Embed Size (px)
Citation preview

Copyright 0 1988 by the Genetics Society for America
Partial Revertants of the Transposable Element-Associated Suppressible Allele white-apricot in Drosophila melanogaster: Structures and
Responsiveness to Genetic Modifiers
Stephen M. Mount,*'? Melvin M. Green?: and Gerald M. Rubin* * Department of Biochemistry, University of California, Berkeley, California 94720, $Department of Genetics, University of California,
Davis, California 95616 and tDepartment of Biological Sciences, Columbia University, New York, New York 10027 Manuscript received July 17, 1987
Revised copy accepted October 8, 1987
ABSTRACT The eye color phenotype of white-apricot (w'), a mutant allele of the white locus caused by the
insertion of the transposable element copia into a small intron, is suppressed by the extragenic suppressor suppressor-of-white-apricot (su(wa)) and enhanced by the extragenic enhancers suppressor- of-forked s u o ) and Enhancer-of-white-apricot (E(w")) . Derivatives of w" have been analyzed molecularly and genetically in order to correlate the structure of these derivatives with their response to modifiers. Derivatives in which the copia element is replaced precisely by a solo long terminal repeat (sLTR) were generated in vitro and returned to the germline by P-element mediated transformation; flies carrying this allele within a P transposon show a nearly wild-type phenotype and no response to either su(f) or su(wn). In addition, eleven partial phenotypic revertants of wn were analyzed. Of these, one appears to be a duplication of a large region which includes w", three are new alleles of su(w'), two are sLTR derivatives whose properties confirm results obtained using transformation, and five are secondary insertions into the copia element within w'. One of these, w'"'*~'', differs from wn by the insertion of the most 3' 83 nucleotides of the I factor. The five insertion derivatives show a variety of phenotypes and modes of interaction with su((f) and su(w'). The eye pigmentation
IS affected by su(f) and E(w") , but not su(wa). These results demonstrate that copia (as opposed to the interruption of white sequences) is essential for the W" phenotype and its response to genetic modifiers, and that there are multiple mechanisms for the alteration of the W" phenotype by modifiers.
of WaR84h '
G ENETIC suppression in Drosophila is charac- terized by suppressor (and enhancer) mutations
which can modify the phenotype of alleles at diverse loci. Many suppressible alleles are of spontaneous origin and associated with the insertion of retrovirus- like transposable elements into noncoding DNA (flanking sequences or introns) within a gene. The genetic modifiers of such alleles are most often recessive, and primarily affect the phenotype of mutations caused by the insertion of a particular transposable element. For example, suppressor-of- Hairy-wing [su(Hw); 3-54.81 acts almost exclusively on insertions of the transposable element gypsy (Mo- DOLELL, BENDER and MESELSON 1983). Suppression of mutations at the forked [f;1-56.71 (MCLACHLIN 1986; PARKHURST and CORCES 1985, 1986b), yellow F; 1-0.01 (PARKHURST and CORCES 1986a), Hairy-wing [Hw; 1-0.01 (CAMPUZANO et al. 1986), white [w; 1-1.51 (LEVIS, O'HARE and RUBIN 1984; ZACHAR et al. 1985), and vermilion [v ; 1-33.01 (SEARLES and VOELKER 1986; WALKER, HOWELLS and TEARLE 1986) loci of Droso- phila melanogaster has been described at the molecular level, but in no case has the mechanism of suppression been elucidated definitively. Available information
Genetics 118: 221-234 (February, 1988)
indicates that suppressor loci encode factors which function at such stages of gene expression as tran- scription or RNA processing (for reviews of suppres- sion in Drosophila, see CHANG et al. 1986 and KUBLI 1986).
Mutations at the white locus affect the pigmentation of eyes, ocelli, Malpighian tubules, and testes, with eye pigmentation in a number of white alleles varying between bleach white and the dark red of wild type (LINDSLEY and GRELL 1968). The white-apricot (u") mutation, which arose spontaneously in 1923 (LIN- DSLEY and GRELL 1968), causes a change in eye color from wild-type red to orange-yellow, and was the first Drosophila mutation shown to be associated with the insertion of copia or a related element (GEHRING and PARO 1980; BINGHAM and JUDD 1981; BINGHAM, LEVIS and RUBIN 1981; GOLDBERG, PARO and GEHRING 1982). The mutant phenotype is caused by the pres- ence of copia in a 74 nucleotide intron, oriented such that the white gene and copia are transcribed in the same direction (O'HARE et al. 1984).
The two most abundant polyadenylated transcripts from wa (other than those which might be derived from sequences entirely within copia) are comparable

222 S. M. Mount, M. M. Green and G. M. Rubin
in abundance to w + RNA, are detected by hybridi- zation probes upstream but not downstream of the insertion, and appear to have 5' termini correspond- ing to that of wild-type white RNA but 3' termini within copia (see Figure 1). It is likely that the residual expression characteristic of wa is due to the generation of small amounts of structurally normal mRNA after the removal of copia by splicing, and such RNA has been detected (LEVIS, O'HARE and RUBIN 1984; PIR- ROTTA and BROCKL 1984; ZACHAR et al. 1985). Thus, w" transcripts initiated at the white promoter are generally polyadenylated at a site, or sites, within copia, and the wa phenotype appears to be caused by whatever molecular events lead to the production of these aberrant RNAs at the expense of RNA with the wild type structure.
The mutant phenotype of wa is enhanced (pigmen- tation is further reduced) by viable recessive alleles of suppressor-of-forked [ s u o ; 1-65.91 and by the dom- inant mutation Enhancer-of-white-a~rzcot [E( w") ; 2- - 1061, but suppressed (pigmentation is increased) by recessive alleles of suppressor-ofwhite-apricot [m(wa) ; 1-0.11 (LINDSLEY and GRELL 1968). Mutations at su(f) suppress a number off alleles, at least some of which are associated with the presence of the transposable element gypsy and are also suppressed by su(Hw) (MODOLELL, BENDER and MESELSON 1983). The effects of m ( w a ) and su(f) are specific for wn among white alleles, implying that these are not simply regulators of white expression. Lethal alleles of su(f) (DUDICK, WRIGHT and BROTHERS 1974; RUSSELI. 1974), and possibly su(wu) (ZACHAR et al. 1985) have been iden- tified. These properties are consistent with the pro- posal that these modifiers play important, even es- sential, roles in gene expression.
The transposable element copia is a member of the large class of structurally homologous genetic elements that includes the transposable T y elements of yeast (ROEDER and FINK 1983), vertebrate retroviral proviruses (VARMUS 1983), and a number of distinct Drosophila transposable elements, including most of those found associated with suppressible mutant al- leles (RUBIN 1983). Complete nucleotide sequencing of two independent copia elements, including the element at wa, found them to be virtually identical (differing at seven noncoding positions in 5146 nu- cleotides) and confirmed the similarity of copia to retroviruses and Ty with regard to coding potential (EMORI et al. 1985; MOUNT and RUBIN 1985). The conserved feature of this class of elements most relevent to this work is their transcription pattern: initiation occurs in the 5' LTR (FLAVELL et al. 198 1) and polyadenylation in the 3' LTR (EMORI et al. 1985).
An important step in the elucidation of the mech- anism of action of genetic modifiers of w' is the identification of sequences within copia which are responsible for these effects. Here we report results
correlating structural alterations of copia with alter- ations in the phenotype and genetic interactions of w". We have found that sequences within copia but not in the LTR are essential for both the mutant phenotype of wa and its response to modifiers, and that insertions within copia can disrupt these phe- nomena in a variety of ways. In particular, the existence of a partial revertant ( w " ~ ' " ~ ) which is sensitive to su(f) and E(w") , but not su(w0), implies that these modifiers have distinct sites of action.
MATERIALS AND METHODS
Stocks: The genotypes and sources of the fly strains used in these experiments are given in Table 1. Recombi- nants described in the text were derived from crosses among the starting strains listed in Table 1 and standard ~ ~ ~ ' ~ f ' s u ( ( f ) , su(wfl) will8 and y su(wa) w"" strains. wI1l8 is a hleach-white allele of white with a deletion of sequences at the 5' end of the locus, including the entire first exon (HAZELRIGG, LEVIS and RUBIN 1984). w f f l X f' su((f) was derived from wh'f5, w l f l X and w" f55u(f) stocks via a w " " ~ f' recombinant. su(w") will8 was derived from su(w") w" and wl"*. y su(w") wl l lX was derived from su(w") w l l l N and y w ' ~ . Recombinations involving SU(W") [ 1-0.11 were followed using y [ 1-0.01 as a marker and confirmed by complemen- tation tests or by regeneration of su(w") w". Recombinations involving su(f) were followed using f ' to identify the su( (f) allele. All recombinants were recovered using the C(I)DX, y wfand FM7c stocks. The effects of E(w") were generally studied by mating males with the genotype of interest either with C( I)RA85, y wrf5; Cy0 lE(w") females (compar- ing Cy and Cy' male progeny) or with wf118; Cy0 lE(w") females (comparing Cy and Cy' female progeny).
wu derivatives: A number of partial revertants of W" are described in the literature (LINDSLEY and GRELL 1968; RASMUSON, GREEN and EWER-ISON 1960) and more have been isolated recently (this work). A brief introduction to each of the w" derivatives or putative W" derivatives ex- amined in this study is appropriate.
