Embed Size (px)
Citation preview

1
Doktori (Ph.D.) értekezés
Poli(ADP-ribóz) polimeráz (PARP) szerepe a bőr ultribolya (UV) fény indukálta károsodásában
Dr. Csete Béla
Interdiszciplinális doktori iskola vezető: Dr. Sümegi Balázs, egyetemi
tanár
Alprogramvezető: Dr. Farkas Beatrix, egyetemi tanár
Dr. Battyáni Zita, egyetemi docens
Pécsi Tudományegyetem
2009.

2
Tartalomjegyzék
Bevezetés 2.
Célkitűzések 10.
I. Anyag és módszer 12.
I/1. Állatkísérletek 12.
I/2. Klinikai tünetek értékelése 24.
II. Eredmények 30.
III. Összefoglalás 45.
III/1. Témában elért új eredmények 46.
Irodalomjegyzék 56.
Rövidítések jegyzéke 66.
Köszönetnyilvánítás 67.
A szerző közleményeinek jegyzéke 68.
A szerző kongresszusi előadásainak jegyzéke 70.
Táblázatok és Ábrák 72.

3
Bevezetés
A bőr multifunkcionális „interface” a külvilág és a szervezet között. A
környezeti tényezők (légköri szennyeződés, inszoláció fokozódása, az
ózonlyuk növekedése, stb.), valamint az életmódbeli szokások (nyári, téli
üdülés, hétvégi szabadtéri tevékenység, mesterséges ultraibolya (UV)
besugárzás rendszeres igénybevétele, stb.) radikális változása az UV-fény
okozta bőrkárosodás pontos patomechanizmusának megismerését, valamint
ezáltal a folyamat megelőzésére és kezelésére szolgáló eljárások
kidolgozását világszerte a bőrgyógyászat egyik vezető kutatási területévé
tette.
Az UV fény három különböző tartományra osztható. Így az UVA 320-400 nm
(ezen belűl UVAI: 340-400 nm, UVAII: 320-340 nm), az UVB: 290-320 nm és
az UVC: 200-290 nm formájában lett meghatározva (Diffey 2002). Ismert,
hogy az UV-sugarak hullámhossz-dependens módon penetrálnak a bőrbe,
vagyis a hullámhossz növekedésével a behatolás mélysége nő. A rövidebb
hullámhosszú UVB-sugárzás főleg az epidermiszben abszorbeálódik, és ott
dominálóan a keratinocitákkal lép interakcióba. A hosszabb hullámhosszú
UVA-tartomány a mélybe penetrál, és az epidermisz mellett érinti a dermális
sejtek funkcióját is (Höningsmann 2002). Az UVA- és UVB-tartomány káros
hatása additív jellegű és kumulálódik (Lim és mtsai. 2001, Kurtmann 2001)
(1.ábra).
Annak ellenére, hogy a Földet érő elektromágneses hullámból az UVA-
tartomány aránya lényegesen magasabb (95%), mint az UVB-é, a
biológiailag káros hatások kiváltását dominálóan az UV-fény UVB-

4
tartományával hozzák összefüggésbe (Elmets és mtsai. 2001, Cole 2001,
Lim és mtsai. 2001).
Az UV-fény legtöbb, bőrben jelentkező kóros biológiai hatásának fő
molekuláris „targetje” a DNS. Az epidermisz számos kromoforral
(nukleinsavak, urokainsav, aromás aminosavak, melanin prekurzorok, stb.)
rendelkezik, melyek a fény UV-tartományát elnyelik, és fotokémiai reakción
keresztül a bőr biológiai károsodását idézhetik elő (Hönigsmann 2002). Az
UV-fény direkt DNS-károsító hatása során dominálóan ciklobután-pirimidin-
dimerek (CPD), valamint pirimidin (6-4) pirimidon fotoproduktumok
képződnek (Freeman és mtsai. 1989, Hemminki és mtsai. 2001). A direkt
DNS-károsodás kiváltásában az UVB- és a rövid UVA- (UVA-II)
tartománynak van szerepe (Young és mtsai. 1998) A fény UVA-I tartománya
főleg indirekt módon, ROS indukció által fejti ki hatását, mely a DNS
oxidációja mellett, a lipid-peroxidációban, a fehérjék oxidációjában, a
transzkripciós faktorok aktivációjában, DNS-törések előidézésében nyilvánul
meg (Krutmann és mtsai. 1997). A fény UVB-tartománya szintén képes ROS
képződést indukálni, direkt hatása mégis a DNS-fototermékek (timin-
dimerek), egyesláncú DNS-törések képződésén, a DNS-repair enzimek
aktivációján keresztül jelentősebb (Katiyar és mtsai. 2000, Berton és mtsai.
1997, Krutmann 2001, Mitchell és mtsai. 2001). Az epidermisz sejtjeiben
bekövetkező DNS-károsodás mértéke jól követhető az akut UV-fény stressz
hatására létre jött apoptótikus keratinocyták („sunburn sejtek”) kvantitatív
meghatározásával (Ziegler és mtsai. 1994).
Az UV-sugárzás direkt és indirekt DNS-t károsító hatása a bőrben gyulladás,
fototoxikus reakció, immunszupresszió, „photoaging”, fotokarcinogenezis és

5
számos egyéb elváltozás formájában nyilvánul meg (Krutmann 2001, Kulms
és mtsa. 2000, Yarosh és mtsai. 2001, Maccarrone és mtsai. 1997,
Steenvoorden és mtsai. 1997) (2. Ábra). Általában az UV-sugárzás akut
tüneteinek kiváltásáért, amely dominálóan az epidermális keratinocitákra
kifejtett különböző mértékű citotoxikus károsodás következménye, a fény
UVB-tartományát teszik felelőssé (Elmets és mtsai. 2001, Lim és mtsai.
2001). A napégés, számos jelátviteli útvonalon keresztül, önmaga elégséges
lehet az évekkel, évtizedekkel később manifesztálódó melanoma és nem-
melanoma bőrtumorok (carcinoma spinocellulare, carcinoma basocellulare)
indukciójához. A krónikus fénykárosodás, vagy „photoaging” kiváltásában az
UV-fény mindkét tartományának szerepe van, hiszen hatásuk additív, mégis
az UVA jelentőségét emelik ki. Az UV-fény okozta öregedés („photoaging”)
és fotokarcinogenezis (melanoma malignum, carcinoma basocellulare illetve
spinocellulare) legkifejezettebben az I-II bőrtípusú (fehér, érzékeny bőr)
kaukázusiaknál jelentkezik (Young és mtsai. 1996, Katiyar és mtsai. 2000,
Kraemer 1997, Farkas és mtsai. 2002).
Az életkor meghosszabbodása, valamint a természetes (napfény) és a
mesterséges (pl. szolárium) UV-expozíció növekedése által kiváltott
fénykárosodás világszerte a jó- és rosszindulatú bőrtumorok előfordulási
gyakoriságának progresszív emelkedéséhez vezetett A malignus bőrtumorok
incidenciája az utóbbi 30 évben hazánkban is drámaian emelkedett, s ez a
tendencia jelenleg is folytatódik. Míg az epidermális eredetű laphám
carcinoma és a basocelluláris carcinoma patogenézisében a krónikus (élet
során összeadódó) napfény expozíciónak, addig malignus melanoma

6
esetében inkább az ismétlődő, rövid expozíciós idejű, intenzív napégéseknek
lehet szerepe.
Fenti tényezők miatt a fényvédő készítmények iránti követelmények
megnőttek. A teljes spektrumú védelem (UVA, UVB) alkalmazása feltétlenül
indokolttá vált (Lim és mtsai. 2001). A jelenleg meglévő, és a fényvédők
használatát sok esetben akadályozó mellékhatások (fototoxikus, fotoallergiás
reakció, színező hatás, nem megfelelő konzisztencia, stabilitási problémák)
kiküszöbölése fontos feladat (Lim és mtsai. 2001). A kutatás az eddigiektől
merőben eltérő, új típusú fényvédő készítmények (pl.: a DNS-repair
kapacitást támogató T4V (T4N5) liposzómális endonukleáz, az IL-12, PARP-
gátlók) előállítása irányába folyik (Yarosh és mtsai. 2001, Schwartz és mtsai
2002, Farkas és mtsai. 2002). A cél a megelőzés és a hatékony védelem
által a bőrtumor-incidencia növekedésének megállítása (Lim és mtsai 2001).
A rendelkezésünkre álló új molekulákkal munkánk célja elsősorban az akut
és krónikus UV-fény okozta károsodás kialakulásának megelőzésében
szerepet játszó szabályzó mechanizmusok vizsgálata, valamint a bőr
védelmét elősegítő illetve károsító folyamatok farmakológiai
befolyásolhatóságának tanulmányozása volt.
A bonyolult védelmi funkció érvényesüléséhez az alapvető celluláris
folyamatokat (proliferáció, differenciálódás, túlélés, apoptózis) szabályozó,
számos jelátviteli rendszer korrekt működése szükséges. Jelen munkában a
nagyszámú jelátviteli folyamatból, melyek az UV-fény indukálta
bőrelváltozások kialakulásában részt vesznek a poli(ADP-ribóz) polimeráz
(PARP) enzim aktivitás és annak szabályozásával összefüggő
mechanizmusokat kívántam vizsgálni.

7
A poli(ADP-ribóz) polimeráz az eukarióta sejtekben (kivéve az
élesztősejteket) nagy mennyiségben (körülbelül 106 molekula/sejt) jelenlevő
nukleáris, 116 kD molekulasúlyú enzim, amely meglehetősen hosszú
féléletidejű fehérje. A PARP három fő doménból áll: az N-terminális DNS-
kötő domén (DBD), az automodifikációs domén (AMD) és a C-terminális
katalitikus domén. A DBD-n belül két cink-ujj felelős a DNS kötésért és
néhány fehérje-fehérje kölcsönhatásért. A DBD ezen kívül tartalmaz egy
nukleáris lokalizációs szignál (NLS) régiót, amelyen belül a kaszpáz hasító
hely (DEVD) is található. Az automodifikációs domén tartalmaz egy BRCT
(breast cancer susceptibility protein C terminus) szakaszt, amely számos
DNS repair és sejtciklus fehérjében közös. A PARP a különböző fehérje-
fehérje kölcsönhatásokban a BRCT szakaszon keresztül vesz részt. Az
enzim C-terminális részén található aktív hely, az eukariótákban rendkívül
konzervált, 50 aminosavból álló szekvencia úgy is ismert, mint „PARP
névjegy” (Virág és mtsa. 2002).
Amennyiben a sejtet különböző behatások (pl.: alkiláló szerek, ionizáló
sugárzás, UV-sugárzás vagy szabadgyökök, stb) károsítják, a PARP gyorsan
odakötődik a DNS lánctörések helyére, automodifikáción megy keresztül,
amely a célfehérjén (leggyakrabban magán az enzimen) hosszú, elágazó
láncú poli(ADP-ribóz) polimerek képződéséhez vezet. A folyamathoz NAD+
használódik fel szubsztrátként, amely végeredményben az intracelluláris
NAD+ deplécióját okozza (Lindahl és mtsai. 1995, Burkle 2001, Virág és
mtsa. 2002). A képződő negatív töltésű PARP azután leválik a DNS-
végekről, elősegítve a DNS-repair folyamatát (Lindahl és mtsai. 1995). A
poli(ADP-ribóz) in vivo rövid életű, mert a poli(ADP-ribóz) glükohidroláz

8
(PARG) gyorsan (2-5 perccel a polimer képződése után) lebontja (Bernardi
és mtsai. 1997). A PARP-nak ez az oda-vissza kötődése a DNS lánctörés
helyén annak a hipotézisnek a megszületéséhez vezetett, hogy a PARP és a
poli-ADP-riboziláció különböző, kromatinhoz kötött működésekben is
szerepet játszik (de Murcia és mtsai. 1997, Kanai és mtsai. 2000). A PARP
automodifikációja megváltoztatja az enzim affinitását a DNS lánctörésekhez,
valamint azt a képességét, hogy más, a DNS-repair-ben részt vevő
fehérjékkel versenyezzen (Lindahl és mtsai. 1995, D’Amours és mtsai. 1999).
A PARP különböző biológiai folyamatokban játszott lehetséges funkciójába
azok az in vitro kísérletek nyújtottak sokoldalú betekintést, amelyekben NAD+
analógokat alkalmaztak a poli-ADP-riboziláció gátlására (például:
nikotinamid, benzamid és származékai). A tanulmányok szerint a kémiai
gátlószerek hatékonyan képesek megakadályozni a NAD+ deplécióját, és
túlérzékennyé tenni a sejteket a DNS-károsító szerekre, valószínűleg a
károsodott DNS kijavítását késleltetve (D’Amours és mtsai. 1999, Shall
1995). Ezek az anyagok valóban gátolják a PARP aktivitást, ugyanakkor a
normális sejtmetabolizmust is befolyásolják más NAD+-dependens
folyamatokat érintve (Milam és mtsai. 1984).
Annak ellenére, hogy az elmúlt évtizedben számos szervvel és szövettel
(szív, vázizom, központi idegrendszer, vese, bél, izület, szem, stb.)
kapcsolatos vizsgálatok történtek a PARP szerepére vonatkozóan(Zhang
2002, Virág és mtsa. 2002, Szabados és mtsai. 2000.). A bőr az ilyen irányú
kutatásokból gyakorlatilag kimaradt. A PARP aktivációnak a bőrelváltozások
patomechanizmusában betöltött jelentőségével kapcsolatban csak nagyon
szórványos adatok találhatók az irodalomban, a PARP aktiváció bőrben

