Embed Size (px)
Citation preview

DOI: 10.1126/science.1239278, 651 (2013);341 Science et al.Pingping Hou
Small-Molecule CompoundsPluripotent Stem Cells Induced from Mouse Somatic Cells by
This copy is for your personal, non-commercial use only.
clicking here.colleagues, clients, or customers by , you can order high-quality copies for yourIf you wish to distribute this article to others
here.following the guidelines
can be obtained byPermission to republish or repurpose articles or portions of articles
): April 2, 2014 www.sciencemag.org (this information is current as of
The following resources related to this article are available online at
http://www.sciencemag.org/content/341/6146/651.full.htmlversion of this article at:
including high-resolution figures, can be found in the onlineUpdated information and services,
http://www.sciencemag.org/content/suppl/2013/07/17/science.1239278.DC1.html can be found at: Supporting Online Material
http://www.sciencemag.org/content/341/6146/651.full.html#relatedfound at:
can berelated to this article A list of selected additional articles on the Science Web sites
http://www.sciencemag.org/content/341/6146/651.full.html#ref-list-1, 6 of which can be accessed free:cites 37 articlesThis article
http://www.sciencemag.org/content/341/6146/651.full.html#related-urls12 articles hosted by HighWire Press; see:cited by This article has been
http://www.sciencemag.org/cgi/collection/developmentDevelopment
subject collections:This article appears in the following
registered trademark of AAAS. is aScience2013 by the American Association for the Advancement of Science; all rights reserved. The title
CopyrightAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. (print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by theScience
on
Apr
il 2,
201
4w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
o
n A
pril
2, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
on
Apr
il 2,
201
4w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
o
n A
pril
2, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
on
Apr
il 2,
201
4w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

16. C. A. Hidalgo, R. A. Castro, C. Rodriguez-Sickert, New J.Phys. 8, 52 (2006).
17. E. Bakshy, B. Karrer, L. A. Adamic, Social influence andthe diffusion of user created content, in Proceedings ofthe 10th ACM Conference on Electronic Commerce(Association for Computing Machinery, New York, 2009),pp. 325–334.
18. J. Leskovec, A. Singh, J. Kleinberg, Patterns of influencein a recommendation network, in Advances in KnowledgeDiscovery and Data Mining, Lecture Notes in ComputerScience (Springer, Berlin, Heidelberg, 2006), vol. 3918,pp. 380–389.
19. F. Wu, B. A. Huberman, Internet and Network Economics5385, 334–341 (2008).
20. N. Archak, A. Ghose, P. G. Ipeirotis, Manage. Sci. 57,1485–1509 (2011).
21. C. F. Manski, Rev. Econ. Stud. 60, 531–542(1993).
22. S. Aral, L. Muchnik, A. Sundararajan, Proc. Natl. Acad.Sci. U.S.A. 106, 21544–21549 (2009).
23. S. Aral, Mark. Sci. 30, 217–223 (2011).24. S. Aral, D. Walker, IEEE Intell. Syst. 26, 91–96
(2011).25. S. Aral, D. Walker, Manage. Sci. 57, 1623–1639
(2011).26. S. Aral, D. Walker, Science 337, 337–341 (2012).27. C. R. Shalizi, A. C. Thomas, Sociol. Methods Res. 40,
211–239 (2011).28. C. L. Apicella, F. W. Marlowe, J. H. Fowler, N. A.
Christakis, Nature 481, 497–501 (2012).29. D. Centola, R. Willer, M. Macy, Am. J. Sociol. 110,
1009–1040 (2005).
Acknowledgments: We thank D. Eckles and D. Watts forinvaluable discussions. This work was supported by a MicrosoftResearch faculty fellowship (S.A.) and by NSF Career Award0953832 (S.A.). The research was approved by the New YorkUniversity institutional review board. There are legal obstaclesto making the data available and revealing the name of theWeb site, but code is available upon request. All of the user
data that we analyzed is publically available, except thetreatment assignment and the random identifier that links adeidentified user to a vote, and therefore does not contain anyother information that cannot be obtained by crawling theWeb site. The randomized testing that the Web site performedis covered by the Web site’s terms of service. Opt-inpermissions were granted by the users when they registeredfor the Web site. No data on nonregistered users was collected.
