Embed Size (px)
Citation preview

From the Department of Clinical Neuroscience
Karolinska Institutet, Stockholm, Sweden
PET IMAGING OF TWO MONOAMINERGIC NEUROTRANSMITTER SYSTEMS IN BRAIN:
STUDIES OF THE NOREPINEPHRINE TRANSPORTER AND DOPAMINE D2 RECEPTOR
Nicholas Seneca
Stockholm 2006

All previously published papers were reproduced with permission from the publisher. Published and printed by Karolinska University Press Box 200, SE-171 77 Stockholm, Sweden © Nicholas Seneca, 2006 ISBN 91-7140-923-8

ABSTRACT Positron emission tomography (PET) has been widely used to study non-
invasively function of the brain, pathophysiology of disease and aid in the development of new drugs. PET and selective radiolabeled molecules allow imaging of certain critical components of neurotransmission, such as pre-synaptic transporters and post-synaptic receptors in living brain. The general aim of the present thesis was (i) to measure neuropharmacological interventions using PET (e.g., competition between synaptic neurotransmitters and radiolabeled tracers provide a useful tool to estimate changes in synaptic levels of neurotransmitters), and (ii) PET determination of drug induced receptor occupancy, examined by the curvilinear relationship between drug plasma concentration and receptor occupancy over a wide dose-range in nonhuman primate brain.
The aim of Papers I and II was to evaluate (S,S)-[18F]FMeNER-D2 as a radiotracer for the norepinephrine transporter (NET). Paper I examined the whole-body biodistribution of (S,S)-[18F]FMeNER-D2 and estimated the resulting radiation exposure to organs of the body in nonhuman primates. The estimated radiation burden of (S,S)-[18F]FMeNER-D2 was found to be comparable to that of other 18F radioligands. In Paper II, the aim was to determine if atomoxetine occupies NET in a dose-dependent fashion using (S,S)-[18F]FMeNER-D2 in nonhuman primate brain. Previous PET studies have failed to demonstrate the feasibility of measuring a dose-dependent NET occupancy. After administration of increasing doses of atomoxetine, a dose-dependent occupancy from 38% to 82% was observed in brain regions known to contain high densities of NET.
The aim of Papers III and IV was to combine PET imaging of the D2 receptor with pharmacological challenges that either increase or decrease concentrations of dopamine. In Paper III, the aim was to assess and compare in nonhuman primate brain the sensitivity of the agonist radioligand [11C]MNPA and antagonist [11C]raclopride to stimulant-induced dopamine release. [11C]MNPA binding potential was ~50% more sensitive than [11C]raclopride to pharmacological induced increases in dopamine. The aim of Paper IV was to estimate the occupancy of D2 receptors in rat brain by endogenous dopamine using PET and [11C]MNPA. The dopamine depletion paradigm increased [11C]MNPA binding potential significantly and the results indicate ~27% of D2 high affinity state receptors are occupied by endogenous dopamine during basal conditions.
The aim of Paper V was to determine D2 and 5-HT1A receptor occupancy in brain after administration of RGH-188. In addition, the intrinsic activity of RGH-188 was estimated in vivo using a dopamine D2 agonist and antagonist radioligand. RGH-188 occupied D2 receptors in a dose dependent and saturable manner, with lowest dose occupying ~5% of receptors, and highest dose more than 90%. RGH-188 was equally potent to displace both antagonist and agonist radioligand. 5-HT1A receptor occupancy was much lower compared to D2 occupancy at the same doses, with a maximal value of ~30%. These results suggest that RGH-188 binds to D2 rather than 5-HT1A receptors, and its equal potency to displace agonist and antagonist radiotracers suggests that RGH-188 is an antagonist.
1

2
“Our life is what our thoughts make of it.”
— Marcus Aurelius, Roman Emperor, A.D. 161-180

LIST OF PUBLICATIONS The thesis is based on the following papers and manuscripts:
I. Seneca N, Andree B, Sjoholm N, Schou M, Pauli S, Mozley PD, Stubbs JB, Liow JS, Sovago J, Gulyas B, Innis R, Halldin C. Whole-body biodistribution, radiation dosimetry estimates for the PET norepinephrine transporter probe (S,S)-[18F]FMeNER-D2 in non-human primates. Nucl Med Commun 2005; 26(8):695-700.
II. Seneca N, Gulyás B, Varrone A, Schou M, Tauscher J, Vandenhende F, Kielbasa W, Farde L, Innis RB, Halldin C. Atomoxetine occupies the norepinephrine transporter in a dose-dependent fashion: A PET study in nonhuman primate brain using (S,S)-[18F]FMeNER-D2. Psychopharmacology 2006; 188(1):119-27.
III. Seneca N, Finnema SJ, Farde L, Gulyas B, Wikstrom HV, Halldin C, Innis RB. Effect of amphetamine on dopamine D2 receptor binding in nonhuman primate brain: A comparison of the agonist radioligand [11C]MNPA and antagonist [11C]raclopride. Synapse 2006; 59(5):260-269.
IV. Seneca N, Zoghbi SS, Liow JS, Hong J, Pike VW, Halldin C, and Innis RB. PET imaging in rat brain of endogenous dopamine occupancy of D2/3 receptors using the agonist radioligand [11C]MNPA. Manuscript.
V. Seneca N, Finnema S, Innis RB, Halldin C, Laszlovszky I, Kiss B, Horváth A, Pásztor G, Kapás M, Gyertyán I, Farkas S, and Gulyás B. Occupancy of dopamine D3/D2 and serotonin 5-HT1A receptors by a novel D3/D2 antipsychotic drug in nonhuman primate brain: A PET study. Manuscript.
3

CONTENTS 1 INTRODUCTION ....................................................................................... 6
1.1 POSITRON EMISSION TOMOGRAPHY (PET) ........................... 6 1.1.1 Principles of PET ................................................................... 6 1.1.2 Applications of PET neuroreceptor imaging......................... 7
1.2 NORADRENERGIC SYSTEM ........................................................ 8 1.2.1 Noradrenergic projections...................................................... 8 1.2.2 Role of norepinephrine and the norepinephrine transporter . 9 1.2.3 PET imaging of the norepinephrine transporter.................... 9
1.3 DOPAMINERGIC SYSTEM.......................................................... 10 1.3.1 Dopaminergic projections.................................................... 10 1.3.2 Dopaminergic receptors ....................................................... 11 1.3.3 Functions of dopamine......................................................... 12 1.3.4 PET imaging of dopamine receptors ................................... 13
1.4 INTERACTION BETWEEN DOPAMINE AND GLUTAMATE15 1.4.1 Glutamate receptors ............................................................. 15 1.4.2 Interactions between dopamine and glutamate ................... 16 1.4.3 PET studies of dopamine glutamate interactions ................ 18
2 AIMS .......................................................................................................... 20 3 MATERIALS AND METHODS .............................................................. 21
3.1 SUBJECTS....................................................................................... 21 3.2 MATERIALS AND RADIOLIGANDS ......................................... 21 3.3 POSITRON EMISSION TOMOGRAPHY .................................... 21 3.4 PET EXPERIMENTAL PROCEDURES ....................................... 22 3.5 EXPERIMENTAL DRUGS AND DOSE SELECTION ............... 23 3.6 IMAGE ANLAYSIS........................................................................ 25 3.7 CALCULATIONS ........................................................................... 25 3.8 NEUROPHARMACOLOGICAL INTERVENTIONS ................. 26 3.9 RECEPTOR OCCUPANCY ........................................................... 27 3.10 STATISTICAL ANALYSIS........................................................ 28
4 RESULTS AND DISCUSSION................................................................ 29 5 SUMMARY OF FINDINGS..................................................................... 39 6 FUTURE PROSPECTS............................................................................. 40 7 ACKNOWLEDGEMENTS....................................................................... 41 8 REFERENCES........................................................................................... 43
4

LIST OF ABBREVIATIONS ADHD Bmax BP CNS DA ED50 EPS fMRI i.v. i.p. Ki L-dopa MAP MeNER MIRD mGluR MNPA MPEP MRI mRNA MRTM2 NET NMDA %ID PET PMOD R1 ROI SA SPECT SPM2 SUV
Attention deficit hyperactivity disorder Receptor density Binding potential Central nervous system Dopamine Effective dose to occupy 50% of receptors Extrapyramidal side effects Functional magnetic resonance imaging Intravenous Intraperitoneal Equilibrium dissociation constant Levodopa Methamphetamine Methyl norethyl reboxetine Medical internal radiation dose Metabotropic glutamate receptors (R)-2-CH3O-N-n-propylnorapomorphine [2-methyl-6-(phenylethynyl)pyridine] Magnetic resonance imaging Messenger ribonucleic acid Multilinear reference tissue model Norepinephrine transporter N-methyl-D-aspartate receptors Percent injected dose Positron emission tomography Image quantification and kinetic modeling software Relative blood flow image Region of interest Specific radioactivity Single photon emission computerized tomography Statistical parametric mapping Standard uptake value
5

1 INTRODUCTION 1.1 POSITRON EMISSION TOMOGRAPHY (PET)
PET has been widely used to study noninvasively function of the brain,
pathophysiology and patient treatment management. Using neuroimaging, we are able
to track biochemical changes in the brain by imaging specific neurotransmitter
systems. Changes detected in certain neurotransmitter systems can then be used as a
lead to reveal the pathophysiology of patients suffering from psychiatric, neurological
and neurodegenerative disorders.
The PET technique utilizes radiotracers labeled with relatively short-lived
positron-emitting radionuclides such as 18F, 11C, or 15O. A low amount of mass can be
used due to the relatively high specific radioactivity (SA). Radioligands allow us to
study several biological variables such as: anatomical distribution, binding in the
brain, drug-related receptor occupancy and metabolism of the tracer. Several criteria
need to be fulfilled for PET radioligands: including high affinity and selectivity for
the target site, minimal metabolism in tissues, permeability across the blood-brain
barrier, and low non-specific binding (Halldin et al., 2001a; Lee and Farde, 2006).
The radioligands typically are evaluated in several steps, from the early evaluation in
rodents, including knockout mice, nonhuman primate PET imaging, and human post
mortem autoradiography, to the study of human physiology initially in healthy
subjects and then as applied to patients with psychiatric and neurological disorders.
1.1.1 Principles of PET PET is an in vivo molecular imaging
technique that uses positron labeled
molecules to image molecular
interactions of biological processes. A
positron-emitting radionuclide ejects a
positron from the nucleus as it decays.
The positron travels a short distance in
tissue and loses energy through
interactions with atoms. The positron
will combine with an electron in tissue
and annihilate, simultaneously
producing two 511 keV gamma rays that are emitted 180º apart (Figure 1). PET
Figure 1. The physics underlying positron emission tomography (PET). (Adapted from Cherry 2001).
6

7
scanners contain several rings of positron-sensitive scintillation detectors, with up to
25,000 individual scintillators. The pair of photons produced from a single
annihilation will register almost simultaneously on opposing pairs of scintillation
detectors as a “coincidence event.” The rings of scintillation detectors register
thousands of coincidence events emitted from the subject per second (Phelps and
Mazziotta, 1985; Cherry, 2001). The data gathered from the coincidence events are
used to determine the source of positron annihilation at a given time. These are then
converted into a tomographic image via reconstruction algorithms.
1.1.2 Applications of PET neuroreceptor imaging Several monoamine neurotransmitter systems have been studied in animals,
healthy subjects and patient populations suffering from psychiatric disorders using
PET. The dopaminergic system has been the most extensively investigated in terms of
both pre-synaptic and post-synaptic biological markers. Exploration of the
serotoninergic system has shed light on new drugs acting on this system and has
played a role in the understanding of the pathophysiology of many psychiatric
disorders. The shortage of radioligands poses a serious limiting factor to PET
researchers. Thus, development of new radioligands is crucial to enable further study
of neurotransmitter systems in vivo such as the norepinephrine system, glutamatergic
neurotransmission pathways as well as dopamine subtype receptors and reuptake
sites.
Monoamine transporter imaging for the dopamine, serotonin and to a lesser
extent the norepinephrine transporter have proven useful in assessing transporter
occupancy of well characterized and novel drugs, and as markers for
neurodegenerative diseases (i.e., evaluation of neuronal cell loss of the dopaminergic
system and the associated loss of radioligand binding to the dopamine transporter)
(Laakso and Hietala, 2000). Biomarkers for pathophysiology of various disorders are
being developed, for example, the binding of [11C]PIB reflects the amount of β-
amyloid plaques in brain (Klunk et al., 2004). Quantification of β-amyloid plaques
provides a useful tool to gain insight into the involvement of β-amyloid plaques in the
pathology of Alzheimer’s disease and monitoring therapeutics under development
(Nichols et al., 2006).

8
Small animal imaging using microPET has several advantages: aid in the
development of novel radiotracers, administration of pre-clinical drugs, study animal
models of human diseases, and cost effective compared to nonhuman primates (Phelps,
2000; Cherry, 2001). Rodent, monkey and human brain weight may vary drastically
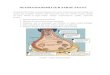
from ~2, ~65, to ~1300 g, however, PET imaging of D2/3 receptors using
[11C]raclopride across species provides a similar outcome measure (Figure 2).
Neuroreceptor PET imaging is just one example of how PET can provide a bridge from
rodent to humans, as the same imaging experiments can be translated across species
(Cherry, 2001).
4.0
0
BP
Rat Brain Human Brain Monkey Brain
Figure 2. Parametric images of [11C]raclopride binding potential estimated with a reference tissue model. (Rat PET data is courtesy of Dr. Tetsuya Suhara, NIRS, Chiba, Japan).
1.2 NORADRENERGIC SYSTEM 1.2.1 Noradrenergic projections
The major noradrenergic nucleus in
the brain is the locus coeruleus, which
is located in the rostral pons (Figure
3). These neurons give rise to most of
the noradrenergic innervation of the in
the cerebral cortex, thalamus,
cerebellum, and brain stem nuclei. In
contrast to dopaminergic neurons,
which show relatively restricted
axonal projections, noradrenergic
neurons give rise to diffuse axonal
projections and innervate virtually all
Figure 3. Noradrenergic projection systems in the brain. The major noradrenergic nuclei of the brain, the locus coeruleus and the lateral tegmental nuclei. (Adapted from Hyman and Nestler, 1993).

