Embed Size (px)
Citation preview

Passive immunization of the newborndairy calf via fermented colostrum
Item Type text; Thesis-Reproduction (electronic)
Authors Snyder, Antes Catherine, 1950-
Publisher The University of Arizona.
Rights Copyright © is held by the author. Digital access to this materialis made possible by the University Libraries, University of Arizona.Further transmission, reproduction or presentation (such aspublic display or performance) of protected items is prohibitedexcept with permission of the author.
Download date 17/06/2018 18:13:45
Link to Item http://hdl.handle.net/10150/554869

PASSIVE IMMUNIZATION OF THE NEWBORN DAIRY CALF VIA FERMENTED COLOSTRUM
byAntes Catherine Snyder
A Thesis Submitted to the Faculty of.theCOMMITTEE ON ANIMAL PHYSIOLOGY (GRADUATE)In Partial Fulfillment of the Requirements
For the Degree ofMASTER OF SCIENCE
In the Graduate College
THE UNIVERSITY OF ARIZONA
19 7 5

STATEMENT OF AUTHOR
This thesis has been submitted in partial fulfillment of requirements for an advanced degree at The University of Arizona and is deposited in the University Library to be made available to borrowers under rules of the Library.
Brief quotations from this thesis are allowable without special permission, provided that accurate acknowledgment of source is made. Requests for permission for extended quotation from or reproduction of this manuscript in whole or in part may be granted by the head of the , major department or the Dean of the Graduate College when in his judgment the proposed use of the material is in the interests of scholarship. In all other instances, however, permission must be obtained from the author.
SIGNED:
APPROVAL BY THESIS DIRECTOR
This thesis has been approved on the date shown below:
f j James D. Schuh Professor of Dairy and Food.Sciences

To my Parents for opening my life
To Dr. Schuh for opening my eyes
Dr. Wegner and Bob Schoenberger for opening the door
and to you for opening my thesis

TABLE OF CONTENTS
PageLIST OF ILLUSTRATIONS , . . . . . . . . ......... vLIST OF TABLES................... viABSTRACT . ............ viiINTRODUCTION , . . . . . . . . . . . . . .......... 1REVIEW OF LITERATURE . . . . . . . . . ....... . . . . . . 3
Introduction . . . . . . . . . . . . . . . . . ........ 3Transmission of Antibodies . . . . . . . . . . . . . . . 3Major Components of Colostrum 5The. .Immune Globulins of Colostrum ....... 6Changes in the Digestive Tract Associated with
Transmission . V . .'. ... . . . . 8Transmission of Globulin from Intestine to the'
Circulation . ............ 10The Decline of Passive Immunity and Development of
Active Immunity............ 11Immune1 Globulin Spectrums and Titers of Colostrum from
Individual Cows . . . . . . . . . 13Fermented Colostrum ........ 15
PROCEDURE........ 18RESULTS AND DISCUSSION . . , . . . . . . . . , .. . ........ 21
APPENDIX 1: DATA ON THE FIVE TREATMENTS- . . . . . . . . . . 35LITERATURE CITED .......... 42
iv

LIST OF ILLUSTRATIONS
Figure . Page1. Electrophoretic serum protein patterns of three calves
show precolostral absence of a globulin and itsvariable appearance in post colostra! serum at 48 hr. y. . 24
2. Mean serum a globulin levels of the different colostrumtreatments contrasted ........ 25
3. Mean serum beta levels of the different colostrum treatmentscontrasted . •. . 26
4. Mean serum alpha levels of the different colostrumtreatments contrasted .............. 27
5. Mean serum albumin levels of the different colostrumtreatments contrasted . . . . . . . .................. 28
6. Schematic guide of precipitin bands for the followingimmunoelectrophoretograms (Figures 7, 8, 9, 10, and 11) . 29
7. Immunoelectrophoretogram of pre- and post-colostra! calfserum 30 '
8. Immunoelectrophoretogram of post colostral serum from calvesfed fresh colostrum . . . . . . . . ......... . . 31,
9. Immunoelectrophoretogram of post colostral serum from a calffed fresh colostrum .......... 32
10. Immunoelectrophoretogram . . . . . . . . . . . . . . . . . . 3311. Immunoelectrophoretogram of calf 437, 48 hr demonstrating
a globulinopathy. . . . . . . . . . . . . . . . . . . . . 34
v

LIST OF TABLES
Table Page1. The Progressive Change of Colostrum into Milk . . . . . . . . 72. Treatment with Fresh Colostrum . . . . . . . . . . . . . . . . 363. Treatment with Frozen Colostrum...............' .......... 374. Treatment with Fermented Colostrum ......... 385. Treatment with Pooled Frozen Colostrum ........ . . . . 396. Treatment with Pooled Fermented Colostrum . . . . . . . . . . 40
7. Average of the Five Treatments . . ........... 41
vi

ABSTRACT
Twenty-five Holstein calves were equally assigned to one of , five colostrum feeding regimens: (1) fresh, (2) frozen, (3) fermented,(4) pooled frozen, and (5) pooled fermented. Each calf received 500 ml of colostrum (first milk) at birth and again at 12 hr. Treatments 1,2, and 3 represented individual portions (1000 ml) of colostrum from each of five cows, while 4 and 5 represented mixed colostrum of equal portions from all five cows. Blood samples were taken and analyzed at birth, 12, 24 and 48 hr for serum immunoglobulins. Mean calf serum IgG concentrations (g/100 ml) in order 1 to 5 were: 24 hr .67, .40, .24, .65, .17; 48 hr .80, .45, .20, .59, .26. Serum IgG values were consistently lower for calves fed fermented colostrum (3, 5). Only one calf (5) failed to absorb the immunoglobulin. Immunoelectrophoretic data will be presented.

CHAPTER 1
INTRODUCTION
Dairy calves are bom agammaglobulinaemic and acquire early postnatal immunity to pathogens through the ingestion and absorption of immunoglobulins (Ig) or antibodies in the colostrum milk during the first 24 hours of life. Whether or not an adequate level of immunoglobulin in the blood of a calf is obtained depends on three factors. First, the mother must yield an adequate amount of colostrum with a sufficiently high concentration and spectrum of immunoglobulin. Second, the calf must have the ability to absorb the immunoglobulin efficiently when an adequate amount is. ingested. Third, calf management practices must be such that the transfer is not blocked or unduly reduced.
It is known that large variations occur in the concentration and spectrum of colostral antibodies among individual cows (Smith 1930, Ingram, Lovell and Wood 1956, Kruse 1958, Johnson and Pierce 1959, Sarwar, Campbell and Petersen 1964, Gay 1965, Smith 1965, Lambert and Femelius 1968, Royal, Robinson and Duganzich 1968, Ingram, Corbiel and
Mai corns m 1970, Ingram and Malcomson 1970, Rankin "and Taylor 1970/Bush j Aguilera and Adams 1971, Watts 1971). Accordingly, the maternal passive immunity acquired by calves may often be too limited in scope to afford any significant degree of protection to the myriad of organisms in their, environment. Low levels of immunoglobulins in calf serum have been
1

2correlated with a higher incidence of disease and death (Fey and Mar- gandent 1962, Dam 1968, Ferris and Thomas 1974),
In addition to its basic protective value colostrum is often stated to have a prophylactic or therapeutic effect when fed to older calves with digestive upsets. A new method of storing colostrum by natural fermentation has been reported to improve nutrition and reduce the incidence and severity of digestive disturbances when fed to young calves.
In identifying the role of colostrum in the epidemiology of disease and acquired passive immunity in new bom calves little attention has been given to the restricted scope of immunoglobulins that may occur in maternal colostrum. Information is limited on the prophylactic value of fresh, frozen and fermented colostrum when fed to young calves. This experiment was designed to test the efficiency of colostral antibody transfer when colostrum milk was fed to new born calves in the fresh, frozen, fermented, pooled frozen and pooled fermented states.

CHAPTER 2
REVIEW OF LITERATURE
IntroductionA considerable amount of information is available concerning
the transmission of immunity in ruminants, particularly the cow, sheep and goat. Basically the processes of antibody transmission .and related factors for these species are similar. Therefore, to avoid redundancy attention in this literature review will concern only the bovine with the understanding that supportative information is available for the sheep and goat. For a more comprehensive review and discussion of this subject including the rabbit, rat, mouse, cat, dog, monkey, man, ruminant, and birds , the reader is referred to a recently published book by Brambell (1970).
. . Transmission of Antibodies Little and Orcutte (1922) observed a rapid absorption of Bacillis
abortus agglutinins into the blood stream of newborn calves within a few hours after the calves had ingested their mother's first milk "Colostrum." Accordingly Smith and Little (1922a) found that newborn calves fed colostrum usually survived with a minimum of difficulties whereas a majority of calves fed raw milk in place of colostrum died in the first week of
life from intestinal infections.
3

At about the same time, Howe (1921) and Orcutt and Howe (1922). observed that agglutinins appearing in calf serum following consumption of colostrum were associated with a globulin precipitable at low concentrations of sodium sulfate. Subsequently, several investigators (Jameson, Alvarez-Tostado and Sortor 1942, San Clemente and Huddleson 1943,Hansen and. Philips 1947) found by electrophoretic analysis that immunoglobulin is generally not present in calf serum at birth but a slow moving globulin appears in the serum after ingestion of colostrum.
Smith (1948) and Smith and Holm (1948) demonstrated that the electrophoretic component appearing in the serum of calves fed colostrum was identical in mobility with immune lactoglobulin in colostrum. Deutsch and Smith (1957) confirmed that lactoglobulin is the component responsible for transferring colostral immunity to newborn calves.
Analysis by electrophoresis and ultracentrifuge of postcolostral calf serum by Johnson and Pierce (1959) showed a rise in total serum protein containing 51-56% immune lactoglobulin. No similar component appeared in precolostral sera. There was no detectable change in the structure of the immune lactoglobulin protein during its absorption and appearance in the calf blood (Jameson et al. 1942, Smith and Holm 1948, Johnson and Pierce 1959).
Other researchers have observed that cows positive to agglutination and complement fixation tests for Bacillis abortus (McAlpine and
Rettger 1925, McDiarmid 1946), Trichomonas foetus (Kerr and Robertson 1946), Rinderpest (Brown 1958), and Foot and Mouth disease (Graves 1963)

5did not transmit agglutinins for these maladies to their calf in uterine but. only after birth through colostrum.
Ratner, Jackson and Gruehl (1927) attribute the lack of anti
bodies in calf serum at birth to the large number of cell layers between the maternal and fetal circulation in the bovine during gestation. Brambell , Hemmings and Henderson (1951) have reported that ruminants with five cell layers and other animals (horse, pig) with six layers between the maternal and fetal blood supply do not transfer immunoglobulins to the fetus. In contrast carnivores (dogs, cats, etc.), man and rodents (rat, guinea pig, etc.) with four, three and one cell layers,
respectively, readily transfer immunoglobulins via the placenta.To determine if the lack of placental transmission in the bovine
was related to differences in species protein Kulangara and Schechtman (1963) injected human serum albumin into three cows 18 to 96 hours before parturition. Although the human albumin was readily detected in the cows' serum none could be found in the calves' serum even when some albumin was injected into the uterine lumen of another cow 70 hours be
fore calving.Since all available evidence indicates that little or no pla
cental transfer of antibodies occurs in the bovine species, the newborn calf must receive these vital components soon after birth via colostrum
to enhance its survival and well being.
Major Components of ColostrumColostrum is defined as the first lacteal secretion after par
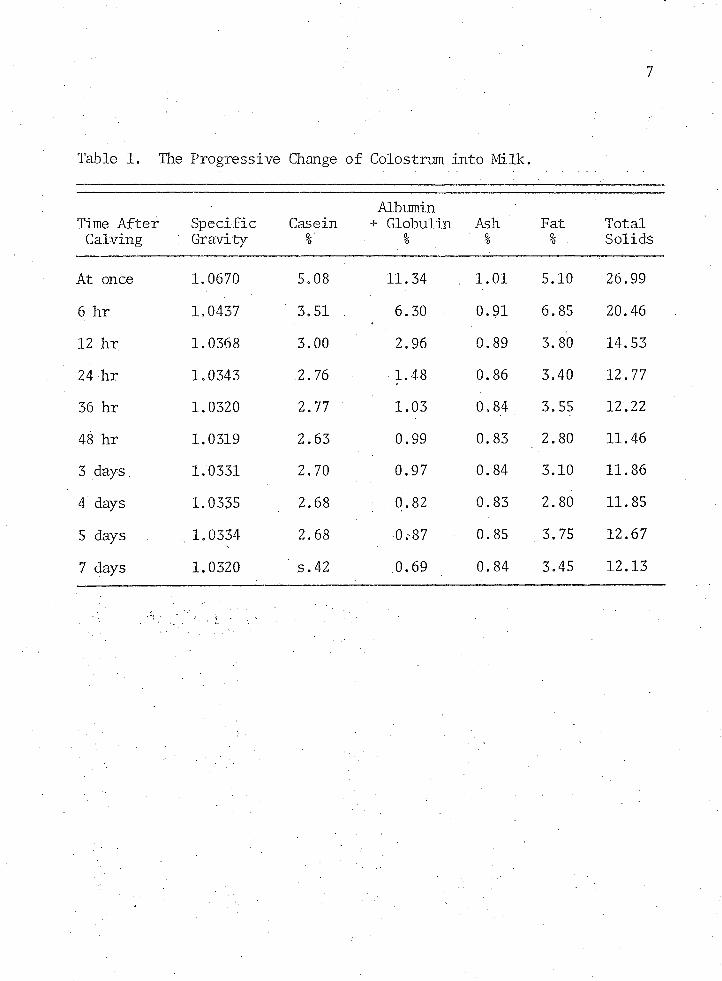
turition i Average composition of colostrum and its progressive change

