Embed Size (px)
Citation preview

Parental memories shape seedsFred Berger1,2 and Abed Chaudhury3
1 Temasek Lifesciences Laboratory, 1 Research Link, 117604 Singapore2 Department of Biological Sciences, National University of Singapore, 14 Science Drive, 117543 Singapore3 VitaGrain, 232 Orchard Road, Level 9, Suite 232, Faber House, 238854 Singapore
Review
It is ten years since imprinting was first demonstrated inArabidopsis, following the realization, five years earlier,that some genetic controls of seed development did notconform to Mendelian inheritance. Sixteen imprintedgenes have since been identified in maize and Arabidop-sis and these are expressed primarily in the endosperm,which nurtures embryo development. Imprinting resultsfrom the regulation of transcriptional silencing by DNAmethylation or by Polycomb Group complex-mediatedhistone methylation. Here we review recent studiessuggesting that imprinting results from global epige-netic changes that occur during female gametogenesis.We also discuss why imprinting has evolved and what itsbiological functions might be.
The discovery of imprinting in plantsIn flowering plants, two female gametes (the egg cell andthe central cell) originate from the haploid female game-tophyte produced by meiosis. The female gametes reside inthe embryo sac in the maternally derived ovule integu-ments [1–3]. After double fertilization, the egg cell and thecentral cell develop into the embryo and the endosperm,respectively (Figures 1,2). Nutrients are delivered from themother plant to the seed through specialized maternaltissues. Once inside the seed, the nutrients pass throughthe endosperm to the developing embryo [4–6]; thus,embryo development depends entirely on maternalsupplies. Maternal-effect mutations affecting thesesupplies are also known to impair seed development [7].
Geneticists working with maize (Zea mays) observednon-Mendelian controls of seed development in crossesinvolving one polyploid parent and one diploid parent,leading to seed lethality [8] (Box 1). In Arabidopsis (Ara-bidopsis thaliana), seeds produced by crosses between atetraploid mother and a diploid father are smaller thanwild-type diploid seeds, whereas seeds produced by atetraploid father and a diploid mother are larger thanwild-type seeds [9]. Thus, seed size is enhanced by anexcess of paternal genomes and restricted by an excessof maternal genomes in the zygotic products. Furthermore,interploidy crosses affect endosperm development specifi-cally [8,9], indicating the existence of genes expressedpreferentially by one of the two parental alleles, also calledimprinted genes [10]. First evidence for imprinting camefrom the demonstration of the maternal expression of cer-tain allele of theR gene responsible for seed pigmentation inmaize [8] (Box 1). The first imprinted genes were eventuallycharacterized after 1999 in Arabidopsis and maize [11–13].
Corresponding authors: Berger, F. ([email protected]); Chaudhury, A.([email protected]).
550 1360-1385/$ – see front matter � 2009 Elsevier Ltd. A
Here, we review rapid advancesmade in our understandingof the molecular controls of imprinting in plants and high-light recent data that point to a global maternal impact ofthe regulationofDNAmethylation in female gametesand inmaternal vegetative components [14].
Imprinted genes and their functionMost imprinted genes known in plants are only or prefer-entially expressed in the endosperm and some of these areessential for endosperm development [15,16]. Sixteenimprinted genes have been identified in Arabidopsis andmaize (Table 1). MEA (MEDEA) [17,18] and FERTILIZA-TION INDEPENDENT SEED 2 (FIS2) [19,20] are mater-nally expressed and encode conserved subunits of aPolycomb Group (Pc-G) complex active in endosperm[21]. The FIS Pc-G complex controls the developmentaltiming of endosperm development [22], the degree ofproliferation and the stage of endosperm cellularization[22–24]. The functions of the other imprinted genes remainunclear (Table 1).
There is a need for new methods to discover newimprinted genes. Analysis of QTL linked with seed de-velopment could lead to the discovery of imprinted genes[25]. Given that all known imprinted genes are expressedin endosperm, genome-wide studies from isolated endo-sperm tissuemight constitute a promising approach for thediscovery of new imprinted genes [26]. This prediction isvalidated by the discovery of five imprinted genes from 50candidates identified using a genome-wide analysis ofDNA methylation from endosperm in Arabidopsis [27].
