Embed Size (px)
Citation preview

Vol. 56, No. 12
Pectin Lyase Activity in a Penicillium italicum StrainAITOR ALANA,t* ITZIAR ALKORTA, JUAN B. DOMINGUEZ,4 MARIA J. LLAMA, AND JUAN L. SERRA
Departamento de Bioquimica y Biologia Molecular, Facultad de Ciencias, Universidad del Pafs VascolEuskal HerrikoUnibertsitatea, Apartado 644, E48080 Bilbao, Spain
Received 11 July 1990/Accepted 3 October 1990
An extracellular pectin lyase (PNL) [poly-(methoxygalacturonide)lyase; EC 4.2.2.10] produced by Penicil-lium italicum CECT 2294 grown on a surface bran (natural medium) or in a submerged (synthetic medium)culture was investigated. Both culture filtrates showed macerating activity at low pH on cucumber, potato, andorange tissues. The physicochemical properties of the enzyme obtained from both culture methods were
identical, as well as its catalytic properties, which were assayed by different methods. The molecular mass ofthe PNL obtained by gel filtration chromatography was 22 kDa; the isoelectric point was 8.6, as determined bychromatofocusing; and the enzyme was able to catalyze the eliminative cleavage of pectins with low (37%) andhigh (from 54 to 82%) degrees of esterification. The PNL produced in liquid medium showed a K,m for pectin(degree of esterification, 70%) of 3.2 mg/ml, and the optimum pH was 6.0 to 7.0. This enzyme was stable at50°C and at pH 8.0. The ability of this PNL to macerate plant tissues in acidic environmental conditions, itsstability at low pH and temperatures up to 50°C (thus preventing mesophilic microbial growth), and theabsence of pectinesterase make this preparation useful for the food industry.
Many plant-pathogenic bacteria and fungi are responsiblefor soft rot diseases of plant products before harvesting andduring storage. The factors which produce such tissue mac-
eration are generally random chain-splitting pectic enzymes,i.e., pectate lyase (PL), pectin lyase (PNL), and polygalac-turonase (8, 9). PL and PNL split the a-1,4 glycosidic bondbetween galacturonic acid residues by trans-elimination,while polygalacturonases catalyze a hydrolytic cleavage. PLprefers galacturonic acid polymers of low C-6 methylation(pectate) as substrates, while PNL cleaves only polygalac-turonic acid polymers in which a high proportion of C-6carboxyl groups is methylated (pectin) (25).
Microbial enzymes catalyzing the degradation of pecticpolysaccharides are also involved in the decay of dead plantmaterial by nonpathogenic microorganisms, assisting in re-cycling carbon compounds in the biosphere. Moreover,these enzymes play an important role in food processing(32). The major industrial applications of these enzymesinclude the extraction and clarification of fruit juices andgrape musts (citrus fruit juice and wine technology), macer-ation of vegetables and fruit, and the production of cloudi-ness in soft drinks (13).The importance of using isolated PNL in fruit juice tech-
nology comes from the fact that this enzyme is the only one
currently known to be able to cleave, without the prioraction of other enzymes, the ot-1,4 glycosidic bond of highlyesterified pectins, decreasing the viscosity of and clarifyingjuices without damaging the volatile ester content responsi-ble for the specific aroma of various fruits. Several otheradvantages of PNL versus other pectolytic enzymes, notonly in fruit juice technology but also in fruit and vegetableliquefaction, have increased interest in studying this enzyme(13; A. Alafia, Ph.D. thesis, University of the BasqueCountry, Bilbao, Spain, 1990).
* Corresponding author.t Present address: Investigaci6n Alimentaria, S.A. (Grupo KAS),
Larragana 12, E-01013 Vitoria-Gasteiz, Spain.: Permanent address: Investigaci6n Alimentaria, S.A. (Grupo
KAS), Larragana 12, E-01013 Vitoria-Gasteiz, Spain.
As far as the postharvest spoilage of citrus products isconcerned, PNL is also of great importance. Among themicroorganisms responsible for this process, Penicilliumspp. (mainly Penicillium italicum and Penicillium digitatum)are the most important examples in terms of both quantityand quality (10). Several studies dealing with production andcharacterization of PNL have been done with Penicilliumspp. over the last twenty years (6, 7, 21, 28, 30, 31).Nevertheless, there is still a lack of information about PNLproperties and possible applications of Penicillium culturefiltrates. In a previous article (1), we described the produc-tion of PNL by P. italicum grown in submerged culture andthe presence of several pectolytic enzymes in the filtrates.To complete this study, this paper describes, in addition toPNL and PL, other nonpectolytic enzymes produced by P.italicum CECT 2294 (ATCC 66636) when grown on surfacebran or in submerged culture. The macerating activity shownby both culture filtrates and some physicochemical andcatalytic properties of the PNL produced in these twogrowth media are also discussed. This work may be ofinterest in the food industry because it describes a fungalPNL that is highly stable and capable of macerating planttissues under acidic conditions.