(RASMUSON, GREEN and EWERTSON 1960) was in- duced by x-rays, and has recently been shown by sequence anal sis (CAKBONARE and GEHRING 1985) to represent a
as spontaneous mutations by MOSSICE (described in RAS- MUSON, GREEN and EWERI-SON 1960), MULLER (1944), BIR- CHLER (1984) and MOUNT (1984), respectively. The origin
is described below. wnr arose from an insertion of the retrovirus-like transposable ele- ment roo within copia, at nucleotide 2195, oriented such that the direction of its transcription is identical to that found in copia and white; the unmodified phenotype of w'" is not distinct from that of w", and the difference between these two alleles was discovered by Southern analysis (DAVIS, SHEN and J U U U 1987).
Removal of wr from the Muller-5 balancer and gener- ation of C(l)RA85, y w'f': wlarose in 1944 as a spontaneous revertant of w' on Zn(l) sc8Zn( I ) S , sc8 w" B (MULLER 1944). It was liberated from the doubly inverted chromosome for use in these studies by recombination in In( I ) scam( 1 ) S, scX w'ly w11i8 f'; CyOl+; T M 3 / + ; females mated to y w1'18
f' males. C(I )RA85 , y w' f 5 , which behaves as a stable compound X , also arose from these females, presumably by recombination within heterochromatin (NOVITSKI 1963). The chromosome was originally heterozygous at f, but a f' homozygote has been obtained (heterozygosity for W'
WaH 5YkI
w~Rd?TR derivative. WnRM, wr, waR84h and W n R X 4 ~ 7 were isolated
of W~R79127 and WaR84r19(13)

Reversion and Suppression of wa 223
TABLE 1
D. melanoguster stocks used in this work"
Genotype Sourceb Notes
wbf f BG Y ufp BJ su(w0) w= BG
W"f s u o RL W a BG
C(I )DX, y w f RL FM7cIKAl4 RL I n ( I ) SF I n ( I ) S, sc8 w' CT
w"'~; CyOlnub b Sco It s t d RL w""; TM3, p' b 2 4 e e" Sblln(3LR)CxD RL C( I ) R M , y w flXY".Y", y wa J B C ( W f 4 , f LC XYL.YS9 w'118 RL y wa/bw+ Y; E(wa)lCyO CT WdM DV
wa4 BG We BG dP55 CT f Wd RL up3 BG WCf RL
JR4 DT sc JR5 DT sc JR6 DT sc z wu RL dal BG sc z W'I RL sc z w'" RL gt wa DV
c -
w I 1 1 8 RL - d
-
W a R 5 9 k I DV
gt W i + A sf1 DV -R h
h
h
- - -
a Descriptions of genotypes, unless footnoted, can be found in LINDSLEY and GRELL (1968).
Abbreviations are as follows: BG, the Mid-America Drosophila Stock Center; BJ, BURKE JUDD; RL, Rubin laboratory; CT, the California Technology Drosophila stock center; JB, JIM BIRCHLER; LC, LORING CRAYMER; DV, Green laboratory; DT, DANIELLE THIERRY-MIEG.
MERRIAM (1968). HAZELRIGG, LEVIS and RUBIN (1984). R. LEVIS (unpublished data).
~ Z A C H A R and BINGHAM (1982). g KARESS and RUBIN (1982).
BUCHETON et al. (1982).
In(1) S on one arm and wl'" on the other is maintained). The reversed acrocentric structure and continued hetero- zygosity for In( I ) S has been confirmed by examination of polytene chromosomes (J. YEUNC and T. LAVERTY, personal communication).
Isolation of wd79127 and w ~ ~ ~ ~ ~ ~ ( ~ ~ ) - . Th ese were de- tected as phenotypic revertants among the progeny of females heteroallelic for gt (f sc gt""/gt w"), a condition which appears to be mutagenic (GREEN 1982). Three other independent alleles isolated in this manner appeared iden- tical to wQR79(27 In ' phenotype, interactions with su(wa) and su(f), and preliminar Southern analysis, but have not been analyzed further. wa'84e'9('3) appears to be unique among the five wa revertants derived from this cross. (A complete description of mutagenesis bJ gt heteroalleles will be pub- lished elsewhere; only and w ~ ~ ~ ~ ~ ' ~ ( ' ~ ) are consid- ered here.)
Eye pigmentation comparisons: Eye coloration was ex- amined in flies which had been raised entirely at 25" and aged for five to eight days following eclosion. The numer- ical scale used in Table 3 was developed starting with the principle that the difference in pigmentation between a fly carrying two copies of a particular white allele and one heterozygous for that allele and the deletion w"18 should be 1.0, with a lower number indicating less pigmentation. In each of two experiments (a and b), all flies were raised simultaneously and compared inter se (see Table 3). Geno- types with widely disparate scores were not generally com- pared, but all distinct genotypes with identical scores were directly compared and found to be identically pigmented, and all pairs of genotypes with similar scores (a difference of less than 1.0) were directly compared to verify the direction and approximate extent of difference indicated by the score. It was empirically observed that the numerical difference between homozygotes and w"" heterozygotes sometimes varied from 1.0. However, confidence in this scale is based on analysis of a large number of phenotypes, including many whose relative level of white expression is known, which reveals an unbroken continuum from white eyes to wild type eyes. Only in the case of w~~~~~~~ were there qualitative differences making it difficult to rank all genotypes on a common scale. I t was consistently observed that, as the level of white expression increased, the pigmen- tation first grew darker in color up to the brown tones characteristic of a score of about 5.5 on this scale. Beyond that, increased white expression led to a redder coloration, typical of wild type. In fact, the phenotype which corre- sponds to a 4.5 on this scale (e.g. , w'"'~/w' ' '~) is similar in color tone to those with scores above 7 (which are indistin- guishable from wild type). However, these two phenotypes are easily distinguished by the presence of an "eye spot" in those flies with the higher levels of expression. Previous experience with pigment assays indicates that careful direct comparison as performed here is more reliable, particularly for genotypes with low levels of white expression (see LEVIS, HAZELRIM and RUBIN 1985b).
Construction of theP[wddTR] allele: p3922a15 (MOUNT and RUBIN 1985) was opened by partial digestion with BalI, which cleaves within the copia LTR at nucleotide 70. The appropriately sized fragment was isolated from low-melting agarose and religated to form p3922.LTR. Regeneration of the Ball site by precise religation was verified by complete digestion of this plasmid with BalI. Next, pClLTR.2 was generated by replacement of the wild-type SacI-XbaI frag- ment (white +432 to -600) of pC1 (LEVIS, HAZELR~CC and RUBIN 1985a) with the corresponding fragment from p3922.LTR. pCILTR.2 therefore contains a single copia LTR at the precise location of the original copia insertion, now in the context of a complete W + gene (-5.1 to +6.6) in the P element vector pCIW4 (RUBIN and SPRADLINC 1983).
Drosophila transformation: The preparation of DNA and microinjection into embryos were as described in SPRADLING and RUBIN (1982). The P[waRSLTR] allele itself was used as a dominant visual marker following injection of wll" flies with pC1.LTR and ~ 1 1 2 5 . 1 ~ ~ (KARESS and RUBIN 1984). The subsequent isolation of isochromosome lines was as described in LEVIS, HAZELRICG and RUBIN (1985a). Transformants were observed among the progeny of three of 35 fertile w'"~ adults resulting from these injections, and strains were established from a single fly derived from each of these three.
Mobilization of the P[wdATR]24 transposon: The P[waRSLTR]24 insertion confers a low level of pigmentation, and is lethal when homozygous or hemizygous. The eye

224 S. M. Mount, M. M. Green and G. M. Rubin
8' E'
Probes:
RNAs:
5.7
I C
- D
+
2.6
1.25
0.9
+ -I"
?
+ I
+ I
+
1 kb
color of progeny of willN I ~ [ ~ ~ ' ~ ~ ' ~ - ~ ~ ~ ] 2 4 , wI1Ix. , P[hsp;w-l/ + females was examined. Transpositions of P [ w ' " " ~ ~ '1 from 2C1 were detected by either pigmentation in males (which would not normally inherit P [ ~ " ~ " ~ * ' ~ ] 2 4 due to its associ- ated lethality) or increased igmentation levels in females. Transpositions of P[waR51,Tq were due to the activity of P [ h p ; w - ] , a single complete P element derived from P[hp;w+] (LASKI, RIO and RUBIN 1986) by F. SPENCER (unpublished results). That only one P element was present in the w"18; P [ h p ; w - ] strain at the time of this experiment was indicated by Southern analysis of a parental wl'IN; P [ h p ; w - ] stock several generations prior to this experiment (F. SPENCER, unpublished data), and the absence of subse- quent instability (see below). However, later attempts to use precisely this stock in similar experiments revealed P activity mapping to multiple chromosomes. In this exper- iment, P [ h p ; w - ] was present on the third chromosome,
mated to will8; TM3,SbICxD males to facilitate rapid sep- aration of the intact P element from any newly induced transpositions. Twenty-five out of 54 single-female matings gave rise to a least one reddish rogeny, and all but 2 of 18 lines carrying new P[waRALTd; insertions on the X or second chromosomes were stable after isolation.
P[Wd" Anal sis of the effect of an extra Y chromosome on ' $22: To analyze the effect of an extra Y chro-
mosome, wI1l8; ~ [ w ~ ~ ~ ~ ~ ~ 121 males were first mated to C ( I ) R M , y w f l 0 females from a C ( Z ) R M , y w f l XUL . Ys y wa stock. The resulting C ( I )RM, y w f 1 Y; P[wUXrLTR]21 i + females were then mated to either XYL . Ys, w ~ " ~ / Y or XYL . Y', will8/ 0 males to generate C ( I ) R M , y w f I 0;
females, whose pigmentation was compared. Molecular biology: RNA blots were performed using
single-stranded probes as described (O'HAKE, LEVIS and
and w1118 1 pIWaRsLTR 124 w 1 I I 8 ; P [ h p ; w - ] I + females were
p[WaR,'LTR 121 I + and C ( l ) R M , y w f I Y ; P [ w ~ ~ ~ ~ - ~ ~ 121 / +
+ I
+ m
+ I
+ I
RUBIN 1983).
