Embed Size (px)
Citation preview

PHYSIOLOGICAL ADAPTATIONS OF MANGROVE FLORA
OF COCHIN
u Introduction
Physiology of S a l t Tolerance
Adaptations to Respirat ion
Adaptations t o Reproduction
o Materials and Methods
Analysis o f P a lan ine
Data Analysis
u Results
u D ~ s c u s s i o n

PHYSlOLOGICkL ADAPTATIONS OF MANGROVE FLORA OF
COCHIN
INTRODUCTION
The physiological processes which allow mangroves to live in the
constantly changing environment where the land meets the sea, are unique among plants.
A mangrove must contend with drying effects of the sun and wind, osmotic imbalance
caused by the high salinity of seawater in which it is immersed, and growth in salty,
oxygen deficient, and waterlogged soils. Mangroves must also have the ability to
withstand the action of tides, exposure to freshwater, destruction effects of storm surges
and the diurnal and seasonal fluctuations of temperature.
Evolutionary adjustments to varying coastal marine environments have
produced some astounding biological characteristics within mangrove plant communities.
Although these plants grow in the areas which are well saturated with water yet they
cannot directly available to them because of high concentration of salts in the soil and
water. Thus mangroves are growing on soils of physically wet but physiologically dry in
nature (Sharrna, 1997). These plants also have to cope with the problems of salinity and
excess of water (Blascol 1075). The latter creates difficulty of anchoring, aeration and
seed germination Mangrove trees have specially adapted aerial and salt-filtering ~oo t s
and salt-secretin2 leaves that enable them to occupy the saline wetlands where other plant
life cannot susvive ( Snedaker, 1993)
In response to environmental extremes, mangroves have developed
specialised means in ordel to establish. grow and reproduce.

Physiology of Salt Tolerance
Mangroves have adapted to life in a saline environment in a number of
ways. Exclusion of sodlium ions and chloride ions at the roots by a process called ultra
filtration allows Brzibwiera g~mt~orhiza, A~~icet~tr ia rnarita. Rhizophora slylosa,
Sorit~eratia sps. fi,xcoecc~ria sps arrd Aegialitis sps to remove salt from seawater as water
is absorbed for normal plant physiological processes (Anonymous,l999b). This is a
passive process which does not require the expenditure of energy by the plant.
Salt exclusi~on mechanism may be performed due to the presence of ultra
filtration mechanism in their roots (Scholander, 1968). By this ultra filtration mechanism
when the water is absorbed the salt ion remain as filtered out (Tomlinson 1994). In
Aegicerns cort~rctrla/rrn, special salt secreting glands are present in their leaves. Salt
secretion is an active process which requires the expenditure of metabolic energy to
proceed. A white film of salt crystals usually forms on the upper leaf surface of salt
secreting mangroves. Salt may also be removed by storage in the older leaves which
eventually fall from the plimt.
Salt extrusion mechanisms controlled by the salt glands are present in the
leaves of A~~icrru~ru ntaiiria and Acutrthrr.~ sp.~. also. Joshi el a/.. 1975 have pointed out
that Avicrtnritr sp. are th~e most efficient salt extruding species and can grow in high saline
conditions. but Acntr/hrc!; sps. and Aeg~certrs .ys. mostly grow in less saline habitat.
Salt resistarlce of mangroves may be controlled by the presence of salt
gland, viviparous gel-mination and succulent nature of leaves (Naskar and Mandal, 1999).
I.rrt~~tritzet~tr racettro.s~r an,d ,Sotrtrr~.~r/~a .sp.s. accumulate extra salt in its salt glands, on both
sides of the leaves. as such leaves are succulent. Salt accumulation mechanisms in

