Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 263, No. 29, Issue of October 15, PP. 1497614983.1988 Printed in U. S. A.
Phosphorylation of @-Crystallin B2 (@Bp) in the Bovine Lens*
(Received for publication, June 1, 1988)
Norman J. KleimanS, Raul Chiesa, Mary Ann Gawinowicz KolksQ, and Abraham Spectorn From the Biochemistry and Molecular Biology Laboratory, Department of Ophthalmology and §Center of Reproductive Sciences, Protein Chemistry Core Laboratory, College of Physicians and Surgeons of Columbia University, New York, New York I0032
Three major “P-labeled polypeptides were found in the soluble fraction of bovine lenses cultured in a me- dium containing [32P]orthophosphate. Two of the poly- peptides corresponded to the phosphorylated A and B chains of a-crystallin. In this communication, the third polypeptide is now identified. This polypeptide is char- acterized by a molecular weight of 27,000 and a PI of 6.6, eluted exclusively in the j 3 ~ ~ ~ fraction of a CL-GB gel filtration separation of lens soluble material, and could be further purified by DE52 anion exchange chromatography. The only “P-labeled amino acid de- tected was phosphoserine. A single 32P-labeled peptide was observed after tryptic digestion and two-dimen- sional mapping. The amino acid sequence of the puri- fied peptide is Gly-Ala-Phe-His-Pro-Ser-Ser. This se- quence exactly matches the expected C-terminal tryp- tic fragment, residues 198-204, of the bovine B- crystallin B2. The results of carboxypeptidase A diges- tion of the 32P-labeled peptide suggest that only SerZo3 is phosphorylated. By using the catalytic subunit of the CAMP-dependent protein kinase, purified BB2 was phosphorylated in vitro, generating a single 32P-la- beled polypeptide with the identical PI as the phospho- rylated polypeptide obtained from lens culture. On the basis of these data, the M, 27,000 “P-labeled polypep- tide is identified as the phosphorylated form of the B- crystallin B2.
The lens of the eye is a unique transparent and avascular tissue that grows throughout life. Beneath the anterior surface of the lens capsule there is a single layer of epithelial cells that differentiate into mature lens fiber cells (1). New fiber cells are constantly formed overlaying older less metabolically active fibers. While most of the proteins found in the epithe- lial cells are common to many cell types, the bulk of the lens dry weight is composed of a specialized set of lens proteins termed crystallins (2-4).
Lens crystallins have traditionally been arranged into three classes, termed a, @, and y (5, 6). @-Crystallin includes a number of aggregates ranging in size from 50,000 to 200,000 Da and can be further divided into a smaller sized subclass (PhW) and a larger sized subclass ( @ ~ ~ ~ h ) . A variety of polypep- tide species, varying in molecular mass from 20,000 to 30,000
* This work was supported by grants from the National Eye Insti- tute, National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accord- ance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of a National Institutes of Health Individual National Research Service Award.
To whom correspondence should be addressed Biochemistry and Molecular Biology Laboratory, Dept. of Ophthalmology, College of Physicians and Surgeons, Columbia University, 630 West 168th St., New York, NY 10032.
Da and including the major constituent @B2 (also known as @Bp), has been identified and characterized (7-11). The com- plete sequence of @B2 has been determined (10).
The crystallins are generally assumed to function exclu- sively as structural proteins in the lens, primarily contributing to the maintenance of lens transparency (12, 13). Several recent findings, however, suggest that the concept that crys- tallins are passive lens structures needs to be reevaluated. These include the observations that “taxon-specific” crystal- lins are very closely related to certain cellular enzymes (14- 16) and that one of the y-crystallins may be involved in redox reactions (17). Furthermore, the finding that a-crystallin is phosphorylated in an apparently CAMP-dependent manner (18) suggests that crystallins are subject to metabolic control.