9
történő szabályozásával (PARP-gátlók hatása) kapcsolatos vizsgálatokra
vonatkozó keresésünk pedig eredménytelenül zárult. A PARP bőrben
betöltött szerepére vonatkozó eddigi ismereteinket a következőkben tudjuk
összefoglalni. Keratinocitákon végzett vizsgálatokkal mutatták ki, hogy a
peroxinitrit és a hidrogén-peroxid a HaCaT sejtekben PARP aktivációt idéz
elő, amely hozzájárul a peroxinitrit által okozott citotoxicitáshoz (Szabó C. és
mtsai. 1998), ill. Hinshaw és mtsai. a kén-mustárral (DNS károsító szer)
kiváltott fokozott PARP-szintézisről számoltak be (Hinshaw és mtsai. 1999).
Később, in vivo oxazolonnal előidézett kontakt dermatitisz kapcsán (Szabó E.
és mtsai. 2001), ill. egérbőr mustárolajjal történő kezelése során (Virág és
mtsai. 2002) észleltek PARP aktivációt. Virág és mtsai. az epidermisz bazális
rétegében, immunhisztokémiai vizsgálattal, monoklonális poli-ADP-ribóz
ellenes antitestet használva észleltek kifejezett keratinocita festődést.
Ugyanezen kísérlet során, a mustárolajjal kezelt bőrben, poliklonális
antinitrotirozin antitestet használva, az epidermális és dermális sejtekben
jelentős nitrotirozin képződést figyeltek meg, mely alapján feltételezték a
peroxinitrit jelenlétét Vizsgálataik alapján arra következtettek, hogy a PARP
aktiváció egyik mechanizmusa a PARP direkt aktivációja lehet az alkilált DNS
által, valamint az intracutan peroxinitrit képződés is hozzájárulhat az enzim
aktivációjához (Virág és mtsai. 2002).
Annak ellenére, hogy az UV-fény DNS-károsító hatása (Hemminki és mtsai.
2001, Freeman és mtsai. 1989), valamint a PARP DNS-lánctörésre történő
aktivációja (Benjamin és mtsai. 1980, Satoh és mtsa. 1992, Lindahl és mtsai.
1995) jól ismert, az UV-besugárzást követően a PARP bőrben betöltött
szerepére vonatkozóan ismereteink hiányosak. Ezért érdemesnek látszott,

10
hogy kutatásom egyik legfontosabb céljává tenni az UV-expozíciót követő
PARP-aktiválás kimutatását, valamint azt, hogy direkt bizonyítékokat
szolgáltassak a PARP-gátlás vagy reguláció protektív hatására.

11
Célkitűzések
Munkánk célja a bőr különböző környezeti stresszhatásokkal szembeni,
illetve a már kialakult kóros állapotok helyreállításában részt vevő poli(ADP-
ribóz) polimeráz (PARP) enzim szerepének és farmakológiai
befolyásolhatóságának tanulmányozása volt.
A környezeti stresszhatások közül érdeklődésünk középpontjába a fény
ultraibolya (UV) tartománya került. Ismert, hogy az élet során elszenvedett
fénykárosodás kumulálódik. A civilizált világban az életkor
meghosszabbodásával a természetes (napfény) és mesterséges (pl.
szolárium) UV-expozíció által kiváltott fénykárosodás mértéke jelentősen nő.
A fentiek következtében, az UV-fény okozta jó- és rosszindulatú (melanoma,
és nem-melanoma típusú) bőrtumorok előfordulási gyakoriságának
progresszív növekedése már jelenleg is világszerte megoldandó problémát
jelent. Az új molekulák (immunstimulánsok, DNS-protektív anyagok, stb.)
előállításával és alkalmazásával várható a hatékonyabb prevenciós
lehetőségek számának növekedése illetve a kóros állapotok jelenleginél
korszerűbb diagnosztikus és terápiás megoldása.
A rendelkezésünkre álló új molekulákkal célunk elsősorban az akut és
krónikus UV-fény károsodás kialakulásának megelőzésében szerepet játszó
szabályzó mechanizmusok vizsgálata, valamint a bőr védelmét elősegítő,
protektív, illetve károsító folyamatok farmakológiai befolyásolhatóságának
tanulmányozása volt.
A megvalósításhoz egyszerű, ugyanakkor jól reprodukálható modelleket
próbáltunk használni, amelyek mind sejtszinten, mind pedig a bőr, mint szerv

12
szintjén biztosítják a fiziológiás és patológiás történések vizsgálhatóságát, és
magukban foglalják a gyakorló klinikus számára az alkalmazás potenciális
lehetőségét.
A fentiek alapján tanulmányozni kívántuk:
1. PARP-regulátor/inhibitor általi szabályzás vizsgálata a bőr akut és Krónikus UV-irradiáció elleni védelmében
• A poli-ADP-riboziláció jelentősége a bőr UV-fény indukálta károsodásában
• PARP aktivitás in vitro és in vivo vizsgálata
• PARP-regulátor/inhibítor antieritematogén, DNS-protektív, immunoprotektív hatásának vizsgálata
2. PARP-regulátor/inhibítor szerepe az UVA- és UVB-fény indukálta
karcinogenezis elleni védelemben

13
I. Anyag és módszer
A vizsgálatok végzéséhez PTE ÁOK Régionális Etikai Bizottsága
engedélyével, valamint beteg információs és beleegyező nyilatkozattal
rendelkeztünk.
I./1. Állatkísérletek
A kísérleteket az „European Community guiding principles for the care and
use of laboratory animals” szabályai szerint, állatetikai engedély birtokában
végeztük. Egy-egy kísérleti sorozatban az állatok életkor és súly szerint
lényeges eltérést nem mutattak.
I./1.1. Akut és krónikus fénykárosodás vizsgálata „hairless”
egérmodellen
A lokálisan alkalmazott PARP-regulátor (különböző koncentrációban BGP-
15M hatóanyagot tartalmazó krém) dermatológiai hatásának vizsgálatához
(akut és krónikus UV-fény expozíciós kísérletek) hairless CRL:hr/hr BR Hr1,
(Charles River Ltd., Németország) szőrtelen egereket használtunk (3. Ábra).
Az általunk használt egerek lényeges tulajdonsága, hogy
immunkompetensek, spontán malignus tumor képződésre nem hajlamosak,
és jóindulatú papillómák is csak nagyon idős korban jelentkeznek az
állatokon (Sundberg 1994). Az egerek hátán az epidermisz vastagsága
megközelítőleg 30 µm (két sejtsoros), az epidermális sejtek „turnover” ideje
1-2 hét. A szőrtelensége miatt kifejezett UV-abszorpciót mutató epidermisz
alkalmassá teszi ezt az egértípust az UV-fény hatásának (pl. az akut, vagy a
krónikus fénykárosodás, fotokarcinogenezis, stb.) modellezésére (De Gruijl

14
és mtsa. 1995). Kísérleteinkhez 5-8 hetes, 18-26 g súlyú, nőstény egereket
használtunk. Az állatokat kórokozó mentes környezetben, 12 órás periódikus
(sötét/világos) megvilágítás alkalmazásával, 22-25°C-on, 50-70%
páratartalom mellett, általánosan használt egértápon és csapvíz itatással
tartottuk. Az állatok egyéni identifikálása a fülön történő jelöléssel, a
ketreceké pedig kártyával történt.
I./1.2. Lokális kezeléshez alkalmazott készítmények
BGP-15M (PARP-regulátor) az O-(2-hidroxi-3-piperidino-propil) piridin-3-
karboxil sav amidoxim monohidroklorid sója. A molekula, amelyet lokális
alkalmazásra állítottak elő, az orális, szisztémás alkalmazásra szánt
dihidroklorid sóval szemben a BGP-15M jelölést kapta (előállító: N-GENE
Res. Lab., New York). A lokális készítményre vonatkozó adatokat az előállító
bocsátotta rendelkezésünkre. Vizes oldata enyhén savas kémhatású (pH
5,6). Por formában fehér, nem higroszkópos kristályos anyag. Vízben
oldódik. A BGP-15M két enantiomert (+ és -) tartalmaz, a molekula chilaris
centruma mentén. A BGP-15M jelölés egyben a racem keverékre utal. A
kísérletekhez előállított krém 5-20% koncentrációban tartalmazott BGP-15M-
t. Az elvégzett stabilitási (fotostabilitási) vizsgálatok alapján a BGP-15M a
krémben stabilnak bizonyult. A preklinikai biztonsági vizsgálatok (toxikológia,
mutagenitás, farmakológia, farmakokinetika) dominálóan orális adást
követően, bizonyos esetekben dermális kezeléshez társulva (pl. akut
dermális toxicitás egérben, szőrtelen egérben, akut dermális irritáció nyúlban,
szubakut (28 napos) dermális toxicitás két hetes követéssel patkányban)

15
történt. Az ismertetett adatok alapján a krém formula a biztonsági vizsgálatok
során lokális (eritéma, ödéma), és szisztémás (letalitás, belszervi
makroszkópos és mikroszkópos eltérések, testsúly csökkenés) kóros
elváltozásokat nem okozott.
BGP-15M lokális készítmény vivőanyaga olaj a vízben (O/V) típusú, kis
zsírtartalmú kenőcs (desztillált víz, glicerin, sztearin, cetil-sztearil-alkohol,
fehér vazelin, folyékony paraffin, lecitin, polietilénglikol-cetilsztearil éter
(etoxiszáma 6) polietilénglikol-cetilsztearil éter (etoxiszáma 25) (Ph.Hg.VII.
szerinti magyar névvel feltüntetve).
Az „AS” jelzésű készítményt kísérleteinkben kontrollként használtuk. A
kereskedelmi forgalomban kapható, Mexoryl® SX komponense által az UVA
tartományban is hatékony, fotostabil fényszűrőt tartalmazó, SPF (Sun
Protection Factor) 30 és anti UVA/UVB jelzésű (széles spektrumú, a fény
UVB és UVA tartományát lefedő), korszerű fényvédőt választottuk ki
referencia készítményként, és jelöltük ”AS”-sel. A kontroll készítmény
összetétele a következő: desztillált víz, octocrilen, ciclopentasiloxan, titán-
dioxid, glicerin, propilénglikol, izohexadekán, sztearin, butil-
metoxidibenzoilmetán, oktil-palmitát, sztearil-heptanoát, PVP/eikozén
kopolimer, nátrium-cetil-foszfát, jojoba, tokoferol-acetát, metilcellulóz
propilénglikol éter, fenoxietilalkohol, sztearil-kaprilát, polietilénglikol
sztearinsav észtere (100 etilénoxid csop.), etil-p-hidroxi benzoát,
trietanolamin, dimethikonol, szilikon olaj, propil-p-hidroxi benzoát,
tereftalidén-dikámfor-szulfon sav, akrilátok/C10-30 alkil-akrilát krosszpolimer,
dinátrium-EDTA, butirospermum parkii, cetil-alkohol, metil-p-hidroxi benzoát,

16
butil-p-hidroxi benzoát, alumínium-hidroxid, glicerilmonosztearát, parfüm
(C9794/1). (Ph.Hg.VII. szerinti magyar névvel feltüntetve).
A szőrtelen egerek törzsén kialakított egy, vagy több tesztterületet BGP-15M
tartalmú krémmel (hatóanyag tartalom: 5%, 10%, 15%, 20% BGP-15M),
illetve hatóanyagmentes vivőanyagával (vehikulum), vagy az ”AS”-sel jelölt
készítménnyel, 2 mg/ testfelszín cm2 krém egyenletes felvitelével kezeltük.
Az UV-fény akut károsító hatásának vizsgálata során az állatokat az UV-fény
expozíció előtt 15 perccel, egyszeri kezelésben részesítettük. Az UV-
besugárzás krónikus hatásának tanulmányozására végzett kísérletekben az
egerek tesztterületeit heti 5 alkalommal, egymást követő napokon, az UV-
irradiáció előtt 15 perccel, azonos időben és módon kezeltük.
I./1.3. Szőrtelen egerek UV-fény kezelése
Az UV-irradiáció kivitelezéséhez mesterséges (UVB- és UVA-tartományban
sugárzó) és természetes fényforrást (Nap) használtunk. Az UVB-
expozícióhoz a széles spektrumú UV21 Philips lámpát (13 cső, csőtípus: F
85/100 W, emissziós spektrum: 285-350 nm, peak: 310-315 nm, Waldmann
Medizintechnik, Villingen-Schwenningen) alkalmaztunk UVB-sugárzóként. Az
UVA-irradiációt Waldmann UV 8001K széles spektrumú UVA-készülékkel
(csőtípus: WF 85/100 W, emissziós spektrum: 320-400 nm, peak: 365 nm,
Waldmann Medizintechnik, Németország) végeztük.
A sugárzás méréséhez IL 700 spektroradiométert (koszinusz korrekciós SEE
detektorral) használtunk.