mangroves are also preisent in the species like Excoecaria agallocha, Sorlrteratia and
Xylocarprrs sr). These mangrove species deposit the sodium and chloride ions in their
stem barks and pneumatophores or knee roots and in their older leaves, as well (Joshi, et
a1 ., 1972). Due to their salt accumulation abilities, these mangrove species mostly bear
succulent leaves (Jeenings, 1968)
Adaptations to Respiration
Some mangroves have specialised root structures to deal with growth in
saline and anaerobic :;oils which are periodically covered by seawater. To enable these
root systems to breathe during times of tidal drying, specialised aerial roots known as
pneumatophores serve: to aerate those parts of mangrove plants which are deprived of an
adequate oxygen supply and in particular those root structures buried in anaerobic soil.
Thick knobbly, 'bent-knee' roots are characteristic of Ceriops t a p / . Aeration in water
logged soils is achieved by the development of a surface systems o f prop roots around the
main trunk (Natarajan., 1'398).
'The prop roots of some mangrove species such as Rhizophora and the
pneumatophores of othel-s such as A~dcrr?tlin contain many small breathing pores, called
lenticels. 'These allow oxygen to diffuse into the plant and down to the underground roots
by means of air space tissue in the cortex called aerenchyma. The lenticels are inactive
durins hi& tide
Stilt ro'3ts are observed in R/tizo(,/tora, Hrr~grrirm arid Crricy).~. The stilt
root becornin3 shallow buttresses in old trees. In Soturera~ia root system including an
extended series of catlle roots giving rise to narrow, shallowly descending lateral roots
and erect pncunlatophores. Pneumatophores at first greenish gray with a flaky back,

extending to as much as 2m. at matuarity, tapering woody via secondary thickening and
with numerous narrow second order roots developing horizontally in the substrate
(Tomlinson, 1994). Root knees are seen in Brttguiera, where the knee is initiated as the
result of primary growth. Plank roots are found in Xylocarpus p z a t u r n and Heritiera.
The following mangroves normally lack any elaborated aerial part to the
root system,: Argiceras, AAegialiris, Exc:oecaria, Katzdelia and Nypa. However, certain
structures may be developed to aerate the root system. The base of the stem in Aegialiiis
is enlarged and tluted, with a very spongy texture. Karzdelia tends to develop aerial roots
in some limiting environments
Stmctulres regarded as hydathodes have been described in mangrove
leaves. I t must be emphasized that there is no record of guttation from mangrove leaves,
but this is not surprising because positive pressures in the xylem are hardly possible with
plants rooted in sea water. Stomata are scarcely sunken, but guard cells are somewhat
thick walled, ofien with prominent or even elaborated ledges, suggesting some increased
resistance to stomata1 1:ranspiration.
Adaptations to Reproduction
Mangroves are flowering plants with male and female flower parts
situated on either the same tree or on separate trees as in Excoecaria agaNocha L.. a
milky rnangrove Flowers and fruits may appear throughout the year according to
variations in regional climate. In I<hi:ophorn slylosa aerial roots usually produce baby
trees. The baby trees can float for a long time before it finally find a good place and settle
down. A V I C ~ I I I I ~ O ninritin and 1,rrgutlcrrlnrio rncmtosn are producing seeds.

Througll viviparity embryo germination begins on the tree itself; the tree
later drops its developed embryos, called seedlings, which may have take root in the soil
beneath. Viviparity may lhave evolved as an adaptive mechanism to prepare the seedlings
for long-distance dispersal, and survival and growth within a harsh saline environment.
During this viviparous development, the propagules are nourished on the parent tree, thus
accumulating the carbohydrates and other compounds required for later autonomous
growth. The structural complexity achieved by the seedlings at this early stage of plant
development helps acclimate the seedlings to extreme physical conditions which
otherwise might preclude normal seed germination (Snedaker, 1993). According to
Gunasekar, (1995) the concentrations of chlorophyll, total sugars and starch and net
photosynthetic rate increased during the viviparous germination of Hhizophora
nirrcrorlnicr and Rhizo,~>/iorn apicr~lntn hypocotyles.
Another special adaptation is the dispersal of certain mangrove propagules
which hang from the branches of mature trees. These fall off and eventually take root in
the soil around the parent tree or are carried to distant shorelines. Depending on the
species, this propagult:~ may float for extended periods upto a year, and still remain
viable Vivipary and the long lived propagules allow these mangrove species to disperse
over wide areas
Under saline conditions more aminoacids are formed in mangroves (Joshi
et (11.. 1977). This en,hances nitrogen metabolism. The nitrogen value in halophytes
ranges from 1 54 to 2.17 (Myers, 1990). High percentage of polyphenols occur in
A~Jcet~tr;<r r?ffici~wll.s. The Nacl- salinity results in accumulation of proline in halophytes.