The present paper extends this concept by reporting that a constituent of second crystallin family, @-crystallin B2, is phosphorylated.‘
EXPERIMENTAL PROCEDURES AND RESULTS*
Fig. 1 shows the CL-GB gel filtration separation of soluble lens crystallins obtained from intact bovine lenses incubated in a medium containing [32P]orthophosphate. The Coomassie Blue-stained and autoradiographic SDS-PAGE3 profiles of the total soluble lens homogenate and each pooled crystallin fraction are shown. Two 32P-labeled M, 20,000 and 22,000 CY-
crystallin species were observed. This supplements previous reports (18, 20) which displayed the gel profiles of lens crys- tallins phosphorylated in uitro. While trichloroacetic acid- precipitable radioactivity was found associated with the a- crystallin fraction, approximately 30% of the total 32P radio- activity in the soluble fraction was associated with the @Low
(DL) peak. No radioactivity was ever associated with the &igb
(@H) material. Variable amounts of a 32P-labeled M, 18,000 polypeptide were sometimes observed in the valley between @L and y-crystallin (see Fig. 1, lanes marked TS) .
When the PI. material was pooled and analyzed by SDS- PAGE and isoelectric focusing, the majority of the radioactiv- ity was associated with a single band of M, 27,000 (Fig. 1) having an isoelectric point of about 6.6 (Figs. 2 and 3). (The major @-crystallin polypeptide B2 has a molecular weight of 27,000 and a PI of about 7.0.) When the pooled a material was also analyzed by isoelectric focusing (Fig. 2), two 32P- labeled species, corresponding to the phosphorylated a-crys- tallin A1 and B1 chains, were observed. SDS-PAGE and isoelectric focusing analyses of individual fractions across the
A preliminary abstract of this work has been published (19). Portions of this paper (including “Experimental Procedures,” part
of “Results,” and Figs. 4 and 5) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press.
The abbreviations used are: SDS-PAGE, sodium dodecyl sulfate- polyacrylamide gel electrophoresis; MEM, minimum essential me- dium.
14978

Phosphorylation of B-Crystallin B2 (BBp)
FIG. 1. Separation of soluble bovine lens crystalline by CL- 6B gel filtration chromatography. The A m profile (-) and trichloroacetic acid (TCA)-precipitable radioactivity (- - -) profile are shown. The total soluble fraction (TS) and the individual pooled CL-GB peak fractions were resolved by SDS-PAGE and the labeled bands detected by autoradiography. Coomassie Blue-stained gels (C) and autoradiograms ( R ) are shown below the respective peaks. The positions of molecular weight standards are indicated.
@L peak suggested that both the relative specific activity of the 3ZP-labeled polypeptide and the relative ratio of the 32P- labeled polypeptide to the major @-crystallin B2 were constant in all fractions (not shown).
Two-dimensional isoelectric focusing x SDS-PAGE of pooled @L from the CL-GB column is shown in Fig. 3. The overall Coomassie Blue-stained protein profile appears similar to that previously reported for @-crystallin (9, 11). The no- menclature for @-crystallin polypeptides proposed by Berbers et al. (11) is used to identify the various polypeptide spots. The 32P-labeled polypeptide has the same molecular weight as the major @-crystallin @B2 (previously known as @Bp) and appears to be an acidic variant. These observations are con- sistent with the interpretation that the 27,000 polypeptide is a phosphorylated form of pB2. The putative phosphorylated @B2 appears to represent approximately 10% of the total @B2 based on densitometric scanning of the Coomassie Blue- stained spots of @B2 and its labeled acidic variant. Based on the ratio between Coomassie stain and 32P label, it appears that the relative specific activity of the M, 27,000 polypeptide is at least as great as that of a-crystallin B1.
The data presented above suggested that the M, 27,000 polypeptide is a form of @-crystallin B2. To explore this possibility, the 32P-labeled M, 27,000 polypeptide was further purified by DE52 anion exchange chromatography in prepa- ration for sequence analysis (Fig. 4) (data presented in Min- iprint). An aliquot of this material was partially hydrolyzed and used to identify phosphoserine as the only 32P-labeled amino acid (Fig. 5 ) (data presented in Miniprint). The re-
C R a
14979
0
3 C R
BL FIG. 2. Analysis of the pooled a- and &,-crystallin fractions
from the CL-GB gel filtration column by pH 5-8 urea isoelec- tric focusing. The Coomassie Blue-stained gel and respective auto- radiogram of the pooled a and D L fractions are shown. The location of the 32P-labeled M, 27,000 polypeptide found in the peak is indicated by an arrow. In addition, the positions of the major cr- crystallin polypeptides A2 and BP and the major p-crystallin polypep- tide B2 is indicated. The positions of the phosphorylated a-crystallin species AI and B, are also shown. C, Coomassie Blue-stained gel; R, autoradiogram.