9
areas of the brain and spinal cord. In addition, the brain also contains smaller
collections of additional noradrenergic neurons that are located in discrete regions of
the pons and medulla. These norepinephrine-containing cells project to basal forebrain,
thalamus, including certain nuclei in the hypothalamus, limbic areas such as the
amygdala, hippocampus, and brain stem.
1.2.2 Role of norepinephrine and the norepinephrine transporter Norepinephrine, also known as noradrenaline, plays an important role in human
physiology and pathology, and is involved in mood regulation, sleep regulation,
expression of behavior and the general degree of alertness and arousal (McCormick et
al., 1991; Mandela and Ordway, 2006; Mazei et al., 2002). Norepinephrine also exerts
its effects over the endocrine system and autonomic nervous system. Outside the CNS
and adrenal glands, norepinephrine is located in the sympathetic nerve endings, and the
norepinephrine content of a particular tissue reflects the extent of its sympathetic
innervation.
The norepinephrine transporter is located in the plasma membrane of
noradrenergic neurons, where it functions to take up synaptically released
norepinephrine (Mandela and Ordway, 2006). The transporter serves as the primary
mechanism for the inactivation of noradrenergic signaling. Reuptake of norepinephrine
by the transporter protein is the primary mechanism by which the biological effects of
norepinephrine in the synapse are terminated. The inactivation process through the
transporter is critical in preventing an excessive increase in the norepinephrine
concentrations in the synaptic cleft, which regulate adrenergic neurotransmission in the
brain, as well as the removal of norepinephrine from the heart and other peripheral
organs. Reuptake of norepinephrine is competitive with a variety of naturally occurring
amines and drugs. Drugs of abuse (e.g., cocaine) and antidepressants (e.g., desipramine,
reboxetine, bupropion) block the transport of norepinephrine and thereby cause an
elevation in the synaptic concentrations of norepinephrine and potentiation of the
activation of postsynaptic receptors (Zhou, 2004; Berton and Nestler, 2006).
Norepinphrine plays an important role in the CNS, and is likely to play a role in various
psychiatric disorders, including major depression and ADHD.
1.2.3 PET imaging of the norepinephrine transporter Imaging agents targeting the norepinphrine transporter (NET) in brain have
lagged due to a lack of selective NET radioligands that give a high signal to noise
ratio. Nisoxetine has high affinity for NET in vitro, nevertheless, [11C]nisoxetine
exhibits high levels of non-specific binding in vivo (Haka et al., 1989). Several

10
analogs of cocaine have been radiolabeled as probes for NET, but due to high affinity
to dopamine transporter and the low density of NET in brain, the radioligands
selectively labeled the dopamine transporter in vivo (Musachio et al., 2006).
Evaluation of 11C-radiolabeled desipramine, talopram, and talsupram demonstrated
low brain uptake and lack of selective NET binding in vivo, even though they exhibit
high selectivity for NET over dopamine and serotonin transporters (Schou et al.,
2006; McConathy et al., 2004).
Recently, 11C and 18F labeled analogs of reboxetine have been prepared and
evaluated as PET radioligands in rodent, monkey and human (Wilson et al., 2003;
Schou et al., 2003; Ding et al., 2003 Andree et al., 2004). In vitro autoradiography and
in vivo PET imaging have demonstrated selective binding of [11C]MeNER and
[18F]FMeNER-D2 to NET. Further evaluation of [11C]MeNER in nonhuman primate
and human brain showed that specific binding to NET did not reach maximal values
(i.e. equilibrium) during the PET measurement. This, together with a somewhat noisy
final signal at later time points, are relative deficiencies of [11C]MeNER for quantitative
studies of NET in brain. These deficiencies lead to the preparation and evaluation of
radiofluorinated analogs of (S,S)-MeNER (Schou et al., 2004). The results indicated
that (S,S)-[18F]FMeNER-D2 is a useful radioligand for imaging NET in monkey and is
superior to [11C]MeNER given that a specific binding peak equilibrium is reached
during the PET experiment at a lower noise level. The advantage of the 18F-radioligand
lead to further development with in vitro human brain autoradiography (Schou et al.,
2005), and calculated dosimetry estimates for (S,S)-[18F]FMeNER-D2 were comparable
to other 18F-labeled radioligands allowing for multiple injections in human subjects
(Seneca et al., 2005). In addition, human NET occupancy studies using [11C]MeNER
showed a lack of complete saturation and dose-dependent occupancy across various
plasma concentrations of reboxetine and atomoxetine, but using (S,S)-[18F]FMeNER-
D2 a dose dependent and complete saturation in brain was demonstrated (Andree et al.,
2004; Wong et al., 2005; Seneca et al., 2006). In total, these data encourage further
PET studies using (S,S)-[18F]FMeNER-D2 in humans.
1.3 DOPAMINERGIC SYSTEM 1.3.1 Dopaminergic projections
Dopaminergic neurons in the substania nigra pars compacta, the ventral
tegmental area, and the hypothalamus give origin to four main pathways, the
nigrostriatal, the mesolimbic, the mesocortical, and the tuberoinfundibular (Figure 4).

11
The nigrostriatal system projects from the substantia nigra to the dorsal
striatum. This system is important for the control of movement and is affected in
Parkinson’s disease and other disorders of movement. The mesolimbic system projects
from the ventral tegmental
area to limbic structures
such as the ventral striatum
(i.e., nucleus accumbens),
hippocampus and amygdala.
The mesocortical system
projects from the ventral
tegmental area to cortical
regions. Both the
mesolimbic and
mesocortical projections are
important for affect,
emotion, and motivation and are affected in schizophrenia. The fourth dopaminergic
projection originates in the arcuate nucleus of the hypothalamus and projects to the
pituitary gland, where it regulates secretion of hormones.
Figure 4. Dopaminergic projection systems in the brain. (Adapted from Hyman and Nestler, 1993).
1.3.2 Dopaminergic receptors Dopamine exerts its action by binding to specific membrane receptors, which
belong to the family of seven transmembrane domain G-protein coupled receptors. Five
distinct dopamine receptors have been divided into two sub families based on their
biochemical and pharmacological properties (Vallone et al., 2000; Missale et al., 1998;
Jaber et al., 1996). The D1-like family comprises of D1 and D5, while the D2-like
includes D2, D3, and D4. Receptors of the D1-like family stimulate adenylate cyclase,
on the other hand, the D2-like family of receptors inhibit this effector.
The D1 and D2 have the most widespread and highest levels of expression of the
dopamine receptors. The D1 receptor is mainly expressed in the striatum, ventral
striatum, cerebral cortex and amygdala. The D2 receptor is expressed predominately in
the striatum and lower densities in thalamus and cortex. The D3 receptor is concentrated
in limbic areas (e.g., islands of Calleja and nucleus accumbens) that may mediate
abnormalities of memory, speech, and focused attention in schizophrenia (Sokoloff et
al., 1990; Suzuki et al., 1998). The highest density of D4 receptors are located in
cerebral cortex, hippocampus, hypothalamus and amygdala (Primus et al., 1997). The

12
D5 highest density is the hippocampus and parafascicular nucleus of the thalamus
(Jackson et al., 1994).
1.3.3 Functions of dopamine Dopamine is the most abundant catecholamine in brain and involved in the
regulation of different physiological functions in the central nervous system, including
locomotor activity, and reward mechanisms (Chinta et al., 2005; Girault et al., 2004).
Dysregulation of dopaminergic neurotransmission has been associated with multiple
neurological and psychiatric conditions such as Parkinson’s disease, and schizophrenia.
Parkinson’s disease
Parkinson’s disease is one of the most common neurodegenerative disorders,
and affects approximately 1.5% of individuals over the age of 65, and 2.6% of
individuals over the age of 80 (Stoessl and Fernandez, 2003). Dopamine deficiency,
caused by the degeneration of nigrostriatal dopaminergic neurons, is the cause of the
major clinical motor symptoms of Parkinson’s disease. Within the substantia nigra,
neuronal loss tends to be estimated to be 60 to 70% at the onset of symptoms (Lang and
Lozano, 1998). This results in severe dopamine depletion in the striatum, responsible
for the motor symptoms associated with Parkinson’s disease. The primary symptoms of
Parkinson’s disease include muscular rigidity, resting tremor, difficulty with movement
initiation (bradykinesia), slowness of voluntary movement, difficulty with balance, and
difficulty with walking.
Levodopa (L-dopa) remains the most effective treatment for Parkinson’s
disease, this drug remains the most effective treatment of the slowness of movement,
increased muscle tone, and tremor that are typical of Parkinson’s disease (Olanow,
2002). The therapeutic response of L-dopa in Parkinson’s disease can be attributed to
several factors: 1) neuronal responses to L-dopa are mediated by its metabolic
transformation to dopamine and noradrenaline, 2) dopamine produced by L-dopa
activates D1-like and D2-like receptors, 3) facilitates noradrenaline mediated
activation of α- and β- adrenoceptors following its conversion to noradrenaline.
Although L-dopa reduces many of the motor symptoms of Parkinson’s disease, it
does not affect non-motor symptoms and does not halt the progression of the
degeneration of dopamine-containing neurons in the substantia nigra. A main
problem associated with Parkinson’s disease is the fact that dopamine-containing
neurons continue to die and ultimately there is not enough release of dopamine, even
after L-dopa treatment, to maintain essential functions (Mercuri et al., 2005).
Schizophrenia

13
Schizophrenia is a debilitating mental illness that affects 1% of the population.
Symptoms of schizophrenia are often divided into positive symptoms that include
hallucinations, delusions, thought disorganization, and negative symptoms such as
blunted affect, social withdrawal, and decreased cognitive function. The “classical”
dopaminergic hypothesis of schizophrenia proposed that excessive release of dopamine
during synaptic transmission underlies at least some aspects of the pathogenesis of
schizophrenia has received its primary support from pharmacological studies (Carlsson
and Lindqvist, 1963). This was supported by two observations 1) the correlation
between antipsychotic potency of neuroleptics and their potency to block D2 receptors,
and 2) drugs that increase the level of dopamine, such as L-dopa, cocaine, and
amphetamine can induce psychotic episodes resembling schizophrenia.
Antipsychotic drugs are effective for the treatment of schizophrenia.
Antagonism at the dopamine D2 receptor is considered an essential component of the
mechanism of action (Creese et al., 1976; Seeman et al., 1975). Classical or “typical”
antipsychotics (e.g. haloperidol, chlorpromazine) are associated with high affinity for
D2 receptors and effectively treat the positive symptoms of schizophrenia. On the other
hand, “atypical” antipsychotics such as clozapine also improve negative symptoms and
cognition, and are useful in patients who have failed treatment with typical
antipsychotics (Casey et al., 1989; Meltzer et al., 1991). In addition, clozapine causes
fewer extrapyrmidal side effects (EPS) than typical antipsychotics, possibly because of
low striatal D2 receptor occupancy and its actions at other targets (e.g. antagonist at
serotonin 5-HT2A receptors).
1.3.4 PET imaging of dopamine receptors In vitro and in vivo neuroreceptor imaging studies using autoradiography and
PET imaging have contributed to a greater understanding of the dopamine system. The
dopamine receptor system is by far the most studied neurotransmitter system of the
brain. This is primarily due to the availability of a number of well-characterized
radiotracers and to great interest in this system, especially in psychiatry. PET
radioligands for the dopamine D1 and D2/3 receptors have been studied extensively. The
present thesis focuses on PET imaging of dopamine D2/3 receptor subtypes:
Dopamine D2/3 receptors
Early brain imaging studies using D2/3 receptor radioligands (N-[methyl 11C]methylspiperone (NMSP), [11C]raclopride, and [123I]IBZM) enabled imaging of
D2/3 receptors in striatum (Halldin et al., 2001a). Studies comparing parameters of D2
receptor binding in patients with schizophrenia and healthy controls have been

14
extensively studied (Wong et al., 1986; Farde et al., 1990; Hietala et al., 1994; Laruelle
et al., 1996).
In vivo neuroreceptor binding techniques can be used to measure acute
fluctuations in the concentration of endogenous transmitters. Competition between
radioligands and agents that increase synaptic dopamine concentrations like
amphetamine have been shown in human and nonhuman primates to decrease D2
receptor radioligand binding (Innis et al., 1992; Farde et al., 1992; Volkow et al., 1994;
Laruelle et al., 1995; Carson et al., 1997; review: Laruelle, 2000). Several studies
reported that amphetamine induced dopamine release is increased in patients with
schizophrenia compared to matched healthy controls, and resulted in an worsening of
positive symptoms (Laruelle et al., 1996; Breier et al., 1997). These results provide
direct evidence that exaggerated activiation of dopamine transmission at D2 receptors
mediates the expression of psychotic symptoms following amphetamine challenge.
Similarly, agents that reduce synaptic dopamine levels (e.g., reserpine or alpha-methyl-
para-tyrosine) decrease competition by the endogenous transmitter and thereby
”unmask” D2 receptors and increase radioligand binding (Ginovart et al., 1997;
Laruelle et al., 1997; Verhoeff et al., 2001). Comparing D2 receptor availability at
baseline and in the dopamine depleted state provides an indirect measure of the
proporation of D2 receptors occupied by dopamine in the baseline state. The results of
pharmacological challenges, both amphetamine stimulated dopamine release and
endogenous dopamine depletion provide direct evidence of the dopamine hypothesis of
schizophrenia.
The majority of PET imaging studies of the D2 receptor have been performed
with antagonist radioligands, such [11C]raclopride. Antagonists of G-protein coupled
receptors, such as the D2 receptor, have equal affinity for receptors in the high (i.e.,
coupled) or low (i.e., uncoupled) affinity state (Creese et al., 1984; George et al., 1985;
Seeman et al., 1985; Sibley et al., 1983). Since dopamine is the endogenous agonist for
the D2 receptor, it would more effectively compete with the binding of an agonist,
compared to an antagonist radioligand. Recent development of three D2 agonist
radioligands, [11C]NPA, [11C]MNPA, and [11C](+)-PHNO make it possible to test this
hypothesis (Hwang et al., 2000; Finnema et al., 2005; Wilson et al., 2005). The use of a
D2 agonist radioligand may shed more light into the pathophysiology of the functional
state of the D2 receptor as initial evaluation studies in rodents and nonhuman primates
can now be measured in healthy control and neuropsychiatric patients.

15
PET can also be used as a tool in drug development in either a direct or indirect
manner to assess the in vivo activities of a drug (Halldin et al., 2001b; Farde and Lee
2006; Farde et al., 1996; Talbot et al., 2002; Brooks et al., 2005). Several well-
characterized PET radioligands have been used to measure receptor occupancies of
antipsychotic agents at several neurotransmitter systems. The main focus has been on
the D2 receptor in striatum using [11C]raclopride and in extrastriatal regions with
[11C]FLB 457 (Halldin et al., 1995; Farde et al., 1986). PET studies using
[11C]raclopride have demonstrated a correlation between the percentage of striatal D2
receptor occupancy and clinical response in patients treated with a range of
antipsychotics (Farde et al., 1988; Farde et al., 1989). The therapeutic effect of typical
antipsychotic medications occurs at striatal D2 receptor occupancies of ∼ 65-70%, with
significantly increased risk of EPS at occupancies >80% (Farde et al., 1992; Nordstrom
et al., 1993; Kapur et al., 2000). In comparison to these standard agents, atypical
antipsychotics, such as clozapine and quetiapine, have a reduced tendency to induce
EPS and have lower occupancy of striatal D2 receptors (Farde et al., 1992; Nordstrom
et al., 1995; Kapur et al., 2000).
1.4 INTERACTION BETWEEN DOPAMINE AND GLUTAMATE 1.4.1 Glutamate receptors
The major excitatory transmitter in brain is the amino acid L-glutamate.
Glutamate is involved in many brain functions, such as differentiation, neuronal cell
survival and death, proliferation and development of neuronal and glial cells, and
Figure 5. Glutamate receptors and their effector systems. (Adapted from Spooren, 2004).

