Embed Size (px)
Citation preview

Plasticity in the adult human central auditory system:evidence from late-onset profound unilateral deafness
Curtis W. Ponton a;*, Juha-Pekka Vasama b, Kelly Tremblay c, Deepak Khosla d,Betty Kwong a, Manuel Don a
a Electrophysiology Laboratory, House Ear Institute, 2100 West Third Street, Los Angeles, CA 90057, USAb Department of Otolaryngology, Helsinki University Central Hospital, Haartmaninkatu 4 E, 00290 Helsinki, Finland
c Department of Speech and Hearing Sciences, 1417 NE 42nd Street, University of Washington, Seattle, WA 98105, USAd HRL Laboratories, LLC, 3011 Malibu Canyon Road, Malibu, CA 90265, USA
Received 7 September 2000; accepted 4 January 2001
Abstract
Experience-related changes in central nervous system (CNS) activity have been observed in the adult brain of many mammalianspecies, including humans. In humans, late-onset profound unilateral deafness creates an opportunity to study plasticity in the adultCNS consequent to monaural auditory deprivation. CNS activity was assessed by measuring long-latency auditory evoked potentials(AEPs) recorded from teens and adults with late-onset (post-childhood) profound unilateral deafness. Compared to monaurallystimulated normal-hearing subjects, the AEPs recorded from central electrode sites located over auditory cortical areas showedsignificant increases in inter-hemispheric waveform cross-correlation coefficients, and in inter-hemispheric AEP peak amplitudecorrelations. These increases provide evidence of substantial changes from the normal pattern of asymmetrical (contralater-als ipsilateral amplitude) and asynchronous (contralateral earlier than ipsilateral) central auditory system activation in the normal-hearing population to a much more symmetrical and synchronous activation in the unilaterally deaf. These cross-sectional analysesof AEP data recorded from the unilaterally deaf also suggest that the changes in cortical activity occur gradually and continue for atleast 2 years after the onset of hearing loss. Analyses of peak amplitude correlations suggest that the increased inter-hemisphericsymmetry may be a consequence of changes in the generators producing the N1 (approximately 100 ms peak latency) potential. Theseexperience-related changes in central auditory system activity following late-onset profound unilateral deafness thus provideevidence of the presence and the time course of auditory system plasticity in the adult brain. ß 2001 Elsevier Science B.V. Allrights reserved.
Key words: Plasticity; Auditory evoked potential ; Unilateral ; Deafness; Human
1. Introduction
Evidence of central nervous system (CNS) plasticity,de¢ned as an experience-related change in function oractivity (Greenough, 1975), has been observed in allsensory systems. In the auditory system, evidence ofCNS plasticity has been derived from studies of exper-imentally induced frequency-speci¢c peripheral hearing
loss or complete monaural deafness in non-humanmammals. Numerous investigations have demonstratedthat frequency-speci¢c hearing loss may change the re-sponse properties of neurons at both subcortical andcortical levels of the central auditory system. In tono-topically organized areas of auditory cortex, regionsdeprived of their normal peripheral input often becomeresponsive to intact adjacent frequencies (e.g., Willott etal., 1984, 1991; Robertson and Irvine, 1989; Kalten-bach et al., 1992; Rajan et al., 1993; Harrison et al.,1993; Schwaber et al., 1993). Evidence of central audi-tory system plasticity has been reported following ex-perimentally induced monaural deafness (e.g., Reale etal., 1987; Popelar et al., 1994). Changes in central audi-
0378-5955 / 01 / $ ^ see front matter ß 2001 Elsevier Science B.V. All rights reserved.PII: S 0 3 7 8 - 5 9 5 5 ( 0 1 ) 0 0 2 1 4 - 3
* Corresponding author. Present address: Neurosoft, Inc., 5700Cromo Drive, Suite 100, El Paso, TX 79912, USA.Tel. : +1-915-845-5600 ext. 28; Fax: +1-915-845-2965;E-mail: [email protected]
HEARES 3638 12-3-01
Hearing Research 154 (2001) 32^44
www.elsevier.com/locate/heares

tory pathway activation tend to be more extensive whensensory experience is modi¢ed soon after birth (e.g.,Popelar et al., 1994). However, experience-relatedchanges in CNS sensory and motor pathways havebeen reported in the adult brain of many mammals,including humans (Donoghue, 1995).
The central auditory system of mammals, includinghumans, includes a¡erents that project cortically on theside of the brain ipsilateral to the stimulated ear as wellas a¡erents that cross the midline at the level of thebrainstem and project to cortex on the contralateralside. The contralateral pathway contains a greater num-ber of nerve ¢bers and represents a more direct routewith fewer synapses to cortex than the ipsilateral path-way (Adams, 1979; Brunso-Bechtold et al., 1981; Cole-man and Clerici, 1987). Physiological studies in non-human mammals have shown that central auditory sys-tem activity evoked by monaural presentation is stron-ger and has lower activation thresholds in the contra-lateral than in the ipsilateral auditory pathway (Popelaret al., 1994; Reale et al., 1987; Kitzes, 1984). Thisasymmetrical activation of the central auditory systemalso occurs in humans. Thus, studies using monauralpresentation have consistently shown earlier and muchlarger evoked electrical potential and magnetoencepha-lographic (MEG) responses over the hemisphere contra-lateral to the stimulated ear (Vaughan and Ritter, 1970;Elberling et al., 1981; Pantev et al., 1986; Reite et al.,1988). Data from fMRI studies suggest an even moreasymmetric pattern of monaurally elicited cortical acti-vation (Sche¥er et al., 1998).
In non-human mammals, experimentally inducedprofound unilateral deafness signi¢cantly alters the nor-mally observed asymmetrical activation of the centralauditory system. Unilaterally deafened animals showmuch larger than normal responses and lower thresh-olds of activation in the pathway ipsilateral to the intactear, with little change in activity along the contralateralpathway. These changes are apparent subcortically inauditory nuclei such as the inferior colliculus (Reale etal., 1987; Kitzes, 1984, 1996), as well as at the level ofauditory cortex (Reale et al., 1987). While most studieshave focused on changes following neonatally induceddeafness, Popelar et al. (1994) demonstrated similarpatterns of change at both subcortical and cortical lev-els in unilaterally deafened adult guinea pigs. Hendryand Jones (1988) have suggested that a loss of inhibi-tory processes might account for the increased ipsilat-eral pathway activity. Alternatively, the data of Kitzes(1996) suggest that, at least in neonatally deafened ani-mals, the increased activity may re£ect the emergence ofadditional a¡erent ¢bers in the ipsilateral pathway.
In humans, the changes in cortical activity followingprofound unilateral deafness are consistent with thosethat follow experimentally induced unilateral hearing
loss in non-human mammals. Thus, MEG studies in-cluding individuals with early- and adult-onset pro-found unilateral deafness have shown both an increasein cortical activation ipsilateral to the intact ear andevidence of additional activation in other cortical areas(Vasama et al., 1994; Vasama and Ma«kela«, 1995; Fu-juki et al., 1998). These ¢ndings are corroborated bybrain imaging data from Sche¥er et al. (1998) whoused fMRI to compare patterns of cortical activationproduced by monaural stimulation in normal-hearingand unilaterally deaf adults. The unilaterally deaf adultshad much more symmetrical cortical activation of audi-tory areas. Sche¥er et al.'s results included data from amixed group containing individuals with early- andlate-onset unilateral deafness. These data would suggestthat changes in central auditory activation patterns oc-cur even with late-onset deafness. However, the extentto which such changes occur exclusively in individualswith late-onset profound deafness is not known.
The goals of this study were to investigate the extentto which cortical activation changes following late-on-set (teen-aged and older) profound unilateral deafnessand to describe the time course of these changes. Theresults from previous studies indicate that while activityin the contralateral pathway is unchanged by unilateraldeafness, excitation of the pathway ipsilateral to theintact ear is enhanced. This suggests that a commonactivation path of the contralateral and ipsilateral path-ways by the intact ear is created or enhanced. Onemanifestation of an increased common activation ofthe contralateral and ipsilateral pathways may be anincrease in the correlation of activity between the con-tralateral and ipsilateral hemispheres. Increased inter-hemispheric correlations can be demonstrated by (1) achange in the timing of activity between the hemi-spheres, and (2) a more consistent pattern of ipsilater-al/contralateral response amplitudes across individuals.
