Embed Size (px)
Citation preview

Platelet-activating factor (PAF)-like activity, localizationof PAF receptor (PAF-R) and PAF-acetylhydrolase
(PAF-AH) activity in bovine endometrium at differentstages of the estrous cycle and early pregnancy<
U. Tiemann*, W. Tomek, F. Schneider, K. Wollenhaupt, W. Kanitz,F. Becker, R. Po¨hland, H. Alm
Unit of Reproductive Biology of Research Institute for the Biology of Farm Animals,18196 Dummerstorf, Germany
Received 11 August 2000; received in revised form 27 December 2000; accepted 22 February 2001
Abstract
PAF-like activity in the endometrium increased from days 2–4 to day 12 and day 20 in both cyclicand pregnant cows. There was an increase in platelet aggregation induced by PAF-like activity in theendometrium of pregnant animals on day 20 as compared to cyclic animals at the same point in time.Two major bands of PAF-R protein at 67 kDa and 97 kDa were detected by Western blot analysis.PAF-R was localized mainly in luminal and glandular epithelium of the endometrium, but the stainingwas markedly increased in the endometrium of pregnant cows on day 20 compared to cyclic animalson the same day. The purified PAF-AH from the endometrium is similar to in plasma. In cyclic cattle,no changes in PAF-AH activity of endometrium were observed, whereas a decrease in enzyme activityoccurred in pregnant cows on day 20 as compared to cyclic animals on the same day. We suggest thatthe bovine endometrium produces PAF-like activity, expresses the PAF-R and possesses a PAF-AHactivity which varies during pregnancy. © 2001 Elsevier Science Inc. All rights reserved.
Keywords:PAF-like activity; PAF-receptor; PAF-acetylhydrolase-activity; endometrium; bovine
< This study was supported by Deutsche Forschungsgemeinschaft (DFG), Ti 189/4–1.* Corresponding author. Tel.:149-38208-68766; fax:149-38208-68752.E-mail address:[email protected] (U. Tiemann).
Prostaglandins & other Lipid Mediators 65 (2001) 125–141
0090-6980/01/$ – see front matter © 2001 Elsevier Science Inc. All rights reserved.PII: S0090-6980(01)00130-7

1. Introduction
Platelet-activating factor (PAF) was discovered from sensitized basophils that causedplatelet aggregation [1]. This factor is not only a mediator of inflammation and allergy, buthas numerous physiological functions, especially those involved in reproduction (for reviewsee: [2]).
The presence of PAF in the uterus was documented in the rabbit [3], rat [4,5], in culturedhuman endometrial cells [6,7], and in explanted ovine endometrial tissue [8]. The endome-trium of the tammar wallaby released PAF into culture medium in vitro varied with stage ofthe luteal phase [9]. PAF has been reported to play a role in maternal recognition ofpregnancy based on decreases in the inactive PGF2a metabolite (PGFM) in jugular venousplasma [10], and it promotes the synthesis of prostaglandin E2 in uterine endometrial cellsof humans [7,11], sheep [12], and bovine [13]. Therefore, it has been suggested that PAFmay play an important role in implantation. Antagonists of PAF inhibit implantation in themouse and rat [14,15].
The response by maternal tissue to PAF may be mediated by endometrial PAF receptor[PAF-R] binding sites. Endometrial PAF-R have been characterized in the rabbit uterus onday 6 of pregnancy [16]. Furthermore, the existence of a PAF-R and its functions have beenshown in a human endometrial cell line HEC-1A [17]. PAF-R immunoreactivity wasdetected in proliferative and secretory human endometrium [18] but the localization of thereceptor in bovine endometrium during preimplantation of pregnancy remains to be deter-mined.
The regulation of PAF degradation is catalyzed by the enzyme PAF-acetylhydrolase(PAF-AH), which converts PAF to the biologically inactive lyso-PAF [19]. Therefore, it hasbeen suggested that PAF-AH may regulate, in part, PAF concentrations in plasma and tissue.PAF-AH was found in the uterine lumen of the ewe [8], mouse [20], and uterus of rat [5,21].
Until now there have been no reports investigating the three parameters (PAF-like activity,PAF-R, PAF-AH) in view of their significance in the fertilization and implantation processes.We investigated whether they change in bovine endometrium at different stages of pregnancycompared to that of nonpregnant animals: on days 2–4 (when the embryo is in the oviduct),day 12 (when the embryo is in the uterus), and day 20 of pregnancy just prior to implantation.The present work may contribute to a better understanding of the potential role of PAF andits receptor and the regulatory role of PAF-AH in the endometrium during the earlypregnancy of cows.
2. Methods
2.1. Animals and treatment
German Holstein heifers, 20 and 22 months of age and weighing between 460 and 540 kg,were used. Estrus was detected by daily observation of the animals. Palpation of the corpusluteum was per rectum 10 to 13 days after estrous detection. At this time point, all heiferswere synchronized by application of 2 ml Cloprostenol forte® (Jenapharm, Jena, Germany).
126 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

After 65 h, injection of 2 ml GnRH agonist Gonavet® (Veyx, Schwarzenborn, Germany) wasadministrated to induce ovulation (estrus, day 0). One group of heifers was artificiallyinseminated 13 h after ovulation induction and the other group was not inseminated. Heifersof both groups were slaughtered by exsanguination at fixed times after induced estrus: days2–4, 12 and 20. The reproductive tracts were removed immediately after slaughtering andtransported on ice to the laboratory. The oviducts and the uterine horns from animalsconsidered to be pregnant were flushed with PBS (pH 7.4). Animals were considered to bepregnant when an embryo was found with at least two blastomeres on day 2.
Endometrial tissues derived from cyclic animals on days 2 to 4 (n 5 5), day 12 (n 5 9), day20 (n 5 5) and pregnant animals on days 2 to 4 (n 5 5), day 12 (n 5 5), day 20 (n 5 6) wereinvestigated. Heifers that were mated, but not pregnant at the time of slaughter were not used.
2.2. Lipid extraction and isolation of phospholipid by thin-layer chromatography
The uterus was opened longitudinally, the endometrium was isolated and samples werestored at280°C. Total lipids from endometrium samples were extracted by the method ofBligh and Dyer [22]. One-two g of tissue was used. Dry phospholipid extract (PL) wasresuspended in 100ml fresh chloroform and submitted to preparative thin layer chromatog-raphy (TLC) on Silica Gel G plates (203 20 cm, Merck, Darmstadt, Germany). OneTCL-plate was divided in five lanes (one for standards and four for two samples); thestandard lane and one lane of endometrial extract were stained with phosphospray (Sigma,Deisenhofen, Germany) to detect the migration of PAF (Sigma) or PAF-like phospholipidsisolated from endometrial tissues. In the unstained sample lane, the area between sphingo-myelin (SM) and lyso-PAF (Sigma), which corresponds to that of synthetic PAF, wasscraped, and PL was extracted by the method of Blight and Dyer [22]. We analyzed twoendometrial samples per plate and each endometrial sample was analyzed twice. Phospho-lipids were evaporated to dryness under N2 and stored at220°C until assayed. The amountof PAF recovered after extraction and purification was determined by radiotracer using200,000 dpm [3H]PAF (Amersham, Braunschweig, Germany) per lane. Mean recovery of[3H]PAF activity from the TLC plate was 64% in the region in which PAF was visualizedwith phosphospray.