17
A természetes fénnyel, a napfénnyel végzett UV-expozíció paramétereinek
meghatározása a beeső teljes fotonszám mérése alapján Brewer MK3
Spektrofotometerrel és Brewer szoftverrel (Sci-Tec, USA) történt. (A mérések
végzéséért ezúttal szeretnénk köszönetet mondani Tóth Zoltánnak,
Országos Meterológiai Szolgálat Légkörfizikai Intézet, Budapest.) Az aktuális
maximális irradianciát és a biológiailag effektív intenzitásokat a fény UVB-
tartományának 287-320 nm hullámhosszúságú, valamint az UVA-tartomány
321-363 nm hullámhosszúságú részére adtuk meg. Hazánkban, a fény UVB-
tartományában az irradiancia maximális értéke: 1,2-1,3 J/cm2/h, az UVA-
tartományában pedig: 8,5-8,6 J/cm2/h.
A vizsgálat időpontjában a fény UVB-tartományában (287-320 nm-re
integrálva) az irradiancia maximális értéke: 1,08 J/cm2/h volt. A vizsgálat
időpontjában a fény UVA-tartományában (321-363 nm-re integrálva) az
irradiancia maximális értéke: 8,3 J/cm2/h-nak bizonyult. Ez az érték 321-380
nm tartományban 13 J/cm2/h.
A biológiailag effektív intenzitások: UVB (287-320 nm): 0,07 J/cm2, UVA
(321-363 nm): 0,015 J/cm2. A számítások végzéséért szeretnénk köszönetet
mondani Erostyák Jánosnak (PTE Kísérleti Fizikai Tanszék, Pécs).
Az állatokat a besugárzáshoz a mesterséges UV-fényforrástól 30 cm-re
helyeztük el. A bőrfelszínen mért sugárzás intenzitás alapján határoztuk meg
az egyes kísérlet során a sugárzó által leadandó dózist. (A sugárzó és a
spektroradiométer kalibrálása, az előírás szerinti rendszeres időszakonként,
a szakszerviz által történt.) Minden egyes kísérlet előtt meghatároztuk a
minimális eritéma dózist (MED), amelyet mJ/cm2, illetve J/cm2-ben adtunk
meg.

18
A szoláris UV-fény kezeléshez az állatokat speciális egérlapon rögzítettük,
amely biztosította a tesztterület standard kezelését és a környező bőr
biztonságos takarásának megoldását. Az UVA-besugárzás, valamint a
szoláris expozíció esetén az állatokat a hőhatástól alulról, átfolyásos
vízhűtéssel működő rendszerrel védtük.
Az UVA-fénnyel, a krónikus fénykárosodás megítélésére (pl.:
fotokarcinogenezis tanulmányozásához) végzett kísérletek során, a
besugárzási idő csökkentése céljából „forszírozott UVA”-kezelést, vagyis
pszorálen (8-metoxi-psoralen (8-MOP), Geroxalen® oldattal fényérzékenyített
tesztterületeket sugaraztunk be (Lowe és mtsai. 1987). A 8-MOP oldattal
történt ecsetelés és az UVA-irradiáció között 30 perc telt el.
A napfény expozíciót felhőmentes nyári napon 12-13 óra között végeztük. Az
előkísérletek során határoztuk meg mindazon feltételeket, amelyek az állatok
számára az általunk megítélt legkíméletesebb eljárást biztosították.
I./1.4. A minimális eritéma dózis (MED) meghatározása szőrtelen
egereken
A minimális eritéma dózis (MED) alatt azt az UVB-fény által közvetített
energiamennyiséget értjük, amely a bőr minimális eritémás reakciójának
előidézéséhez szükséges (Lowe 1990). A MED meghatározásához az
egerek fedetlen bőrén kialakított 6 db, egyenként 0,25 cm2 területű
tesztterületet emelkedő dózisú (0,12 J/cm2-től induló) UVB-fénnyel
sugaraztuk be. A MED leolvasása 24 órával az UVB-besugárzást követően,

19
ugyanazon személy által történt (az interperszonális eltérések elkerülése
céljából).
I./1.5. UVB-expozíció akut hatásának vizsgálata PARP-regulátorral
kezelt egérbőrön
Az egerek hátán (thoracalis régió), a reakciómentes, ép bőrön, 2 x 1,44 cm2-
es nagyságú, egymás alatt elhelyezkedő (egymástól legalább 0,8 cm
távolságra levő), tesztterületet alakítottunk ki, és használtunk a kísérletekben
(antieritematogén koncentráció meghatározásra, fényvédő hatás
összehasonlító vizsgálatára UVB-tartományban, stb.) - az állatok számát az
egyes kísérleteknél tüntettük fel. Az egerek kezeléséhez a PARP-regulátor
BGP-15M tartalmú krémet (hatóanyagtartalom: 5%, 10%, 15% és 20%)
alkalmaztuk, illetve vivőanyagát használtuk. Az egerek krémmel történő
előkezelését 1.2. pontban leírtak szerint végeztük. Ezt követően az állatokat
egyszeri, eritémát okozó UVB-irradiációban részesítettük. Az alkalmazott
MED dózisának meghatározása 24 órával a kísérlet előtt 3 állaton történt. A
környező bőrfelszíneket az UV-behatástól takarással védtük. A negatív
kontroll csoportot a krémmel nem kezelt, UV-expozícióban nem részesült
állatok képezték. Az irritatív hatás vizsgálatára 20% BGP-15M tartalmú
krémmel kezelt, UV-expozícióban nem részesült állatok csoportja szolgált.
Az UVB-irradiáció után közvetlenül, a BGP-15M krémet, illetve vivőanyagát
az állatok bőréről eltávolítottuk.
A vizsgálatokat az egerek I.-VI. csoportjával (jelölés: /1= felső, /2= alsó
tesztterület) végeztük, melyek a következő kezelésben részesültek: I.: I./1.:

20
vivőanyag + UVB, I./2: 5% BGP-15M + UVB; II.: II./1.: vivőanyag + UVB, II./2:
10% BGP-15M + UVB; III.: III./1.: vivőanyag + UVB, III./2: 15% BGP-15M +
UVB; IV.: IV./1.: vivőanyag + UVB, IV./2: 20% BGP-15M + UVB; V.: 20%
BGP-15M; VI.: kezeletlen, negatív kontroll. A tesztterületek elrendezésével
biztosítani kívántuk az esetleges individuális különbségekből adódó eltérések
minimálisra csökkentését. A lokálisan alkalmazott PARP-regulátor hatásának
vizsgálatához a bőrmintákat az UVB-besugárzás után közvetlenül, vagy 24
óra elteltével excízióval nyertük. Az állatokat cervicalis dislocatioval
pusztítottuk el.
I./1.6. Egérbőr kezelése a PARP-regulátor és ismert fényvédő faktorú
készítmény antieritematogén hatásának összehasonlító
vizsgálatához
A BGP-15M antieritematogén hatását a forgalomban kapható, széles
spektrumú (UVA és UVB tartományban ható), általunk ”AS”-sel jelölt,
fényvédő készítménnyel hasonlítottuk össze.
Az egerek háti felszínén kialakított 1,44 cm2-es tesztterületek közül a
proximálisat „AS” jelzésű krémmel, a disztálisat pedig 10%, 15%, vagy 20%
BGP-15M tartalmú krémmel kezeltük az egymáshoz viszonyított
antieritematogén hatás megítéléséhez. Az egyes készítmények, kísérleti
körülményeink között, az eritéma kialakulására kifejtett gátló hatásának
meghatározásához a proximális tesztterületet UVB-irradiációban
részesítettük (pozitív kontroll), a disztális tesztterületet pedig „AS” jelzésű,
vagy 15% BGP-15M krémmel kezeltük. A tesztterületeket egyszeri, 2 MED
dózissal UVB-irradiációban részesítettük, a környező bőr takarással
biztosított védelme mellett. A MED meghatározása (I./1.4. pont) a kísérlet

21
előtt 24 órával, 3 állaton történt. Negatív kontrollként kezeletlen, UVB-
expozícióban nem részesült egerek csoportja szolgált. A krémet az állatok
bőréről az UVB-irradiáció után közvetlenül eltávolítottuk.
A kezelés alapján a vizsgálatokhoz az állatok következő csoportjait
használtuk: I.: I/1.:AS + UVB, I./2.:10% BGP-15M+UVB; II.: II./1.:AS + UVB,
II./2.: 15% BGP-15M+UVB; III.: III/1.: AS+UVB, III./2.: 20% BGP-15M+UVB;
IV.: IV./1.: UVB, IV./2.: AS+UVB; V.: V./1.: UVB, V./2.: 15% BGP-15M+UVB;
VI.: kezeletlen, negatív kontroll csoport. A tesztterületek elrendezésével
biztosítani kívántuk az esetleges individuális reakciókülönbségből adódó
eltérések minimálisra csökkentését. Az antieritematogén hatást az UVB-
besugárzás után 24 és 48 órával vizsgáltuk.
I./1.7. Szőrtelen egerek kezelése PARP-regulátorral szoláris-expozíció akut
hatásának vizsgálatához
A PARP-regulátor, a napfény akut károsító hatásával szembeni fotoprotektív
tulajdonságának vizsgálatához a szőrtelen egerek csoportjait 10%, 15%,
vagy 20% BGP-15M tartalmú krémmel, valamint vivőanyagával és az
ismertettek alapján kezeltük. Az állatok felhőtlen nyári napon, délben (12-13
óra), speciálisan erre a célra kialakított (hőhatást csökkentő rendszer)
körülmények között (I./1.3. pont) részesültek a tesztterületeken 1 órán
keresztül természetes napfény expozícióban. Az egerek a napoztatási idő
alatt mért irradiancia alapján összesen 8,3 J/cm2/h UVA és 1,08 J/cm2/h UVB
dózisú sugárzást kaptak. Az állatokat a tesztterületek kivételével az UV-
fénytől védtük. Negatív kontrollként kezeletlen, az UV-fénytől a teljestest

22
takarásával védett (sham-irradiált) egerek szolgáltak. A BGP-15M krémet,
illetve vivőanyagát az állatok bőréről az UV-irradiáció után közvetlenül
eltávolítottuk.
A kezelés alapján az egerek következő csoportjait használtuk: I.: I./1: 10%
BGP-15M + UV, I./2.: vivőanyag + UV; II.:II/1.: 15% BGP-15M + UV, II./2.:
vivőanyag + UV; III.: III./1: 20% BGP-15M + UV, III./2.: vivőanyag + UV; IV.:
15% BGP-15M; V.: kezeletlen, negatív kontroll. Az állatok egy részét
közvetlenül az UV-expozíció után cervicalis dislocatioval elpusztítottuk, a
tesztterületekből nyert bőrmintákat további feldolgozásig fagyasztva (-70°C)
tároltuk. A fennmaradó egerek bőrén az UV-irradiáció után 24 órával
végeztünk vizsgálatokat.
I./1.8. Szőrtelen egerek kezelése PARP-regulátorral UVB-expozíció
krónikus hatásának vizsgálatához
A PARP-regulátor, BGP-15M protektív hatását a krónikus UVB-irradiáció
bőrelváltozásokat előidéző tulajdonságával szemben, azonos életkorú (6
hetes), 18-21g súlyú, nőstény szőrtelen egereken vizsgáltuk. Az egerek
hátán (n=20 állat/csoport), a középvonalban 12 x 18 mm-es tesztterületet
alakítottunk ki, melyet 15% BGP-15M tartalmú krémmel, vagy annak
vivőanyagával (n=20 állat) előkezeltünk. Az állatok az előkezelés után napi
egyszeri, 1 MED UVB-besugárzást kaptak, heti öt egymást követő napon
keresztül. A MED dózisának meghatározása 24 órával a kísérlet elkezdése
előtt 5 egéren történt. Az állatok tesztterületen kívül eső bőrét takarással
védtük az UV-sugárzással szemben. Tíz egér nem részesült kezelésben. Ez

23
a csoport volt a negatív kontroll. A kontroll csoportba tartozó egerek
bőrállapota (makroszkópos és mikroszkópos) szolgált a „photoaging” (fény
általi öregedés) mentes kronológiai öregedés („aging”) követésére. Öt állatot
a tesztterületnek megfelelően 15% BGP-15M tartalmú krémmel kezeltünk
(UVB-fénytől védve) az irritatív vagy allergiás reakciók vizsgálata céljából. Az
UVB-irradiáció után közvetlenül a BGP-15M krémet, ill. vivőanyagát az
állatok bőréről eltávolítottuk.
A kialakított vizsgálati csoportok a következők: I.: 15% BGP-15M + UVB; II.:
vivőanyag + UVB; III.: 15% BGP-15M; IV.: kezeletlen, negatív kontroll. Az
állatokat 32 héten át kezeltük és követtük. A 6. héttől kezdve az egyes
csoportokból cervicalis dislocatioval elpusztított egerek bőrmintáin
hisztológiai, immunhisztokémiai és elektronmikroszkópos vizsgálatokat
végeztünk.
I./1.9. Szőrtelen egerek kezelése PARP-regulátorral UVA-expozíció
krónikus hatásának vizsgálatához
A PARP-regulátor BGP-15M potenciális fotoprotektív tulajdonságát a
krónikus UVA-irradiáció által a szőrtelen egerek bőrében előidézett
változásokra („photoaging” és fotokarcinogenezis) „forszírozott-UVA”
kezelésben részesített (1.3. pont), azonos korú (7 hetes), nőstény egereken
vizsgáltuk. Az egerek hátán 2,25 cm2 (1,5x1,5 cm) tesztterületek (n=2)
kerültek kialakításra. Az állatokat 8-metoxipsoralen (8-MOP, Geroxalen®,
Gerot Pharmazeutika, Wien) 0,15%-os oldatának használatával
fényérzékenyítettük. Az egerek előkészítése a kísérlethez a