Supplementary Materialswww.sciencemag.org/cgi/content/full/341/6146/647/DC1Materials and MethodsSupplementary TextTables S1 to S21Figs. S1 to S16References (30–57)
14 May 2013; accepted 16 July 201310.1126/science.1240466
Pluripotent Stem Cells Inducedfrom Mouse Somatic Cellsby Small-Molecule CompoundsPingping Hou,1* Yanqin Li,1* Xu Zhang,1,2* Chun Liu,1,2* Jingyang Guan,1* Honggang Li,1*Ting Zhao,1† Junqing Ye,1,2† Weifeng Yang,3† Kang Liu,1† Jian Ge,1,2† Jun Xu,1† Qiang Zhang,1,2†Yang Zhao,1‡ Hongkui Deng1,2‡
Pluripotent stem cells can be induced from somatic cells, providing an unlimited cellresource, with potential for studying disease and use in regenerative medicine. However,genetic manipulation and technically challenging strategies such as nuclear transferused in reprogramming limit their clinical applications. Here, we show that pluripotentstem cells can be generated from mouse somatic cells at a frequency up to 0.2% using acombination of seven small-molecule compounds. The chemically induced pluripotentstem cells resemble embryonic stem cells in terms of their gene expression profiles, epigeneticstatus, and potential for differentiation and germline transmission. By using small molecules,exogenous “master genes” are dispensable for cell fate reprogramming. This chemicalreprogramming strategy has potential use in generating functional desirable cell typesfor clinical applications.
Pluripotent stem cells, such as embryonicstem cells (ESCs), can self-renew anddifferentiate into all somatic cell types.
Somatic cells can be reprogrammed to becomepluripotent via nuclear transfer into oocytes orthrough the ectopic expression of defined factors(1–4). However, exogenous pluripotency-associatedfactors, especially Oct4, are indispensable for es-tablishing pluripotency (5–7), and previous repro-gramming strategies have raised concerns regardingthe clinical applications (8, 9). Small moleculeshave advantages because they can be cell perme-able, nonimmunogenic, more cost-effective, andmore easily synthesized, preserved, and standard-
ized. Moreover, their effects on inhibiting andactivating the function of specific proteins areoften reversible and can be finely tuned by varyingthe concentrations. Here, we identified small-molecule combinations that were able to drive thereprogramming of mouse somatic cells towardpluripotent cells.
To identify small molecules that facilitate cellreprogramming, we searched for small moleculesthat enable reprogramming in the absence ofOct4usingOct4 promoter-driven green fluorescent pro-tein (GFP) expression (OG) mouse embryonicfibroblasts (MEFs), with viral expression of Sox2,Klf4, and c-Myc. After screening up to 10,000 smallmolecules (table S1A), we identified Forskolin(FSK), 2-methyl-5-hydroxytryptamine (2-Me-5HT), and D4476 (table S1B) as chemical “sub-stitutes” for Oct4 (Fig. 1, A and B, and figs. S1and S2). Previously, we had developed a small-molecule combination “VC6T” [VPA,CHIR99021(CHIR), 616452, Tranylcypromine], that enablesreprogramming with a single gene, Oct4 (6). Wenext treated OG-MEFswith VC6T plus the chem-
ical substitutes of Oct4 in the absence of trans-genes. We found that VC6T plus FSK (VC6TF)induced some GFP-positive clusters expressingE-cadherin, a mesenchyme-to-epithelium transi-tion marker, reminiscent of early reprogrammingby transcription factors (10, 11) (Fig. 1C and fig.S3). However, the expression ofOct4 andNanogwas not detectable, and their promoters remainedhypermethylated, suggesting a repressed epige-netic state (fig. S3).