MATERIALS AND METHODSMaterials. Mono P and Superose 12 columns, Polybuffer
96, and fast-performance liquid chromatography (FPLC)equipment were obtained from Pharmacia (Uppsala, Swe-den). Commercial preparations of pectinases were suppliedby Grindsted Products A/S, Brabrand, Denmark (PektolasePA); Miles Kali-Chemie GmbH, Nienburg, Federal Republicof Germany (Opticlar-P); Novo Industri A/S, Laufen, Swit-zerland (Ultrazym 100 G); and Societd Rapidase, Sdclin,France (Rapidase CNP). DEAE-cellulose (DE-52) and glassfiber filters (GF/C) were from Whatman (Maidstone, UnitedKingdom). Azocoll, carboxymethyl cellulose, bovine serum
albumin, polygalacturonic acid, sodium polypectate, andcitrus pectin (grade I, with a degree of esterification [DE] of70%) were purchased from Sigma (St. Louis, Mo.). Applepectin was kindly supplied by OBI Pektin (Bischofszell,Switzerland). Gel filtration molecular weight markers were
3755
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 1990, p. 3755-37590099-2240/90/123755-05$02.00/0
on January 12, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
obtained from Boehringer Mannheim (Mannheim, FederalRepublic of Germany). The potato dextrose agar mediumwas from bioMerieux (Charbonnieres les bains, France).The protein assay dye reagent was from Bio-Rad (Munich,Federal Republic of Germany). All other chemicals werereagents of analytical grade supplied by Merck (Darmstadt,Federal Republic of Germany).Growth of microorganisms. P. italicum was obtained from
Coleccion Espafiola de Cultivos Tipo (CECT), Valencia,Spain, and maintained in potato dextrose agar medium.Surface bran cultures were grown in 2-liter Erlenmeyerflasks containing 100 g of wheat bran and 100 ml of water at28°C in a constant-temperature room. The wet bran-contain-ing flasks were sterilized for 15 min at 1.3 atm (1 atm =101.29 kPa) and inoculated with a suspension of germinatedspores (106/ml in 20% [vol/vol] glycerol) grown for 4 days inpotato dextrose agar. Filtrates were prepared after 72 h ofgrowth by adding distilled water to the bran (100 ml of waterto 10 g of dry bran) and allowing the suspension to standwhile being gently shaken. After 1 h, the liquid was de-canted, filtered through a nylon filter, and finally centrifugedat 10,000 x g and 4°C for 30 min.Submerged cultures were grown at 28°C with shaking (160
rpm) in 2-liter Erlenmeyer flasks containing 1 liter of CzapekDox medium (Oxoid Ltd., London, England) (containing, ingrams per liter, the following ingredients: NaNO3, 2; KCl,0.5; magnesium chloride, 0.2; FeSO4, 0.01; sucrose, 30; andK2S04, 0.35) supplemented with 1% (wt/vol) citrus pectin.The medium was sterilized for 15 min at 1.3 atm andinoculated with a suspension of germinated spores (104/mlin 20% [vol/vol] glycerol) grown for 4 days in potato dextroseagar. Filtrates from submerged cultures were prepared,after 48 h of growth, by filtering on glass fiber filters underreduced pressure.Enzyme and macerating assays. PNL (EC 4.2.2.10) activity
was routinely determined spectrophotometrically with aShimadzu UV-260 spectrophotometer by monitoring theincrease in A235 as described by Albersheim and Killias (3).The reaction mixture (1.25 ml) contained 0.25 M citrate-phosphate buffer (pH 6.0), 0.5% (wt/vol) citrus pectin (DE,70%; for experiments in which apple pectin was used, itsconcentration and DE are stated below), and the appropriatevolume of crude extract (final concentration, 1 ,ug of proteinper ml). Preincubations were carried out at 40°C for 15 min,and the reaction was started by adding pectin. In theseconditions, PNL activity varied linearly with respect to thecrude extract up to 10 ,ug of protein per ml. Control tubescontained the enzyme previously inactivated by incubationfor 10 min at 100°C. One unit of activity is the amount ofenzyme which produces an increase of 1 U of A235 per min.For PNL assays, stock solutions of 1.25% (wt/vol) pectin indistilled water were prepared and heated for 10 min at 100°C.These solutions were filtered through glass fiber and storedat 4°C for not longer than 1 week.PNL was also assayed by monitoring changes in substrate
viscosity. Reduction in viscosity was measured in a Cannon-Fenske viscometer (size 100) at 40°C with apple pectin(0.4%, wt/vol) in 0.2 M citrate-phosphate buffer (pH 6.0),with 80 mU of PNL per ml in the reaction mixture.PL (EC 4.2.2.2) activity was measured spectrophotomet-
rically as described for PNL. The reaction mixture (1.25 ml)contained 0.3 mM CaCl2 in 0.1 M Tris hydrochloride buffer(pH 8.5) containing 0.5% (wt/vol) polygalacturonic acid andthe appropriate volume of crude extract (final concentrationof 1 ,g of protein per ml). Preincubation and assay condi-
tions and the unit of activity were the same as thosedescribed for PNL.