Changes in cortical activity associated with profoundunilateral deafness were assessed by measuring thelong-latency auditory evoked potentials (AEPs). Long-latency AEPs provide non-invasive, objective measuresof human CNS activity that primarily re£ect synchro-nous neural activation of structures in the thalamic-cor-tical segment of the central auditory system. The sub-ject population included teens and adults with late-onset (post-childhood) profound unilateral deafnesswhose AEPs were compared with those of normal-hear-ing adults. Two approaches were used to assess inter-hemispheric correlations of evoked auditory activity.First, cross-correlation coe¤cients (zero time lag) werecalculated between AEP waveforms from electrodepairs located at homologous locations over each hemi-sphere. Cross-correlation coe¤cients are, because of theinherent normalization, amplitude-independent. Thesecoe¤cients provided an indication of whether there
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^44 33

were signi¢cant di¡erences between the unilaterally deafand normal-hearing subjects in the AEP waveformmorphology and timing of inter-hemispheric activity.Second, peak-to-peak and baseline-to-peak amplitudecorrelations were generated for homologous electrodepairs located over each hemisphere. Third, the resultsof MEG and fMRI studies (Vasama et al., 1994; Vasa-ma and Ma«kela«, 1995; Fujuki et al., 1998; Sche¥er etal., 1998) indicated that the measured changes in hu-man cortical activity following profound unilateraldeafness originate from auditory cortical areas. There-fore, analyses of inter-hemispheric peak amplitude cor-relations focused on AEPs recorded from three pairs ofcentral scalp electrodes located over ipsilateral and con-tralateral auditory cortical areas. These analyses exam-ined the inter-hemispheric amplitude correlations of thecommonly measured peaks of the AEP including P1
(mean latency = 50 ms), N1 (mean latency = 100 ms),and P2 (mean latency = 150 ms). The P1 and N1 peaksare generally considered to re£ect activation of neuralgenerators speci¢c to the auditory thalamic-corticalpathway (e.g., Na«a«ta«nen and Picton, 1987; Lie«geois-Chauvel et al., 1994; Ma«kela« and McEvoy, 1996). Inparticular, evidence suggests that neural generators ofthe N1 peak at least partly re£ect re-entrant or recur-rent activation of auditory cortical areas between hemi-spheres (Ma«kela« and Hari, 1992; Ma«kela« and McEvoy,1996). In contrast, the P2 peak does not appear to orig-inate from the temporal lobes. This peak may at leastpartly re£ect output from the mesencephalic reticularactivating system, which responds to input from allsensory modalities (Yingling and Skinner, 1977; Skin-ner and Yingling, 1977; Knight et al., 1980, 1988; Na«a«-ta«nen and Picton, 1987; Rif et al., 1991; Woods et al.,1993). Since these commonly measured AEP peaks ap-pear to have di¡erent underlying neural generators, thee¡ects of profound unilateral deafness may not be thesame. Di¡erences in the correlations may provide infor-mation regarding the origin(s) of observed changes inhuman cortical activity.
2. Materials and methods
2.1. Subjects
AEPs were recorded from 15 teens and adults withprofound unilateral deafness: seven with right-sidedand eight with left-sided deafness. These subjectsranged in age from 17 to 67 years (mean age 43.7years). Profound unilateral deafness was the result ofsurgical treatment for acoustic neuroma or other oto-logic disorders (12 subjects) as well as sudden sensori-neural hearing loss and meningitis (three subjects). Theaverage age at onset of deafness was 40.8 years (ranging
from 14.0 to 65.8 years). The duration of unilateralhearing loss prior to AEP testing ranged from lessthan 1 year (for three subjects) to 13.7 years. Theaudiograms for their intact ears were normal (9 25 dBHL) from 0.25 to 4.0 kHz and none of the subjectsreported any history of other central neurological dis-orders. The normal-hearing group consisted of nineneurologically intact adults 20^38 years of age (meanage 32.8 years). Informed consent was obtained fromeach subject prior to testing. The test protocol for thisinvestigation was approved by an institutional reviewboard for human subjects. All testing was performedin accordance with the guidelines of the Declarationof Helsinki.
2.2. Procedure
Subjects were stimulated with clicks generated by ap-plying 100 Ws voltage pulses to a Sony earbud head-phone. Each stimulus presentation consisted of a brieftrain of 10 clicks which was repeated at a rate of 1.3/s.The interval between the onsets of successive clickswithin the train was 2 ms. All subjects perceived thisstimulus train as a single auditory event. Stimuli werepresented monaurally at the same physical level for allsubjects; this level was approximately 65 dB SL abovethreshold based on testing of a small number of nor-mal-hearing subjects. For normal-hearing subjects, eachear was tested separately.
All subjects were tested while seated in a comfortablereclining chair in an electrically shielded, sound-attenu-ated booth. The AEPs were recorded at 30 standardelectrode locations on the scalp. Four electrodes werelocated on the midline of the head (Fz, Cz, Pz, and Oz),13 electrodes were located over the hemisphere contra-lateral to the stimulated ear, and 13 electrodes werelocated over the ipsilateral hemisphere at locations ho-mologous to those on the contralateral side. Activityfrom each electrode was recorded with reference to aforehead electrode located at Fpz. To record the AEPs,single-sweep EEG epochs were sampled and recorded ata rate of 1 kHz from 100 ms prior to stimulus onset to400 ms after stimulus onset using a Neuroscan1 elec-trophysiological data acquisition system. O¡-line, thesingle-sweep epochs were baseline-corrected by sub-tracting the average DC o¡set from the prestimulusinterval (3100^0 ms) from the entire AEP epoch. Thesingle AEP epochs were then subjected to an automaticartifact rejection algorithm: an epoch in which anychannel containing activity exceeding þ 150 WV was re-jected from further processing. To minimize the impactof eye movements on the AEP data, a regression algo-rithm was applied that reduces the e¡ects of ocularcontamination on a¡ected AEP epochs. The acceptedsweeps (at least 1500 for each subject) were then aver-
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^4434
aged. Prior to analysis, the averages were bandpass-¢l-tered from 1 to 70 Hz (24 dB/octave) and average-ref-erenced (the average of activity measured at all electro-des was subtracted from each electrode).
2.3. Data analyses
The AEP data were analyzed with respect to whetherthe activity was recorded over the hemisphere contra-lateral or ipsilateral to the stimulated ear. Thus, ear ofstimulation (i.e., left or right) was not considered in thisanalysis. To determine whether the data obtained inthis study were consistent with previous ¢ndings (i.e.,increased ipsilateral activation), the ¢rst analyses com-pared root mean square (RMS) ipsilateral and contra-lateral AEP amplitudes both within and across subjectgroups. The AEP RMS amplitudes were measured fromstimulus onset to the end of the recording epoch 400 mslater. The contralateral/ipsilateral amplitude contrastswere performed by generating for each subject average
RMS values of all electrodes over each hemisphere (ex-cluding midline electrodes).