change to normal milk is shown in Table 1 on the following page (Henderson 1971).
The greater amount of total solids in colostrum when compared to milk is primarily reflected in the additional protein content, especially the albumin-globulin fraction. The immune globulin may represent as much as 50 to 60% of the total colostral protein or 90% of the albumin-globulin fraction (Smith 1948) .
Although colostrum is often referred to as milk produced during the first 3 to 5 days after calving (Crowley 1973) only milk drawn within the first 6 hours contains the high level of antibodies needed to protect the young calf (Smith and Little 1922a,Smith 1959, Graves 1963, Bush et al. 1971). Graves (1963) observed a significant difference in antibody titer to Hoof and Mouth disease in colostrum from different quarters and a marked drop of antibody in all quarters following first milking. More recently Bush et al. (1971) have reported a decline in colostral immunoglobulins from an initial level of 6% to 2.4% in 24 hours. In addition, colostrum has about 10 times more vitamin A which also does not diffuse across the placental barrier (Stewart and McCallum 1942, Hansen, Phillips and Smith 1946, Watts 1971).
The Immune Globulins of ColostrumWork with radioactive materials has shown immune globulins in
colostrum and milk are not synthesized in the udder but are derived directly from the blood serum (Askonas et al. 1954, Larson and Gillespie 1957). The mammary gland selectively secretes serum globulins during all phases of lactation but the rate of infiltration is especially high
7
Table 1. The Progressive Change of Colostrum into Milk.
Time After Calving
Specific ; Gravity
Casein%
Albumin + Globulin
%Ash%
Fat%
TotalSolids
At once 1.0670 5.08 11.34 . 1.01 5.10 26.99
6 hr 1.0437 3.51 . 6.30 0.91 6.85 20.46
12 hr 1.0368 3.00 2.96 0.89 3.80 14.5324 hr 1.0343 2.76 1.48 0.86 3.40 12.7736 hr 1.0320 2.77 1.03 0.84 . 3.55 12.22
48 hr 1.0319 2; 63 0.99 0.83 2.80 11.46
3 days 1.0331 2.70 0.97 0.84 3.10 11.86
4 days 1.0335 2.68 0.82 0.83 2.80 11.85
5 days . 1.0334 2.68 ,0.-87 0.85 . 3.75 12.67
7 days 1.0320 s.42 0.69 0.84 3.45 12.13

near parturition when colostral a-globulin levels may exceed the serum by five times (Pierce and Feinstein 1965, Klaus, Bennett and Jones 1969).
Smith (1948) and Smith and Holm (1948) reported that the electrophoretic properties of colostral lactoglobulin corresponded closely with the a-̂ globulins from the maternal blood serum and were identical to the component observed in calf serum following the ingestion of colostrum.In contrast little similarity was shown between the immune globulines of colostrum and the (^globulins isolated from the maternal blood serum.
Larson and Kendall (1957) and Larson (1958) demonstrated that the level of and aglobulins are the same as the two immune globulins that appear in colostrum and subsequently in the newborn calf after ingestion. Why the B^ and a-̂ globulins pass selectively into the udder to a greater degree at parturition than their relative serum concentration is not known.
Changes in the Digestive Tract Associated with Transmission
Comline, Roberts and Titchen (1951) found absorption of colostral globulin takes place entirely in the small intestine. Lecce (1966) found, during the period, of intestinal permeability, whole proteins are absorbed by a process of pinocytosis through the intestinal epithelium.
Numerous_investigators have shown that permeability of the intestinal wall of the newborn calf to colostrum globulins is transitory (Hansen and Phillips 1947, Anonymous 1949, Smith and Erwin 1959, Balfour and Comline 1962, Gay 1965, Pierce and Feinstein 1965, McCoy, Hunter and
Williams 1968, Bush et al. 1971).

Howe (1921) , Hansen and Phillips (1947) observed that colostra! globulin is absorbed into the serum of the newborn calf only during the first 24 hours of life. Other workers (Comline et al. 1951, Bush et al. 1971) report intestinal permeability to immune globulins may persist to 48 hours after which "closure" is presumed. On the other hand Gay (1965) has indicated this special function may be lost in some, calves within 6 to 8 hours after birth. Wide variations, including a lack of serum aglobulins have been observed in calves known to consume colostrum within a few hours after birth (Gay 1965, Smith, O'Neil and Simmons 1967, Klaus, Bennett and Jones 1969, Ingram and Mai corns on 1970). Klaus et al. (1969) state that Fey and Margadant (1962) were unable to find aglobulin in 10% of calves fed colostrum soon after birth. Klaus et al. (1969) also reported marked hypogammaglobulinaemia in calves allowed to nurse colostrum from their dams but they could find no cor
relation between colostral antibody titers and Calf serum immunoglobulin levels.
Recently Bush et al. (1971) determined that approximately 68% of the variation in calf serum immunoglobulin at 24 hours could be attributed to differences in colostral antibody concentration when calves were fed colostrum according to body weight. However, the concentration of colostral globulin consumed could not be related to hypogamma- globulinaemia in some calves. Maximum serum immunoglobulin levels were not closely related to incidence or severity of diarrhea during the seven month study. " In contrast, other workers (Fey and Margadent 1962, Dam 1968, Ferris and Thomas 1974) have noted that calves with low blood

10serum immmoglobulin succumb to infection more readily than calves with higher blood values.
The factors involved in cessation of intestinal permeability - are unknown, but digestive degradation of the proteins does not appear to play a part (Deutsch and Smith 1957, Smith and Erwin 1959) as was originally hypothesized (Laskowski and Laskowski 1951, Hill 1956). Several researchers (Gay 1965, Ingram and Maicornson 1970, Bush et al. 1971) indicate that low serum titers in calves fed adequate amounts of colostrum may be related to intestinal malabsorption. Balfour and Comline (1962) suggest changes in properties of the intestinal epithelial cells may be responsible for the disappearance of this form of absorption. Gay (1965) has reported that Kaeckenbeeck, Colinet and Schoenaers (1961) found absorption of antibody to a somatic antigen of E. coli, is reduced almost 50% at 16 hours after birth. This suggests that cessation of intestinal permeability to colostra! immune globulins is progressive. Possible prenatal or accelerated postnatal closure of the intestinal epithelium to immunoglobulin absorption in some calves reduces significance of the 24-48 hour limitation and emphasizes need for the calf to consume colostrum immediately after birth.
Transmission of Globulin from Intestine to the Circulation .
After absorption from the small intestine the unchanged colos- tral globulins do not enter the portal circulation in appreciable amounts but are carried in lymph to the peripheral blood (Little and Orcutte 1922, McDiarmid 1946, Alamercery, Hamner and Latus 1951, Comline et al. 1951, Balfour and Comline 1962). Comline et al. (1951)

11found an interval of 60 to 120 minutes elapsed between introduction of whey into the duodenum and appearance of colostrum proteins in the lymph. Subsequently, agglutinins were detected in the blood serum 120 to 140 minutes after they first appeared in the lymph. This is in contrast to the smaller molecules of .serum albumin of which a considerable portion directly enters and is transported by the blood (Balfour and Comline 1959) and of even smaller molecules of insulin which almost entirely enter the blood directly (Pierce, Risdall and Shaw 1964).
The Decline of Passive Immunity and Development of Active Immunity
Ingram and Mai corns on (1970) found that colostral antibodiesagainst E. coli attained a serum level in calves of approximately 25%of the amount ingested. McEwan, Fisher and Selman (1970) also observedan efficiency of 25% absorption using actual plasma volume of each ani-
1mal. Balfour and Comline (1962) recovered 12 to 25% of the I-labelled bovine serum globulin from calf lymph within 5 hours after its administration into the duodenum in fresh colostral whey.
Bush, Aguilera and Adams (1971) calculated blood volume according to body weight at one day of age and determined that 45% of the ingested immunoglobulin consumed at birth and at 12 hours was absorbed in the first 24 hours of life. MacDougall and Mulligan (1969) using isotcpically labelled IgG measured absorption in the intravascular compartment and the extravascular pool. A high absorption efficiency of 55% was calculated from their data since no previous account had been
taken of globulin in the extravascular pool (McEwan et al. 1970).

12
Following ingestion of colostrum the serum immunoglobulin reaches its maximum peak about 24 hours and then gradually declines (Klaus et al.
1969, Bush et al. 1971). Smith (1948) noted that globulin acquired from colostrum declined steadily in calves after 2 days of age to about one- half its initial maximum level at 20 days. Thereafter, the rate of decrease was gradual and some globulin persisted in the serum for several months. Accordingly MacDougall and Mulligan (1969) reported the half- life of aglobulin to be about 18 days.
More specifically Smith and Holm (1948) observed that diphtheria antitoxin and two antibodies in colostrum from hyperimmunized cows, fed to newborn calves, declined to one-half their initial serum values in about 16 and 50 days, respectively. Kerr and Robertson (1946) and Pierce (1955a and 1955b) studying immunity to trichomonas foetus determined that the passively transmitted colostra! antibody disappeared from the serum between 17 and 60 days. Graves (1963) calculated the .. antibody half - life of Hoof and Mouth disease to be 15-19 days in three calves. Graves (1963) reported that duration of detectable antibody titer at 80, 130 and about 190 days was dependent on the initial titer achieved from ingested colostrum. True serum half-life of passively ac
quired colostral globulin may be longer since determinations have not accounted for dilution from calf growth (Smith and Holm 1948) .and expansion of plasma volume the first few days of life (McEwan et al. 1970).
The normal calf that has taken colostrum and acquired its maximum concentration of maternal immune globulin by the second day, enters upon a period when the decline in the passively acquired globulin "

13overlaps and tends to obscure the rise in autogenous a-globulin. A point is reached sometime after birth when the a-globulin, concentration . is at a minimum and the loss of passively acquired a-globulin is . equalled by the production of autogenous a-globulin or active immunity (Brambell 1970). Passively acquired immunity is of real importance to the health of the newborn calf for the first 30 days before it is able to synthesize its own active immunity (Hansen and Phillips 1947,Tennant, Harrold, P.eina-Guerra and Laben 1969, Ingram and Malcomson 1970, Ingram, Corbel 1 and Malcomson 1970, Boyd 1972).
Immune Globulin Spectrums and Titers of Colostrum from Individual Cows
In addition to the variability among cows in yield of colostrum and immunoglobulin concentration several researchers (Ingram, Lovell and Wood 1956, Kruse 1958, Gay 1963, Smith 1965, Lambert and Femelius 1968, Royal, Robinson and Duganzich 1968, Ingram et al. 1970 , Rankin ..and Taylor 1970, Watts 1971) have demonstrated that marked voids may occur in the spectrum of colostral antibodies from individual cows resulting in limited or no protection for calves to common bovine pathogens in
their environment.Sarwar, Campbell and Petersen's (1964) observations showed that
in colostrum specimens taken from representative cows from dairy herds in Minnesota, large gaps occur in the spectrum of antibodies, at least as shown by the agglutination technique utilized. They concluded such colostrum may be limited in protective properties insofar as these species of pathogens were concerned. The range of immunoglobulins in the colostrum of different cows immediately after parturition is