Molecular control of imprinting by DNA methylationBoth alleles ofFERTILIZATION INDEPENDENTSEED2(FIS2), MATERNALLY EXPRESSED PAB C-TERMINAL(MPC) and FLOWERING WAGENINGEN (FWA) aresilenced throughout the plant life cycle until gametogen-esis (Figure 2). Silencing of FWA, FIS2 and MPC ismediated by the DNA METHYLTRANSFERASE 1(MET1), which maintains DNA methylation of CpG sites[19,28,29]. Silencing marks on FWA, FIS2 and MPC aremaintained in the sperm cells during male gametogenesis[19,29]. During endosperm development, the inheritedpaternal copy of maternally expressed imprinted genesremains silenced by MET1, whereas the maternal copyis inherited as transcriptionally active, resulting in mono-parental expression [19,28–31]. Analysis of the maternallyexpressed Fertilization independent endosperm1 (Fie1)locus in maize shows that DNA methylation is presentin sperm cells and is specifically removed in the centralcell, (the female progenitor of endosperm) but not in the egg
ll rights reserved. doi:10.1016/j.tplants.2009.08.003 Available online 11 September 2009
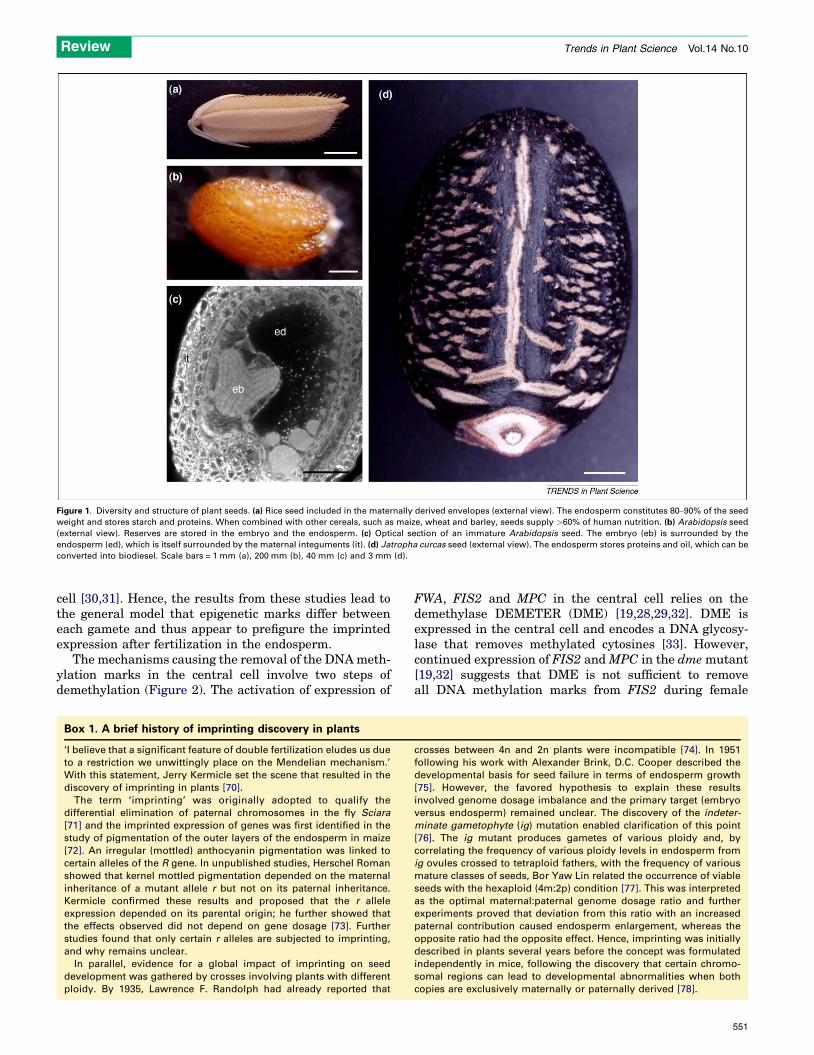
Figure 1. Diversity and structure of plant seeds. (a) Rice seed included in the maternally derived envelopes (external view). The endosperm constitutes 80–90% of the seed
weight and stores starch and proteins. When combined with other cereals, such as maize, wheat and barley, seeds supply >60% of human nutrition. (b) Arabidopsis seed
(external view). Reserves are stored in the embryo and the endosperm. (c) Optical section of an immature Arabidopsis seed. The embryo (eb) is surrounded by the
endosperm (ed), which is itself surrounded by the maternal integuments (it). (d) Jatropha curcas seed (external view). The endosperm stores proteins and oil, which can be
converted into biodiesel. Scale bars = 1 mm (a), 200 mm (b), 40 mm (c) and 3 mm (d).
Review Trends in Plant Science Vol.14 No.10
cell [30,31]. Hence, the results from these studies lead tothe general model that epigenetic marks differ betweeneach gamete and thus appear to prefigure the imprintedexpression after fertilization in the endosperm.
The mechanisms causing the removal of the DNAmeth-ylation marks in the central cell involve two steps ofdemethylation (Figure 2). The activation of expression of
Box 1. A brief history of imprinting discovery in plants
‘I believe that a significant feature of double fertilization eludes us due
to a restriction we unwittingly place on the Mendelian mechanism.’
With this statement, Jerry Kermicle set the scene that resulted in the
discovery of imprinting in plants [70].
The term ‘imprinting’ was originally adopted to qualify the
differential elimination of paternal chromosomes in the fly Sciara
[71] and the imprinted expression of genes was first identified in the
study of pigmentation of the outer layers of the endosperm in maize
[72]. An irregular (mottled) anthocyanin pigmentation was linked to
certain alleles of the R gene. In unpublished studies, Herschel Roman
showed that kernel mottled pigmentation depended on the maternal
inheritance of a mutant allele r but not on its paternal inheritance.
Kermicle confirmed these results and proposed that the r allele
expression depended on its parental origin; he further showed that
the effects observed did not depend on gene dosage [73]. Further
studies found that only certain r alleles are subjected to imprinting,
and why remains unclear.
