Embed Size (px)
Citation preview

This article was downloaded by: [University of Wisconsin-Milwaukee]On: 11 October 2014, At: 02:06Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
New Zealand Journal of BotanyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tnzb20
Patterns of species composition, recruitment, andgrowth within canopy gaps in two New Zealand kauri(Agathis australis) forestsN. J. Enright a , R. M. Bartlett b & C. R. De Freitas ba Department of Geography , University of Melbourne , Parkville, Australia , 3052b Department of Geography , University of Auckland , Private Bag 92 019, Auckland, NewZealandPublished online: 19 Dec 2011.
To cite this article: N. J. Enright , R. M. Bartlett & C. R. De Freitas (1993) Patterns of species composition, recruitment,and growth within canopy gaps in two New Zealand kauri (Agathis australis) forests, New Zealand Journal of Botany, 31:4,361-373, DOI: 10.1080/0028825X.1993.10419514
To link to this article: http://dx.doi.org/10.1080/0028825X.1993.10419514
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

New Zealand Journal oj Botany, 1993, Vol. 31: 361-373 0028-825X/93/3104-0361 $2.50/0 © The Royal Society of New Zealand 1993
361
Patterns of species composition, recruitment, and growth within canopy gaps in two New Zealand kauri (Agathis australis) forests
N. 1. ENRIGHT
Department of Geography University of Melbourne Parkville, Australia 3052
R. M. BARTLETT
C. R DE FREITAS
Department of Geography University of Auckland Private Bag 92 019 Auckland, New Zealand
Abstract Within-gap patterns of forest species composition, tree growth, and associated microclimate were investigated in two kauri (Agathis australis) forests in northern New Zealand. Detrended correspondence analysis (DCA) of sites at Trounson Kauri Park revealed a gradient of changing species composition from gap centre to forest understorey. However, while some species were typical either of the understorey (e.g., Dysoxlylum spectabile) or the centre of large gaps (e.g., Knightia excelsa, LeucopogonJasciculata), no species was restricted solely to gap edge sites.
Growth of potted tree seedlings over 6 months at different locations in and near a large canopy gap revealed a significant positive relationship between percent canopy openness (%CO) and height increase for each of the five species tested. The early successional tree Kunzea ericoides showed the greatest height increase at gap centre, while the longlived conifer Dacrydium cupressinum had the greatest height increase in the forest understorey.
DCA of within-gap vegetation at Huapai Reserve over the first 5 years following gap formation revealed several trends: (I) sample plots in the root zone on the southern side of the gap were most
B92038 Received 20 August 1992; accepted 25 May 1993
similar to gap centre plots, while those on the northern side were most similar to the forest understorey; (2) species composition (DCA loadings on the first axis) was significantly correlated with %CO; (3) at 9 of 10 sample locations within the gap, resprouts and individuals from the seedling bank were more likely to capture the available canopy space than seedlings established after gap creation.
Keywords canopy gap; canopy openness; microclimate; Agathis australis; tree species diversity
INTRODUCTION
Canopy gaps created by tree-fall are important sites for the regeneration of shade-intolerant tree species in both tropical and temperate forests (e.g., Runkle 1981, 1982, 1989; Hibbs 1982; Brokaw 1982; Putz 1983). Denslow (1980,1987) and Whitmore (1982) note that seedlings of most shade-tolerant species also benefit from the higher light intensities provided by such tree-falls. Many workers have contended that the turnover of sites at the forest floor due to tree-fall provides a major mechanism for maintaining woody species diversity and a relatively stable forest composition (Hartshorn 1978; Oldeman 1978; Whitmore 1978; Connell 1978; Denslow 1980). They argue that tree mortality results in the regular provision of canopy gaps as sites for tree species recruitment, and that gap size influences the microclimate relevant to seedling recruitment and growth. Variation between species in requirements for light and nutrients, and tolerance of heat and moisture stress (Denslow et aL 1990), might thus select which species is most likely to "capture" a given gap.
The hypothesised link between canopy gap dynamics and maintenance of tree species diversity has led Lo a concentration of research on speciesrich tropical forests (e.g., Pickett 1983; Barton 1984; Brokaw & Scheiner 1989; Denslow 1990; Brown & Whitmore 1992). However, canopy gap dynamics are important in temperate forests also and may play
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

362
a similar role within communities less diverse than tropical rainforest (Barden 1979; Runkle 1981, 1982, 1989; Hibbs 1982; Whitmore 1982; Ogden et al. 1987).
Position within gaps may also be important in selecting between an array of tree species available to colonise a site, since microclimate will vary between centre and edge (Ricklefs 1977). Orians (1982) further proposed that differences in the chemical environments of the root, bole, and canopy zones of individual gaps may influence species recruitment and survival patterns within gaps in tropical rainforests. Few studies have examined the potential role of these within-gap variations in the partitioning of establishment sites between species. While Brandani et al. (1988) and Nunez-Farfan & Dirzo (1988) found differences in species recruitment between the root and crown zones of gaps in Costa Rican and Mexican tropical forests, respectively, Popma et al. (1988) found no consistent differences between centre and edge. However, they noted that the area invaded by gap species included the adjacent forest understorey, and concluded that Brokaw's (1982) commonly used measure of gap size underestimated the affected area.
Temperate rainforests in New Zealand may contain up to 30 tree speciesiha, many of which are shade-intolerant and require canopy disturbance for successful regeneration (Ogden 1985). The present paper reports on three different approaches to the study of forest canopy gap dynamics and its role in the maintenance of forest composition and tree species diversity in New Zealand kauri (Agathis australis) forest: (1) ordinations of plant species composition within gaps, (2) field pot trials of tree seedling growth at different locations in and near a large tree-fall gap, and (3) the monitoring of species recruitment and recovery within a newly formed gap.
We sought to answer the following questions: (1) Does species composition within canopy
gaps differ such that gap edges can be differentiated both from the gap centre and forest understorey?
(2) Do the growth responses of tree species seedlings, ranging from those typical offorest understorey to those of large gaps and disturbed areas, differ when grown in the understorey and different parts of a gap?
(3) Do the patterns of plant species recruitment and growth correlate with environmental variations within gaps?
New Zealand Journal of Botany, 1993, Vol. 31
METHODS
The study sites
The study was carried out in two areas of temperate rainforest dominated by the emergent gymnosperm, Agathis australis* (D. Don) Lindl. (kauri): (1) the University of Auckland Huapai Reserve, a 15 ha forest remnant located in the Waitakere Ranges (36°47'S, 174°29'E) c. 30 km north-west of Auckland City; and (2) Trounson Kauri Park, a 273 ha forest reserve near Donnellys Crossing, Northland (35°44'S, 173°39'E), New Zealand. These forests each contain up to 6 gymnosperm and 25 angiosperm tree species. Canopy height is generally c. 20 m, with kauri emergent to 40 m. Both sites are characterised by the presence of large kauri (>2 m dbh), which are probably in excess of 600 years old (Ogden 1983), and a number of these have fallen within the last 30 years.
The Huapai site receives an average annual rainfall of 1328 mm with a winter maximum. Temperatures are mild all year round but are highest in February (mean daily temperature 18.9°C) and lowest in July (10A°C). Soils are brown granular clays (Parau clay) derived from andesitic tuffs (Thomas & Ogden 1983). Trounson Kauri Park is slightly wetter and more cloudy than Huapai, but the seasonal patterns of rainfall and temperature, and soil properties, are similar.
Ordination of canopy gap vegetation at Trounson Kauri Park
The forest is characterised by a continuous angiosperm canopy dominated by Beilschmiedia tarairi (Lauraceae), and large, emergent kauri. Five canopy gaps of different size (measured as percent canopy openness) and age, resulting from kauri treefall, were used for the ordination study (Table 1). Vegetation data were collected along 2 m wide transects running north-south and east-west through the gap centre and into the forest understorey. Density of all plant species >20 cm height was recorded for contiguous 2 x 2 m quadrats along these transects.
