 62, 480-483.
HANZLIKOVA, V. & SCHIAFFINO, S. (1977). Mitochondrial changes in ischemic skeletal muscle.J. Ultrastruct. Res. 60, 121-133.
JESSEN, J. (1968). The morphology and distribution of mitochondria in ameloblasts with specialreference to a helix containing type. J. Ultrastruct. Res. 22, 120-135.
KISLEV, N., SWIFT, H. & BOGORAD, L. (1965). Nucleic acids of chloroplasts and mitochondriain Swiss chard. J. Cell Biol. 25, 327-344.
KORMAN, E. F., ADDINK, A. D. F., WAKABAYASHI, R. & GREEN, D. E. (1970a). A unified modelof mitochondrial morphology. Bioenergetics 1, 9-32.
KORMAN, E. F., HARRIS, R. A., WILLIAMS, C. H., WAKABAYASHI, T., GREEN, D. E. & VALDIVIA,
E. (19706). Paracrystalline arrays in mitochondria. Bioenergetics 1, 387-404.LARSON, W. J. (1970). Genesis of mitochondria in insect fat body. J. Cell Biol. 47, 373-
383.LEDUC, E. H., BERNARD, W. & TOURNIER, P. (1966). Cyclic appearance of atypical mitochondria
containing DNA fibers in cultures of an adenovirus. 12. Induced hamster tumour. Expl CellRes. 42, 597-616.
LEE, K. J., KIM, D. N. & LEE, K. T. (1974). Ultrastructural study of hepatic mitochondrialabnormality in swine treated with clofibrate. Expl molec. Path. 20, 387-396.
LUCK, D. J. L. (1963). Formation of mitochondria in Neurospora crassa. A quantitative radio-autographic study. J. Cell Biol. 16, 483-499.
MARTINEZ-MANAUTOU, J., AZNAR-RAMOS, R., BAUTISTA-O'FARRIL, J. & GONZALEZ-AUGULO, A.(1970). The ultrastructure of liver cells in women under steroid therapy. Acta endocr.,Copnh. 65, 207-211.
MASSOVER, W. H. (1971). Intramitochondrial yolk crystals of frog oocytes. II . Expulsion ofintramitochondrial yolk crystals to form single membrane bound hexagonal crystalloids.J. Ultrastruct. Res. 36, 603-620.
MUCNAINI, E. (1964). Helical filaments in astrocytic mitochondria of the corpus striatum of therat. J. Cell Biol. 23, 173-182.
MUNN, E. A. (1974). The Structure of Mitochondria. London, New York: Academic Press.NASS, M. M. K. & NASS, S. (1963). Intramitochondrial fibers with DNA characteristics.
I. Fixation and electron staining reactions. J. Cell Biol. 19, 593-611.NATHANIEL, D. R. (1976). Helical inclusions and atypical cristae in the mitochondria of the
rabbit thyroid gland. J. Ultrastruct. Res. 57, 194-203.NATHANIEL, D. R. (1978). Effect of gonadectomy on the follicular cell and inclusions in mito-
chondria of rabbit thyroid gland. Am. J. Path. 91, 137-148.NEWCOMBE, E. H., STEER, N. W., HEPLER, P. K. & WERGIN, W. P. (1968). An atypical crista
resembling a 'tight junction' in bean root mitochondria. J. Cell Biol. 39, 35-42.NUNEZ, E. A., GRIEF, R. L. & GERSHON, M. D. (1975). Paracrystalloids from mitochondria in
thyroid follicular cells. Normal occurrence and experimental induction. Lab. Invest. 33, 352-361.
PARSONS, J. A. &RUSTARD,R. C. (1968). The distribution of DNA among dividing mitochondriaof Tetrahymena pyriformis. J. Cell Biol. 37, 683-693.
POLLOCK, E. G. & CASSELL, R. Z. (1977). An intracristal component of fucus sperm mito-chondria. J. Ultrastruct. Res. 58, 172-177.
SUZUKI, T. & MOSTOFI, F. K. (1967). Intramitochondrial filamentous bodies in the thick limbof henle of the rat kidney. J. Cell Biol. 33, 605-623.
TAKASAN, H., YAMADA, T., OZAWA, K. & HONJO, I . (1968). Morphological alteration throughchanges in respiratory enzyme levels in the liver mitochondria of jaundiced patients. JlElectron Microsc, Tokyo 24, 184-207.

32 D. R. Nathaniel
TANDLER, B., ERLANDSON, R. A., SMITH, A. L. & WUNDER, E. L. (1969). Riboflavin and mousehepatic cell structure and function. II. Division of mitochondria during recovery from simpledeficiency. J. Cell Biol. 41, 477-493.
TANDLER, B. & MOPPEL, C. L. (1970). Division of cardiac mitochondria. Anat. Rec. 166, 388.TANDLER, B. & HOPPEL, C. L. (1972). Mitochondria. New York, London: Academic Press.TANDLER, B., HUTTER, R. V. P. & ERLANDON, R. A. (1970). Infrastructure of oncocytoma of the
parotid gland. Lab. Invest. 23, 567-580.VALDIVIA, E. (1970). Unpublished observations cited Korman et al. in Paracrystalline arrays in
mitochondria. Bioenergetics 1, 387-404.WILLIAMS, C. H., WILLIAM, J. V., HARRIS, R. A., CALDWELL, M., GREEN, D. E. & VALDIVIA, E.
(1970). Confirmational basis of energy transduction in membrane systems. Configurationalchanges of mitochondria. In situ and in vitro. Bioenergetics 1, 147-180.
WILLIS, E. J. & WALTON, B. (1978). A morphological study of unexplained hepatitis followinghalothane anesthesia. Am. J. Path. 91, 11-32.
YOUSON, J. H. (1971). Prismatic cristae and matrix granules in mitochondria of the kidneys ofammocoetes. J. Cell Biol. 48, 189-191.
(Received 7 August 1979)