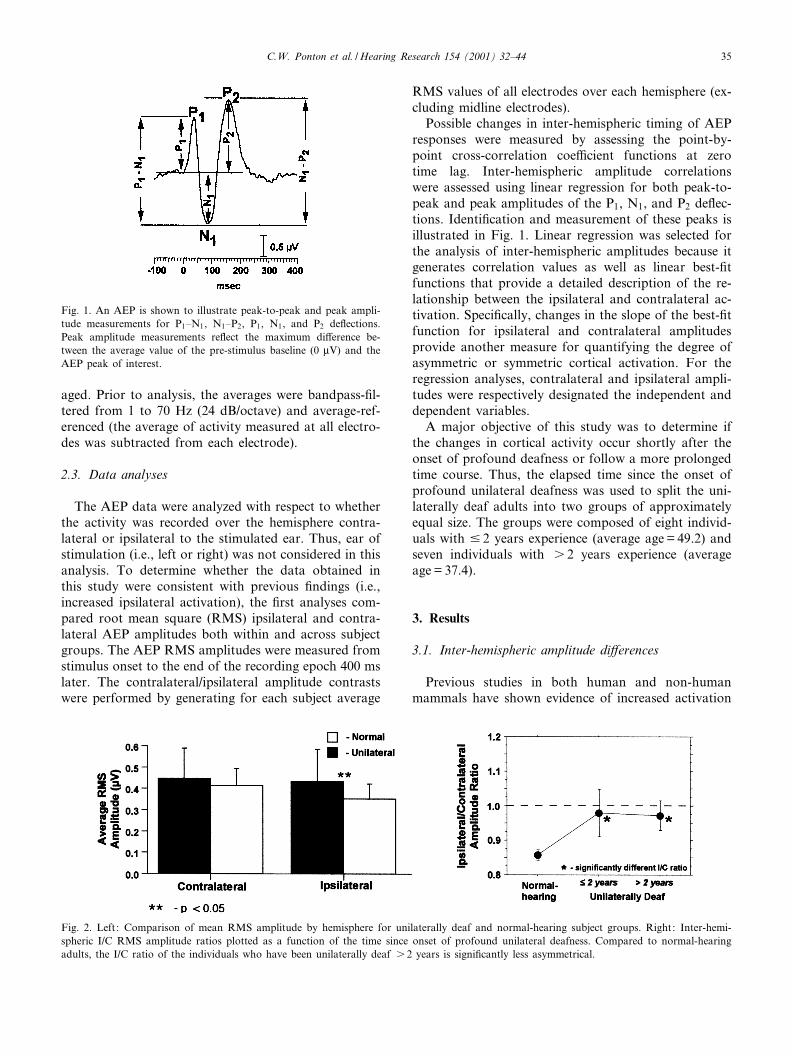
Possible changes in inter-hemispheric timing of AEPresponses were measured by assessing the point-by-point cross-correlation coe¤cient functions at zerotime lag. Inter-hemispheric amplitude correlationswere assessed using linear regression for both peak-to-peak and peak amplitudes of the P1, N1, and P2 de£ec-tions. Identi¢cation and measurement of these peaks isillustrated in Fig. 1. Linear regression was selected forthe analysis of inter-hemispheric amplitudes because itgenerates correlation values as well as linear best-¢tfunctions that provide a detailed description of the re-lationship between the ipsilateral and contralateral ac-tivation. Speci¢cally, changes in the slope of the best-¢tfunction for ipsilateral and contralateral amplitudesprovide another measure for quantifying the degree ofasymmetric or symmetric cortical activation. For theregression analyses, contralateral and ipsilateral ampli-tudes were respectively designated the independent anddependent variables.
A major objective of this study was to determine ifthe changes in cortical activity occur shortly after theonset of profound deafness or follow a more prolongedtime course. Thus, the elapsed time since the onset ofprofound unilateral deafness was used to split the uni-laterally deaf adults into two groups of approximatelyequal size. The groups were composed of eight individ-uals with 9 2 years experience (average age = 49.2) andseven individuals with s 2 years experience (averageage = 37.4).
3. Results
3.1. Inter-hemispheric amplitude di¡erences
Previous studies in both human and non-humanmammals have shown evidence of increased activation
Fig. 1. An AEP is shown to illustrate peak-to-peak and peak ampli-tude measurements for P1^N1, N1^P2, P1, N1, and P2 de£ections.Peak amplitude measurements re£ect the maximum di¡erence be-tween the average value of the pre-stimulus baseline (0 WV) and theAEP peak of interest.
Fig. 2. Left: Comparison of mean RMS amplitude by hemisphere for unilaterally deaf and normal-hearing subject groups. Right: Inter-hemi-spheric I/C RMS amplitude ratios plotted as a function of the time since onset of profound unilateral deafness. Compared to normal-hearingadults, the I/C ratio of the individuals who have been unilaterally deaf s 2 years is signi¢cantly less asymmetrical.
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^44 35
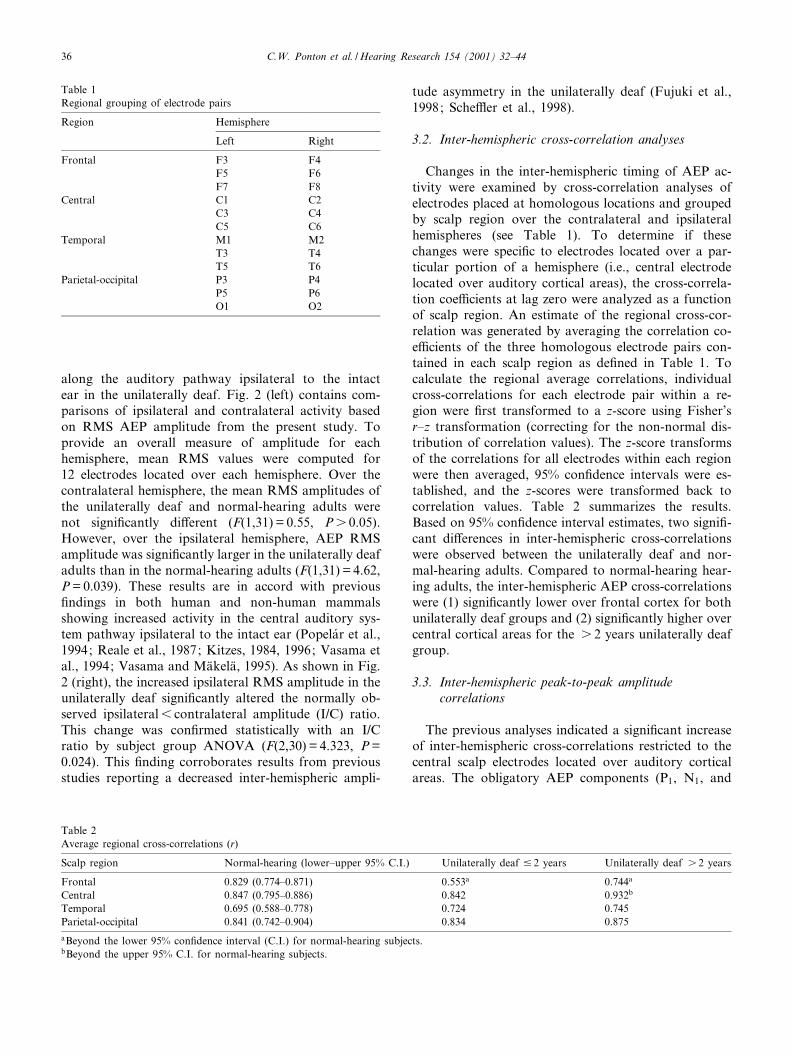
along the auditory pathway ipsilateral to the intactear in the unilaterally deaf. Fig. 2 (left) contains com-parisons of ipsilateral and contralateral activity basedon RMS AEP amplitude from the present study. Toprovide an overall measure of amplitude for eachhemisphere, mean RMS values were computed for12 electrodes located over each hemisphere. Over thecontralateral hemisphere, the mean RMS amplitudes ofthe unilaterally deaf and normal-hearing adults werenot signi¢cantly di¡erent (F(1,31) = 0.55, Ps 0.05).However, over the ipsilateral hemisphere, AEP RMSamplitude was signi¢cantly larger in the unilaterally deafadults than in the normal-hearing adults (F(1,31) = 4.62,P = 0.039). These results are in accord with previous¢ndings in both human and non-human mammalsshowing increased activity in the central auditory sys-tem pathway ipsilateral to the intact ear (Popelar et al.,1994; Reale et al., 1987; Kitzes, 1984, 1996; Vasama etal., 1994; Vasama and Ma«kela«, 1995). As shown in Fig.2 (right), the increased ipsilateral RMS amplitude in theunilaterally deaf signi¢cantly altered the normally ob-served ipsilateral6 contralateral amplitude (I/C) ratio.This change was con¢rmed statistically with an I/Cratio by subject group ANOVA (F(2,30) = 4.323, P =0.024). This ¢nding corroborates results from previousstudies reporting a decreased inter-hemispheric ampli-
tude asymmetry in the unilaterally deaf (Fujuki et al.,1998; Sche¥er et al., 1998).