Quadrat data were combined to produce sites of equal size (20 m2) representing the vegetation of gap centres, gap edges, forest edges, and forest under-
*Nomenclature for New Zealand plants follows Allan (1961), Brownsey et al. (1985), and Connor & Edgar (1987).
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

Enright et al.-Canopy gaps in kauri forest
o 10 +~ F
,-----=------,' F
C Canopy gap centre
~ Gap edge • Fores! edge
F Forest understorey
Fig. 1 Arrangement of contiguous plots in and near canopy gaps at Trounson Kauri Park, and method of combining data into sampling units of approximately equal size that represent different parts of the canopy gap.
storey near gaps (Fig. 1). Stem density per site for woody species ranged from 50 to 120. One gap (Small gap, Table 1) was too small to contain all four site types and the gap edge category was not used. Forest understorey data for three additional sites> 10 m away from the nearest gap edge were also collected. Data for the 22 "sites" created in this way were ordinated using detrended correspondence analysis (DCA) (Hill & Gauch 1980) in order to reveal any underlying association between species composition and microsite location in and near canopy gaps. Species that occurred in only one site were excluded from the DCA, which was run using the default settings, including down weighting for rare species.
Field pot trials
Five tree species that occurred naturally in the study areas were selected for use in seedling growth
Table 1 Identity, approximate age, and percentage canopy openness for the five canopy gaps at Trounson Kauri Park, Northland, used in the DCA ordination.
Site identity Code'
Large gap L Small gap S Wardle gap W Cabbage tree gap C Tree stump gap T
Canopy Age (yr) openness (%)
22 36.9 22 5.5
6 16.1 >10 11.1 >10 5.7
*Codes used to identify sites on ordination diagrams.
363
experiments at Trounson Kauri Park: (l) Beilschmiedia tarairi (Lauraceae), the dominant angiosperm canopy tree; (2) Kunzea ericoides (Myrtaceae), a small, secondary tree; (3) Agathis australis (Araucariaceae) (kauri); (4) Dacrydium cupressinum (Podocarpaceae), a large and long-lived emergent gymnosperm; and (5) Dacrycarpus dacrydioides (Podocarpaceae), an emergent gymnosperm. These species were chosen partly because of the availability of seedlings, but also because they characterise different stages of succession in kauri forest. Ogden (1983) describes K. ericoides as a light-demanding, early secondary species (longevity c. 150 yr) which often enters sites shortly after disturbance. Kauri and D. dacrydioides commonly show massive regeneration beneath the open canopy of kanuka, while D. cupressinum (longevities for all three gymnosperms >600 yr) and B. tarairi (longevity c. 300 yr) are hypothesised to be more shade-tolerant and usually appear later in the succession.
Six-month-old seedlings of these species were obtained from a commercial nursery. Individual plants had been growing in potting mix in 15 cm plastic tubes for several months before the start of the experiment and remained in these tubes throughout. Ten seedlings of each species were randomly assigned to positions within blocks of 50 plants at each of nine locations: (1) the centre of Large gap (Table 1); (2-5) the northern, southern, eastern, and western gap edges; (6-7) the northern and western forest edges; (8) the undisturbed understorey nearby; and (9) the centre of Small gap. Seedlings were grown for 6 months from midsummer to mid-winter. Individual plant heights were recorded at the start of the experiment, and survival and height increase (including relative growth rate for height in cm/cm over the 6 months) were calculated at the end. Wet weight of plastic tubes containing potting mix only was determined monthly at each location (using a portable electronic balance) to check on differences in soil moisture level.
Species recruitment and recovery within a large gap at Huapai
A large gap (LG) developed at the Huapai Reserve in September 1985 by the fall of a kauri tree c. 2 m in diameter and 35 m in height. An almost circular clearing occurred where the tree crown landed, while a narrow "slot" and small opening appeared above the stem and root zone, respectively, producing a wine-glass-shaped gap, or "chablis" (sensu Oldeman
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

364
F1 •
F2 •
t N
Fig.2 Plan drawing of the canopy gap at Huapai Forest reserve showing the IO vegetation sample sites within the gap, the two forest understorey sites, and the five photo locations (letters) described in the text.
1978), overall (Fig. 2). The tree fell in a northerly direction down an 80 slope.
Ten vegetation reference points were located within the newly formed gap in October 1985 (Fig. 2). Species composition, density, and height were recorded (of all individuals >20 cm height) within a 1.25 m radius (c. 5 m2) of each reference point in 1986, 1988, and 1990. Forest understorey species composition was recorded at two locations 10-15 m from the eastern edge of the gap in 1988. Data from the vegetation reference points for each of the four measurement times were analysed using DCA.
Gap size and canopy openness Hemispherical photographs (177 0 field of view) of all gaps at Trounson Kauri Park were taken with a fish-eye lens positioned 1.5 m above ground level at the gap centre. Photographs were converted to computer images using a scanning digitiser, and contrast between foliage and clear sky increased by modifying the computer image subjectively. The percentage weighted canopy openness (percentage
New Zealand Journal of Botany, 1993, Vol. 31
of open sky visible from the camera position weighted by the relative contribution to total illumination of each open point) was determined using the SOLARCALC computer program of Chazdon & Field (1987). They contend that this is an estimate of the diffuse site factor. Canopy openness thus describes each site using a variable that is likely to be strongly correlated with both the light environment and other aspects of local microclimate. This measure also provides the most appropriate index of gap size (Brown & Whitmore 1992) since it standardises for surrounding vegetation height, so that a small gap in low forest may have a canopy openness equivalent to a large gap in tall forest. This overcomes some of the problems of between-gap comparison noted by Popma et al. (1988) and links the measure of gap size more directly to the environmental differences hypothesised to control species partitioning.
Hemispherical photographs of the forest canopy were taken directly above each of the seedling plots used in the pot growth experiment, and %CO determined.
Dimensions of the canopy gap at Huapai were mapped in the field by measuring the long axis of the gap and its width at six equidistant points along this axis. The position of gap edges was determined using the vertical projection definition of Brokaw (1982). Gap area was calculated both by digitising the survey diagram and by fitting an ellipse to the gap dimensions (Runkle 1981). Size of the canopy gap was remeasured in 1990 to determine the degree of canopy closure caused by lateral growth of individuals along the gap edge. Fish-eye lens photographs of the canopy/sky hemisphere were taken at a height of 1.5 m for the gap centre, and at points defined to characterise the north, south, east, and west of the gap (each 2 m from the gap edge). Photographs were also taken for the two understorey locations. The %CO was calculated for each position using SOLARCALC (Chazdon & Field 1987) as described above. Although SOLARCALC also allows calculation of predicted photosynthetically active photon flux density (PFD), this measure was discarded since a computation error has been discovered in the program (T. C. Whitmore pers. comm.).
Microclimate at Huapai Temporary microclimate recording stations were established at the centre of the large tree-fall gap and at one understorey site. At each site a Stephenson Screen at 1.5 m height housed a hygrothermograph
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014
Enright et al.-Canopy gaps in kauri forest
and maximum/minimum thermometer. Vapour pressure deficit (VPD) was calculated from the temperature and humidity records. Wooden stands were established to a height of 2.5 m for spot measurement of other variables at sites in the northern, eastern, southern, and western parts of the gap used to estimate %CO.
Net radiation (Q*), relative humidity, temperature, and vertical temperature gradients (VTG) were recorded over each of four clear sky days close to summer solstice in 1985/86. Microclimatic gradients across the gap should be steepest at this time of year and under these conditions (Poulson & Platt 1989). Net radiation was measured at 1.5 m height at 2 hourly intervals using Fritschen-type net radiometers. Output was read in mV using a digital voltmeter and values converted to W/m2. Net radiation was recorded for each of the 4 days in the gap (LG) sites, but for 2 days only in the two forest understorey sites due to instrument limitations. Mean daytime net radiation centred around solar noon (0700-1700 h) was estimated from these readings.