To identify small molecules that facilitate latereprogramming, we used a doxycycline (DOX)–inducible Oct4 expression screening system,addingDOXonly in the first 4 to 8 days (6). Small-molecule hits, including several cAMP agonists(FSK, Prostaglandin E2, and Rolipram) and epi-genetic modulators [3-deazaneplanocin A (DZNep),5-Azacytidine, sodium butyrate, and RG108], wereidentified in this screen (fig. S4 and table S1B).
To achieve complete chemical reprogrammingwithout the Oct4-inducible system, these smallmolecules were further tested in the chemical re-programming of OG-MEFs without transgenes.When DZNep was added 16 days after treatmentwith VC6TF (VC6TFZ), GFP-positive cells wereobtained more frequently by a factor of up to 65than those treated with VC6TF, forming compact,epithelioid, GFP-positive colonies without clear-cut edges (Fig. 1, D and E, and fig. S5). In thesecells, the expression levels of most pluripotencymarker genes were elevated but were still lowerthan in ESCs, suggesting an incomplete repro-gramming state (fig. S6). After switching to 2i-medium with dual inhibition (2i) of glycogensynthase kinase-3 and mitogen-activated proteinkinase signaling after day 28 posttreatment, cer-tain GFP-positive colonies developed an ESC-likemorphology (domed, phase-bright, homogeneouswith clear-cut edges) (Fig. 1F) (12, 13). These col-onies could be further cultured for more than 30passages, maintaining an ESC-like morphology(Fig. 1, G and H).We refer to these 2i-competent,ESC-like, and GFP-positive cells as chemicallyinduced pluripotent stem cells (CiPSCs).
Next, we optimized the dosages and treatmentduration of the small molecules and were ableto generate 1 to 20 CiPSC colonies from 50,000
1College of Life Sciences and Peking-Tsinghua Center for LifeSciences, Peking University, Beijing 100871, China. 2Schoolof Chemical Biology and Biotechnology, Peking UniversityShenzhen Graduate School, Shenzhen 518055, China. 3BeijingVitalstar Biotechnology Co., Ltd., Beijing 100012, China.
*These authors contributed equally to this work.†These authors contributed equally to this work.‡Corresponding author. E-mail: [email protected](H.D.); [email protected] (Y.Z.)
www.sciencemag.org SCIENCE VOL 341 9 AUGUST 2013 651
REPORTS

initially platedMEFs (fig. S7). After an additionalscreen, we identified some small-molecule boostersof chemical reprogramming, among which, a syn-thetic retinoic acid receptor ligand, TTNPB, en-hanced chemical reprogramming efficiency upto a factor of 40, to a frequency comparable totranscription factor–induced reprogramming (upto 0.2%) (fig. S8 and table S1B). Furthermore,
using the small-molecule combination VC6TFZ,we obtained CiPSC lines from mouse neonatalfibroblasts (MNFs),mouse adult fibroblasts (MAFs),and adipose-derived stem cells (ADSCs) withOG cassettes by an efficiency lower by a factorof ~10 than that obtained from MEFs (fig. S9and table S3). Moreover, we induced CiPSCsfrom wild-type MEFs without OG cassettes or
any other genetic modifications by a comparableefficiency to that achieved from MEFs with OGcassettes (fig. S9). The CiPSCswere also confirmedto be viral-vector free by genomic polymerasechain reaction (PCR) and Southern blot analysis(fig. S10).
The established CiPSC lines were then fur-ther characterized. They grew with a doubling
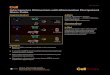
Fig. 1. Generation of CiPSCs by small-molecule compounds.(A and B) Numbers of iPSC colonies induced from MEFs infectedby SKM (A) or SK (B) plus chemicals or Oct4. Error bars, mean TSD (n = 3 biological repeat wells). (C) Morphology of MEFs forchemical reprogramming on day 0 (D0) and a GFP-positivecluster generated using VC6TF on day 20 (D20) after chemicaltreatment. (D) Numbers of GFP-positive colonies induced afterDZNep treatment on day 36. Error bars, mean T SD (n = 2biological repeat wells). (E to G) Morphology of a compact,epithelioid, GFP-positive colony on day 32 (D32) after treatment(E), a primary CiPSC colony on day 40 (D40) after treatment (F),and passaged CiPSC colonies (G). (H) Schematic diagram illustrat-ing the process of CiPSC generation. Scale bars, 100 mm. For(D), cells for reprogramming were replated on day 12.