3.2. Inter-hemispheric cross-correlation analyses
Changes in the inter-hemispheric timing of AEP ac-tivity were examined by cross-correlation analyses ofelectrodes placed at homologous locations and groupedby scalp region over the contralateral and ipsilateralhemispheres (see Table 1). To determine if thesechanges were speci¢c to electrodes located over a par-ticular portion of a hemisphere (i.e., central electrodelocated over auditory cortical areas), the cross-correla-tion coe¤cients at lag zero were analyzed as a functionof scalp region. An estimate of the regional cross-cor-relation was generated by averaging the correlation co-e¤cients of the three homologous electrode pairs con-tained in each scalp region as de¢ned in Table 1. Tocalculate the regional average correlations, individualcross-correlations for each electrode pair within a re-gion were ¢rst transformed to a z-score using Fisher'sr^z transformation (correcting for the non-normal dis-tribution of correlation values). The z-score transformsof the correlations for all electrodes within each regionwere then averaged, 95% con¢dence intervals were es-tablished, and the z-scores were transformed back tocorrelation values. Table 2 summarizes the results.Based on 95% con¢dence interval estimates, two signi¢-cant di¡erences in inter-hemispheric cross-correlationswere observed between the unilaterally deaf and nor-mal-hearing adults. Compared to normal-hearing hear-ing adults, the inter-hemispheric AEP cross-correlationswere (1) signi¢cantly lower over frontal cortex for bothunilaterally deaf groups and (2) signi¢cantly higher overcentral cortical areas for the s 2 years unilaterally deafgroup.
3.3. Inter-hemispheric peak-to-peak amplitudecorrelations
The previous analyses indicated a signi¢cant increaseof inter-hemispheric cross-correlations restricted to thecentral scalp electrodes located over auditory corticalareas. The obligatory AEP components (P1, N1, and
Table 1Regional grouping of electrode pairs
Region Hemisphere
Left Right
Frontal F3 F4F5 F6F7 F8
Central C1 C2C3 C4C5 C6
Temporal M1 M2T3 T4T5 T6
Parietal-occipital P3 P4P5 P6O1 O2
Table 2Average regional cross-correlations (r)
Scalp region Normal-hearing (lower^upper 95% C.I.) Unilaterally deaf 9 2 years Unilaterally deaf s 2 years
Frontal 0.829 (0.774^0.871) 0.553a 0.744a
Central 0.847 (0.795^0.886) 0.842 0.932b
Temporal 0.695 (0.588^0.778) 0.724 0.745Parietal-occipital 0.841 (0.742^0.904) 0.834 0.875aBeyond the lower 95% con¢dence interval (C.I.) for normal-hearing subjects.bBeyond the upper 95% C.I. for normal-hearing subjects.
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^4436
P2) are typically most robust at these scalp electrodelocations. Therefore, linear regression analyses for in-ter-hemispheric peak-to-peak AEP amplitudes focusedon electrodes from the central scalp region. To providea more representative measure of inter-electrode ipsilat-
eral/contralateral amplitude variability within the cen-tral scalp region, peak amplitude data for all electrodepairs (C1^C2, C3^C4, and C5^C6) from this regionwere included in these analyses. Fig. 3 shows individualipsilateral peak-to-peak amplitudes plotted as a func-
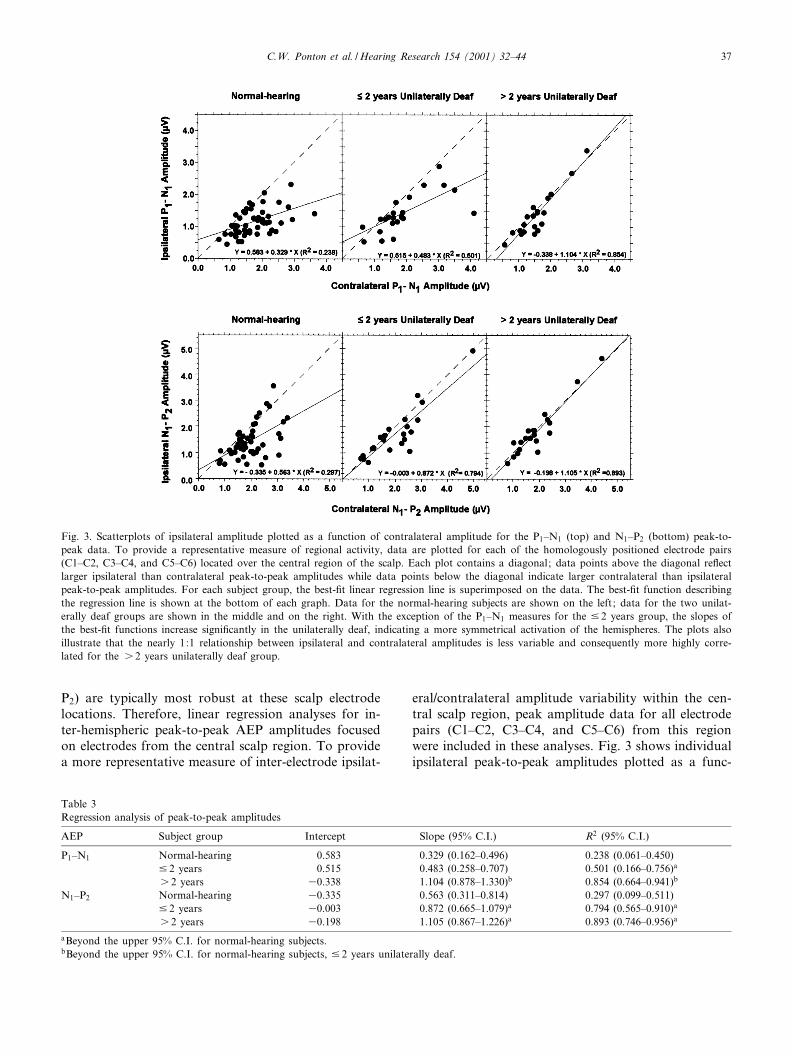
Fig. 3. Scatterplots of ipsilateral amplitude plotted as a function of contralateral amplitude for the P1^N1 (top) and N1^P2 (bottom) peak-to-peak data. To provide a representative measure of regional activity, data are plotted for each of the homologously positioned electrode pairs(C1^C2, C3^C4, and C5^C6) located over the central region of the scalp. Each plot contains a diagonal; data points above the diagonal re£ectlarger ipsilateral than contralateral peak-to-peak amplitudes while data points below the diagonal indicate larger contralateral than ipsilateralpeak-to-peak amplitudes. For each subject group, the best-¢t linear regression line is superimposed on the data. The best-¢t function describingthe regression line is shown at the bottom of each graph. Data for the normal-hearing subjects are shown on the left; data for the two unilat-erally deaf groups are shown in the middle and on the right. With the exception of the P1^N1 measures for the 9 2 years group, the slopes ofthe best-¢t functions increase signi¢cantly in the unilaterally deaf, indicating a more symmetrical activation of the hemispheres. The plots alsoillustrate that the nearly 1:1 relationship between ipsilateral and contralateral amplitudes is less variable and consequently more highly corre-lated for the s 2 years unilaterally deaf group.
Table 3Regression analysis of peak-to-peak amplitudes
AEP Subject group Intercept Slope (95% C.I.) R2 (95% C.I.)
P1^N1 Normal-hearing 0.583 0.329 (0.162^0.496) 0.238 (0.061^0.450)9 2 years 0.515 0.483 (0.258^0.707) 0.501 (0.166^0.756)a
s 2 years 30.338 1.104 (0.878^1.330)b 0.854 (0.664^0.941)b
N1^P2 Normal-hearing 30.335 0.563 (0.311^0.814) 0.297 (0.099^0.511)9 2 years 30.003 0.872 (0.665^1.079)a 0.794 (0.565^0.910)a
s 2 years 30.198 1.105 (0.867^1.226)a 0.893 (0.746^0.956)a
aBeyond the upper 95% C.I. for normal-hearing subjects.bBeyond the upper 95% C.I. for normal-hearing subjects, 9 2 years unilaterally deaf.