Maximum/minimum thermometers were placed at 1.5 m height for all positions within the gap, and at the two understorey sites. Readings were taken at 1500 h each day. Air temperature at 0.2 m and 2.5 m height was measured using a Gemwave electric psychrometer, and VTG estimated as the difference between readings at these two heights. These direct measures of canopy gap microclimate were correlated with %CO estimated using SOLARCALC, and site loadings for species composition from DCA. The non-parametric Spearman's rank correlation (r,) was used in all cases since sample sizes were generally too small to adequately test for normality of data.
RESULTS
Trounson Park
Ordination of canopy gap vegetation
The five canopy gaps sampled ranged in age from 6 years to about 30 years since their formation (Park Ranger pers. comm.). In each case the gap resulted from the death and known time of tree-fall of a large kauri. Canopy openness varied markedly, ranging from 6% to 37% in the gaps down to a very low 0.4% in the forest understorey (Table 1).
A total of 35 species was used in the DCA of the 22 sample sites, which described the species composition (density) of vegetation in relation to position in and near canopy gaps. There were 19
365
250
200
150
Ul (') ·x F(') «
100 (')
c 50
0 0 50 100 150 200 250
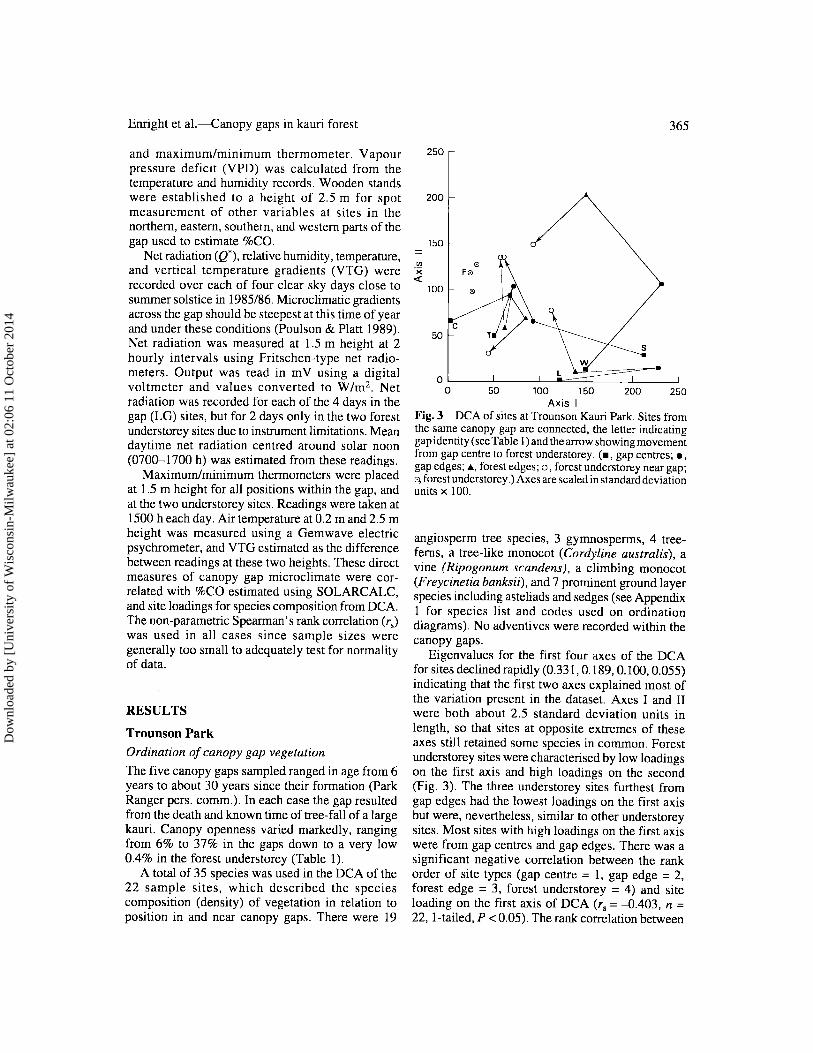
Axis I Fig. 3 DCA of sites at Trounson Kauri Park. Sites from the same canopy gap are connected, the letter indicating gap identi ty (see Table I ) and the arrow showing movement from gap centre to forest understorey. (., gap centres;., gap edges;.a., forest edges; 0, forest understorey near gap; C'l forest understorey.) Axes are scaled in standard deviation units x 100.
angiosperm tree species, 3 gymnosperms, 4 treeferns, a tree-like monocot (Cordyline australis), a vine (Ripogonum scandens), a climbing monocot (Freycinetia banksii), and 7 prominent ground layer species including asteliads and sedges (see Appendix 1 for species list and codes used on ordination diagrams). No adventives were recorded within the canopy gaps.
Eigenvalues for the first four axes of the DCA for sites declined rapidly (0.331, 0.189, 0.100, 0.055) indicating that the first two axes explained most of the variation present in the dataset. Axes I and II were both about 2.5 standard deviation units in length, so that sites at opposite extremes of these axes still retained some species in common. Forest understorey sites were characterised by low loadings on the first axis and high loadings on the second (Fig. 3). The three understorey sites furthest from gap edges had the lowest loadings on the first axis but were, nevertheless, similar to other understorey sites. Most sites with high loadings on the first axis were from gap centres and gap edges. There was a significant negative correlation between the rank order of site types (gap centre = I, gap edge = 2, forest edge = 3, forest understorey = 4) and site loading on the first axis of DCA (r, = -0.403, n = 22, I-tailed, P < 0.05). The rank correlation between
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014
366
350
240
130
'" X «
20
-90
NEPcor
DYSspe.
BEllaw.
BEl tar_
ACKros
D\Csqu II NESlan
PHOten II ALSmac
II CVAde8
CORaus II IAGAaus GHAxan
CYAsmi II II HEDarb
II RIPsca
FREban. D II
ASTtrl
II MYRaus
• PSEcra
CYAmed II
MELram l
PODtot.