Cellulase (EC 3.2.1.4) and a-amylase (EC 3.2.1.1) activi-ties were assayed by measuring the reducing groups releasedin the reaction mixture by the Nelson-Somogyi copperreduction method described by Spiro (29) using Somogyi'scopper reagent (27) and Nelson's arsenomolybdate reagent(20). The reaction mixtures contained 50 ,ul of 0.1 M acetatebuffer (pH 5.0), 50 ,ul of crude extract, and 50 ,ul of 1%(wt/vol) carboxymethyl cellulose or starch for cellulase ora-amylase activity, respectively. Preincubations were car-ried out at 37°C for 10 min. The reaction was started byadding the substrate, and incubation was carried out at 37°Cfor 10 min. One unit of cellulase or ac-amylase activity is theamount of enzyme which releases 1 ,umol equivalent ofglucose per min.
General proteinase activity was assayed with Azocoll asthe substrate, as described by Asahi et al. (4). One unit ofproteinase activity is the amount of enzyme that caused anincrease in A540 equal to 0.01 optical density units per min.
Macerating activity was determined by incubating thinwedges of plant tissue (0.5 g of fresh fruit) with 17 mU ofPNL per ml in 0.25 M citrate-phosphate buffer (pH 4.0 or6.0) at 25°C for 18 h. Loss of tissue cohesiveness was testedwith a spatula. For all enzyme assays, enzyme boiled for 10min at 100°C was used as a control.
Fractional precipitation of PNL. After the culture filtrateswere adjusted to pH 8.0, PNL was salted out with ammo-nium sulfate as previously described (1) and finally stored at4°C until use. These concentrated culture filtrates, consid-ered as crude extracts, were free of pectin.
Preparation of highly methoxylated pectin. Pectin wasesterified by the method described by Morell et al. (19).
Analytical methods. Molecular mass determination wascarried out by gel filtration in a FPLC-Superose 12 (HR10/30) column equilibrated with 100 mM KCl in 50 mM Trishydrochloride buffer (pH 8.0). Elution was done with thesame buffer at a flow rate of 0.5 ml/min. The column waspreviously calibrated with bovine serum albumin (66 kDa),ovalbumin (45 kDa), myoglobin (17 kDa), and cytochrome c(12.5 kDa). Isoelectric point studies were carried out byFPLC chromatofocusing in a Mono P (HR 5/5) column. Aflow rate of 1 ml/min and a pH gradient from 9 to 6 wereused. After equilibration of the column with 75 mM 1,3-diaminopropane-HCl buffer (pH 10.5), a crude PNL samplewas injected onto the column and the pH gradient wasformed by applying 30 ml of 1:10 diluted Polybuffer 96-HCl(pH 6.0). One-milliliter fractions were collected, and thePNL activity and pH were measured.
Protein was determined by the method of Bradford (5)using serum albumin as the standard. The DE and themethoxy content of pectins were determined as described bySchultz (26) and McCready (18). The galacturonic acidcontent was estimated as anhydrogalacturonic acid by thecarbazole colorimetric method (12) as modified by Dietz andRouse (11).
RESULTS
Enzyme and macerating activities shown by P. italicum in asubmerged (synthetic) and on a surface bran (natural) culture.Table 1 summarizes the presence of several enzyme activi-ties in P. italicum submerged (after 48 h) and surface bran(after 72 h) cultures. Although PNL activity is much higherwhen the fungus grows in a semisolid medium (620 mU/ml)than when it grows in liquid (in which it never rises above
3756 ALA&A ET AL.
on January 12, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
PNL ACTIVITY IN P. ITALICUM 3757
TABLE 1. Enzyme activities present in culturefiltrates of P. italicuma
Culture Enzyme activity (U/mg of protein)method PNL PL Proteinase Cellulase a-Amylase
Submerged 17.5 0.0 1.3 7.7 2.5Surface bran 5.8 0.2 0.2 0.9 6.7
a Enzyme activities were assayed as described in Materials and Methods.The fungus was grown in submerged or surface bran culture for 48 or 72 h,respectively.