A
FIGURE 1 .-Structure of the white-apricot allele and its transcripts. Exons of the wild- type white mRNA are indicated by black boxes; the copia insertion is indicated by stippled box, except for the long terminal repeats, which are indicated by white boxes. Immediately below the map are shown re- striction fragments used as hybridization probes A through F. Below that is a listing of observed transcripts from white-apricot to- gether with their hypothesized structures (based on data in LEVIS, O'HAKE and KUBIN 1984; P I R R O J T A and BROCKL 1984; Z A C H A K
et al. 1985; and this work). A plus or minus sign indicates whether each transcript is de- tected by each of the three hybridization probes A through C (Figure 5 and data not shown). Restriction sites are: A, Apal; B, BamHI; H, HindIII; Hp, HpaI; Ps, PstI; Pv, PvuI; Sc, SacI; SI, SalI; Xb, XbaI; and Xh, XhoI.
Figure 5 shows blots probed with DNA synthesized from-an XbaI-Nad fragment (B in Figure I) , white nucleotides +68 to +432, cloned in M13mp19.
Isolation of DNA and Southern blots were performed as described previously (HAZELRIGG, LEVIS and RUBIN 1984). The copy number and structure of P[waRSLTR] insertions was determined (data not shown) by digestion of DNA from transformed flies with BamHI plus SacI (revealing the copy number) and HindIII plus SacI (revealing any rearrangements within the transposon); the probe used was lambda m 2.2, which encompasses the entire region shown in Figure 1 (white coordinates -5.8 to +5.5; LEVIS, BINGHAM and RUBIN 1982). The structure of w" derivatives was analyzed (always in arallel with w") using the following digests and probes: waK P and w~~~~~~ were digested with XbaI and SacI and probed with pm12.5 (white coordinates + 3.17 to - 0.67; D in Figure 1). w ~ ~ ~ ~ ~ ~ , UnRM and waRSyk1 were digested with BamHI plus EcoRI and BamHI alone and probed with pm12.5 (D). w ~ ~ ~ ~ ~ ~ , wCtRM and WaR5Ykl
were digested separately with Sac1 and HindIII and Smai and probed first with pm12.5 (D) and later with pm12.3 (white coordinates - 1.5 to - 0.67; E in Figure 1). w ~ ' ~ ~ ~ ~ ~ ,
with lambda m 2.2. w~~~~~~ was analyzed alongside
with lambda m 2.2. w ~ ~ ~ ~ ~ ~ , waRM and wnRgYk1 were digested separately with SacI and HindIII and SmaI and probed first with 11112.5 (D) and later with pm12.3 (E). w ~ ~ ~ ~ ~ ~ ,
and wr were digested with HindIII plus Sal1 and probed first with lambda m 2.1 (white coor- dinates - 5.0 to + 9.5) and later with pm 12.5. wnR7y127 and w ~ ~ ~ ~ ' ~ ~ ~ ~ ) were digested with EcORI plus BamHI plus SacI and probed first with lambda m 2.1 and later with pm12.5
probed first with lambda m 2.1 and later with pm12.5.
WaRM and WaRSYkl were digested with HindIII and probed
p[waRJLTR ] by digestion with BamHI and SacI and probing
WaRM w&ge7 WaR84h
(D). Wa1z7y127 was digested with EcoRI alone and A d 1 and

Reversion and Suppression of w" 225
WaR79L27 WaRM and WaR84e19(13) were digested with HindIII plus ApaI, plus NarI, plus HpaI or HindIII alone; waRM
AccI or SalI plus BamHI plus ApaI; waR 9127 was digested with Sal1 plus BamHI plus HpaI, or SalI plus BamHI plus PstI, or SalI plus BamHI plus StuI; wURM was digested with BalI, ApaI or NarI plus StuI; these digests were all probed with single stranded radiolabeled DNA from the BamHI- SalI fragment of white (coordinates + 1.4 to - 0.7; F in Figure 1).
For sequencing, w~~~~~ was cloned as an XhoI fragment in the SalI site of lambda Charon 35 (LOENEN and BLAIINER 1983), subcloned as a BamHI-Sal1 fragment (white coordi- nates + 1.4 to -0.7) in pBR322 and sequenced by shotgun cloning random subfragments of the 1 kb. HincII fragment (copia nucleotide 4777 to white nucleotide -675) into m13mp19 (see BANKIER and BARREL 1983).
and WaR84rI ( 1 3 ) were digested with Sal1 lus BamHI plus ?
RESULTS
Transformation with a solo LTR derivative of wa: We originally sought to study suppression phenom- ena in a simplified system by replacing the copia element of W" with a single copia LTR. Polyadeny- lation of copia RNA occurs within the 3' LTR (Emori et al. 1985). Because polyadenyiation within copia is likely to be responsible for the mutant phenotype of w", and possibly for its interaction with modifiers as well, it was of interest to see whether these properties required sequences other than those within the LTR. A derivative of w" in which copia is replaced by a single LTR was generated from cloned w" DNA by precise in vitro recombination between the two LTRs at a BalI restriction site. The solo LTR derivative was inserted into a P element vector and three indepen- dent germline transformants were recovered (see MATERIALS AND METHODS). These will be referred to as carrying the insertions P[wnRaLTR]3, P[w"R'i.?'R 12 1 and P[wURJLTR]24 (abbreviated V3, V21 and V 2 4 ) . Genomic DNA blotting experiments (see MATERIALS
AND METHODS) and in situ hybridization (see Table 2) revealed that each of these strains carries one un- rearranged copy of P[waRJLTR].
V 3 , an insertion on the second chromosome, con- ferred strong pigmentation: homozygous males were wild type in appearance; females and heterozygous males were almost so (less pigmentation in females is the expected result of dosage compensation; HA- ZELRIGG, LEVIS and RUBIN 1984). These properties are typical of P[waRALTR] transposons. Distinct pheno- types were observed in V21 and V24 , but were shown to be caused by position effects; a description of their properties and the evidence for this conclusion is presented in a later section.
Structure of additional wa derivatives: Partial rev- ertants of wa, namely w ~ ~ ~ ~ ~ ~ ~ , wr, wURM, w ~ ~ ~ ~ ~ ~ ~ ~ ( ~ ~ ) , @79127, and w ~ ~ ~ ~ ~ , were analyzed by Southern gen- omic blot analysis using probes from the wild type
TABLE 2
Transduced wddTR transposons ~~ ~~~~
Abbreviation position" Eye color Cytological
v 3 v 2 1
V24 V24- 12 V24-20C
V24-20D V24-35 V24-55 V24-B4B V24-B8
47D 81F
2c 1 2 c 1 84F
52A 60D 2 c 1 34A(puff) 2 c 1
Reddish Variable and
variegated Dark apricot Reddish Yellow, red
spots Reddish Reddish Reddish Reddish Reddish
a Assignments by T. LAVERTY (personal communication).
(Canton S) allele (see Figure 2 for sample data and MATERIALS AND METHODS for experimental details).
the basis of co-migration of 2.3-kb SacI-BamHI frag- ments (from coordinates - 0.60 to + 1.38) in DNA
after, the sequence of this region from w~~~~~~~ was published, confirming this suspicion (CARBONARE and GEHRING 1985). wr is believed to be a waRsLTR deriv- ative on the basis of the co-migration of 4.1-kb SalI- HindIII fragments (from coordinates -0.67 to + 3.17) in DNA from wr and P[wURLTR] stocks (com- pare lanes 2 and 3 in Figure 2A; this fragment is 3.8 kb in wild-type strains, and is split into two fragments of 5.4 and 3.4 kb in w").
comparison to wfL following digestion with enzymes having recognition sites within copia and hybridiza- tion with white probes flanking copia (see Figure 2 for an example and MATERIALS AND METHODS for details). wURM was found to be an insertion of roughly 2.3 kb into the copia element at w", between the BaZI site within the 5' LTR at copia nucleotide 70 and the ApaI site just outside of the 5' LTR at copia nucleotide 288. Thus, it is likely that this insertion is within the 5' LTR (which extends from copia nucleotide 1 to copia nucleotide 276). w ~ ~ ~ ~ ~ ~ ~ ( ~ ~ ) was found to be an insertion of roughly 6 kb between the AccI site at copia nucleotide 1348 and the HindIII site at copia nucleotide 2253. waR79127 was found to be an insertion of roughly 4.5 kb between the PstI site at copia nucleotide 4038 and the HpaI site at copia nucleotide 4777. These results are summarized schematically in Figure 3.