HYPten
PRWer II
CQPaus
GENrup
~RAPyr • WElrac
II LEUfas
II IXEbre
BLEcap II II KNlexc
-200 '--__ ---'-___ -'--__ ---.J ___ --'--__ -----J
-200 -90 20 130 240 350 Axis I
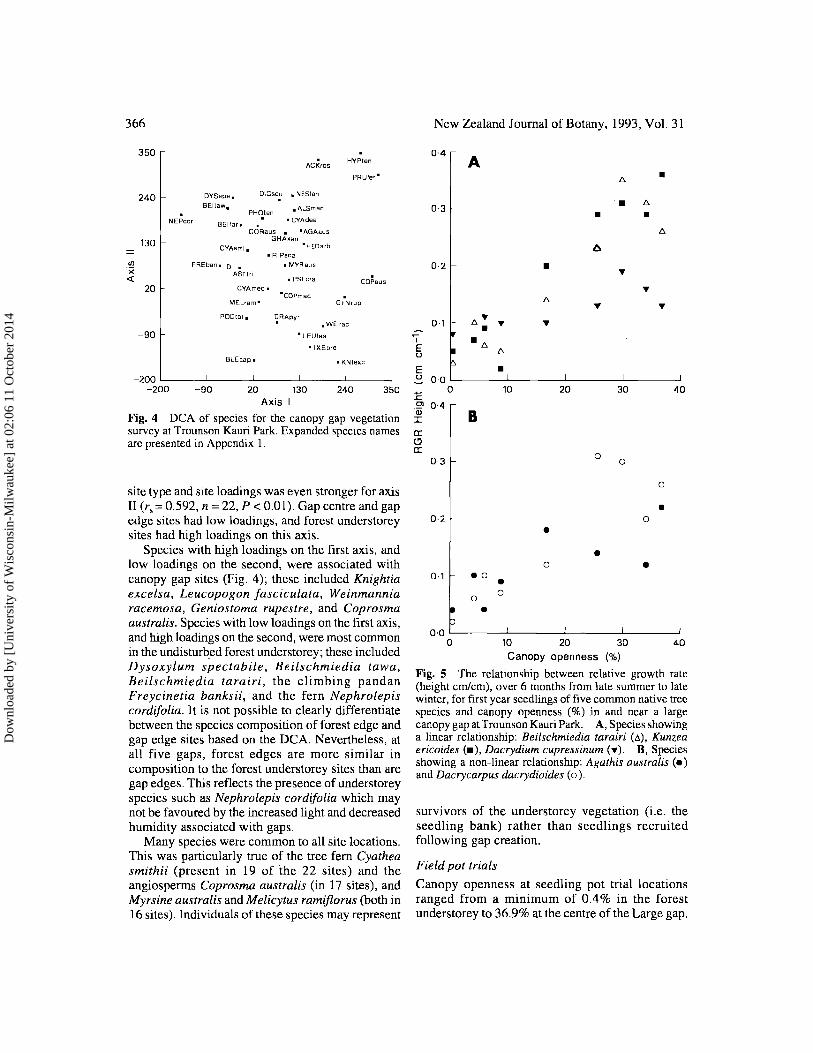
Fig. 4 DCA of species for the canopy gap vegetation survey at Trounson Kauri Park. Expanded species names are presented in Appendix 1.
site type and site loadings was even stronger for axis II (rs = 0.592, n = 22, P < 0.01). Gap centre and gap edge sites had low loadings, and forest understorey sites had high loadings on this axis.
Species with high loadings on the first axis, and low loadings on the second, were associated with canopy gap sites (Fig. 4); these included Knightia excelsa, Leucopogon Jasciculata, Weinmannia racemosa, Geniostoma rupestre, and Coprosma australis. Species with low loadings on the first axis, and high loadings on the second, were most common in the undisturb.ed forest understorey; these included Dysoxylum spectabile, Beilschmiedia tawa, Beilschmiedia tarairi, the climbing pandan Freycinetia banksii, and the fern Nephrolepis cordi/olia. It is not possible to clearly differentiate between the species composition of forest edge and gap edge sites based on the DCA. Nevertheless, at all five gaps, forest edges are more similar in composition to the forest understorey sites than are gap edges. This reflects the presence of understorey species such as Nephrolepis cordi/olia which may not be favoured by the increased light and decreased humidity associated with gaps.
Many species were common to all site locations. This was particularly true of the tree fern Cyathea smithii (present in 19 of the 22 sites) and the angiosperms Coprosma australis (in 17 sites), and Myrsine australis and Melicytus ramiflorus (both in 16 sites). Individuals of these species may represent
New Zealand Journal of Botany, 1993, Vol. 31
0·4 A
" •
• " • • 0·3
" t::.
0·2 • ... ...
" ... ... ... 0·1 ,,'" ... •
'I ." E u " E • u 0·0
1: 0 10 20 30 40
.Ql 0-4 Q) B I
a: (!l a:
0·3 o o
o
• 0·2 o
• •
0 • 0·1 .0 •
0 0
10 20 30 40
Canopy openness (%)
Fig. 5 The relationship between relative growth rate (height cm/cm), over 6 months from late summer to late winter, for first year seedlings of five common native tree species and canopy openness (%) in and near a large canopy gap at Trounson Kauri Park. A, Species showing a linear relationship: Beilschmiedia tarairi C~), Kunzea ericoides C-), Dacrydium cupressinum C ... )· B, Species showing a non-linear relationship: Agathis australis C.) and Dacrycarpus dacrydioides Co).
survivors of the understorey vegetation (i.e. the seedling bank) rather than seedlings recruited following gap creation.
Field pot trials
Canopy openness at seedling pot trial locations ranged from a minimum of 0.4% in the forest understorey to 36.9% at the centre of the Large gap.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014
Enright et al.-Canopy gaps in kauri forest
All five species used in the pot trials showed increases in both total height, and RGR for height, as %CO increased. The early secondary tree Kunzea ericoides showed by far the greatest increase in height over the 6 months of field study, growing by an average (±SE) of 22.4 ± 2.9 cm in the centre of the Large gap. Dacrydium cupressinum showed the greatest height increase in the forest understorey (3.1 ± 0.8 cm). Wet weight of tubes containing potting mix only did not vary significantly either between months or between locations, indicating little variation in soil moisture during the course of the experiment.
RGR for height (cm/cm) over the 6 month period was positively correlated with %CO for all species. However, while linear models best described the relationship for K. ericoides (rs = 0.87, P < 0.001), D. cupressinum (rs = 0.84) and B. tarairi Crs = 0.87) (Fig. 5A), non-linear models were best for kauri (log RGR = log 0.338 SYF - 3.055, F = 10.59, P < 0.02) and D. dacrydioides (log RGR = log 0.595 SYF -3.519, F = 66.30, P < 0.001), which both showed a levelling-off in growth response for very high %CO (Fig. 5B).
Where CO exceeded about 5%, K. ericoides maintained a faster absolute growth rate than the other species. This encompassed all positions within the large canopy gap, where the lowest measured CO was 16.7%. K. ericoides did not show the same response when RGR for height was compared among species. D. cupressinum showed the best RGR for sites with low CO (<8%) (e.g., in the forest understorey and forest edge sites). As CO increased beyond about 10%, K. ericoides displayed an increasing rate of height growth relative to both D. cupressinum and kauri, but not D. dacrydioides and B. tarairi.
Huapai Reserve
Seedling recruitment and growth in a large gap
The large canopy gap (LG) at Huapai had an estimated area of 348 m2 (digitised from field map) and ellipse area of 327 m 2. Ellipse area was calculated using a mean gap diameter rather than the maximum diameter proposed by Runkle (1981) since the gap was irregular in shape. CO was greatest for the gap centre (41 %) and about twice that for gap edge situations, excepting LG south (29%), which retained a relatively high CO owing to its position on the upslope side of the gap. This increased CO by reducing the obstruction effect of vegetation on the northern side of the gap. CO for
367
250
200
150
Ul x «
100
50
0 0 100 200 300 400
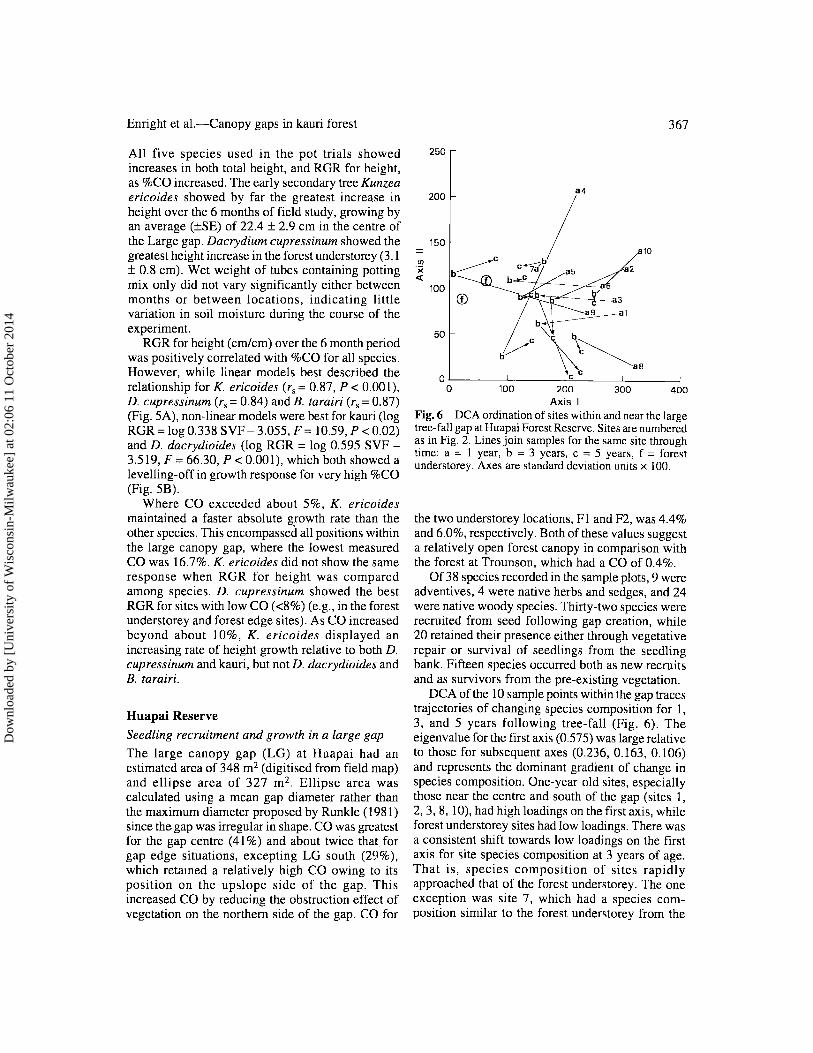
Axis I Fig.6 DCA ordination of sites within and near the large tree-fall gap at Huapai Forest Reserve. Sites are numbered as in Fig. 2. Lines join samples for the same site through time: a = 1 year, b = 3 years, c = 5 years, f = forest understorey. Axes are standard deviation units x 100.