143 mU/ml [1]), the specific activity is about threefold higherin the latter. PL activity was detected only when wheat branwas used as the carbon and energy source.
P. italicum displayed proteinase, cellulase, and a-amylaseactivities when grown not only in a natural medium but alsoin a synthetic medium.The macerating activities of both culture filtrates were
similar when assayed with several plant tissues (cucumber,potato, and orange) at pH 4.0 and 6.0. The activity at thelower pH value was slightly more effective, and both culturefiltrates showed the highest macerating activity on cucumberwedges. Although the effectiveness in cell separation waslower with potato and orange tissues as substrates, bothplant tissues were also affected by P. italicum filtrates.
Comparative study of the PNL produced in a submergedculture and on a surface bran culture. As previously stated,PNL production in a semisolid medium is more effectivethan in a liquid one, in spite of the higher specific activityvalues in the latter. To demonstrate the existence of only onePNL form in both media, a comparative study of submergedand surface bran culture filtrates was carried out. Prepara-tions of the enzyme obtained from both growth mediashowed identical behavior when they were chromatographedwith the ion exchangers DEAE-cellulose and carboxymethylcellulose at pH 8.0 and 6.0, respectively. Both preparationsalso yielded the same elution pH value (8.6) after chromato-focusing through an FPLC-Mono P column and the sameelution volume (15.1 ml) when the crude extracts (22 kDa)were applied onto an FPLC-gel filtration column (Superose12).When apple pectin was used as the substrate, the PNL
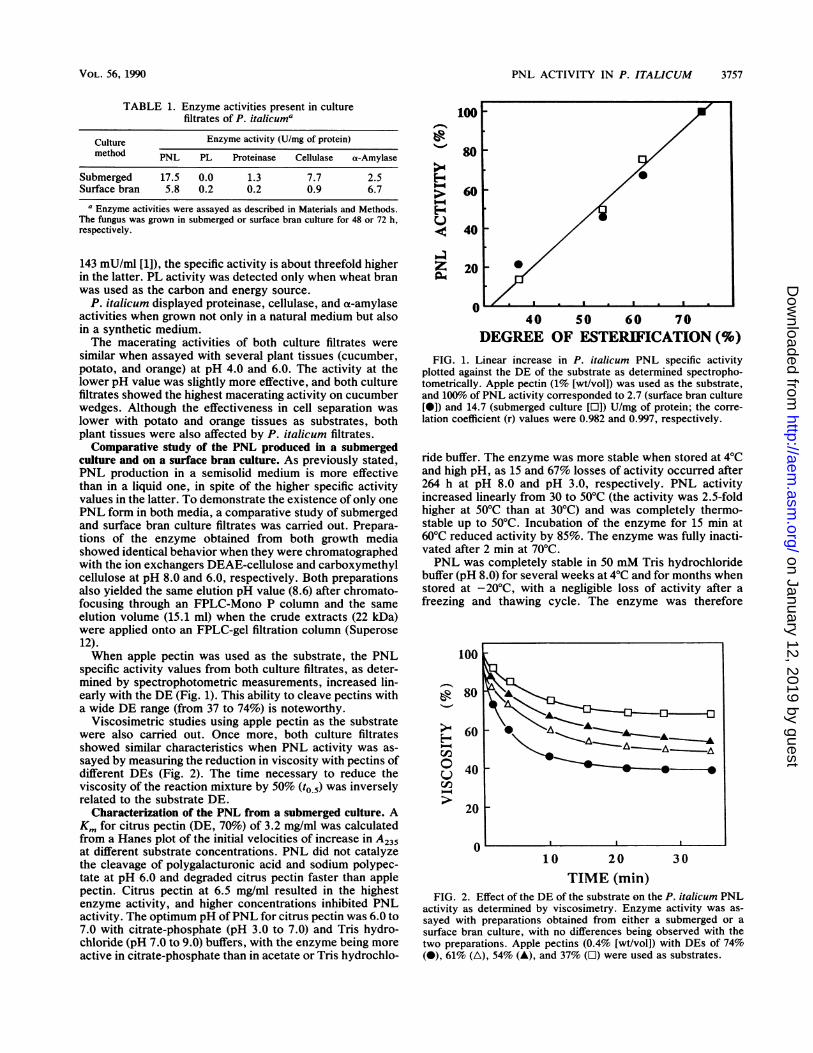
specific activity values from both culture filtrates, as deter-mined by spectrophotometric measurements, increased lin-early with the DE (Fig. 1). This ability to cleave pectins witha wide DE range (from 37 to 74%) is noteworthy.