B
Num
ber
of iP
SC
col
onie
spe
r 40
,000
cel
ls
(-) co
ntro
l0
2-M
e-5H
T
D4476
FSKOct4
10
20
30
40
50SK plusA
(-) co
ntro
l
2-M
e-5H
T
D4476
FSKOct4
Num
ber
of iP
SC
col
onie
spe
r 30
,000
cel
ls
100
75
50
25
0
SKM plus
(Replate)
0 12 16-20
MEFs Epithelioid and compact GFP+ colonies
CiPSCs
Chemical Reprogramming Medium 2i-Medium
GFP- colonies
Pick up
GFP+ cells/clusters
28-36
VC6TFZVC6TF
Plate
-1 (Days)
D G
H
C D0
D20
E D32
D40F
VC6TFZ
VC6TF
Num
ber
of G
FP
+ c
olon
ies
per
500,
000
repl
ated
cel
ls
20
15
10
5
0
Fig. 2. Characterization of CiPSCs. (A and B)Pluripotency marker expression as illustrated by im-munofluorescence (A, clone CiPS-25) and RT-PCR (B).Scale bars, 100 mm. (C) Hierarchical clustering ofglobal transcriptional profiles. 1-PCC, Pearson corre-lation coefficient. (D) Bisulfite genomic sequencing ofthe Oct4 and Nanog promoter regions. MNF-CiPS-7,MNF-derived CiPSC line no. 7; MAF-CiPS-1, MAF-derived CiPSC line no. 1.
DNanog Oct4
MEFs
CiPS-21
MAF-CiPS-1
MNF-CiPS-7
CiPS-36
ESCs
CA
Oct4
Nanog
Sox2
Utf1
Rex1
Fbxo15
Cripto
Dax1
Gapdh
MEFs
ESCs
OSKM-iP
S
CiPS-4
7
CiPS-3
0
CiPS-2
1
CiPS-3
4
CiPS-5
0
CiPS-4
5
CiPS-5
6B
UTF1
SSEA1 NANOG
OCT4
0.1
0.2
MEFs
CiPS-3
4
MNF-C
iPS-7
CiPS-5
0
CiPS-2
1
OSKM-iP
SESCs
1-P
CC
9 AUGUST 2013 VOL 341 SCIENCE www.sciencemag.org652
REPORTS
time (14.1 to 15.1 hours) similar to that of ESCs(14.7 hours), maintained alkaline phosphataseactivity, and expressed pluripotency markers, asdetected by immunofluorescence and reversetranscription (RT)-PCR (Fig. 2, A and B, andfig. S11). The gene expression profiles were sim-ilar in CiPSCs, ESCs, and OSKM-iPSCs (iPSCsinduced by Oct4, Sox2, Klf4, and c-Myc) (Fig. 2Cand fig. S12).DNAmethylation state and histonemodifications at Oct4 and Nanog promoters inCiPSCs were similar to that in ESCs (Fig. 2Dand fig. S13). In addition, CiPSCs maintained anormal karyotype and genetic integrity for up to13 passages (fig. S14 and table S2).