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^44 37
tion of the corresponding homologous contralateralamplitudes for P1^N1 (top) and N1^P2 (bottom). Abest-¢t linear regression line is superimposed on eachof the data sets. The parameters for each best-¢t linearfunction (intercept, slope, and R2), as well as 95% con-¢dence intervals (95% C.I.) for slope and R2, are shownin Table 3. For P1^N1, shown at the top of Fig. 3, theregression analyses indicated signi¢cant linear correla-tions between contralateral and ipsilateral amplitudes
for the normal-hearing adults and for each of the uni-laterally deaf subject groups (P6 0.001 or better foreach group). The analyses also revealed di¡erences be-tween the normal-hearing and unilaterally deaf groups.As shown in Table 3, the slope of the best-¢t regressionline was signi¢cantly steeper (beyond the upper 95%C.I.) for the s 2 years unilaterally deaf group thanfor the normal-hearing adults and the 9 2 years unilat-erally deaf group. In addition, there was a signi¢cant
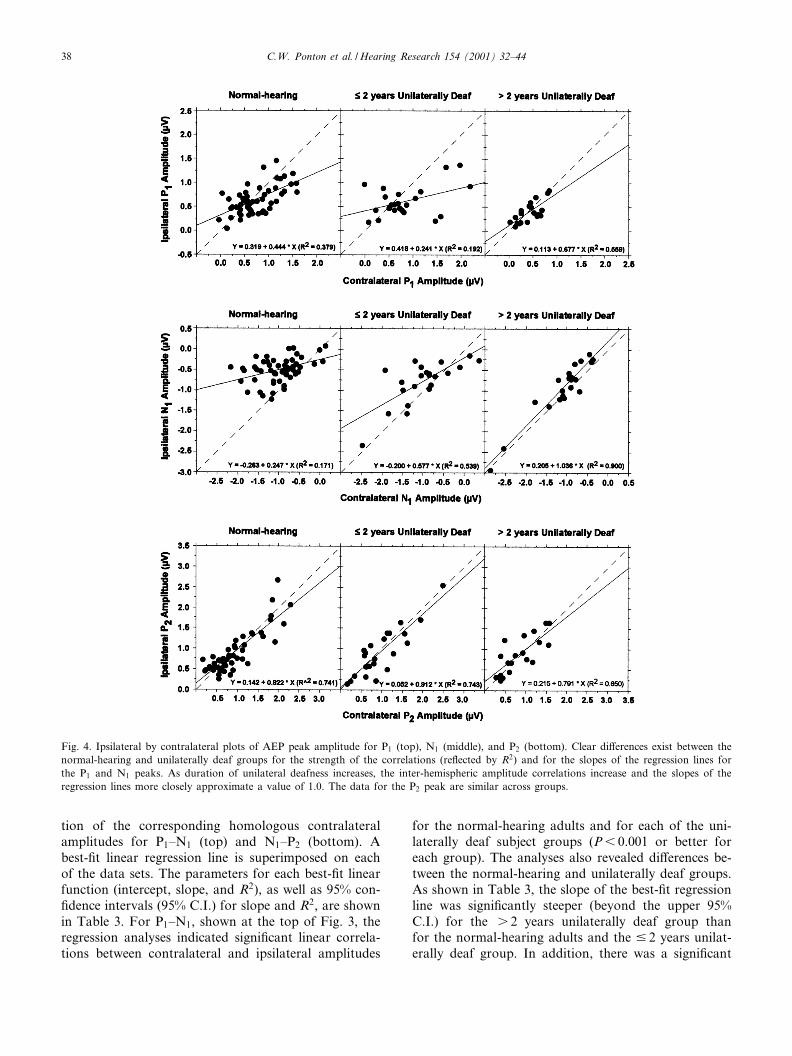
Fig. 4. Ipsilateral by contralateral plots of AEP peak amplitude for P1 (top), N1 (middle), and P2 (bottom). Clear di¡erences exist between thenormal-hearing and unilaterally deaf groups for the strength of the correlations (re£ected by R2) and for the slopes of the regression lines forthe P1 and N1 peaks. As duration of unilateral deafness increases, the inter-hemispheric amplitude correlations increase and the slopes of theregression lines more closely approximate a value of 1.0. The data for the P2 peak are similar across groups.
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^4438

increase in the inter-hemispheric correlation (as repre-sented by R2) for P1^N1 peak-to-peak amplitude be-tween the normal-hearing group and each of the unilat-erally deaf groups. Moreover, the correlation wassigni¢cantly stronger (beyond the upper 95% C.I.) forthe s 2 years deaf than the 9 2 years deaf.
For the N1^P2 peak-to-peak comparisons, the inter-hemispheric amplitude correlations were signi¢cant forthe normal-hearing adults and for each of the unilater-ally deaf subject groups (P6 0.001 or better for eachgroup). For the s 2 years deaf group, the 1.105 slopevalue of the N1^P2 best-¢t regression line was signi¢-cantly steeper than that obtained for the 9 2 years deafor the normal-hearing groups. Furthermore, this valuewas virtually the same as the slope of the best-¢t func-tion for inter-hemispheric P1^N1 analysis (slope =1.104). As shown in Table 3, the N1^P2 inter-hemi-spheric amplitude correlations were signi¢cantly stron-ger (beyond the upper 95% C.I.) in the two unilaterallydeaf groups than in normal-hearing subjects. Whilethere was a trend toward an experience-related increasein slope and correlation values for the N1^P2 peak-to-peak amplitude data, the di¡erences between the twounilaterally deaf groups were not statistically signi¢-cant.
3.4. Individual AEP peak inter-hemispheric amplitudecorrelations
The inter-hemispheric peak amplitude correlationsfor the individual AEP peaks P1, N1, and P2, are shownin Fig. 4 and described in Table 4. Similar to the resultsobtained for the analysis of peak-to-peak amplitude,inter-hemispheric amplitude correlations for each indi-vidual peak were strongly signi¢cant for all groups(P6 0.001 for each peak, each group). However, anal-yses of the inter-hemispheric peak amplitude correla-tions suggested that the di¡erences between subjectgroups were peak-dependent.
For the P1 peak, di¡erences between the normal-
hearing and unilaterally deaf subjects were mixed. Com-pared to the normal-hearing subjects, the slope of theregression line was signi¢cantly more shallow for thoseindividuals deaf 9 2 years, but signi¢cantly steeper forthose deaf s 2 years. There were no signi¢cant di¡er-ences in the P1 inter-hemispheric amplitude correlationsbetween subject groups as re£ected by the R2 valuescontained in Table 4. The results of the analyses forinter-hemispheric N1 peak amplitude correlationssharply contrast with those for P1. Compared to thedata for the normal-hearing adults, the best-¢t func-tions for both unilaterally deaf groups had signi¢cantlysteeper slopes with values much closer to 1.0, indicatingmore symmetrical ipsilateral/contralateral activation.For the unilaterally deaf subjects, the slope of thebest-¢t function increased signi¢cantly from 0.577 forthe 9 2 years group to 1.036 for the s 2 years group.Three-point slope values for contralateral/ipsilateralamplitudes were also calculated on a per subject basis.This analysis showed that ¢ve of seven subjects unilat-erally deaf for s 2 years had slope values greater than0.5, compared to two of eight unilaterally deaf for 9 2years, and one of nine normal-hearing subjects. Basedon R2 measures (and 95% con¢dence intervals), contra-lateral and ipsilateral N1 amplitudes were signi¢cantlymore correlated in the s 2 years deaf subjects than inthe normal-hearing subjects. The contralateral and ipsi-lateral N1 amplitudes were also signi¢cantly more cor-related in s 2 years deaf than in the 9 2 years deafgroup, suggesting an experience-related change in thegenerator(s) of the N1 AEP. As shown for the P2
peak in Table 4, the linear regression analyses indicatedno di¡erences in inter-hemispheric correlations betweenany of the subject groups. The slopes of the best-¢tfunctions for P2 amplitude were uniformly high(0.791^0.902), as were the R2 values (0.741^0.844).
3.5. Individual and grand mean AEP waveforms
The AEP waveforms in Fig. 5a (individual subjects)
Table 4Regression analysis of peak amplitudes
AEP Subject group Intercept Slope (95% C.I.) R2 (95% C.I.)
P1 Normal-hearing 0.319 0.444 (0.282^0.605) 0.379 (0.169^0.579)9 2 years 0.418 0.241 (0.017^0.464)a 0.192 (0.001^0.518)s 2 years 0.133 0.677 (0.379^0.975)c 0.559 (0.208^0.799)
N1 Normal-hearing 30.263 0.247 (0.093^0.401) 0.171 (0.025^0.381)9 2 years 30.200 0.577 (0.328^0.826)b 0.539 (0.205^0.779)b
s 2 years 0.205 1.036 (0.871^1.202)c 0.900 (0.767^0.958)c
P2 Normal-hearing 0.142 0.822 (0.681^0.963) 0.741 (0.586^0.844)9 2 years 0.030 0.912 (0.668^1.155) 0.743 (0.485^0.883)s 2 years 0.215 0.791 (0.504^1.079) 0.650 (0.320^0.848)
aBeyond the lower 95% C.I. for normal-hearing subjects.bBeyond the upper 95% C.I. for normal-hearing subjects.cBeyond the upper 95% C.I. for normal-hearing subjects and 9 2 years unilaterally deaf.