maining 32P-labeled material was digested with trypsin, and two-dimensional peptide maps were prepared. Only one major 32P-labeled tryptic peptide was detected (Fig. 6). Quantities of this phosphopeptide sufficient for peptide sequencing were obtained by preparative thin-layer chromatography followed by preparative thin-layer electrophoresis at pH 1.9 as de- scribed under “Experimental Procedures.” The purity of the resultant peptide was confirmed by analytical thin-layer elec- trophoresis at pH 5.6. A single fluorescamine positive spot was detected, which matched exactly the spot on the corre- sponding autoradiogram (not shown). Sequence analysis of this peptide identified the unequivocal sequence Gly-Ala-Phe- His-Pro-Ser-Ser. This sequence exactly matches the expected C-terminal tryptic fragment of @-crystallin B2.
It was reported previously that the a-crystallin A and B chains are phosphorylated in their C- and N-terminal do- mains, respectively (21). In this regard, it is interesting to note that @-crystallin is also phosphorylated in a terminal domain. The physiological significance of this observation is unknown.
The identification of the M , 27,000 polypeptide as phos- phorylated @-crystallin B2 if further strengthened by the following experiment. Unphosphorylated purified @B2 was isolated by DE52 anion exchange chromatography and incu- bated with the catalytic subunit of the CAMP-dependent protein kinase. Polypeptide species were resolved by isoelec- tric focusing and compared to the profiles obtained from both the total soluble supernatant and the @L fraction isolated from intact lenses labeled with [32P]orthophosphate. A single 32P- labeled band with the identical isoelectric point as that of the 32P-labeled M , 27,000 polypeptide from the intact lens was detected (Fig. 7).

14980 Phosphorylation of &Crystallin B2 (BBp)
0 pH 6.6 I
2 7 K D -
2 7 K D -
FIG. 3. Two-dimensional isoelectric focusing X SDS-PAGE separation of the pooled fractions from the 1 9 ~ peak of a CL- 6B gel filtration separation. The corresponding position of the phosphorylated protein in the Coomassie Blue-stained gel (top) and the autoradiogram (bottom) is indicated by an arrow. The molecular weight and the isoelectric point of the 3ZP-labeled M, 27,000 polypep- tide and the nomenclature used to identify various @-crystallin poly- peptides are indicated.
-- electrophoresis - + FIG. 6. Two-dimensional tryptic peptide map of peak 2 ma-
terial from the DE52 column. The first dimension was electro- phoresis a t pH 1.9 and the second dimension ascending chromatog- raphy. The autoradiogram is shown.
0 C R C R C R ws P L m 2
FIG. 7. I n vitro phosphorylation of purified 19B2 by the catalytic subunit of the CAMP-dependent kinase. For compari- son, 32P-labeled polypeptides in the total water-soluble homogenate ( l a n e WS) and the @L peak ( l a n e @I,), both obtained from lens organ culture incubated with [32P]orthophosphate, are shown. These sam- ples and purified @B2 incubated in vitro with the catalytic subunit of the CAMP-dependent protein kinase ( l a n e @B2) were resolved in a pH 6-8 isoelectric focusing gel. Both the Coomassie Blue-stained gel (C) and the autoradiogram (R) are shown.
It was of interest to determine the precise location of the phosphorylated serine(s). Because the two serine residues in the 32P-labeled tryptic peptide are the penultimate and ter- minal amino acids, it seemed unlikely that accurate measure- ments of the release of 32P label could be made using stepwise Edman degradation. Therefore, to determine the position(s) of the 32P-labeled serine(s), the purified peptide was digested with carboxypeptidase A. This enzyme would be expected to only remove the terminal serine because of the presence of a proline at residue 202 (22). In these experiments, free serine and/or phosphoserine as well as the remaining peptide were resolved by thin-layer chromatography or thin-layer electro- phoresis.
By using a peptide:enzyme ratio of 101 and digestion times ranging from 4 to 16 h, the time-dependent appearance of a fluorescamine-positive spot corresponding to the position of serine was observed. Even after 16 h of digestion, however, the fluorescent intensity of this spot appeared less than that of the remaining 32P-labeled peptide, suggesting that the digestion was incomplete. Phosphoserine was never detected (Fig. 8). The same results were obtained using the two- dimensional chromatographic system described under “Ex- perimental Procedures.” Similar experiments, using carboxy- peptidase Y, were completely ineffective in removing the terminal serine.