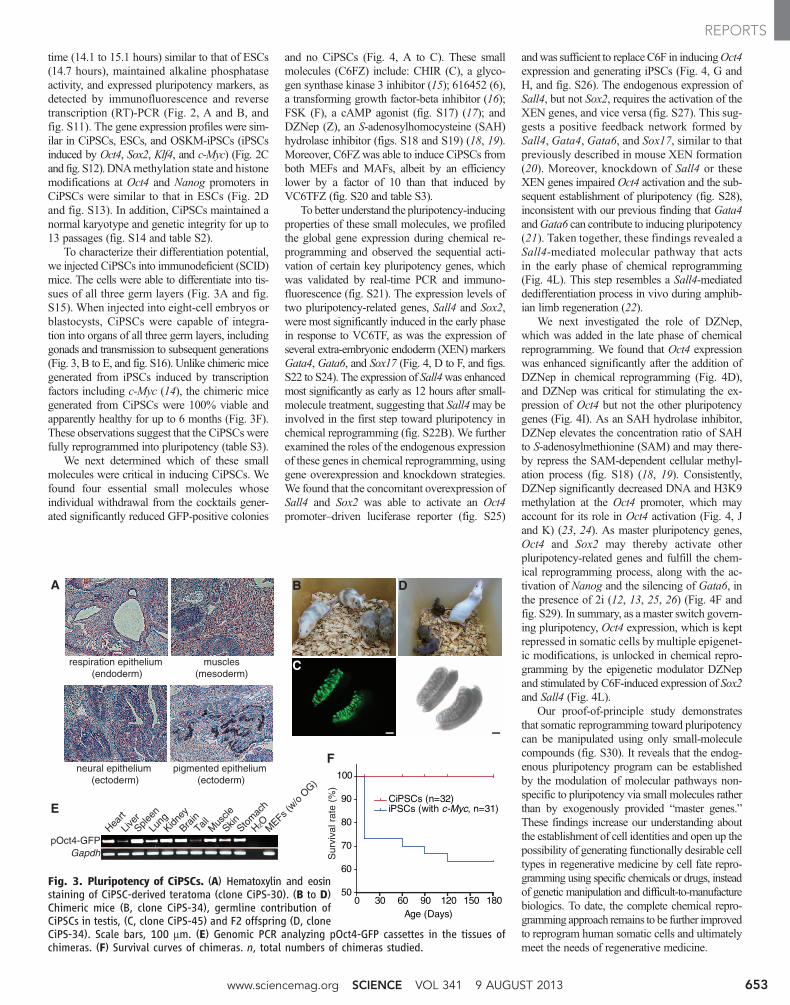
To characterize their differentiation potential,we injected CiPSCs into immunodeficient (SCID)mice. The cells were able to differentiate into tis-sues of all three germ layers (Fig. 3A and fig.S15). When injected into eight-cell embryos orblastocysts, CiPSCs were capable of integra-tion into organs of all three germ layers, includinggonads and transmission to subsequent generations(Fig. 3, B to E, and fig. S16). Unlike chimeric micegenerated from iPSCs induced by transcriptionfactors including c-Myc (14), the chimeric micegenerated from CiPSCs were 100% viable andapparently healthy for up to 6 months (Fig. 3F).These observations suggest that the CiPSCs werefully reprogrammed into pluripotency (table S3).
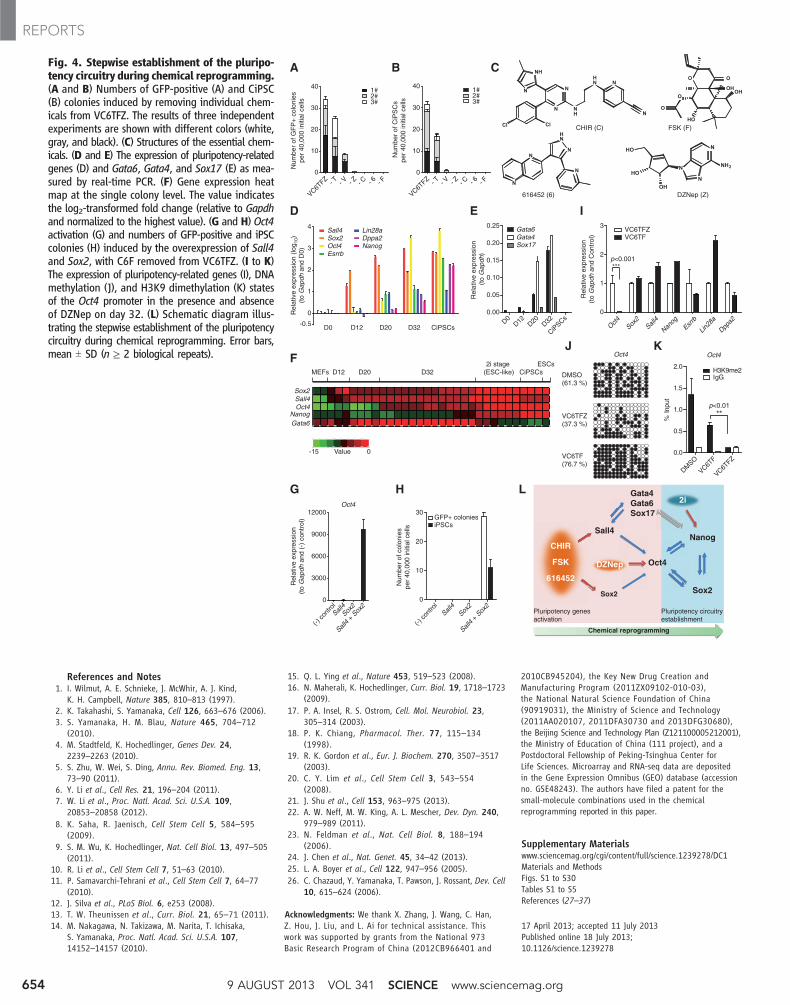
We next determined which of these smallmolecules were critical in inducing CiPSCs. Wefound four essential small molecules whoseindividual withdrawal from the cocktails gener-ated significantly reduced GFP-positive colonies
and no CiPSCs (Fig. 4, A to C). These smallmolecules (C6FZ) include: CHIR (C), a glyco-gen synthase kinase 3 inhibitor (15); 616452 (6),a transforming growth factor-beta inhibitor (16);FSK (F), a cAMP agonist (fig. S17) (17); andDZNep (Z), an S-adenosylhomocysteine (SAH)hydrolase inhibitor (figs. S18 and S19) (18, 19).Moreover, C6FZ was able to induce CiPSCs fromboth MEFs and MAFs, albeit by an efficiencylower by a factor of 10 than that induced byVC6TFZ (fig. S20 and table S3).