16
plastic changes in efficacy of synaptic transmission (Nakanishi, 1992). Glutamate
neurotoxicity can result in neurodegeneration and neuronal cell death in disorders
such as Alzheimer and Huntington’s disease. Glutamate activates two families of
receptors: ionotropic, which are ligand-gated ion channels and metabotropic
receptors, G-protein coupled receptors linked to second messenger pathways (Conn et
al., 1997; Schoepp et al., 1999) (Figure 5).
Ionotropic and metabotropic glutamate receptors
Ionotropic glutamate receptors are glutamate-gated ion channels that when
activated increase cellular excitability. The NMDA receptor is one subgroup of
ligand-gated channel receptors, which is highly permeable to Ca2+, Na+, and K+, and
the resultant increase of intracellular Ca2+ is thought to be responsible for evoking
both neuronal plasticity and neurotoxicity (Nakanishi et al., 1994). On the other hand,
metabotropic glutamate receptors (mGluR) have been classified into three groups
based on sequence similarities, signal transduction pathways and pharmacological
characterization (Pin et al., 2003). Group I receptors (mGluR 1 and 5) are linked to
the activation of phospholipase C and generally mediate postsynaptic excitatory
effects, whereas group II (mGluR 2 and 3) and group III (mGluR 4, 6, 7 and 8)
receptors are negatively coupled to adenylyl cyclase and generally mediate
presynaptic inhibitory influences on neurotransmitter release (Hermans et al., 2001).
While glutamate binds to the large extracellular region, the action of non-competitive
antagonists such as MPEP, are thought to bind within the seven-transmembrane
domain (Pagano et al., 2000). It is speculated that the action of the non-competitive
antagonists binding within the seven-transmembrane domain of mGluR5s act to
stabilize the inactive state of the receptor and therefore inhibit the constitutive
activity.
1.4.2 Interactions between dopamine and glutamate Abnormalities of various neurotransmitter systems have been proposed in
patients suffering from schizophrenia. Research has been primarily driven by the
dopamine hypothesis of schizophrenia proposing a hyperactivity of dopamine
transmission that is responsible for the positive symptoms associated with this disorder.
Nevertheless, some patients fail to respond to treatment with dopamine antagonists and
while amphetamine causes psychosis, administration of ketamine or PCP have been
found to induce both positive and negative symptoms similar to that of patients
suffering from schizophrenia. NMDA receptor hypofunction has been suggested with

17
patients suffering from schizophrenia (Olney and Farber, 1995; Tamminga et al., 1995).
NMDA receptor blockade by acute administration of ketamine has been shown to
induce both positive and negative symptoms in healthy and schizophrenic patients.
NMDA hypofunction in schizophrenic patients most likely occurs chronically rather
than acutely, sustained dysfunction may lead to alterations in dopamine transmission
(cortical dopamine deficit and subcortical dopamine hyperactivity).
A multi-factorial view has been proposed in which neurotransmitter interactions
may shed more light into the pathophysiology of schizophrenia (Carlsson et al., 1999;
Carlsson et al., 2000). The interaction between glutamate and dopamine has lead to the
theory in which a ‘feedback loop’ exists to protect the cerebral cortex against any
elevated concentrations of both neurotransmitters regulated via a brake and accelerator.
For example, if dopamine is enhanced with amphetamine, a negative feedback
regulatory circuit appears to be activated leading to counter act the excess release. It has
been proposed that the ‘brake’, which counteracts the excess of dopamine tone fails to
function properly in patients suffering from schizophrenia. The interaction between
dopamine and glutamate has not been limited to ionotropic glutamate receptors, more
recently interactions between mGluR have been shown to affect dopamine-mediated
responses.
In vitro microdialysis studies by Golembiowska et al., 2003 demonstrated that
the effects of a systematic injection of MPEP reduced pharmacological induced
dopamine release. This inhibition reduced dopamine levels by nearly 50% compared to
methamphetamine only pretreatment. These data suggest that blockade of mGluR5 may
protect striatum terminals of dopamine neurons against methamphetamine induced
toxicity. Nevertheless, local administration of MPEP in the striatum did not decrease
but even increased dopamine levels. Thus, the modulation of excessive dopamine
release may be due to the actions of mGluR5 located outside the striatum. Further data
by Pietraszek et al., 2004 supported the interactions of mGluR5 inhibition of the
amphetamine effect in a rodent model of locomoter hyper-activity induced by
amphetamine. Labeling of dopaminergic synapses in monkey striatum have found
significant post-synaptic labeling for mGluR5 in terminals of nigrostriatal axons
(Paquet et al., 2003), while anterograde labeling determined these receptors originate
from presynaptic mGluR5 located in the primary motor cortex and other cortical
regions.

18
1.4.3 PET studies of dopamine glutamate interactions The modulation of amphetamine induced dopamine release by pretreatment of
glutamatergic drugs has been studied in nonhuman primate and human brain in vivo
with PET (van Berckel et al., 2006; Kegeles et al., 2000). The effects of LY354740
(mGluR group II agonist) on the modulation of amphetamine-induced dopamine
release using PET and [11C]raclopride was studied in baboons under ketamine and
gaseous anesthesia (van Berckel et al., 2006). Acute administration of LY354740
increased the effect of amphetamine on striatal [11C]raclopride binding potential.
Amphetamine (0.5 mg/kg) alone reduced the binding potential by approximately
28%, and after pretreatment with LY354740, the amphetamine-induced reduction in
binding potential was 35%. Although these results demonstrate a greater reduction in
the outcome measure, several factors should be taken into account when interpreting
these results: (1) ketamine was used to initially to immobilize the monkey and
isoflurane was used during the PET measurements, (2) the effects of LY354740 alone
on [11C]raclopride binding was only tested twice and in the same monkey, resulting in
an 13% increase in the binding potential, (3) LY354740 is a group II agonist, which
has been found to have no effect on amphetamine induced locomotor activity
(Cartmell et al., 1999).
Sustained NMDA transmission deficiency, induced by prolonged infusion of
ketamine has been studied to determine if this leads to an increase in amphetamine
induced dopamine release in humans (Kegeles et al., 2000). Amphetamine (0.25
mg/kg) alone was found to decrease [123I]IBZM binding potential by approximately
5%. Ketamine enhanced the amphetamine induced decrease in [123I]IBZM binding
potential to approximately 12%. Acute alteration of NMDA transmission by ketamine
increased amphetamine induced dopamine release in humans. This observation
supports the hypothesis that, in schizophrenia, a deficiency of glutamatergic control of
dopamine activity might underlie the increase in amphetamine induced dopamine
release previously reported in schizophrenic patients (Breier et al., 1997; Laruelle et al.,
1996). Two important limitations should be considered when interpreting these results.
First, acute NMDA transmission deficiency most likely does not simulate the chronic
NMDA transmission deficiency occurring in patients suffering from schizophrenia
(Olney and Farber, 1995). Second, amphetamine induced a rather small change (-5.5%)
in [123I]IBZM binding potential compared to previously published brain imaging
studies using similar paradigms (Laruelle et al., 1999; Kegeles et al., 1999). The small

19
reduction in binding potential after amphetamine administration is within the range of
test-retest reproducibility (5-10%) at baseline for PET radioligands (Volkow et al.,
1993; Kim et al., 2006). Nevertheless, the increase in amphetamine induced dopamine
release induced by ketamine (greater than two-fold) was comparable in magnitude to
the exaggerated response seen in patients with schizophrenia. Due to limitations in
previous brain imaging studies on modulation of amphetamine induced striatal
dopamine release by glutametergic drugs, we performed a PET study to further
understand dopamine glutamate interactions in vivo.

2 AIMS The main objectives of the work presented in this thesis were as follows:
1. Evaluation of (S,S)-[18F]FMeNER-D2 as a potential radiotracer for the norepinephrine transporter in nonhuman primate brain (Papers I and II).
2. To measure neuropharmacological interventions using PET (e.g., competition
between synaptic neurotransmitters and radiolabeled tracers provide a useful tool to estimate changes in synaptic levels of neurotransmitters) (Papers III and IV).
a. To assess and compare the sensitivity of the agonist radioligand
[11C]MNPA and antagonist [11C]raclopride to stimulant-induced dopamine release in nonhuman primate brain (Paper III).
b. To estimate the occupancy of D2 receptors by endogenous dopamine in
rat brain using PET and [11C]MNPA (Paper IV). 3. PET determination of drug induced receptor occupancy, examined by the
curvilinear relationship between drug plasma concentration and receptor occupancy over a wide dose-range in nonhuman primate brain (Papers II and V).
a. To determine if atomoxetine (a NET reuptake inhibitor) occupies
norepinephrine transporter in a dose-dependent fashion using (S,S)-[18F]FMeNER-D2 in nonhuman primate brain (Paper II).
b. To determine dopamine D2 and serotonin 5-HT1A receptor occupancy in
nonhuman primate brain after administration of RGH-188 (a novel antipsychotic drug). In addition, the intrinsic activity of RGH-188 was estimated in vivo using a dopamine D2 agonist ([11C]MNPA) and antagonist ([11C]raclopride) radioligand (Paper V).
20

3 MATERIALS AND METHODS Detailed description of the utilized methods is provided in the individual papers. More
general characterization of the methods is presented here.
3.1 SUBJECTS Nonhuman primates
In papers I, II, III and V, Astrid Fagraeus Laboratory, Swedish Institute for
Infectious Disease Control, Solna, Sweden supplied seven (5 male and 2 female)
cynomolgus monkeys (Macaca fascicularis) weighing 2.5 – 4.0 kg. The Animal
Research Ethical Committee of the Northern Stockholm Region approved the studies.
Rodents
In paper IV, male Sprague-Dawley rats (300 - 400 g) were obtained from
Taconic Farms (Germantown, NY, USA). All animal procedures were performed in
accordance with the Guide for Care and Use of Laboratory Animals and approved by
the National Institute of Mental Health Animal Care and Use Committee.
3.2 MATERIALS AND RADIOLIGANDS The precursor and standard for FMeNER-D2, and atomoxetine (brand name
Strattera®) were obtained from Eli Lilly, Indianapolis, USA (Papers I and II). Gedeon
Richter Ltd. (Budapest, Hungary) provided RGH-188 (Paper V). We obtained
chemicals and drugs of analytical grade from various commercial sources. The
solutions were prepared as an intravenous stock-solution batch according to Good
Laboratory Practice standards.
Radiosynthesis of (S,S)-[18F]FMeNER-D2, [11C]raclopride, [11C]MNPA, and
[carbonyl-11C]WAY-100635 were prepared in high specific radioactivity (SA) as
described previously (Schou et al., 2004; Langer et al., 1999; Finnema et al., 2005;
Krasikova et al., 2003).
3.3 POSITRON EMISSION TOMOGRAPHY PET system
Nonhuman primate studies
Whole-body transmission and emission scans were acquired on a Siemens
ECAT EXACT HR, which was run in 2D mode (paper I). The spatial resolution is
about 6.0 mm full width half maximum and the field of view equal to 10.8 cm. All
other nonhuman primate studies measured radioactivity in brain with the Siemens
ECAT Exact HR 47 in 3D-mode (Wienhard et al., 1994). A three ring detector block
architecture gives a 15 cm wide field of view. The transversal resolution in the
21

22
reconstructed image is about 3.8 mm full width at half maximum and an axial
resolution of 3.125 mm. The attenuation correction of the data was obtained with three
rotating 68Ge line sources. Raw PET data were then reconstructed using the standard
filtered back projection consisting of the following parameters: 2-mm Hanning Filter,
scatter correction, a zoom factor of 2.17 and a 128 x 128 matrix size (Wienhard et al.,
1994).
Rodent studies
We measured radioactivity in rat brain (Paper IV) with the small animal PET
scanner (NIH Advanced Technology Laboratory Animal Scanner) with an effective
transaxial field of view of 6.0 cm and an axial field of view of 2.0 cm (Seidel et al.,
2003). PET data were reconstructed into 17 coronal slices by 3D Ordered-Subset
Expectation Maximization algorithm, achieving a 1.6 mm full width at half maximum
resolution at the center (Johnson et al., 2002; Liow et al., 2003). The reconstructed
voxel size was 0.56 x 0.56 x 1.12 mm. No attenuation or scatter corrections were
applied.
3.4 PET EXPERIMENTAL PROCEDURES Nonhuman primate studies
In papers, I, II, III and V anesthesia was induced and maintained by repeated
intramuscular injections of a mixture of ketamine hydrochloride (3.75 mg/kg-1h-1
Ketalar , Pfizer) and xylazine hydrochloride (1.5 mg/kg-1h-1 Rompun Vet., Bayer).
The head was immobilized with a fixation device (Karlsson et al., 1993). Body
temperature was maintained with a forced-air heated air blanket (Bair Hugger-Model
505, Arizant Healthcare Inc, MN) and monitored by a rectal thermometer (Precision
Thermometer, Harvard Apparatus, MA). Cardiac and respiratory rates were measured
every 20 min.
In paper I, whole-body transaxial images were acquired for a total of approx.
4.5 hours from six bed-positions from head to mid-tail. The acquisition sequence for
each frame consisted of the following: starting an emission scan at the first bed position
for the head, moving the bed caudally to the next section, scanning a total of 6 sections
consecutively for the same period of time. Each of the 6 sections was imaged 21 times
with the following sequence of frame acquisitions: 4 x 0.50 min; 6 x 1 min; 5 x 2 min,
6 x 4 min. Emission data for paper II were collected continuously for 240 min
according to a pre-programmed series of 34 frames. The five initial frames were 1 min
each, followed by five scans of 3 min each, five scans of 6 min each and the remaining