Viscosimetric studies using apple pectin as the substratewere also carried out. Once more, both culture filtratesshowed similar characteristics when PNL activity was as-sayed by measuring the reduction in viscosity with pectins ofdifferent DEs (Fig. 2). The time necessary to reduce theviscosity of the reaction mixture by 50% (t05) was inverselyrelated to the substrate DE.
Characterization of the PNL from a submerged culture. AKm for citrus pectin (DE, 70%) of 3.2 mg/ml was calculatedfrom a Hanes plot of the initial velocities of increase in A235at different substrate concentrations. PNL did not catalyzethe cleavage of polygalacturonic acid and sodium polypec-tate at pH 6.0 and degraded citrus pectin faster than applepectin. Citrus pectin at 6.5 mg/ml resulted in the highestenzyme activity, and higher concentrations inhibited PNLactivity. The optimum pH ofPNL for citrus pectin was 6.0 to7.0 with citrate-phosphate (pH 3.0 to 7.0) and Tris hydro-chloride (pH 7.0 to 9.0) buffers, with the enzyme being moreactive in citrate-phosphate than in acetate or Tris hydrochlo-
100
80
>60
40 -
Z 20 -
040 50 60 70
DEGREE OF ESTERIFICATION (%)FIG. 1. Linear increase in P. italicum PNL specific activity
plotted against the DE of the substrate as determined spectropho-tometrically. Apple pectin (1% [wt/vol]) was used as the substrate,and 100% of PNL activity corresponded to 2.7 (surface bran culture[0]) and 14.7 (submerged culture [I]) U/mg of protein; the corre-lation coefficient (r) values were 0.982 and 0.997, respectively.
ride buffer. The enzyme was more stable when stored at 4°Cand high pH, as 15 and 67% losses of activity occurred after264 h at pH 8.0 and pH 3.0, respectively. PNL activityincreased linearly from 30 to 50°C (the activity was 2.5-foldhigher at 50°C than at 30°C) and was completely thermo-stable up to 50°C. Incubation of the enzyme for 15 min at60°C reduced activity by 85%. The enzyme was fully inacti-vated after 2 min at 70°C.PNL was completely stable in 50 mM Tris hydrochloride
buffer (pH 8.0) for several weeks at 4°C and for months whenstored at -20°C, with a negligible loss of activity after afreezing and thawing cycle. The enzyme was therefore
100
e 80
>; 60
0- 40° 40Icn
2U
010 20
TIME (min)30
FIG. 2. Effect of the DE of the substrate on the P. italicum PNLactivity as determined by viscosimetry. Enzyme activity was as-sayed with preparations obtained from either a submerged or asurface bran culture, with no differences being observed with thetwo preparations. Apple pectins (0.4% [wt/vol]) with DEs of 74%(0), 61% (A), 54% (A), and 37% (I) were used as substrates.
VOL. 56, 1990
on January 12, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
routinely stored at 4°C in diluted form (20 ,ug of protein perml) in the previously mentioned buffer. Ca2+, Mg2+, Mn2+,and Zn2+ did not affect PNL activity at 5 mM, while Co2+reduced it by 14%. However, Cu2+ and Fe2+, at the sameconcentration, produced complete inhibition. When a che-lating agent (ethylenedinitrilotetraacetic acid) was added at0.2 mM to the incubation mixture, PNL activity was notaffected.When highly methoxylated citrus pectin (DE, 82%) was
used as substrate, PNL activity was higher than with citruspectin of a lower DE (62%). The same characteristic wasexhibited by the PNL component of several commercialpreparations of pectinases (Opticlar-P, Pektolase PA, Rapi-dase CNP, and Ultrazym 100 G), with the increase in activitybeing dependent on the preparation (this increase variedfrom twofold in Ultrazym 100 G to fivefold in RapidaseCNP). This effect could be interpreted in terms of differentenzymatic action patterns.