the two understorey locations, F1 and F2, was 4.4% and 6.0%, respectively. Both of these values suggest a relatively open forest canopy in comparison with the forest at Trounson, which had a CO of 0.4%.
Of 38 species recorded in the sample plots, 9 were adventives, 4 were native herbs and sedges, and 24 were native woody species. Thirty-two species were recruited from seed following gap creation, while 20 retained their presence either through vegetative repair or survival of seedlings from the seedling bank. Fifteen species occurred both as new recruits and as survivors from the pre-existing vegetation.
DCA of the 10 sample points within the gap traces trajectories of changing species composition for 1, 3, and 5 years following tree-fall (Fig. 6). The eigenvalue for the first axis (0.575) was large relative to those for subsequent axes (0.236, 0.163, 0.106) and represents the dominant gradient of change in species composition. One-year-old sites, especially those near the centre and south of the gap (sites 1, 2,3, 8, 10), had high loadings on the first axis, while forest understorey sites had low loadings. There was a consistent shift towards low loadings on the first axis for site species composition at 3 years of age. That is, species composition of sites rapidly approached that of the forest understorey. The one exception was site 7, which had a species composition similar to the forest understorey from the
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014
368
start, and showed little change through time. This site was located at the north-western edge of the canopy gap.
Species compositional shifts along the second axis of DCA were less uniform. Several sites showed a trend towards lower loadings on this axis at 5 years of age (sites 1, 2, 3). These sites were all in the southern part of the canopy gap.
The species ordination (Fig. 7) showed adventives (e.g., Solanum mauritianum, Cirsium vulgare, and Phytolacca octandra) and a native herb (Solanum americanum) with high loadings on the first axis. Seedlings of these species dominated the early stages of regrowth, especially in the centre and south of the gap. Other species with high loadings were also characteristic of disturbed habitats but were present in low numbers only (e.g., Hypochoeris radicata, ]uncus effusus, Kunzea ericoides). Species with low loadings on the first axis were all native woody species characteristic of the surrounding forest vegetation (e.g., Vitex lucens, Melicytus ramiflorus, Rhopalostylis sapida, and Cyathea dealbata).
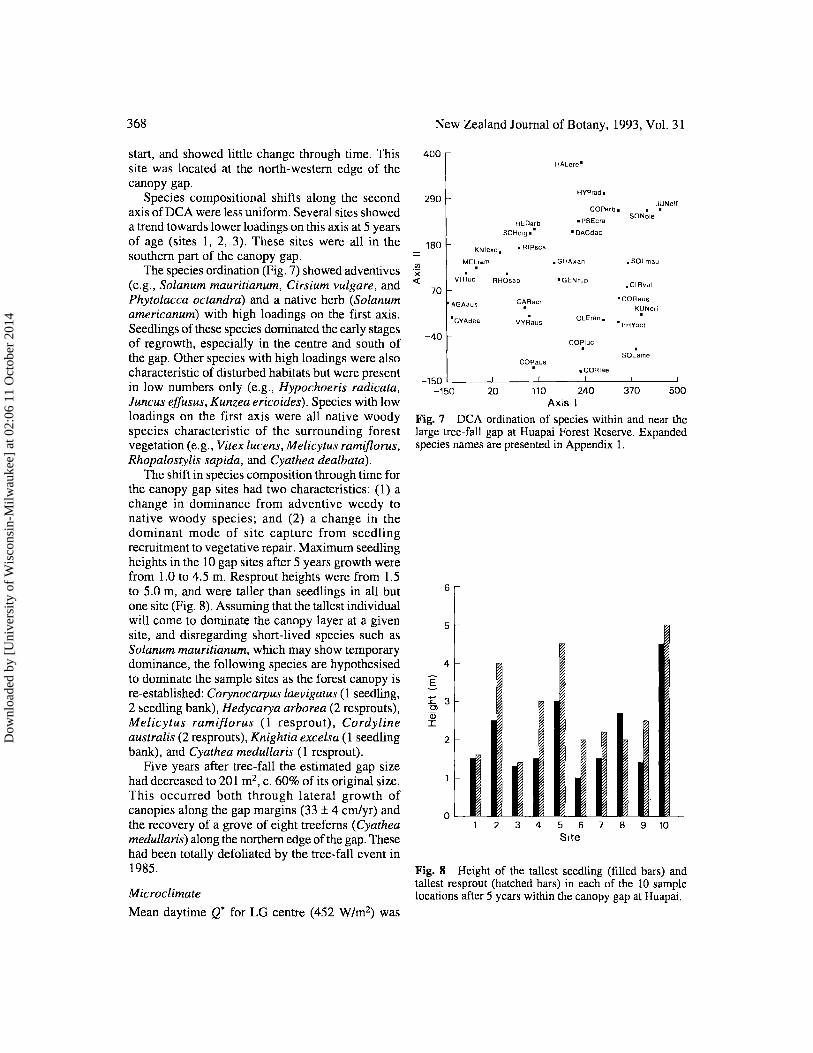
The shift in species composition through time for the canopy gap sites had two characteristics: (1) a change in dominance from adventive weedy to native woody species; and (2) a change in the dominant mode of site capture from seedling recruitment to vegetative repair. Maximum seedling heights in the 10 gap sites after 5 years growth were from 1.0 to 4.5 m. Resprout heights were from 1.5 to 5,0 m, and were taller than seedlings in all but one site (Fig. 8). Assuming that the tallest individual will come to dominate the canopy layer at a given site, and disregarding short-lived species such as Solanum mauritianum, which may show temporary dominance, the following species are hypothesised to dominate the sample sites as the forest canopy is re-established: Corynocarpus laevigatus (1 seedling, 2 seedling bank), Hedycarya arborea (2 resprouts), Melicytus ramiflorus (l resprout), Cordyline australis (2 resprouts), Knightia excelsa (1 seedling bank), and Cyathea medullaris (1 resprout).