The halophytes show 0.4 to 46% sodium and 0.24 to 2.72 % of Potassium (Dagar et
a1.,1991).
Joshi rt al. (1975) studied the photosynthetic carbon metabolism in the
prominent Indian mangrove species. They found that aspartate and alanine are the major
product of less than 10 seconds of photosynthesis in most of the Indian mangroves. These
initially synthesized aminoacids, provide carbon for production of larger organic
materials. Studies with carboxylating enzymes showed that phosphoenol pyruvate
carboxylase was more active and incorporates more carbon, than ribulose diphosphate
carboxylase Mangroves .are Cq types of plants and possibly of aspartate formers,
however they lack kranz type of anatomy unlike other Cq plants (Dagar et a/., 1991).
Stomata of mangroves open between 8 and 10 a.m. to facilitate maximum carbon
assimilation, but in the early afternoon as the temperature increases, they open to a slight
extent and in the evening they again close and remain closed all night.
Mangroves develop a tolerance to soil salinity because they maintain a
high cellular water potential and are relatively insensitive to salt toxicity. In India, studies
have also shown that different mangrove species display tolerance for a range of salinity
(Rao, 1986) The mechanisms of salt tolerance are complex and variable and involve
factors such as ionic potentials across membranes, osmotic relationships, enzyme
activation and protein synthesis (Tomlinson, 1994). The salt exclusion mechanism of
martgroves therefore must be selective. Decreased potassium concentration at high
salinities are noticed in Ai'icei~nitr and Argicerns. This is significant to their studies of
photosynthetic responses to high salinities since potassium ions are involved in the

stomata1 mechanism and indirectly the potassium concentration could affect gas
exchanse
In the present investigation quantity of P alanine content in different
species of mangroves in Cochin area were determined to find out the influence of P
alanine in their distribution. Alanine is a non protein aminoacid which has been detected
from natural sources. Many of these are of physiological importance and interfere in
important metabolic processes. The molecular formula of alanine is C,H702N. In P
alanine NH2 assumes the position of P . The structural formula of 13 alanine is
NH2-CHZ-CH2-COOH. Seasonal variation of P alanine content in the Acanthus ilicifolirrs
with respect to salinity istratss was also determined during this investigation.
MATERIALS AND METHODS
The fresh leaf samples of all the true mangrove species identified from the
sampling stations were collected and analysed for f3 alanine during premonsoon season
1999. Prenlonsoon season showed maximum water salinity during the investigation
period. Sampling was done from the stations where the mangrove species were
represented in rnaxirnum density 411 sample collections were made from the trees of 2
10 years oltl Ac.~urr/~rrs i:lIc~fi)Ii~i.s L was found distributed in maximum area at Cochin.
So monthly analysis of ,lclrt~/hrrs il/c!foli~r.s L. leaves were conducted for an year to find
out the seasonal variation of p alanine content. Leaves were collected 1 m above the
water level In order to find out the variation of !3 alanine content, 3 replications were
taken fro111 different plants of the same species at each station. Water salinity