14variable. Concentrations range from 1.7 to 8.7% immunoglobulin on a whole colostrum basis with the lower values not associated with any particular breed (Bush et al. 1971).
Highly significant differences between breeds were found in yield of colostrum at first milking after calving and in the concentration of immunoglobulin in colostrum by Kruse (1958). Kruse (1958) also noted that heifers had a lower colostrum and immunoglobulin yield than cows in their second and later lactations. No effect of seasonal variation was found on colostrum yield, immunoglobulin percent and immunoglobulin yield.
The range of immunoglobulin in the colostrum of different cows immediately after parturition was of considerable interest to Bush et al. (1971). They found approximately 68% of the variation in blood serum immunoglobulins in the calves could be attributed to dif
ferences in amount of immunoglobulins consumed per unit of weight. ' The . importance of immunoglobulin content of colostrum from individual cows was readily apparent.
The association between the amount of immune globulin-ingested and the serum immune globulin levels has also been shown by McEwan et al. (1970) and Boyd (1972). The results show a significant relationship between low serum immune globulins in the calves and a higher incidence
of disease and death.Attempts to increase passive immunity in the neonatal calf
via maternal immunization, blood transfusions and oral administration of blood serum from adult animals have given variable results (Smith

15and Little 1922b, Smith and Little 1923, Peterson and Campbell 1955, Graves 1963, Lotan et al. 1964, Watts 1971). Studies show that bovine serum injected sub cutaneously (Smith and Little 1923, Watts 1971) or administered orally (Smith and Little 1922b, Smith and Little 1923, Graves 1963) replaced colostrum in promoting calf survival but colostrum was the most efficient antibody transporting agent.
Although attempts have been made to use pooled blood to enhance passive immunity in calves, similar information is limited relative to the prophylactic value of pooling colostrum for this purpose. McCoy et al. (1968) and Lotan et al. (1964) tried to increase serum immune globulin levels in newborn calves by pooling colostrum. No significant results were obtained by either group because.the emphasis of their experiments was only on increasing serum IgG levels and not increasing the spectrum of immunity in the newborn calves.
Fermented ColostrumPreserving pooled colostrum from several cows to increase the
spectrum of IgG received by newborn calves has been impractical and costly for small dairies. Until recently the only way to extend the colostrum feeding period to 4 or 6 weeks was done by refrigerating and/ or freezing the extra colostrum milk. A new method of storing colostrum by . natural fermentation has, been reported by dairymen (Crowley 1973a and 1973b) both to improve nutrition and reduce the incidence and severity, of digestive disturbances when fed to young calves (Bonewitz 1973, Crowley 1973a, Crowley 1973b, Hutjens 1973) between 2 days of age
and weaning.

16Interest in the use of sour or fermented colostrum for nutri
tional purposes has developed rapidly since researchers in England showed that extra colostrum could be stored by allowing it to ferment. The usual souring or fermentation of milk that occurs under natural conditions results in a breakdown of protein. It is generally recognized that protein denaturation does not occur as rapidly in colostrum as in milk. Why colostrum keeps and why the desirable acid fermentation occurs still need further study. Possibilities suggested are that the acid condition of colustrum (pH 6.0 to 4.0) encourages growth of acid-producing bacteria. The antibodies or protective qualities in the colostrum actually may protect the colostrum from growth of harmful bacteria. Perhaps the colostrum is inoculated naturally with the right bacteria and the composition of the colostrum promotes the right fermentation (Crowley 1973a)»
Otterby and Dutton (1974) have recently observed some of the changes fermenting colostrum goes through. They noted a decrease in pH from 6.1 to 4.0 while the total titratable acidity rose from .04 meq/g to .40 meq/g over a 28-day period. Total solids decreased from 17.00% to 15.00% while total nitrogen also decreased slightly from 10.7 mg/g to 10.2 mg/g. Soluble nitrogen increased over the 28-day period from .44 mg/g to 3.5 mg/g.
The new method of storing colostrum by natural fermentation has
been reported (Morril, Mickelsen and Dayton 1974, Plog, Huber and Oxen- der 1974, Polzin, Johnson and Otterby 1974, White et al; 1974) to inw prove nutrition and reduce the incidence and severity of digestive

17disturbance when' fed to young calves. In this sense the term "colostrum" is generally a misnomer since the fermented product usually represents the colostrum, and other milk collected for several milkings or days after calving. This practice greatly dilutes the IgG concentration.Also calves fed this milk are usually 24 hours of age and have generally lost the ability to absorb colostral antibodies for passive immunity.

CHAPTER 3
PROCEDURE
Twenty-five Holstein male and female calves from The .University of Arizona Dairy Herd were used to conpare the efficiency of immunoglobulin transfer from colostrum milk. The calves were equally assigned to one of the five colostrum feeding regimes: (1)fresh, (2) frozen, (3) fermented, (4) pooled frozen, (5) pooled fermented. Each calf received 500 ml of its assigned colostrum at birth and again at .12 hours. The calves were individually housed for the next 50 days, maintained on milk and allowed water and dry feed ad lib.
'Four blood samples were taken from each calf by jugular vena- puncture using 10 ml vacutainers. (Beeton-Dickinson), Blood was sampled at birth, 12 hours, 24 hours, and 48 hours. The serum was prepared (Brambell 1970) and frozen until analyzed.
Blood serum protein fractions were determined by published methods (Beckman 1963, Beckman 1971) on a Beckman model R-101 Microzone electrophoresis cell. The stained electrophonetograms were quantitatively scanned on a Beckman/Spinco Analytrol model RB fitted with a micro zone scanning attachment model R-102. Total serum protein was determined by refractive index on a Bausch and bomb ABBE-3L Refractometer model RBj using purified beef serum albumin standards.
Specific calf immunoglobulins were determined by immunoelectro- phoresis as outlined by Campbell et al. (1970). Calf serum was
18

19electrcphoretically separated on ion agar (Difco Laboratories) and reacted against specific antiserum (Miles Lab., Inc.): (1) Rabbitanti-bovine whole serum and (2) Rabbit anti-bovine serum IgG. The nomenclature for immunoglobulins for the domesticated bovine will conform to human nomenclature (Rowe 1970) .
The colostrum milk used in this experiment was collected from four University of Arizona Holstein cows and one Holstein cow from Shamrock Dairy, a local commercial dairy herd. The ages of these cows ranged from 2 to 8 years. . These cows calved within 5 days of one another.
Colostrum milk from the first milking after parturition was used from each cow. The colostrum milk was divided into five feeding regimens: (1) fresh colostrum, 1000 ml from each cow, (2) frozencolostrum, 1000 ml from each cow, (3) fermented colostrum, 1000 ml from each cow, (4) pooled frozen colostrum, 1000 ml from each cow, (5) pooled fermented colostrum, 1,500 ml from each cow. The fresh colostrum was kept refrigerated for no longer than 24 hours until it was fed to the calves in the first group. The frozen colostrum samples were immediately frozen in individual 500 ml plastic containers, and thawed out just prior to feeding.
The colostrum used for the third and fifth groups was allowed to ferment after the addition of one-half pint of buttermilk, at room temperature for 2 weeks, or until it reached pH 4.5. One thousand ml of colostrum from each cow was allowed to ferment in individual cloth- covered glass gallon containers. The pooled fermented colostrum was

. 20 obtained by allowing 1,500 ml of colostrum from each cow to ferment in one covered ten-gallon plastic bucket.
Colostrum samples were analyzed for percent protein (Udy 1956) and pH on a Perkin-Elmer Metrion IV pH meter, Coleman 28 C.
Qualitative estimation of specific immunoglobulins in colostrum milk was determined by immunoelectrophoretic separation of colostrum milk serum proteins. Identification was carried out by reacting colostrum milk serum proteins against specific antisera: (1) Rabbit anti-bovine serum serum, and (2) Rabbit anti-bovine serum IgG (Miles Laboratory International) . Refer to publication on nomenclature (Rowe 1970) .
All data was subjected to one-way analysis of variance and significance was computed by student T-test (Steel and Torrie 1960).

CHAPTER 4
RESULTS AND DISCUSSION
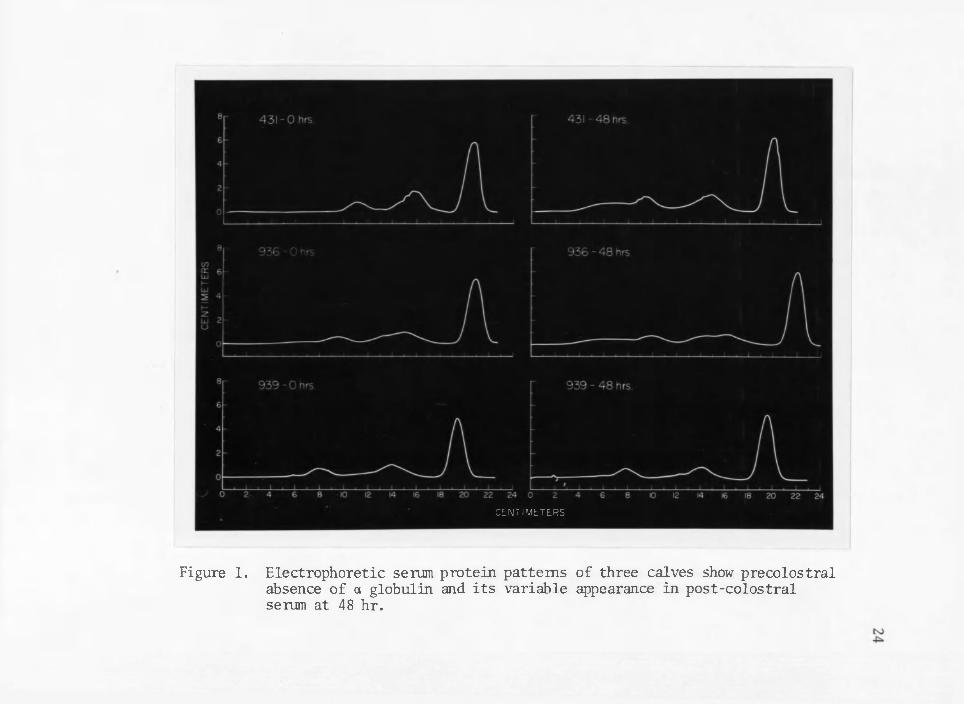
Electrophoretic blood serum protein patterns of three calves (Figure 1) show the characteristic absence, of a globulin in pre- colostral calf serum and its variable appearance in the post-colostra! serum at 48 hours. This data supports the observations of previous researchers (Hansen and Phillips 1947, Smith 1948, Smith and Holm 1948, Johnson and Pierce 1959, Smith and Erwin 1959, Graves 1963, Klaus et al. 1969, Ingram and Mai corns on 1970, Ingram et al. 1970). Although the 48-hour electrophoretic patterns, represent different colostral treat
ments they are presented here to show contrasting differences in serum levels of aglobulins among calves. Calf 431 received fresh.colostrum and absorbed one of the highest concentrations of immunoglobulins observed in this experiment. In contrast calf 939 fed pooled fermented colostrum was the only calf in the study that apparently failed to absorb any a globulin. Calf 936 received pooled frozen colostrum and the electrophoretic pattern demonstrates a serum a globulin level be
tween the two previously mentioned calves.. The calf serum protein fractions are listed by treatment in
Tables 2, 3, 4, 5, and 6 (presented in Appendix I). Serum proteins, globulin, alpha, beta and albumin levels taken from calves at birth,12 hr, 24 hr and 48 hr are included in these tables along with information concerning birth weights and type of colostrum consumed by each
a .