2.3. Platelet aggregation assay
Platelets were obtained from clinically normal heifers and processed as described byLiggitt et al. [23]. Briefly, platelet-rich plasma (PRP) was obtained by centrifugation (3003g for 30 min at 22°C) of 10-mL aliquots of whole blood containing one mL 0.11 mol/litersodium citrate at 3003 g for 30 min at 22°C. The supernatant consisted of plasma containingplatelets. Platelet-poor plasma (PPP) was made by centrifugation of citrate-blood at 20003g for 15 min at 22°C. A final concentration of 2–43 107 platelets/mL was used foraggregation experiments. The aggregation response of bovine platelets was evaluated usinga dual-channel aggregometer (Model APACT produced by LAFIBRINTIMER, Ahrensburg,Germany). The aggregation assay varied depending on the individual bovine platelet prep-
127U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

aration. To minimize the variation in PL levels in TLC-fractions, PL isolated from endo-metrium of each cycle or pregnant stages was assayed with the same platelet preparation. Astandard curve for synthetic PAF concentration (final concentration, 1.3–41.6 nmol/liter)was carried out for each bovine platelet preparation. Lyso-PAF or SM were used at a finalconcentration of 83 nmol/liter.
A 200 ml aliquot of the platelet suspension and 100ml PBS (0.25% BSA) were added toaggregometer tubes and stirred for 1 min at 37°C prior to the addition of 5ml PAF-standarddissolved in 0.25% BSA-PBS or PL-containing endometrial extract dissolved in ethanol (5ml). The dry PL residues were recovered in 60% ethanol for the aggregation assay [24].Phospholipids in the purified endometrial samples caused platelet aggregation (increasedlight transmission) was determined to be PAF-like activity and standardized to 1g endo-metrial tissue. Specificity of the synthetic PAF or PAF-like activity for action on bovineplatelet aggregation was assessed by experiments conducted in the presence of the specificPAF receptor antagonist WEB 2086 (Boehringer, Ingelheim, Germany) [25]. The PAFreceptor antagonist WEB 2086 was dissolved in 0.25% BSA-PBS (60 or 120 nmol/liter, finalconcentration) and 5ml of each solution was added to the cuvette 1 min prior to the additionof synthetic PAF (10.4 nmol/liter, final concentration) or PAF-like containing endometrialtissue samples.
2.4. Western blotting
Equal amounts of endometrial protein (30mg) were subjected to 10% SDS polyacryl-amide gel electrophoresis (PAGE) (SDS-PAGE) under reducing conditions. Electro transferof proteins to nitrocellulose membranes (Schleicher and Schuell, Dassel, Germany) wascarried out for 1 h at 0.2 mA/cm2, followed by blocking in TBST (Tris-buffered saline [TBS:20 mmol/liter Tris, 500 mmol/liter NaCl, pH 7.5]1 1% Tween 20) supplemented with 5%horse serum over night at 4°C. Membranes were then incubated with rabbit polyclonalanti-PAF-R antibody (1:500; Alexis, Gru¨nberg, Germany) in TBST supplemented with 2.5%horse serum for 3 h at20°C, washed both two times in TBST supplemented with 0.5% horseserum and then incubated with donkey anti-rabbit antibody (1:10,000) conjugated withhorseradish peroxidase (Chemicon International, Hofheim, Germany) for 1.5 h at 20°C. Afterwashing with TBST, the blots were developed using a chemiluminescence system (ECL-kit;Amersham) according to the manufacturer’s protocol.
2.5. Immunohistochemical staining
Endometrial tissues were fixed in Bouin’s fixative at room temperature for 7 h; they werethen washed in 70% ethanol at room temperature overnight. After fixation, tissue wasdehydrated in graded ethanol solutions, cleared in xylene, and embedded in paraffin forsectioning; 5mm sections were cut and mounted on poly-L-lysine slides. Briefly, sectionswere deparaffinized and dehydrated. Peroxidase quenching was performed with 0.3% H2O2
in TBS for 10 min. Nonspecific protein binding sites were blocked with 20% normal horseserum in TBS for 25 min at room temperature in a humidified chamber. Sections wereincubated with rabbit polyclonal anti-PAF-R antibody at 1:200 dilution in TBS (2.5%
128 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

BSA10.05% Tween) for 60 min at room temperature. Subsequently, the sections wereincubated with anti-rabbit IgG biotinylated secondary antibody (Chemicon International) at1:1000 dilution in TBS (0.5% BSA10.05% Tween) for 60 min at room temperature.Localization of PAF-R was detected using streptavidin-immunoperoxidase (Chemicon In-ternational) at 1:1000 dilution for rabbit IgG at room temperature for 60 min. After a 10 minwash in TBS, localization of the primary antibody was visualized with diaminobenzidine for3–5 min, producing a brown-colored stain. The specificity of the staining reaction wasmonitored in control sections where the primary antibody had been omitted or was replacedby normal rabbit serum. Sections were mounted in glycerin jelly and examined using a Zeissmicroscope. Sections were stained with Harris hematoxylin solution for morphological investigation.
2.6. Preparation of endometrial PAF-acetylhydrolase
Endometrium (1g) was homogenized in six volumes of 0.1 mol/liter Tris-HCl (pH 7.2)buffer (buffer A) at 4°C with a PT-K/PCU-11 Polytron homogenizer. This homogenate wascentrifuged at 5003 g for 15 min and then at 105,0003 g for 60 min. The resultingsupernatant was applied to a Mono-Q anionic exchanger in a Smart-chromatography system(Pharmacia, Sweden). The column was washed extensively with buffer A until all nonbinding material has passed the system. Bound proteins were eluted by increasing the ionicstrength in one step up to 300 mmol/liter KCl in buffer A. This procedure separated nearlyhalf of the protein content (non binding material) and a 10-fold concentration of the sample(bound material). This concentration step is necessary to reduce the sample volume for thefollowing PAF-AH assay because the PAF-AH activity was too low in crude extracts.
Thereafter, the sample (300 mmol/liter fraction) was transferred to buffer A by gelfiltration using a fast desalting column. The final purification was performed by Mono-Qchromatography using a flat gradient up to 300 mmol/liter KCl. Purity of the fractions wasdetermined by SDS-PAGE.
2.7. Electrophoresis
SDS-PAGE was performed on 12.5% gels according to Laemmli [26]. The gels werestained with Coomassie blue R 250 and destained with methanol:acetic acid:water (4:1:4).
2.8. Protein estimation
Protein concentration was determined [27]. Approximately 100–200mg protein was usedin each PAF-AH assay.