These results indicate that some fraction of the total 32P- labeled peptide is phosphorylated only at the penultimate serine, SerZo3. It is unlikely that both SeP3 and Ser2M are phosphorylated in the remaining undigested peptide, because a difference in charge contributed by an additional phosphate moiety would have resulted in the appearance of a second 32P- labeled component upon thin-layer electrophoresis. It would seem likely, although not certain, that in the remaining intact peptide only Ser203 is phosphorylated.
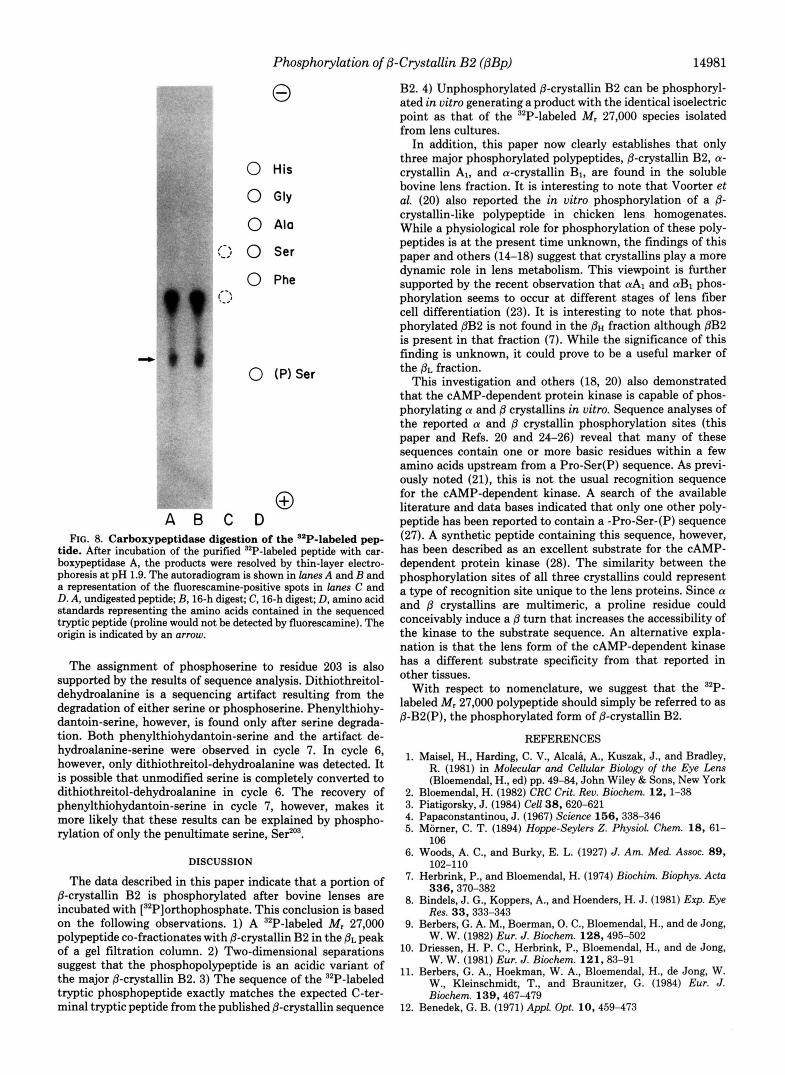
Phosphorylation of @-Crystallin B2 (@Bp) 14981
0 (PI Ser
0 A B C D
FIG. 8. Carboxypeptidase digestion of the '"P-labeled pep- tide. After incubation of the purified "P-labeled peptide with car- boxypeptidase A, the producta were resolved by thin-layer electro- phoresis at pH 1.9. The autoradiogram is shown in lanes A and B and a representation of the fluorescamine-positive spots in lanes C and D. A, undigested peptide; B, 16-h digest; C, 16-h digest; D, amino acid standards representing the amino acids contained in the sequenced tryptic peptide (proline would not be detected by fluorescamine). The origin is indicated by an arrow.