23
frames of 10 min. In papers III and V emission data were collected continuously for 93
min according to a pre-programmed series of 20 frames starting immediately after
injection of radioligand. The three initial frames were 1 min, followed by four scans of
3 min, and the remaining frames of 6 min.
Rodent studies
In paper IV, anesthesia was induced and maintained using 1 - 1.5% isoflurane
and 100% O2 inhalation through a noise cone. Radioligand was administered as a bolus
followed by a continuous infusion via penile vein catheter (Intramedic PE-10
polyethylene catheter; Aster Industries, Harmony, PA, USA) using a syringe pump
(Harvard PhD 2000, Harvard Apparatus, Holliston, MA, USA). Emission data were
collected continuously for 90 min, according to a pre-programmed series of six initial
frames of 20 s, followed by five scans of 1 min, four scans of 2 min, three scans of 5
min, three scans of 10 min, and the remaining frames of 20 min. Body temperature in
the anesthetized rats was monitored with a rectal temperature probe and maintained at
36.5-37.5°C with a heating lamp and heating pad.
3.5 EXPERIMENTAL DRUGS AND DOSE SELECTION Atomoxetine is a selective NET reuptake inhibitor, which has been found to be
effective in the treatment of ADHD in both children and adults (Spencer et al., 1998;
Michelson et al., 2001). The aim of paper II was to determine if atomoxetine occupies
NET in a dose-dependent fashion using PET in nonhuman primate brain. Each monkey
participated in a total of four PET measurements: one at baseline and one each after
administration of atomoxetine at three different doses. A prolonged intravenous
infusion design was utilized rather than a bolus injection to avoid high initial
atomoxetine concentrations, to better mimic a human oral absorption profile and to
reach plasma steady state during the PET measurements. Monkey doses were based on
a therapeutic concentration range in adults. As such, an examination of NET occupancy
in monkey at clinically-relevant exposures could be examined during the PET study.
In papers III and IV, pharmacological challenges that either increase or decrease
synaptic concentrations of dopamine are combined with PET imaging of the D2
receptor. Agents that increase synaptic dopamine concentrations like amphetamine
have been shown in human and nonhuman primates to decrease D2 receptor radioligand
binding (Innis et al., 1992; Farde et al., 1992; Volkow et al., 1994; Laruelle et al., 1995;
Carson et al., 1997; review: Laruelle, 2000). While agents that reduce synaptic
dopamine levels (e.g., reserpine or α-methyl-para-tyrosine) decrease competition by the

24
endogenous transmitter and thereby ”unmask” D2 receptors and increase radioligand
binding (Ginovart et al., 1997; Laruelle et al., 1997; Verhoeff et al., 2001).
In paper III, amphetamine was administered intravenously as a 30 s bolus
approximately 20 min prior to radioligand injection at four doses (0.1, 0.2, 0.5, and 1.0
mg/kg). Previous studies utilizing in vivo microdialysis in nonhuman primates have
demonstrated that after administration of 0.1 mg/kg of amphetamine produced an
increase of 100% over basal extracellular dopamine levels, 0.2 mg/kg (increase of 459 -
549%), 0.5 mg/kg (~1300%) and the maximum dose assessed 1.0 mg/kg (1600-2000%)
(Moghaddam et al., 1993; Saunders et al., 1994; Laruelle et al., 1997; Breier et al.,
1997). The maximum increase after amphetamine enhanced extracellular dopamine
levels in striatum by more than 20-fold, which reached a peak 40 min after
administration.
Dopamine depleting agents such as α-methyl-para-tyrosine and reserpine are
typically administered at low doses because of side effects, including hypotension. α-
methyl-para-tyrosine is a competitive and reversible inhibitor of tyrosine hydroxylase
(Mignot et al., 1985; Bennett et al., 1981) and has restricted dose limits in humans due
to excretion of crystals in urine at higher doses (Engleman et al., 1968). Reserpine acts
as an inhibitor of vesicular uptake of catecholamines and causes prolonged depletion
because of its irreversible binding to the vesicular transporter (Ponzio et al., 1984;
Guldberg et al., 1971). In paper IV, dopamine depletion was achieved in rats by
administration of reserpine (5 mg/kg, i.p.) given 24 h prior to the PET measurement.
On the day of the PET experiment, the reserpine-treated rats received two injections of
α-methyl-para-tyrosine (200 mg/kg, i.p.) at 4 and 1 h before radioligand injection. This
dual dopamine depletion paradigm has been found to reduce dopamine tissue
concentrations by 95% in rat striatum (Guo et al., 2003).
In paper V, we determined receptor occupancy of RGH-188, a novel
antipsychotic drug that exhibits high affinity for the D2 and D3 receptors, and moderate
affinity for 5-HT1A receptors. Monkeys participated in three consecutive PET
measurements per experimental day. The first PET measurement was performed at
baseline conditions, the second after a low dose (1 - 5 µg/kg) of RGH-188, and the
third after administration of a high dose (30 - 300 µg/kg). RGH-188 was injected
intravenously over 30 s, 15 min prior to radioligand injection.

25
3.6 IMAGE ANLAYSIS In paper I, estimates of whole-body biodistribution over time of (S,S)-
[18F]FMeNER-D2 were performed on compressed planar images as previously
described (Tipre et al., 2004). The planar images were analyzed with PMOD software
(PMOD technologies Ltd, Adliswil, Switzerland). Regions of interest (ROI) were
drawn on planar images and a single generous sized ROI was drawn over the total
body, urinary bladder, brain, kidneys, liver, vertebra, heart and lungs. The use of
planar images for the data analysis provided conservative estimates of radiation
exposure, since the large regions of interest included overlying tissues
In paper II, mean PET images of (S,S)-[18F]FMeNER-D2 were generated with
the highest radioactivity from cortical and sub-cortical brain structures and the least
contribution from skull. PET measurements of experiments with low, medium and high
doses of atomoxetine were co-registered to baseline measurement using the “co-register
and re-slice function” in SPM2 (Statistical Parametric Mapping, Wellcome Department
of Cognitive Neurology, UK). The mean image of the baseline PET measurement was
transformed into standard anatomical space using the monkey version of the Human
Brain Atlas developed at Karolinska Institute (Roland et al., 1994). A standard template
of ROIs was generated on an average monkey MRI scan and applied to each PET
study. Anatomical regions of interest were delineated for the thalamus, locus
coeruleus, whole brain, caudate, mesencephalon, cingulate gyrus, frontal, parietal,
temporal and occipital cortex.
In papers III-V, parametric images of binding potential (BP) and relative blood
flow (R1) were generated from the original reconstructed PET data using a reference
tissue model (Ichise et al., 2003). The R1 PET image was fused with the BP image
using a tool in PMOD software (PMOD Technologies Ltd, Adliswil, Switzerland).
Anatomical regions of interest were manually defined on the fused image for left and
right striatum and cerebellum for dopamine D2 radioligands; temporal and frontal
cortex (defined as forebrain), raphe nuclei, and cerebellum for the 5HT1A radioligand.
All parametric imaging was performed in PMOD (Mikolajczyk et al., 1998) installed
on a PC workstation.
3.7 CALCULATIONS Dosimetry
Activity in the source organs (not decay-corrected) were expressed as a
percentage of injected dose and plotted versus time (Paper I). The residence times

26
from the monkey were calculated by converting the corresponding human values by
multiplication with a factor to scale organ and body weights (in kg) as
(wm,b/wm,o)(wh,o/wh,b), where wm,b is the monkey body weight, wm,o, is the monkey
organ weight, wh,b, is the human body weight, and wh,o is the human organ weight.
This allometric scaling factor is identical to that using SUV (standard uptake value),
which expresses uptake as (%ID/g organ)* g body weight. Thus, the scaling used in
this paper assumes the SUV in the monkey organ is equal to that in the human organ.
Target organ absorbed radiation doses, effective dose and effective dose equivalent
were calculated by MIRDOSE 3.1 (Loevinger et al., 1991). The dynamic bladder
model, implemented in MIRDOSE 3.1 software, was applied to calculate residence
time of the urinary activity with voiding intervals of 2.4 h, 4.8 h and no urine voiding.
3.8 NEUROPHARMACOLOGICAL INTERVENTIONS In paper III, regional radioactivity was normalized to injected activity and body
weight by use of a standardized uptake value (%SUV) [%SUV = (% injection dose/cm3
brain) x body weight (g)], which normalizes for injected activity and body weight. In
paper IV, regional time-activity curves were normalized to injected activity by use of
%(IA per h) per cm3 = [% injected radioactivity per h/cm3 brain].
The radioactivity in the cerebellum was used as an approximate value for free
and non-specifically bound radioligand concentration in the brain. The time curve for
the ratio of radioactivity was calculated for each ROI to the cerebellum. Specific
binding to D2 receptors in striatum was defined as the difference between the total
radioactivity concentration in striatum and cerebellum.
In paper IV, two approaches were used to calculate the outcome measure: 1.)
An equilibrium analysis was applied to PET data obtained from 17.5 to 90 min. The
averaged values obtained during these time frames for the striatum and cerebellum
uptake were applied using a specific to non-displaceable ratio: BPequil = (striatum –
cerebellum)/cerebellum. Second, BP was also estimated by the reference tissue model
MRTM2 (Ichise et al., 2003).
The change in the binding potential after administration of amphetamine or
dopamine depleting agents was calculated according to the equation:
Change (%) = ((BPpost-drug – BPbaseline)/ BPbaseline)*100
Under depleted conditions by α-methyl-para-tyrosine plus resperine, one can
assume that the total pool (100%) of D2/3 receptors are available for radioligand
binding. The baseline estimation is confounded by competition between endogenous

27
dopamine and the radioligand. By comparing the BPdepleted to the BPbaseline, the
difference in the signal is related to the amount of endogenous dopamine occupying
D2/3 receptors during basal conditions. We can estimate the occupancy of endogenous
dopamine by the following equation: (BPdepleted – BPbaseline)/ BPdepleted)*100
3.9 RECEPTOR OCCUPANCY Paper II and V, regional radioactivity was expressed as a standardized uptake
value (%SUV), which normalizes for injected activity and body weight. To calculate
dopamine and serotonin receptor occupancy, two quantitative methods were applied.
The first approach was the peak ratio method (Farde et al., 1992, Andree et al., 1998,
Nyberg et al., 1996). Briefly, the peak ratio method calculates the ratio of
(Bound)/(Free) between the total radioactivity in the region of interest (Bound) and the
total radioactivity in a reference region (Free) with negligible density of D2 and 5-HT1A
receptors. The radioactivity in the cerebellum was used as an approximate value for
free and non-specifically bound radioligand concentration, bearing in mind the
negligible density of D2 (Hall et al., 1994; Hall et al., 1996; Civelli et al., 1993) and 5-
HT1A receptors in cerebellum (Hall et al., 1997). The ratio B/F obtained at the peak of
specific binding time-activity curve is assumed to represent transient equilibrium
(Farde et al. 1989). To calculate D2 and 5-HT1A receptor occupancy, the striatum, and
forebrain and raphe nuclei were used as the region of interest, respectively. Receptor
occupancy was defined as the percentage reduction in the ratio of B/F after drug
administration as compared to B/F at baseline.
The second method to calculate receptor occupancy was the percent reduction
of BP (estimated by MRTM2) after administration of RGH-188 compared to BP at
baseline:
Receptor occupancy (% reduction in BP)
= ((BPpost-drug – BPbaseline)/ BPbaseline)*100%
A hyperbolic curve fit (one-site binding) of NET occupancy plotted as a
function of steady state N-desmethyl-atomoxetine concentration (ng/mL). Transporter
occupancy (%) was defined by change in BP (MRTM2) after drug administration
compared to at baseline. Points represent values estimated from monkey 1 and 2. The
calculation is represented as: Occupancy (%) = Occmax*[Concdrug]/Kd + [Concdrug] in
which Occmax is the maximal occupancy that can be induced by the drug (was set to
100), [Concdrug] is the Css of N-desmethyl-atomoxetine and Kd expresses the drug
concentration at which 50% transporter occupancy is induced.

28
3.10 STATISTICAL ANALYSIS In paper III, a comparison of baseline [11C]MNPA (a selective D2 agonist) and
[11C]raclopride (an antagonist at D2 and D3 receptors) binding potential were assessed
by a repeated measured analysis of variance, with the radioligand as the repeated
condition (n = 8 for each radioligand). For each radioligand the between monkey
differences at baseline were investigated by a one-way analysis of variance (ANOVA),
with monkey as group factor (n = 8 for each radioligand). In addition, the effect of
amphetamine on the change in binding potential for each radioligand was evaluated by
ANOVA. The change in binding potential for each radioligand was designated as the
dependent variable and the main effect assessed for dose, with an interaction between
the radioligands and ligand*dose. The minimum level of significance was designated as
p < 0.05.
In paper IV, a two-tailed independent t-test was applied to the PET image data
and ex vivo radiometabolite data comparing the control and dopamine depletion groups
with a minimum level of significance designated as p < 0.05. All statistical analyses
were performed using SPSS version 13.

4 RESULTS AND DISCUSSION Whole-Body Biodistribution, Radiation Dosimetry Estimates for the PET
Norepinephrine Transporter Probe (S,S)-[18F]FMeNER-D2 in Non-Human
Primates (PAPER I)
An important safety as well as limiting factor for the clinical usability of a
radioligand is set by the relationship between radiation absorbed doses in different
source organs of the body following the radioactivity dose injected. The amount of
radiation absorbed dose delivered by internal administration radiopharmaceuticals is
based on the fact that radiopharmaceuticals have a certain biological, physical and
effective half-life based on the radionuclide half-lives. Paper I measured the whole-
body biodistribution over time of (S,S)-[18F]FMeNER-D2 and estimated the resulting
radiation exposure to organs of the body. The use of planar images for the data
analysis provided conservative estimates of radiation exposure, since the large regions
of interest included overlying tissues. Absorbed doses were calculated from
cynomolgus monkey biodistribution data. The MIRD scheme was applied as an
accurate determination of the time dependent activity of the target regions of the body.
Figure 6. Whole-body images demonstrating biodistribution of (S,S)-[18F]FMeNER-D2 in monkey at 3, 8, 30 and 110 minutes after radioligand injection.
In planar images, urinary bladder, brain, kidneys, liver, lungs, entire abdomen
(GI tract) and vertebra were visually identified as organs with moderate to high
activity (Figure 6). The peak values of the percent injected dose (%ID) at time after
radioligand injection were calculated for the lungs (26.76% ID at 1.42 min), kidneys
(13.55% ID at 2.18 min), whole brain (5.65% ID at 4.48 min), liver (7.20% ID at 2
min), red bone marrow (5.02% ID at 2.06 min), heart (2.36% ID at 1.42 min) and
29