In parallel, evidence for a global impact of imprinting on seed
development was gathered by crosses involving plants with different
ploidy. By 1935, Lawrence F. Randolph had already reported that
FWA, FIS2 and MPC in the central cell relies on thedemethylase DEMETER (DME) [19,28,29,32]. DME isexpressed in the central cell and encodes a DNA glycosy-lase that removes methylated cytosines [33]. However,continued expression of FIS2 andMPC in the dmemutant[19,32] suggests that DME is not sufficient to removeall DNA methylation marks from FIS2 during female
crosses between 4n and 2n plants were incompatible [74]. In 1951
following his work with Alexander Brink, D.C. Cooper described the
developmental basis for seed failure in terms of endosperm growth
[75]. However, the favored hypothesis to explain these results
involved genome dosage imbalance and the primary target (embryo
versus endosperm) remained unclear. The discovery of the indeter-
minate gametophyte (ig) mutation enabled clarification of this point
[76]. The ig mutant produces gametes of various ploidy and, by
correlating the frequency of various ploidy levels in endosperm from
ig ovules crossed to tetraploid fathers, with the frequency of various
mature classes of seeds, Bor Yaw Lin related the occurrence of viable
seeds with the hexaploid (4m:2p) condition [77]. This was interpreted
as the optimal maternal:paternal genome dosage ratio and further
experiments proved that deviation from this ratio with an increased
paternal contribution caused endosperm enlargement, whereas the
opposite ratio had the opposite effect. Hence, imprinting was initially
described in plants several years before the concept was formulated
independently in mice, following the discovery that certain chromo-
somal regions can lead to developmental abnormalities when both
copies are exclusively maternally or paternally derived [78].
551

Figure 2. A model for the control of maternally expressed imprinted genes regulated by DNA methylation. The RBR1/MSI1 pathway activates the maternal expression of
FIS2 and FWA via the transcriptional repression of the DNA methyltransferase MET1 in the central cell (in yellow within the embryo sac). The imprinted genes FIS2 and FWA
(each allele is shown as a rectangle) are repressed throughout the vegetative stage by MET1-dependent DNA methylation on their promoters. Transcriptional repression is
maintained by MET1 in the male gametes carried by pollen. At the end of female gametogenesis, the low level of MET1 activity during female gametogenesis causes a
gradual dilution of the DNA methylation marks after each cycle of DNA replication (loss of methylation is indicated by pink curved arrow pointing to CH3). Following the
third cycle of syncytial division, the central cell and the egg cell become isolated from each other. In the egg cell, the remaining marks are sufficient to maintain FIS2 and
FWA silenced before and after fertilization in the embryo. By contrast, DME is expressed in the central cell and previous enrichment in hemi-methylated DNA at the FIS2 and
FWA loci primes DME to remove remaining DNA methylation, leading to activation of FIS2 and FWA. After fertilization, the endosperm inherits an active allele from the
central cell and an inactive allele from one of the two sperm cells. The demethylated maternal allele (red rectangle) remains active (black arrow) in the endosperm, whereas
the paternal allele (blue rectangle) remains silent, leading to imprinted expression.
Review Trends in Plant Science Vol.14 No.10
gametogenesis and an additional mechanism relying onthe Retinoblastoma pathway was identified [34]. The Ara-bidopsis homolog RETINOBLASTOMA RELATED (RBR)interacts with the WD40 domain-containing protein MUL-TICOPYSUPPRESSOR OF IRA 1 (MSI1) and directlyrepresses MET1 transcription during the late phase of
Table 1. Imprinted genes and their function
Species Name
Maize (Zea mays) R (Certain alleles only)
Allele MO17 of the dzr1 locus
Fertilization independent endosperm 1 (Fie1)
No apical meristem related protein 1 (Nrp1)
Maize Enhancer of Zeste1 (Mez1)
Maternally expressed gene1 (Meg1)
Arabidopsis thaliana MEDEA (MEA)
FLOWERING WAGENINGEN (FWA)
PHERES1 (PHE1)
FERTILIZATION INDEPENDENT SEED 2 (FIS2
MATERNALLY EXPRESSED PAB
C-TERMINAL (MPC) 43
HD-ZIP GENE9 (HDG9)
HD-ZIP GENE8 (HDG8)
HD-ZIP GENE3 (HDG3)
ATMYBR2
AT5G62110
552
female gametogenesis. FIS2 and FWA expression in thecentral cell is completely inhibited when the Retinoblas-toma pathway is inactive. This is probably the consequenceof the hypermethylation of FIS2 and FWA, although thishas not been demonstrated directly. These results lead tothe model depicted in Figure 2. Owing to transcriptional
Potential function Refs
Transcription factor [73]
Reserve protein [79]
Pc-G chromatin remodeling factor [44,45,80]
Unknown [37]
Pc-G chromatin remodeling factor [36]
Cys-rich peptide [81]
Pc-G chromatin remodeling factor [17,19,31]
Homeobox transcription factor [33,42,48]
Type1 MADS-box transcription factor [35,40]
) Pc-G chromatin remodeling factor [19,33,39,48]
C-terminal domain of poly(A) binding
proteins (PABPs); probably
controls mRNA stability and translation
[43]
Transcription factor [27]
Transcription factor [27]
Transcription factor [27]
Transcription factor [27]
Putative Transcription factor [27]

Review Trends in Plant Science Vol.14 No.10
inhibition by the Retinoblastoma pathway, MET1 activitydrops below a point where DNA methylation is not main-tained, producing hemi-methylated DNA, the preferredsubstrate for DME [35]. The synergistic action of passivedemethylation by repression of MET1 activity followed byactive demethylation by DME completely demethylatesthe cis elements in FIS2 and FWA promoters, causingexpression of these genes in the central cell. After fertiliza-tion, the active maternal allele is inherited with ademethylated cis element, whereas the inactive paternalallele is inherited with a fully methylated cis element.MET1 is active in endosperm and maintains this imbal-anced pattern of methylation, causing imprinted expres-sion in endosperm. Such a mechanism is likely to apply toall maternally expressed imprinted genes silenced byMET1 in sperm cells [27].