Sequence of the w~~~~ insertion: w~~~~~ was orig- inally observed to be associated with a very slight alteration relative to w" in the mobility of the HindIII- Sal1 fragment containing the 3' end of copia (copia nucleotide 2253 to white coordinate -0.67). T o in-
WuR59ki was suspected to be a WaRsLTR derivative on
from WuR59kI and P[waRJLTR] stocks. Shortly there-
WaRM WaR84e19( i3) and wuR79127 were analyzed by
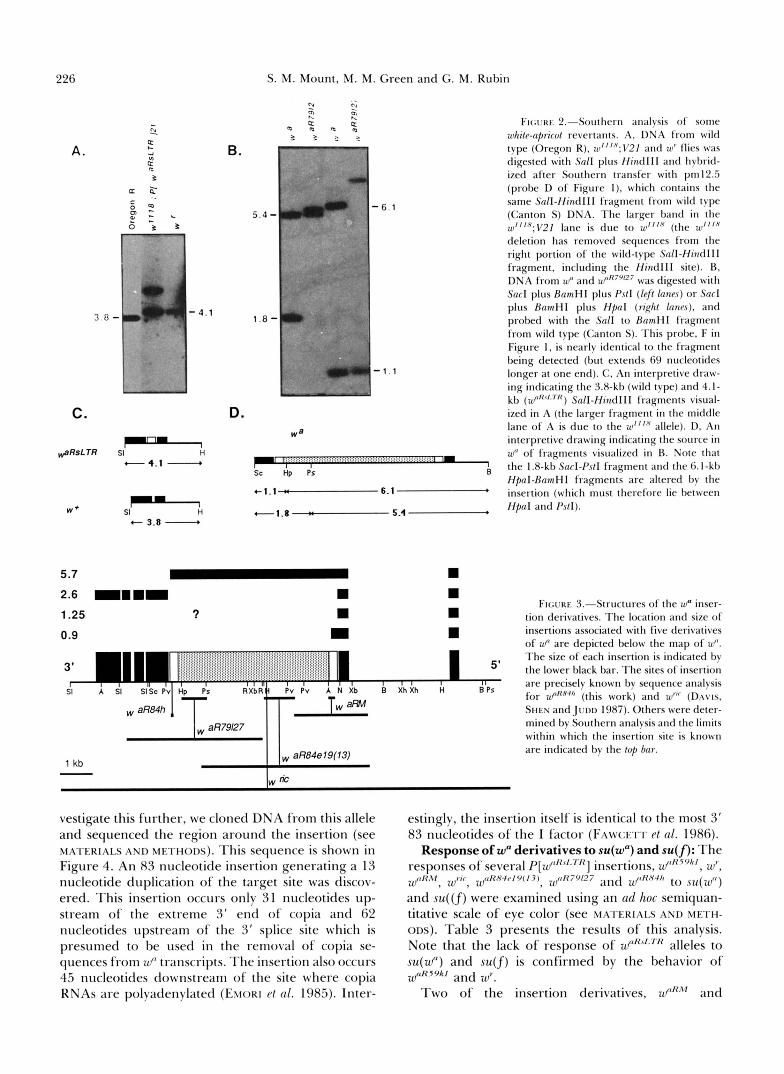
226 S. M. Mount, M. M. Green and G. M. Rubin
A. I
: 4 v1
C. - d R s L T R SI H - 4.1 -
W * SI - H
+ 3.8 -
B.
5 .4
1.8
D.
- 6.1
- 1 . 1
I 1 B Sc Hp Ps
........................................................................... ,I
61.1 :: 6.1 *
FIGURF. 2.-Southern analysis of some u!hite-nprico/ revertants. A, DNA from wild type (Oregon K), 10"'~:V21 and ulr flies was digested with Snll plus HindIII and hybrid- ized after Southern transfer with pm12.5 (probe D of Figure I), which contains the same Salt-HindIII fragment from wild type (Canton S) DNA. The larger band in the
deletion has removed sequences from the right portion of the wild-type SnlI-HindlII fragment, including the HinclIlI site). B. DNA from ui' and UP^^^'^^ was digested with Sac1 plus RnmHI plus PsfI (left /ones) or Sac1 plus RnmHI plus HpnI (right lanes), and probed with the Sal1 to RnmHI fragment from wild type (Canton S). This probe, F in Figure I , is nearly identical to the fragment being detected (but extends 69 nucleotides longer at one end). C, An interpretive draw- ing indicating the 3.8-kb (wild type) and 4.1-
ized in A (the larger fragment in the middle lane of A is due to the w"'"' allele). D, An interpretive drawing indicating the source in ui' of fragments visualized in R. Note that the 1.8-kb SncI-PstI fragment and the 6. I-kb HpnI-BnmHI fragments are altered by the insertion (which must therefore lie between HpaI and PstI).
?"I I / x . ,V21 lane is due to u!"" (the u~""
kb (&*/<>I.'//< ) SnlI-Hind111 fragments visual-
FIGURE 3.-Structures of the w" inser- tion derivatives. The location and size of insertions associated with five derivatives of up are depicted below the map of uf'. The size of each insertion is indicated by
5' the lower black bar. The sites of insertion are precisely known by sequence analysis for dRx4" (this work) and w''~ (DAVIS, Stlm and J U D D 1987). Others were deter- mined by Southern analysis and the limits within which the insertion site is known are indicated by the top bnr.
;ips
aR84e19(13) 1 kb -
W k
vestigate this further, we cloned DNA from this allele and sequenced the region around the insertion (see MATERIALS AND METHODS). This sequence is shown in Figure 4. An 83 nucleotide insertion generating a 13 nucleotide duplication of the target site was discov- ered. This insertion occurs only 31 nucleotides up- stream of the extreme 3' end of copia and 62 nucleotides upstream of the 3' splice site which is presumed to be used in the removal of copia se- quences from w'' transcripts. The insertion also occurs 45 nucleotides downstream of the site where copia RNAs are polyadenylated ( E M O K I et 01. 1985). Inter-
estingly, the insertion itself is identical to the most 3' 83 nucleotides of the I factor (FAWCEIT e& al. 1986).
Response of wa derivatives to su(wa) and su(f): The responses of several P[w~~"\ '~? '~ ' ] insertions, wnH5Vkl , w', Wnf<*\l w r ~ c , u,nRX4clY(13) W~~f~7Y127 and wnRXJh to su(w'I)
and su((f) were examined using an ad hoc serniquan- titative scale of eye color (see MATERIALS AND METH- ODS). Table 3 presents the results of this analysis. Note that the lack of response of alleles to su(wf') and su(f) is confirmed by the behavior of
Two of the insertion derivatives, w''~'''' and Wr~R59kl and wr.

Reversion and Suppression of w" 227
GGTCGTGGTG CTGGTGTTGC A G T T ~ T A A C T T ~ T A T A A A T ~ T M FIGURE 4,"Sequence of the wYRXJh insertion. The se- copial quence of the w'~"' ' allele from copia nucleotide 5029
TAAACTTGAC TATTTATTTA TTTATTqBEB AAGGAAATAT et al. 1984) is presented. T h e 83 nucleotide insertion labeled (MOUNT and R U B I N 1985) to white nucleotide -36 (O'HAKE
[I element "I element" is identical to the 3' end of the 1 factor in dRl TATTATTATT ATTATTATTA TGATAGATAG AATAGTTTAC A A A A C T W (FAWC:KI-I et al. 1986; nucleotides 5297-5375) except that
I element] [copia the TAA repeat is four nucleotides longer in waRX4" than in TAACTT-T AGTAT-T T ~ T A c ~ ET- W U ~ T w"' and the opposite strand is indicated. The 13 nucleotide
target site duplication surrounding the I element insertion laon is underlined. The site of polyadenylation of copia tran-
*>>
copia J [white AAcATAAAGG GTccAATTAC cAATTTGAAA CTmTTTGC scripts ( E M O K I et a[. 1986) is indicated by carets.
W a R 8 4 e / Y ( / 3 ) , do respond to both SU(W") and s u o ,
suggesting that although these reversion events have affected the phenotype of w", they have not signifi- cantly interfered with the sequences necessary for the interaction of W" with the products of the su(w") and su(f) loci. wnC, a derivative which does not differ from w" phenotypically, shows a clear but lessened response to both su(w") and s u o . The effect of these modifiers on w~~~~~~~ was examined in many exper- iments. Although response to both su(w") and su(f) was occasionally observed (Table 3 and data not shown), this was not always the case. Thus, it appears that in w ~ ' ~ ~ ~ ~ ~ the response to both su(w") and su(f) is largely, but perhaps not completely, eliminated. In contrast, the results with wfiH8"', a strong revertant, were unambiguous. Expression of w"'"~" is unaf- fected by S U ( W " ) , but is greatly reduced by s u o .
Enhancer of white-apricot: The allele specificity of E(w") was investigated in females heterozygous for various white alleles and wII1". In accordance with previous results (VON HAUE 1969; BIRCHLEK 1986), the white alleles a, a4, e and $155, but not +, u2, a3, bJ 4 i + A , IR4, IR5, IRG, i s , sat, sp', zl or zm showed markedly less pigmentation in combination with a single copy of E(w") . All known responsive alleles are associated with transposable element insertions into the transcribed region, and this pattern of interaction supports a restriction of the effects of E(w") to a subset of transposable element insertions at white. We tested the action of this dominant extragenic modifier on the various wn derivatives described here: the insertion derivatives w('RM, w~~~~~~~~ and w ~ ' ~ ~ ~ ~ ~ were affected by E(w") , but w ~ ~ ~ ~ ~ ~ ' , w' and several
results show that sequences in addition to the copia LTR are necessary for the response of w" to yet a third extragenic modifier.
Structure of polyadenylated RNA products: A number of studies on the structure of polyadenylated RNAs from wn and the effects of su(wf') and su(f) on these RNAs have been published (LEVIS, O'HARE and RUBIN 1984; PIRROTTA and BROCKI. 1984; ZACHAR et al. 1985). These analyses are complicated by the low abundance of white RNA, by the difficulty of obtain- ing RNA from the appropriate tissue (the eye disc) at the appropriate time in development (the early pupal period) and by the complexity of the RNA
P[W['K.'LTR ] transformed lines were unaffected. These
pattern itself; we have not attempted to reexamine the results of those studies, which are summarized in the introduction. However, we have looked at the gross effects on RNA structure of some of the rever- sion events described here.