The assignment of phosphoserine to residue 203 is also supported by the results of sequence analysis. Dithiothreitol- dehydroalanine is a sequencing artifact resulting from the degradation of either serine or phosphoserine. Phenylthiohy- dantoin-serine, however, is found only after serine degrada- tion. Both phenylthiohydantoin-serine and the artifact de- hydroalanine-serine were observed in cycle 7. In cycle 6, however, only dithiothreitol-dehydroalanine was detected. It is possible that unmodified serine is completely converted to dithiothreitol-dehydroalanine in cycle 6. The recovery of phenylthiohydantoin-serine in cycle 7, however, makes it more likely that these results can be explained by phospho- rylation of only the penultimate serine, Seflo3.
DISCUSSION
The data described in this paper indicate that a portion of 8-crystallin B2 is phosphorylated after bovine lenses are incubated with [32P]orthophosphate. This conclusion is based on the following observations. 1) A 32P-labeled M, 27,000 polypeptide co-fractionates with @-crystallin B2 in the / 3 ~ peak of a gel filtration column. 2) Two-dimensional separations suggest that the phosphopolypeptide is an acidic variant of the major @-crystallin B2.3) The sequence of the 32P-labeled tryptic phosphopeptide exactly matches the expected C-ter- minal tryptic peptide from the published @-crystallin sequence
B2. 4) Unphosphorylated 8-crystallin B2 can be phosphoryl- ated in vitro generating a product with the identical isoelectric point as that of the 32P-labeled M, 27,000 species isolated from lens cultures.
In addition, this paper now clearly establishes that only three major phosphorylated polypeptides, j3-crystallin B2, a- crystallin Al, and a-crystallin B1, are found in the soluble bovine lens fraction. It is interesting to note that Voorter et al. (20) also reported the in vitro phosphorylation of a B- crystallin-like polypeptide in chicken lens homogenates. While a physiological role for phosphorylation of these poly- peptides is at the present time unknown, the findings of this paper and others (14-18) suggest that crystallins play a more dynamic role in lens metabolism. This viewpoint is further supported by the recent observation that aA1 and aB1 phos- phorylation seems to occur at different stages of lens fiber cell differentiation (23). It is interesting to note that phos- phorylated @B2 is not found in the @H fraction although @B2 is present in that fraction (7). While the significance of this finding is unknown, it could prove to be a useful marker of the @L fraction.
This investigation and others (18, 20) also demonstrated that the CAMP-dependent protein kinase is capable of phos- phorylating a and @ crystallins in vitro. Sequence analyses of the reported a and @ crystallin phosphorylation sites (this paper and Refs. 20 and 24-26) reveal that many of these sequences contain one or more basic residues within a few amino acids upstream from a Pro-Ser(P) sequence. As previ- ously noted (21), this is not the usual recognition sequence for the CAMP-dependent kinase. A search of the available literature and data bases indicated that only one other poly- peptide has been reported to contain a -Pro-Ser-(P) sequence (27). A synthetic peptide containing this sequence, however, has been described as an excellent substrate for the CAMP- dependent protein kinase (28). The similarity between the phosphorylation sites of all three crystallins could represent a type of recognition site unique to the lens proteins. Since a and B crystallins are multimeric, a proline residue could conceivably induce a 8 turn that increases the accessibility of the kinase to the substrate sequence. An alternative expla- nation is that the lens form of the CAMP-dependent kinase has a different substrate specificity from that reported in other tissues.
With respect to nomenclature, we suggest that the 32P- labeled M, 27,000 polypeptide should simply be referred to as @-B2(P), the phosphorylated form of @crystallin B2.