2.9. Bioassay of PAF-acetylhydrolase activity
To confirm whether PAF-AH activity is present in the purified cytosolic fractions, 50mlof PAF solution (0.5 and 1.0mmol/liter final concentration) were incubated with 50ml PBS,PAF-AH negative or positive fractions at 37°C for 5 min. After incubation, the mixtures werecooled rapidly in an ice bath. Platelet aggregation activity was measured by adding 20ml of
129U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

the incubation mixture to 300ml bovine platelet suspension. Changes in light transmissionwere monitored using the aggregometer system described above.
2.10. PAF-acetylhydrolase assay
The activity of purified PAF-AH was assayed by using a commercially available assay kit(Alexis). The enzyme activity was expressed as nmol/min/mg protein. The detection rangeof the assay is from 10 to 200 nmol/min/mg protein, and the color development is linear forat least 30 min. The PAF-AH activity was calculated with the values measured after 10 min.For characterization of the two isoenzymes of PAF-AH, a plasma (extracellular) and acytosolic (intracellular) form, the purified cytosolic fractions of endometrium (n 5 3) andbovine purified plasma (n 5 2) were treated without or with sodium fluoride (NaF), trypsin,EGTA, p-bromophenacylbromide (p-BPB) (Sigma, final concentration: 0.04%, 40 mmol/liter, 20 mmol/liter, 0.15 mmol/liter, respectively) for 45 min at 37°C before assaying forPAF-AH activity.
2.11. Statistical analysis
The data are expressed as the mean6 SEM of duplicate measurements of 3 to 9 animalsper group. All data were first examined by ANOVA. Differences between means weredetermined by Student-Newman-Keuls test;P values,0.05 were considered significant. Therelative data were transformed with SAS statistical software for estimation ofEC50 forPAF-induced aggregation of platelets. Transforming model was the ‘Three Parametric logTransforming Model’.
3. Results
3.1. Platelet aggregation assay
The aggregation response of bovine platelets to exogenous synthetic PAF occurred overa PAF concentration range of 1.3–41.6 nmol/liter (final concentration), with a maximumaggregation occurring at 20.8–41.6 nmol/liter and with theED50 at 2.44 nmol/liter of PAF(n 5 7). Lyso-PAF, PC and SM did not cause any aggregation of bovine platelets. WEB 2086(final concentration, 120 nmol/liter) completely inhibited platelet aggregation induced by10.4 nmol/liter synthetic PAF.
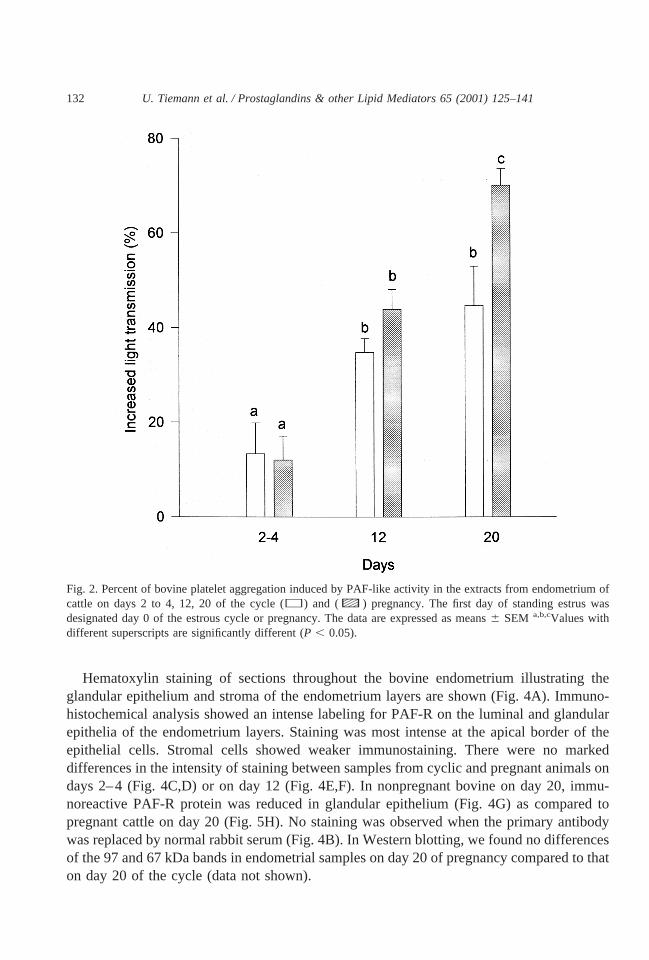
Platelet aggregation occurred in endometrial samples from cyclic and pregnant cattlefollowing PL extraction and purification by TLC. To assess the specificity of the PL extractsas PAF-like activity, experiments were conducted in the presence of WEB 2086. Fig. 1demonstrates that WEB 2086 inhibited platelet aggregation induced by purified PL fromendometrium. The antagonism of purified PL was equivalent to that of the synthetic PAF.Therefore, we can assume that we detected in endometrium after extraction and partialpurification by TLC a phospholipid that possesses PAF-like activity. Fig. 2 shows that theability for aggregation was lower in endometrial extracts from both cyclic (13.36 6.53%,
130 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141
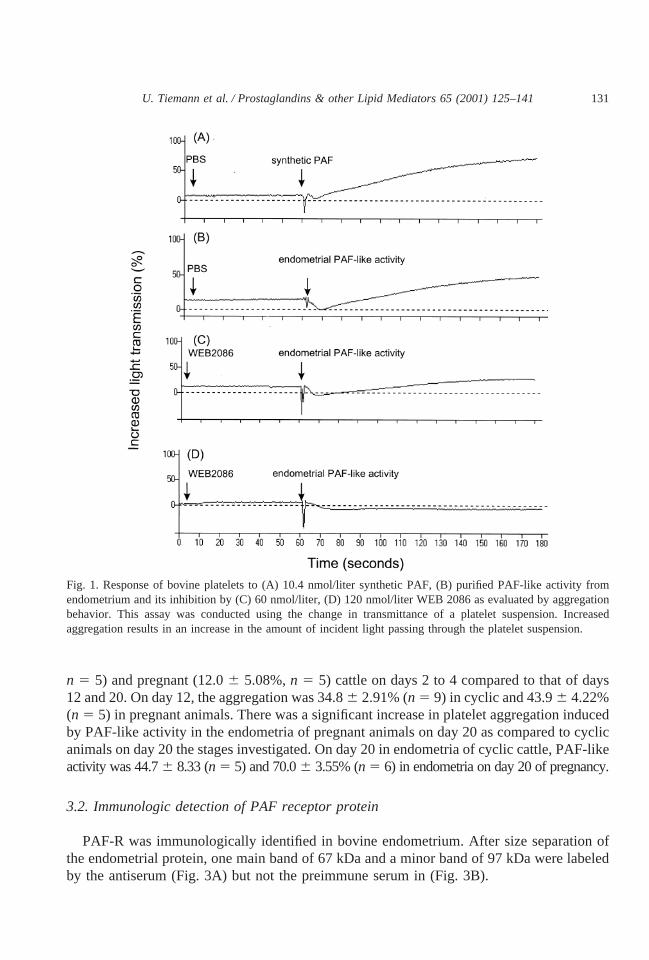
n 5 5) and pregnant (12.06 5.08%,n 5 5) cattle on days 2 to 4 compared to that of days12 and 20. On day 12, the aggregation was 34.86 2.91% (n 5 9) in cyclic and 43.96 4.22%(n 5 5) in pregnant animals. There was a significant increase in platelet aggregation inducedby PAF-like activity in the endometria of pregnant animals on day 20 as compared to cyclicanimals on day 20 the stages investigated. On day 20 in endometria of cyclic cattle, PAF-likeactivity was 44.76 8.33 (n 5 5) and 70.06 3.55% (n 5 6) in endometria on day 20 of pregnancy.