Five years after tree-fall the estimated gap size had decreased to 201 m2, c. 60% of its original size. This occurred both through lateral growth of canopies along the gap margins (33 ± 4 cm/yr) and the recovery of a grove of eight treeferns (Cyathea medullaris) along the northern edge ofthe gap. These had been totally defoliated by the tree-fall event in 1985.
Microclimate
Mean daytime Q* for LG centre (452 W/m2) was
New Zealand Journal of B otan y, 1993, Vol. 31
400 HALere-
HYPrad. 290
JUNeff COParb.
SON~le HEDarb • PSEcra
SCHdlg. • DACdac
180 KNlexc. • RIPsca
.~ MELram • GHAxan • SOLmau
)(
<l:: VITluc RHOsap -GENrup
70 .CIRvul
CARser • CORaus AGAaus
KUNen
·CYAdea MYRaus OLEran.
• PHYoct
-40 CDPluc .
SOLame CDPaus . .CORlae
-150 -150 -20 110 240 370 500
Axis I
Fig. 7 DCA ordination of species within and near the large tree-fall gap at Huapai Forest Reserve. Expanded species names are presented in Appendix I.
6
5
4
E
- 3 ..c:: 01 ill I
2
o
r
I--
I--
- '-- I 2345678910
Site
Fig. 8 Height of the tallest seedling (filled bars) and tallest resprout (hatched bars) in each of the 10 sample locations after 5 years within the canopy gap at Huapai.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

Enright et al.-Canopy gaps in kauri forest
approximately six times that for Fl, while LG east and west received more Q' than LG north and south (Table 2). At these positions, Q* was intermediate between values for the understorey and SG centre. However, there was no significant correlation between Q* and %CO.
Daily maximum temperature was significantly corr~lated with both %CO and Q* (rs' P < 0.01). MaXImum temperatures were highest at LG centre (27.0°C) and lowest at FI (23.3°C). Temperatures on the periphery of the large gap were within the range 24.2 and 24.9°C with lowest values near the northern gap edge. Minimum temperature was lowest at LG centre but was not significantly correlated with any other variable measured.
VTG was highly variable between days; overall, it was highest at LG centre, intermediate around the periphery of the gap, and was lowest at Fl and F2. However, values were not significantly correlated with the other variables. LG centre showed maximum daily values for VPD nearly twice those for the forest understorey (paired t-test, P < 0.02).
CO was positively correlated with axis I loadings for species composition of sample plots at 1, 3, and 5 years (r" P < 0.01 in all cases). None ofthe other measured microclimate variables showed consistent correlation with species composition within the treefall gap.
Table 2 Percentage canopy openness and mean microclimatic information for four clear sky days near summer solstice 1985/86, within the large canopy gap and nearby forest understorey at Huapai.
Location co Q* VPD (%) (W/m2) (kPa)
LG centre 41 452 1.85 LG south 29 123 LG west 12 241 LG east 25 316 LG north 18 94 Forest 1 4 74 1.07 Forest 2 6 1.07
VTG Max Min (0C) (0C) (0C)
1.35 27.0 12.5 0.78 24.7 12.8 0.87 24.9 13.3 0.96 24.9 12.8 1.15 24.2 13.2 0.58 23.3 12.7 0.66 23.7 12.8
Q* is mean total daytime net radiation (0700-1700 h)' VPD is maximum recorded value over the 4 days of ~eas.urements; Max is maximum temperature; and Min IS minimum temperature recorded over the 4 days of measurements.
369
DISCUSSION
The res~lts present~d here show that within-gap patterns In plant speCIes composition are identifiable. The DCA ordination of gap and forest vegetation at Trounson Kauri Park showed gap edge and forest edge sites to lie between gap centre and forest understorey sites in species composition.
However, much overlap exists, and it is not possible to identify a unique set of species that characterise gap edge and forest edge locations. Rather, there is a continuum of change from gap centres to forest understoreys. A few species with intermediate loadings on the DCA axes may show opt~mum recruitment in the intermediate light enVIronments offered by gap edges, or the centres of small gap~ (e.g . .' Coprosma australis), but many are located In thIS part of the ordination space because they have a broad distribution from gap centre to forest understorey (i.e. they show no micro site preference).
Ogden et al. (1987) surveyed 20 canopy gaps created by tree-falls at Trounson Kauri Park and considered that nearly all (85-95%) of these gaps would eventually show dominance by one or other of the following species: Beilsehmiedia tarairi Weinmannia si/vieo/a, lxerba brexioides, Quintini~ serrata, or Prumnopitys ferruginea. The DCA described here identified Weinmannia silvieola Ixerba brexioides, and Prumnopitys ferruginea a~ characteristic of gap centres. Beilsehmiedia tarairi was more common in the forest understorey seedling bank but may require canopy gaps for successful recruitment to the canopy layer.
Popma et al. (1988) concluded that there was no difference in species composition between gap centre and gap edge sites in their study of a Mexican tropical forest. However, they enumerated only early colonists of newly formed gaps, and defined centre and edge in such a way that gap centres often encompassed the whole gap as well as parts of the adjacent forest understorey.
At Huapai we identified sites in the centre, north, south, east, and west of a large gap and in the nearby forest understorey. This arrangement of sites was based on the premise that, at temperate latitudes, the low solar angle (relative to tropical locations) would result in differences in growing conditions between northern and southern sides of the gap. Poulson & Platt.(l989~ speculated on the potential significance of thIS gradIent at temperate latitudes, while Collins & Pickett (1987) used a similar approach in a northern hardwood forest in Pennsylvania. They
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

370
concluded that their canopy gap may have been too small « 150 m2) to show pronounced environmental differences.
Species composition within the kauri tree-fall gap at Huapai was dominated in the first year by the adventives Solanum mauritianum, Phytolacca octandra, and Cirsium vulgare and the native herb Solanum americanum. Enright & Cameron (1988) recorded these species as dormant, viable seed in soils of the undisturbed forest in this reserve. Rates of seed input were insufficient to explain their presence through post-disturbance immigration (Enright & Cameron 1988; Sem 1991).
These weedy species occurred primarily at the centre, and in the south, of the gap. These locations had the highest %CO and so may have favoured both their germination and growth. In addition, the southern edge (site 8) was in the root zone where mineral soil was exposed. Sites located in the crown zone were covered by a thick layer ofleaf and branch debris. Where thick debris coincided with low %CO (i.e., towards the downslope, northern edge of the gap), no weedy species were recorded. None of these adventives was recorded in canopy gaps at Trounson Park, probably because of the advanced age of these gaps at the time of sampling.
By the time the gap at Huapai was 3 years old, species composition had shifted markedly towards that of the surrounding forest understorey. This was due both to the recruitment and growth of seedlings of native woody species, and to the recovery of damaged woody plants through basal res prouts and epicormic shoots. After 5 years it was apparent that gap closure would result mostly from lateral expansion of canopy trees around the gap edge, and the rapid vertical growth of plants damaged, but not killed, by the tree-fall. The estimated rate of lateral closure (33 cm/yr) is slightly higher than those obtained for some northern hardwood forests in North America, which range from 6 to 26 cm/yr (Hibbs 1982), and for montane forests of central North Island, New Zealand (Ogden et al. 1991), perhaps owing to a longer growing season.
Maximum rates of seedling height increase ranged from 0.2 to 0.9 m/yr. Runkle (1982) suggested a minimum rate of 0.5-1.0 m/yr was needed if seedlings were to keep pace with the rate of canopy closure for gaps in northern hardwood forests. Given the rapid rate of lateral growth, and competition from resprouters (which appears rare in other forests; e.g., Runkle 1982; Brandani et al. 1988; R. Dirzo pers. comm.), seedlings appear to have a low probability of being recruited to the
New Zealand Journal of Botany, 1993, Vol. 31
canopy in this gap. Only seedlings of the birddispersed species Corynocarpus laevigatus were growing quickly enough to compete with resprouting plants, and in some cases these may have been from the pre-gap seedling bank.