quantification i the plants was also carried out along with the P alanine determination
in the plants
Tree size, morphological peculiarities, leaf size and texture of the samples
collected are given below.
Rhizophoru mucrongtay Lamk :- A glabrous evergreen tree with characteristic aerial stilt
roots, appearing butressed by the mud. Bark brown, with vertical clefts. The leaves are
typically much larger simple entire, opposite, blade elliptical, mucronate up to 10 cm
broad. Leaf texture coriaceous, glabrous, but with numerous microscopic cork warts on
the lower surface visible on older leaves as fine black dots. The height of the tree was
12 m and age of the tree was approximately 15 years.
Rhizc~phoru candelaria, DC. :- A glabrous small evergreen tree, with large dark glossy
green leaves. Bark brown. Height of the tree was 20 m and the age was 20 years
approximately Size of the leaves measured as 8.65 cm.
Bruguiera gymnorhiza L. :- Trees to 20 m high with short buttresses. Bark rough, black,
fissured pattern Shoots plagiotropic by apposition, developing terminal short shoots.
Leaves elliptic, oblong.. coriacious, I0 X 6 cm, apex bluntly pointed, petiole up to 4 cm
long often glaucous with a white wax. The age of the tree was approximately 20 years.
Ilruguieru cylindricu L. :- Tree to about 18 m high with short butresses, bark greyish.
Leaves elliptic. 8 X 5 cm, with a bluntly pointed apex and cuneate base, petiole to 4 cm
long. The age of the tret: was approximately 12 years.
Kundeliu currrlel L :- Small tree, ~r-oxiny to 5 m. Butresses and pneumatophores absent.
Bark smooth. greyish. Leaves opposite. 10 X 5 cm oblong elliptic, apex obtuse, margins
entire, petiole 1 5 crn long, age of the tree was aproximatelyltj years.

Sonneratia caseoiaris L :- Trees to 10 m with continuos growth but diffuse branching,
branches horizontal. Leaves glabrous, opposite, shortly petiolate to almost sessile. Adult
leaves broadly ovate ~isually with a blunt apex some what fleshy with an entire margin,
4 X 6 cm, Leaves lanciolate. with extended red petioles, root system including an
extended series of cable roots giving rise to narrow, shallowly descending lateral roots
and erect pneumatophores. Pneumatophores extending to as much as 2 m at maturity.
Age of the tree was 1 I years aproximately.
A~~iccnniu r$fcinalis L. :- A small tree to 9 m high much branched, dense crowned.
Aerial stilt roots presenlt. Under ground roots extended, cable like and supporting as
leteral branches, erect exposed preumatophores and descending absorbing roots. The bark
is smooth. lenticellate liight coloured and not fissured. Leaf shape is ovate and with
rounded apex The tree was approximately 12 years old.
Acanthus ilicifolius L. :- A low sprawling herb to a height of 2 m. Branching infrequent
and commonly from older parts Aerial roots from lower surface of reclining stems.
Leaves decussate. us~ially with a pair of spines at the insertion of each leaf Leaves
glabrous, petiole short (Icm), blade upto 10 cm long. Gradually tapered below. The apex
usually with a sinuous, spiny margin, broadly tridentate including an apical spine. Age of
the herb was approxiriately 20 years.
l%coecuriu ugall~~-lzo~ LA :- A dioecious tree to 14 m high with abundant white latex.
E3ark grey. lenticels PI-orninent on younger twigs. Leaves spirally arranged, shoots with
infrequent ditt'use brartchiny. Leaves simple, coriaceous with a terete petiole 2 cm long,
blade ovate-elliptic, up t'o 6 cm long and 4 cm wide. Apex rounded slightly emarginate.
Margin incotlspicuously notched. with a minute gland in the notch in young leaves. Basal

