Embed Size (px)
Citation preview

PHAR2811 Dale’s lecture 3
Genome Structure
COMMONWEALTH OF AUSTRALIA Copyright Regulation
WARNING This material has been reproduced and communicated to you by or on behalf of the University of Sydney pursuant to Part VB of
the Copyright Act 1968 (the Act). The material in this communication may be subject to copyright
under the Act. Any further reproduction or communication of this material by you may be the subject of copyright protection
under the Act.
Do not remove this notice

Questions from the last lecture
• The role of methotrexate in the inhibition of thymidylate synthase
• Methotrexate is a competitive inhibitor of the enzyme that converts dihydrofolate (DHF) to tetrahydrofolate (THF) (dihydrofolate reductase, DHFR)
• It binds to the enzyme with ~1000 times the affinity of DHF...making the inhibition almost irreversible
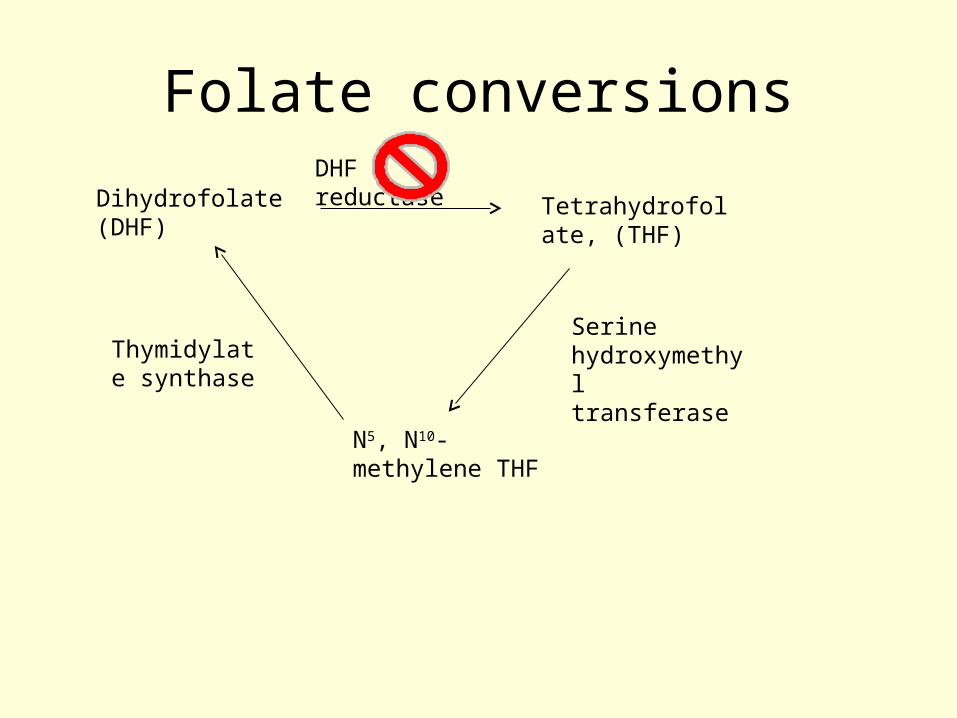
Folate conversions
Dihydrofolate (DHF)
Tetrahydrofolate, (THF)
DHF reductase
N5, N10-methylene THF
Serine hydroxymethyl transferase
Thymidylate synthase

N
NNH2N
O
H
H
CH2
N
H R etc
H
Dihydrofolate, DHF
N
NNH2N
O
H
H
CH2
N
H R etc
H
H
H
NADPH + H+
NADP+
Tetrahydrofolate, THF
DHF reductase
N
NNH2N
O
H
H
CH2
N
R etc
H
H
CH2
Serine
Glycine
Serine Hydroxmethyl Transferase
ThymidylateSynthase
This methylene group is transferred to dUMP at C6
dUMP
TMP

Prokaryotes
• The genome of prokaryotes is extremely efficient.
• There are 4.6 million base pairs in your average E. coli
• If the average bacterial protein has a molecular weight of ~40,000 D how many different proteins does the average E. coli make?

Prokaryotes
• To do this calculation you need to know:
• The average mol. Wt. of an amino acid ~100
• This means the average protein has 400 amino acids
• Which means 1200 bases + promoter and terminator sequences ~1500 bp.
• 4.6 X 106/1500 = ~3000 different proteins.

In humans if the whole genome was coding
• The genome has 3*109 bp and the average protein subunit is 50,000
• So 500 aa = 1500 bp = 3000 bp with large promoter regions which makes the maths easy
• 3*10 ^9/3*10^3 = 1*10^6 or 1 million different proteins. We only make about 30,000 different proteins so there is a discrepancy

Prokaryotes versus Eukaryotes
• Prokaryotes have no room for redundant sequences.
• Their survival depends on rapid proliferation when nutrients are available
• Complex multi-cellular eukaryotes depend for survival on quick responses, adjusting to changes in the environment.

Prokaryotes versus Eukaryotes
• E. coli can divide every 20 min if conditions are optimal
• The human cell takes 18 to 24 h to go through the cell cycle once.
• The human genome only has about 2% coding regions.
• The gene density is much lower!!

Chromosome Characteristics
• Chromosomes vary in number between species. The chromosome number is a combination of the haploid number (n) X the number of sets. Algae and fungi are haploid; most animals and plants are diploid. The number of pairs of chromosomes in different species’ genomes is bizarre.

What do these life forms have in common?