Field growth experiments provided a controlled approach to the study of within-gap variations in growing conditions. The five species used in the experiment at Trounson included species supposedly adapted to a wide range of light environments. Seedlings of all species showed increased height growth with the provision of more light than is available in the forest understorey. Denslow et al. (1990) report similar findings for seven tropical sh..'LJb species from the genera Miconia and Piper grown in gap centre, gap edge, and understorey locations. They found that RGRstems was positively correlated with available light but showed no significant response to nutrient treatments. Light appeared to be the most critical resource limiting growth.
The early secondary tree Kunzea ericoides showed by far the greatest rate of increase in height when located in gaps with CO >20%, and a marked decline in growth for locations where CO was <20%. The shade tolerant species Beilschmiedia tarairi also showed best growth in locations with high %CO, perhaps confirming a recruitment strategy of persistence in the seedling bank and release when canopy disturbance provides increased access to light. Dacrydium cupressinum showed the least response to increased %CO and the greatest growth, relative to other species, in the most heavily shaded locations. Morton et al. (1984) describe this species as "relatively shade tolerant".
Kauri (Agathis australis) and Dacrycarpus dacrydioides showed a non-linear relationship between height increase and %CO, rate of growth increasing initially as %CO increased, but then declining for the most open sites. Ogden et al. (1987), reviewing the findings of many earlier workers, noted that kauri can act as an early secondary species, colonising K. ericoides (or Leptospermum scoparium) stands after disturbance, and found that it could also regenerate in large treefall gaps. However, even in the latter case, individuals appeared to colonise only after a "nurse canopy" (Ogden et al. 1987) was established (i.e., not in the highest %CO situations). D. dacrydioides also appears dependent upon canopy disturbance for successful recruitment (Ogden 1985), but less is known of its specific requirements within the kauri forests described tere.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

Enright et al.-Canopy gaps in kauri forest
Studies of gap microclimate have generally contrasted conditions at the centre of gaps of different sizes with those in the forest understorey (e.g., Schulz 1960; Bazzaz & Pickett 1980; Denslow 1980; Chazdon & Fetcher 1984; Fetcher et al. 1985; Oberbauer et al. 1988). Photosynthetic light and maximum air temperature increases, and humidity decreases, as gap size increases (i.e., measures of gap microclimate approach those for open sites). Whitmore (1982) notes that light-intolerant species in tropical forests are commonly restricted to the centres of gaps >1000 m2.
The range of microclimatic environments within gaps is also likely to increase as gap size increases so that so-called small gap specialists (Denslow 1980) might occur around the edges of large gaps. Nunez-Farfan & Dirzo (1988) illustrated differences in species composition, diversity, and dominance by Cecropia obtusifolia Bertol. and Heliocarpus appendiculatus Turcz., in the root and canopy zones of two tree-fall gaps for a Mexican tropical forest. Cecropia was more abundant and showed better survival in the centre of a large crown gap, while Heliocarpus was favoured in a smaller opening near the roots of the fallen tree. They identified a number of possible explanations for this pattern, including differences in levels of both light and herbivory.
Results presented here for relationships between species composition, site locations, and microclimate variables must be regarded as hypothesis-generating only, given the small sample sizes. They reveal correlations between DCA loadings, %CO, and maximum temperature for sites in and near the large gap at Huapai. Other workers have illustrated a strong relationship between gap size, %CO, and photosynthetically active radiation (e.g., Denslow et al. 1990; Brown & Whitmore 1992).
Net radiation (Q*) was not correlated with either vegetation or %CO at the Huapai gap. This is hard to explain but may partly be attributed to small sample size and the shape and orientation of the gap. Despite a high %CO because of its upslope position, the southern part (root zone) of the gap was narrow and subject to shading by the surrounding canopy both early and late in the day. Nevertheless, in combination with the other measured (or estimated) variables, Q* provides a description of the total above-ground plant environment. While microclimate varies both within gaps, and between the understorey and gap, there is little evidence of finescale niche partitioning along this environmental gradient.
371
CONCLUSIONS
Ordination of gap vegetation at Trounson Park revealed a gradient of changing plant species composition from gap centre, through gap and forest edge, to the forest understorey. However, gap and forest edge locations were characterised more by the presence of widely distributed species than of those that might show optimum recruitment at these locations. There is little evidence of niche partitioning within gaps beyond that separating early secondary species of large gap centres from other fo~es.t species. Some variation in species composition withm the large canopy gap at Huapai was evident but is hypothesised to be short-lived. Woody and herbaceous. adventives established only at the gap centre and m exposed soil in the root zone (south) of the gap. They are short-lived, and native woody speCIes had overtopped them within the first 5 years of regrowth.
All five na.tive tree species used in the seedling growth expenment showed a positive relationship between RGRheight and %CO. The shorter lived angiosperm tree species showed a greater response to canopy opening than did the longer lived conifer species, despite major differences in their tolerance o~ shading. This might indicate that repeated canopy dIsturbance is more critical for successful recruitment than is some hypothetical specialisation on a partic~lar envelope of light regimes. The great longeVIty of the three conifer species ensures that t~ey will, on average, experience more canopy dIsturbances than their shorter lived neighbours. Denslow et al. (1990) and Brown & Whitmore (1992) have recently proposed this same idea.
ACKNOWLEDGMENTS
We thank Andrew Price and Brian Hurst for assistance in the field and with photography. This research was partIally funded by the University Grants Committee, Ne":,, Zealand. John Ogden, Rodolfo Dirzo, Tim WhItmore, and an anonymo.us referee provided useful comments on an earlIer verStOn of this paper.
REFERENCES
Allan, H. H. 1961: Flora of New Zealand. Vol. 1. Wellington, Government Printer.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

372
Barden, L. S. 1979: Tree replacement in small canopy gaps of a Tsuga canadensis forest in the southern Appalachians, Tennessee. Oecologia (Berlin) 44: 141-142.
Barton, A. M. 1984: Neotropical pioneer and shadetolerant tree species: do they partition treefall gaps? Tropical ecology 25: 196-202.
Bazzaz, F. A.; Pickett, S. T. A 1980: Physiological ecology of tropical succession: a comparative review. Annual review of ecology and systematics 11: 287-310.
Brandani, A.; Hartshorn, G. S.; Orians, G. H. 1988: Internal heterogeneity of gaps and tropical tree species richness. Journal of tropical ecology 4: 99-119.
Brown, N. D.; Whitmore, T. C. 1992: Do dipterocarp seedlings really partition tropical rain forest gaps? Philosophical transactions of the Royal Society of London B 335: 369-378.
Brokaw, N. V. L. 1982: The definition oftreefall gap and its effect on measures of forest dynamics. Biotropica 14: 158-160.
Brokaw, N. V. L.; Scheiner, S. M. 1989: Species composition in gaps and structure of a tropical forest. Ecology 70: 538-541.
Brownsey, P. J.; Given, D. R.; Lovis, J. D. 1985: A revised classification of New Zealand pteridophytes with a synonymic checklist of species. New Zealand journal of botany 23: 431-489.
Chazdon, R. L.; Fetcher, N. 1984: Photosynthetic light environments in a lowland tropical rainforest in Costa Rica. Journal of ecology 72: 553-564.
Chazdon, R. L.; Field, C. B. 1987: Photographic estimation of photosynthetically active radiation: evaluation of a computerized technique. Oecologia (Berlin) 73: 525-532.
Collins, B. S.; Pickett, S. T. A. 1987: Influence of canopy opening on the environment and herb layer in a northern hardwoods forest. Vegetatio 70: 3-10.
Connell, J. H. 1978: Diversity in tropical rainforests and coral reefs. Science 199: 1302-1310.