3.2. Immunologic detection of PAF receptor protein
PAF-R was immunologically identified in bovine endometrium. After size separation ofthe endometrial protein, one main band of 67 kDa and a minor band of 97 kDa were labeledby the antiserum (Fig. 3A) but not the preimmune serum in (Fig. 3B).
Fig. 1. Response of bovine platelets to (A) 10.4 nmol/liter synthetic PAF, (B) purified PAF-like activity fromendometrium and its inhibition by (C) 60 nmol/liter, (D) 120 nmol/liter WEB 2086 as evaluated by aggregationbehavior. This assay was conducted using the change in transmittance of a platelet suspension. Increasedaggregation results in an increase in the amount of incident light passing through the platelet suspension.
131U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141
Hematoxylin staining of sections throughout the bovine endometrium illustrating theglandular epithelium and stroma of the endometrium layers are shown (Fig. 4A). Immuno-histochemical analysis showed an intense labeling for PAF-R on the luminal and glandularepithelia of the endometrium layers. Staining was most intense at the apical border of theepithelial cells. Stromal cells showed weaker immunostaining. There were no markeddifferences in the intensity of staining between samples from cyclic and pregnant animals ondays 2–4 (Fig. 4C,D) or on day 12 (Fig. 4E,F). In nonpregnant bovine on day 20, immu-noreactive PAF-R protein was reduced in glandular epithelium (Fig. 4G) as compared topregnant cattle on day 20 (Fig. 5H). No staining was observed when the primary antibodywas replaced by normal rabbit serum (Fig. 4B). In Western blotting, we found no differencesof the 97 and 67 kDa bands in endometrial samples on day 20 of pregnancy compared to thaton day 20 of the cycle (data not shown).
Fig. 2. Percent of bovine platelet aggregation induced by PAF-like activity in the extracts from endometrium ofcattle on days 2 to 4, 12, 20 of the cycle (h) and (c) pregnancy. The first day of standing estrus wasdesignated day 0 of the estrous cycle or pregnancy. The data are expressed as means6 SEM a,b,cValues withdifferent superscripts are significantly different (P , 0.05).
132 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

3.3. Endometrial PAF-AH-activity
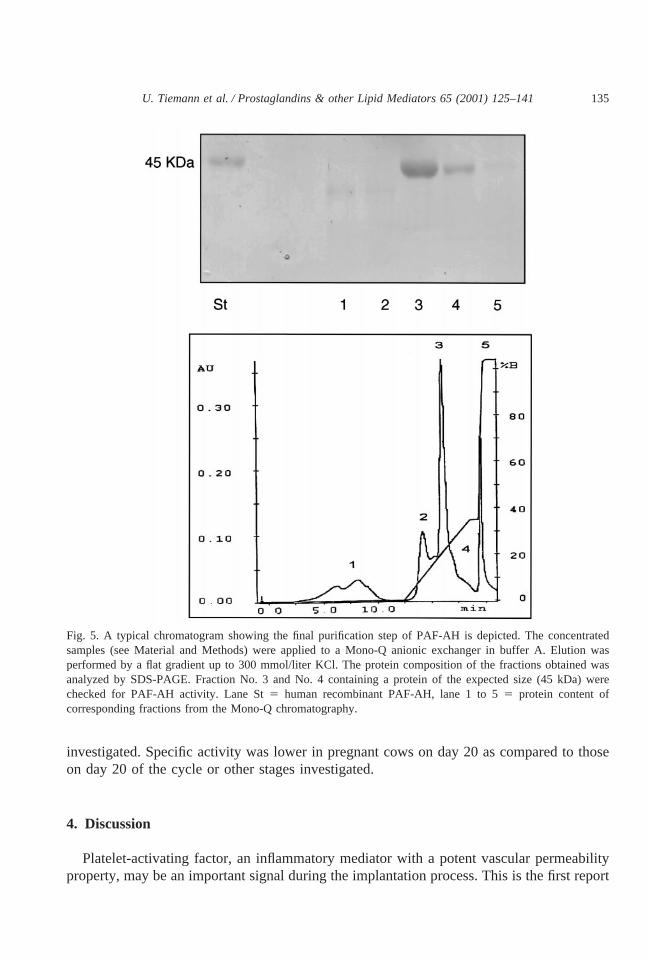
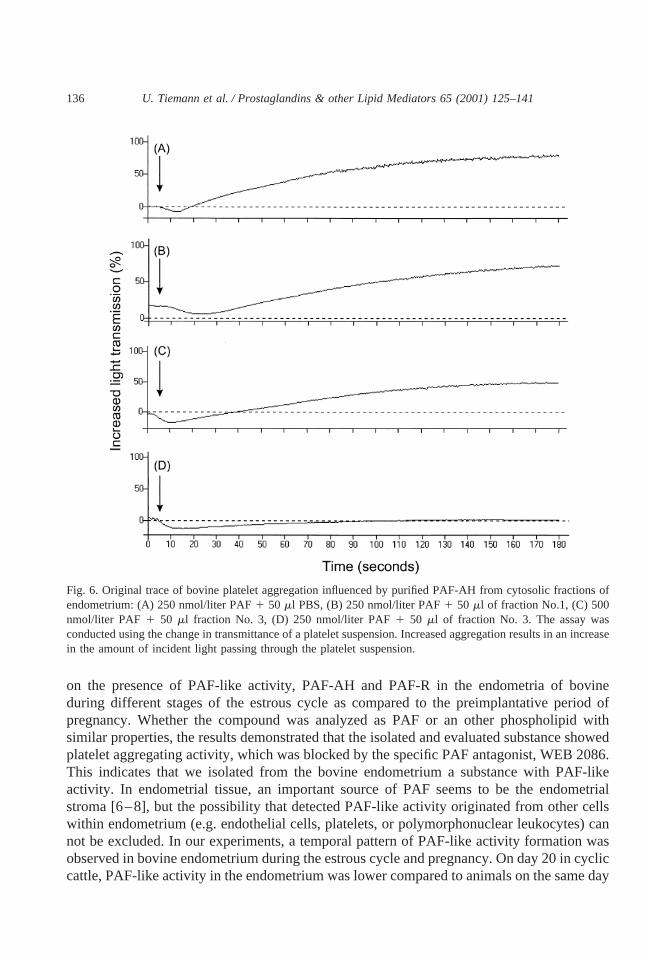
The separation of PAF-AH fractions by anionic exchange chromatography of endometrialsubcellular fraction protein demonstrates the pattern of protein elution (Fig. 5). From allisolated fractions, PAF-AH enzyme activity in endometrium was found mainly in fractionNo. 3 and was lower in fraction No. 4 and not detected in fraction Nos. 1, 2, 5. The activitycorresponded to a single major 45 kDa band on SDS-PAGE of fractions No. 3 and 4, whichis in the band of human recombinant PAF-AH. The characteristic 45 kDa band of fractionNo. 3 corresponded with biologic activity of PAF-AH measured by aggregometry. Afterincubation with 50ml of PAF-AH-positive fraction No. 3 together with 500 nmol/liter PAF,a decrease of aggregation to 41.46 1.20% (n 5 2) occurred, whereas after incubation with250 nmol PAF together with 50ml of fraction No. 3 no aggregation was detected. As acontrol, platelet aggregation could be detected after addition of 250 nmol/liter PAF solutionwith 50 ml PBS, it was 70.96 2.60% (n 5 3). Platelet aggregation was not changed byPAF-AH-negative fraction Nos. 1, 2, 5 (68.46 2.47%,n 5 9). The original trace of thisPAF-AH-caused inhibition of PAF-induced platelet aggregation is shown (Fig. 6). ThePAF-AH standard (human recombinant PAF-AH) did not cause aggregation (data notshown).