30
urinary bladder (23% ID at 250 min). The cumulative urinary excretion of (S,S)-
[18F]FMeNER-D2 was 20.8% at 150 min. Human residence times were extrapolated
from the average of two monkeys calculated from whole-body planar images.
Radiation absorbed dose estimates were calculated with MIRDOSE 3.1 computer
program, with urine voiding intervals of 2.4 h, 4.8 h and no voiding. Assuming a
urine voiding interval of 2.4 h, the four organs with highest exposures in µGy/MBq
(mrad/mCi) were: kidneys 126 (468), heart wall 108 (399), lungs 88.4 (327) and
urinary bladder 114 (422). The effective doses were estimated with and without urine
voiding at a range of 123 (33) and to 131 (35.5) mrad/mCi (µGy/MBq).
This study indicated that (S,S)-[18F]FMeNER-D2 may be a suitable
radioligand for studying NET in brain. The calculated dosimetry results seem
comparable with those for other 18F labeled brain imaging agents. These results in
primates can be used to estimate the limit of radioactivity that can be administered at
a low risk to human subjects. Nevertheless, a human biodistribution study would
provide more accurate estimation of organ radiation absorbed doses.
Atomoxetine Occupies the Norepinephrine Transporter in a Dose-Dependent
Fashion: A PET Study in Non-Human Primate Brain Using (S,S)-[18F]FMeNER-
D2 (PAPER II)
PET can be used as a tool in drug development in either a direct or indirect
manner to assess the in vivo activities of a drug (Farde et al., 1996; Talbot et al., 2002;
Brooks et al., 2005). A direct approach is to radiolabel a potential new drug with 11C or 18F and trace its uptake, anatomical distribution, and binding in brain. An indirect
approach measures the degree to which administration of the new drug competes with
the specific binding of a conventional PET radioligand. Thus, PET studies can confirm
brain distribution, and drug binding selectivity to various subtypes of receptors.
Furthermore, correlation between plasma drug concentration and receptor occupancy
may provide valuable data to establish clinical dose levels (Halldin et al., 2001b; Farde
and Lee 2006; Farde et al., 1996; Talbot et al., 2002; Brooks et al., 2005).
The aim of paper II was to determine if atomoxetine occupies NET in a dose-
dependent fashion in nonhuman primate brain. In this PET study, we used (S,S)
[18F]FMeNER-D2 to measure NET occupancy of atomoxetine in the locus coeruleus,
thalamus, mesencephalon and cingulated gyrus. PET data was analyzed to define
relationships between doses, plasma concentrations and NET occupancy for
atomoxetine. Doses were based on the therapeutic concentration range of atomoxetine

31
in humans. As such, the transporter occupancy data may be informative to guide dosing
strategies in drug development or in determining the occupancy level necessary to
induce clinical effects.
During baseline conditions, (S,S)-[18F]FMeNER-D2 uptake was highest in NET
rich regions such as the locus coeruleus, thalamus, mesencephalon and the cingulate
gyrus, whereas the radioactivity in the
caudate was low. Quantification of
cortical uptake of (S,S)-[18F]FMeNER-D2
was difficult due to defluorination
resulting in skull-bound radioactivity that
increased cortical uptake over time. After
administration of low, medium and high
doses of atomoxetine, there was a dose-
dependent reduction of the binding
potential estimated by a reference tissue
model. The relationship between N-
desmethyl-atomoxetine concentration and NET occupancy in the locus coeruleus was
described by a hyperbolic function (Figure 7), resulting in 51% at the lowest dose (0.03
mg/kg/h) to 91% at the highest dose (0.12 mg/kg/h).
0 20 40 600
20
40
60
80
100
Locus coeruleus
N-desmethylatomoxetine (ng/mL)
NET
occ
upan
cy (%
)
80
Figure 7. A hyperbolic curve fit (one-site binding) of NET occupancy plotted as a function of steady-state N- desmethylatomoxetine
This study demonstrated that steady-state infusion of atomoxetine occupies
NET in non-human primate brain in a dose-dependent fashion ranging from 38 to 82
%. NET occupancy using the longer half-life of (S,S)-[18F]FMeNER-D2 compared to
[11C]MeNER allows a more realiable determination of NET occupancy since transient
specific binding equilibrium is attained during the time frame of the PET measurement.
This is the first in vivo PET study to successfully demonstrate the ability to measure a
dose-dependent change in NET occupancy in brain using (S,S)-[18F]FMeNER-D2.
Effect of Amphetamine on Dopamine D2 Receptor Binding in Non-Human
Primate Brain: A Comparison of the Agonist Ligand [11C]MNPA and Antagonist
Radioligand [11C]Raclopride (PAPER III)
Pharmacological challenges that increase synaptic concentrations of dopamine
are combined with PET imaging of the D2 receptor using tracer doses of the
radioligand. For example, agents that increase synaptic dopamine concentrations like
amphetamine or methylphenidate have been shown in human and nonhuman primates
to decrease D2 receptor radioligand binding. These endogenous competition studies
have typically been performed with antagonist radioligands, such as [11C]raclopride.

32
Antagonists of G-protein coupled receptors, such as the D2 receptor, have equal affinity
for receptors in the high (i.e., coupled) and low (i.e., uncoupled) affinity states. In
contrast, agonists bind preferentially to the high affinity state. Since dopamine is the
endogenous agonist for the D2 receptor, it would more effectively compete with the
binding of an agonist, compared to an antagonist radioligand. The aim of paper III was
to examine whether the striatal uptake of the agonist radioligand [11C]MNPA is more
sensitive than the antagonist [11C]raclopride to stimulant-induced dopamine release.
Cynomolgus monkeys were examined before and after increasing doses of intravenous
amphetamine. Finally, the data were used in an attempt to estimate the proportion of D2
receptors in the high and low affinity states in the anesthetized nonhuman primate.
Following injection of [11C]MNPA in the baseline condition, activity
concentrated in striatum, with lower levels in the thalamus that were still higher than
the nonspecific uptake in cerebellum. Under baseline conditions, [11C]raclopride BP
(5.76 ± 0.95, n = 8) was significantly higher than that for [11C]MNPA BP (1.31 ± 0.21,
n = 8). When comparing the difference between the four monkeys BP values for each
radioligand, no significant between subject effects were found for [11C]MNPA or
[11C]raclopride.
The effect of amphetamine on the binding of [11C]MNPA and [11C]raclopride
was investigated by administering amphetamine intravenously (0.1, 0.2, 0.5 and 1.0
mg/kg) approximately 20 min prior to injection of radioligand. Amphetamine caused a
reduction in [11C]MNPA binding potential of 4% at 0.1, 23% at 0.2, 25% at 0.5 and
46% at 1.0 mg/kg. [11C]Raclopride binding potential was reduced to a lesser extent by
2% at 0.1, 16% at 0.2, 15% at 0.5 and 23% at 1.0 mg/kg of amphetamine (Figure 8). A
statistically significant greater change in BP for [11C]MNPA compared to
[11C]raclopride was observed (P = 0.024; ANOVA).
Based on calculations proposed by Narendran et al. (2004) and the assumption
that 10% of receptors are occupied by endogenous dopamine at baseline, our results
suggest that 61% of the D2 receptors are configured in the high affinity state. This
calculation is based upon the assumption that 10% of the receptors are occupied by
dopamine at baseline and that extrasynaptic receptors cannot be displaced by stimulant-
induced dopamine release. This 61% in the high affinity state is distributed as: 10%
occupied by dopamine at baseline, 23% are synaptic and 28% are extra-synaptic. By
this calculation the remaining receptors (39%) would be in the low affinity state (ie.,
39% = 100% - 61% in the high affinity state).

33
Paper III provides evidence that the agonist radioligand [11C]MNPA is more
sensitive than the antagonist [11C]raclopride to displacement by endogenous dopamine.
At all doses examined, the change in [11C]MNPA BP was approximately 50% more
sensitive than [11C]raclopride to pharmacological induced increases in synaptic
dopamine. These results agree with those of Cumming et al. (2002) and Narendran et
al. (2004), who similarly investigated the effect of amphetamine on the binding of
agonist and antagonist radioligands in mice and non-human primates.
Figure 8. Parametric images of [11C]raclopride and [11C]MNPA BP estimated by a reference tissue model at baseline and postamphetamine conditions. Images represent the same monkey for each dose of amphetamine.
The current results can been interpreted based on the following assumptions: 1)
with the understanding that [11C]raclopride binds to a greater number of D2 receptors
than [11C]MNPA, because the antagonist binds to both the high and low affinity states
of the receptor. In fact, there may be other reasons for differences in Bmax′ (available
receptor density) between these two tracers. For example, the ligands may differ in
affinity for the monomer and dimer forms of the receptor. 2) effects of anesthesia
should be taken into account, a mixture of ketamine and xylazine was administered
every 40 min at baseline and during amphetamine challenge conditions. It cannot be
ruled out that ketamine has some effect on [11C]MNPA and [11C]raclopride binding at
both baseline and pretreatment conditions. 3) the greater reduction in BP after
amphetamine administration for [11C]MNPA compared to [11C]raclopride may also be
secondary to agonist induced receptor internalization. The decreased uptake of
radioligand may be the result of the loss of available binding sites by internalization

34
and a subsequent loss of appropriate three dimensional binding to antagonists as well as
agonists. In summary, [11C]MNPA is a promising radioligand for PET-imaging of the
high affinity state of D2 receptors in the primate brain. Stimulation of dopamine release
with amphetamine caused a marked decreases in BP of [11C]MNPA in a dose-
dependent fashion, thus demonstrating that the radioligand is sensitive to the
endogenous dopamine concentration.
Imaging Endogenous Dopamine Occupancy of the D2 Receptor in Rat Brain With
[11C]MNPA (PAPER IV)
Endogenous dopamine levels during basal conditions have been estimated using
PET and SPECT in human and nonhuman primate brain. Dopamine depletion induced
by reserpine and/or α-methyl-para-tyrosine decreases competition by endogenous
dopamine to the D2 receptor and increases radioligand binding. The increase in binding
compared to baseline measurements provides an estimate of the number of D2 receptors
occupied by dopamine under basal conditions. Occupancy of the D2 receptor by
endogenous dopamine in brain using antagonist radioligands has ranged from
approximately 10% to 30%. The aim of paper IV was to estimate the occupancy of D2
receptors by endogenous dopamine in rat brain in vivo using PET and the agonist
radioligand [11C]MNPA. In addition, ex vivo radiometabolite studies were performed to
determine if the increase in radioligand binding after dopamine depletion was due to a
change in radiometabolites entering brain.
Following injection of [11C]MNPA in the baseline condition, activity
concentrated in striatum, with lower levels in cerebellum. [11C]MNPA baseline BP
values were similar between the outcome
measures BPequil and MRTM2, 1.04 ± 0.14
and 1.08 ± 0.12, respectively.
Nonradioactive raclopride administered
during steady state, 2 and 4 mg/kg, induced
a significant displacement in striatal activity
by ~ 60% and ~ 80%, respectively. The
specific binding of [11C]MNPA was nearly
completely inhibited by this D2/3-antagonist,
consistent with the radioligand also binding
to D2/3 receptors. Depletion of endogenous
dopamine with reserpine and α-methyl-
Control Depletion0.5
1.0
1.5
2.0
[11C
]MN
PA B
P
Figure 9. [11C]MNPA BP measured at baseline and after dopamine depletion. Dopamine depletion resulted in a significant increase in striatal [11C]MNPA binding (two-tailed p = 0.0005).

35
para-tyrosine significantly increased activity in striatum but had little effect in
cerebellum. Under baseline conditions [11C]MNPA BP was 1.08 ± 0.12 (n = 5) and
after dopamine depletion resulted in a significant increase of 38% in BP 1.49 ± 0.12 ((n
= 5); p = 0.0005), Figure 9. Assuming a complete depletion of endogenous dopamine in
rat striatum following reserpine and α-methyl-para-tyrosine administration, these
results indicate a baseline D2/3 high affinity state receptor occupancy by endogenous
dopamine of ~ 27%, calculated by comparing the BP during baseline conditions and
after acute dopamine depletion. The effect of dopamine depletion was also
demonstrated by ex vivo studies where the brain to plasma parent concentration ratios
in the depleted rats was higher than that in control rats by 30%. Brain radioactivity
composition of radiometabolite in control condition was not significantly different after
dopamine depletion. Thus, the increase in radioligand binding after dopamine depletion
is not attributed to an increase in radiometabolite binding. Both NPA and MNPA had
high binding affinities for D2 receptors (Ki = 4.7 and 9.4 nM, respectively) (Paper IV,
unpublished data). PHNO binding affinity was highest to D3 receptors (Ki = 0.53),
consistent with previous reports (Parker et al., 2006; Freedman et al., 1994). Thus, NPA
and MNPA have similar binding affinities for D2/D3 receptors, while PHNO bound
preferentially to D3 receptors.
Our PET imaging in rats confirm prior studies in monkeys (Finnema et al.,
2005; Seneca et al., 2006) that [11C]MNPA is a useful agonist radioligand for D2/3
receptors. One potential reason for increased activity in striatum after dopamine
depletion can be attributed to an increase of radiometabolites entering brain.
Nevertheless, our ex vivo experiments under control and dopamine depletion conditions
found no significant difference in radiometabolite composition or parent radioactivity
in whole brain samples. Instead, the increase in striatal binding could have been caused
by at least four factors: 1) an increase in the number of D2 receptors, 2) trafficking of
the receptor to the surface membrane, assuming that the internalized receptor does not
bind MNPA, 3) an increase in the percentage of the D2 receptors in the high affinity
state, 4) removal of competition by endogenous dopamine. Several homogenate
binding studies have found no increase in the density of D2 receptors after acute
dopamine depletion (Laruelle et al., 1997; Ross et al., 1989). In agreement with the
homogenate binding studies, in vivo Scatchard analysis using [11C]raclopride binding
reflected an increase in receptor affinity but no change in D2 receptor density in
nonhuman primate brain (Ginovart et al., 1997). Thus, the first possibility (i.e., an

36
increase in the number of D2 receptors) seems unlikely. Nevertheless, the remaining
three mechanisms are plausible and cannot be excluded by our results. In conclusion,
we estimate that dopamine occupies ∼ 27% of high affinity D2/3 receptors in brains of
rats under gaseous anesthesia.
Occupancy of Dopamine D2 and Serotonin 5-HT1A Receptors by a Novel D2/D3
Antipsychotic Drug in Non-Human Primate Brain: A PET Study (PAPER V)
RGH-188 is a novel antipsychotic drug that binds with high selectivity and
affinity to D2 and D3 and moderate affinity to 5-HT1A receptors. The aim of paper V
was to determine if RGH-188 occupies dopamine D2 and serotonin 5-HT1A receptors
in nonhuman primate brain. We used both an antagonist [11C]raclopride and agonist
[11C]MNPA radioligands to measure D2 receptor occupancy in striatum and
[carbonyl-11C]WAY-100635 to measure 5HT1A receptor occupancy in raphe nuclei
and forebrain. Following the injection of [11C]raclopride or [11C]MNPA, striatum
showed the highest activity, with lower levels in cerebellum. RGH-188 occupied D2
receptors in a dose dependent and saturable manner, with lowest dose (1 µg/kg)
occupying ~5% of receptors, and highest dose (300 µg/kg) more than 90% (Figure
10A). A comparison of antagonist/agonist radioligand ratio of receptor occupancy
may provide an in vivo estimation of intrinsic activity of RGH-188. RGH-188 was
equally potent to displace both [11C]raclopride and [11C]MNPA with an ED50 of ~5
µg/kg i.v., which suggests that RGH-188 is an antagonist.
A B.
0 50 100 150 200 250 300 3500
20
40
60
80
100
Binding PotentialPeak Ratio
RGH-188 (µg/kg)
5-H
T 1A o
ccup
ancy
(%)
0 50 100 150 200 250 300 3500
20
40
60
80
100
[11C]Raclopride (Binding Potential)[11C]Raclopride (Peak Ratio)
EPS
[11C]MNPA (Binding Potential)[11C]MNPA (Peak Ratio)
RGH-188 (µg/kg)
2D
occ
upan
cy (%
)
Figure 10. A hyperbolic curve fit of dopamine D2 (A) and serotonin 5-HT1A (B) receptor occupancy plotted as a function of RGH-188 dose (µg/kg). Receptor occupancy (%) was estimated by two approaches: the percent reduction of B/F (Peak ratio method: shown as triangles with dotted line) and change in BP estimated by a reference tissue model (shown as circles with solid line).