22
calf. Table 6 (see Appendix I) includes the mean values for a globulin, alpha, beta and albumin from each colostrum treatment.
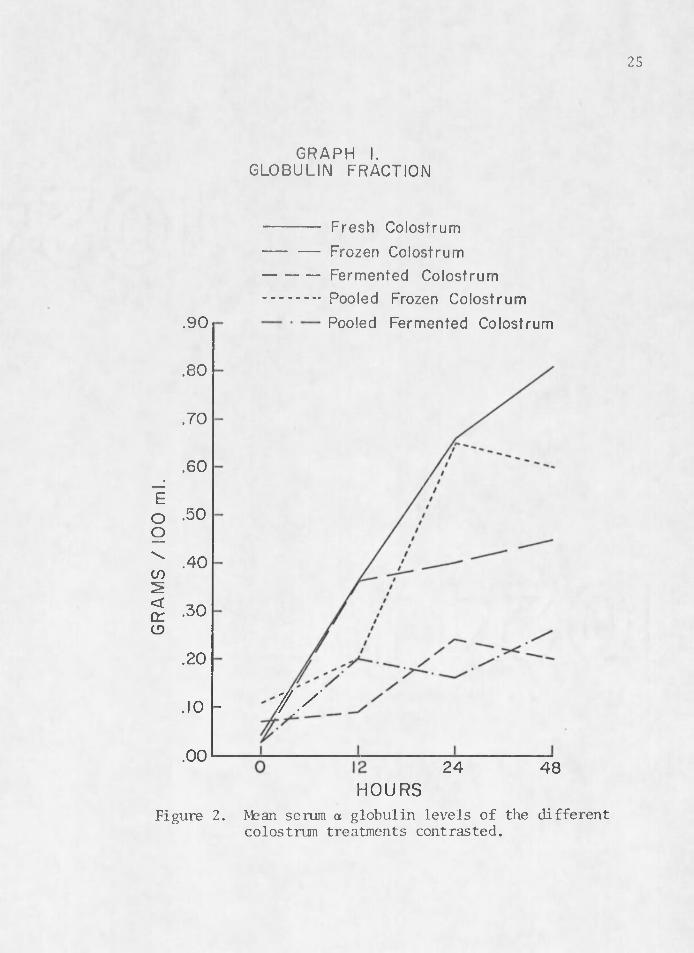
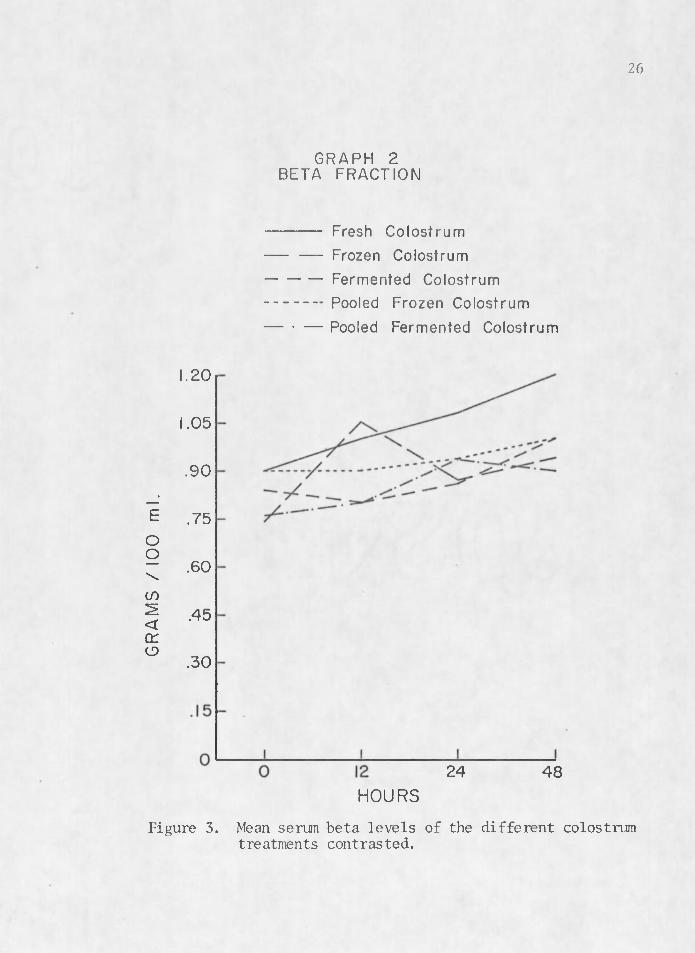
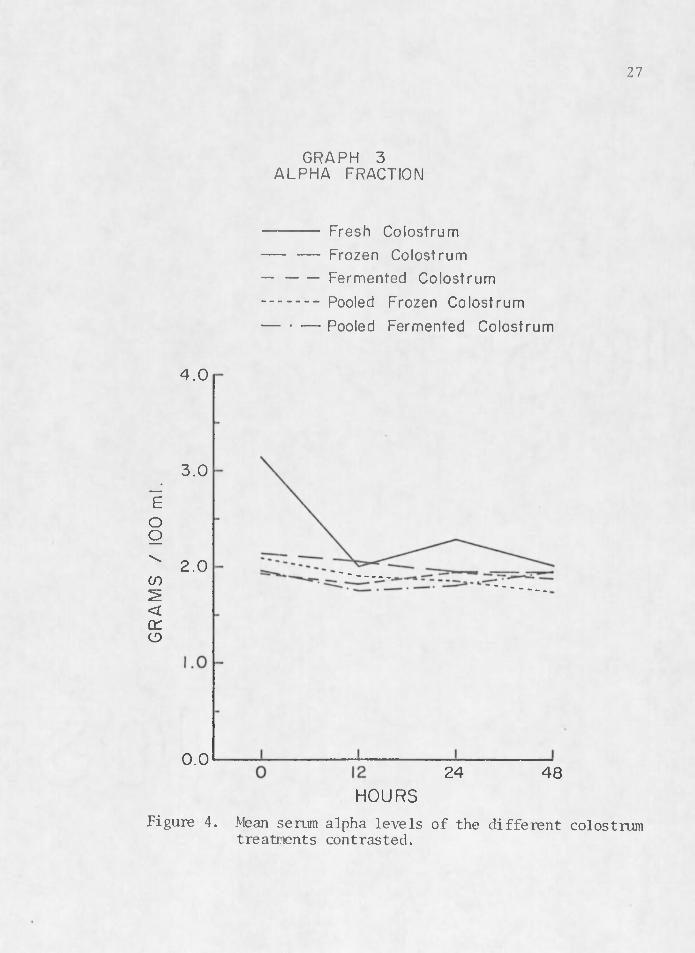
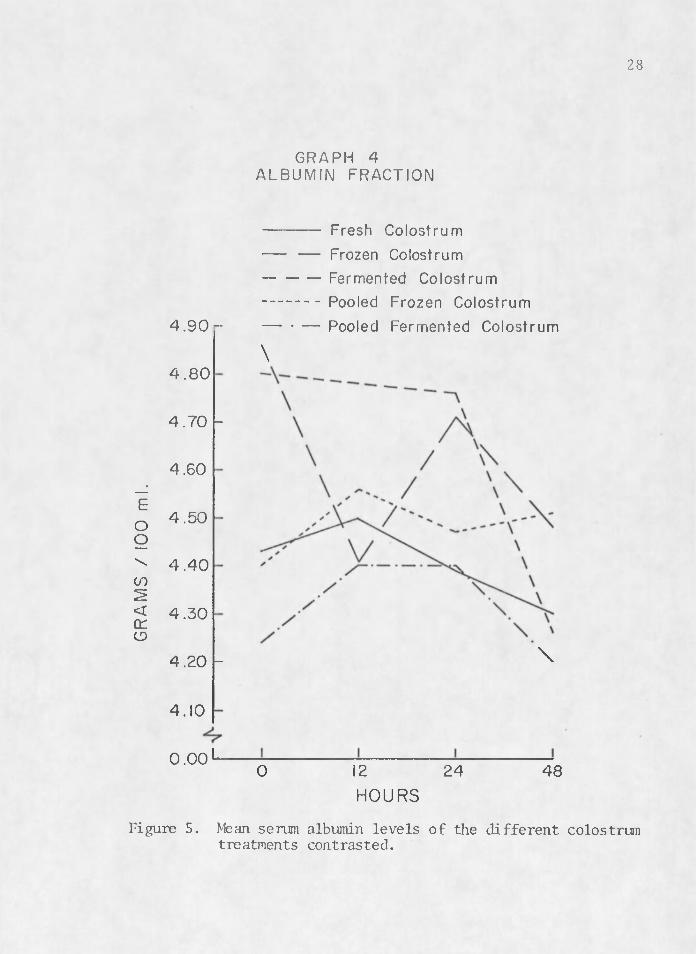
The rise and fall of serum proteins noted by previous researchers (Aschaffenburg 1949, Pierce 1955a, Johnson and Pierce 1959, Tennant et al. 1969) is illustrated in Figures 2, 3, 4, and 5. These graphs depict a rise in serum a globulin and beta levels with a decline in alpha and albumin levels between birth and 48 hr. Figure 2 contrasts mean serum globulin levels between colostrum treatments. Fresh colostrum (1) seems to be the most efficient transporting media for colostral antibodi'es of immunoglobulins. Pooled frozen colostrum (4) also tends to manifest higher titers of serum a globulin in calves which received it. Fermented (3) and pooled fermented colostrum (5) seem to be very ineffective mediums for the transfer of acquired passive im- . munity in the newborn calf.
Immunoelectrcphoretograms of pre- and post-colostral calf
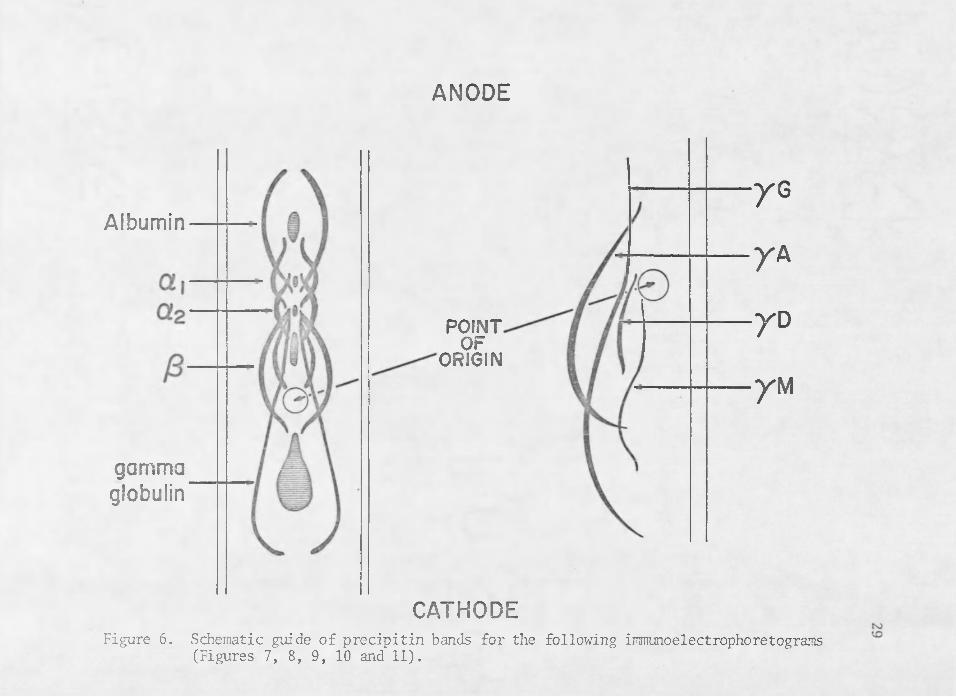
serum (Figure 7) confirm and augment the . results of microzone electrophoretic data (Figure 1) by showing some qualitative changes in immunoglobulins conferred in post-colostral calf serum. In general pre- colostral calf serum shows the often reported condition of agammaglobulinemia to be the result of extremely low levels of IgG and a complete absence of the fractions IgM and IgA (Figure 7). It is worth noting, however, that pre-colostral calf serum does contain precipitin arcs coincident with the electrophoretic mobilities of transferrin and two of the a-globulin protein fractions of serum (Figure 7), as well