Northern blot analysis of RNA from adult males (0-24 hr post-eclosion) has shown that the previously described wn RNAs of 5.7 and 1.25 kb are eliminated
see Figure 5, lanes 5 and 6 of the right panel; identical data were obtained for V? but are not shown). Also seen in these revertants are extremely low levels of a group of RNAs of approximately 900 nucleotides that hybridize to DNA probes covering both the first and second white exons. These RNAs appear on closer examination to be present in wn and all revertants examined, but are absent from wild type. Their size is consistent with polyadenylation within the 5' copia LTR. The level of these RNAs is not greatly affected by su(w") or su(f) (Figure 5 and data not shown). Thus, the polyadenylation activity of the 5' LTR, or a solo LTR, appears to be very low and unaffected by modifiers.
In the insertion revertant w " ~ ~ ~ ~ ~ ~ (right panel, lane 4) both the 5.7- and 1.25-kb RNAs are eliminated, and a band of wild type mobility is visible. In contrast, the 5.7-kb RNA, but not the 1.25-kb RNA, is elimi- nated in the insertion revertant waRM (lanes 3 and 4). In this case, however, the 5.7-kb RNA is replaced by an RNA of approximately 7.9 kb, the size expected if the waRM insertion were to be simply incorporated into the body of the 5.7-kb RNA. waRXJh (lanes 1 and 2) differs from w" only in the presence of increased levels of an RNA or RNAs in the 2.6-kb size range.
Position effects observed with P[wnRsLTR] inser- tions: Of the three original transformant lines, two, 1/21 and V24, had phenotypes different from V3 and from each other. These phenotypes appear to be due to the influence of sequences around the site of insertion.
V21 flies have variegated eyes. The insertion is present at the very base of the right arm of the third chromosome (Table 2), and variegation is presumably due to the proximity of the insertion to centromeric heterochromatin (SPOFFORD 1976). Interestingly, ex- treme variation in the overall level of pigmentation was observed among V21 flies, and this variation
in the WuR.sLTR derivatives studied (for w " ~ ~ ~ ~ '

228 S. M. Mount, M. M. Green and G. M. Rubin
TABLE 3
Phenotypes of white-apricot derivatives
W" 3.0".' 4.0".' 1.0".' 4.0".' 5.5"*' W"RM
2.00.'
w7ic 3.0' 3.7' 2.7' waR79127
4.5' 5.0' 3.0' 5.8' 6.3' 3.7'
4.0' #84~19(13) 3.7' 4.1' 2.0' 4.2' 5.7' 3.0'
4.4' 3.7' 4.3' 5.2' 4.3' 6.0' 6.4' 6.0' 5.5" 5.5" 3.4" 6.8" 6.8" 4.1" 6.1' 6.1' 6.1' >6.8' >6.8' >6.8'
w&84h
WaR59kl
W? 5.5" 5.5" 5.5. >6.8" >6.8" >6.8" P[W-=R]3 4.5Q.b 4.5..' 4.ja.' 5. 50.6 5 , 50.6 P[wnR'L=R]24
5.5".' 4.0" NA 4.0" SA SA
P [ f l L T R ] 2 4 - 3 5 5.0" NA 5.0" 6.3" S A 6.3" N A
W& refers to the white allele indicated at left. Superscripts a and b refer to two independent experiments. See text for an explanation of the numerical scale.
FIGURE 5.-Northern analysis of uto and some revertants. Po- lyadenylated RNA prepared from newly eclosed adults of the indicated genotypes was prepared, electrophoresed in agarose, transferred to nitrocellulose and hybridized as described (O'HARE, LEVIS and RUBIN 1983) using a "'P-labeled single-stranded probe prepared from an M13 clone containing the XbnI-Nor1 fragment of white (probe B of Figure 1). RNAs diagramed in Figures 1 and 3 are indicated. The W" samples in the two rightmost lanes of the left panel and the first and third lanes of the right panel are identical. Note (1) the absence of any strong effects of-either su(uf') or su((f) on RNA from this stage, (2) the overall similarity of preparations from W" and wf~"'', (3) the presence in u~"~"" of a 7.9-kb RNA, which is larger than the 5.7-kb RNA by the size of the insert found in that revertant, and (4) the presence of the 5.7- and 1.25-kb RNAs (or, in the case of w"~"', the corresponding 7.9- and 1.25-kb RNAs) in samples from W", w"~" ' and ufJRX4" , but not
or wild type (Oregon R) flies. The P [ W ' ' ~ ' ~ ~ ~ ] insertion V3 produces RNAs which are identical to those seen in
Wd779L27 WnR59kl
d R 5 9 k I (data not shown).
appeared to be strongly influenced by their precise genetic background. For example, when flies carrying the second chromosome balancer Cy0 and homozy- gous for V21 were backcrossed to W"", the W"";
Cy0 l+ ; V21 / + progeny were as a class much less pigmented than their w' I I8 ; +l+ ; V21 /+ siblings (greater than 90% of the Cy+ flies had more pigment than the median level of pigmentation among the
Cy0 flies). Similar results were obtained with other chromosome substitutions. In another experiment, selection in vials for ten generations, at a population size of three pairs of flies per generation, resulted in two V21 sublines, one of which had nearly bleach- white eyes with a small number of lightly pigmented spots and one of which was resembled V3, showing only slight variegation.

Reversion and Suppression of wa 229
Position effect mutations associated with chromo- somal rearrangements placing cell autonomous eu- chromatic genes under the influence of centromeric heterochromatin are generally responsive to in- creased temperature and the presence of additional Y chromosomes (both of these factors generally in- crease the amount of wild type tissue; see SPOFFORD (1976) for a review of position effect variegation). To see if V21 behaved like these rearrangements, the effects of extra Y chromosomes and increased tem- perature was investigated. C( 1 )RM, y w f / Y; V21 /+ females displayed considerably more eye pigmenta- tion than C(I )RM, y w f; V21 I + females, with no overlap in expression. However, w l J r x ; V2I flies raised at 25" were not obviously more pigmented than those raised at 18" (small effects would have been obscured by the large amount of variation between individuals characteristic of this stock). Thus, the V2I insertion appears to place the P[wflRJLTR] allele under the influence of a position effect which displays some, but not all, of the features typically seen in classical position effect variegation.
V24 results in a pigmentation level commensurate with a twofold increase in gene expression relative to wa (;.e., V24 w ~ ~ ~ ~ I w ~ ~ ~ ~ and homozygous wn females are pigmented identically; w ~ ' ~ w ~ ~ ~ ~ females are, of course, less pigmented-see Table 3). Thus, V24 shows significantly less expression than do other P[waRSLTR]-containing lines, and differs from V21 in having a constant non-variegating phenotype. This insertion is into the euchromatic region 2C1 and confers hemizygous lethality.
The effect of su(w") and su(f) on these P[W"~""'~ 1 insertions was investigated, but this analysis was ren- dered impossible in the case of V22 by extreme variability in the phenotype. V24 was shown to be unaffected by su(f) by comparison of V24 w l r r X f' su(f) lw"" f'su(f) and V24 wlIrx f' 1 ~ ~ ~ ~ ' f'su((f) females. Since recombinants between SU(W") and V24 were not obtained, the effect of su(wn) on V24 could not be tested. Definitive results on the effects of su(w") and su(f) on P [ w ~ ~ . ' ~ ~ ~ ] were obtained with V? and the mobilized derivatives of V24.
Mobilization of the V24 insertion by a single gen- omic P factor: In a large number of experiments involving P element mediated transformation it has been observed that the majority of insertions exhibit relatively uniform expression, but a minority (on the order of 10-20%) show variant levels of expression [see, for example, HAZELRIGG, LEVIS and RUBIN (1984) and SPRADLING and RUBIN (1983)l. We there- fore considered it likely that the variation in pheno- type among the three original P [ w " ~ . ' ~ ~ ~ ] transfor- mant lines (V?, V22 and V24) was due to a chance occurrence of two cases of strong position effects in these three. In order to investigate whether V24 was indeed a position effect, and provide additional
fTWURbLTR ] insertions for study, we induced the V24 insertion to transpose to new sites, as had been done previously for white transposons showing a position effect (LEVIS, HAZELRIGG and RUBIN 1985b).
For this experiment, we crossed the V24 w1'18 strain with a w' l rX strain carrying a derivative of an insertion of P[h.sP;w+] (LASKI, RIO and RUBIN 1986) on the third chromosome. This element had lost the heat shock promoter and white sequences by deletion and was essentially a single wild-type P element (F. SPEN- CER, unpublished data) but could provide functional P transposase activity, inducing the transposition of
the active P element was separated from these new insertions by additional crosses (see MATERIALS AND
METHODS for experimental details). As a result of these mobilization crosses, 25 inde-
pendent reversions of the V24 position effect were observed. Of these, 24 were phenotypically similar to V3, demonstrating that in most genomic positions
version events were characterized to varying extents; most were mapped to a chromosome and discarded. In all, there were eight transpositions to the second chromosome, five transpositions to the third chro- mosome and 12 cases of X-linked reversion. Of the latter, six were homozygous viable (the lethality as- sociated with the original copy of V24 was not ob- served) and six were not. Of those which were not homozygous viable, three (V24-12, V24-55 and V24- 88) were examined by in situ hybridization (see Table 2), with the result that in each case only a single band of hybridization at 2C1 was visible. While this hy- bridization could result from transpositions to a site adjacent to the original insertion, it is more likely that rearrangements which alter the flanking se- quences responsible for the position effect were the basis for the phenotypic reversion in 1/24-12, V24-55 and V24-B8 (see LEVIS, HAZELRICC and RUBIN 1985b).