Chromosome CharacteristicsSpecies Genome size in
Megabases (Mb)# Haploid chromosomes
E. coli 4.6 1 circular
yeast 13 16
cow 39
human 3 000 23
alligator 16
carp 52
salamander 90 000 14

Chromosome Characteristics• Chromosomes vary in size within a
species. Within the human genome there is a four fold difference in the size of the chromosomes.
• Centromere: the region of the chromosome where the spindle fibres attach. Repetitive satellite DNA is often found around the centromere.
• Telomere: ends of the chromosome, containing a distinct repeating sequence, which enables the ends of the chromosome to replicate.

Centromere Characteristics
• The relative position of the centromere is constant, which means that the ratio of the lengths of the two arms is constant for each chromosome. This ratio is an important parameter for chromosome identification, and also, the ratio of lengths of the two arms allows classification of chromosomes into several basic morphologic types:

Centromere Characteristics

Chromosome Banding
• Chromosomes can be stained with special dyes which give a consistent and unique pattern like a bar-code for each chromosome; so much so that the bands have been numbered.
• The most common stain used is a Giesma stain. This stain, when applied after mild proteolytic treatment (trypsin) gives light (G-light) and dark (G-dark) bands.

The Human Genome


The ps and qs of chromosomes
• There are 2 arms on the chromosome denoted p and q
• For most chromosomes the short arm is the petit or p arm
• The longer arm is the q or queue arm
• Numbering is done from the centromere along one of the arms

Chromosome Banding
• When viewed at the lowest resolution only a few bands appear. These are numbered p1, p2, p3 etc counting from the centromere.
• If the stained chromosomes are viewed at higher resolution many sub-bands are revealed. So the labelling then goes p11, p12, p13.
• So if your DNA marker may be given a position on the chromosome with a set of numbers like 17p23. This means the locus is on chromosome 17 on the short p arm in sub-band 23.

Some terms:
• The general material which makes up the chromosomes is called chromatin This is composed of DNA and protein.
• Heterochromatin contains DNA which is more tightly packaged or condensed and probably is transcriptionally inert.
• Euchromatin contains most active genes; those actively transcribed.

Chromosome packaging at the molecular level.
• Each chromosome contains a single molecule of DNA
• This DNA is wound around small proteins called histones
• These proteins have lots of lysine and arginine residues, making them very positively charged at pH 7 (and high pIs ~12)

Histones
• There are 5 major histone variants: H1, H2A, H2B, H3 and H4.
• Two molecules each of H2A + H2B + H3 + H4 make up an octamer which the DNA wraps around with 1.7 turns.
• This structure is known as a nucleosome.• Each nucleosome has an H1 associated
and a linker section of DNA, like beads on a thread.

Histone Octamer

Histones
• The major force holding the association of histones to DNA is electrostatic.
• To separate the histones from the DNA, chromatin is treated with high ionic strength solutions. The high salt reduces the electrostatic interactions and the protein dissociates from the DNA.

Higher order Packaging


Figure 11.28A model for chromosome structure, human chromosome 4. The 2-nm DNA helix is wound twice around histone octamers to form 10-nm nucleosomes, each of which contains 160 bp (80 per turn). These nucleosomes are then wound in solenoid fashion with six nucleosomes per turn to form a 30-nm filament. In this model, the 30-nm filament forms long DNA loops, each containing about 60,000 bp, which are attached at their base to the nuclear matrix. Eighteen of these loops are then wound radially around the circumference of a single turn to form a miniband unit of a chromosome. Approximately 106 of these minibands occur in each chromatid of human chromosome 4 at mitosis.

The cell cycleAt interphase (G1, S and G2) the chromosomes look like a plate of spaghetti, entangled and dispersed throughout the nucleus
At M phase the newly replicated daughter chromatids condense and line up.

Chromosomes at interphase

The role of histones
• Shield the negative charges of the phosphates
• Allow bending and DNA wrapping
• Restrict access to transcription
• The interaction between the histones and the DNA is dynamic and non-base sequence specific

Histone Remodeling
• Influences the DNA accessibility for transcription
• Can be one of the first events when switching on a set of genes.
• Nucleosome remodeling complexes
• Nucleosome positioning
• Histone modification

Histone modifications
• Phosphorylation; serine residues
• Methylation, adding –CH3
• Acetylation, lysine residues

Histone modifications
• Phosphorylation; serine residues
• Methylation, adding –CH3
• Acetylation, lysine residues

Acetylation
• Transferring an acetyl group to the amino side chain of lysine residues
• Histone acetyl Transferases (HATs)
• Histone deacetylases (HDAs)

Lysine
NH
CH
C
H2C
NH
O
H2C
H2C
H2C +NH3
C O

Acetylated Lysine
NH
CH
C
H2C
NH
O
H2C
H2C
H2C NH
C O
C CH3
O

What effect would acetylation have on DNA accessibility?
• It neutralises the positive charge of the lysine side chain
• The histone will not have as much affinity for the DNA phosphates (negative)
• The nucleosome packing will be looser
• DNA more accessible for transcription
• Deacetylases will pack it up again!





![[PPT]Dale’s cone of experience - WordPress.com · Web viewCone of Experience is a visual model meant to summarize Dale’s classification system for the varied types of mediated](https://img.pdfslide.net/doc/110x75/5b3623427f8b9a8b4b8dea1c/pptdales-cone-of-experience-web-viewcone-of-experience-is-a-visual-model.jpg)