REFERENCES 1. Maisel, H., Harding, C. V., Alcali, A., Kuszak, J., and Bradley,
R. (1981) in Molecdur and Cellular Biology of the Eye Lens (Bloemendal, H., ed) pp. 49-84, John Wiley & Sons, New York
2. Bloemendal, H. (1982) CRC Crit. Rev. Biochem. 12,l-38 3. Piatigorsky, J. (1984) Cell 38,620-621 4. Papaconstantinou, J. (1967) Science 156,338-346 5. Morner, C . T. (1894) Hoppe-Seylers 2. Physwl. Chem. 18, 61-
6. Woods, A. C., and Burky, E. L. (1927) J. Am. Med. Assoc. 89,
7. Herbrink, P., and Bloemendal, H. (1974) Biochim. Biophys. Acta
8. Bindels, J. G., Koppers, A., and Hoenders, H. J. (1981) Exp. Eye Res. 33,333-343
9. Berbers, G . A. M., Boerman, 0. C., Bloemendal, H., and de Jong, W. W. (1982) Eur. J. Biochem. 128,495-502
10. Driessen, H. P. C., Herbrink, P., Bloemendal, H., and de Jong, W. W. (1981) Eur. J. Biochem. 121.83-91
11. Berbers, G. A., Hoekman, W. A., Bloemendal, H., de Jong, W. W., Kleinschmidt, T., and Braunitzer, G. (1984) Eur. J. Biochem. 139,467-479
106
102-110
336,370-382
12. Benedek, G. B. (1971) Appl. Opt. 10,459-473

14982 Phosphorylation of @-Crystallin B2 (@Bp)
13. Delaye, M., and Tardieu, A. (1983) Nature 302,415-417 14. Wistow, G. H., Mulders, J. W. M., and de Jong, W. W. (1987)
15. Wistow, G., and Piatigorsky, J. (1987) Science 236 , 1554-1556 16. Carper, D., Nishimura, C., Shinohara, T., Dietzchold, B., Wistow,
G., Craft, C., Kador, P., and Kinoshita, J. H. (1987) FEES
17. McDermott, M. J., Gawinowicz-Kolks, M. A., Chiesa, R., and Spector, A. (1988) Arch. Biochem. Biophys. 262,609-619
18. Spector, A., Chiesa, R., Sredy, J., and Garner, W. (1985) Proc. Natl. Acad. Sci. U. S. A. 82,4712-4716
19. Kleiman, N. J., Chiesa, R., Gawinowicz-Kolks, M. A., and Spec- tor, A. (1988) Invest. Ophthalmol. Visual Sci. 2 9 , (suppl.) 186
20. Voorter, C. E., Mulders, J. W. M., Bloemendal, H., and de Jong, W. W. (1986) Eur. J. Biochem. 160,203-210
21. Chiesa, R., Gawinowicz-Kolks, M. A., Kleiman, N. J., and Spec- tor, A. (1988) Exp. Eye Res. 4 6 , 199-208
22. Ambler, R. P. (1972) Methods Enzymol. 2 5 , 143-154 23. Chiesa, R., McDermott, M. J., Kleiman, N. J., and Spector, A.
Nature 326,622-624
Lett. 220 , 209-213
(1988) Invest. Ophthalmol. Visual Sci. 2 9 , (suppl.) 72
24.
25.
26.
27.
28.
29. 30.
31. 32. 33. 34. 35. 36.
Chiesa, R., Gawinowicz-Kolks, M. A., and Spector, A. (1987) J. Bid. Chm. 262,1438-1441
Chiesa, R., Gawinowicz-Kolks, M. A., Kleiman, N. J., and Spec- tor, A. (1987) Curr. Eye Res. 6,539-542
Chiesa, R., Gawinowicz-Kolks, M. A., Kleiman, N. J., and Spec- tor, A. (1987) Biochem. Bwphys. Res. Commun. 144, 1340- 1347
Cohen, P., Rylatt, D. B., and Nimmo, G. A. (1977) FEES Lett.
Chessa, G., Borin, G., Marchiori, G., Meggio, F., Brunati, A. M.,
Li, L.-K. (1978) Exp. Eye Res. 27, 553-566 Herbrink, P., Van Westreenen, H., and Bloemendal, H. (1975)
Laemmli, U. K. (1970) Nature 227 , 680-685 O'Farrell, P. H. (1975) J. Bwl. Chem. 250,4007-4021 Ui, N. (1971) Biochirn. Biophys. Acta 229,567-581 Ushiro, H., and Cohen, S. (1980) J. Biol. Chem. 255,8363-8365 Hayashi, R. (1977) Methods Enzymol. 47, 84-93 Reimann, E. M., and Beham, R. A. (1983) Methods Enzymol. 9 9 ,
76,182-186
and Pinna, L. A. (1983) Eur. J. Biochem. 135 , 609-614
Exp. Eye Res. 20, 541-548
51-55

Phosphorylation of P-Crystallin B2 (PBp) 14983
0
- Pi
- P-lyr
"P-Thr
"P-Ser





