24
következőképpen történt: 5 µl/cm2 8-MOP oldattal a tesztterületeket
bekentük, az egereket sötétbe helyeztük, majd 30 perc múlva, a 15% BGP-
15M tartalmú krémmel, vagy annak vivőanyagával kezeltük és 15 perc
elteltével UVA-irradiációban részesítettük. Az állatok UVA- és a „forszírozott-
UVA” kezeléssel (8-MOP + UVA) szembeni érzékenységének vizsgálatára
külön állatcsoportot állítottunk be (krémkezelést nem kaptak). További, 15%
BGP-15M tartalmú krémmel kezelt (UVA-expozíciótól védett) egerek
szolgáltak az esetlegesen fellépő irritatív, vagy allergiás reakció vizsgálatára.
A fentiek mellett kezeletlen kontroll csoportot használtunk. Az egerek heti 5
alkalommal, napi 0,5 J/cm2 UVA-besugárzást kaptak 26 héten keresztül. Az
állatok tesztterületen kívül eső bőrét takarással védtük. Az UVA-irradiáció
után közvetlenül a kezelésre használt krémeket (BGP-15M tartalmú, illetve
vivőanyag) az állatok bőréről eltávolítottuk.
A kezelés szerint az állatokat a következő csoportokba osztottuk: I.:I./1.: 8-
MOP + UVA, I./2.: UVA; II.: II./1.: 8-MOP + vivőanyag + UVA; II./2.: 8-MOP +
15% BGP-15M krém + UVA ; III.: III./1.: 8-MOP + vivőanyag, III./2.: 15%
BGP-15M krém; IV.: kezeletlen kontoll (n=6 egér/csoport). A kontroll
csoportba tartozó egerek bőrállapota szolgált a kronológiai öregedés
követésére. A rendszer összeállítása lehetővé tette az egerek UVA- és
„forszírozott-UVA” érzékenységének (makro- és mikromorfólógiai)
megítélését, valamint az önkontrollos elhelyezés az esetleges individuális
különbségekből adódó eltérések minimálisra csökkentését. Az egereket 26
héten át kezeltük és követtük. A 26. hét végén cervicalis dislocatioval
elpusztított egerekből vett bőrmintákat használtuk a hisztológiai és
immunhisztokémiai vizsgálatokhoz.

25
I./2. Klinikai tünetek értékelése
I./2.1. Klinikai vizsgálatok akut és krónikus UV-expozícióban
A különböző kezelésben részesült egerek tesztterületeinek bőrállapotát
vizuálisan és dermatoszkóppal vizsgálva összehasonlítottuk a kezeletlen
kontroll állatokéval. Az akut UV-expozíció által kiváltott fénykárosodás klinikai
tüneteit (eritéma, ödema, vezikula, bulla, erózió) az UV-besugárzás után
közvetlenül és 24 órával azt követően, öt fokozatú skála (0-4 pont) szerint
értékeltük (I/2.2. pont) és tüntettük fel az egérkövetési lapokon.
A krónikus UVB- és UVA-besugárzás során a szőrtelen egerek tesztterületeit
hasonló módon, a kezelések előtt, hetente 5 alkalommal, valamint a
kísérletek befejezésekor vizsgáltuk. A klinikai tünetek (eritéma, pigmentáció,
erózió, exulceráció) intenzitását (I/2.2. pont) értékeltük. Az átmérő ≥1 mm
nagyságú tumorokat vettük figyelembe és tüntettük fel. Az észlelteket az
egérkövetési lapon regisztráltuk. Az egyes csoportok bőrstátuszában
bekövetkezett változásokat átlagosan 4 hetente, valamint a kísérlet végén
fotódokumentációval rögzítettük.
I./2.2. Klinikai tünetek értékelése pontszámmal
A klinikai tünetek meglétének és intenzitásának értékeléséhez öt fokozatú
skálarendszert használtunk. A tünetek súlyosságát 0-4 ponttal: 0 = nincs, 1 =
nagyon enyhe, 2 = enyhe, 3 = közepesen súlyos, 4 = súlyos fejeztük ki.

26
Eritémára vonatkoztatva: 0 = nincs, 1 = nagyon enyhe (alig észlelhető), 2 =
enyhe (jól látható), 3 = közepesen súlyos (vörös), 4 = súlyos (mélyvörös);
ödémára: 0 = nincs, 1 = nagyon enyhe (alig megítélhető), 2 = enyhe (érintett
terület előemelkedő széllel bír), 3 = közepesen súlyos (a bőr szintje fölé
emelkedik <1mm-rel), 4 = súlyos (>1mm-rel emelkedik a bőr szintje fölé).
A krónikus UVB- és UVA-besugárzás során a szőrtelen egerek
tesztterületein a klinikai tüneteket (eritéma, pigmentáció, erózió, exulceráció)
hasonló módon értékeltük. A krónikus UVA-besugárzás során hat fokozatú
skálát alkalmaztunk (0 = nincs - 5 = nagyon súlyos).
I./3. PARP enzim auto-ADP-ribozilációjának vizsgálata Western-blot
analízissel
A PARP auto-ADP-ribozilációját különböző bőrmintákban (kezeletlen, nem
besugárzott; 15% BGP-15M tartalmú krémmel kezelt; 15% BGP-15M
tartalmú krémmel, vagy vivőanyaggal előkezelt és UV-fénnyel besugárzott)
határoztuk meg (Szabados és mtsai. 2000). A bőrmintákat (20-22 mg/minta)
250 µl 50 mM Tris pufferben (pH 7,8) Ultra-Turrax készülékben
homogenizáltuk, majd 8 M karbamid tartalmú 2x Laemmli puffer 250 µl-ének
hozzáadását követően Potter-Elvehjem teflonos homogenizátorban
(Wheaton) tovább homogenizáltuk, majd centrifugáltuk (5 perc, 10.000 rpm).
A mintákban levő fehérjéket SDS-poliakrilamid gélelektroforézissel (8%-os
gél) szeparáltuk és nitrocellulóz membránra blottoltuk (Western-blot analízis)
(Szabados és mtsai. 2000). Az ADP-ribozilált fehérjék kimutatására anti-
ADP-ribóz monoklonális antitestet (Dr. Alexander Berkley, Heidelberg és Dr.
Masanao Miva, Tsukuba bocsátotta rendelkezésünkre) és anti-egér IgG

27
peroxidáz komplexet (Sigma, München) használtunk. Az antitest-antigén
komplex megjelenítését ECL (Enhanced Chemiluminescence) módszerrel
végeztük. A Western-bloton kapott jelek kvantitatív meghatározása „Image
Tool (Version 1.27) image processing” programmal (University of Texas,
Health Science Center, San Antonio) történt.
I./4. „Sunburn” sejtek kvantítatív meghatározása
A „sunburn” sejtek UV-fény expozicíó hatására a bőrben megfigyelhető
apoptótikus keratinociták. A „sunburn” keratinociták a környező sejtektől
különálló, lekerekedett alakú, zsugorodott, eozinofil citoplazmával,
vakuolumokkal rendelkező, kondenzált, piknotikus sejtmaggal bíró sejtek
(Ziegler és mtsai. 1994). A jellegzetes morfológiai elváltozásokkal rendelkező
„sunburn” sejteket felhasználják a fénykárosodás mértékének
megítélésében.
Az eozinofilen festődő, nukleusz nélküli, vagy piknotikus maggal rendelkező
„sunburn” sejtek számát a szövetminták 10%-os pufferolt formalinban fixált,
paraffinba ágyazott, He-festett, 4 µm-es metszetein, a bazálmembrán 250
µm-es szakasza (kalibrált ocular, Periplan GF 10x/20 Leitz, Nürnberg) feletti
interfollikuláris epidermiszben, 400x nagyítás mellett (Diaplan
fénymikroszkóp, Leitz, Nürnberg) határoztuk meg. Az állatonként 16
látótérben (80 látótér/csoport) vizsgált „sunburn” sejtek számának átlagát 1
mm hosszúságú epidermiszre vonatkoztatva (átlag ± SEM/mm epidermis)
adtuk meg.

28
I./5. Hisztológiai vizsgálatok
A vizsgálatokhoz a szövetmintákat narkózisban, vagy az állatok cervicalis
dislocatioval történő elpusztítása után nyertük. Az így kapott anyagot, vagy
azonnal feldolgoztuk, vagy a további feldolgozásig -70°C-on tároltuk.
Kimetszés után közvetlenül a bőrminták egy részét Tissue Freezing
Mediumot (Cambridge Instruments, Heidelberg) használva beágyazóként,
folyékony nitrogénben lefagyasztottuk. A blokkokat feldolgozásig -70°C-on
tároltuk. Az immunhisztokémiai vizsgálatokhoz 6 µm-es metszeteket
készítettünk (-24°C, Reichert-Jung Cryocut 1800 kriosztát, Leica, Nusslock).
Másrészt, a szövetmintákból formalinos fixálás (fedeles beágyazó
kazettában, (Bio-Optica, Milánó), 4,5% pufferolt formalin, pH 7,0, 9-24 h),
paraffinba való beágyazás (Citadel 1000 víztelenítő automata (Shandon,
Runcorn), 22 h, felszálló alkohol sor: 50% etilalkohol, 70% etilalkohol, 80%
etilalkohol, 96% etilalkohol, aceton, amilacetát + paraffinolaj (1:1), xylol
(Reanal, Budapest), paraffin (tissuewax, olvadáspont: 52-54°C, (Medite,
Burgdorf)) történt. Rotációs mikrotómmal (Leica RM 2135 rotációs mikrotóm,
Nusslock) 4 µm-es (felvétel és tárgylemezre olvasztás, 56°C, 30 min.)
metszeteket készítettünk. A festések előtt deparaffinálást és rehidrálást (xilol
2x10 min., abszolut alkohol, 96% alkohol 2x, 80% alkohol, 70% alkohol, 50%
alkohol), majd 3x csapvizes öblítést végeztünk (Mikkel 1994, Luna 1968).

29
I./5.1. Hematoxilin-eozin (He) festés
A metszeteket hematoxilin oldattal (Merck, Darmstadt) 8 percig festettük,
majd kékítést (2 min., csapvíz) követően a további festést eozin oldattal
(Merck, Darmstadt) szintén 8 percig végeztük. A víztelenítéshez felszálló
alkoholsort (70% etilalkohol 2x, 90% etilalkohol, 96% etilalkohol 2x, abszolut
alkohol 2x, abszolút alkohol+xilol 1:1 arányú keveréke, xilol 2x) használtunk.
A készítményt Pertex oldattal (Medite, Burgdorf) fedtük (Mikkel 1994, Luna
1968).
I./5.2. Fontana-Masson festés
A melanin kimutatására paraffinos metszeteken argentaffin reakciót
végeztünk Fontana-Masson módszere szerint (Krobock és mtsai. 1978).
Deparaffinálást és rehidrálást követően a metszeteket 2%-os AgNO3 oldattal
(Reanal, Budapest) inkubáltuk (szobahő, 2h), majd mosás után hidegen
telített (5%) nátriumtioszulfát oldattal (Reanal, Budapest) kezeltük (szobahő,
2 min.), folyó csapvizes mosás (30 min.) után háttérfestésként Mayer-féle
kármint (Merck, Darmstadt) alkalmaztunk (3 min.). Dehidrálást követően a
fedés Pertex oldattal (Medite, Burgdorf) történt.
I./5.3. Orcein-Giemsa festés
A rugalmas rostok vizsgálatára paraffinos metszeteken Orcein-Giemsa
festést végeztünk (Krobock és mtsai. 1978). Deparaffinálást és rehidrálást

30
követően a metszeteket orcein (Merck, Darmstadt) savanyított alkoholos
oldatával festettük (20 min.). Abszolut alkohollal való differenciálás, Giemsa
oldattal (Reanal, Budapest) történő háttérfestés (5 min.) és dehidrálás után, a
fedés Pertex oldattal (Medite, Burgdorf) történt.
I./6. Immunhisztológiai vizsgálatok
Az egérbőrből nyert, 6 µm-es kriosztátos metszeteket üveg tárgylemezen,
hideg acetonban (4°C, 10 min.) fixáltuk. A szövetminták minőségének
ellenőrzésére a metszeteken hematoxilin-eozin festést végeztünk. Az
immunhisztokémiai reakcióhoz anti-poli-ADP-ribóz poliklonális antitestet
(hígítás: 1:1000 TBS-ben, Biomol, Hamburg) használtunk, az inkubációt
szobahőmérsékleten, 60 percig végeztük, és a streptavidin-biotin-peroxidáz
módszert alkalmaztuk (H2O2/aminoetilkarbazol (AEC) előhívás, Immunotech
Universal kit (Immunotech, Marseilles)). Negatív kontrollként nem immunizált
egér hasűri folyadéka szolgált (Mikel 1994).
I./7. Elektronmikroszkópos vizsgálatok
Elektronmikroszkópos vizsgálathoz a frissen eltávolított bőrmintákat 12-24
órán át, 4°C-on Karnovszky fixálóban (1% paraformaldehid és 2,5%
glutáraldehid keveréke, Reanal R, SERVA) inkubáltuk, majd 1%-os
ozmiumtetroxidban utófixáltuk (90 min.). Dehidratáció után az anyagot
Araldit-be (Fluka) ágyaztuk. Az ultravékony metszeteket uranil acetáttal és
ólom citráttal (MERCK) kontrasztosítottuk, és JEOL 1200 EX II
transzmissziós elektromikroszkópon vizsgáltuk (Pease 1964).