Global control of seed development by MET1MET1 is expressed strongly in sperm cells [34] and its lossduring male gametogenesis inhibits endosperm growthand results in smaller seeds [14,36–39]. This is probablycaused by ectopic activation of the paternal allele ofregulated imprinted genes, such asFWA andFIS2. Crossesbetween wild-type pollen and ovules from plants carryingan antisense directed against MET1 (MET1a/s) showedthe opposite effect [38,39]. However, this was not observedin crosses between wild-type pollen and ovules frommet1/1plants [36,37]. The difference between these two exper-iments resides in the dominant effect of the antisensedirected againstMET1. The increase in seed size observedin seeds from crosses between wild-type pollen and ovulesfrom MET1a/s plants could result from the loss of MET1on the female gametes, the integuments, or both. Thishypothesis was tested using mutant recessivemet1 alleles,which enable distinction between the two potential devel-opmental origins, because whereas all ovules produced bya met1/1 plant have wild-type integuments, half of themcarry a met1 mutant female gamete. However, crossesbetween wild-type pollen and ovules from met1/1 plantsdid not produce two classes of seeds size, as had beenpredicted for a direct effect of a loss of MET1 during femalegametogenesis [14,36]. This result could be explained bythe reduced expression of MET1 in female gametes [34].However crosses between wild pollen and ovules frommet1/met1 plants produced larger seeds, as shown forMET1a/s plants. Seed integuments of MET1a/s andmet1/met1 ovules contain more cells and elongateautonomously in absence of MET1 function [14]. MET1might thus prevent proliferation and elongation of ovulesinteguments, which would explain the dominant maternaleffect of the loss of MET1 on seed size [14,38,39]. Overall,the epigenetic control of seed size by MET1 appears toresult primarily from maternal controls. These derivedirectly from the action of MET1 on the sporophytic vege-tative tissues and indirectly through the role of MET1acting during male gametogenesis.
Genome-wide demethylation during femalegametogenesis?Whereas DNA demethylation mediated by DME duringfemale gametogenesis is likely to affect only a discrete
number of loci [40], DNA demethylation regulated bythe Retinoblastoma pathway [34] could affect the entiregenome, leading to a reduction of the constitutive hetero-chromatin fraction in the central cell, which could beinherited in the endosperm. A decrease of maternalDNAmethylation in the endospermwas suggested to occurin maize based on the analysis of methylation-sensitiveamplified polymorphism [41] and in Arabidopsis based onthe reduction of the heterochromatin fraction [42]. Thisglobal decrease has been demonstrated recently using deepsequencing after immunoprecipitation against methylcy-tosine residues of DNA extracted from purified endospermor embryos [27,43]. The genome-wide cytosine methylationlevel is decreased in endosperm by 15–20%when comparedwith levels measured in embryos or vegetative tissues.Endosperm from dme mutants showed only a partialincrease of DNA methylation on CG, implying that aDME-independent pathway causes demethylation. Accord-ingly, retinoblastoma-dependent repression of MET1expression in the central cell is likely to contribute todemethylation of DNA in the endosperm precursor centralcell [34]. Interestingly, low DNA methylation levels inendosperm are observed not only at imprinted loci, butalso at repeats and transposons [27,43].
We propose that a global demethylation of the maternalgenome occurs in the central cell by the combined action ofpassive demethylation owing to reduced MET1 activityand active demethylation by DME in the central cell. Thisdemethylated status is inherited in endosperm, and DNAmethylation associated with imprinted loci might thusfollow general dynamics of DNA methylation in thegametes (maintained in sperms and reprogrammed inthe central cell). If this hypothesis is correct, imprintingmight not involve dedicated epigenetic mechanisms butendosperm-specific transcriptional activation of locidemethylated on the maternal allele by a global phenom-enon that does not affect male gametes.
Although MET1 does not appear to be expressed in theegg cell, global demethylation is not expected to affect theegg cell DNA significantly because it has been establishedthat silencing by DNAmethylation is transmitted from onegeneration to the next [44,45]. It is thus possible thatspecific mechanisms counter the loss of MET1 activity inthe egg cell and ensure the transmission of DNA methyl-ation patterns to the offspring. Such mechanisms couldinvolve DNA methyltransferases other than MET1. Thishypothesis is supported by the unusual highmethylation ofCHH contexts in isolated embryos compared with vegeta-tive tissues [43]. Large amounts of small non-coding RNAsmaternally inherited in the endosperm and the embryo,might also influence the global level of DNA methylationthrough their association with de novo DNA methyltrans-ferases [46].
The globalmaternal effect ofMET1 suggests additionalquestions. Wild-type seeds derived from crosses betweenwild-type pollen and ovules from met1/1 plants are largerthanwild-type seeds [14]. This suggests that the reductionofMET1 dosage in thematernal vegetative tissue has alsoan impact on seed size. Thus, any impact onMET1 activityduring vegetative development might affect seed size.Given that environmental stresses modulate DNA
553

Review Trends in Plant Science Vol.14 No.10
methylation [47,48], it is possible that DNA methylationprovides amemory of environmental conditions, resultingin altered seed size, a crucial trait for survival and adap-tation [49].
Molecular controls of imprinting by histone methylationSeveral studies have shown that DNA methylation marksare present at the MEA locus in the promoter and atrepeats at the 3’end of the gene, the MEA ISR domain[50–52] (and also refer to online data at http://epigen-omics.mcdb.ucla.edu/cgi-bin/hgTracks). In addition, MEAexpression depends DME [53,54]. Whether MET1 andDME regulate MEA imprinting directly is unclear. Theloss of MET1 in vegetative tissues and in pollen does notcause ectopic expression of MEA as could be expected ifMEA expression were to be directly repressed by DNAmethylation. In addition the loss of MET1 in male gametesdoes not deregulate MEA imprinted status in endosperm[53,55]. These results imply that MEA is repressed by amechanism independent of DNA methylation.