glands 2, usually I on each side of blade at its insertion on the petiole. Approximately
tree was with 10 ye;m old.
Excuecoriu indicu Willd :- A tree to about 15m high with thorny bark. Regularly
crenulate, almost lanceolate leaves. The age of the tree was approximately 20 years. Leaf
size of the tree was slightly bigger than the leaves of Fxcmcaria agailocha L.
(approximately 7cm 1.
ANALYSIS OF 0 A,LANINE
& ulunine sepurution 1y TIdC :- 10 gnis of raw leaves were taken and the tissue was first
pounded in a pestle and mortar into a pulp and then extracted with 80 % ethanol in water
directly. The mixtur'e was heated to about 70' to 80' during extraction. 'The pooled
extracts were centrifuge'd and the clear supernatant was concentrated. (Jayaraman, 1988).
6-alanine were separated by thin layer chromatography using silica gel G
coated plates. The solvent used was 96% ethanol - water (7:3 vfv) The spray reagent
used was 0 3 % solut~~on of Ninhydrin in butanol containing 3 ml acetic acid. The
coloured spots &ere developed by heatlng the plates at 110 '~ for 10 minutes. Ones the
colour had developed., the plates were exposed to vopours of concentrated ammonium
hydroxide which helped in the stabilization of colours. The Rf value was calculated. The
standard Rf value of D--alanine listed is 0.47. Then identified the spot of p-alanine
comparing with the standiud.
Quantitative Estimation of p- alanine (Colorimetric method):
Scratched the identified alanine spot completely and taken in a test tube.
The sttlve~it used was hlethylcellosolve. The concentration of it was calculated by

colorimetric analysils, by applying a derivation of Beer Lambert law (Wilson, 1995;
Concentration of Optical dencity of test solution X Concentration of standard the test solution :.
Optical dencity o f standard solution.
DATA ANALYSlS
Mean anti standard deviation of salinity and p-alanine were calculated for
the 3 sampling seasons (Snedecor and Cochran, 1967) ANOVA for seasonal data were
also computed using excel programme Results of the investigation is given in the form
tables and figures
RESCJLTS
Accordin!; to the nature of salt resistant adaptations the Cochin mangroves
can be categorized into three different types. They are described below:-
Hrrig~iiercr cvlir~driccr WA. i- Salt exclus~on 1 ype I~hiroj~horcr niricrorrn~tr Lamk
( I<hizophorn ccrr~u'rliirro DC.
Salt extrus~on type
ACLIII~/~II.Y ilic1f0llii.s I , .
Av~cr~i~r i t r t?fficirrcrli.s I L-