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^44 39
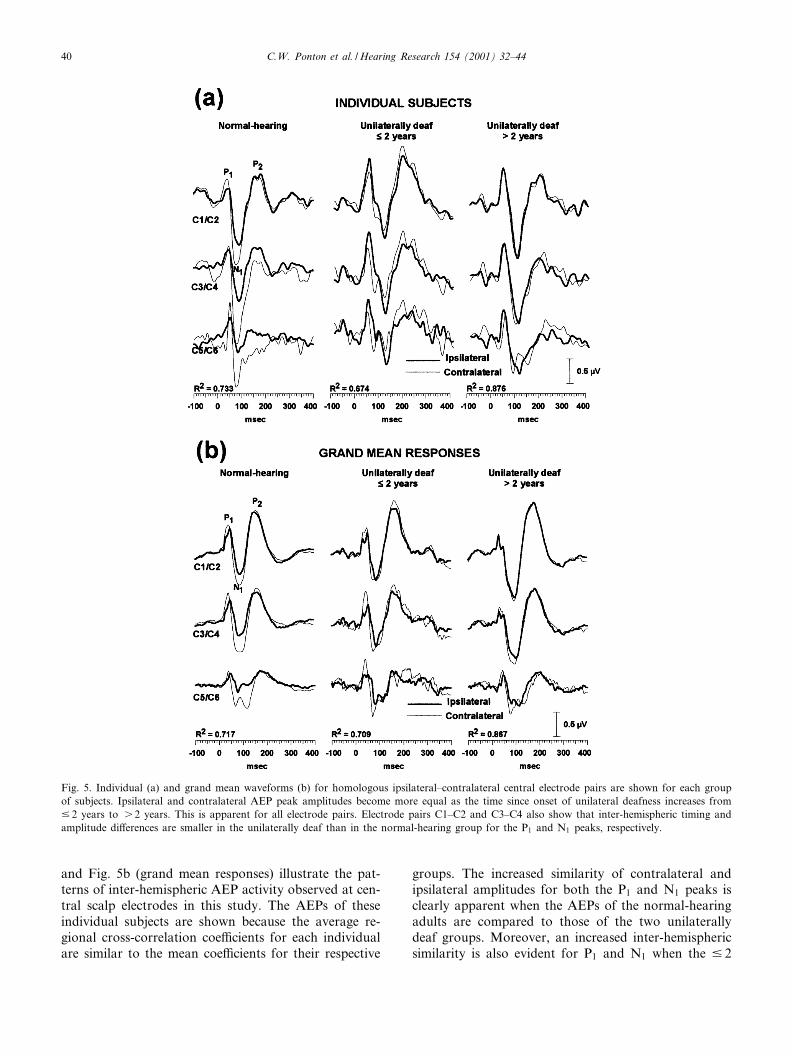
and Fig. 5b (grand mean responses) illustrate the pat-terns of inter-hemispheric AEP activity observed at cen-tral scalp electrodes in this study. The AEPs of theseindividual subjects are shown because the average re-gional cross-correlation coe¤cients for each individualare similar to the mean coe¤cients for their respective
groups. The increased similarity of contralateral andipsilateral amplitudes for both the P1 and N1 peaks isclearly apparent when the AEPs of the normal-hearingadults are compared to those of the two unilaterallydeaf groups. Moreover, an increased inter-hemisphericsimilarity is also evident for P1 and N1 when the 9 2
Fig. 5. Individual (a) and grand mean waveforms (b) for homologous ipsilateral^contralateral central electrode pairs are shown for each groupof subjects. Ipsilateral and contralateral AEP peak amplitudes become more equal as the time since onset of unilateral deafness increases from9 2 years to s 2 years. This is apparent for all electrode pairs. Electrode pairs C1^C2 and C3^C4 also show that inter-hemispheric timing andamplitude di¡erences are smaller in the unilaterally deaf than in the normal-hearing group for the P1 and N1 peaks, respectively.
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^4440

years deaf are compared to the s 2 years deaf. As in-dicated by the statistical analyses, the timing and am-plitude of the P2 peak appears quite similar across sub-ject groups.
4. Discussion
The results of this study provide evidence of signi¢-cant experience-related changes in the central nervoussystem following adult-onset, partial sensory depriva-tion due to profound unilateral deafness. Based onRMS amplitude, the AEPs of the unilaterally deaf in-dicate a clear increase in activation of the hemisphereipsilateral to the stimulated ear. The di¡erences incross-correlation coe¤cients suggest a decrease in in-ter-hemispheric synchrony over frontal cortex but in-creased synchronization over central cortical areas.The data also suggest that inter-hemispheric peak-to-peak and peak amplitude correlation changes are expe-rience-dependent. Across the unilaterally deaf subjects,the strength of the inter-hemispheric amplitude correla-tions, particularly for the N1 peak, increases for at least2 years after the onset of profound unilateral deafness.
4.1. Comparison to previous studies
When tested under identical monaural listening con-ditions, mean AEP RMS amplitude recorded over thehemisphere ipsilateral to the stimulated ear was signi¢-cantly larger in unilaterally deaf adults than in normal-hearing adults. Signi¢cant between-group di¡erenceswere not found for RMS amplitude measured overthe contralateral hemisphere. These results are consis-tent with those from animal experiments showing am-plitude increases in auditory pathway activity ipsilateralto the intact ear after unilateral destruction of one co-chlea (Popelar et al., 1994; Reale et al., 1987; Kitzes,1984, 1996). The increase in ipsilateral activation in theabsence of any change over the contralateral hemi-sphere signi¢cantly reduces the normally observed in-ter-hemispheric amplitude asymmetry produced bymonaural stimulation. Consequently, compared withthe normal-hearing group, the inter-hemispheric RMSI/C ratios of both unilaterally deaf groups were closerto 1.0, indicating more symmetrical activation. TheRMS I/C ratio changes were reinforced by similarchanges in the slopes of the best-¢t linear regressionfunctions derived from the inter-hemispheric correla-tions for AEP peak amplitude. These ¢ndings con¢rmprevious results of Vasama et al. (1994), Fujuki et al.(1998), and Sche¥er et al. (1998) showing more sym-metrical activation of the hemispheres by monauralsound presentation in the unilaterally deaf, comparedto normal-hearing subjects.
4.2. Origin of the increased inter-hemisphericcorrelations
Inter-hemispheric AEP waveform cross-correlationcoe¤cients and peak amplitude correlations were sig-ni¢cantly stronger in the unilaterally deaf than in thenormal-hearing subjects. It is important to note that allinter-hemispheric correlations examined for the normal-hearing and unilaterally deaf groups were signi¢cant atlevels well beyond chance. Depending on the distancefrom the scalp, orientation, and spatial extent, a singleneural generator may produce measurable evoked ac-tivity over large areas of the head, both contralateraland ipsilateral to the underlying source (Scherg, 1990).Thus, some degree of inter-hemispheric correlation isexpected simply due to volume conduction and the spa-tial smearing e¡ects of the meninges, cerebrospinal £u-id, scalp, and skull on scalp-recorded AEP activity. Themajority of unilaterally deaf subjects tested in this studyunderwent acoustic neuroma surgery, requiring a cra-niotomy on the side contralateral to the intact ear.There is some post-operative ¢ll-in of the cranial open-ing, but the reduced skull thickness at the site of thecraniotomy could alter the ¢eld of the scalp-recordedAEPs, particularly at electrode locations proximal tothe site of surgery, contralateral to the intact ear. How-ever, the di¡erences found in absolute RMS amplitudewere only observed ipsilateral, not contralateral, to theintact ear. Thus, it is unlikely that di¡erences in volumeconduction and spatial smearing e¡ects can account forthe observed AEP di¡erences between the unilaterallydeaf and the normal-hearing groups.