31
II. Eredmények
1. PARP-regulátor szerepe a bőr UV-fény károsodása elleni
védelemben
2. PARP-regulátor hatásának vizsgálata UV-fény indukálta
karcinogenezisben
Napjainkban a bőr vonatkozásában a legjelentősebb környezeti DNS
károsító faktor a napfény UV tartománya. Egyszeri, magas dózisú napfény-
expozíció alkalmazásával vizsgáltuk a PARP-regulátor (5-20% BGP-15M
hatóanyag tartalmú krém) potenciális DNS-protektív hatását
immunkompetens hairless (szőrtelen) egérmodellen. Az UV-sugárzás által az
egérbőrben indukált akut DNS-károsodás mértékét a bőrben képződött
egyes láncú DNS-törések mennyiségének meghatározásával, továbbá az
UV-irradiáció által okozott DNS-károsodás markereként nyilvántartott
epidermális „sunburn” sejtek (apoptótikus keratinociták) képződésének
kvantitatív, valamint ultrastruktúrális vizsgálatával jellemeztük.
II./1.1. PARP-regulátor hatása az UV-sugárzás indukálta egyes láncú
DNS törések képződésére a bőrben
Az egyes láncú DNS törések meghatározását DNS-lánc szétcsavarodásán
alapuló fluoreszcenciás módszerrel végeztük. Mint ismert, a DNS a törések
környezetében szétcsavarodik és csak a sérülés mentes régiókban marad
meg kettős láncú formában. Ugyanakkor a kettős láncú DNS-hez kötődő

32
ethidium-bromid fluoreszcenciája sokkal magasabb, mint hasonló
körülmények között az egyes láncúé.
Az egyszeri, magas dózisú, természetes UV-expozíció (napfény) akut bőr
DNS-károsító hatását szőrtelen egérmodellen vizsgáltuk.
Rendszerünkben a napfény expozícióban nem részesült, kezeletlen egerek,
valamint a 15% BGP-15M tartalmú krémmel kezelt csoport bőrmintáiban a
DNS nagy része (átlagosan 80%) károsodást nem szenvedett, kettős láncú
DNS-nek bizonyult. Ugyanakkor az intenzív napfény expozíciónak (egy óra
alatt 8,3 J/cm2 UVA és 1,08 J/cm2 UVB) kitett egérbőrben a vivőanyaggal
kezelt mintákban az egyes láncú DNS törések mennyiségének jelentős
megnövekedése következtében, a nem károsodott DNS aránya 30% alá
csökkent (1. Táblázat). A napfény behatás előtt PARP-regulátorral (15%
BGP-15M tartalmú krém) végzett lokális előkezelés az intenzív UV-sugárzás
által indukált nagyfokú DNS-károsodás mértékét szignifikánsan (p<0,01)
csökkentette, a mintákban (n=5) az ép DNS aránya meghaladta az 50%-ot
(4. Ábra).
II./1.2. PARP-regulátor hatása az akut UVB-sugárzás indukálta
„sunburn” sejtek képződésére
Az apoptótikus sejthalál egyik prototípusának tartott „sunburn” sejtképződés
kvantitatív meghatározásával vizsgáltuk egérbőrben, az elszenvedett UV-
fény károsodás mértékét. Rendszerünkben (I./1.5. pont) a PARP-regulátorral
vagy a hatóanyagot nem tartalmazó vivőanyaggal előkezelt egércsopotok
tesztterületeiben egyszeri, eritémát okozó (2 MED) UV-irradiáció hatására

33
jelentkezett „sunburn” sejtek (apoptótikus keratinociták) számát hasonlítottuk
össze az UV-sugárzástól védett (kezeletlen) kontroll állatok bőrében
észleltekkel. A „sunburn” sejtek kvantitatív meghatározását 24 órával az
UVB-expozíció után vett bőrmintákból készült, hematoxilin-eozinnal festett
hisztológiai készítményekben vizsgáltuk.
Az UVB-besugárzástól védett, kontroll állatok bőrmintáiban az epidermisz 1
milliméteres bazálmembrán feletti szakaszán a „sunburn” sejtek átlagos
száma 0,5 ± 0,2 volt (2. Táblázat). Az UV-expozíció hatására a vivőanyaggal
kezelt bőrben, az ugyanekkora epidermisz szakaszon megfigyelhető
„sunburn” sejtek száma hetvenötszörösére (37,4 ± 1,4 sejt/mm) nőtt, ami
kifejezett DNS-károsodásra utalt (5. Ábra). A PARP-regulátorral történt
előkezelés már az 5% BGP-15M hatóanyag tartalom mellett szignifikánsan
(p<0,01) csökkentette az UVB-besugárzás által indukált apoptótikus
keratinociták képződését (8,8 ± 0,4 sejt/mm epidermisz), ami a szer
magasabb koncentrációinál (10% és 15%) még kifejezettebben érvényesült
(1,6 ± 0,3 és 1,1 ± 0,4 sejt/mm epidermisz) (6. Ábra). A 15% BGP-15M
tartalmú krém előkezelés 34-szeres védelmet mutatott az egyszeri, eritémát
okozó (2 MED) UVB-sugárzás apoptótikus sejthalált kiváltó hatásával
szemben. Amennyiben az UVB védelmi képesség megítélésének elterjedt
módját, a protektív faktort használjuk, úgy a PARP-regulátor (15% BGP-15M
tartalmú krém) protektív faktora a „sunburn” sejt képződésre: 34 (2.
Táblázat).
Az apoptótikus keratinociták számával meghatározott, UVB-expozíció
indukálta DNS-károsodásra vonatkozó eredmények összhangban vannak a
napfény által okozott, a bőrben képződött egyes láncú DNS törések

34
kvantitatív vizsgálatával kapott adatokkal, és a PARP-regulátor feltételezett
fotoprotektív tulajdonságát véli alátámasztani.
II./1.3. PARP-regulátor hatásának ultrastruktúrális vizsgálata az akut
UVB-sugárzás indukálta DNS-károsodásra a bőrben
A lokálisan alkalmazott PARP-regulátor hatását az akut UVB-irradiáció által
okozott fénykárosodás ultrastruktúrális eltéréseit frissen eltávolított bőrminták
ultravékony metszeteiben transzmissziós elektronmikroszkóppal vizsgáltuk.
A kezeletlen, UV-fénytől védett szőrtelen egerek bőrmintáiban kóros
ultrastruktúrális eltérést nem észleltünk. Ugyanakkor az eritémát okozó (2
MED) UVB-besugárzásban részesült (vivőanyaggal előkezelt) egérbőrben a
felszíni hámréteg bazális és szuprabazális sejtrétegében az apoptózis
ultrastruktúrális jeleit mutató sejteket detektáltunk (7. Ábra). A kromatin
marginális kondenzációját, részleges nukleáris fragmentációt, a mag
zsugorodását, perinukleáris ödémát és az intercelluláris kapcsolat részleges
felbomlását lehetett megfigyelni. Az elváltozás megfelelt a „sunburn” sejtekre
jellemzőknek (7. Ábra). A lokálisan alkalmazott PARP-regulátorral (BGP-15M
tartalmú krémmel) előkezelt, UVB-irradiációban részesített állatok
bőrmintáiban apoptózisra jellemző sejtkárosodást az elektronmikroszkópos
feldolgozás során nem észleltünk (8. Ábra).
Az akut UV-expozícióval végzett ultrastruktúrális vizsgálatok eredményei
további adatot szolgáltattak a PARP-regulátor DNS-protektív szerepére.

35
II./1.4. PARP auto-ADP-ribozilációja és regulációja UV-besugárzott
bőrben
A poli(ADP-ribóz) polimeráz, egy olyan evoluciónárisan konzervatív nukleáris
enzim, mely az eukarióta sejtekben a környezeti (pl. UV-sugárzás) és
endogén genotoxikus hatásokkal szembeni DNS-károsodást elhárító
rendszer ismert szabályzó faktora (D’Amours és mtsai. 1999). A nagyfokú
DNS-károsodás, amely direkt módon, túlméretezett PARP-aktivációhoz
vezet, a nukleáris fehérjék fokozott poli-ADP-ribozilációja, valamint a PARP
auto-ADP-ribozilációja által előidézi a NAD+ és az ATP deplécióját, a sejt
energia metabolizmusának károsodását, és ezáltal a sejtfunkciók különböző
mértékű zavarát eredményezheti.
Az egyszeri, magas dózisú, természetes UV-expozíció akut DNS-károsító
hatása által a szőrtelen egérbőrben indukált PARP-aktiváció mértékét az
enzim auto-ADP-ribozilációjának mérésével határoztuk meg. Kísérleti
körülményeink között (I./1.7. pont) az egér csoportok tesztterületeinek az UV-
expozíció előtt különböző koncentrációjú BGP-15M tartalmú krémmel történő
kezelésével a PARP-regulátor enzimaktivitásra kifejtett hatását vizsgáltuk. A
PARP auto-ADP-ribozilációjának meghatározását az UV-irradiáció (napfény
expozíció) után közvetlenül eltávolított bőrmintákban végeztük. Western-blot
technikát alkalmaztunk, az ADP-ribozilált fehérjéket anti-poli(ADP-ribóz)
monoklonális antitesttel (peroxidáz komplex) detektáltuk, „ECL”-módszerrel
láthatóvá tettük, valamint „Image-Tool (verzió 1.27) image processing”
programmal a jelintenzitás erősségének mértékét kvantitatív módon

36
meghatároztuk (3. Táblázat). A továbbiakban immunhisztokémiai technikával
vizsgáltuk a bőrminták epidermális sejtjeiben a PARP ADP-ribozilációját.
II./1.5. PARP auto-ADP-ribozilációjának vizsgálata Western-blot
analízissel akut UV-fény expozícióban részesült egérbőrben
Kísérleti körülményeink között (I./1.7. pont) a kezeletlen szőrtelen egerek
bőrmintáiban az ADP-riboziláció mértéke (Western-blot jel) a nehezen
detektálható tartományba esett, ami arra utalt, hogy az UV-irradiációtól
védett állatok bőrében a PARP aktivációs szignál intenzitása (DNS törések
mennyisége) gyakorlatilag elenyésző volt (9. Ábra és 3. Táblázat). Az egy
órás napfény expozícióban részesült, vivőanyaggal előkezelt állatokból
származó bőrmintákban a PARP enzim auto-ADP-ribozilációja az UV-
sugárzástól védett kontroll bőrben meghatározott érték 25x-re nőtt.
Ugyanakkor a ≥ 10% BGP-15M tartalmú krémmel végzett előkezelés
hatására az UV-expozíció által kiváltott fokozott ADP-riboziláció mértéke, a
Western-blot analízissel kapott jelek intenzitásának kvantitatív
meghatározása alapján, több mint 50%-kal csökkent. A BGP-15M lokális
alkalmazásával, UV-besugárzás hatására a bőrben létrejött extrém magas
PARP-aktivitás (a PARP-enzim auto-ADP-ribozilációjával mérve) szignifikáns
mértékben (p<0,01) csökkent.

37
II./2. PARP-regulátor a bőr akut fénykárosodására kifejtett hatásának
klinikai vizsgálata
II./2.1. PARP-regulátor hatásának vizsgálata egyszeri UVB-expozíció
indukálta akut klinikai tünetekre
A lokális BGP-15M-kezelés potenciális fotoprotektív hatását szőrtelen egerek
bőrén, az egyszeri, eritematogén (2 MED) UVB-irradiáció által kiváltott, akut
fénykárosodás (dermatitis solaris) klinikai tüneteinek öt fokozatú skálán
történő értékelésével vizsgáltuk. A különböző koncentrációjú (5%, 10%, 15%
és 20%) BGP-15M tartalmú krémmel, illetve vivőanyaggal előkezelt
tesztterületeken az UVB-irradiáció után 24 órával jelentkezett klinikai tünetek
(eritéma, ödéma, vezikula, bulla, erózió) súlyosságát vizuálisan és
dermatoszkóppal határoztuk meg (10. Ábra). Az egyszeri, 2 MED UVB-
besugárzás hatására a vivőanyaggal kezelt csoportban 5 állatnál a
tesztterületen intenzív bőrpír (eritéma pont: 3), valamint enyhe ödéma
(pontérték: 2), egy állatnál pedig jól definiálható eritéma (pontérték: 2)
jelentkezett. Az alkalmazott kísérleti feltételek mellett > 5% BGP-15M
tartalmú krémmel végzett előkezelés teljes mértékű védelmet mutatott az
UVB-irradiáció indukálta akut klinikai tünetek kialakulásával szemben. Az
egerek 5% BGP-15M lokális kezelésben részesült tesztterületén egy állatban
jól definiálható eritéma és nagyon enyhe ödéma (pontérték: 2, illetve 1),
valamint két további esetben bizonytalan eritéma (pontérték: 1) jelentkezett.
A vizsgálat során a tesztterületeken (BGP-15M-krémmel kezelt UVB-
besugárzott és nem besugárzott) irritációra utaló elváltozást nem észleltünk.