37
After administration of [11C]WAY-100635, activity concentrated in forebrain
and raphe nuclei, with lower levels in cerebellum. Administratrion of 30 µg/kg of
RGH-188 slightly reduced activity in raphe nuclei compared to baseline and had no
effect in forebrain. 5-HT1A receptor occupancy could be described by a hyperbolic
funtion and was much lower compared to D2 occupancy at the same doses, with a
maximal value of ~30% for the raphe nuclei (Figure 10B).
In conclusion, RGH-188 occupies dopamine D2 receptors in a dose-dependent
and saturable manner and has much lower occupancy of serotonin 5-HT1A receptors.
The finding of a relatively low maximal 5-HT1A receptor occupancy suggests that
RGH-188 will have minimal pharmacological actions via this target. These PET data
demonstrated an ED50 value of 5 µg/kg at the D2 receptor and a 5 times greater dose
induced only 11-18% occupancy at the 5-HT1A receptor. RGH-188 primary CNS
actions are related to the D2 and D3 receptor, with minimal 5-HT1A related effects at
therapeutic dose levels. Imaging studies using both dopamine D2 receptor agonist and
antagonist radioligands may be used to define the affinity of a drug to the low and
high affinity states of a receptor and thereby provide an in vivo estimate of intrinsic
activity. RGH-188 had similar occupancy measured with both an agonist
([11C]MNPA) and an antagonist ([11C]raclopride) radioligand, confirming in vitro
studies that RGH-188 is an antagonist.
Interactions between Dopamine and Glutamate: A PET study in rat and non-
human primate brain to elucidate the enhancement of endogenous dopamine with
amphetamine by Group I mGluR antagonist, MPEP. (Unpublished work
performed by Seneca together with the PET imaging facility at NIRS, Chiba,
Japan. Manuscript is under preparation by Seneca, Maeda, Tokunaga, Higuchi,
Innis, Halldin, and Suhara).
The aim of this study was to explore the inhibition of methamphetamine
induced dopamine release by pretreatment with an mGluR group I antagonist, MPEP,
using the D2 agonist radioligand, [11C]MNPA. In awake conditions, cynomolgus
monkeys (n = 4) and rats (n = 3) were examined with [11C]MNPA at baseline (test-
retest conditions), following MPEP pretreatment, after methamphetamine
administration and MPEP plus methamphetamine conditions. Additionally,
microdialysis was applied to rat in order to quantify extracellular dopamine content in
the drug challenge conditions identical to those in the PET study.

38
[11C]MNPA baseline binding potential values were similar between monkey
and rat, 1.00 ± 0.05 and 0.82 ± 0.06, respectively. The test–retest binding potential
variability was relatively small, ranging from 3% to 6%. MPEP alone did not
significantly change the D2 receptor availability for the radioligand. Methamphetamine
caused a reduction in [11C]MNPA binding potential by 17 ± 6% and 25 ± 8% in
monkeys and rats, respectively. Meanwhile, pretreatment with MPEP reversed
methamphetamine induced reduction of striatal [11C]MNPA binding to baseline levels
(Figure 11A). The extracellular dopamine concentration in rat striatum was increased
by approximately 10 fold at 10 min after methamphetamine administration. MPEP
pretreatment nearly halved the impact of methamphetamine administration on the
dopamine contents (Figure 11B). This observation is in line with the PET data,
substantiating the interference of MPEP with modulation of dopamine release by
pharmacological increase in extracellular dopamine release.
0 6TestRetest
MPEP MAP
MPEP+MAP60
80
100
120
RatMonkey
BP
Cha
nge
(% B
asel
ine)
-20 -10 0 10 20 30 40 50 60 70 80 90 1000
200400600800
10001200140016001800
MAPMPEP+MAP
MPEP (1.0 mg/kg i.v.)
MAP (0.5 mg/kg i.v.)
Time (min)
DA
rele
ase
(% o
f bas
elin
e)
A. B.
Figure 11. Change in striatal BP of [11C]MNPA in conscious monkey and rat during modulation of dopamine release (A). Changes in extracellular dopamine levels in rat striatum during microdialysis (B). (MAP methamphetamine; MPEP mGluR group I antagonist).
In conclusion, these results provide evidence for the modulation of striatal
dopamine release by Glutamate-Dopamine interactions across species. The mechanistic
view that MPEP inhibits mGluR group I located on dopaminergic neurons and
suppressively acts on stimulation of dopamine release from the striatal terminals
without altering the baseline state of dopamine transmission.

5 SUMMARY OF FINDINGS The present thesis focused on examining two monoaminergic neurotransmitter
systems in rodent and nonhuman primate brain.
The estimated radiation burden of (S,S)-[18F]FMeNER-D2 is comparable to
that of other 18F radioligands. These results in primates can be used to
estimate the limit of radioactivity that can be administered at a low risk to
human subjects.
Peak equilibrium measurements were achieved using (S,S)-[18F]FMeNER-D2
in contrast to the previously reported data for (S,S)-[11C]MeNER. After
administration of atomoxetine, a dose-dependent occupancy from 38 to 82%
was observed for brain regions known to contain high densities of NET.
The agonist radioligand [11C]MNPA, was found to be more sensitive than the
antagonist [11C]raclopride to displacement by endogenous dopamine. Since an
antagonist radioligand binds with equal affinity to both high- and low-affinity
states, a radiolabeled agonist would be advantageous since it only binds to
receptors in the high- affinity state, likely leading to more sensitivity to
endogenous dopamine.
PET imaging in rats confirm prior studies in monkeys that [11C]MNPA is a
useful agonist radioligand for D2/3 receptors. We estimate that dopamine
occupies ∼ 27% of high affinity D2/3 receptors in brains of rats under gaseous
anesthesia.
The administration of increasing doses of RGH-188 (a novel antipsychotic
drug) resulted in a dose-dependent and saturable reduction in the specific
binding of [11C]raclopride and [11C]MNPA. Administration of RGH-188 at 30
µg/kg induced a higher striatal D2 receptor occupancy of ∼ 87% compared to
only 21% for 5-HT1A receptor. RGH-188 had similar occupancy measured
with both an agonist and an antagonist radioligand, confirming in vitro studies
that RGH-188 is an antagonist.
39

6 FUTURE PROSPECTS Future prospects for NET radioligands: (S,S)-[18F]FMeNER-D2 has the
potential to be a useful radioligand for quantitative imaging of NET in brain.
Nevertheless, a radioligand with a higher signal to noise ratio and lower non specific
binding would be advantageous. NET radioligands have more challenges than other
transporter radioligands such as the dopamine or serotonin transporter. A high basal
occupancy of NET by norepinephrine and other endogenous monoamines such as
serotonin and dopamine may compete for the same binding site as the radioligand. A
future study using a reserpine depletion paradigm (depletes not only dopamine
concentrations, but norepinephrine) can be performed to determine the occupancy of
NET by endogenous monoamines. New radioligands with higher affinity for NET
may help improve the signal since sparse densities of NET in brain compared to other
monoamine transporters. The use of a higher resolution PET camera may help
delineate smaller brain regions such as the caudate for the reference region and
regions known to contain high densities of NET (e.g. the locus coeruleus).
Future prospects with pharmacological challenges: Based on the results from
paper III and IV that [11C]MNPA was more sensitive to changes in endogenous
dopamine than antagonist radioligands, the findings should be confirmed in healthy
subjects and patients suffering from altered dopamine neurotransmission.
Future prospects with receptor occupancy studies: PET studies employing dual
agonist and antagonist radioligands may lead to a novel approach to further understand
the in vivo mechanisms of action of antipsychotic drugs. The data provided in paper V
should be expanded in both animal and human studies to determine if one can assess
the intrinsic activity of a drug. Studies employing, antagonists, partial agonists and
agonists would help lead further support into the notion that PET imaging using dual
radioligands can determine intrinsic activity.
40

7 ACKNOWLEDGEMENTS I would like to express my sincere gratitude and appreciation to everyone who has contributed to this work. Especially I would like to thank: Prof. Christer Halldin, my KI mentor, for his optimism, endless enthusiasm and his keen eye for developing new radioligands. Dr. Robert B. Innis, my NIH mentor, for accepting and supporting me as a NIH/KI student in the graduate partnership program, and for teaching me how to be precise in scientific thinking, studies and writing. Prof. Lars Farde, for teaching me that too few words is much better than too many. So I will keep it short!! National Institute of Radiological Sciences (NIRS) in Chiba, Japan. I wanted to thank Dr. Tetsuya Suhara for accepting the proposed study and allowing me the opportunity to perform PET studies at NIRS. Drs. Tokunaga and Maeda for being gracious hosts and for making my stay fruitful and enlightening. PET imaging group at Karolinska Institute, Dr. Bengt Andree for making my transition to Sweden and KI PET group as smooth as possible, Dr. Per Karlsson for his discussions about sports and PET imaging, Kjerstin Lind for making the workplace feel like home during the holiday season, Dr. Andrea Varrone for his friendship and valuable insight into data analysis, Julio Gabriel for reconstructing the PET data as quickly as possible! Nils Sjoholm for making sure the PET camera is always working and Dr. Stefan Pauli for his volume rendering images. Jacqueline Borg, Drs. Judit Sóvágó, Zsolt Cselenyi, Johan Lundberg and Simon Cervenka for many interesting discussions about research and life in general. The members of the PET radiochemistry group at Karolinska, for what seems like a 99% success track record on radioligand synthesis, and endless commitment to producing the highest specific activity radiotracers. I appreciated their help and collaboration: Magnus Schou, Anu Airaksinen, Phong Truong, Arsalan Amir, and Jan Andersson. Assoc. Prof. Balázs Gulyás – For all of your help with the monkey PET experiments, teaching me how to handle and work with nonhuman primates, and fruitful discussions about life and science. My office-mate, Sjoerd Finnema, fellow student and friend, the many hours, no years of working together to figure out the intricacies of PET imaging, no stone should be left unturned. Ulla-Kajsa Pehrsson and Karin Zahir – for valuable Swedish translation and helping with the many administrative issues that have arisen over the years.
41

42
Thanks to all the members of the radiochemistry and PET imaging group at NIMH. PET imaging group at the molecular imaging branch (NIMH/NIH), Dr. Jeih-San Liow for his help with microPET studies and for his friendship, Garth Terry for his assistance during the microPET studies, and Dr. Sami Zoghbi for performing the radiometabolite analysis and for his attention to detail. My family and friends – For never giving up hope and always being there when I needed them! Zoya, for your love and endless support, and for the great life we have together.
The National Institutes of Health and Karonlinska Institutet for financial support.

8 REFERENCES Andree B, Nyberg S, Ito H, Ginovart N, Brunner F, Jaquet F, Halldin C, Farde L.
1998. Positron emission tomographic analysis of dose-dependent MDL 100,907 binding to 5-hydroxytryptamine-2A receptors in the human brain. J Clin Psychopharmacol 18(4):317-323.
Andree B, Seneca N, Schou M, Mozley PD, Potter WZ, Farde L, Gulyas B, Halldin C. 2004. Regional central norepinephrine transporter occupancy induced by reboxetine determined in man using [11C]MeNER. Neuroimage 22 Suppl 2:30.
Bennett BA, Sundberg DK. 1981. Catecholamine biosynthesis in specific brain areas of the rat as determined by liquid chromatography and amperometric detection. Life Sci 28(25):2811-2817.
Berton O, Nestler EJ. 2006. New approaches to antidepressant drug discovery: beyond monoamines. Nat Rev Neurosci 7(2):137-151.
Breier A, Su TP, Saunders R, Carson RE, Kolachana BS, de Bartolomeis A, Weinberger DR, Weisenfeld N, Malhotra AK, Eckelman WC, Pickar D. 1997. Schizophrenia is associated with elevated amphetamine-induced synaptic dopamine concentrations: evidence from a novel positron emission tomography method. Proc Natl Acad Sci U S A 94(6):2569-2574.
Brooks DJ. 2005. Positron emission tomography and single-photon emission computed tomography in central nervous system drug development. NeuroRx 2(2):226-236.
Carlsson A, Hansson LO, Waters N, Carlsson ML. 1999. A glutamatergic deficiency model of schizophrenia. Br J Psychiatry Suppl(37):2-6.
Carlsson A, Lindqvist M. 1963. Effect of Chlorpromazine or Haloperidol on Formation of 3methoxytyramine and Normetanephrine in Mouse Brain. Acta Pharmacol Toxicol (Copenh) 20:140-144.
Carlsson A, Waters N, Waters S, Carlsson ML. 2000. Network interactions in schizophrenia - therapeutic implications. Brain Res Brain Res Rev 31(2-3):342-349.
Carson RE, Breier A, de Bartolomeis A, Saunders RC, Su TP, Schmall B, Der MG, Pickar D, Eckelman WC. 1997. Quantification of amphetamine-induced changes in [11C]raclopride binding with continuous infusion. J Cereb Blood Flow Metab 17(4):437-447.
Casey DE. 1989. Clozapine: neuroleptic-induced EPS and tardive dyskinesia. Psychopharmacology (Berl) 99 Suppl:S47-53.
Cartmell J, Monn JA, Schoepp DD. 1999. The metabotropic glutamate 2/3 receptor agonists LY354740 and LY379268 selectively attenuate phencyclidine versus d-amphetamine motor behaviors in rats. J Pharmacol Exp Ther 291(1):161-170.
Cherry SR. 2001. Fundamentals of positron emission tomography and applications in preclinical drug development. J Clin Pharmacol 41(5):482-491.
Cherry SR, Gambhir SS. 2001. Use of positron emission tomography in animal research. Ilar J 42(3):219-232.
Chinta SJ, Andersen JK. 2005. Dopaminergic neurons. Int J Biochem Cell Biol 37(5):942-946.
43