In vegetative tissues, both alleles ofMEA are silenced byHistone 3 Lysine 27 (H3K27) trimethylation, which ismediated by Pc-G complexes [53,55]. Genome-wide arraysof DNA methylation and H3K27 trimethylation haveshown that the MEA locus is covered with H3K27 tri-methylation [56]. Compromising H3K27 trimethylationin mutants for Pc-G activity causes MEA ectopic expres-sion in pollen and vegetative tissues and compromisesMEA imprinted status in endosperm [53,55]. It is thuslogical to conclude that MEA is silenced by H3K27 meth-ylation. This implies that transcriptional activation ofMEA requires removal of trimethylated H3K27 duringfemale gametogenesis. Although the mechanism causingsuch a removal is unknown, DME is required for MEAtranscriptional activation [32] and it is possible that MET1and DME indirectly activate a pathway that removesH3K27 trimethylation marks from MEA locus, leadingto its activation. Alternatively, maternal MEA expressionmight require a transcriptional activator that is itselfdirectly controlled by DNAmethylation and DME activity.A third hypothesis could involve the DNA methylationmarks on the ISR. The methylated ISR domain mightprevent MEA activation. DME would then have a moredirect role, removing DNAmethylation fromMEA ISR andenabling H3K27 methylation to be removed from theMEAlocus, leading to transcriptional activation. Such a complexinteraction between a methylated element in 30 of thepaternally expressed imprinted gene PHERES1 andH3K27 methylation has been recently discovered and isreported below [57].
One of the maize homologs of MEA, Maize enhancer ofzeste 1 (Mez1) is imprinted [58]. Instead of DNA methyl-ation, the silenced paternal allele carries H3K27 trimethy-lation on the paternal allele [59]. Similar to the self-regulation of MEA imprinting, disruption of PcG functionprovided by the Mez1 maternal allele causes expression ofthe Mez1 paternal allele [60], suggesting a conservation ofthe mechanisms that regulate imprinting of MEA and itshomolog Mez1.
PHERES1 (PHE1) [60] and two recently identifiedgenes HD-ZIP GENE 3 and At5G2110 [27] are the only
554
known paternally expressed imprinted genes in Arabidop-sis. The silencing of the maternal allele of PHE1 ismediated by the maternal action of Pc-G in endosperm[61]. However, the PHE1 maternal allele is expressed atvariable levels depending on the natural accessions, indi-cating that a Pc-G independent mechanism regulatesPHE1 [57]. The expression of the maternal allele dependson DNA methylation of an element that is present 2.5 Kbdownstream of the 30 end of PHE1. This element containsCpG residues methylated by MET1. DNA methylationusually silences expression but surprisingly the elementmust be demethylated to ensure the proper silenced statusofPHE1maternal allele. This demethylationmight be partof the global changes of the DNA methylation occurring inthe central cell and might thus depend on RBR and DMEpathways.
Although histone methylation by Pc-G is involved inimprinting in mammals [11,62], it does not appear to act asthe essential repressor of the silenced allele of imprintedgenes, as shown for certain imprinted genes in plants. Amajor challenge is to understand which mechanismsremove the H3K27 methylation mark from the expressedallele ofMEA andPHE1. Understanding suchmechanismswill further provide means to identify other imprintedgenes controlled by the Pc-G pathway.
What is the biological significance of imprinting?Imprinting arose independently in plants and mammals,and is characterized by a mode of reproduction involvingdirect maternal nutrition of the developing embryo. If amother produces offspring from different fathers, all off-spring are genetically related to the mother but carryunrelated paternal genomes. According to this scenario,paternal genomes compete for resource allocation from themother. Consequently, the ‘goal’ of the paternal genome isto derive as much maternal resource as possible for theembryos of which it is a part. When kin selection isconsidered, it is advantageous for the mother to down-regulate such opposing interference from the paternalgenomes to ensure equitable distribution of nutrients toeach offspring. These considerations led to the parentalconflict theory, which predicts a positive selection of mater-nally expressed growth inhibitors and paternallyexpressed growth enhancers [63]. The parental conflicttheory is supported in plants by the result of crossesbetween plants of different ploidies [9,10]. Additionaldosage of paternal genome causes increased endospermsize and additional maternal dosage reduces endospermsize. Furthermore, the maternal action of MET1 supportsan overwhelming maternal control of size during seeddevelopment. This scenario could still be considered as aconflict but what, if any, is the opposing role of the paternalgenome remains unclear.
The reproductive scenario associated with the parentalconflict theory applies to certain mammalian and out-crossing plant species but perhaps not so much to self-fertilizingArabidopsis, in which imprinting is still found tobe active. Imprinting could thus be related to an out-cross-ing reproductive strategy linked to its role in response tohybridity [64]. Interspecific crosses are possible in relatedplant species but often lead to reduced seed fertility. In a

Review Trends in Plant Science Vol.14 No.10
study of crosses between Arabidopsis relatives, it wasshown that PHERES1 and other AGAMOUS-LIKE TYPEI MADS-box genes have an important role in the viabilityof the hybrid seeds [65,66]. It is thus possible that imprint-ing has been selected as a mechanism contributing tohybridity and as such would be crucial for speciation.