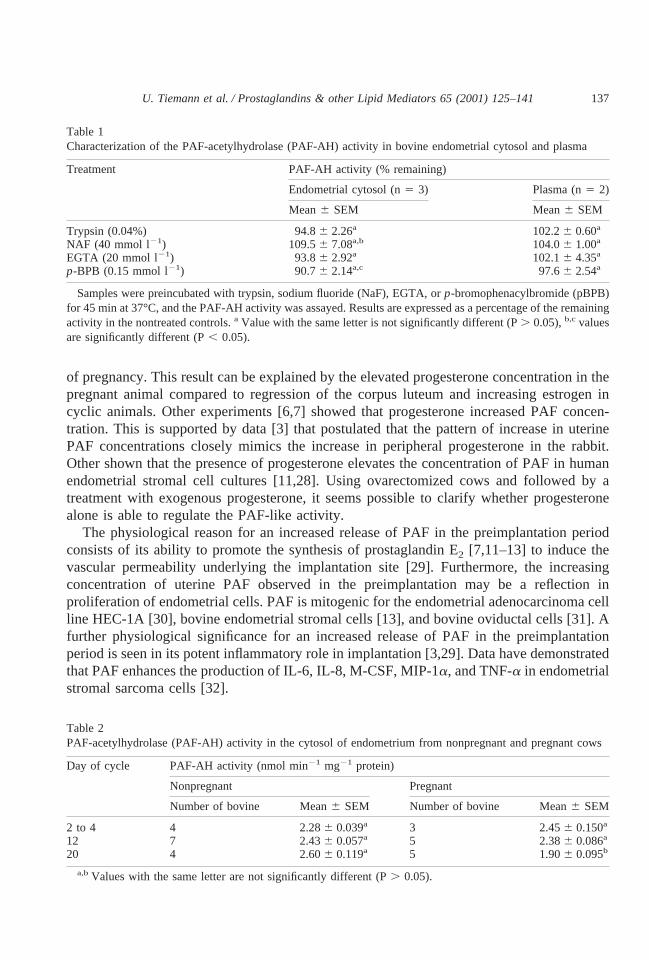
As shown in Table 1, PAF-AH purified and isolated from bovine endometrial homoge-nates and plasma by anionic exchange chromatography were resistant to trypsin, NaF,p-BPB, and EGTA. There are no differences between the control and different treatedgroups. This suggests that PAF-AH in the cytosol of the endometrium is apparently identicalto that present in plasma. The SDS-PAGE identified PAF-AH of plasma as a protein with themain band in the range of 45 kDa on SDS-PAGE (data not shown).
The specific activity of endometrial PAF-AH is given in Table 2. In the endometria ofcyclic cattle, changes in enzyme activity were not observed during the estrous cycle
Fig. 3. Detection of PAF-R (10% SDS-PAGE) in bovine endometrial cytosol using Western blotting. 30mg oftotal protein from lysate of endometrial tissue were incubated with (A) the anti-hPAF-R antiserum at a dilutionof 1:500, (B) the preimmune serum at a dilution of 1:500.
133U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141
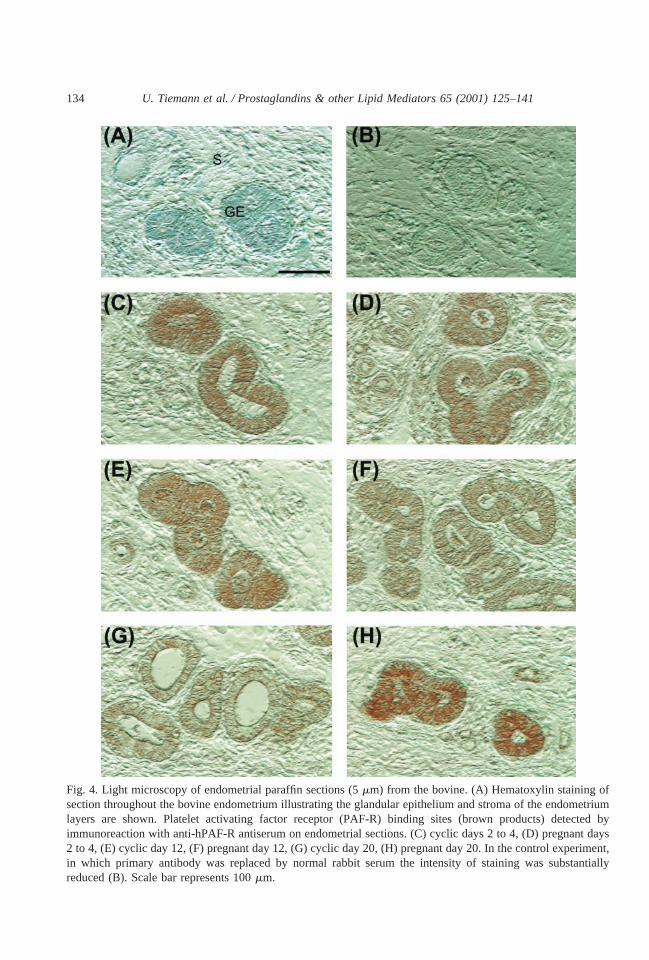
Fig. 4. Light microscopy of endometrial paraffin sections (5mm) from the bovine. (A) Hematoxylin staining ofsection throughout the bovine endometrium illustrating the glandular epithelium and stroma of the endometriumlayers are shown. Platelet activating factor receptor (PAF-R) binding sites (brown products) detected byimmunoreaction with anti-hPAF-R antiserum on endometrial sections. (C) cyclic days 2 to 4, (D) pregnant days2 to 4, (E) cyclic day 12, (F) pregnant day 12, (G) cyclic day 20, (H) pregnant day 20. In the control experiment,in which primary antibody was replaced by normal rabbit serum the intensity of staining was substantiallyreduced (B). Scale bar represents 100mm.