In this experiment, transpositions of P[wnRaLTR] from the X chromosome would have been detected regardless of their phenotype (so long as it were not bleach white). The phenotypic similarity of the ma- jority of the derivatives of V24 therefore allows us to conclude that the phenotype of the original 1/24 insertion was due to a nonvariegating euchromatic postion effect, and to take V? (which they resemble) as a standard.
The strains listed in Table 2 were retained for further analysis. One transposition, to cytological position 84F (V24-2OC), conferred yellow eyes with red spots, and appears to be another case of a strong euchromatic position effect. The structural integrity of P[waRSLTR] in this strain has not been confirmed by genomic blotting, but further transpositions re- sulting in the reddish phenotype were observed in unstable sublines of V24-2OC.
P[WURSLTR ] to new sites. Following the mobilization,
P[W'rRrLTR 3 confers reddish pigmentation. These re-

230 S. M. Mount, M. M. Green and G. M. Rubin WaR84e7 appears to be a duplication: A spontaneous
partial revertant of u" (phenotypically like su(w')w") was observed in the su(wa) wa f'su(f) stock. Recom- bination analysis revealed that the alteration was linked to the white locus, and the revertant was named
large region including the white locus was provided by (1) Southern analysis, (2) a study of interactions with su(wn) and su(f), ( 3 ) the observation of effects on recombination in the vicinity of white, (4) a study of interactions with zeste, and (5) the detection of phenotypic revertants (to apricot) in association with recombination (see ROBERTS 1976). Accordingly, the increased pigmentation seen in this derivative can be attributed to increased gene copy number rather than an alteration of the properties of the gene, and
was not informative with regard to the mo- lecular mechanisms by which the phenotype of wn is modified by suppressors or enhancers.
New su(wa) alleles: su( u " ) ~ ~ * , su( Z U " ) ~ ~ ~ and s ~ ( w " ) ~ . ' were first identified as single darker-eyed males in un stocks by J. BIRCHLER (unpublished data), K. RENDAHL (unpublished data) and B. JAFFE (unpub- lished data), respectively. These alleles were passed on to us as putative u" revertants by their discoverers, and were identified as new alleles of su(wf'). The possibility that these apparently new alleles of su(wN) are in fact su( wn)' contaminants cannot be completely excluded, but seems unlikely, particularly in the case of s ~ ( w ' ~ ) ~ J , which arose on a marked chromosome (y un f ) in a laboratory which does not maintain any su(w")' stocks (B. JAFFE, personal communication). The phenotypes of su(w'L)xJz u", S U ( U ~ ' ) ~ ~ ~ w", and su(w")e' ua are indistinguishable from su(wn)' u", demonstrating that the mutation of su( w") + to a state like su(w')' is not a novel event. This is particularly interesting in light of the occurrence of much more extreme alleles of su(w") (ZACHAR et al. 1985).
WaRN4e7 . Evidence that uuR84e7 is a duplication of a
unRX4e7
DISCUSSION
Models of suppression: It is useful to describe three general hypotheses for the molecular basis of suppression (or enhancement) of wu:
1. The majority of RNAs from z d L are polyadeny- lated within copia and probably do not function in pigmentation, but a small amount of RNA with wild- type structure is produced by the removal of copia sequences via RNA splicing (LEVIS, O'HARE and RUBIN 1984; PIRROTTA and BROCKL 1984; ZACHAR et al. 1985; see Figure 1). These observations lead us to favor models in which genetic modifiers act through an effect on RNA processing (either poly- adenylation or splicing). In these models, modifier gene products could play roles in either polyadeny- lation or splicing and mutations in the modifiers
would affect the efficiency of polyadenylation at sites within copia or splicing between sites flanking the insertion. (Note that these two events are mutually exclusive fates of a single primary transcript.)
2. A related, but distinct, hypothesis holds that actual termination of transcription occurs within the copia insertion. In this model modifiers are genes whose products are termination (or antitermination) factors. It should be stressed that since termination of transcription by eukaryotic RNA polymerase I1 is an ill-defined process which typically occurs well downstream of the polyadenylation sites which form the mature 3' ends of messenger RNAs (BIRNSTIEL, BUSSLINGER and STKUU 1985), the presence of poly- adenylated 3' ends within copia does not, by itself, constitute evidence in favor of this model.
3. Copia could provide cis-acting transcriptional control elements which alter the initiation of tran- scription. This could be either a direct effect of transcriptional enhancers or repressors within copia or a product of competition between promoters in copia and white. Modifiers, in this model, are genes whose products act in trans to allow the functioning of cis-acting transcriptional control sequences, and mutations in the modifiers affect either the ability of the white promoter to respond to these sequences or the competition between promoters. In yeast, suppression by spt? of his4 mutations caused by the insertion of Ty elements has been traced to a require- ment for SPT? in T y transcription (WINSTON et al. 1984; WINSTON, DURBAN and FINK 1984). Similar hypotheses advanced to explain suppression in Dro- sophila by su((f) and su(Hw) have found support in that these suppressors affect transcription from gypsy (PARKHURST and CORCES 1986a, b). In the case of u", where any transcriptional alteration of RNA levels would be superimposed upon a defect in RNA struc- ture, copia has been found to alter the temporal pattern of white expression (ZACHAR et al. 1985).
Behavior of waRsLTR derivatives shows that se- quences within the internal portion of copia are nec- essary for both the severity of the wa phenotype and its response to trans-acting modifiers: The u"R".TR allele was originally conceived of as a simplified version of wfL. (Note that zunR.",'l'*' IS ' used here to refer to both the specific allele generated in vitro and studied as P [ U " ~ . ' ~ ~ ~ ] transposons, and alleles, such
and ur, which arose in flies and have the identical structure.) Polyadenylation of white tran- scripts within copia appears to interfere with the synthesis of wild type ut RNA from u", and the
as WuR5YK/
WaR.~LTR allele contains the copia polyadenylation site. ~ l ~ ~ , WnR.\LTR contains a single LTR at precisely the same location as the copia element in w". Thus, any defect which is caused by the disruption of white sequences, by the creation of novel sequences at the junction between white and copia, or by the action o f

Reversion and Suppression of wa 23 1
regulatory sequences contained entirely within the LTR, would be evident in waRJLTR. Finally, Zachar et al. (1985) describe significant and similar alterations in the developmental pattern of white expression in w" and the waRsLTR derivative waLTRI. If any of these factors were the basis of the interaction between w' and trans-acting modifiers of its expression, then one would have expected to have seen a parallel inter- action between these modifiers and waRILTR alleles.
In fact, no effect of su(wa), su(f) or E(w") on the eye pigmentation of any of several independently derived waRJLTR derivatives ( w ~ ~ ~ ~ ~ ~ , wr and several distinct P[waRbLTR] insertions) was observed. We are led to conclude that sequences within the central portion of copia (or some kind of interaction between the two LTRs) are essential, not only for the bulk of the inhibitory effect of copia on white expression, but also for the effect of three trans-acting modifiers of w" expression. It may be relevent that polyadenylation of white-initiated transcripts occurs in the 3' LTR of copia with a much higher frequency than in the 5' LTR (see ZACHAR et al. 1985), possibly because copia sequences upstream of the 3' LTR are necessary for the function of this polyadenylation site.
Secondary insertion appears to be the major route of spontaneous intragenic reversion of this insertion mutation: In addition to the P [ w " ~ . ' ~ ~ ~ ] derivatives (which were constructed in vitro) and the three new su(w") alleles (which are extragenic), we have exam- ined seven derivatives of we. The wnRILTR derivative
was x-ray induced, and the roo insertion wnr is not revertant. That leaves five spontaneous rev-
> f and wr) , of which four are insertion mutations. This result is consistent with previous observations that the major- ity of spontaneous mutations in Drosophila are asso- ciated with the insertion of DNA (see, for examples, CHANC; et al. 1986; LEVIS, O'HARE and RUBIN 1984; ZACHAR and BINCHAM 1982).
Nevertheless, w e were surprised to find that a mutation caused by the insertion of a 5-kb transpos- able element into a 74-nucleotide intron could be alleviated by the insertion of yet more DNA. Fur- thermore, these reversions are not consistently asso- ciated with the insertion of additional DNA into a single region of copia, and they appear (based on their interactions with trans-acting modifiers and the structure of their transcripts) to affect wa expression in different ways. These are not the results which would have been expected if there were a particular region of copia, such as a polyadenylation site, whose disruption by the second insertion was explanation for the revertant effect.
Partial reversion of mutations by a second insertion within an initial insertion is not unprecedented. For example, the allele w' is caused by the insertion of a transposable element near the w + transcriptional
WnR59kI
ertants (WuRM, WnKX4~I Y ( I 3 ) WnR7Y127 WnRX4h
start site (O'HARE, LEVIS and RUBIN 1984; O'HARE et al. 1984; PIRROTTA and BROCKL 1984). Revertants of w' include we and wh, which are associated with insertions of a 190-nucleotide portion of the element pogo and the 8.7-kb B104/roo element, respectively (O'HARE, cited in FINNECAN and FAWCETT 1986), and dR2, which is due to an insertion of an intact 5.4-kb 1 factor (FAWCETT et al. 1986). A similar phenomenon was observed by MIZROKHI et ai. (1985), who found the 3.1-kb element Jockey within the mdg4/gypsy insertion of cPR2pNI',, an allele of ct with a less extreme phenotype than the grandparental allele cPR2, which carries only gypsy at precisely the same location.