Even if one assumes that ancestors of mammals andflowering plantswere obligate out-breeders with no restric-tion on the numbers of male partners, the parental conflicttheory suggests that imprinted genes should not be underpositive selection in species that always self fertilize. Ara-bidopsis thaliana usually self-fertilizes to a greater degreein contrast to closely related species Arabis lyrata andArabis petraea. Three studies have evaluated the type ofselection exerted on MEA in Arabidopsis and its homologsin related species [67–69]. Although there are some limiteddiscrepancies between the studies, it appears that MEAhas not been subjected to a strong positive selection inArabidopsis. Similarly, no positive selection was detectedfor the MEA maize homolog Mez1 [58]. Together, thesestudies do not provide strong support for the conflicttheory. However, it is difficult to reach an unambiguousconclusion since no similar data have been gathered forother imprinted genes in plants. The recent identificationof imprinting in three members of the clade of class IVhomeodomain transcription factors containing FWA,shows that all contain 50 methylated elements derivedfrom helitrons, suggesting that transposon insertions aretightly linked with the selection of imprinting [27]. Thistype of approach will significantly improve our under-standing of how imprinting evolved in plants.
Although only a limited number of imprinted geneshave been identified so far, the fact that they are allexpressed in the endosperm together with the endo-sperm-specific effect of interploid crosses in Arabidopsisand maize, leads us to envisage that imprinting function isintimately linked to the endosperm lineage during femalegametogenesis. Recent molecular insights also suggestthat the central cell might experience chromatin re-arrangement at the genomic scale including DNA andhistone methylation [27,34,42,43], which might causespecific expression of certain genes, the maternal alleleof which remains active in the endosperm. This endo-sperm-specific expression mechanism logically leads toimprinting if the paternal allele remains silenced. Thus,imprinting of maternally expressed genes might be a‘sequel’ of the global remodeling of chromatin marks occur-ring in the central cell.
Conclusion and perspectivesThese past few years have been marked by progress inunderstanding of the molecular mechanisms involved inimprinting and the discovery that global levels of DNAmethylation are reduced in the endosperm. However, themajor questions that remain include how the alleles ofgenes silenced by Pc-G marks are activated and howmaintenance of the imprinted status is maintained. It isessential to identify new imprinted genes to test whetherthe mechanisms outlined by the studies of MEA, PHE1,FIS2 and FWA can be generalized. Such identificationshould also lead to a better understanding of the functional
role of imprinting. In this respect, a change of scale of theanalysis of imprinting might be necessary, with involve-ment of population genetics and the study of the impact ofthe environment on imprinting.
AcknowledgementsThis manuscript is dedicated to the memory of Melissa Spielman, whosework and remarkable essays inspired researchers in the field. F.B. isfunded by Temasek LifeSciences Laboratory, Singapore MilleniumFoundation and affiliated to the Department of Biological Sciences, atNational University of Singapore. A.C. is supported by VitaGrain Ltd, aglobal Agro-tech company based in Singapore.
References1 Drews, G.N. and Yadegari, R. (2002) Development and function of the
angiosperm female gametophyte. Annu. Rev. Genet. 36, 99–1242 Faure, J.E. et al. (2002) Fertilization in Arabidopsis thalianawild type:
Developmental stages and time course. Plant J. 30, 481–4883 Berger, F. et al. (2008) Double fertilization - caught in the act. Trends
Plant Sci. 13, 437–4434 Berger, F. et al. (2007) Arabidopsis as a model for understanding the
basics of endosperm development. Plant Cell Monogr. 8, 91–1105 Lopes, M.A. and Larkins, B.A. (1993) Endosperm origin, development,
and function. Plant Cell 5, 1383–13996 Olsen, O.A. (2004) Nuclear endosperm development in cereals and
Arabidopsis thaliana. Plant Cell 16 (Suppl), S214–S2277 Chaudhury, A.M. and Berger, F. (2001) Maternal control of seed
development. Semin. Cell. Dev. Biol. 12, 381–3868 Kermicle, J.L. and Allemand,M. (1990) Gametic imprinting inmaize in
relation to the angiosperm life cycle. Development (Suppl.) 1, 9–149 Scott, R.J. et al. (1998) Parent-of-origin effects on seed development in
Arabidopsis thaliana. Development 125, 3329–334110 Spielman, M. et al. (2001) The epigenetic basis of gender in flowering
plants and mammals. Trends Genet. 17, 705–71111 Scott, R.J. and Spielman, M. (2004) Epigenetics: imprinting in plants
and mammals - the same but different? Curr. Biol. 14, R201–R20312 Gehring, M. et al. (2004) Imprinting and seed development. Plant Cell
16 (Suppl.), S203–S21313 Garnier, O. et al. (2008) Genomic imprinting in plants. Adv. Exp. Med.