134 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141
investigated. Specific activity was lower in pregnant cows on day 20 as compared to thoseon day 20 of the cycle or other stages investigated.
4. Discussion
Platelet-activating factor, an inflammatory mediator with a potent vascular permeabilityproperty, may be an important signal during the implantation process. This is the first report
Fig. 5. A typical chromatogram showing the final purification step of PAF-AH is depicted. The concentratedsamples (see Material and Methods) were applied to a Mono-Q anionic exchanger in buffer A. Elution wasperformed by a flat gradient up to 300 mmol/liter KCl. The protein composition of the fractions obtained wasanalyzed by SDS-PAGE. Fraction No. 3 and No. 4 containing a protein of the expected size (45 kDa) werechecked for PAF-AH activity. Lane St5 human recombinant PAF-AH, lane 1 to 55 protein content ofcorresponding fractions from the Mono-Q chromatography.
135U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141
on the presence of PAF-like activity, PAF-AH and PAF-R in the endometria of bovineduring different stages of the estrous cycle as compared to the preimplantative period ofpregnancy. Whether the compound was analyzed as PAF or an other phospholipid withsimilar properties, the results demonstrated that the isolated and evaluated substance showedplatelet aggregating activity, which was blocked by the specific PAF antagonist, WEB 2086.This indicates that we isolated from the bovine endometrium a substance with PAF-likeactivity. In endometrial tissue, an important source of PAF seems to be the endometrialstroma [6–8], but the possibility that detected PAF-like activity originated from other cellswithin endometrium (e.g. endothelial cells, platelets, or polymorphonuclear leukocytes) cannot be excluded. In our experiments, a temporal pattern of PAF-like activity formation wasobserved in bovine endometrium during the estrous cycle and pregnancy. On day 20 in cycliccattle, PAF-like activity in the endometrium was lower compared to animals on the same day
Fig. 6. Original trace of bovine platelet aggregation influenced by purified PAF-AH from cytosolic fractions ofendometrium: (A) 250 nmol/liter PAF1 50 ml PBS, (B) 250 nmol/liter PAF1 50 ml of fraction No.1, (C) 500nmol/liter PAF 1 50 ml fraction No. 3, (D) 250 nmol/liter PAF1 50 ml of fraction No. 3. The assay wasconducted using the change in transmittance of a platelet suspension. Increased aggregation results in an increasein the amount of incident light passing through the platelet suspension.
136 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141
of pregnancy. This result can be explained by the elevated progesterone concentration in thepregnant animal compared to regression of the corpus luteum and increasing estrogen incyclic animals. Other experiments [6,7] showed that progesterone increased PAF concen-tration. This is supported by data [3] that postulated that the pattern of increase in uterinePAF concentrations closely mimics the increase in peripheral progesterone in the rabbit.Other shown that the presence of progesterone elevates the concentration of PAF in humanendometrial stromal cell cultures [11,28]. Using ovarectomized cows and followed by atreatment with exogenous progesterone, it seems possible to clarify whether progesteronealone is able to regulate the PAF-like activity.
The physiological reason for an increased release of PAF in the preimplantation periodconsists of its ability to promote the synthesis of prostaglandin E2 [7,11–13] to induce thevascular permeability underlying the implantation site [29]. Furthermore, the increasingconcentration of uterine PAF observed in the preimplantation may be a reflection inproliferation of endometrial cells. PAF is mitogenic for the endometrial adenocarcinoma cellline HEC-1A [30], bovine endometrial stromal cells [13], and bovine oviductal cells [31]. Afurther physiological significance for an increased release of PAF in the preimplantationperiod is seen in its potent inflammatory role in implantation [3,29]. Data have demonstratedthat PAF enhances the production of IL-6, IL-8, M-CSF, MIP-1a, and TNF-a in endometrialstromal sarcoma cells [32].
Table 1Characterization of the PAF-acetylhydrolase (PAF-AH) activity in bovine endometrial cytosol and plasma
Treatment PAF-AH activity (% remaining)
Endometrial cytosol (n5 3) Plasma (n5 2)
Mean6 SEM Mean6 SEM
Trypsin (0.04%) 94.86 2.26a 102.26 0.60a
NAF (40 mmol l21) 109.56 7.08a,b 104.06 1.00a
EGTA (20 mmol l21) 93.86 2.92a 102.16 4.35a
p-BPB (0.15 mmol l21) 90.76 2.14a,c 97.66 2.54a
Samples were preincubated with trypsin, sodium fluoride (NaF), EGTA, orp-bromophenacylbromide (pBPB)for 45 min at 37°C, and the PAF-AH activity was assayed. Results are expressed as a percentage of the remainingactivity in the nontreated controls.a Value with the same letter is not significantly different (P. 0.05),b,c valuesare significantly different (P, 0.05).
Table 2PAF-acetylhydrolase (PAF-AH) activity in the cytosol of endometrium from nonpregnant and pregnant cows
Day of cycle PAF-AH activity (nmol min21 mg21 protein)
Nonpregnant Pregnant
Number of bovine Mean6 SEM Number of bovine Mean6 SEM
2 to 4 4 2.286 0.039a 3 2.456 0.150a
12 7 2.436 0.057a 5 2.386 0.086a
20 4 2.606 0.119a 5 1.906 0.095b
a,bValues with the same letter are not significantly different (P. 0.05).
137U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

The action of PAF is regulated via its receptors. In the present study, Western blots fromlysates of endometrium tissue showed a main band of PAF-R with a size of 67 kDa. Thisfinding supports previous results obtained from human platelets reported by [33] and inlysates from human leukocytes by [34]. We found an additional band at 97 kDa. Thediscrepancy between these results may be due to the predicted glycosylation [35]. Multipleforms of PAF-R have also been reported previously [36,37]. However, Ahmed et al. [18]were not able to detect PAF-R protein in human endometrial tissue by Western blotting. Theauthors assumed that the receptor could be too hydrophobic to migrate adequately on gels.
We found by immunohistochemistry a specific staining of PAF-R that is mainly distrib-uted in glandular and luminal epithelia of the endometrium, but in the stromal cells consistentbinding was less. This result is in agreement with the data reported for human glandularepithelium [38] and in glandular epithelial cells of pregnant rabbit endometrium [16]. In thepresent study, there was an increase in PAF-R immunohistochemical staining on day 20 ofpregnancy compared to day 20 of the estrous cycle. This result was confirmed by immuno-cytochemical staining measured by flow cytometry in uterine stromal cells [39]. Thediscrepancy between immunohistochemistry and Western blotting regarding day effectscould be due to the method used for Western blotting. An equal amount of protein per lanewas loaded on gels, it is possible that different amount of tissue equivalents were loaded ineach lane. Therefore, we can assume that comparisons between Western blotting and theother methods would be inappropriate.