The w~~~~ insertion is a truncated Z element: The I factor is a transposable element of approximately 5.4-kb which is responsible for the ZIR system of hybrid dysgenesis (SANG et al. 1984; for a review see BREGLIANO and KIDWELL 1983). Active I factors are found in I strains, but not R strains, and appear to cause partial or complete sterility, chromosomal rear- rangements and mutations following the mating of I strain males and R strain females. Recently, the sites of insertion of several I factors, and one complete I factor, have been sequenced (FAWCETT et al. 1986). The I factor appears similar in sequence organization and coding potential to mammalian LINEs or L1 sequences, long interspersed repeated sequences found in the mammalian genome and believed to be nonviral retrotransposons (reviewed by WEINEK, DEININCER and EFSTRATIADIS 1986).
Despite its extremely short length, the w~~~~~ in- sertion appears to be a fairly typical Z element inser- tion. The seven target site duplications sequenced by FAWCETT et al. (1986) were all 10-14 bp in length and AIT rich. The 13-bp target site duplication of
The sequence of the 83 nucleotide wnRXJh insertion is identical to the last 83 nucleotides of the complete I factor (except for the variable number of TAA repeats), and can be thought of as an I factor which is severely truncated at its 5' end. The insertion of 5' truncated incomplete I elements has been de- scribed recently as the cause of two mutations at the yellow locus, f R 3 and yIR4 (FINNEGAN et al. 1987); this observation is consistent with the similarities between I factors and LINEs, which are often observed to be truncated at their 5' ends (WEINER, DEININGER and EFSTRATIADIS 1986).
extensive backcrossing of an unrelated P element insertion mutation into an M strain (J. BIRCHLER, personal communication). It is interesting to specu- late that this backcross was in fact an I/R dysgenic cross, and was responsible for mobilizing Z elements, leading to the w ~ ' ~ ~ ~ ~ reversion.
Altered response of w~~~~ to su(w'), but not sum
W11R84h , ATATTTCCTTTCT, is therefore typical.
WuRX4h arose during, or immediately following,

232 S. M. Mount, M. M. Green and G. M. Rubin
or E(w"), implies different sites of action for these trans-acting modifiers: We think it likely that the trans-acting modifiers of w" act to alter the efficiency of rate-limiting steps in the elaboration of white mRNA from the white-apricot allele. These can either be steps (such as initiation of transcription or the removal of copia sequences by splicing) which lead to the production of functional wild type RNA, or steps (such as RNA degradation, polyadenylation within copia, or termination of transcription) which lead to the production of nonfunctional products. A primary goal of this research is the identification of precise steps in the expression of w' at which trans- acting modifiers of this gene act.
It seems possible, a priori, that all known modifiers of wU expression might act at the same step. In this context, it is particularly interesting that wu can mutate to a state (represented by the w~~~~~ allele) which is responsive to su(f) but unresponsive to su(wf'). Does it follow from this observation that su(wu) and su(f) act independently? Let us suppose that su(w") and su(f) do act at the same step in w" expression. For example, su(f) could act by increasing the con- centration of the su(w') gene product, in which case their antagonistic effects would be due to different effects on the rate of some process dependent on the su(w') gene product. T o reconcile this model with the altered response of w ~ ~ ~ ~ ' ~ to su(wu) but not su(f), one could suppose that w~~~~~ had an increased threshold (above wild type levels) for the SU(W") gene product, and decreasing it further (the su(wU) muta- tion) has no effect, but increasing it (the su(f) muta- tion) does have an effect. This hypothesis is experi- mentally testable. The su(wa) wn su(f) genotype confers a level of pigmentation very similar to that of wU alone (LEVIS, O'HARE and RUBIN 1984). Thus, if this model is true, the su(w") su(f) combination leads to a level of su(w') gene product approximately equal to that in wild type flies and su(wn) w"RR4h su(f) should resemble waRX4 alone. However, su(w") wuRX4h su(f(f) flies are pigmented exactly as are w~~~~~~ su(f) flies (data not shown). su(wU) has no effect on w " ~ ~ ~ ~ , even in the presence of the su((f). From this we conclude that su( wa) and su(f) do indeed have differ- ent sites of action, and that the insertion of a small I element in the w~~~~~ revertant has specifically altered the response of W" to su(w").
This logic can be extended to E(w"), which affects transposable element-induced mutations unaffected by su(wu) and su(f) (e.g., wa4 and we; GREEN 1959), but similarly fails to interact with the majority of white alleles, including waRsLTR alleles. It appears that the insertion of copia has created an allele, wa, whose phenotype is subject to modulation by mutations in at least three unlinked genes acting through three separable mechanisms.
We thank J I M BIRCHLER, BETSY JAFFE, BURKE JUDD, ROBERI
LEVIS, FORREST SPENCER, DANIELLE THIERRY-MIEG, K A n E KEN- DAHL, and both the Mid-America and Cal Tech Drosophila Stock Centers for providing Drosophila stocks. We thank J I M BIRCHLER, DAVID FINNEGAN, BURKE JUDD, FORREST SPENCER and ALLAIN BUCHETON for communication of results prior to publication. We especially thank TODD LAVERTY for performing the in situ hybrid- izations, JOE YEUNG for cytology, ROBERT LEVIS for sharing his encyclopedic knowledge of white locus molecular biology, and FORREST SPENCER for her significant contributions to the mobili- zation of P[WnRsLTR]. TULLE HAZELRIM;, ROBERT LEVIS, MAKK DWORKIN and PA.I.RIC;K Lo are appreciated for their careful reading of the manuscript. S.M. was supported by a postdoctoral fellowship from the National Institutes of Health. This work was supported by grants from the National Institutes of Health and American Cancer Society to G.M.R.
LITERATURE CITED
BANKIER, A. T., and B. G. BARREL, 1983 Shotgun DNA sequenc- ing. In: Techniques in Nucleic Acid Biochemistry, Val B5, pp 1- 33. Edited by R. A. FLAVELL. Elsevier Scientific Publishers, Ireland, Ltd.
BINGHAM, P. M., and B. H. JUDD, 1981 A copy of. the copia transposable element is very tightly linked to the w" allele at the white locus of D. melanogester. Cell 25: 705-7 1 1.
BINGHAM, P. M., R. LEVIS and G. M. RUBIN, 1981 Cloning of DNA sequences from the white locus of D. melanogaster by a novel and general method. Cell 25: 693-704.
BIRCHLER, J., 1986 Dominant genetic enhancers interact with transposable element insertion mutants at the white locus in Drosophila. Genetics 113: s47.
BIRNSTIEL, M., M. BUSSLINGER and K. S r ~ u u , 1985 Transcription termination and 3' end processing: the end is in site! Cell 41: 349-359.
BREGLIANO, J.-C., and M. G. KIDWELI., 1983 Hybrid dysgenesis determinants. In: Mobile Genetic Elements. pp. 363-4 10, Edited by J. A. SHAPIRO. Academic Press, Orlando, Fla.
CAMPUZANO, S., L. BALCELLS, R. VILLARES, L. CAKRAMOLINO, L. GARCIA-ALONSO and J . MODOLELL, 1986 Excess function Hairy-wing mutations caused by gypsy and copia insertions within structural genes of the achaete-scute locus of Droso- phila. Cell 44: 303-312.
CARBONARE, B. D., and W. J. GEHRING, I985 Excision of copia element in a revertant ofthe white-apncot mutation of Drosophila rnelanogmter leaves behind one long terminal repeat. Mol. Gen. Genet. 199: 1-6.
CHANC;, D.-Y., B. WISELY, S.-M. HUANG and R. A. VOELKEK, 1986 Molecular cloning of suppressor-of-sable, a Drosophila rnelanogmter transposon-mediated suppressor. Mol. Cell. Biol.
DAVIS, P. S., M. W. SHEN and B. H. JUDD, 1987 Asymmetrical pairings of transposons in and proximal to the white locus of Drosophila account for four classes of regularly occurring exchange products. Proc. Natl. Acad. Sci. USA 84: 174-178.
DUDICK, M. E., T. R. F. WKIGHT and L. L. BKOI H E M , 1974 The developmental genetics of the temperature-sensitive lethal allele of the suppressor-of~orked, 1 ( l ) s ~ ( j ) " " ~ , in Dmwphih mc- lanogaster. Genetics 76: 487-5 10.
EMORI, Y., T. SHIBA, S. KANAYA, S. INOUYE, S. YUKI and K. SAIW, 1985 The nucleotide sequences of copia and copia-related RNA in Drosophila virus-like particles. Nature 315: 773-776.
FAWCEIT, D. H., C. K. LISTEK, E. KELLETT' and D. J. FINNEGAN, 1986 Transposable elements controlling I-R hybrid dysge- nesis in D. rnelanogester are similar to mammalian LINES. Cell
FINNEGAN, D. J., I. BUSSEAU, M. LYNCH, D. H. FAWCETT, C. K. LISTER, A. PELISSON, H. M. SANG and A. BUCHETON, 1987 The structure of mutations induced by I-R hybrid dysgenesis in Drosophila melanogaster. In: Tmmposable ElemPnt.5
6: 1520- 1528.
47: 1007-1015.

Reversion and Suppression of wa 233
as Mutagenic Agents. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
FINNEGAN, D. J., and D. H. FAWCETT, 1986 Transposable ele- ments in Drosophila melanogaster. In: Oxford Surveys on Eukaryotic Genes, Vol. 3. Oxford, University Press, New York.