44
Civelli O, Bunzow JR, Grandy DK. 1993. Molecular diversity of the dopamine receptors. Annu Rev Pharmacol Toxicol 33:281-307.
Conn PJ, Pin JP. 1997. Pharmacology and functions of metabotropic glutamate receptors. Annu Rev Pharmacol Toxicol 37:205-237.
Creese I, Burt DR, Snyder SH. 1976. Dopamine receptor binding predicts clinical and pharmacological potencies of antischizophrenic drugs. Science 192(4238):481-483.
Creese I, Sibley DR, Leff SE. 1984. Agonist interactions with dopamine receptors: focus on radioligand-binding studies. Fed Proc 43(13):2779-2784.
Cumming P, Wong DF, Dannals RF, Gillings N, Hilton J, Scheffel U, Gjedde A. 2002. The competition between endogenous dopamine and radioligands for specific binding to dopamine receptors. Ann N Y Acad Sci 965:440-450.
Daggett LP, Sacaan AI, Akong M, Rao SP, Hess SD, Liaw C, Urrutia A, Jachec C, Ellis SB, Dreessen J, et al. 1995. Molecular and functional characterization of recombinant human metabotropic glutamate receptor subtype 5. Neuropharmacology 34(8):871-886.
Ding YS, Lin KS, Garza V, Carter P, Alexoff D, Logan J, Shea C, Xu Y, King P. 2003. Evaluation of a new norepinephrine transporter PET ligand in baboons, both in brain and peripheral organs. Synapse 50(4):345-352.
Engelman K, Horwitz D, Jequier E, Sjoerdsma A. 1968. Biochemical and pharmacologic effects of alpha-methyltyrosine in man. J Clin Invest 47(3):577-594.
Farde L, Hall H, Ehrin E, Sedvall G. 1986. Quantitative analysis of D2 dopamine receptor binding in the living human brain by PET. Science 231(4735):258-61.
Farde L, Wiesel FA, Halldin C, Sedvall G. 1988. Central D2-dopamine receptor occupancy in schizophrenic patients treated with antipsychotic drugs. Arch Gen Psychiatry 45(1):71-76.
Farde L, Eriksson L, Blomquist G, Halldin C. 1989. Kinetic analysis of central [11C]raclopride binding to D2-dopamine receptors studied by PET--a comparison to the equilibrium analysis. J Cereb Blood Flow Metab 9(5):696-708.
Farde L, Wiesel FA, Stone-Elander S, Halldin C, Nordstrom AL, Hall H, Sedvall G. 1990. D2 dopamine receptors in neuroleptic-naive schizophrenic patients. A positron emission tomography study with [11C]raclopride. Arch Gen Psychiatry 47(3):213-219.
Farde L, Hall H. 1992. Positron emission tomography--examination of chemical transmission in the living human brain. Development of radioligands. Arzneimittelforschung 42(2A):260-264.
Farde L. 1996. The advantage of using positron emission tomography in drug research. Trends Neurosci 19(6):211-214.
Finnema SJ, Seneca N, Farde L, Shchukin E, Sovago J, Gulyas B, Wikstrom HV, Innis RB, Neumeyer JL, Halldin C. 2005. A preliminary PET evaluation of the new dopamine D2 receptor agonist [11C]MNPA in cynomolgus monkey. Nucl Med Biol 32(4):353-360.
Freedman SB, Patel S, Marwood R, Emms F, Seabrook GR, Knowles MR, McAllister G. 1994. Expression and pharmacological characterization of the human D3 dopamine receptor. J Pharmacol Exp Ther 268: 417-26.

45
George SR, Watanabe M, Seeman P. 1985. Dopamine D2 receptors in the anterior pituitary: a single population without reciprocal antagonist/agonist states. J Neurochem 44(4):1168-1177.
Ginovart N, Farde L, Halldin C, Swahn CG. 1997. Effect of reserpine-induced depletion of synaptic dopamine on [11C]raclopride binding to D2-dopamine receptors in the monkey brain. Synapse 25(4):321-325.
Ginovart N, Galineau L, Willeit M, Mizrahi R, Bloomfield PM, Seeman P, Houle S, Kapur S, Wilson AA. 2006. Binding characteristics and sensitivity to endogenous dopamine of [11C]-(+)-PHNO, a new agonist radiotracer for imaging the high-affinity state of D2 receptors in vivo using positron emission tomography. J Neurochem 97(4):1089-1103.
Girault JA, Greengard P. 2004. The neurobiology of dopamine signaling. Arch Neurol 61(5):641-644.
Golembiowska K, Konieczny J, Wolfarth S, Ossowska K. 2003. Neuroprotective action of MPEP, a selective mGluR5 antagonist, in methamphetamine-induced dopaminergic neurotoxicity is associated with a decrease in dopamine outflow and inhibition of hyperthermia in rats. Neuropharmacology 45(4):484-492.
Guldberg HC, Broch OJ, Jr. 1971. On the mode of action of reserpine on dopamine metabolism in the rat striatum. Eur J Pharmacol 13(2):155-167.
Guo N, Hwang DR, Lo ES, Huang YY, Laruelle M, Abi-Dargham A. 2003. Dopamine depletion and in vivo binding of PET D1 receptor radioligands: implications for imaging studies in schizophrenia. Neuropsychopharmacology 28(9):1703-1711.
Haka MS, Kilbourn MR. 1989. Synthesis and regional mouse brain distribution of [11C]nisoxetine, a norepinephrine uptake inhibitor. Int J Rad Appl Instrum B 16(8):771-774.
Hall H, Farde L, Halldin C, Hurd YL, Pauli S, Sedvall G. 1996. Autoradiographic localization of extrastriatal D2-dopamine receptors in the human brain using [125I]epidepride. Synapse 23(2):115-123.
Hall H, Lundkvist C, Halldin C, Farde L, Pike VW, McCarron JA, Fletcher A, Cliffe IA, Barf T, Wikstrom H, Sedvall G. 1997. Autoradiographic localization of 5-HT1A receptors in the post-mortem human brain using [3H]WAY-100635 and [11C]way-100635. Brain Res 745(1-2):96-108.
Hall H, Sedvall G, Magnusson O, Kopp J, Halldin C, Farde L. 1994. Distribution of D1- and D2-dopamine receptors, and dopamine and its metabolites in the human brain. Neuropsychopharmacology 11(4):245-256.
Halldin C, Farde L, Hogberg T, Mohell N, Hall H, Suhara T, Karlsson P, Nakashima Y, Swahn CG. 1995. Carbon-11-FLB 457: a radioligand for extrastriatal D2 dopamine receptors. J Nucl Med 36(7):1275-1281.
Halldin C, Gulyas B, Langer O, Farde L. 2001a. Brain radioligands--state of the art and new trends. Q J Nucl Med 45(2):139-152.
Halldin C, Gulyas B, Farde L. 2001b. PET studies with carbon-11 radioligands in neuropsychopharmacological drug development. Curr Pharm Des 7(18):1907-1929.
Hietala J, Syvalahti E, Vuorio K, Nagren K, Lehikoinen P, Ruotsalainen U, Rakkolainen V, Lehtinen V, Wegelius U. 1994. Striatal D2 dopamine receptor characteristics in neuroleptic-naive schizophrenic patients studied with positron emission tomography. Arch Gen Psychiatry 51(2):116-123.

46
Hermans E, Challiss RA. 2001. Structural, signalling and regulatory properties of the group I metabotropic glutamate receptors: prototypic family C G-protein-coupled receptors. Biochem J 359(Pt 3):465-484.
Hwang DR, Kegeles LS, Laruelle M. 2000. (-)-N-[(11)C]propyl-norapomorphine: a positron-labeled dopamine agonist for PET imaging of D(2) receptors. Nucl Med Biol 27(6):533-539.
Hyman SE, Nestler EJ. 1993. The molecular foundations of psychiatry. Washington, DC: American Psychiatric Press. xix, 239.
Ichise M, Liow JS, Lu JQ, Takano A, Model K, Toyama H, Suhara T, Suzuki K, Innis RB, Carson RE. 2003. Linearized reference tissue parametric imaging methods: application to [11C]DASB positron emission tomography studies of the serotonin transporter in human brain. J Cereb Blood Flow Metab 23(9):1096-1112.
Innis RB, Malison RT, al-Tikriti M, Hoffer PB, Sybirska EH, Seibyl JP, Zoghbi SS, Baldwin RM, Laruelle M, Smith EO, et al. 1992. Amphetamine-stimulated dopamine release competes in vivo for [123I]IBZM binding to the D2 receptor in nonhuman primates. Synapse 10(3):177-184.
Jaber M, Robinson SW, Missale C, Caron MG. 1996. Dopamine receptors and brain function. Neuropharmacology 35(11):1503-1519.
Jackson DM, Westlind-Danielsson A. 1994. Dopamine receptors: molecular biology, biochemistry and behavioural aspects. Pharmacol Ther 64(2):291-370.
Johnson CA, Seidel J, Vaquero JJ, Pascau M, Desco M, Green MV. 2002. Exact positioning for OSEM reconstructions on the ATLAS depth-of-interaction small animal scanner. Mol Imaging Biol 4 (suppl 1): S85.
Kapur S, Zipursky R, Jones C, Remington G, Houle S. 2000. Relationship between dopamine D(2) occupancy, clinical response, and side effects: a double-blind PET study of first-episode schizophrenia. Am J Psychiatry 157(4):514-520.
Karlsson P, Farde L, Halldin C, Swahn CG, Sedvall G, Foged C, Hansen KT, Skrumsager B. 1993. PET examination of [11C]NNC 687 and [11C]NNC 756 as new radioligands for the D1-dopamine receptor. Psychopharmacology (Berl) 113(2):149-156.
Kegeles LS, Abi-Dargham A, Zea-Ponce Y, Rodenhiser-Hill J, Mann JJ, Van Heertum RL, Cooper TB, Carlsson A, Laruelle M. 2000. Modulation of amphetamine-induced striatal dopamine release by ketamine in humans: implications for schizophrenia. Biol Psychiatry 48(7):627-640.
Kegeles LS, Zea-Ponce Y, Abi-Dargham A, Rodenhiser J, Wang T, Weiss R, Van Heertum RL, Mann JJ, Laruelle M. 1999. Stability of [123I]IBZM SPECT measurement of amphetamine-induced striatal dopamine release in humans. Synapse 31(4):302-308.
Kim JS, Ichise M, Sangare J, Innis RB. 2006. PET Imaging of Serotonin Transporters with [11C]DASB: Test-Retest Reproducibility Using a Multilinear Reference Tissue Parametric Imaging Method. J Nucl Med 47(2):208-214.
Klunk WE, Engler H, Nordberg A, Wang Y, Blomqvist G, Holt DP, Bergstrom M, Savitcheva I, Huang GF, Estrada S, Ausen B, Debnath ML, Barletta J, Price JC, Sandell J, Lopresti BJ, Wall A, Koivisto P, Antoni G, Mathis CA, Langstrom B. 2004. Imaging brain amyloid in Alzheimer's disease with Pittsburgh Compound-B. Ann Neurol 55(3):306-319.

47
Krasikova RN, Truong P, Halldin C. 2003. A fully automated production of [carbonyl-11C]WAY-100635 for clinical PET studies. J Label Compd Radiopharm 46(Suppl 1):S244.
Laakso A, Hietala J. 2000. PET studies of brain monoamine transporters. Curr Pharm
Des 6(16):1611-1623. Lang AE, Lozano AM. 1998. Parkinson's disease. First of two parts. N Engl J Med
339(15):1044-1053. Langer O, Halldin C, Dolle F, Swahn CG, Olsson H, Karlsson P, Hall H, Sandell J,
Lundkvist C, Vaufrey F, Loc'h C, Crouzel C, Maziere B, Farde L. 1999. Carbon-11 epidepride: a suitable radioligand for PET investigation of striatal and extrastriatal dopamine D2 receptors. Nucl Med Biol 26(5):509-518.
Laruelle M. 2000. Imaging synaptic neurotransmission with in vivo binding competition techniques: a critical review. J Cereb Blood Flow Metab 20(3):423-451.
Laruelle M, Abi-Dargham A, van Dyck CH, Rosenblatt W, Zea-Ponce Y, Zoghbi SS, Baldwin RM, Charney DS, Hoffer PB, Kung HF, et al. 1995. SPECT imaging of striatal dopamine release after amphetamine challenge. J Nucl Med 36(7):1182-1190.
Laruelle M, D'Souza CD, Baldwin RM, Abi-Dargham A, Kanes SJ, Fingado CL, Seibyl JP, Zoghbi SS, Bowers MB, Jatlow P, Charney DS, Innis RB. 1997. Imaging D2 receptor occupancy by endogenous dopamine in humans. Neuropsychopharmacology 17(3):162-174.
Laruelle M, Abi-Dargham A, Gil R, Kegeles L, Innis R. 1999. Increased dopamine transmission in schizophrenia: relationship to illness phases. Biol Psychiatry 46(1):56-72.
Laruelle M, Abi-Dargham A, van Dyck CH, Gil R, D'Souza CD, Erdos J, McCance E, Rosenblatt W, Fingado C, Zoghbi SS, Baldwin RM, Seibyl JP, Krystal JH, Charney DS, Innis RB. 1996. Single photon emission computerized tomography imaging of amphetamine-induced dopamine release in drug-free schizophrenic subjects. Proc Natl Acad Sci U S A 93(17):9235-9240.
Lee CM, Farde L. 2006. Using positron emission tomography to facilitate CNS drug development. Trends Pharmacol Sci 27(6):310-316.
Liow JS, Johnson CA, Toyama H, Green MV, Innis RB. 2003. A single slice rebinning/2D exact positioning OSEM reconstruction for the NIH ATLAS small animal PET scanner. J Nucl Med 44 (suppl): 163P.
Loevinger R, Watson E. 1991. MIRD primer for absorbed dose calculations. The Society of Nuclear Medicine.
Mandela P, Ordway GA. 2006. The norepinephrine transporter and its regulation. J Neurochem 97(2):310-333
Martin LJ, Blackstone CD, Huganir RL, Price DL. 1992. Cellular localization of a metabotropic glutamate receptor in rat brain. Neuron 9(2):259-270.
Mazei MS, Pluto CP, Kirkbride B, Pehek EA. 2002. Effects of catecholamine uptake blockers in the caudate-putamen and subregions of the medial prefrontal cortex of the rat. Brain Res 936(1-2):58-67.
McConathy J, Owens MJ, Kilts CD, Malveaux EJ, Camp VM, Votaw JR, Nemeroff CB, Goodman MM. 2004. Synthesis and biological evaluation of