The expression of PAF-R in the endometrium during the preimplantation period is likelyto be regulated, at least in part, by progesterone and furthermore the up-regulation of PAF-Rexpression following exposure to PAF, was reported in human platelets [40], human B-cells[41], Kupffer cells [42] or human monocytes [43]. At the present time, the biologic relevanceof the PAF-mediated up-regulation of its own receptor remains open for discussion.
Biologically active PAF is converted to the biologically inactive lyso-PAF by PAF-AH.The present work demonstrates a simple and reproducible procedure for the isolation ofbovine PAF-AH from the cytosolic fraction of endometrium. Our purified PAF-AH from theendometrium showed a single 45 kDa protein band in the SDS-PAGE which agrees with thesingle band of human recombinant PAF-AH used as standard. PAF-AH was purified firstfrom human plasma [44]. It showed a single 43 kDa protein band [45]. We suggest that thePAF-AH purified from the bovine endometrium is apparently identical to the plasma form ofthe enzyme, since treatment with trypsin, NaF, EGTA, orp-BPB its PAF-activity was in thesame range as that occurring in the bovine plasma. In contrast to our findings, significantquantities of the cytosolic form of PAF-AH in rat uterus have been reported [5]. The reasonfor the different results may be due to species differences (bovine, rat) or some conditionsunder which the experiments were conducted. After our isolation method by means ofMono-Q anionic exchanger, we obtained a protein band at 45 kDa in the fractions of bothendometrial cytosol and plasma.
We reported on the PAF-AH activity in the bovine endometrium, which has already beendescribed in the uterus of the ewe [8], mouse [20], and rat [5,21]. In the present study, nochanges in PAF-AH activity were observed in endometrial fractions of nonpregnant bovineat different stages of the cycle. This agrees with results by Ohisge et al. [46] who did not findvariations in plasma PAF-AH activity in rats over a 14 day period. We observed a decrease
138 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

in endometrial PAF-AH activity in pregnant cows on day 20 compared to cyclic animals onthis day, which confirmed previous observation [20] that the mouse uterine PAF-AH activitywas lower in the preimplantation period compared to its preovulatory level. It does appearthat pregnancy suppresses this enzyme when compared to earlier days. The mechanism bywhich steroids are involved in regulation of PAF-AH in the uterus of cows remains to beelucidate.
The increase in PAF concentrations in endometrium during the estrous cycle and preg-nancy was higher than the observed changes of PAF-AH activity. A similar phenomenon wasfound [5]. PAF-AH contributes to regulation of the PAF level in the endometrium at the timeof implantation. One would predict that the PAF concentration would be elevated in thepregnant uterus, which was in fact the case. It has been suggested that PAF-AH may regulate,in part, PAF concentrations in tissues. The effect of PAF or PAF-like substance on theimplantation process seems to be restricted to a short, critical period in embryo development.The precise mechanisms of PAF involvement in implantation, as well as the primary cellularorigin and targets of its action, remain to be determined. In summary, on the basis of ourresults it is suggested that PAF, its receptor and the PAF-AH are involved in the physio-logical process of implantation in the bovine.
References
[1] Benveniste J, Henson PM, Cochrane CG. Leukocyte-dependent histamine release from rabbit platelets. Therole of IgE, basophils and platelet-activating factor. J Exp Med 1972;136:1356–77.
[2] Yasuda K, Johnston JM. The hormonal regulation of platelet-activating factor acetylhydrolase in the rat.Endocrinology 1992;130:708–16.
[3] Angle MJ, Jones MA, McManus LM, Pinckard RN, Harper MJK. Platelet-activating factor in the rabbituterus during early pregnancy. J Reprod Fertil 1988;83:711–22.
[4] Yasuda K, Satouchi K, Saito K. Platelet-activating factor in normal rat uterus. Biochem Bioph Res Co1986;138:1231–6.
[5] Li L, Yasuda K, Matsubara T, Okada H, Nakajima T, Sanezumi M, Kanzaki H. Estrogen effects onplatelet-activating factor and platelet-activating factor acetylhydrolase activity in rat uterus during the latestages of pregnancy. Prost Lipid Med 1999;57:219–30.
[6] Alecozay AA, Casslen BG, Riehl RM. Platelet-activating factor in human luteal phase endometrium. BiolReprod 1989;41:578–86.
[7] Alecozay AA, Harper MJK, Schenken RS, Hanahan DJ. Paracrine interactions between platelet-activatingfactor and prostaglandins in hormonally-treated human luteal phase endometrium in vitro. J Reprod Fertil1991;91:301–12.
[8] Battye KM, Evans G, O’Neill C. Ovine endometrium synthesizes and releases platelet-activating factorwhich can cause the release of prostaglandin F2a by the uterus in situ. Biol Reprod 1996;54:355–63.
[9] Koijma T, Hinds LA, Muller WJ, O’Neill C, Tyndale-Biscoe CH. Production and secretion of progesteronein vitro and presence of platelet-activating factor (PAF) in early pregnancy of the marsupial, Marcopuseugenii. Reprod Fertil Dev 1993;5:15–26.
[10] O’Neill C. Embryo-derived platelet-activating factor: a preimplantation embryo mediator of maternalrecognition of pregnancy. Domest Anim Endocrin 1987;4:69–86.
[11] Smith SK, Kelly RW. Effect of platelet-activating factor on the release of PGF2a and PGE2 by separatedcells of human endometrium. J Reprod Fertil 1988;82:271–6.
[12] Salamonsen LA. Effect of platelet-activating factor on prostaglandin release from ovine endometrial cellsin culture. Prostagland Leukotr Ess Fatty Acids 1991;44:67–70.
139U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

[13] Tiemann U, Davidson JA, Hansen PJ. Regulation of prostaglandin secretion and proliferation of bovineendometrial stromal cells by platelet-activating factor. Reprod Dom Anim 1995;30:141–3.
[14] Acker G, Hecquet F, Etienne A, Braquet P, Mencia-Huerta JM. Role of platelet-activating factor (PAF) inthe ovoimplantation in the rat: effect of the specific PAF-acether antagonist, BN 52021. Prostaglandins1988;35:233–41.
[15] Spinks NR, Ryan JP, O’Neill C. Antagonists of embryo-derived platelet-activating factor act by inhibitingthe ability of mouse embryo to implant. J Reprod Fertil 1990;88:241–8.
[16] Kudolo GB, Kasamo M, Harper MJK. Autoradiographic localization of platelet-activating factor (PAF)binding sites in the rabbit endometrium during the peri-implantation period. Tissue Res 1991;265:231– 41.
[17] Baldi E, Bonaccorsi L, Finetti G, Luconi M, Muratori M, Susini T, Forti G, Serio M, Maggi M.Platelet-activating factor in human endometrium. J Steroid Biochem Mol Biol 1994;49:359–63.
[18] Ahmed A, Dearn S, Shams M, Li XF, Sangha RK, Rola-Pleszczynski M, Jiang J. Localization, quantifi-cation and activation of platelet-activating factor receptor in human endometrium during the menstrualcycle: PAF stimulates NO, VEGF, and FAK. FASEB J 1998;12:831–43.