23as an arc for serum albumin. This was also indicated in the micro zone electrophoretic data.
Post-colostral calf serum shows some qualitative variations in electrophoretograms which may be attributable to the type of colostrum fed in this e:xperiment. Post-colostral calf serum from calves fed whole fresh or frozen colostrum show all of the precipitin bands found
in adult bovine sera (Figure 8 and 9). However, post-colostral serum from calves fed fermented colostrum demonstrate an absence in the nega-' tive moving conponent of IgG (Figure 10) as well as a seeming complete absence of a precipitin arc for IgM, This apparent abnormality in the IgG component and.the apparent disgammaglobulinemia for IgM could indicate that these serum globulin components are labile to either acid or bacterial degradation during the fermentation process. Further evidence of gammaglobul inop athy (Burnet) is shown when anti-sera prepared in rabbits for each of the colostrum types, fresh, frozen and fermented, was electrophoresed against each 48 hr post-colostral calf sera (Figure 11). Here scaloping of the precipitin arc towards the trough and apparent altered electrophoretic mobilities and improperly shaped precipitin bands of IgG component again indicates a possible partial degradation or qualitative alteration in the IgG fraction of fermented colostrum. Rabbit anti-fermented colostrum sera also showed a complete absence of precipitin bands coincident with the a-globulins and transferrin fractions (Figure 10) while the rabbit anti-fresh and a anti-frozen colostrum contained these fractions. Again this could be evidence of the lability of these components to acid and/or bacterial degradation.
CENTIM ETER S
Figure 1. Electrophoretic serum protein patterns of three calves show precolostral absence of a globulin and its variable appearance in post-colostral serum at 48 hr.
GRA
MS
/ 10
0 m
l.
25
GRAPH I.GLOBULIN FRACTION
----------- Fresh Colostrum— — Frozen Colostrum----------- Fermented Colostrum..............Pooled Frozen Colostrum
.90 Pooled Fermented Colostrum
.80
.70
.60
.50
.40
.30
.20
7//. 1 0 -
.0024 48
HOURSFigure 2. Mean serum a globulin levels of the different
colostrum treatments contrasted.
GRA
MS
/ 10
0 m
l.
26
GRAPH 2BETA FRACTION
-----------Fresh Colostrum— — Frozen Colostrum----------- Fermented Colostrum----------- Pooled Frozen Colostrum— • — Pooled Fermented Colostrum
1.20
1.05
.90
.75
.60
.45
.30
24 48HOURS
Figure 3. Mean serum beta levels of the different colostrum treatments contrasted.
GRA
MS
/ 100
m
l.
27
GRAPH 3ALPHA FRACTION
-----------Fresh Colostrum— — Frozen Colostrum — Fermented Colostrum - Pooled Frozen Colostrum— • — Pooled Fermented Colostrum
4 .0
3.0
2.0
0.024 48
HOURSFigure 4. Mean serum alpha levels of the different colostrum
treatments contrasted.
GRA
MS
/ 10
0 m
l.
28
GRAPH 4ALBUM IN FRACTION
r4 .9 0
4 .8 0
4 .7 0 h
4 .60
4 .5 0
4 .4 0
4 .3 0
4 .20 b
----------- Fresh Colostrum— — Frozen Colostrum —-Fermented Colostrum-----------Pooled Frozen Colostrum— • — Pooled Fermented Colostrum
\
\4.10
0.00 L-0 12 24
HOURS48
Figure 5. Mean serum albumin levels of the different colostrum treatments contrasted.
ANODE
Albumin
gammaglobulin
POINTOF
ORIGIN
rG7A7°7M
Figure 6.CATHODE
Schematic guide of precipitin bands for the following immunoelectrophoretograms (Figures 7, 8, 9, 10 and 11).
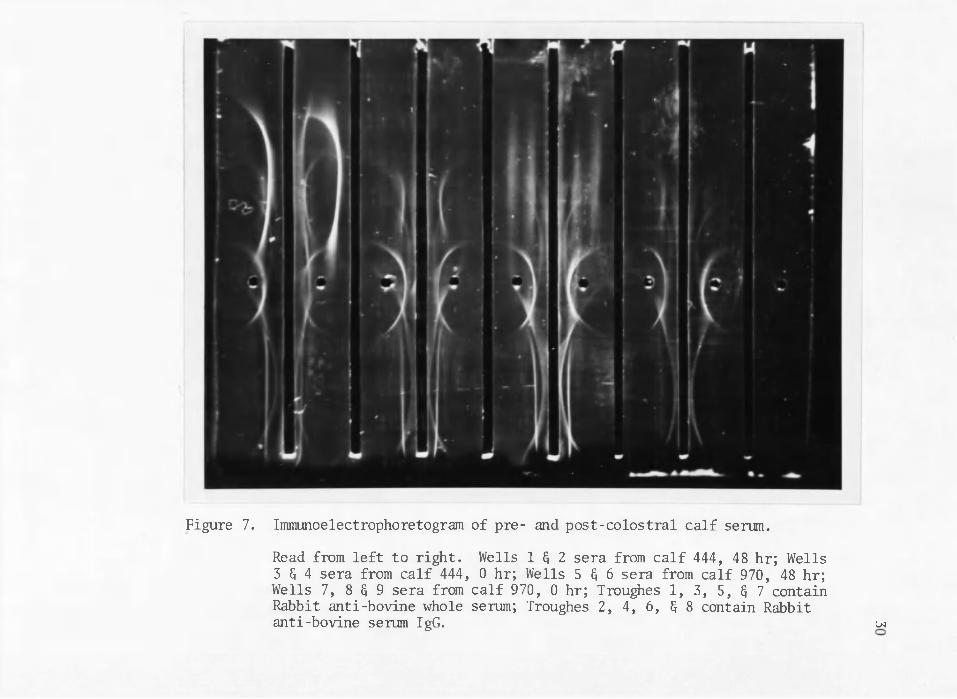
Figure 7. Immunoelectrophoretogram of pre- and post-colostral calf serum.
Read from left to right. Wells 1 G 2 sera from calf 444, 48 hr; Wells 3 G 4 sera from calf 444, 0 hr; Wells 5 § 6 sera from calf 970, 48 hr;Wells 7, 8 G 9 sera from calf 970, 0 hr; Troughes 1, 3, 5, § 7 containRabbit anti-bovine whole serum; Troughes 2, 4, 6, $ 8 contain Rabbitanti-bovine serum IgG. cm
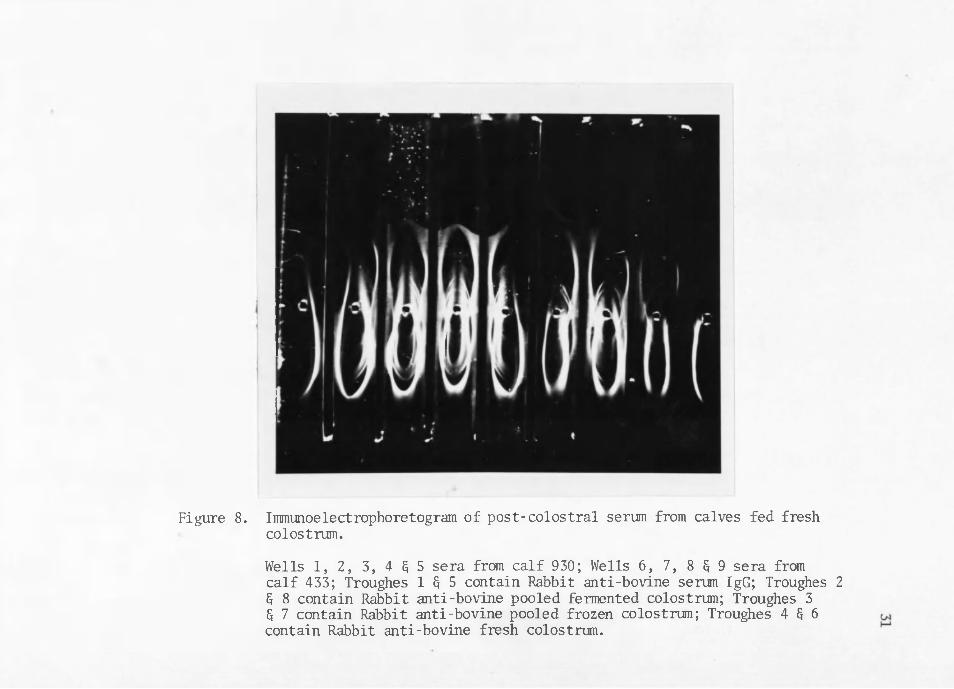
Figure 8. Immunoelectrophoretogram of post-colostral serum from calves fed fresh colostrum.Wells 1, 2, 3, 4 § 5 sera from calf 930; Wells 6, 7, 8 G 9 sera from calf 433; Troughes 1 § 5 contain Rabbit anti-bovine serum IgG; Troughes 2 § 8 contain Rabbit anti-bovine pooled fermented colostrum; Troughes 3 G 7 contain Rabbit anti-bovine pooled frozen colostrum; Troughes 4 § 6 contain Rabbit anti-bovine fresh colostrum.
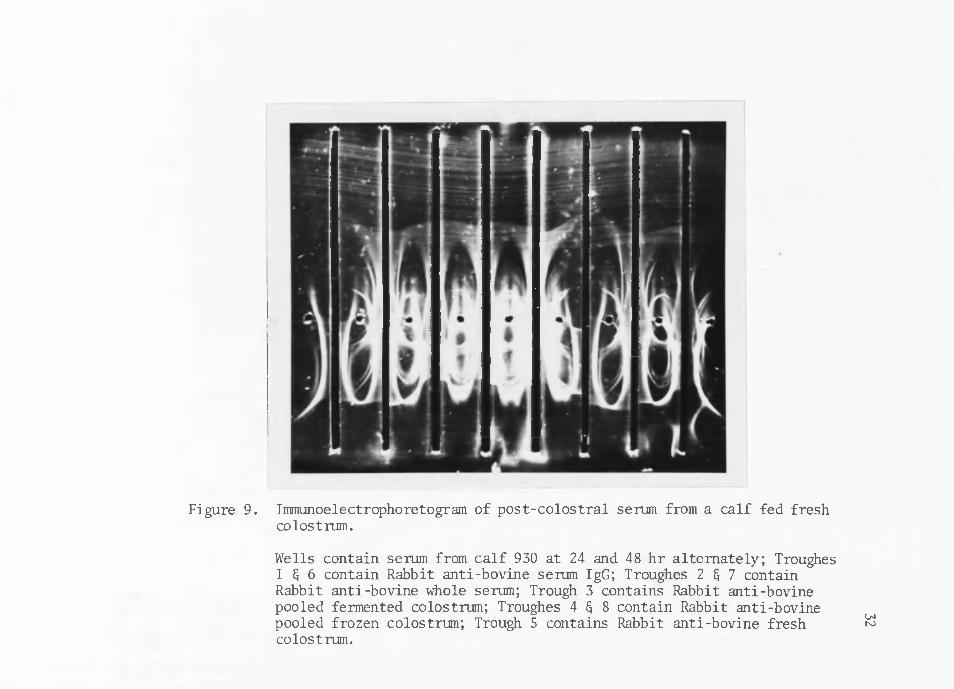
Figure 9. Immunoelectrophoretogram of post-colostral serum from a calf fed fresh colostrum.
Wells contain serum from calf 930 at 24 and 48 hr alternately; Troughes 1 § 6 contain Rabbit anti-bovine serum IgG; Troughes 2 § 7 contain Rabbit anti-bovine whole serum; Trough 3 contains Rabbit anti-bovine pooled fermented colostrum; Troughes 4 § 8 contain Rabbit anti-bovine pooled frozen colostrum; Trough 5 contains Rabbit anti-bovine fresh ^colostrum.

Figure 10. Immunoelectrophoretogram.Wells contain serum from calf 437, 48 hr. This calf received fermented colostrum. Troughes 1 § 8 contain Rabbit anti-bovine whole serum j Troughes 2 § 7 contain Rabbit anti -bovine serum IgG, Troughes 3 S 4 contain Rabbit anti-human transferin; Troughes 5 G 6 contain Rabbit anti-colostrum.
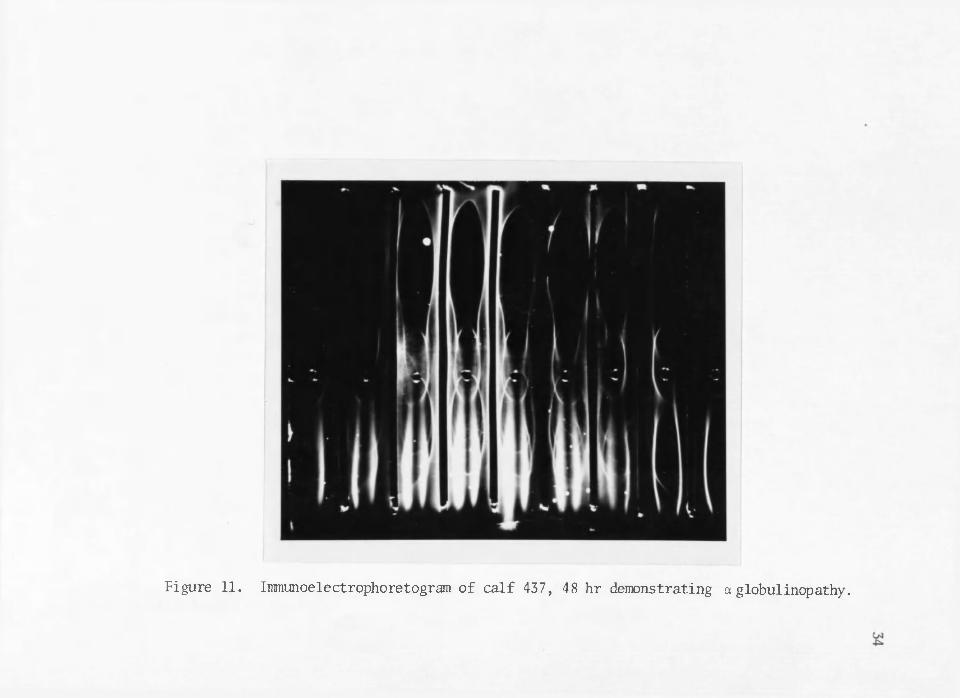
Figure 11. Immunoelectrophoretogram of calf 437, 48 hr demonstrating a globulinopathy.