Connor, H. E.; Edgar, E. 1987: Name changes in the indigenous New Zealand flora, 1960-1986 and nomina nova IV, 1983-1986. New Zealand journal of botany 25: 115-170.
Denslow, J. S. 1980: Gap partitioning among tropical rainforest trees. Biotropica 12 (Supp!.): 47-55.
Denslow, J. S. 1987: Tropical rainforest gaps and tree species diversity. Annual review of ecology and systematics 18: 431-451.
Denslow, J. S.; Schulz, J. c.; Vitousek, P. M.; Strain, B. R. 1990: Growth responses of tropical shrubs to treefall gap environments. Ecology 71: 165-179.
New Zealand Journal of Botany, 1993, Vol. 31
Enright, N. J.; Cameron, E. C. 1988: The soil seed bank of a kauri (Agathis australis) forest remnant near Auckland, New Zealand. New Zealandjournal of botany 26: 223-236.
Fetcher, N.; Oberbauer, S. F.; Strain, B. R. 1985: Vegetation effects on microclimate in lowland tropical forest in Costa Rica.1nternationaljournal ofbiometeorology 29: 145-155.
Hartshorn, G. S. 1978: Tree falls and tropical forest dynamics. In: Tomlinson, P. B.; Zimmerman, M. H. ed. Tropical trees as living systems. New York, Cambridge University Press. pp. 617--638.
Hibbs, D. E. 1982: Gap dynamics in a hemlock-hardwood forest. Canadian journal of forest research 12: 522-527.
Hill, M. 0.; Gauch, H. G. Jun. 1980: Detrended correspondence analysis: an improved ordination technique. Vegetatio 42: 47-58.
Morton, J.; Ogden, J.; Hughes, T. 1984: To save a forestWhirinaki. Auckland, David Bateman Ltd.
Nunez-Farfan, J.; Dirzo, R. 1988: Within-gap spatial heterogeneity and seedling performance in a Mexican tropical forest. Oikos 51: 274-284.
Oberbauer, S. F.; Clark, D. B.; Clark, D. A.; QuesaJa, M. 1988: Crown light environments of saplings of two species of rainforest emergent trees. Oecologia (Berlin) 75: 207-212.
Ogden, J. 1983: The scientific reserves of Auckland University. II. Quantitative vegetation studies. Tane 29: 163-180.
Ogden, J. 1985: An introduction to plant demography with special reference to New Zealand trees. New Zealand journal of botany 23: 751-772.
Ogden, J.; Wardle, G. M.; Ahmed, M. 1987: Population dynamics of the emergent conifer Agathis australis (D. Don) Lind!. (kauri) in New Zealand. II. Seedling population size and gap-phase regeneration. New Zealand journal of botany 25: 231-242.
Ogden, ].; Fordham, R. A; Pilkington, S.; Serra, R. G. 1991: Forest gap formation and closure along an altitudinal gradient in Tongariro National Park, New Zealand. Journal of vegetation science 2: 165-172.
Oldeman, R. A. A 1978: Architecture and energy exchange of dicotyledonous trees in the forest. In: Tomlinson, P. B.; Zimmerman, M. H. ed. Tropical trees as living systems. New York, Cambridge University Press. Pp. 535-560.
Orians, G. H. 1982: The influence of tree-falls in tropical forests on tree species richness. Tropical ecology 23: 255-279.
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014

Enright et al.-Canopy gaps in kauri forest
Pickett, S. T. A. 1983: Differential adaptation of tropical tree species to canopy gaps and its role in community dynamics. Tropical ecology 24: 68-84.
Popma, J.; Bongers, F.; Martinez-Ramos, M.; Veneklaas, E. 1988: Pioneer species distribution in treefall gaps in Neotropical rain forest; a gap definition and its consequences. Journal of tropical ecology 4: 77-88.
Poulson, T. L.; Platt, W. J. 1989: Gap light regimes influence canopy tree diversity. Ecology 70: 553-555.
Putz, F. E. 1983: Treefall pits and mounds, and the importance of soil disturbance to pioneer trees on Barro Colorado Island, Panama. Ecology 64: 1069-1074.
Ricklefs, R. E. 1977: Environmental heterogeneity and plant species diversity: a hypothesis. American naturalist 11 i: 376--381.
Runkle, J. R. 1981: Gap regeneration in some old-growth forests of the eastern United States. Ecology 62: 1041-1051.
Runkle, J. R. 1982: Patterns of disturbance in some oldgrowth mesic forests of eastern North America. Ecology 63: 1533-1546.
Runkle, J. R. 1989: Synchrony of regeneration, gaps and latitudinal differences in tree species diversity. Ecology 70: 546-547.
Schulz, J. P. 1960: Ecological studies on rainforest in Northern Surinam. Proceedings of the koninklijke Nederlandse academie van wetenschappen 253: 1-267.
Sem, G. 1991: The soil seed bank in Agathis australis (D. Don) Lind!. (kauri) forests of northern New Zealand and its potential role in secondary successions. Unpublished Ph.D. thesis, lodged in the Library, University of Auckland, New Zealand.
Thomas, G. M.; Ogden, J. 1983: The scientific reserves of Auckland University. 1. General introduction to their history, vegetation, climate and soils. Tane 29: 143-162.
Whitmore, T. C. 1978: Gaps in the forest canopy. in: Tomlinson, P. B.; Zimmerman, M. H. ed. Tropical trees as living systems. New York, Cambridge University Press. Pp. 639-655.
Whitmore, T. C. 1982: On pattern and process in forests. In: Newman, E. 1. ed. The plant community as a working mechanism. Blackwell, Oxford. Pp. 45-59.
373
APPENDIX 1
Species list and codes used in DCA diagrams (Fig. 4 and 7). Species Code
Ackama rosllefolia ACKros Agathis australis AGAaus Alseuosmia macrophylla ALSmac Astelia trinervia ASTtri Beilschmiedia tarairi BEltar Beilschmiedia tawa BEltaw Blechnum capense BLEcap Carpodetus serratus CARser Cirsium vulgare CIRvul Coprosma arborea COParb Coprosma australis COPaus Coprosma lucida COPluc Coprosma macrocarpa COPmac Cordy line australis CORaus Corynocarpus laevigatus CORlae Cyathea dealbata CYAdea Cyathea medullaris CYAmed Cyathea smithii CYAsmi Dacrycarpus dacrydioides DACdac Dicksonia squarrosa DICsqu Dracophyllum pyramidale DRApyr Dysoxylum !,pectabile DYSspe Freycinetia banksii FREban Geniostoma rupestre GENrup Gahnia xanthophyllum GAHxan Hedycarya arborea HEDarb Hypochoeris radicata HYPrad Hypolepis tenuifolium HYPten lxerba brexioides IXEbre Juncus effusus lUNefu Knightia excelsa KNlexc Kunzea ericoides KUNeri Leucopogon fasciculata LEUfas Lobelia anceps LOBanc Melicytus ramiflorus MELram Myrsine australis MYRaus Nestegis lanceolata NESlan Nephrolepis cordifolia NEPcor Olea ria rani OLEran Phormium tenax PHOten Phytolacca octandra PHYoct Podocarpus tutara PODtot Prumnopitys ferruginea PRUfer Pseudopanax crassifolius PSEcra Rhopalostylis sapida RHOsap Ripogonum scandens RIPsca Schefflera digitata SCHdig Solanum americanum SOLame Solanum mauritianum SOLmau Sonchus uleracea SONole Vitex lucens VITluc Weinmannia racemosa WEIrac
Dow
nloa
ded
by [
Uni
vers
ity o
f W
isco
nsin
-Milw
auke
e] a
t 02:
06 1
1 O
ctob
er 2
014





