48
[11C]talopram and [11C]talsupram: candidate PET ligands for the norepinephrine transporter. Nucl Med Biol 31(6):705-718.
McCormick DA, Pape HC, Williamson A. 1991. Actions of norepinephrine in the cerebral cortex and thalamus: implications for function of the central noradrenergic system. Prog Brain Res 88:293-305.
Meltzer HY, Nash JF. 1991. Effects of antipsychotic drugs on serotonin receptors. Pharmacol Rev 43(4):587-604.
Mercuri NB, Bernardi G. 2005. The 'magic' of L-dopa: why is it the gold standard Parkinson's disease therapy? Trends Pharmacol Sci 26(7):341-344.
Michelson D, Faries D, Wernicke J, Kelsey D, Kendrick K, Sallee FR, Spencer T. 2001. Atomoxetine in the treatment of children and adolescents with attention-deficit/hyperactivity disorder: a randomized, placebo-controlled, dose-response study. Pediatrics 108(5):E83.
Mignot E, Laude D. 1985. Study of dopamine turnover by monitoring the decline of dopamine metabolites in rat CSF after alpha-methyl-p-tyrosine. J Neurochem 45(5):1527-1533.
Mikolajczyk K, Szabatin M, Rudnicki P, Grodzki M, Burger C. 1998. A JAVA environment for medical image data analysis: initial application for brain PET quantitation. Med Inform (Lond) 23(3):207-214.
Missale C, Nash SR, Robinson SW, Jaber M, Caron MG. 1998. Dopamine receptors: from structure to function. Physiol Rev 78(1):189-225.
Moghaddam B, Berridge CW, Goldman-Rakic PS, Bunney BS, Roth RH. 1993. In vivo assessment of basal and drug-induced dopamine release in cortical and subcortical regions of the anesthetized primate. Synapse 13(3):215-222.
Musachio JL, Hong J, Ichise M, Seneca N, Brown AK, Liow JS, Halldin C, Innis RB, Pike VW, He R, Zhou J, Kozikowski AP. 2006. Development of new brain imaging agents based upon nocaine-modafinil hybrid monoamine transporter inhibitors. Bioorg Med Chem Lett 16(12):3101-3104.
Nakanishi S. 1992. Molecular diversity of glutamate receptors and implications for brain function. Science 258(5082):597-603.
Nakanishi S. 1994. The molecular diversity of glutamate receptors. Prog Clin Biol Res 390:85-98.
Narendran R, Hwang DR, Slifstein M, Talbot PS, Erritzoe D, Huang Y, Cooper TB, Martinez D, Kegeles LS, Abi-Dargham A, Laruelle M. 2004. In vivo vulnerability to competition by endogenous dopamine: comparison of the D2 receptor agonist radiotracer (-)-N-[11C]propyl-norapomorphine ([11C]NPA) with the D2 receptor antagonist radiotracer [11C]-raclopride. Synapse 52(3):188-208.
Nichols L, Pike VW, Cai L, Innis RB. 2006. Imaging and in vivo quantitation of beta-amyloid: an exemplary biomarker for Alzheimer's disease? Biol Psychiatry 59(10):940-947.
Nordstrom AL, Farde L, Nyberg S, Karlsson P, Halldin C, Sedvall G. 1995. D1, D2, and 5-HT2 receptor occupancy in relation to clozapine serum concentration: a PET study of schizophrenic patients. Am J Psychiatry 152(10):1444-1449.
Nyberg S, Nakashima Y, Nordstrom AL, Halldin C, Farde L. 1996. Positron emission tomography of in-vivo binding characteristics of atypical antipsychotic drugs. Review of D2 and 5-HT2 receptor occupancy studies and clinical response. Br J Psychiatry Suppl(29):40-44.

49
Olanow CW. 2002. The role of dopamine agonists in the treatment of early Parkinson's disease. Neurology 58(4 Suppl 1):S33-41.
Olney JW, Farber NB. 1995. Glutamate receptor dysfunction and schizophrenia. Arch Gen Psychiatry 52(12):998-1007.
Pagano A, Ruegg D, Litschig S, Stoehr N, Stierlin C, Heinrich M, Floersheim P, Prezeau L, Carroll F, Pin JP, Cambria A, Vranesic I, Flor PJ, Gasparini F, Kuhn R. 2000. The non-competitive antagonists 2-methyl-6-(phenylethynyl)pyridine and 7-hydroxyiminocyclopropan[b]chromen-1a-carboxylic acid ethyl ester interact with overlapping binding pockets in the transmembrane region of group I metabotropic glutamate receptors. J Biol Chem 275(43):33750-33758.
Paquet M, Smith Y. 2003. Group I metabotropic glutamate receptors in the monkey striatum: subsynaptic association with glutamatergic and dopaminergic afferents. J Neurosci 23(20):7659-7669.
Parker C, Clarke K, Gee AD, Rabiner E. 2006. In vitro characterisation of the high affinity D2/D3 agonist (+)PhNO. NeuroImage 31:T29
Phelps ME. 2000. PET: the merging of biology and imaging into molecular imaging. J Nucl Med 41(4):661-681.
Phelps ME, Mazziotta JC. 1985. Positron emission tomography: human brain function and biochemistry. Science 228(4701):799-809.
Pietraszek M, Rogoz Z, Wolfarth S, Ossowska K. 2004. Opposite influence of MPEP, an mGluR5 antagonist, on the locomotor hyperactivity induced by PCP and amphetamine. J Physiol Pharmacol 55(3):587-593.
Pin JP, Galvez T, Prezeau L. 2003. Evolution, structure, and activation mechanism of family 3/C G-protein-coupled receptors. Pharmacol Ther 98(3):325-354.
Ponzio F, Achilli G, Calderini G, Ferretti P, Perego C, Toffano G, Algeri S. 1984. Depletion and recovery of neuronal monoamine storage in rats of different ages treated with reserpine. Neurobiol Aging 5(2):101-104.
Primus RJ, Thurkauf A, Xu J, Yevich E, McInerney S, Shaw K, Tallman JF, Gallagher DW. 1997. II. Localization and characterization of dopamine D4 binding sites in rat and human brain by use of the novel, D4 receptor-selective ligand [3H]NGD 94-1. J Pharmacol Exp Ther 282(2):1020-1027.
Roland PE, Zilles K. 1994. Brain atlases--a new research tool. Trends Neurosci 17(11):458-467.
Romano C, Sesma MA, McDonald CT, O'Malley K, Van den Pol AN, Olney JW. 1995. Distribution of metabotropic glutamate receptor mGluR5 immunoreactivity in rat brain. J Comp Neurol 355(3):455-469.
Ross SB, Jackson DM. 1989. Kinetic properties of the in vivo accumulation of 3H-(-)-N-n-propylnorapomorphine in mouse brain. Naunyn Schmiedebergs Arch Pharmacol 340(1):13-20.
Saunders RC, Kolachana BS, Weinberger DR. 1994. Local pharmacological manipulation of extracellular dopamine levels in the dorsolateral prefrontal cortex and caudate nucleus in the rhesus monkey: an in vivo microdialysis study. Exp Brain Res 98(1):44-52.
Schoepp DD, Jane DE, Monn JA. 1999. Pharmacological agents acting at subtypes of metabotropic glutamate receptors. Neuropharmacology 38(10):1431-1476.
Schou M, Halldin C, Pike VW, Mozley PD, Dobson D, Innis RB, Farde L, Hall H. 2005. Post-mortem human brain autoradiography of the norepinephrine

50
transporter using (S,S)-[18F]FMeNER-D2. Eur Neuropsychopharmacol 15(5):517-520.
Schou M, Halldin C, Sovago J, Pike VW, Gulyas B, Mozley PD, Johnson DP, Hall H, Innis RB, Farde L. 2003. Specific in vivo binding to the norepinephrine transporter demonstrated with the PET radioligand, (S,S)-[11C]MeNER. Nucl Med Biol 30(7):707-714.
Schou M, Halldin C, Sovago J, Pike VW, Hall H, Gulyas B, Mozley PD, Dobson D, Shchukin E, Innis RB, Farde L. 2004. PET evaluation of novel radiofluorinated reboxetine analogs as norepinephrine transporter probes in the monkey brain. Synapse 53(2):57-67.
Schou M, Sovago J, Pike VW, Gulyas B, Bogeso KP, Farde L, Halldin C. 2006. Synthesis and positron emission tomography evaluation of three norepinephrine transporter radioligands: [C-11]desipramine, [C-11]talopram and [C-11]talsupram. Mol Imaging Biol 8(1):1-8.
Seeman P, Chau-Wong M, Tedesco J, Wong K. 1975. Brain receptors for antipsychotic drugs and dopamine: direct binding assays. Proc Natl Acad Sci U S A 72(11):4376-4380.
Seeman P, Watanabe M, Grigoriadis D, Tedesco JL, George SR, Svensson U, Nilsson JL, Neumeyer JL. 1985. Dopamine D2 receptor binding sites for agonists. A tetrahedral model. Mol Pharmacol 28(5):391-399.
Seidel J, Vaquerro JJ, Green MV (2003). Resolution uniformity and sensitivity of the NIH ATLAS small animal PET scanner: comparison to simulated LSO scanners without depth-of-interaction capability. IEEE Trans Nucl Sci 50: 1347-50.
Seneca N, Andree B, Sjoholm N, Schou M, Pauli S, Mozley PD, Stubbs JB, Liow JS, Sovago J, Gulyas B, Innis R, Halldin C. 2005. Whole-body biodistribution, radiation dosimetry estimates for the PET norepinephrine transporter probe (S,S)-[18F]FMeNER-D2 in non-human primates. Nucl Med Commun 26(8):695-700.
Seneca N, Finnema SJ, Farde L, Gulyas B, Wikstrom HV, Halldin C, Innis RB. 2006a. Effect of amphetamine on dopamine D2 receptor binding in nonhuman primate brain: a comparison of the agonist radioligand [11C]MNPA and antagonist [11C]raclopride. Synapse 59(5):260-269.
Seneca N, Gulyas B, Varrone A, Schou M, Airaksinen A, Tauscher J, Vandenhende F, Kielbasa W, Farde L, Innis RB, Halldin C. 2006b. Atomoxetine occupies the norepinephrine transporter in a dose-dependent fashion: a PET study in nonhuman primate brain using (S,S)-[18F]FMeNER-D2. Psychopharmacology (Berl) 188(1):119-127.
Shigemoto R, Nomura S, Ohishi H, Sugihara H, Nakanishi S, Mizuno N. 1993. Immunohistochemical localization of a metabotropic glutamate receptor, mGluR5, in the rat brain. Neurosci Lett 163(1):53-57.
Sibley DR, Mahan LC, Creese I. 1983. Dopamine receptor binding on intact cells. Absence of a high-affinity agonist-receptor binding state. Mol Pharmacol 23(2):295-302.
Sokoloff P, Giros B, Martres MP, Bouthenet ML, Schwartz JC. 1990. Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 347(6289):146-151.

51
Spencer T, Biederman J, Wilens T, Prince J, Hatch M, Jones J, Harding M, Faraone SV, Seidman L. 1998. Effectiveness and tolerability of tomoxetine in adults with attention deficit hyperactivity disorder. Am J Psychiatry 155(5):693-695.
Spooren W, Gasparini F. 2004. mGlu5 receptor antagonists: a novel class of anxiolytics? Drug News Perspect 17(4):251-257.
Stoessl AJ, de la Fuente-Fernandez R. 2003. Dopamine receptors in Parkinson's disease: imaging studies. Adv Neurol 91:65-71.
Suzuki M, Hurd YL, Sokoloff P, Schwartz JC, Sedvall G. 1998. D3 dopamine receptor mRNA is widely expressed in the human brain. Brain Res 779(1-2):58-74.
Talbot PS, Laruelle M. 2002. The role of in vivo molecular imaging with PET and SPECT in the elucidation of psychiatric drug action and new drug development. Eur Neuropsychopharmacol 12(6):503-511.
Tamminga CA, Holcomb HH, Gao XM, Lahti AC. 1995. Glutamate pharmacology and the treatment of schizophrenia: current status and future directions. Int Clin Psychopharmacol 10 Suppl 3:29-37.
Tipre DN, Fujita M, Chin FT, Seneca N, Vines D, Liow JS, Pike VW, Innis RB. 2004. Whole-body biodistribution and radiation dosimetry estimates for the PET dopamine transporter probe 18F-FECNT in non-human primates. Nucl Med Commun 25(7):737-742.
Vallone D, Picetti R, Borrelli E. 2000. Structure and function of dopamine receptors. Neurosci Biobehav Rev 24(1):125-132.
van Berckel BN, Kegeles LS, Waterhouse R, Guo N, Hwang DR, Huang Y, Narendran R, Van Heertum R, Laruelle M. 2006. Modulation of amphetamine-induced dopamine release by group II metabotropic glutamate receptor agonist LY354740 in non-human primates studied with positron emission tomography.
Verhoeff NP, Kapur S, Hussey D, Lee M, Christensen B, Psych C, Papatheodorou G, Zipursky RB. 2001. A simple method to measure baseline occupancy of neostriatal dopamine D2 receptors by dopamine in vivo in healthy subjects. Neuropsychopharmacology 25(2):213-223.
Volkow ND, Fowler JS, Wang GJ, Dewey SL, Schlyer D, MacGregor R, Logan J, Alexoff D, Shea C, Hitzemann R. 1993. Reproducibility of repeated measures of carbon-11-raclopride binding in the human brain. J Nucl Med 34(4):609-613.
Volkow ND, Wang GJ, Fowler JS, Logan J, Schlyer D, Hitzemann R, Lieberman J, Angrist B, Pappas N, MacGregor R, et al. 1994. Imaging endogenous dopamine competition with [11C]raclopride in the human brain. Synapse 16(4):255-262.
Wienhard K, Dahlbom M, Eriksson L, Michel C, Bruckbauer T, Pietrzyk U, Heiss WD. 1994. The ECAT EXACT HR: performance of a new high resolution positron scanner. J Comput Assist Tomogr 18(1):110-118.
Wilson AA, Johnson DP, Mozley D, Hussey D, Ginovart N, Nobrega J, Garcia A, Meyer J, Houle S. 2003. Synthesis and in vivo evaluation of novel radiotracers for the in vivo imaging of the norepinephrine transporter. Nucl Med Biol 30(2):85-92.
Wilson AA, McCormick P, Kapur S, Willeit M, Garcia A, Hussey D, Houle S, Seeman P, Ginovart N. 2005. Radiosynthesis and evaluation of [11C]-(+)-4-propyl-3,4,4a,5,6,10b-hexahydro-2H-naphtho[1,2-b][1,4]oxazin-9 -ol as a potential radiotracer for in vivo imaging of the dopamine D2 high-affinity state with positron emission tomography. J Med Chem 48(12):4153-4160.

52
Wong DF, Wagner HN, Jr., Tune LE, Dannals RF, Pearlson GD, Links JM, Tamminga CA, Broussolle EP, Ravert HT, Wilson AA, Toung JK, Malat J, Williams JA, O'Tuama LA, Snyder SH, Kuhar MJ, Gjedde A. 1986. Positron emission tomography reveals elevated D2 dopamine receptors in drug-naive schizophrenics. Science 234(4783):1558-1563.
Wong D, Kuwabara H, Mozley PD, Dannals R, Kumar A, Ye W, Brasic J, Alexander M, Mathews W, Holt D, Vandenhende F, Gjedde A. 2005. Characterization of dose dependent norepinephrine transporter blockade by atomoxetine in human brain using [11C]MeNER PET. Brain and Brain PET 2005: BP-12.
Zhou J. 2004. Norepinephrine transporter inhibitors and their therapeutic potential. Drugs Future 29(12):1235-1244.