APPENDIX 1 DATA ON THE FIVE TREATMENTS
35
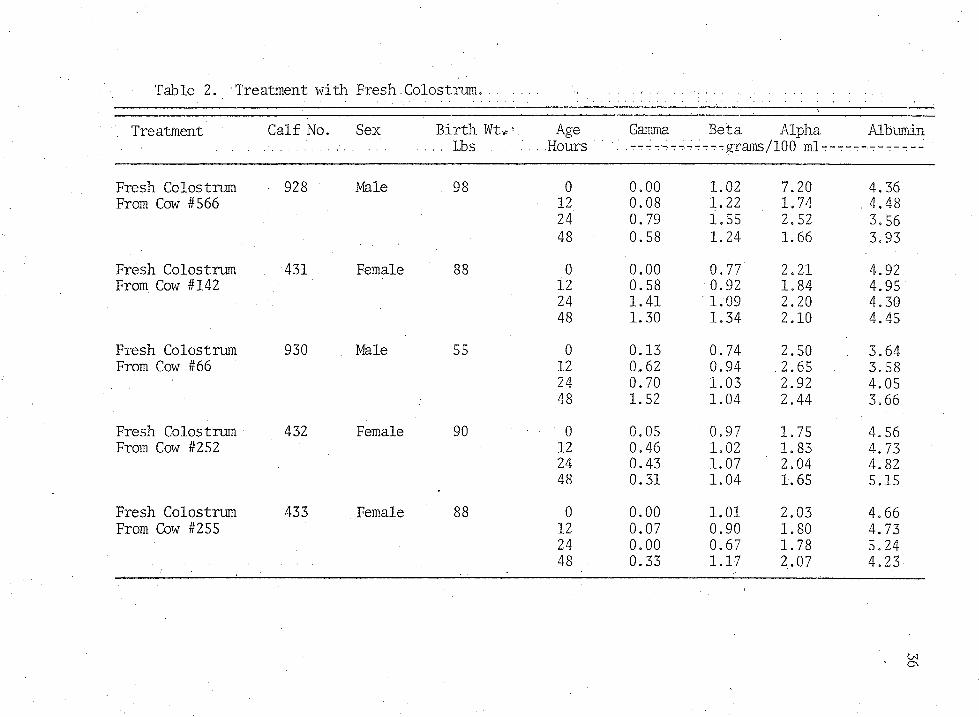
Table 2. Treatment with.Fresh. Colostrum.
Treatment Calf No. Sex Birth Wt*-. Age Gamma Beta Alpha Albumin . . . . Lbs Hours .---:-:----:-:-:-:-grams/100 ml-:--:---'-?------
Fresh Colostrum 928 Male 98 0 0.00 1.02 7.20 4.36From Cow #566 12 0.08 1.22 1.74 . 4.48
24 0.79 1.55 2.52 3,5648 0.58 1.24 1.66 3.93
Fresh Colostrum . 431 Female 88 0 0.00 0.77 2.21 4.92From Cow #142 12 0.58 0.92 1.84 4.95
24 1.41 1.09 2.20 4.3048 1.30 1.34 2.10 4.45
Fresh Colostrum 930 . Male 55 0 0.13 0.74 2.50 . 3.64From Cow #66 12 0.62 0.94 .2.65 3.58
24 0.70 1.03 2.92 4.0548 1.52 1.04 2.44 3,66
Fresh Colostrum 432 Female 90 0 0.05 0.97 1.75 4.56From Cow #252 12 0.46 1.02 1.83 4.73
24 0.43 1.07 2.04 4.8248 0.31 1.04 1.65 5.15
Fresh Colostrum 433 Female 88 0 0.00 1.01 2.03 4.66From Cow #255 12 0.07 0.90 1.80 4.73
24 0.00 0.67 1.78 5.2448 0.33 1.17 2.07 4.23
Cn$Gh
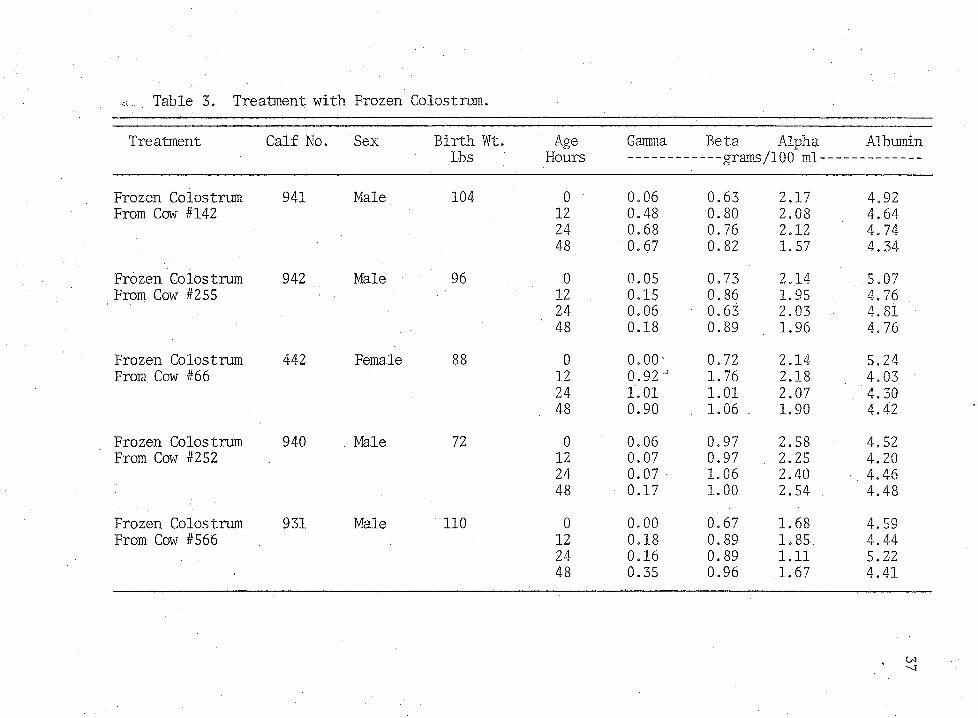
Table 3. Treatment with Frozen Colostrum.
Treatment Calf No. Sex Birth Wt. Lbs
AgeHours
Gamma Beta Alpha --- grams/100 ml- -
Albumin
Frozen Colostrum 941 Male 104 0 0.06 0.63 2.17 4.92From Cow #142 12 0.48 0.80 2.08 4.64
24 0.68 0.76 2.12 4.7448 0.67 0.82 1.57 4.34
Frozen Colostrum 942 Male 96 0 0.05 0.73 2.14 5.07From Cow #255 ■ 12 0.15 0.86 1.95 4.76
24 0.06 0.63 2.03 4.8148 0.18 0.89 1.96 4.76
Frozen Colostrum 442 Female 88 0 o.oo- 0.72 2.14 5.24From Cow #66 12 0.92" 1.76 2.18 4.03
24 1.01 1.01 2.07 4.30. 48 0.90 . 1.06 . 1.90 . 4.42
Frozen Colostrum 940 . Male 72 0 0.06 0.97 2.58 4,52From Cow #252 12 0.07 0.97 . 2.25 4.20
24 0.07 1.06 2.40 4.4648 0.17 1.00 2.54 4.48
Frozen Colostrum 931 Male 110 0 0,00 0.67 1.68 4.59From Cow #566 12 0.18 0.89 1.85 4.44
24 0.16 0.89 1.11 5.2248 0.35 0.96 1.67 4.41
04
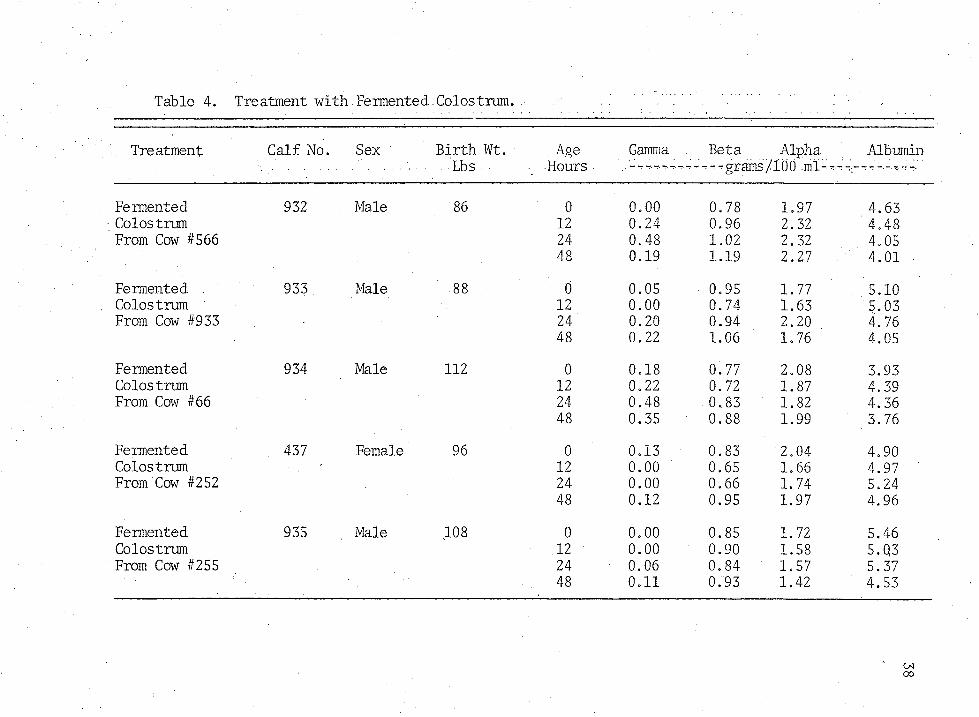
Table 4. Treatment with.Fermented.Colostrum.
Treatment Calf No. Sex Birth Wt, . ......'Lbs...
. Age Gamma Beta Alpha Albumin ... - -TT. --r.-7.~ - - - -r grams/100 ml - - - - t -.-.77. -
Fermented 932 Male 86 0 0.00 0.78 1.97 4.63Colostrum 12 0.24 0.96 2.32 4.48From Cow #566 24 0.48 1.02 2.32 4.05
48 0.19 1.19 2,27 . 4.01Fermented . 933 Male 88 0 0.05 0.95 1.77 5.10Colostrum 12 0.00 0.74 1.63 5.03From Cow #933 24 0.20 0.94 2.20 4.76
48 0.22 1.06 1.76 4.05Fermented 934 Male 112 0 0.18 0.77 2.08 3.93Colostrum 12 0.22 0.72 1.87 4.39From Cow #66 24 0.48 0.83 ' 1.82 4.36
48 0.35 0.88 1.99 3.76Fermented . 437 Female 96 0 0.13 0.83 2.04 4.90Colostrum 12 0.00 0.65 1.66 4.97From Cow #252 24 0.00 0.66 1.74 5.24
48 0.12 0.95 1.97 4.96Fermented 935 Male 108 0 0.00 0.85 1.72 5.46Colostrum 12 0.00 0.90 1.58 5.0,3From Cow #255 . 24 0.06 0.84 1.57 5.37
48 0.11 0.93 1.42 4.53
0300
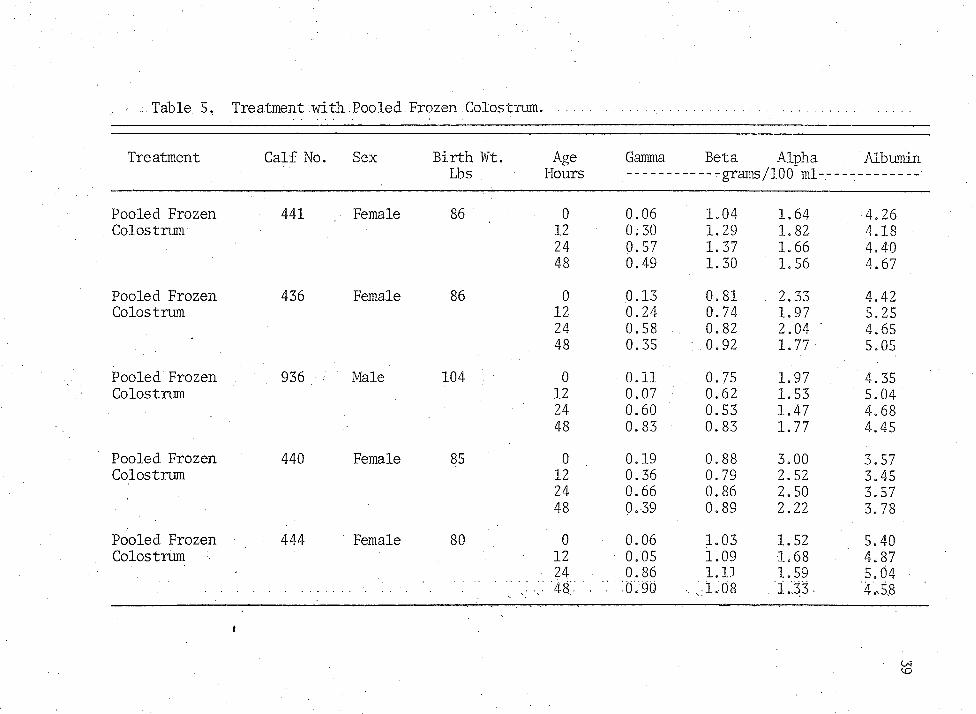
Table 5. Treatment with .Pooled .Frozen.Colostrum.
Treatment Calf No. Sex Birth Wt. Lbs
AgeHours
Gamma Beta Alpha -- -grams/100 ml— -
Albumin
Pooled Frozen 441 Female 86 0 0.06 1.04 1.64 4.26Colostrum 12 0.30 1.29 1.82 4.18
24 0.57 1.37 1.66 4,4048 0.49 1.30 1.56 4.67
Pooled Frozen 436 Female 86 0 0.13 0,81 . 2.33 4.42Colostrum 12 0.24 0.74 1.97 5.25
24 0.58 0.82 2.04 " 4.6548 0.35 : 0.92 1.77 5.05
Pooled Frozen 936 < . Male 104 ' 0 0.11 0.75 1.97 4.35Colostrum 12 0.07 0.62 1.53 5.04
24 0.60 0.53 1.47 4.6848 0.83 0.83 1.77 4.45
Pooled Frozen 440 Female 85 0 0.19 0.88 3.00 3.57Colostrum 12 0.36 0.79 2.52 3.45
24 0.66 0.86 2.50 3.5748 0.39 0.89 2.22 3.78
Pooled Frozen 444 Female 80 0 0.06 1.03 1.52 5.40Colostrum 12 0.05 1.09 1.68 4.87
24 0.86 l.ll 1.59 5.04'48/ . .0.90 .. vi.08 1 ..33 ■ '4,5.8
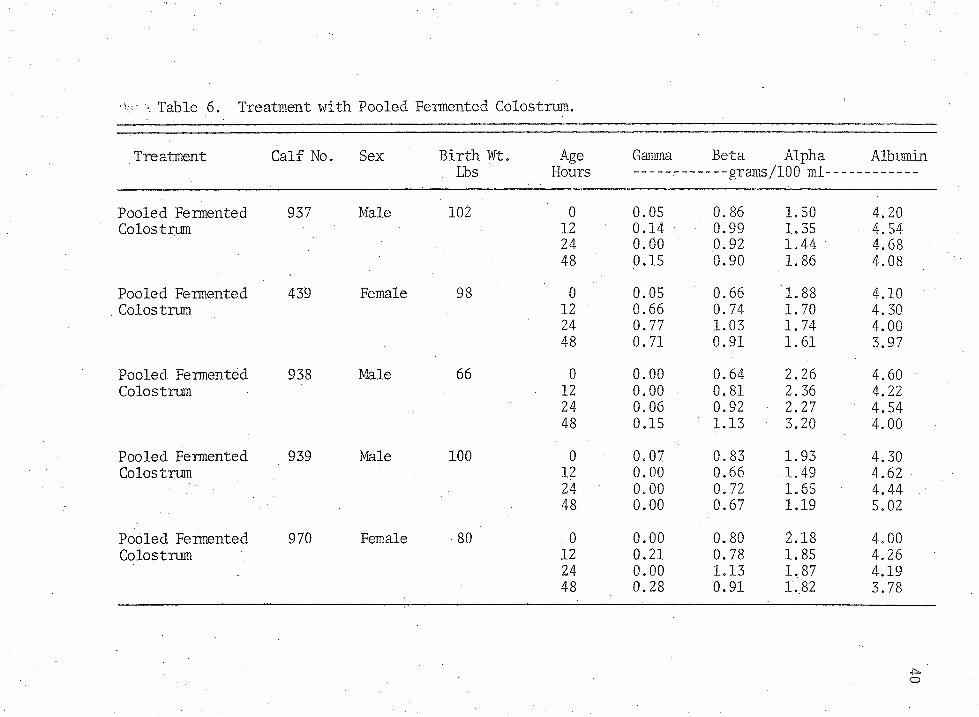
Table 6. Treatment with Pooled Fermented Colostrum.
Treatment Calf No. Sex Birth Wt. . Lbs
AgeHours
Gamma Beta Alpha ---- grams/100 ml—
Albumin
Pooled Fermented 937 Male 102 0 0.05 0.86 1.50 4.20Colostrum 12 0.14 0.99 1.35 4.54 .
24 0.00 0.92 1.44 4.6848 0.15 0.90 1.86 4.08
Pooled Fermented 439 Female 98 0 0.05 0.66 1.88 4.10Colostrum 12 0.66 0.74 1.70 4.30
24 0.77 1.03 1.74 4.0048 0.71 0.91 1.61 3.97
Pooled Fermented 938 Male 66 0 0.00 0.64 2.26 4.60Colostrum 12 0.00 0.81 2.36 4.22
24 0.06 0.92 • 2.27 4.5448 0.15 ' 1.13 • 3.20 4.00
Pooled Fermented 939 Male 100 0 0.07 0.83 1.93 4.30Colostrum 12 0.00 0.66 . 1.49 4.62
24 0.00 0.72 1.65 4.4448 0.00 0.67 1.19 5.02
Pooled Fermented 970 Female ■ 80 . 0 0.00 0.80 2.18 4.00Colostrum 12 0.21 0.78 1.85 4.26
24 0.00 1.13 1.87 4.1948 0.28 0.91 1.82 3.78
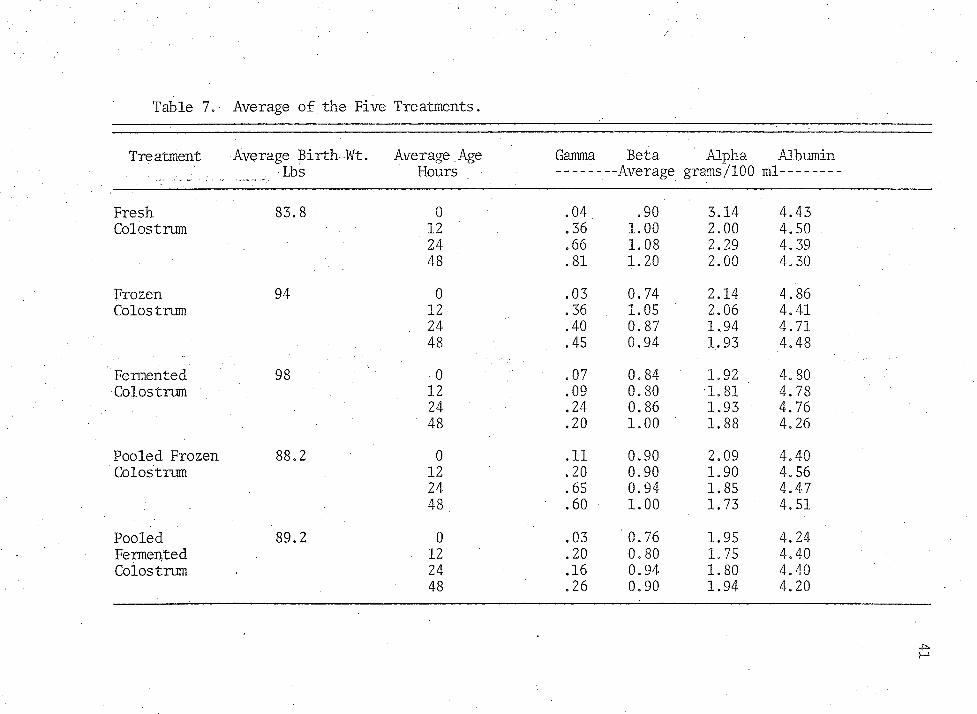
Table 7,- Average of the Five Treatments.
Treatment Average Birth-Wt. Average .Age Gamma Beta Alpha AlbuminLbs Hours --------Average grams/100 ml------
Fresh 83.8 0 • 04. .90 3.14 4.43Colostrum 12 .36 1.00 2.00 4.50
24 .66 1.08 2.29 4.3948 .81 1.20 2.00 4.30
Frozen 94 0 .03 0.74 2.14 4.86Colostrum 12 .36 1.05 2.06 4.41
24 .40 0.87 1,94 4.7148 .45 0.94 1, 93 4.48
Fermented 98 0 .07 0.84 1.92 4.80Colostrum 12 .09 0.80 •1.81 4.78
24 .24 0.86 1,93 4.7648 .20 1.00 1.88 4.26
Pooled Frozen 88.2 0 .11 0.90 2.09 4.40Colostrum 12 .20 0.90 1.90 4.56
24 .65 0.94 1.85 4.4748 . .60 1.00 1.73 4.51
Pooled 89.2 0 .03 0.76 1.95 4.24Fermented 12 .20 0.80 1.75 4.40Colostrum 24 .16 0.94 1.80 4.40
48 .26 0.90 1.94 4.20