Other factors that might contribute to the obtainedpattern of results were also considered. For example, anincrease in AEP amplitude ipsilateral to the stimulatedear could account for the reduced inter-hemispheric am-plitude asymmetry. However, this cannot account forthe increased inter-hemispheric cross-correlation valuesbecause these measures are amplitude-independent. It isalso possible that age di¡erences between subjectgroups might be an important factor. In the rat, Casp-ary et al. (1995) demonstrated signi¢cant age-relateddeclines of the inhibitory neurotransmitter Q-aminobu-tyric acid (GABA). In humans, a similar age-relateddecline in GABA-based inhibitory processes might re-duce inter-hemispheric amplitude asymmetries and in-crease inter-hemispheric cross-correlations. However, itwas the 9 2 years deaf group (mean age = 49.2 years)who were older than the s 2 years deaf group (meanage = 37.4 years) group and the normal-hearing group(mean age = 32.8 years). Thus, age may be an importantfactor, but not in the manner suggested above. Rather,an alternative e¡ect of age must be considered: whenprofound unilateral deafness occurs later in life, thechanges in cortical activity may be less extensive. It
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^44 41

must also be noted that within the existing data set,there were no within-group trends for smaller di¡eren-ces with later age of deafness onset.
Compared to normal-hearing subjects, inter-hemi-spheric AEP cross-correlation coe¤cients were weakerover frontal cortex, stronger over central cortex, andshowed no signi¢cant di¡erences over temporal or pa-rietal-occipital regions. Over the central regions, signi¢-cantly stronger inter-hemispheric peak-to-peak ampli-tude correlations (both P1^N1 and the N1^P2) werefound in the unilaterally deaf. For the unilaterallydeaf subjects, signi¢cantly elevated inter-hemisphericpeak amplitude correlations were found for the P1
peak (s 2 years deaf), but were most robust for theN1 peak (9 2 years and s 2 years deaf). This may in-dicate that the di¡erences between the unilaterally deafand normal-hearing adults primarily re£ect changes inthe neural processes generating the N1 peak. Whiledeafness-related alterations in the neural generatorsunderlying P1 cannot be excluded, a portion of thechanges in P1 amplitude may represent the e¡ects ofvolume-conducted, partly temporally overlapped P1
and N1 activity as measured from the scalp. As previ-ously described, the P2 peak most likely re£ects activa-tion of the mesencephalic reticular activating systemand its thalamo-cortical projections that may be drivenby input from all sensory modalities. This system ap-pears to terminate on thalamic or cortical structures ator near to the midline of the brain rather than auditorycortical areas in the temporal lobe (Skinner andYingling, 1977; Yingling and Skinner, 1977; Knightet al., 1980, 1988; Rif et al., 1991; Woods et al.,1993). On the other hand, P1 and N1 are presumed topredominately re£ect activation of neural generatorsspeci¢c to the auditory thalamic-cortical pathways(Na«a«ta«nen and Picton, 1987; Lie«geois-Chauvel et al.,1994; Ma«kela« and McEvoy, 1996). The distinct en-hancement of the inter-hemispheric correlation for N1
amplitude in the unilaterally deaf is of particular inter-est since a major source of N1 activity may be re-en-trant or reciprocal auditory cortical activity as a resultof inter-hemispheric activation (Ma«kela« and Hari,1992; Ma«kela« and McEvoy, 1996).
4.3. Time course and possible mechanisms of theincreased inter-hemispheric correlations
Unfortunately presurgical AEPs were not obtainedfrom any of the acoustic neuroma patients. In addition,AEP data were not collected from any of the hearing-impaired subjects immediately after the onset of pro-found unilateral deafness. Therefore, the possibilitythat some changes in central auditory system activityoccur immediately after the onset of profound unilat-eral deafness cannot be excluded. Nevertheless, clear
evidence was obtained indicating that major changesin central auditory system activation occur across aprolonged time course of 2 years or more. Signi¢cantdi¡erences in central auditory system activity werefound between the normal-hearing adults and those in-dividuals unilaterally deaf 9 2 years at the time of test-ing, but were much more robust for those individualswho were unilaterally deaf s 2 years. This pattern ofchange suggests that as the time since the onset of uni-lateral deafness increases, the magnitude of the changesin cortical activity also increases. However, as indicatedpreviously, the subjects unilaterally deaf for 6 2 yearswere signi¢cantly older than those unilaterally deaf s 2years. Therefore, the possibility remains that the smallermagnitude e¡ects noted for the older (on average) 9 2years deaf group may actually re£ect less extensive cor-tical plasticity with increased age. This issue needs to beaddressed by a longitudinal investigation of changes incortical activity that spans at least 2^3 years post-onsetof deafness.
One mechanism that could account for more syn-chronous and symmetrical hemispheric activation isthe emergence of new projections in the auditory path-ways ipsilateral to the stimulated ear. This process hasbeen observed in other mammals, but only in thoseunilaterally deafened at a very young age (Kitzes,1996). An enhancement of projections in the auditorybrainstem ipsilateral to the intact ear could increase thecommon and synchronous drive of the ipsilateral andcontralateral pathways from the brainstem through tocortex, and thus account for the enhanced inter-hemi-spheric correlations. In addition, changes in excitatory/inhibitory (EI) processes at the cortical level must beconsidered. For instance, it is not clear how cortical EIcells that should be at least temporarily silenced by theonset of profound unilateral deafness should functionin the long term. Further investigations need to exam-ine the inter-hemispheric correlations of shorter latencyAEP peaks that may represent earlier stages of auditorycortex activation, such as Pa of the middle latency re-sponse.
4.4. Behavioral implications
The results demonstrated signi¢cant di¡erences in theinter-hemispheric correlations of auditory cortical activ-ity between groups of unilaterally deaf and normal-hearing subjects. At this point, one can only speculateabout the implications these changes might have forauditory behavioral abilities. Under natural listeningconditions, the unilaterally deaf perform much worsethan normal-hearing subjects on tests of speech recog-nition in the presence of competing background noise(e.g., Bess et al., 1986; Colletti et al., 1988). Whencompared to normal-hearing subjects who are tested
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^4442

binaurally, the unilaterally deaf also perform muchworse on tests of sound localization (e.g., Jongkeesand van der Veer, 1957; Slattery and Middlebrooks,1994). However, when both groups were tested undermonaural listening conditions, some of the congenitallyunilaterally deaf adults studied by Slattery and Middle-brooks (1994) localized sound better than the normal-hearing subjects. This suggests that for at least somecongenitally unilaterally deaf adults, compensatory pro-cesses exist that dynamically respond to a change insensory input. Whether such compensatory processesalso operate in adults with late-onset unilateral deafnessis not known. Further studies are planned to determinewhether the experience-related neurophysiologicalchanges associated with late-onset profound unilateraldeafness correlate with possible changes in behavioralabilities such as sound localization.
5. Conclusions
The ¢ndings from this and other studies of unilateraldeafness demonstrate that when sensory pathways inthe human brain are deprived of their normal sourceof activation, the capacity exists for experience-basedchange to respond to other sources of activity. Theunilaterally deaf subjects tested in this study all hadlate-onset profound hearing losses. The results thus in-dicate that the capacity for experience-related changesin human central auditory system persists in the adultbrain.
Acknowledgements
J.J. Eggermont made valuable comments on themanuscript. J.-P.V. received support from the FinnishCultural Foundation. We would also like to acknowl-edge the helpful comments from the reviewers.
References
Adams, J.C., 1979. Ascending projections to the inferior colliculus.J. Comp. Neurol. 183, 519^538.
Bess, F.H., Tharpe, A.M., Gibler, A.M., 1986. Auditory performanceof children with unilateral sensorineural hearing loss. Ear Hear. 7,20^26.
Brunso-Bechtold, J.K., Thompson, G.C., Masterson, R.B., 1981.HRP study of the organization of auditory a¡erents ascendingto the central nucleus of the inferior colliculus in cat. J. Comp.Neurol. 197, 705^722.
Caspary, D.M., Milbrandt, J.C., Helfert, R.H., 1995. Central auditoryaging: GABA changes in the inferior colliculus. Exp. Gerontol. 30,349^360.
Coleman, J.R., Clerici, W.J., 1987. Sources of projections to subdivi-
sions of the inferior colliculus in the rat. J. Comp. Neurol. 262,215^226.