FLAVELL, A. J., R. LEVIS, M. A. SIMON and G. M. RUBIN, 1981 The 5’ termini of RNAs encoded by the transposable element copia. Nucleic Acids Res. 9: 6279-6291.
GEHRING, W. J., and R. PARO, 1980 Isolation of a hybrid plasmid with homologous sequences to a transposing element of Dro- sophila. Cell 19: 897-904.
GOLDBERG, M. L., R. PAKO and W. J. GEHRING, 1982 Molecular cloning of the white locus region of Drosophila melanogaster using a large transposable element. EMBO J. 1: 93-98.
GREEN, M. M., 1959 Spatial and functional properties of psue- doalleles at the white locus in Drosophila melanogaster. Heredity
GREEN, M. M., 1982 Genetic instability in Drosophila melanogaster: deletion induction by insertion sequences. Proc. Natl. Acad. Sci. USA 79: 5367-5369.
HAZELRIGG, T., R. LEVIS and G. M. RUBIN, 1984 Transformation of white locus DNA in Drosophila: dosage compensation, ze,ste interaction and position effects. Cell 36: 469-48 1.
KARESS, R. E., and G. M. RUBIN, 1982 A small tandem duplication is responsible for the unstable white-ivory mutation in Droso- phila. Cell 30: 63-69.
KARESS, R. E., and G. M. RUBIN, 1984 Analysis of P transposable element functions in Drosophila. Cell 38: 135-146.
KUBLI, E., 1986 Molecular mechanisms of suppression in Dro- sophila. Trends Genet. 2: 204-209.
LASKI, F. A., D. C. R l o and G. M. RUBIN, 1986 Tissue specificity of Drosophla P element transposition is regulated at the level of mRNA splicing. Cell 44: 7-19.
LEVIS, R., P. M. BINGHAM and G. M. RUBIN, 1982 Physical map of the white locus of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 79: 564-568.
LEVIS, R., T. HAZELRIGG and G. M. RUBIN, 1985a Separable cis- acting control elements for expression of the white gene of Drosophila. EMBO J . 4: 3489-3499.
LEVIS, R., T. HAZELRIG(; and G. M. RUBIN, 1985b Effects of genomic position on the expression of the white gene of‘ Drosophila. Science 229: 558-56 1.
LEVIS, R., K. OHARE and G. M. RUBIN, 1984 Effects of trans- posable element insertions on RNA encoded by the white gene of Drosophila. Cell 38: 47 1-481.
LINDSLEY, D. L., and E. H. GRELL, 1968 Genetic variations of Drosophila melanogaster. Carnegie Inst. Wash. Publ. 627.
LOENEN, V., and F. BLATTNER, 1983 Lambda Charon vectors (Ch32, 33, 34 and 35) adopted for DNA cloning in recombi- nation-deficient hosts. Gene 26: 17 1- 179.
MCLACHLAN, A,, 1986 Drosophila forked locus. Mol. Cell. Biol. 6:
MERRIAM, J. R., 1968 Drosophila Information Service 43: 64. MIZROKHI, L. J., L. A. OBOLENKOVA, A. F. PRIIMAGI, Y. V. ILYIN,
T. I. GERASIMOVA and G. P. GEORGIEV, 1985 The nature of unstable insertion mutations and reversions in the locus cut of Drosophila melanogaster: Molecular mechanism of transposition memory. EMBO J. 4: 3781-3787.
MODOLELL, J., W. BENDER and M. MESELSON, 1983 Drosophila melanogaster mutations suppressible by the suppressor of Hairy- wing are insertions of a 7.3 kilobase mobile element. Proc. Natl. Acad. Sci. USA 80: 1678-1682.
MOUNT, S. M., and G. M. RUBIN, 1985 Complete nucleotide sequence of the Drosophila transposable element copia: ho- mology between copia and retroviral proteins. Mol. Cell. Biol. 5: 1630-1638.
MULLER, H. J., 1944 Reddish-a new near-normal allele of while. Drosophila Inform. Serv. 18: 57.
NOVITSKI, E., 1963 The construction of new chromosomal types
13: 303-3 15.
1-6.
in Drosophila melanogaster. pp. 381-403. In: Methodology in Basic Genetics, Edited by W. J. BURDETTE. Holden Day, San Francisco.
O’HARE, K., R. LEVIS and G. M. RUBIN, 1983 Transcription of the white locus in Drosophila melanogaster. Proc. Nat. Acad. Sci. USA 80: 69 17-692 1.
O’HARE, K., C. MURPHY, R. LEVIS and G. M. RUBIN, 1984 DNA sequence of the white locus of Drosophila melanogaster. J . Mol. Biol. 180: 437-455.
PARKHURST, S. M., and V. G. CORCES, 1985 forked, gypsys and suppressors in Drosophila. Cell 41: 429-437.
PARKHURST, S. M., and V. G. CORCES, 1986a Interactions among the gypsy transposable element and the yellow and the Sup- pressor of Hairy-wing loci in Drosophila melanogaster. Mol. Cell. Biol. 6: 47-53.
PARKHURST, S. M., and V. G. CORCES, 1986b Mutations at the suppressor of forked locus increase the accumulation of gypsy- encoded transcripts in Drosophila melanogaster. Mol. Cell. Biol.
PIRROTTA, V., and C. BROCKL, 1984 Transcription of the Dro- sophila white locus and some of its mutants. EMBO J. 3: 563- 568.
RASMUSON, B., M. M. GREEN and G. EWERTSON, 1960 Qualitative and quantitiative analysis of eye pigments and pteridines in back-mutations of the mutant wn in Drosophila melanogaster. Hereditas 46: 635-650.
ROIIERTS, P. A., 1976 The genetics of chromosome aberration. pp. 132-146. In: The Genetics and Biology ofDrosophila, Vol. 1A. Edited by M. ASHBUKNER and E. NOVITSKI. Academic Press, New York.
ROEDER, G. S., and G. R. FINK, 1983 Transposable elements in yeast. pp. 299-328. In: Mobile Genetic Elements, Edited by J. A. SHAPIRO. Academic Press, Orlando, Fla.
RUIIIN, G. M., I983 Dispersed repetitive DNAs in Drosophila. pp. 329-361. In: Mobile Genetic Elements, Edited by J . A. SHAPIKO. Academic Press, Orlando, Fla.
RUBIN, G . M., and A. C. SPRADLING, 1982 Genetic transformation of Drosophila with transposable element vectors. Nucleic Acids Res. 11: 634 1-635 1 .
RUBIN, G. M., and A. C. SPRADLING, 1983 Vectors for P-element mediated gene transfer in Drosophila. Nucleic Acids Res. 11: 634 1-635 1.
RUSSELL, M. A,, 1974 Pattern formation in the imaginal discs of a temperature-sensitive cell-lethal mutant of Drosophila melan- ogmter. Dev. Biol. 40: 24-39.
SANG, H. M., A. PELISSON, A. BUCHETON and D. FINNEGAN, 1984 Molecular lesions associated with white gene mutations induced by I-R hybrid dysgenesis in Drosophila melanogaster.
SEARLES, L., and R. A. VOELKER, 1986 Molecular characterization of the Drosophila vermilion locus and its suppressible alleles. Proc. Natl. Acad. Sci. USA 83: 404-408.
SPOFFORD, J. B., 1976 Position-effect variegation in Drosophila. pp. 955-1018. In: The Genetics and Biology of Drosophila, Vol. IC, Edited by M. ASHBURNER and E. NOVITSKI. Academic Press, London.
SPRADLING, A. C., and G. M. RUBIN, 1982 Transposition of cloned P elements into Drosophila germ line chromosomes. Science
SPRADLING, A. C., and G. M. RUBIN, 1983 The effect of chro- mosomal position on the expression of the Drosophila xanthine dehydrogenase gene. Cell 34: 47-57.
VARMUS, H. E., 1983 Retroviruses. pp. 41 1-503. In: MobileGenetic Elements, Edited by J. A. SHAPIRO. Academic Press, Orlando, Fla.
6: 227 1-2274.
EMBO J. 3: 3079-3085.
218: 342-347.
VON HALLE, E. S., 1969 Drosophila Inform. Serv. 44: 119. WALKER, A. R., A. J. HOWELLS and R. G. TEARLE, 1986 Cloning
and characterization of the vermilion gene of Drosophila melan- ogaster. Mol. Gen. Genet. 202: 102-107.
WEINER, A. M., P. L. DEININCER and A. EFSTRATIADIS,

234 S. M. Mount, M. M. Green and G. M. Rubin
1986 Nonviral retrotransposons: genes, pseudogenes and transposable element generated by the reverse flow of genetic information. Annu. Rev. Biochem. 55: 631-661.
WINSTON, F., K. J. DURBAN and G. R. FINK, 1984 The SPT3 gene is required for normal transcription of Ty elements in S. cereviriue. Mol. Cell. Biol. 39: 675-682.
WINSTON, F., D. T. CHALEFF, B. VALERT and G. R. FINK, 1984 Mutations affecting Ty-mediated expression of the HIS4 gene of Saccharomyces cerevisiae. Genetics 107: 179-197.
ZACHAR, Z., and P. M. BINGHAM, 1982 Regulation of white locus expression: the structure of mutant alleles at the white locus of Drosophila melanogaster. Cell 30: 529-54 1.
ZACHAR, Z., D. DAVISON, D. GARZA and P. M. BINGHAM, 1985 A detailed developmental and structural study of the transcrip- tional effects of insertion of the copia transposon into the white locus of Drosophila melanogaster. Genetics 111: 495-515.
Communicating editor: W. M. GELBART





