Biol. 626, 89–10014 FitzGerald, J. et al. (2008) DNA methylation causes predominant
maternal controls of plant embryo growth. PLoS ONE 3, e229815 Berger, F. et al. (2006) Endosperm: an integrator of seed growth and
development. Curr. Opin. Plant Biol. 9, 664–67016 Kinoshita, T. et al. (2008) Genomic imprinting: a balance between
antagonistic roles of parental chromosomes. Semin. Cell. Dev. Biol. 19,574–579
17 Kinoshita, T. et al. (1999) Imprinting of the MEDEA polycomb gene inthe Arabidopsis endosperm. Plant Cell 11, 1945–1952
18 Grossniklaus, U. et al. (1998) Maternal control of embryogenesis byMEDEA, a polycomb group gene in Arabidopsis. Science 280, 446–450
19 Jullien, P.E. et al. (2006) Maintenance of DNA methylation during theArabidopsis life cycle is essential for parental imprinting. Plant Cell 18,1360–1372
20 Luo, M. et al. (1999) Genes controlling fertilization-independent seeddevelopment inArabidopsis thaliana. Proc. Natl. Acad. Sci. U. S. A. 96,296–301
21 Guitton, A.E. and Berger, F. (2005) Control of reproduction byPolycomb Group complexes in animals and plants. Int. J. Dev. Biol.49, 707–716
22 Ingouff, M. et al. (2005) Polycomb group genes control developmentaltiming of endosperm. Plant J. 42, 663–674
23 Guitton, A.E. et al. (2004) Identification of new members ofFertilisation Independent Seed Polycomb Group pathway involvedin the control of seed development in Arabidopsis thaliana.Development 131, 2971–2981
24 Kiyosue, T. et al. (1999) Control of fertilization-independent endospermdevelopment by theMEDEA polycomb gene in Arabidopsis. Proc. Natl.Acad. Sci. U. S. A. 96, 4186–4191
25 Li, Y. et al. (2008) A statistical model for estimating maternal-zygoticinteractions and parent-of-origin effects of QTLs for seed development.PLoS ONE 3, e3131
555

Review Trends in Plant Science Vol.14 No.10
26 Day, R.C. et al. (2008) Transcriptome analysis of proliferatingArabidopsis endosperm reveals biological implications for thecontrol of syncytial division, cytokinin signaling, and geneexpression regulation. Plant Physiol. 148, 1964–1984
27 Gehring, M. et al. (2009) Extensive demethylation of repetitiveelements during seed development underlies gene imprinting.Science 324, 1447–1451
28 Kinoshita, T. et al. (2004) One-way control of FWA imprinting inArabidopsis endosperm by DNA methylation. Science 303, 521–523
29 Tiwari, S. et al. (2008) MATERNALLY EXPRESSED PAB C-TERMINAL, a novel imprinted gene in Arabidopsis, encodes theconserved C-terminal domain of polyadenylate binding proteins.Plant Cell 20, 2387–2398
30 Hermon, P. et al. (2007) Activation of the imprinted Polycomb GroupFie1 gene in maize endosperm requires demethylation of the maternalallele. Plant Mol. Biol. 64, 387–395
31 Gutierrez-Marcos, J.F. et al. (2006) Epigenetic asymmetry of imprintedgenes in plant gametes. Nat. Genet. 38, 876–878
32 Choi, Y. et al. (2002) DEMETER, a DNA glycosylase domain protein, isrequired for endosperm gene imprinting and seed viability inArabidopsis. Cell 110, 33–42
33 Gehring, M. et al. (2009) DNA demethylation by DNA repair. TrendsGenet. 25, 83–90
34 Jullien, P.E. et al. (2008) Retinoblastoma and its binding partner MSI1control imprinting in Arabidopsis. PLoS Biol. 6, e194
35 Morales-Ruiz, T. et al. (2006) DEMETER and REPRESSOR OFSILENCING 1 encode 5-methylcytosine DNA glycosylases. Proc.Natl. Acad. Sci. U. S. A. 103, 6853–6858
36 Xiao, W. et al. (2006) Regulation of seed size by hypomethylation ofmaternal and paternal genomes. Plant Physiol. 142, 1160–1168
37 Garcia, D. et al. (2005) Maternal control of integument cell elongationand zygotic control of endosperm growth are coordinated to determineseed size in Arabidopsis. Plant Cell 17, 52–60
38 Adams, S. et al. (2000) Parent-of-origin effects on seed development inArabidopsis thaliana require DNA methylation. Development 127,2493–2502
39 Luo, M. et al. (2000) Expression and parent-of-origin effects for FIS2,MEA, and FIE in the endosperm and embryo of developingArabidopsisseeds. Proc. Natl. Acad. Sci. U. S. A. 97, 10637–10642
40 Penterman, J. et al. (2007) DNA demethylation in the Arabidopsisgenome. Proc. Natl. Acad. Sci. U. S. A. 104, 6752–6757
41 Lauria, M. et al. (2004) Extensive maternal DNA hypomethylation inthe endosperm of Zea mays. Plant Cell 16, 510–522
42 Baroux, C. et al. (2007) The triploid endosperm genome of Arabidopsisadopts a peculiar, parental, dosage-dependent chromatin organization.Plant Cell 19, 1782–1794
43 Hsieh, T.F. et al. (2009) Genome-wide demethylation of Arabidopsisendosperm. Science 324, 1451–1454
44 Saze, H. (2008) Epigenetic memory transmission through mitosis andmeiosis in plants. Semin. Cell. Dev. Biol. 19, 527–536
45 Cubas, P. et al. (1999) An epigenetic mutation responsible for naturalvariation in floral symmetry. Nature 401, 157–161
46 Mosher, R.A. et al. (2009) Uniparental expression of PolIV-dependentsiRNAs in developing endosperm of Arabidopsis. Nature 460, 283–286