To better understand the pluripotency-inducingproperties of these small molecules, we profiledthe global gene expression during chemical re-programming and observed the sequential acti-vation of certain key pluripotency genes, whichwas validated by real-time PCR and immuno-fluorescence (fig. S21). The expression levels oftwo pluripotency-related genes, Sall4 and Sox2,were most significantly induced in the early phasein response to VC6TF, as was the expression ofseveral extra-embryonic endoderm (XEN) markersGata4, Gata6, and Sox17 (Fig. 4, D to F, and figs.S22 to S24). The expression of Sall4was enhancedmost significantly as early as 12 hours after small-molecule treatment, suggesting that Sall4may beinvolved in the first step toward pluripotency inchemical reprogramming (fig. S22B). We furtherexamined the roles of the endogenous expressionof these genes in chemical reprogramming, usinggene overexpression and knockdown strategies.We found that the concomitant overexpression ofSall4 and Sox2 was able to activate an Oct4promoter–driven luciferase reporter (fig. S25)
andwas sufficient to replaceC6F in inducingOct4expression and generating iPSCs (Fig. 4, G andH, and fig. S26). The endogenous expression ofSall4, but not Sox2, requires the activation of theXEN genes, and vice versa (fig. S27). This sug-gests a positive feedback network formed bySall4, Gata4, Gata6, and Sox17, similar to thatpreviously described in mouse XEN formation(20). Moreover, knockdown of Sall4 or theseXEN genes impaired Oct4 activation and the sub-sequent establishment of pluripotency (fig. S28),inconsistent with our previous finding that Gata4andGata6 can contribute to inducing pluripotency(21). Taken together, these findings revealed aSall4-mediated molecular pathway that actsin the early phase of chemical reprogramming(Fig. 4L). This step resembles a Sall4-mediateddedifferentiation process in vivo during amphib-ian limb regeneration (22).