38
Az alkalmazott kísérleti körülmények között a BGP-15M ≥ 10%
koncentrációban fotoprotektívnek bizonyult (10. Ábra). A továbbiakban ezt
tekintettük a BGP-15M antieritematogén koncentrációjának.
II./2.2. PARP-regulátor hatásának vizsgálata UVB-expozíció indukálta
akut hisztológiai elváltozásokra bőrben
A különböző koncentrációjú BGP-15M tartalmú krémmel, illetve vivőanyaggal
előkezelt (I/1.5. pont) egerek tesztterületének bőréből 24 órával az UV-
besugárzás után vett, valamint a negatív kontrollként szolgáló kezeletlen
állatok bőréből nyert szövetmintákat formalinos fixálást, paraffinos
beágyazást, He-festést követően vizsgáltuk. A vivőanyaggal előkezelt UVB-
expozícióban részesített bőrminták hisztológiai feldolgozása során a
kezeletlen kontrollhoz képest különböző mértékű, fénykárosodásra utaló
eltéréseket lehetett megfigyelni, melyek a következők voltak: számos,
eozinofil citoplazmával, piknótikus sejtmaggal rendelkező, vagy mag nélküli
apoptótikus keratinocita. A dermisz felső rétegében részben
perivaszkulárisan, részben a dermo-epidermális határ mentén ödémásan
fellazult alapállományban neutrofil granulociták láthatók, a papilláris réteg
erei dilatáltak, vér alakos elemeivel kitöltöttek, endotéljük duzzadt (11. Ábra).
A ≥ 10% BGP-15M tartalmú krémmel előkezelt állatok csoportjaiban a
kezeletlen kontroll bőrben észleltekhez képest lényeges kóros hisztológiai
eltérést nem találtunk.
A ≥ 10% BGP-15M tartalmú krémmel előkezelt, 2 MED UVB-besugárzott
bőrben a „sunburn” sejtek száma a bazális membrán 1 mm-es szakasza

39
feletti epidermiszben átlagosan egy-egy volt (2. Táblázat). Ugyanakkor a
vivőanyaggal kezelt, UVB-besugárzott állatok azonos hosszúságú
epidermisz szakaszában átlagosan 37 „sunburn” sejtet lehetett detektálni.
Amennyiben az UV-károsodás mértékét az egységnyi epidermiszben jelen
levő „sunburn” sejtek számával fejezzük ki, úgy a BGP-15M ≥ 10%
koncentrációban alkalmazva több mint harmincszoros védelmet jelent, az
egyszeri, eritematogén UVB-irradiáció károsító hatásával szemben (11.
Ábra).
II./2.3. PARP-regulátor antieritematogén hatásának összehasonlító
vizsgálata ismert fényvédő faktorú (SPF) készítménnyel
A BGP-15M tartalmú krémet ≥ 10% koncentrációban alkalmazva, egy
forgalomban levő, jól ismert, SPF (sun protection factor) 30 jelzésű, az UVB
és UVA tartományban védelmet biztosító (általunk ”AS”-sel jelölt)
készítménnyel hasonlítottuk össze antieritematogén hatás vonatkozásában.
A kísérletben egyszeri, eritematogén (2MED) UVB-besugárzást alkalmaztunk
(1.5. pont). Az egerek pozitív kontrollként használt, UVB-vel irradiált
tesztterületein (n=6) dermatitis szoláris klinikai tüneteit (eritéma pont: 3-2,
ödéma pont: 2-1) lehetett megfigyelni (12. Ábra). Az „AS”, illetve BGP-15M
előkezelésben részesült tesztterületeken az UVB-expozíció klinikai tünetet
nem okozott, a környező (UV-fénytől takarással védett) bőrrel, illetve a
negatív kontrollként használt állatok bőréhez hasonlítva makromorfológiai
eltérést nem észleltünk. Hisztológiai vizsgálattal „AS”, ill. BGP-15M
kezelésben részesített bőrmintákban lényeges kóros eltérést nem találtunk.

40
Az alkalmazott kísérleti körülmények között a ≥ 10% BGP-15M tartalmú krém
fotoprotektív hatása megegyezett az „AS” jelzésű, 30-as fényvédő faktorú
(SPF 30) készítményével.
II./2.4. PARP-regulátor fotoprotektív hatásának vizsgálata a bőr akut
szoláris károsodására
Szőrtelen egerek tesztterületeit magas dózisú szoláris UV-irradiációban
részesítettük. A maximális irradiancia a kísérlet időpontjában a fény UVB-
tartományában (287-320 nm-re integrálva): 1,08 J/cm2/h volt (hazánkban az
irradiancia maximális értéke: 1,2-1,3 J/cm2). Az UVA-tartományában (321-
363 nm-re integrálva): 8,3 J/cm2/h volt (hazánkban az irradiancia maximális
értéke: 8,5-8,6 J/cm2). A biológiailag effektív intenzitások: UVB (287-320
nm): 0,07 J/cm2, UVA (321-363 nm): 0,015 J/cm2. (A mérések az Országos
Meterológiai Szolgálat Légkörfizikai Intézetben, Budapest, a számítások a
PTE Kísérleti Fizikai Tanszékén történtek, Pécs).
Az állatok tesztterületei a napfény expozíció előtt 10%, 15%, illetve 20%
BGP-15M tartalmú krém, illetve vivőanyaggal előkezelésben részesültek
(1.5. pont) UV-besugárzástól védett, kezeletlen állatok csoportja szolgált
kontrollként. A tesztelt bőrterületek állapotát 24 órával az UV-irradiációt
követően vizsgáltuk.
A vivőanyaggal előkezelt, intenzív UV-sugárzásnak kitett tesztterületeken a
bőr a napégés klinikai tüneteit (lividvörös eritéma, súlyos ödéma, két esetben
bullaképződés mutatta (13. Ábra). Hisztológiailag a klinikai tüneteknek
megfelelően erodált, illetve ödémásan fellazult, csökkent magfestésű,

41
nekrotikus hámfelszínen nagy mennyiségű fibrincsapadék volt megfigyelhető.
A dermisz legfelső részében elhelyezkedő neutrofil granulociták és
magtörmelékeik denz, széles sávot képeztek (14. Ábra). Másrészről a ≥ 10%
BGP-15M tartalmú krémmel végzett előkezelés az intenzív UV-sugárzás által
indukált súlyos klinikai tüneteket kivédte (13. Ábra). A BGP-15M tartalmú
krémmel kezelt tesztterületeken a bőr sem a környező, UV-besugárzástól
takarással védett, sem az UV-expozícióban nem részesült állatok bőréhez
képest szabadszemmel megfigyelhető, vagy dermatoszkóppal detektálható
eltérést nem mutatott. A bőrminták hisztológiai feldolgozása során normális
anatómiai struktúrának megfelelő szövettani képet láttunk: két rétegből álló
hám, szerkezetében megtartott dermisz, az akut UV-károsodásra utaló
lobosodás jelei nélkül. Az UV-besugárzásban nem részesült kontroll
bőrmintákban kóros hisztológiai eltérést nem észleltünk.
II./3. PARP-regulátor hatása a bőr UVB-sugárzás indukálta
karcinogenezisére (hairless egérmodell)
A PARP-regulátor BGP-15M potenciális fotoprotektív tulajdonságát a
krónikus UVB-sugárzás hatására, a szőrtelen egerek bőrében kialakuló,
„photoaging”-hez, fotokarcinogenezishez vezető változások követésével
határoztuk meg. Klinikai, hisztológiai, immunhisztokémiai és ultrastruktúrális
vizsgálatokat végeztünk a következő egércsoportokban: I.: 15% BGP-15M +
UVB; II.: vivőanyag + UVB; III.: 15% BGP-15M; IV.: kezeletlen kontroll. Az
egereket 32 héten át, heti 5 alkalommal, 15% BGP-15M krém, vagy
vivőanyag előkezelés után 0,25 J/cm2/nap UVB-besugárzásban

42
részesítettük. A klinikai tüneteket a kezelések előtt vizsgáltuk, és átlagosan
2-4 hetente fotodokumentációval rögzítettük. A kezelés második hetétől a
vivőanyaggal előkezelt, UVB-besugárzott tesztterületen az eritémás, ödémás
tüneteket, az egyre mélyülő színű pigmentáció és infiltráció váltotta fel (15.
Ábra). A ≥ 1 mm tumorok időbeni jelentkezését, valamint a tumoros állatok
prevalenciáját a 16. Ábrán tüntettük fel. Az első tumort (átmérő: 1,1 mm)
vivőanyaggal kezelt, UVB-besugárzott szőrtelen egerek csoportjában (II.
csoport) a kezelés 18. hetében észleltük. A tumoros állat az észlelés
időpontjáig 22,5 J/cm2 összdózisú UVB-besugárzást kapott. A 15% BGP-
15M krémmel előkezelt egerek bőre elváltozást nem mutatott (17. Ábra). A
19. héten már 3 egéren lehetett 1-2 mm nagyságú, vörösesbarna színű,
félgömbszerűen kiemelkedő tumor jelentkezését megfigyelni, és a
feldolgozás során hisztológiailag igazolni. A 19. héttől kezdve a vivőanyaggal
kezelt, krónikus UVB-besugárzásban részesített egerekben progresszív
tumorképződést észleltünk. A vivőanyaggal kezelt, UVB-irradiált csoport
minden tagja a 23. hétre egy vagy több, klinikailag és hisztológiailag
verifikálható, spinocellularis carcinomá-val, illetve keratosis solaris-nak
megfelelő bőrelváltozással rendelkezett, ugyanakkor a PARP-regulátor, 15%
BGP-15M krémmel előkezelt egerek bőre enyhe fokú pigmentáción kívül más
eltérést nem mutatott. A 32. héten a vivőanyaggal előkezelt, krónikus UVB-
besugárzásban részesült állatok bőre a tesztterületen jelentősen
megvastagodott, sötétbarna-szürkés pigmentációt mutatott, felszíni
egyenetlenség jeleivel, különösen a multiplex tumorok környezetében (18.
Ábra). A tumorok fejlődése során az adott területen hiperpigmentáció és
depigmentáció egyaránt megfigyelhető volt. A 32. héten, a kísérlet

43
lezárásával, a 15 % BGP-15M krémmel előkezelt, és krónikus UVB-
expozícióban (összdózis: 39 J/cm2) részesített egércsoportban, a tesztterület
bőrét összehasonlítva a kezeletlen kontroll állatokéval, illetve a környező,
takarással védett bőrfelszínnel, barnás-vörös színű pigmentáción kívül egyéb
klinikailag értékelhető eltérést nem találtunk (18. Ábra). 8 hónap elteltével
tumorképződésre utaló klinikai tünetet az UVB-expozíciótól védett, BGP-15M
krémmel kezelt (III. csoport), illetve kezeletlen kontroll állatokon (IV. csoport),
valamint a krónikus UVB-expozícióban részesített, PARP-regulátorral (BGP-
15M krémmel) előkezelt szőrtelen egereken (I. csoport) nem lehetett
megfigyelni. Kísérleteink eredményei alapján megállapíthatjuk, hogy a
PARP-regulátor a krónikus UVB-expozíció fotokarcinogén hatását kivédte.
Az UV-expozíciótól védett, 15% BGP-15M krémmel kezelt állatokon (III.
csoport) a 32 hetes kezelés alatt irritatív vagy allergiás reakcióra utaló
eltérést nem észleltünk (18. Ábra).
A klinikai vizsgálattal megfigyelt tumorképződés hisztológiailag igazolást
nyert.
A 15% BGP-15M krémmel előkezelt, UVB-expozícióban részesített egerek
szövetmintáinak vizsgálati eredménye alapján, a PARP-regulátor gátolta az
UVB-fény által, a vivőanyaggal előkezelt mintákban megfigyelt, súlyos
fénykárosodásra (epidermisz kiszélesedése, akantózis, sejtes infiltráció,
rugalmas rostok fragmentációja, melaninakkumuláció) (19-22. Ábra) és
tumorképződésre (23. Ábra) utaló hisztológiai, immunhisztológiai (IL-10,
TNFα, és p53) (24-26. Ábra) és szubmikroszkópos (27. Ábra) elváltozások
kialakulását.