47 Chinnusamy, V. and Zhu, J.K. (2009) Epigenetic regulation of stressresponses in plants. Curr. Opin. Plant Biol. 12, 133–139
48 Lukens, L.N. and Zhan, S. (2007) The plant genome’s methylationstatus and response to stress: implications for plant improvement.Curr. Opin. Plant Biol. 10, 317–322
49 Gomez, J.M. (2004) Bigger is not always better: conflicting selectivepressures on seed size in Quercus ilex. Evolution 58, 71–80
50 Cokus, S.J. et al. (2008) Shotgun bisulphite sequencing of theArabidopsis genome reveals DNA methylation patterning. Nature452, 215–219
51 Lister, R. et al. (2008) Highly integrated single-base resolution maps ofthe epigenome in Arabidopsis. Cell 133, 523–536
52 Zhang, X. et al. (2006) Genome-wide high-resolution mapping andfunctional analysis of DNA methylation in Arabidopsis. Cell 126,1189–1201
53 Gehring, M. et al. (2006) DEMETER DNA glycosylase establishesMEDEA Polycomb gene self-imprinting by allele-specificdemethylation. Cell 124, 495–506
556
54 Xiao, W. et al. (2003) Imprinting of the MEA Polycomb gene iscontrolled by antagonism between MET1 methyltransferase andDME glycosylase. Dev. Cell 5, 891–901
55 Jullien, P.E. et al. (2006) Polycomb Group complexes self-regulateimprinting of the polycomb group gene MEDEA in Arabidopsis.Curr. Biol. 16, 486–492
56 Zhang, X. et al. (2007) The Arabidopsis LHP1 protein colocalizeswith histone H3 Lys27 trimethylation. Nat. Struct. Mol. Biol. 14,869–871
57 Makarevich, G. et al. (2008) Mechanism of PHERES1 imprinting inArabidopsis. J. Cell. Sci. 121, 906–912
58 Haun, W.J. et al. (2007) Genomic imprinting, methylation andmolecular evolution of maize Enhancer of zeste (Mez) homologs.Plant J. 49, 325–337
59 Haun, W.J. and Springer, N.M. (2008) Maternal and paternal allelesexhibit differential histone methylation and acetylation at maizeimprinted genes. Plant J. 56, 903–912
60 Kohler, C. et al. (2005) The Arabidopsis thaliana MEDEA Polycombgroup protein controls expression ofPHERES1 by parental imprinting.Nat. Genet. 37, 28–30
61 Makarevich, G. et al. (2006) Different Polycomb group complexesregulate common target genes in Arabidopsis. EMBO Rep. 7, 947–
95262 Feil, R. and Berger, F. (2007) Convergent evolution of genomic
imprinting in plants and mammals. Trends Genet. 23, 192–19963 Haig, D. (2004) Genomic imprinting and kinship: how good is the
evidence? Annu. Rev. Genet. 38, 553–58564 de Jong, T.J. and Scott, R.J. (2007) Parental conflict does not
necessarily lead to the evolution of imprinting. Trends Plant Sci. 12,439–443
65 Josefsson, C. et al. (2006) Parent-dependent loss of gene silencingduring interspecies hybridization. Curr. Biol. 16, 1322–1328
66 Walia et al. (2009) Dosage-dependent deregulation of an AGAMOUS-like gene cluster contributes to interspecific incompatibility.Curr. Biol.19, 1128–1132
67 Kawabe, A. et al. (2007) High diversity due to balancing selection in thepromoter region of the Medea gene in Arabidopsis lyrata. Curr. Biol.17, 1885–1889
68 Miyake, T. et al. (2009) Possible diversifying selection in the imprintedgene, MEDEA, in Arabidopsis. Mol. Biol. Evol. 26, 843–857
69 Spillane, C. et al. (2007) Positive darwinian selection at the imprintedMEDEA locus in plants. Nature 448, 349–352
70 Kermicle, J.L. (1978) Imprinting of gene action in maize endosperm. InMaize Breeding and Genetics (Walden, E.B., ed.), pp. 357–371, Wiley &Sons
71 Crouse, H.V. (1960) The controlling element in the sex chromosomebehaviour in Sciara. Genetics 45, 1429–1443
72 Kermicle, J.L. (1970) Dependence of the R-mottled aleurone phenotypein maize on mode of sexual transmission. Genetics 66, 69–85
73 Kermicle, J.L. (1970) Dependence of the r-mottled aleurone phenotypein maize on mode of sexual transmission. Genetics 66, 69–85
74 Randolph, L. (1935) Cytogenetics of tetraploid maize. J. Agr. Res. 50,591–595
75 Cooper, D. (1951) Caryopsis development following matingsbetween diploid and tetraploid strains of maize. Am. J. Bot. 38,702–710
76 Kermicle, J.L. (1971) Pleiotropic effect on seed development of theindeterminate gametophyte gene in maize. Am. J. Bot. 58, 1–9
77 Lin, B-Y. (1984) Ploidy barrier to endosperm development in maize.Genetics 107, 103–115
78 Surani, M.A. et al. (1984) Development of reconstituted mouse eggssuggests imprinting of the genome during gametogenesis. Nature 308,548–550
79 Chaudhuri, S. and Messing, J. (1994) Allele-specific parentalimprinting of dzr1, a posttranscriptional regulator of zeinaccumulation. Proc. Natl. Acad. Sci. U. S. A. 91, 4867–4871
80 Danilevskaya, O.N. et al. (2003) Duplicated fie genes in maize:expression pattern and imprinting suggest distinct functions. PlantCell 15, 425–438
81 Gutierrez-Marcos, J.F. et al. (2004) maternally expressed gene1 is anovel maize endosperm transfer cell-specific gene with a maternalparent-of-origin pattern of expression. Plant Cell 16, 1288–1301





