[19] Blank ML, Lee T-C, Fitzgerald V, Snyder FA. A specific acetylhydrolase for 1-alkyl-2-acetyl-sn- glycero3-phosphocholine (a hypotensive and platelet-activating lipid). J Biol Chem 1981;256:175–8.
[20] O’Neill C. Activity of platelet-activating factor acetylhydrolase in the mouse uterus during the estrous cycle,throughout the preimplantation phase of pregnancy, and throughout the luteal phase of pseudopregnancy.Biol Reprod 1995;52:965–71.
[21] Matsubara T, Yasuda K, Johnston JM, Sanezumi M, Okada H, Matsuoka S, Kanzaki H. Platelet-ActivatingFactor (PAF) and PAF acetylhydrolase activity in rat uterus and placenta during the late stages of pregnancy.Biol Reprod 1997;56:885–90.
[22] Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purification. Can J Biochem Phys1959;37:911–7.
[23] Liggitt HD, Leid RW, Huston L. Aggregation of bovine platelets by acetyl glyceryl ether phosphorylcholine(platelet-activating factor). Vet Immun Immunpath 1987;7:81–7.
[24] Dupuis F, Rougier F, Trimoreau F, Ostyn E, Dulery C, Praloran V, Denizot Y. Production and metabolismof platelet-activating factor by human bone marrow cells. Res Immunol 1997;148:119–26.
[25] Casals-Stenzel J, Muaceivic G, Weber KH. Pharmacological actions of WEB 2086, a new specificantagonist of platelet-activating factor. J Pharm Exp Therap 1987;241:974–81.
[26] Laemmli VK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature1970;227:680–6.
[27] Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the folin phenol reagent. J BiolChem 1951;193:265–9.
[28] Schatz F, Markiewicz, Gurpide E. Differential effects of estradiol, arachidonic acid, and A23187 onprostaglandin F2a output by epithelial and stromal cells of human endometrium. Endocrinology 1987;120:1465–71.
[29] Harper MJK. Platelet-activating factor: a paracrine factor in preimplantation stages of development? BiolReprod 1989;40:907–13.
[30] Maggi M, Bonaccorsi B, Finetti G, Carloni V, Muratori M, Laffi G, Forti G, Serio M, Baldi E. Platelet-activating factor mediates an autocrine proliferative loop in the endometrial adenocarinoma cell lineHEC-1A. Cancer Res 1994;54:4777–84.
[31] Tiemann U, Neels P, Po¨hland R, Walzel H, Lo¨hrke B. Influence on inhibitors on increase in intracellular freecalcium and proliferation induced by platelet-activating factor in bovine oviductal cells. J Reprod Fertil1999;116:63–72.
[32] Nasu K, Narahara H, Matsui N, Kawano Y, Tanaka Y, Miyakawa I. Platelet-activating factor stimulatescytokine production by human endometrial stromal cells. Mol Hum Reprod 1999;5:548–53.
[33] Shen TY, Hussaini I, Hwang S-B, Chang MN. Recent development of platelet-activating factor antagonists.Adv Prosta Throm Leuko Res 1989;19:359–62.
140 U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 125–141

[34] Muller E, Dagenais P, Alami N, Rola-Pleszczynski M. Identification and functional characterization ofplatelet-activating factor receptors in human leukocyte populations using polyclonal anti-peptide antibody.Proc Natl Acad Sci USA 1993;90:5818–22.
[35] Kunz D, Gerard NP, Gerard C. The human leukocyte platelet-activating factor receptor. J Biol Chem1992;267:9101–6.
[36] Nakamura M, Honda Z, Izumi T, Sakanaka C, Mutoh H, Minami M, Bito H, Seyama Y, Matsumoto T,Noma M, Shimizu T. Molecular cloning and expression of platelet-activating factor receptor from humanleukocytes. J Biol Chem 1991;266:20400–5.
[37] Predescu D, Ihida K, Predescu S, Palada GE. The vascular distribution of the platelet-activating factorreceptor. Euro J Cell Biol 1996;69:86–98.
[38] Ahmed A, Dearn S. The role of platelet-activating factor and its receptor in endometrial receptivity. In:Platelet-activating factor and related lipid mediators. Nigram et al., editors. New York: Plenum Press, 1996.p. 277–90.
[39] Tiemann U, Viergutz T, Jonas L, Wollenhaupt K, Po¨hland R, Kanitz W. Fluorometric detection of plateletactivating factor receptor in cultured oviductal epithelial and stromal cells and endometrial stromal cellsfrom bovine at different stages of the oestrus cycle and early pregnancy. Domest Anim Endocrin 2001 (inpress).
[40] Kloprogge E, Akkerman JWN. Binding kinetics of PAF-acether [1–0-alkyl-2-acetyl-sn-glycero-3-phospho-choline] to intact human platelets. Biochem J 1984;223:901–9.
[41] Nguer CM, Pellegrin O, Galanaud P, Benveniste J, Thoman Y, Richard Y. Regulation of PAF-acetherreceptor expression in human B cells. J Immunol 1992;149:2742–8.
[42] Chao W, Lui H, Hanahan DJ, Olson MS. Platelet-activating factor stimulated protein tyrosine phosphory-lation and eicosanoid synthesis in rat Kuppfer cells. J Biol Chem 1989;264:20448–57.
[43] Shirasaki H, Adocok IM, Kwon OJ, Nishikawa M, Mak JC, Barnes PJ. Agonist induced up-regulation ofplatelet-activating factor receptor messenger RNA in human monocytes. Eur J Pharmacol 1994;268:263–6.
[44] Stafforini DM, Prescott GA, McIntyre TM. Human plasma platelet-activating factor acetylhydrolase.Purification and properties. J Biol Chem 1987;262:4223–30.
[45] Akiyama M, Sugatani J, Suzuki T, Suzuki Y, Miwa M. Identification of a major PAF acetylhydrolase inhuman serum/plasma as a 43 kDa glycoprotein containing about 9 kDa asparagine-conjugated sugarchain(s). J Biochem 1998;123:786–9.
[46] Ohshige A, Masaharu I, Koyama T, Maeda T, Yoshihiro T, Okamura H. Effects of oestrogen andprogesterone on platelet-activating factor acetylhydrolase activity in ovarectomized rats. Artery 1994;21:234–42.
141U. Tiemann et al. / Prostaglandins & other Lipid Mediators 65 (2001) 000–000


![[ Home ] [ PAF News ] [ Checklist ] [ PAF Feedback ] [ PAF ...€¦ · [ Home ] [ PAF News ] [ Checklist ] [ PAF Feedback ] [ PAF Search Page ] [ General aspects ] [ Case studies](https://img.pdfslide.net/doc/110x75/5f2c6cae89a8d014356437ba/-home-paf-news-checklist-paf-feedback-paf-home-paf-news.jpg)


