Salt accumulation type I t.;xcorcarra npNochn L
L
Morphological adaptations for the physiological need were also observed
in different mangrove species of the Cochin area. In Karumallor stations Soti~~rraila
c ~ . s c o l ~ ~ ~ . L exhibited branched and unbranched lengthy pneumatophores. The entire
leaf surtice of .lciri,r/hrr.s iltcif~litcs I ~ . . was covered by salt and the plant had stunted
gromth 31 Putll~li ypu station
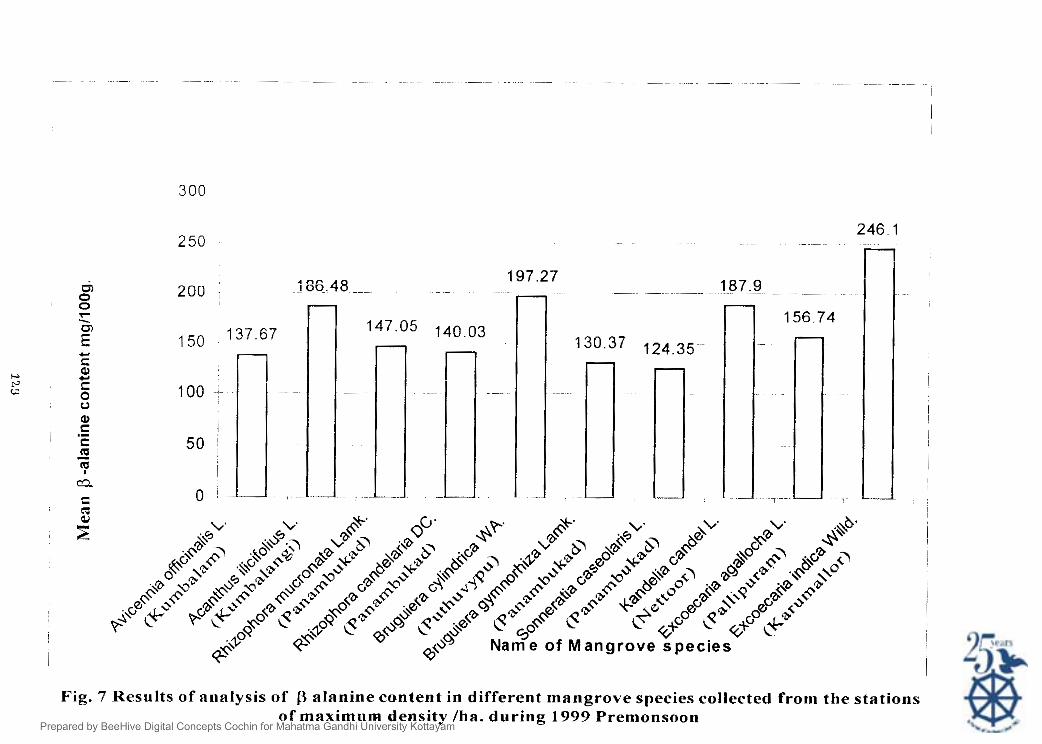
I he aminoacid 0 alanine was present in all the leaf samples analysed The
maximutii quantity of f3 alanine was obsened in k;xcorcnritr itrd;c.rr L, i r . . 246.10
i n t i e I ) . The next higher quantity was noticed in b'rrrg11ter.a cyIN~drictr
WA . Kur~tlrl~~r i<r11dt>l L and Arlnrlhr~s ilic~follrrs L. The rninimum !3 alanine content was
observed in Sotr~rrrtr/icr c n ~ ~ ~ o l ~ r ~ r . s L.: which was exclusively represented at stations such
as Nettoor. l'anarnbukad and Kammallor [Table XVII]. These stations showed a mean
salinity ol'7. 26 I and 7 9s 10~ ' l-espectively during the investigation period.
\ariation between the 13 alanine sample replications of all the mangrove
species showed insigfiifi~cant values. Leaf samples collected from the mangroves of the
same species and froni the same station showed more or less same P alanine content. The
amount 131' 11 alaliine content in the different mangrove species were dissimilar. Amino
acid content i r l iilansrove varies according to the salinity variation. alanine content of
Ac1t1 t11 I I I ~ I J / I I 1 . was lo\+ in tile salinity regions of stations 10 and 4. At station 9,
where sa l i~ i~ t \ \ \ d s the lwwest (1- 0 x 10~") Acotrtl~rr.~ ilic!fblirw L. was absent [Table XVlll
and Fi~ul-i. S /
4 ",- 4 " 100.40 --
I-! !
Fig. 7 Results o f analysis of alanine content in different mangrove species collected from the stations o f maximom density /ha. during 1999 Prenlonsoon

Table XVll
Results of a11alyse:s o'f p-alanine content (mgI100g.) in leaf samples collected during
True mangrove species and the Sample Sample Sample sites of collection -
1 I,I 1 Mean 1 Standard 1 I I 1 deviation
A13iceinricr offici~ru/i.c. . . L.
(Kumbalam)
Acotrthi~t 11r~~1filir1.s L.
(Kumbaiaiigi) ~ ~.--~
Hl~i:(y)hr~rrr n~~rcrot~u ln Lamk.
(Panambukad) - - Khlzophor~r ctr~rde/trrln DC.
(Panambuhad)
Hrtrguirlrr c:slirtdr.icr~ WA. 0.265
(Puthuvypu) - -~
Hrrrgrl~er~r mnlrrorhiza Lamk.
(Panambukad) ~ ~
Soinrrrnl~u ccr.s~~o1uri.c L.
(Panambukad) -~ -~
Klrirdellcr ccrit~/el I .
(Nettoor) - ~~
F;xcoecur-111 i ~ ~ ~ d l o c h r r L
i m r a 111 ) - - --- . -
Excoectwitr ~ialictr Wi lld.
(Karumallor) - ------ -