44
II./4. PARP-regulátor hatása a bőr UVA-sugárzás indukálta
karcinogenezisére („forszírozott UVA”-kezelés)
A PARP-regulátor BGP-15M potenciális fotoprotektív tulajdonságát a
krónikus „forszírozott UVA”-kezelés hatására a szőrtelen egerek bőrében
kialakuló, „photoaging”-hez, fotokarcinogenezishez vezető változások
követésével vizsgáltuk.
A klinikai tünetek (eritéma, pigmentáció, hámlás, erózió, kifekélyesedés)
súlyosságát 0-5 pontos skálán (nincs elváltozás - igen súlyos), a tumorok
(átmérő ≥ 1mm) jelentkezését naponta, a kezelések előtt szabad szemmel és
dermatoszkóppal végzett vizsgálat alapján regisztráltuk (4. Táblázat). Az
állatok I-IV. csoportjában (I.:I./1.: 8-MOP + UVA, I./2.: UVA; II.: II./1.: 8-MOP
+ vivőanyag + UVA; II./2.: 8-MOP + 15% BGP-15M krém + UVA; III.: III./1.: 8-
MOP + vivőanyag, III./2.: 15% BGP-15M krém; IV.: kezeletlen kontroll)
huszonhat héten keresztül értékelve a makromorfológiai elváltozásokat a
következő eredményt kaptuk: az első hónap végén, amikor az állatok 10
J/cm2 összdózisú UVA-besugárzást kaptak, a 8-MOP-pal fényérzékenyített
tesztterületeken intenzív (8-MOP + UVA: 4,0 ± 0,8) és (8-MOP + vivőanyag +
UVA:3,8 ± 0,7) eritéma, valamint egy-egy állaton erózió jelentkezett. A 15%
BGP-15M krémmel előkezelt, fényérzékenyített, UVA-besugárzott állatok
bőrén kóros elváltozást nem észleltünk (28. Ábra és 4. Táblázat). Irritatív,
vagy allergiás reakciót nem tudtunk megfigyelni.
Az idő előrehaladtával, a 3. hónap végére, az UV-besugárzott állatokon a
15% BGP-15M krémmel előkezeltek kivételével, a gyulladás mértéke egyre
kifejezettebbé vált (pontszám: 3 - 5) és egyre több állaton lehetett

45
exulcerációt (pontszám: 2 – 3) megfigyelni (29. Ábra és 4. Táblázat). Az első
tumor a 8-MOP + UVA-kezelt állaton a 14. héten jelentkezett (30. Ábra és 5.
Táblázat). A 4. hónap végére a 8-MOP + UVA-kezelésben részesült
csoportban mind a 6 állaton, a fényérzékenyítést nem kapott, UVA-
besugárzott csoportban 4/6 állaton és a vivőanyaggal előkezelt,
fényérzékenyített, és UVA-besugárzott csoportban pedig 5/6 állaton egy,
vagy több tumor jelentkezett (31. Ábra és 5. Táblázat). Az UVA-val kezelt
csoportban (I./2) a tumorok átmérőjének nagysága alig haladta meg az 1
mm-t és ebben a csoportban egy állat még az 5. hónap végén is tumor
mentes volt. Ugyanekkor végzett klinikai vizsgálattal a 15% BGP-15M
krémmel előkezelt, fényérzékenyített, UVA-besugárzott egerek bőre továbbra
is tünet mentesnek bizonyult (32. Ábra).
Több mint fél év elteltével (26. hét) az UVA-kezelésben részesült
csoportokban kifejezett pigmentációt találtunk (pontérték: 3,7± 0,5 ; 3,2 ± 1,1;
3,7± 0,4). A 15% BGP-15M krémmel előkezelt, UVA-besugárzott csoportban
enyhe fokú pigmentációt észleltünk (pontérték: 1,8 ± 0,4) (31. Ábra, 4.
Táblázat). A fényérzékenyített, UVA-besugárzott állatokon különböző
mértékű hegképződést lehetett megfigyelni a korábban exulcerált
területeken. A 15% BGP-15M krémmel előkezelt, fényérzékenyített, UVA-
besugárzott állatok kivételével az UVA-expozícióban részesült csoportokban
különböző nagyságú tumorokat észleltünk.
Önmagában, besugárzás nélkül a 8-MOP, a vivőanyag, vagy a PARP-
regulátor bőrelváltozást nem okozott (irritatív, vagy allergiás reakció nem
jelentkezett) (31. Ábra). A hisztológiai, immunhisztológiai és elektron-
mikroszkópos eredmények a klinikai képpel korreláltak (33-37. Ábra).

46
III. Összefoglalás
III/1. A PARP-regulátor BGP-15M DNS-protektív hatása a bőrben
Az UV-sugárzás által az egérbőrben indukált akut DNS-károsodás
mértékét a képződött egyes láncú DNS-törések mennyiségének
vizsgálatával azt találtuk, hogy az intenzív napfény expozíció a
szövetmintákban az ép, nem károsodott DNS-arányát 30% alá
csökkentette. Ugyanakkor a PARP-regulátor lokális alkalmazása kifejezett
védelmet biztosított a napfény UV-tartományának DNS-károsító hatásával
szemben. Az UV-irradiáció által okozott DNS károsodás hisztológiai
markereként nyilvántartott epidermális „sunburn” sejtek (apoptótikus
keratinociták) kvantitatív meghatározása során, az UV-besugárzott,
vivőanyaggal kezelt bőrben a kontrollhoz képest hetvenötszörösére
emelkedett „sunburn” sejtek száma. A „sunburn” sejt képződéssel
szemben a PARP-regulátor használata DNS-protektívnek bizonyult. A
PARP-regulátorral előkezelt bőrmintákban az apoptótikus keratinociták
száma közel azonos volt a kontroll bőrben észleltekkel. Az ultrastrukturális
vizsgálatok további adatokat szolgáltattak a PARP-regulátor DNS-
protektív szerepére.
III/2. A PARP-regulátor (BGP-15M) hatása az UV-besugárzás által
bőrben indukált immunszupresszióra
Az UV-fény ismert, negatív tulajdonságai között kiemelt jelentőségű a
fotokarcinogenezis „előszobája”-ként nyilvántartott fotoimmunszupresszív
hatás. A korszerű UV-tartományban fényvédő anyagokkal szemben elvárás,
hogy a bőr, mint immunszerv funkciójának fiziológiás működését biztosítsák,
vagyis megakadályozzák a fotoimmunszupresszió/moduláció kialakulását
(Lim és mtsai. 2001). Az UVB-besugárzás után észlelhető immunszupresszió

47
és gyulladás az epidermális sejtek által termelt citokinek (pl. IL-10, TNFα, IL-
6, IL-1, IL-12,stb) és biológiailag aktív anyagok modulációja (up-,down-
regulációja) révén manifesztálódik. Egérbőrben az IL-10 és TNFα citokineket
az UVB-fény indukálta immunszupresszió markereként tartják számon
(Kripke és mtsai. 1992). A PARP-regulátor (BGP-15M) fotoimmunprotektív
hatásának meghatározását az IL-10 és TNFα citokinek immunhisztokémiai
vizsgálatával végeztük. A vivőanyaggal előkezelt, UVB-besugárzott
mintákban az epidermális sejtek anti-IL-10 poliklonális antitesttel kifejezett
citoplazmatikus festődést mutattak az epidermisz teljes vastagságában.
Ugyanakkor a PARP-regulátorral kezelt mintákban szórványosan csak egy-
egy sejt festődött. A vivőanyaggal előkezelt, UVB-besugárzott bőrből készült
metszeteken anti-TNFα poliklonális antitesttel intenzív citoplazmatikus és
membránfestődést észleltünk. Az ödémásan kiszélesedett hámban részben
a bazális rétegben perinukleárisan intenzív, a felső sejtsorokban pedig
kevésbé kifejezett, diffúz, intracitoplazmális reakciót figyeltünk meg. A
papilláris dermisz legfelső rétegében közvetlenül a hám alatt elhelyezkedő
makrofágok is kifejezett festődést mutattak. A PARP-regulátorral előkezelt,
UVB-expozícióban részesült bőr epidermiszében, hasonlóan az UVB-
besugárzástól védett kontrollhoz, immunhisztokémiailag detektálható TNFα
szintézist nem észleltünk. Ezek a adataink azt mutatják, hogy a PARP-
reguláror (BGP-15M) lokális alkalmazása fotoimmunprotektív hatásúnak
bizonyult az UVB-irradiáció indukálta bőr-immunszupresszióval szemben.

48
III/3. A PARP-regulátor szerepe a bőr akut UVB-fény károsodás
elleni védelmében (klinikai és hisztológiai vizsgálatok)
A szőrtelen egerek bőrén különböző koncentrációjú BGP-15M tartalmú
krémmel előkezelt tesztterületeken, 2 MED UVB-irradiáció után 24 órával
jelentkezett klinikai tünetek (eritéma, ödéma, vezikula, bulla, erózió) alapján
határoztuk meg a PARP-regulátor antieritematogén koncentrációját.
A bőrminták hisztológiai feldolgozása során a vivőanyaggal előkezelt, UVB-
expozícióban részesített bőrben különböző mértékű, akut fénykárosodásra
utaló eltérést észleltünk. Számos, eozinofil citoplazmával, piknótikus
sejtmaggal rendelkező, vagy mag nélküli apoptótikus keratinocitát („sunburn”
sejtet), a dermisz felső rétegében, részben perivaszkulárisan, részben a
dermo-epidermális határ mentén ödémásan fellazult alapállományban
neutrofil granulocitákat, a papilláris rétegben dilatált, duzzadt endotélű, vér
alakos elemeivel kitöltött ereket lehetett látni. A ≥ 10% BGP-15M tartalmú
krémmel előkezelt állatok csoportjaiban lényeges kóros hisztológiai eltérést
nem találtunk. Amennyiben az UVB- károsodás mértékét az egységnyi
epidermiszben jelen levő „sunburn” sejtek számával fejezzük ki, úgy a BGP-
15M ≥ 10% koncentrációban alkalmazva több mint harmincszoros védelmet
jelent, az egyszeri, eritematogén UVB-irradiáció (vivőanyaggal kezelt bőrben
kifejtett) károsító hatásával szemben. Így az itt bemutatott eredmények a
lokálisan alkalmazott PARP-regulátor fotoprotektív hatását igazolták a bőr
akut UVB-fénykárosodása esetén.
III/4. PARP-regulátor antieritematogén hatásának összehasonlító
vizsgálata ismert fényvédő faktorú (SPF) készítménnyel
A BGP-15M tartalmú krém (≥ 10% koncentrációban alkalmazva)
antieritematogén hatását hasonlítottuk össze egy, a forgalomban levő, SPF
(Sun Protection Factor) 30 jelzésű, az UVB és UVA tartományban védelmet
biztosító (általunk ”AS”-sel jelölt) készítménnyel. Az „AS”, illetve BGP-15M

49
krém előkezelésben részesült tesztterületeken az UVB-expozíció klinikai
tüneteket nem okozott. Hisztológiai vizsgálattal az ”AS”-sel jelölt, ill. BGP-
15M krém kezelésben részesített bőrmintákban lényeges kóros eltérést nem
találtunk Ezen adatok jelzik, hogy az alkalmazott kísérleti körülmények között
a ≥ 10% BGP-15M tartalmú krém fotoprotektív hatása megegyezett az „AS”
jelzésű, 30-as fényvédő faktorú (SPF 30) készítményével.
III/5. PARP-regulátor hatása a szoláris UV-expozíció klinikai
tüneteire és a hisztológiai elváltozásokra
A vivőanyaggal előkezelt, intenzív napfény sugárzásnak kitett
tesztterületeken a bőr a napégés klinikai tüneteit (lividvörös eritéma, súlyos
ödéma, két esetben bullaképződés) mutatta. Hisztológiailag a klinikai
tüneteknek megfelelően erodált, illetve ödémásan fellazult, csökkent
magfestésű, nekrotikus hám felszínen nagy mennyiségű fibrincsapadék volt
megfigyelhető. A dermisz legfelső részében elhelyezkedő neutrofil
granulociták és magtörmelékeik sötét, széles sávot képeztek. Ugyanakkor a
≥ 10% BGP-15M tartalmú krémmel végzett előkezelés az intenzív szoláris
UV-sugárzás által indukált súlyos klinikai tüneteket kivédte. A PARP-
regulátor (BGP-15M krém ≥ 10%) fotoprotektívnek bizonyult, meggátolta az
intenzív UV-expozíció által indukált dermatitis solaris (napégés) klinikai és
hisztológiai tüneteinek megjelenését.

50
III/6. A BGP-15M krém protektív a krónikus UV-besugárzás indukálta
fotokarcinogenezis klinikai, hisztológai, immunhisztológiai és ultrastuktúrális
tüneteivel szemben
A 32 héten át, heti 5 alkalommal, 15% BGP-15M krém (vagy vivőanyag)
előkezelés után 0,25 J/cm2/nap UVB-besugárzásban részesített szőrtelen
egerek tesztterületein vizsgáltuk a PARP-regulátor hatását a krónikus
fénykárosodás elleni védelemben. Kísérleteink eredményei alapján a PARP-
regulátor lokális alkalmazása a krónikus UVB-expozíció (32 hét, UVB
összdózis: 39 J/cm2) fotokarcinogén hatását kivédte. A vivőanyaggal
előkezelt tesztterületeken a 18. héttől kezdve (UVB-dózis: 22,5 J/cm2 )
klinikailag 1-2 mm nagyságú, vörösbarna színű, félgömbszerűen kiemelkedő
tumorok jelentkeztek, amelyek hisztológiai vizsgálattal carcinoma
spinocellulare-nak bizonyultak. A 15% BGP-15M krémmel előkezelt egerek
bőrmintáinak hisztológiai feldolgozása során a kontroll bőrhöz képest az
epidermisz másfél-kétszeresére történő kiszélesedését lehetett megfigyelni,
a dermiszben lényeges kóros eltérést nem észleltünk. A vivőanyaggal kezelt,
UVB-irradiált csoport minden tagja a 23. hétre egy vagy több, klinikailag és
hisztológiailag verifikálható, spinocellularis carcinomával, illetve keratosis
solarisnak megfelelő bőrelváltozással rendelkezett, ugyanakkor a PARP-
regulátor, 15% BGP-15M krémmel előkezelt egerek bőre enyhe fokú
pigmentáción kívül más eltérést nem mutatott. A kísérlet befejezésekor a 32.
héten a vivőanyaggal előkezelt, krónikus UVB-besugárzásban részesült
állatok bőre a tesztterületen jelentősen megvastagodott, sötétbarna-szürkés
hiperpigmentációt és depigmentációt mutatott, felszíni egyenetlenség jeleivel,
különösen a multiplex tumorok környezetében. A 15% BGP-15M krémmel