DISCUSSION
As previously reported (1), P. italicum grown in sub-merged culture produces an extracellular PNL, the produc-tion of which is induced at low pHs by the presence of highlyesterified pectins. This PNL release is a common feature infilamentous fungi (23), although in Penicillium paxilli thisactivity has been found not only in the medium but alsoassociated with the mycelium (31). Besides PNL, polygalac-turonase is present in P. italicum culture filtrates, but PL andpectinesterase activities were not detected. To complete thisprevious study we have assayed, in addition to PNL and PLactivities, other enzyme activities produced not only in asubmerged culture but also in a surface bran culture. HigherPNL activity was routinely observed in a semisolid medium,but the specific activity of the filtrates was lower than thatobserved in a liquid one, because of the high protein contentof the former. The presence of polygalacturonase and pec-tinesterase and the absence of PL activity in a P. italicumsurface bran culture were reported by Bush and Codner (7).In our study, we have detected PL activity and observed thatits production depends on the growth medium composition;PL is released only when wheat bran is used as the carbonand energy source. Both growth media were effective in theproduction of other nonpectolytic enzymes, such as cellu-lase, a-amylase, and proteinase. The presence of theseactivities is important because of possible applications ofthese culture filtrates (13).The macerating activity observed by both P. italicum
culture filtrates adds new information to that reported byother authors (7) with orange tissue. This capacity to reducetissue cohesiveness has also been described for Penicilliumexpansum with potato tissue (28) and for Penicillium citri-num and P. paxilli preparations with potato and cucumbertissues (21, 30).
P. italicum PNL obtained from either submerged or sur-face bran cultures shows a basic isoelectric point (8.6), asdetermined by FPLC chromatofocusing. Although basicPNL has been described mainly for bacteria, this propertyhas also been reported in two isoenzymes separated fromculture filtrates of Colletotrichum lindemuthianum (33). Themolecular mass (22 kDa) of P. italicum PNL from bothgrowth media is lower than those (30 to 35 kDa) reported forthe enzyme isolated from a fungal source (Alania, Ph.D.thesis), although a PNL with a molecular mass of 18.2 kDahas been found in culture filtrates of Botrytis cinerea (17). Acommon feature of PNL enzymes is increasing activity as
pectin esterification increases (15). This feature has alsobeen observed, as determined by spectrophotometry andviscosimetry, in our preparations. Nevertheless, the exist-ence of two PNL isoenzymes in partially purified prepara-tions from Aureobasidium pullulans has been described,with one of these showing its maximum activity whenlow-DE pectins were used as substrates (22).Although PNL production by P. italicum growing in liquid
media has already been reported (1), no data about theproperties of the enzyme produced under these growthconditions are available. To this end, we performed variousstudies on the catalytic and physicochemical properties ofthe PNL produced in a submerged culture. This enzymecatalyzes the cleavage of highly esterified pectins and isunable to depolymerize polygalacturonic acid. Despite thedifficulties in comparing PNL catalytic parameters fromdifferent microorganisms, mainly because of the differentenzyme assays and substrates used, the calculated Km (3.2mg/ml) for citrus pectin (DE, 70%) is similar to thosedescribed for the PNLs of Aspergillus niger, P. citrinum,and P. paxilli (2.5 to 3.3 mg/ml) for the same substrate (21,30, 34). The observed decrease in the P. italicum PNLactivity when assayed at high citrus pectin concentrationshas also been reported (at 4 to 10 mg of pectin per ml) forcrude preparations from A. niger (34), B. cinerea (24), P.citrinum (21), and P. expansum (28). Although severalauthors have argued that this activity decrease is due toproduct inhibition during catalysis, some contradictory datasuggest that it is produced by the presence of unsaturateduronic acids in commercial pectins, as initially suggested byAlbersheim (2).The temperature effect on P. italicum PNL activity con-
sisted of an increase up to 50°C. This value is higher thanthose reported for filtrates of A. niger (34), Aureobasidiumpullulans (16), B. cinerea (24), and P. paxilli (30, 31), inwhich the maximum PNL activity is obtained at tempera-tures ranging from 35 to 40°C. PNL from P. italicum is morethermostable than other PNLs described for Penicilliumculture filtrates (21, 28, 30, 31), Aspergillus sojae (14), and C.lindemuthianum (33), in which thermoinactivation is ob-served in the range of 30 to 45°C.The enzyme from P. italicum is within a group of PNLs
with a slightly low optimum pH (6.0 to 7.0), as are thosedescribed for culture filtrates from A. niger (34), Aureoba-sidium pullulans (16), and P. expansum (28). No bacterialPNL with a maximum activity at a low pH has been reportedto date. The effect of pH on the stability of PNL from P.italicum culture filtrates shows that this enzyme is morestable at high pH values. These results show a wider range ofstability than that reported for P. paxilli (30, 31), in whichPNL is only stable in the pH range of 4.0 to 6.0. The stabilityshown by the P. italicum PNL activity after several monthsof storage at 4°C or freezing at -20°C is also remarkable.As observed in P. paxilli (30), and B. cinerea (24), the
presence of Ca2", Mg2+, Mn2+, and Zn2+ (at 5 mM) did notaffect the PNL activity of P. italicum. Nevertheless, in P.citrinum culture filtrates either monovalent or divalent (at2.5 mM) cations increase PNL activity, changing this effectto inhibition at fourfold-higher calcium concentrations (21).In P. italicum this negative effect on PNL activity is pro-duced by Co2+, Cu2+, and Fe2+ (at 5 mM). The presence ofethylenedinitrilotetraacetic acid at 0.2 mM did not affect thePNL activity of P. italicum, suggesting that cations were notessential for activity.Data about isoelectric point, molecular mass, and PNL