We next investigated the role of DZNep,which was added in the late phase of chemicalreprogramming. We found that Oct4 expressionwas enhanced significantly after the addition ofDZNep in chemical reprogramming (Fig. 4D),and DZNep was critical for stimulating the ex-pression of Oct4 but not the other pluripotencygenes (Fig. 4I). As an SAH hydrolase inhibitor,DZNep elevates the concentration ratio of SAHto S-adenosylmethionine (SAM) and may there-by repress the SAM-dependent cellular methyl-ation process (fig. S18) (18, 19). Consistently,DZNep significantly decreased DNA and H3K9methylation at the Oct4 promoter, which mayaccount for its role in Oct4 activation (Fig. 4, Jand K) (23, 24). As master pluripotency genes,Oct4 and Sox2 may thereby activate otherpluripotency-related genes and fulfill the chem-ical reprogramming process, along with the ac-tivation of Nanog and the silencing of Gata6, inthe presence of 2i (12, 13, 25, 26) (Fig. 4F andfig. S29). In summary, as a master switch govern-ing pluripotency, Oct4 expression, which is keptrepressed in somatic cells by multiple epigenet-ic modifications, is unlocked in chemical repro-gramming by the epigenetic modulator DZNepand stimulated by C6F-induced expression of Sox2and Sall4 (Fig. 4L).
Our proof-of-principle study demonstratesthat somatic reprogramming toward pluripotencycan be manipulated using only small-moleculecompounds (fig. S30). It reveals that the endog-enous pluripotency program can be establishedby the modulation of molecular pathways non-specific to pluripotency via small molecules ratherthan by exogenously provided “master genes.”These findings increase our understanding aboutthe establishment of cell identities and open up thepossibility of generating functionally desirable celltypes in regenerative medicine by cell fate repro-gramming using specific chemicals or drugs, insteadof genetic manipulation and difficult-to-manufacturebiologics. To date, the complete chemical repro-gramming approach remains to be further improvedto reprogram human somatic cells and ultimatelymeet the needs of regenerative medicine.
0 30 60 90 120 150 18050
60
70
80
90
100
iPSCs (with c-Myc, n=31)CiPSCs (n=32)
Age (Days)
Sur
viva
l rat
e (%
)
F
Heart
LiverSple
en
Lung
Kidney
BrainTail M
uscle
SkinStom
ach
H2OMEFs (
w/o O
G)
pOct4-GFPGapdh
B D
C
A
respiration epithelium (endoderm)
muscles (mesoderm)
neural epithelium (ectoderm)
pigmented epithelium (ectoderm)
E
Fig. 3. Pluripotency of CiPSCs. (A) Hematoxylin and eosinstaining of CiPSC-derived teratoma (clone CiPS-30). (B to D)Chimeric mice (B, clone CiPS-34), germline contribution ofCiPSCs in testis, (C, clone CiPS-45) and F2 offspring (D, cloneCiPS-34). Scale bars, 100 mm. (E) Genomic PCR analyzing pOct4-GFP cassettes in the tissues ofchimeras. (F) Survival curves of chimeras. n, total numbers of chimeras studied.
www.sciencemag.org SCIENCE VOL 341 9 AUGUST 2013 653
REPORTS
References and Notes1. I. Wilmut, A. E. Schnieke, J. McWhir, A. J. Kind,
K. H. Campbell, Nature 385, 810–813 (1997).2. K. Takahashi, S. Yamanaka, Cell 126, 663–676 (2006).3. S. Yamanaka, H. M. Blau, Nature 465, 704–712
(2010).4. M. Stadtfeld, K. Hochedlinger, Genes Dev. 24,
2239–2263 (2010).5. S. Zhu, W. Wei, S. Ding, Annu. Rev. Biomed. Eng. 13,
73–90 (2011).6. Y. Li et al., Cell Res. 21, 196–204 (2011).7. W. Li et al., Proc. Natl. Acad. Sci. U.S.A. 109,
20853–20858 (2012).8. K. Saha, R. Jaenisch, Cell Stem Cell 5, 584–595
(2009).9. S. M. Wu, K. Hochedlinger, Nat. Cell Biol. 13, 497–505
(2011).10. R. Li et al., Cell Stem Cell 7, 51–63 (2010).11. P. Samavarchi-Tehrani et al., Cell Stem Cell 7, 64–77
(2010).12. J. Silva et al., PLoS Biol. 6, e253 (2008).13. T. W. Theunissen et al., Curr. Biol. 21, 65–71 (2011).14. M. Nakagawa, N. Takizawa, M. Narita, T. Ichisaka,