Colletti, V., Fiorino, F.G., Carner, M., Rizzi, R., 1988. Investigationof the long-term e¡ects of unilateral hearing loss in adults. Br. J.Audiol. 22, 113^118.
Donoghue, J.P., 1995. Plasticity of adult sensorimotor representa-tions. Curr. Opin. Neurobiol. 5, 749^754.
Elberling, C., Bak, C., Kofoed, B., Lebech, J., Saermark, K., 1981.Auditory magnetic ¢elds from the human cortex. Scand. Audiol.10, 203^207.
Fujuki, N., Naito, Y., Nagamine, T., Shiomi, Y., Hirano, S., Honjo,I., Shibasaki, H., 1998. In£uence of unilateral deafness on evokedmagnetic ¢eld. NeuroReport 9, 3129^3133.
Greenough, W.T., 1975. Experiential modi¢cation of the developingbrain. Am. Sci. 63, 37^46.
Harrison, R.V., Stanton, S.G., Ibrahim, D., Nagasawa, A., Mount,R.J., 1993. Neonatal cochlear hearing loss results in developmentalabnormalities of the central auditory pathways. Acta Otolaryngol.113, 296^302.
Hendry, S.H., Jones, E.G., 1988. Activity-dependent regulation ofGABA expression in the visual cortex of adult monkeys. Neuron8, 701^712.
Jongkees, L.B.W., van der Veer, R.A., 1957. Directional hearing ca-pacity in hearing disorders. Acta Otolaryngol. 48, 465^474.
Kaltenbach, J.A., Czaja, J.M., Kaplan, C.R., 1992. Changes in thetonotopic map of the dorsal cochlear nucleus following inductionof cochlear lesions by exposure to intense sound. Hear. Res. 59,213^223.
Kitzes, L.M., 1984. Some physiological consequences of neonatal co-chlear destruction in the inferior colliculus of the gerbil, Merionesunguiculatus. Brain Res. 306, 171^178.
Kitzes, L.M., 1996. Anatomical and physiological changes in thebrainstem induced by neonatal ablation of the cochlea. In: Salvi,R., Henderson, D., Fiorino, F., Colletti, V. (Eds.), Auditory Sys-tem Plasticity and Regeneration. Thieme Medical, New York, pp.256^274.
Knight, R.T., Hillyard, S.A., Woods, D.L., Neville, H.J., 1980. Thee¡ects of frontal and temporal-parietal lesions on the auditoryevoked potentials. Electroencephalogr. Clin. Neurophysiol. 50,112^124.
Knight, R.T., Scabini, D., Woods, D.L., Clayworth, C., 1988. Thee¡ects of lesions of superior temporal gyrus and inferior parietallobe on temporal and vertex components of the human AEP.Electroencephalogr. Clin. Neurophysiol. 70, 499^509.
Lie«geois-Chauvel, C., Musolino, A., Badier, J.M., Marquis, P., Chau-vel, P., 1994. Evoked potentials recorded from the auditory cortexin man: evaluation and topography of the middle latency compo-nents. Electroencephalogr. Clin. Neurophysiol. 92, 204^214.
Ma«kela«, J.P., Hari, R., 1992. Neuromagnetic auditory evoked re-sponses after a stroke in the right temporal lobe. NeuroReport3, 94^96.
Ma«kela«, J.P., McEvoy, L., 1996. Auditory evoked ¢elds to illusorysound source movements. Exp. Brain Res. 110, 446^454.
Na«a«ta«nen, R., Picton, T.W., 1987. The N1 wave of the human electricand magnetic response to sound: a review and analysis of thecomponent structure. Psychophysiology 24, 375^425.
Pantev, C., Lu«tkenho«ner, B., Hoke, M., Lenhaertz, K., 1986. Com-parison between simultaneously recorded auditory-evoked mag-netic ¢elds and potentials elicited by ipsilateral, contralateral,and binaural sound burst stimulation. Audiology 25, 54^61.
Popelar, J., Erre, J.-P., Aran, J.-M., Cazals, Y., 1994. Plastic changesin the ipsi-contralateral di¡erences of auditory cortex and inferiorcolliculus evoked potentials after injury to one ear in the adultguinea pig. Hear. Res. 72, 125^134.
Rajan, R., Irvine, D.R., Wise, L.Z., Heil, P., 1993. E¡ect of unilateral
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^44 43

partial cochlear lesions in adult cats on the representation of le-sioned and unlesioned cochleas in primary auditory cortex.J. Comp. Neurol. 338, 17^49.
Reale, R.A., Brugge, J.F., Chan, J.C., 1987. Maps of auditory cortexin cats reared after unilateral cochlear ablation in the neonatalperiod. Brain Res. 431, 281^290.
Reite, M., Teale, P., Zimmerman, J., Davis, K., Whalen, J., 1988.Source location of a 50 ms latency auditory evoked ¢eld compo-nent. Electroencephalogr. Clin. Neurophysiol. 70, 490^498.
Rif, J., Hari, R., Hamalainen, M.S., Sams, M., 1991. Auditory atten-tion a¡ects two di¡erent areas in the human supratemporal cortex.Electroencephalogr. Clin. Neurophysiol. 79, 464^472.
Robertson, D., Irvine, D.R., 1989. Plasticity of frequency organiza-tion in auditory cortex of guinea pigs with partial unilateral deaf-ness. J. Comp. Neurol. 282, 456^471.
Sche¥er, K., Bilecen, D., Schmid, N., Tschopp, K., Seelig, J., 1998.Cereb. Cortex 8, 156^163.
Scherg, M., 1990. Fundamentals of dipole source potential analysis.In: Grandori, F., Romani, G.L., Hoke, M. (Eds.), AuditoryEvoked Potentials and Fields, Advances in Audiology, Vol. 6,Karger, Basel, pp. 40^69.
Schwaber, M.K., Garraghty, P.E., Kaas, J.H., 1993. Neuroplasticityof the adult primate auditory cortex following cochlear hearingloss. Am. J. Otol. 14, 252^258.
Skinner, J.E., Yingling, C.D., 1977. Central gating mechanisms thatregulate event-related potentials and behavior: a neural model forattention. In: Desmedt, J.E. (Ed.), Attention, Voluntary Contrac-tion and Event-Related Cerebral Potentials. Progress in ClinicalNeurophysiology, Karger, Basel, pp. 30^69.
Slattery, W.H., Middlebrooks, J.C., 1994. Monaural sound localiza-tion: acute versus chronic unilateral impairment. Hear. Res. 75,38^46.
Vasama, J.-P., Ma«kela«, J.P., 1995. Auditory pathway plasticity inadult humans after unilateral idiopathic sudden sensorineuralhearing loss. Hear. Res. 87, 132^140.
Vasama, J.-P., Ma«kela«, J.P., Parkkonen, L., Hari, R., 1994. Auditorycortical responses in humans with congenital unilateral conductivehearing loss. Hear. Res. 78, 91^97.
Vaughan, H.G., Ritter, W., 1970. The sources of auditory evokedresponses recorded from the human scalp. Electroencephalogr.Clin. Neurophysiol. 28, 360^367.
Willott, J.F., Kulig, J., Satter¢eld, T., 1984. The acoustic startle re-sponse in DBA/2 and C57BL/6 mice: relationship to auditoryneuronal response properties and hearing impairment. Hear.Res. 16, 161^167.
Willott, J.F., Parham, K., Hunter, K.P., 1991. Comparison of theauditory sensitivity of neurons in the cochlear nucleus and inferiorcolliculus of young and aging C57BL/6J and CBA/J mice. Hear.Res. 53, 78^94.
Woods, D.L., Knight, R.T., Scabini, D., 1993. Anatomical substratesof auditory selective attention: behavioral and electrophysiologicale¡ects of posterior association cortex lesions. Cognit. Brain Res. 1,227^240.
Yingling, C.D., Skinner, J.E., 1977. Gating of thalamic input to cer-ebral cortex by nucleus reticularis laminaris. In: Desmedt, J.E.(Ed.), Attention, Voluntary Contraction and Event-Related Cere-bral Potentials. Progress in Clinical Neurophysiology, Karger,Basel, pp. 70^96.
HEARES 3638 12-3-01
C.W. Ponton et al. / Hearing Research 154 (2001) 32^4444





