activity with respect to the substrate DE suggest that the two
3758 ALA&A ET AL.
on January 12, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

PNL ACTIVITY IN P. ITALICUM 3759
PNL activities found in semisolid and liquid media corre-sponded to the same enzyme and that P. italicum producesonly a single PNL. This argument is also corroborated by thefact that data for optimum pH and thermostability of P.italicum PNL produced in a surface bran culture (7) coincidewith our data for the same activity obtained in a liquidmedium. Many phytopathogenic microorganisms, dependingon the growth media composition, produce multiple forms ofPNL. Although this feature has been thought to be advan-tageous because some isoenzymes can be more stable undercertain conditions than other isoenzymes, the strategyshown by P. italicum could be more effective, as this fungusis able to produce a single PNL active in growth media ofdifferent composition and in wide ranges of pH and temper-ature.The importance of P. italicum in spoilage of citrus prod-
ucts and the role played by PNL in phytopathological andindustrial processes, as well as its increasing application tothe study of pectic polysaccharides and cell wall structures,seem to justify future attempts to obtain homogeneous PNLpreparations. This achievement will let us fill the gap inphysicochemical and catalytic data about this pectolyticenzyme.
ACKNOWLEDGMENTS
A.A. and I.A. were the recipients of scholarships from theSpanish Ministry of Education and Science (PFPI) and the BasqueGovernment, respectively. This work was partially supported bygrants from Diputaci6n Foral de Vizcaya (47-1/85) and Universidaddel Pafs Vasco/Euskal Herriko Unibertsitatea (310-0122/88).
LITERATURE CITED1. Alana, A., A. Gabilondo, F. Hernando, M. D. Moragues, J. B.
Donuinguez, M. J. Llama, and J. L. Serra. 1989. Production ofpectin lyase by a Penicillium italicum strain. Appl. Environ.Microbiol. 55:1612-1616.
2. Albersheim, P. 1963. Auxin induced product inhibition of pectintranseliminase as shown by ozonolysis. Plant Physiol. 38:426-429.
3. Albersheim, P., and U. Killias. 1962. Studies relating to thepurification and properties of pectin transeliminase. Arch. Bio-chem. Biophys. 97:107-115.
4. Asahi, M., R. Lindquist, K. Fukuyama, G. Apodaca, W. L.Epstein, and J. H. McKerrow. 1985. Purification and character-ization of major extracellular proteinases from Trichophytonrubrum. Biochem. J. 232:139-144.
5. Bradford, M. M. 1976. A rapid and sensitive method for thequantification of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72:248-254.
6. Bush, D. A., and R. C. Codner. 1968. The nature of maceratingfactor of Penicillium digitatum saccardo. Phytochemistry7:863-869.
7. Bush, D. A., and R. C. Codner. 1970. Comparison of theproperties of the pectin transeliminases of Penicillium digitatumand Penicillium italicum. Phytochemistry 9:87-97.
8. Chesson, A. 1980. Maceration in relation to the post-harvesthandling and processing of plant material. J. Appl. Bacteriol.48:1-45.
9. Collmer, A., and N. T. Keen. 1986. The role of pectic enzymesin plant pathogenesis. Annu. Rev. Phytopathol. 24:383-409.
10. Diaz-Borras, M. A., R. Vila Aguilar, and E. Hernandez Gimenez.1987. Comparaci6n de la actividad pectinolftica in vitro entrecepas de Penicillium sensibles y resistentes a fungicidas comer-ciales. Alimentaria 24:57-58.
11. Dietz, J. H., and A. H. Rouse. 1953. A rapid method for
estimating pectic substances in citrus juices. Food Res. 18:169-177.