S. Yamanaka, Proc. Natl. Acad. Sci. U.S.A. 107,14152–14157 (2010).
15. Q. L. Ying et al., Nature 453, 519–523 (2008).16. N. Maherali, K. Hochedlinger, Curr. Biol. 19, 1718–1723
(2009).17. P. A. Insel, R. S. Ostrom, Cell. Mol. Neurobiol. 23,
305–314 (2003).18. P. K. Chiang, Pharmacol. Ther. 77, 115–134
(1998).19. R. K. Gordon et al., Eur. J. Biochem. 270, 3507–3517
(2003).20. C. Y. Lim et al., Cell Stem Cell 3, 543–554
(2008).21. J. Shu et al., Cell 153, 963–975 (2013).22. A. W. Neff, M. W. King, A. L. Mescher, Dev. Dyn. 240,
979–989 (2011).23. N. Feldman et al., Nat. Cell Biol. 8, 188–194
(2006).24. J. Chen et al., Nat. Genet. 45, 34–42 (2013).25. L. A. Boyer et al., Cell 122, 947–956 (2005).26. C. Chazaud, Y. Yamanaka, T. Pawson, J. Rossant, Dev. Cell
10, 615–624 (2006).
Acknowledgments: We thank X. Zhang, J. Wang, C. Han,Z. Hou, J. Liu, and L. Ai for technical assistance. Thiswork was supported by grants from the National 973Basic Research Program of China (2012CB966401 and
2010CB945204), the Key New Drug Creation andManufacturing Program (2011ZX09102-010-03),the National Natural Science Foundation of China(90919031), the Ministry of Science and Technology(2011AA020107, 2011DFA30730 and 2013DFG30680),the Beijing Science and Technology Plan (Z121100005212001),the Ministry of Education of China (111 project), and aPostdoctoral Fellowship of Peking-Tsinghua Center forLife Sciences. Microarray and RNA-seq data are depositedin the Gene Expression Omnibus (GEO) database (accessionno. GSE48243). The authors have filed a patent for thesmall-molecule combinations used in the chemicalreprogramming reported in this paper.
Supplementary Materialswww.sciencemag.org/cgi/content/full/science.1239278/DC1Materials and MethodsFigs. S1 to S30Tables S1 to S5References (27–37)
17 April 2013; accepted 11 July 2013Published online 18 July 2013;10.1126/science.1239278
Fig. 4. Stepwise establishment of the pluripo-tency circuitry during chemical reprogramming.(A and B) Numbers of GFP-positive (A) and CiPSC(B) colonies induced by removing individual chem-icals from VC6TFZ. The results of three independentexperiments are shown with different colors (white,gray, and black). (C) Structures of the essential chem-icals. (D and E) The expression of pluripotency-relatedgenes (D) and Gata6, Gata4, and Sox17 (E) as mea-sured by real-time PCR. (F) Gene expression heatmap at the single colony level. The value indicatesthe log2-transformed fold change (relative to Gapdhand normalized to the highest value). (G and H) Oct4activation (G) and numbers of GFP-positive and iPSCcolonies (H) induced by the overexpression of Sall4and Sox2, with C6F removed from VC6TFZ. (I to K)The expression of pluripotency-related genes (I), DNAmethylation (J), and H3K9 dimethylation (K) statesof the Oct4 promoter in the presence and absenceof DZNep on day 32. (L) Schematic diagram illus-trating the stepwise establishment of the pluripotencycircuitry during chemical reprogramming. Error bars,mean T SD (n ≥ 2 biological repeats).
9 AUGUST 2013 VOL 341 SCIENCE www.sciencemag.org654
REPORTS