Table XVIlI
Station wise Seasonal Variation of Water Salinity and [3 Alanine content in
Acanthus ilicifulius observed during 1999
~~ -- .- I~
Salinity ppt. 1 a Alanine mg1100 gm
Mon- Postman- Mean soon soon soon soon
187.0 , 187.74
1 185.75 ) 185.54
2 6 % "6.80 " i ~ 8 . 0 1 187.40 1 187.5' 8 7 . 6 4
26 07 r 2 6 . I 1 187.20 187.01 187.00 187.07
186.48 186.51
- -~
2 8; .4cantl?us 111cifi)lius L. \\as 11ot found in this station -- -~
-370 3.70 1 130.53 1 130.09 1 130.10 / 130.24 I 19.78 --T19101i 180.50 j 180.64 180.59
__~ 1 - L -

STATIONS
SERIES 1 0 p-alanine mg/100g
1 SERIES 2 .+ Mean salinity ppt. 1 Fig 8 Station wise variation of water salinity and B alanine content in the leaves of
Acnntitus iZicifoNous L observed during 1999

The observation shows that salt extrusion type mangroves are most widely
distributed in Cochin area when compared to other species. The salt exclusion types like
Ka~tdrlru c<u~dt.f L. and Rhkopizorn c~zt~delc~riu DC. are found to have almost disappeared
from the Cochin area being limited to very restricted area. They require special protection
in this location otherwise they may be eliminated from the system. Similarly salt
accumulation types like fixcoecn/.irr it~dicn Willd. and So~r~~erafia careolari L. are also
rare.
'The p alanine content of different mangrove species differed. The analyses
ofAcatrrl~rr.s tlic.ifi,lir~s 1.. illustrates that alanine content is not constant for a species. It
varies with the salinity of the aquatic environment of mangrove ecosystem. Higher
salinity promoted the accumulation o f f \ alanine. It was also noted that irrespective of the
species, fi alanine content was constant in a particular locality. These results indicate a
strong correlation between salinity, f3 alanine and the distribution of the mangroves. j3
alanine does have a decissive role in the salt tolerance mechanism of the mangroves.
Physical conditions such as salinity and oxygen gradients determine the distribution of
plants (Robertson L,/ a / , 1991)
'Mangro.ves thrive in a very peculiar environment and serve as a bridging
ecosystem between lieshwater and marine systems. This has imposed several
modifications in these plants Accordins to Bhosale and Mulik (1992), aminoacids,
carbohydsates and polyphenols appear to be modified under saline conditions in
mangroves I'olyphenols seen to have a protective role to play under polluted conditions.

The polyphenoles are related to pollution stress, the nature of action is obscure (Kadam
and Bhosale. 1989). Similarly Mulik (1989) and Mulik and Bhosale (1989) reported that
the anlinoacid (3 alanine is found to play an important role in salt tolerance of these
plants The present results also suppons this view. In the investigation alanine content
in the leaves of ~L,'xcoecnr;tr itrdiccr Willd., Acuttfhr~s i/ic!folirts L., and Brtrgrriera
L ~ / / ~ I ~ / ~ I L ~ L I WA were found to be highest. The saline resistance of Acatrthrrs may be due
to its !?I alanine concentration. Nrrrgt~iertr cylitldrica WA, is observed in station 1, 6 and 7
where salinity value ranges between 29 to 33.9 x 10" . So this plant also has tolerated
high sal~nity stress. (3 alanine content is comparatively low in Sotrrrerafia ca.seolrrris L.
and Brrrprr~era jynr~~or,hizu Lamk. Both these species are found only in 3 stations.





