12. Dische, Z. 1947. A new specific color reaction of hexuronicacids. J. Biol. Chem. 167:189-198.
13. Fogarty, W. M., and C. T. Kelly. 1983. Pectic enzymes, p.131-181. In W. M. Fogarty (ed.), Microbial enzymes andbiotechnology. Applied Science Publishers, London.
14. Ishii, S., and T. Yokotsuka. 1971. Maceration of plant tissues bypectin transeliminase. Agric. Biol. Chem. 35:1157-1159.
15. Ishii, S., and T. Yokotsuka. 1975. Purification and properties ofpectin lyase from Aspergillus japonicus. Agric. Biol. Chem.39:313-321.
16. Manachini, P. L., C. Parini, and M. G. Fortina. 1988. Pecticenzymes from Aureobasidium pullulans LV 10. Enzyme Mi-crob. Technol. 10:682-685.
17. Martinez, M. J., R. Martinez, and F. Reyes. 1988. Effect ofpectin on pectinases in autolysis of Botrytis cinerea. Myco-pathologia 102:37-43.
18. McCready, R. M. 1970. Pectin, p. 565-599. In M. A. Joslyn(ed.), Methods in food analysis. Academic Press, Inc. (Lon-don), Ltd., London.
19. Morell, S., L. Bauer, and K. P. Link. 1934. The methylglyco-sides of the naturally occurring hexuronic acids. III. Polygalac-turonic acid-methylglycosides derived from pectin. J. Biol.Chem. 105:1-13.
20. Nelson, N. 1944. A photometric adaptation of the Somogyimethod for the determination of glucose. J. Biol. Chem. 153:375-380.
21. Olutiola, P. O., and 0. A. Akintunde. 1979. Pectin lyase andpectin methyl esterase production by Penicillium citrinum.Trans. Br. Mycol. Soc. 72:49-55.
22. Parini, C., M. G. Fortina, and P. L. Manachini. 1988. Propertiesof two pectin lyases produced by Aureobasidium pullulans LV10. J. Appl. Bacteriol. 65:477-481.
23. Rexova-Benkova, L., and 0. Markovic. 1976. Pectic enzymes.Adv. Carbohydr. Chem. Biochem. 33:323-385.
24. Reyes, F., M. J. Martinez, R. La Hoz, and S. A. Archer. 1984.Characterization as glycerol of an inhibitor of pectin lyases fromautolysing cultures of Botrytis cinerea. Trans. Br. Mycol. Soc.82:689-696.
25. Rombouts, F. M., and W. Pilnik. 1980. Pectic enzymes, p.227-282. In A. H. Rose (ed.), Economic microbiology. Micro-bial enzymes and bioconversions. Academic Press, Inc. (Lon-don), Ltd., London.
26. Schultz, T. H. 1965. Determination of the degree of esterificationof pectin. Methods Carbohydr. Chem. 5:189-194.
27. Somogyi, M. 1952. Notes on sugar determination. J. Biol. Chem.195:19-23.
28. Spalding, R. T., and A. A. Abdul-Baki. 1973. In vitro and in vivoproduction of pectin lyase by Penicillium expansum. Phytopa-thology 63:231-235.
29. Spiro, R. G. 1966. Analysis of sugars found in glycoproteins.Methods Enzymol. 8:3-26.
30. Szajer, I., and C. Szajer. 1982. Pectin lyase of Penicilliumpaxilli. Biotechnol. Lett. 4:549-552.
31. Szajer, I., and C. Szajer. 1985. Formation and release of pectinlyase during growth of Penicillium paxilli. Biotechnol. Lett.7:105-108.
32. Voragen, A. G. J., and W. Pilnik. 1989. Pectin-degradingenzymes in fruit and vegetable processing, p. 93-115. In J. R.Whitaker and P. E. Sonnet (ed.), Biocatalysis in agriculturalbiotechnology. American Chemical Society, Washington, D.C.
33. Wiesundera, R. L. C., J. A. Bailey, and R. J. W. Byrde. 1984.Production of pectin lyase by Colletotrichum lindemuthianum inculture and in infected bean (Phaseolus vulgaris) tissue. J. GenMicrobiol. 130:285-290.
34. Zetelaki-Horvwith, K. 1982. Factors affecting pectin lyase activ-ity. Acta Aliment. 11:21-29.
VOL. 56, 1990
on January 12, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from












![ŠTETNICI SMILJA Helichrysum italicum G. Don.] U PLANTAŽNOM](https://img.pdfslide.net/doc/110x75/626ef272b00e9b56707b4c1a/tetnici-smilja-helichrysum-italicum-g-don-u-plantaznom-.jpg)
