LITERATURE CITED
Alamercery, J., C. L, Hamner and M. Latus. 1951. Histological Changesin the Epithelium of the Small Intestine During Protein Absorption in the Newborn Animal. Nature, Vol. 168, No. 4263; 84.
Aschaffenburg, R. 1949.. Changes in the Serum Protein of the NewbornCalf Following the Ingestion of Small Quantities of the Nonfatty Fraction. Brit. J..Nutrit., Vol. 3; 200.
Askonas, B. A., P. N. Campbell, J. H. Humphrey and T. S. Work. 1954.The Source of Antibody Globulins in Rabbit ML Ik and Goat Colostrum. Biochem. J. 56; 597.
Balfour, W. E. and R. S. Comline. 1959. The Specificity of the Intestinal Absorption of Large Molecules by the Newborn Calf. "J". Physiol., London 148; 77.
Balfour, W. E. and R. S. Comline. 1962. Acceleration of the Absorption of Unchanged Globulin in the New-born Calf by Factors in Colostrum. J. Physiol., 160; 234.
Beckman. 1963. Parts List. Published by Beckman Instruments, Inc.,2500 Harbor Blvd., Fullerton, California.
Beckman. 1971. Methods Manual and Instruction Manual.. Published bySpinco Division of Beckman Instruments, Inc., Stanford Industrial Park, Palo Alto, California.
Becton-Dickinson Vacutainer. Division of Becton, Dickinson and Company, Rutherford, New Jersey.
Bonewitz, R. 1973. First Five Hours Important for New Calves. The Modem Dairyman, March 17; 35.
Boyd, J. W. 1972. The Relationship between Serum Immune Globulin Deficiency and Disease in Calves; A Farm Survey. Veterinary Record, 90; 645.
Brambell, F,. W. R. 1970. The Transmission of Passive Immunity from Mother to Young. American Elsevier Publishing Company, Inc.,New York.
Brambell, F. W. R. , W. A. Hemmings and M. Henderson. 1951. Antibodies and Embryos. University of London Athlone Press.
42

43
Brown, R. D. 1958. Rinderpest Immunity in Calves. I. The Acquisition and Persistence of Maternally Derived Antibody. J. Hyg., Camb. 56; 427. '
Burnet, M. 1959. The clonal selection theory of antibody production. Oxford University Press, London and New York.
Bush, L. J., M. A. Aguilera and G. D. Adams . 1971. Absorption ofColostral Immunoglobulines by Newborn Dairy Calves. J. of Dairy Sci.., Vol. 54, No. 10: 1547.
Campbell, D. H., J. S. Garvey, N. E. Cremer and D. H. Sussdorf. 1970. Methods in Immunology. W. A. Benjamin. Inc., New.York.
Comline, R. S., H. E. Roberts and D. A. Titchen. 1951. Route of•Absorption of Colostrum Globulin in the Newborn Animal. Nature, Vol. 167, No. 4249; 561.
Crowley, J. W. 1973a. Interest Grows in "Sour" or "Pickled" Colostrum. Hoard's Dairyman, Vol. 118; 614.
Crowley, J. W. 1973b, The Feeding Value of Colostrum Varies, Hoard's Dairyman, Vol. 188; 685.
Dam, A. 1968. Studies on the Gammaglobulin Levels in Sera of Calves from Herds with Colisepticaemia as a Problem, and Same Investigations on the Content of Specific Antibodies in Colostrum. Nord. Vet. Med. 20; 449.
Deutsch, H. F. and V. R. Smith. 1957. Intestinal Permeability toProtein in the. Newborn Herbivore. Amer. J. Physiol. 191; 271.
Difco Laboratories, Detroit Michigan, U.S.A.Ferris, T. A. and J. W. Thomas. 1974. Relationship of Immunoglobulin
to Dairy Calf M rtality and Influence of Herd Environment,. J. of Dairy Sci. Vol. 57, No..5; 641.
Fey, H. and A. Margandent. 1962. Hypergamrnaglobulinemic' Bie Der Der.Colisepsis Des Kalbes. Path. Microbiol. 24; 970.
Gay, C. C. 1965. Escherichia Coli and Neonatal Disease of Calves. Bacteriological Reviews. Vol. 29, No. 1; 75.
Graves, J. H. 1963. Transfer.of Neutralizing Antibody by Colostrum to Calves Bom of Foot-and-mouth Disease Vaccinated. Dams. J.Immun. 91; 251.

44Hansen, R. G. and P. H. Phillips. 1947. Studies on Proteins from
Bovine Colostrum. J. Biol. Chem., 171; 223.Hansen, R. G., P. H. Phillips and V. R. Smith. 1946. Colostrum Milk
and Its Vitamin A Content. J. Dairy Sci. 29; 809.Henderson, J. L. 1971. The Fluid-MiIk Industry. Third Edition.
Westport, Connecticut, The Avi Publishing Company, Inc. 28.Hill, K. J. 1956. Gastric Development and Antibody Transference in
the Lamb with Some Observations on the Rat and Guinea Pig. Quart. J. Exp. Physiol. 42; 72.
Howe, P. E. *1921. An Effect of the Ingestion of Colostrum upon the Composition of the Blood of New-Born Calves. J. Biol. Chem.,49; 115.
Hutjens, M. F. 1973. Sour Colostrum. Animal Nutrition and Health,May; 10.
Ingram, D. G., L. B. Corbeil and M. E. Mai corns on. 1970. Antibodies toEscherichia Coli in Young Calves; K Antigens. Am. J. Vet.Res., Vol. 31, No. 1; 71.
Ingram, P. L., R. Lovell and P. C. Wood. 1956. Bacterium Coli Antibodies in Colostrum and Their Relation to Calf Survival. J.Path. Bact., Vol. LXXII; 561.
Ingram, D. G., M. E. Mai corns on. 1970. Antibodies to EscherichiaColi in Young Calves: 0 Antigens. Am. J. Vet. Res.Vol. 31, No. 1; 61.
Jameson, E., C. Alvarez-Tostado and H. H. Sortor. 1942. ElectrophoreticStudies on New-Born Calf Serum. Proc. Soc. Exp. Biol. Med. ,51; 163.
Johnson, P. and A. E. Pierce. 1959. Ultracentrifugal and Electrophoretic Studies on Neonatal Calf Sera and Maternal Colostrum.J. Hyg., Vol. 57; 309.
Kaeckenbeeck, A., G. Colinet, and F. Schoenaers. 1961. Evolution de 1’aptitude de 1'intestine du veaux nouveau-ne a resorber les anti corps apportes par le colostrum. Ann. Med. Vet. 105: 197.
Kerr, W. R. and M. Robertson. 1946. A study of the Passively Acquired Antibody to Tr. Foetus in the Blood of Young Calves and Its Behaviour in Agglutination Tests and Intradermal Reactions. J.Comp. Path. Ther. 56; 38.

45Klaus, G. G. B. , A. Bennett and E. W, Jones. 1969. A Quantitative
Study of the Transfer of Colostral Immunoglobulins to the Newborn Calf. Immunology, Vol. 16; 293.
Kruse, V. 1958. Yield of Colostrum gnd Immunoglobulin in Cattle at the First Milking after Parturition. Anim. Prod. 12; 619.
Kulangara, A. C. and A. M. Schechtman. 1963. Do Heterologous Proteins Pass from Mother to Fetus in Cow, Cat and Guinea Pig? Proc.Soc, Exp. Biol. Med. 112; 220.
Lambert, G. and A. L. Femelius. 1968. Bovine Viral Diarrhea Virus and Escherichia Coli in Neonatal Calf Enteritis. Can, J.Comp. Med., Vol. 32, No. 2; 440,
Larson, B. L. 1958. Transfer of Specific Blood Serum Proteins to Lacteal Secretions Near Parturition. J. of Dairy Sci. 41;1045.
Larson, B. L. and D. C. Gillespie. 1957. Origin of the Major Specific Proteins in Milk. . J. Biol. Chem. 227; 565.
Larson, B. L. and K. A. Kendall. 1957. Changes in Specific BloodSerum Protein Levels Associated with Parturition in the Bovine. J. Dairy Sci. 40; 659.
Laskowski, M. and M. Laskowski. 1951. Crystalline Trypsin Inhibitor from Colostrum. J. Biol. Chem. 190; 563.
Lecce, J. G. 1966. Absorption of Macromolecules by Neonatal Intestine. Biol. Neonat. 9; 50.
Little, R. B., M. L. Orcutte. 1922. The Transmission of Agglutinins of Bacillus Abortus from Cow to Calf in the Colostrum. J. Exp.Med., 35; 161.
Lotan, E., A.. Berman, A. Tadmor and K. Perk. 1964. The Efficiency of Pooled Gamma Globulins in Preventing Scours in Israeli-Holstein Calves. Brit. Vet. J., Vol. 120, No. 2; 576,
MacDougall, D. F. and W. Mulligan. 1969. The Distribution and Metabolism of Fast IgG Immunoglobulin in the Neonatal Calf. J. Physiol., London. 201; 77.
McALpine, J. G. and L. F. Rettger. 1925. Serological Studies on Bovine Infectious Abortion. J. Immun. 10; 811.

46McCoy, G. C., A. G. Hunter, and J, B. Williams. 1968. Effects of Diet
and Time on the Circulating Gamma Globulin Concentration in thi Newborn Calf. Journal of Dairy Science, Vol. 51, No. 6; 964.
McDiarmid, A. 1946. The Transference of Agglutinins for Brucella Abortus from Cow to Calf and Their Persistence in the Calf's Blood. Vet. Rec. 58; 146.
McEwan, A. D. , E. W. Fisher and I. E. Selman. 1970. An Estimation ofthe Efficiency of the Absorption of Immune Globulins from Colostrum by Newborn Calves. Res. Vet. Sci., Vol. 11; 239.
Miles Laboratories, Inc., Research Products Div., Kankakee, Illinois,60901.
Morril, J. L., R. Mickelsen, and A. D. Dayton. 1974. Sour Colostrum,Cultured Milk, and Antibiotic for Young Calves. J. Dairy Sci.Vol. 57, No. 5; 643.
Orcutt, M. L. and P. E. Howe. . 1922. The Relation Between the Accumulation of Globulins and the Appearance of Agglutinins in the Blood of Newborn Calves. J. Exp. Med., Vol. 36; 291.
Otterby, D. E. and R._ E. Dutton. 1974. Comparative Fermentations of Cows' Colostrum. J. of Dairy Sci. Vol. 57, No. 5; 642.
Peterson, W. E. and B. Campbell. 1955. The use of Protective Principle in Milk and Colostrum in Prevention of Diseases in Man and Animals. J. Lancet. 75; 491.
Pierce, A. E. 1955a. Electrophoretic and Immunological Studies on Sera from Calves from Birth to Weaning. I, Electrophoretic Studies. J. Hyg., Camb. Vol. 53, No, 3; 247.
Pierce, A. E. 1955b. Electrophoretic and Immunological Studies on Sera from Calves from Birth to Weaning. II. Electrophoretic and Serological Studies with Special Reference to the Normal and Induced Agglutinins to Trichomonas Foetus. J. Hyg.,. Camb. 53; 261.
Pierce, A. E. and A. Feinstein. 1965. Biophysical and Immunological Studies on Bovine Immune Globulins with Evidence for Selective Transport within the Mammary Gland from Maternal Plasma to Colostrum. Immunology, Vol. 8; 106.
Pierce, A. E., P. C. Risdall and B. Shaw. 1964. Absorption of Orally Administered Insulin by the Newly Born Calf. J. Physiol.,London, 171; 203.

47Plog, J., J. T. Huber and W. Oxender. 1974. Growth, Diarrhea and
Gamma Globulin of Calves fed Frozen and Fermented Colostrum. J. of Dairy Sci. Vol. 57, No. 5; 642.
Polzin, H. W., D. G. Johnson and D. E. Otterby. 1974. Sour Colostrum or Milk Replacer for Rearing Calves. J. Dairy Sci. Vol. 57, No.5; 642.
Rankin, J. D. and R. J. Taylor. 1970. An Attempt Passively to Immunise Calves Against Salmonella Infection by Vaccination of Their Dams. The Veterinary Record, Vol. 86; 254.
Ratner, B., H. C. Jackson and H. L. Gruehl. 1927. Transmission ofProtein Hypersensitiveness from Mother to Offspring IV Passive Sensitivization in Utero. J. Immun. 14; 291.
Rowe, D. S. 1970. Human Nomenclature. Nature,. London. 228; 509,Royal, W. A., R. A. Robinson and D. M. Duganzich. 1968. Colostral
Immunity against Salmonella Infection in Calves. New Zealand Veterinary Journal, Vol. 16; 141.
San Clemente, C. L. and I. F. Huddleson. 1943. Electrophoretic Studies of the Proteins of Bovine Serums with Respect to Brucella. Michigan Agr. Exp. Sta. Bull., 182; 3.
Sarwar, M., B. Campbell and W. E. Petersen. 1964. Observations on the Presence and Concentration of Natural Antibodies in Colostrum.
: Can. J. Comp. Med. Vet. Sci., Vol. 28; 157.Smith, E. L. .1948. The Isolation and Properties of the Immune Proteins
of Bovine Milk and Colostrum and Their Role in Immunity; A Review. J. of Dairy Sci., Vol. 31; 127.
Smith, E, L. and August Holm. 1948. The. Transfer of Immunity to the Newborn Calf from Colostrum. J. of Biochem., Vol. 175, No.1; 349.
Smith, H. C. 1965. The Perennial Problem of Calf Scours. Modem Veterinary Practice, Vol. 46, No. 6; 51.
Smith, H. W., J. A. O'Neil and E. J. Simmons. 1967. The Immune GlobulinContent of the Serum of Calves in England. Veterinary Record, Vol. 80, No. 23; 664. ,
Smith, T. 1930. The. Immunological Significance of Colostrum. J. of Exp. Med., Vol. 51; 473.

48Smith, T. and R. B. Little. 1922a. The Significance of Colostrum to
the New-Born Calf. J. Exp. Med. 36; 181.Smith, T. and R. B. Little. 1922b. Cow serum as a Substitute for
Colostrum in Newborn Calves. J. Exp. Med. 36; 453.Smith, T. and R. B. Little. 1923. The Absorption of Specific Aggluti
nins in Homologous Serum Fed to Calves During the Early Hours of Life. J. Exp. Med. Vol. 37, No. 3; 671.
Smith, V. R. 1959. Physiology of Lactation. Fifth Edition. IowaState University Press, Ames Iowa, 172.
Smith, V. R. and E. S. Erwin. 1959. Absorption of Colostrum Globulins Introduced Directly into the Duodenum. J. of Dairy Sci.,Vol. 42, No. 2; 364.
Steel, R. and J. H. Torrie. 1960. Principles and Procedures of Statistics. McGraw-Hill Book Company, Inc., New York.
Stewart, J. and J. W. McCallum. 1942. The Effect of a Vitamin ARich Diet on the Vitamin A Content of the Colostrum of Dairy Cows. J. of Dairy Res. 13; 1.
Tennant, B., D. Harm Id, M. Re in a-Guerra and R. C. Laben. 1969.Neonatal Alterations in Serum Gamma Globulin Levels of Jersey and Holstein-Friesian Calves. Am. J. Vet. Res. Vol. 30, No.3; 345.
Transfer of Immunity from Colostrum to the Newborn Calf. Nutrition Review, Vol. 7, No. 3., 73.
Udy, D. A. 1956. A Rapid Method for Estimating Total Protein in Milk. Nature 178; 314.
Watts, R. E. 1971. Calf Mortality Reduction through Blood Transfusions. Feedstuffs, Vol. 43, No. 45; 28.
White, W. , D. H. Yungblut, J. L. Albright, B. W. Crowl and F. J.Babel. 1974. Composition and Nutritive Value of Fermented Colostrum for Feeding Dairy Calves. J. of Dairy Sci., Vol.57, No. 5; 643.






