51
előkezelt, és krónikus UVB-expozícióban (összdózis: 39 J/cm2) részesített
egércsoportban, a tesztterület bőrét összehasonlítva a kezeletlen kontroll
állatokéval, illetve a környező, takarással védett bőrfelszínnel, barnás-vörös
színű pigmentáción kívül egyéb klinikailag értékelhető eltérést nem találtunk.
Nyolc hónap elteltével tumor képződésre utaló klinikai, vagy hisztológiai
eltérést a BGP-15M krémmel előkezelt csoportban nem észleltünk. Az UVB-
expozíciótól védett (BGP-15M, ill kontroll csoport) állatokon spontán tumor
képződést nem lehetett megfigyelni. Irritatív, vagy allergiás reakciót a kísérlet
során nem észleltünk.
A krónikus fénykárosodás a vivőanyaggal előkezelt mintákban a 26. héten a
rugalmas rostok Orcein-Giemsa festéssel kimutatható károsodásában (nagy
gócokban fragmentáció és összecsapzódás), a 32. héten a tumorok körül az
elasztikus rostok megfogyatkozásában, és a kollagén rostok
felszaporodásában nyilvánult meg. A 15% BGP-15M krémmel előkezelt, és
UVB-irradiációban részesült egerek bőrmintáiban ugyanakkor megtartott, ill.
enyhén megfogyatkozott rugalmas rosthálózatot figyelhettünk meg, amit a
kronológiai öregedés hisztológiai jeleként értékeltünk. Amíg a vivőanyaggal
előkezelt mintákban a krónikus UV-expozíció hatására a dermiszben a
melanin a makrofágokban durva szemcsés formában, esetenként szabadon
a kötőszöveti alapállományban is megtalálható volt, addig a PARP-
regulátorral előkezelt egerek bőrében a melanin pigment a bazális réteg
egyes sejtjeiben gócosan jelent meg.
Immunhisztokémiai vizsgálattal a PARP-regulátorral előkezelt, krónikus UVB-
besugárzásban részesített bőrmintákban a bazális keratinociták p53
antitesttel csak elvétve adtak immunhisztológiai reakciót. A vivőanyaggal

52
előkezelt állatok bőrében a keratosis solarisnak megfelelő területeken foltos
p53 festődést kaptunk. Az invazív tumorokban a sejtek nagy része intenzív
p53 magpozitivitást adott. A precancerosus elváltozásokban megfigyelhető
„patch”-jelleg a klonális expanzióra utalt.
Elektronmikroszkópos vizsgálattal a vivőanyaggal előkezelt, 39 J/cm2 UVB-
expozícióban részesült egérbőrben a normál keratinocita struktúra részbeni
felbomlását, tonofibrillum fragmentumokhoz kötődve dezmoszóma struktúrát,
a perinukleáris régióban számfeletti centroszómákat, a sejtmagok nagy
részében durva, szabálytalanul összecsapzódott kromatin állományt
észleltünk. A szubmikroszkópos elváltozások jelentős része megfelelt a
carcinoma spinocellulare-ban látottaknak. A BGP-15M krémmel előkezelt,
UV-besugárzott bőrminták elektronmikroszkópos vizsgálata során egy
esetben sem észleltünk in situ, vagy invazív karcinomát.
A fenti eredmények némileg nem várható eredményt okoztak azaz a PARP
feltehetően nem komplett részleges gátlása A 32. héten az UVB expozicíót
megelőzően lokálisan alkalmazott PARP-regurátorral a krónikus UVB-
expozíció fotokarcinogén hatásának kivédését eredményezte. A hatás
feltehetően az UV expozicíóra bekövetkező, mások által is megfigyelt extra
PARP-aktiváció a BGP-15M-nek közel a normális szintre történő down-
regulációjával magyarázható.

53
III/7. PARP-regulátor hatásának vizsgálata UVA-sugárzás indukálta
fotokarcinogenezis klinikai tüneteire szőrtelen egérmodellen
A PARP-regulátor BGP-15M potenciális fotoprotektív tulajdonságát a
krónikus UVA- besugárzott, ill. 8-MOP (Geroxalen®) fényérzékenyített és
UVA- besugárzott szőrtelen egerek bőrében kialakuló, „photoaging”-hez,
fotokarcinogenezishez vezető változások követésével vizsgáltuk. A szőrtelen
egerek tesztterületei 26 héten át, heti 5 alkalommal, 15% BGP-15M krém
(vagy vivőanyag) előkezelés után 0,5 J/cm2/nap UVA-besugárzásban
részesültek. A klinikai tünetek (eritéma, pigmentáció, hámlás, erózió,
kifekélyződés) súlyosságát 0-5 pontos skálán, valamint a tumorok (átmérő ≥
1mm) jelentkezését vizsgáltuk a kezelések előtt. Az első tumort a 14. héten,
a fényérzékenyített és UVA-besugárzott a vivőanyaggal kezelt állaton
figyeltük meg. A 4. hónap végére a vivőanyaggal előkezelt, 8-MOP+UVA-
irradiációban részesült állatokon különböző nagyságú szoliter tumorokat
észleltünk. A PARP-regulátor lokális előkezelés fotoprotektívnek bizonyult
mind a 8-MOP-pal fényérzékenyített, mind pedig a „csak” UVA-besugárzott
(26 hét, összdózis: 65 J/cm2) állatok bőrében. A PARP-regulátor kivédte a
vivőanyaggal előkezelt, hasonló módon UV-expozíciónak kitett állatokban
klinikailag megfigyelt, és hisztológiailag igazolt tumorképződést.
A PARP-regulátorral előkezelt, UVA-besugárzott bőrmintákban hisztológiai
vizsgálattal a hám enyhe kiszélesedését (5-8 sejtréteg) és megtartott hám
struktúrát találtunk. Ugyanakkor a vivőanyaggal előkezelt, UVA-besugárzott
mintákban az epidermisz egyenetlenül akantótikusnak és
hiperkeratótikusnak, szegmentálisan parakeratótikusnak bizonyult a
hematoxilin-eozinnal festett metszetekben. Az epidermisz alsó részein a
szerkezet felbomlott, a magok polaritása megszűnt. Ezeken a területeken a

54
sejtmagok általában anapláziát, hiperkrom kromatin szerkezetet mutattak, és
nagyobb számban sejtoszlások is észlelhetők voltak. A bazális rétegben
kifejezett sejtproliferációt láttunk. A dermo-epidermális határ mentén főként
limfocitákból és hisztiocitákból álló lobosodást észleltünk.
A vivőanyaggal kezelt, UVA-besugárzott bőrmintákból készült hisztológiai
metszeteken Orcein-Giemsa festéssel a rugalmas rostok fragmentálódását
és összecsapzódását figyeltük meg a dermisz középső részében. A PARP-
regulátorral előkezelt UVA besugárzott bőrben (8-MOP+BGP-15M+UVA) az
elasztikus rostok száma minimális mértékben csökkent, a kollagén rostok
száma változatlan volt a kezeletlen, azonos korú kontroll bőrhöz viszonyítva.
Fontana-Masson festéssel kisebb mennyiségű melanin pigmentet tudtunk
fokálisan kimutatni a bazális és szuprabazális keratinocitákban. A
vivőanyaggal előkezelt, fényérzékenyített, UVA-besugárzott egérbőrben
Fontana-Masson festéssel fokálisan nagy mennyiségű melanint lehetett
kimutatni a bazális és szuprabazális keratinocitákban, az epidermisz alsóbb
sejtsoraiban, és a dermális makrofágokban tömeges melanin pigmentet
észleltünk. Ugyanakkor az UVA-fénnyel besugárzott bőrben kisebb
mennyiségben lehetett melanin pigmentet a hámban és az irhában
megfigyelni.
Immunhisztológiai vizsgálattal a 15% BGP-15M krémmel előkezelt, UVA-
besugárzott (8-MOP+BGP-15M+UVA), valamint az UV-expozícióban nem
részesült (8-MOP; 15% BGP-15M és kezeletlen kontroll) bőrmintákban p53
festődést nem észleltünk A vivőanyaggal előkezelt, UVA-besugárzott (8-
MOP+vivőanyag+UVA) részesített állatok bőrében a bazális sejtréteg
magjainak és a megbomlott szerkezetű hámrészletek tumorosan átalakult
sejtjeinek p53 akkumulációját igazoltuk.
Eletronmikroszkópos vizsgálattal a PARP-regulátorral (8-MOP+BGP-
15M+UVA) előkezelt és UVA expozicíóban részesült bőrben megtartott
szerkezetet láttunk, kóros ultrastruktúrális eltérés nélkül. A vivőanyaggal

55
előkezelt, UVA-besugárzott (8-MOP+vivőanyag+UVA) bőrminták
feldolgozása során már 4 hetes UVA-expozíciót (UVA-összdózis: 10J/cm2)
követően az intercelluláris rések kiszélesedését lehetett megfigyelni.
Az UVA-expozícióban nem részesült (8-MOP; 15% BGP-15M) bőrmintákban,
valamint a kezeletlen azonos korú kontroll állatok bőrében a mikroszkópos és
ultramikroszkópos vizsgálatok során kóros eltérést nem észleltünk.Ezek az
adatok jelzik, hogy PARP-regulátorral végzett kezelés protektívnek bizonyult
a több mint féléves UVA-besugárzás fotokarcinogén hatásával szemben.
A fenti adatok azt mutatják, hogy a PARP részleges gátlása jelentős
védelmet nyújthat akut és krónikus UV károsodás tüneteinek kialakulásával,
valamint fotokalcinogenésissel szemben, jelezve egy rendkívül fontos
területet PARP-inhibitorok klinikai alkalmazására. A BGP-15M krém jelenleg
NG-SUN néven fázis kettő vizsgálatokban vesz rész melynek kezdeti része
bíztató, így remélhetőleg belátható időn belül megjelenhet a klinikai
felhasználásban. Azaz az értekezésben bemutatott vizsgálatok ténylegesen
hozzájárulhatnak egy új típusú UV protektív anyag kifejlesztéséhez és a
gyógyászatba történő megjelenéséhez.

56
A témában elér új eredmények összefoglalása:
Igazolást nyert a BGP-15M PARP-regulátor
a, DNS-protektív hatása
b, fotoimmunoprotektív hatása UVB tartományban
c, fotoprotektív hatása megegyezik 30-as SPF faktorú gyári
készítménnyel
d, fotoprotektív hatása szoláris UV expozicióval szemben
e, protektív krónikus UVA-besugárzás indukálta carcinogenezisben

57
Irodalomjegyzék
[1] Affar E. B., Germain M., Winstall E., Vodenicharov M., Shah R. G.,
Salvesen G. S., Poirer G. G.: Caspase-3- mediated processing
poly(ADP-ribose) glycohydrolase during apoptosis. J. Biol. Chem.
(2001) 276:2935-2942.
[2] Benjamin R. C., Gill D. M.: ADP-ribosylation in mammalian cell ghosts.
Dependence of poly(ADP-ribose) synthesis on strand breakage in
DNA. J. Biol. Chem. (1980) 255:10493-10501.
[3] Bernardi R., Rossi L., Poirier G. G., Scovassi A. I.: Analysis of
poly(ADP-ribose) glycohydrolase activity in nuclear extracts from
mammalian cells, Biochem. Biophys. Acta. (1997) 1338:60-68.
[4] Bernstein E. F., Chen Y. Q., Kopp J. B., Fisher L., Brown D. B., Hahn P.
J., Robey F. A., Lakkakorpi J., Uitto J.: Long-term sun exposure
alters the collagen of the papillary dermis. Comparison of sun-
protected and photoaged skin by northern analysis,
immunohistochemical staining, and confocal laser scanning
microscopy. J. Am. Acad. Dermatol. (1996) 34:209-218.
[5] Bernstein E. F., Chen Y. Q., Tamai K., Shepley K. J., Resnik K. S.,
Zhang H., Tuan R., Mauviel A., Uitto J.: Enhanced elastin and fibrillin
gene expression in chronically photodamaged skin. J. Invest.
Dermatol. (1994) 103:182-186.
[6] Berton T. R., Mitchell D. L., Fischer S. M., Locniskar M. F.: Epidermal
proliferation but not the quantity of DNA photodamage is correlated
with UV-induced mouse skin carcinogenesis, J. Invest. Dermatol.
(1997) 109:340-347.
[7] Brash D. E., Ziegler A., Jonason A. S., Simon J. A., Kunala S., Leffell D.
J.: Sunlight and Sunburn in Human Skin Cancer: p53, Apoptosis, and
Tumor Promotion. Journal of Investigative Dermatology Symposium
Proceedings (1996) 1:136-142.
[8] Burkle A.: Physiology and pathophysiology of poly(ADP-ribosyl)ation.
Bioessays. (2001) 23:795-806.

58
[9] Cole C.: Sunscreen protection in the ultraviolet A region: how to
measure the effectinevess. Photodermatol. Photoimmunol.
Photomed. (2001) 17:2.
[10] Cruz P. D. Jr., Leverkus M., Dougherty I., Gleason M. J., Eller M., Yaar
M., Gilchrest B. A.: Thymidine dinucleotides inhibit contact
hypersensitivity and activate the gene for tumor necrosis factor α, J.
Invest. Dermatol., (2000) 114:253-258.
[11] D’Amours D., Desnoyers S., D’Silva I., Poirier G. G.: Poly(ADP-
ribosyl)ation reactions in the regulation of nuclear functions.
Biochem. J. (1999) 342:249-268.
[12] D’Amours D., Sallmann F. R., Dixit V. M., Poirer G. G.: Gain-of-fuction
of poly(ADP-ribose) polymerase-1 upon cleavage by apoptotic
proteases: implications for apoptosis. J. Cell. Sci. (2001) 114:3771-
3778.
[13] Dantzer F., Schreiber V., Niedergang C., Trucco C., Flatter E., De La
Rubia G., Oliver J., Rolli V., Menissier-de Murcia J., de Murcia G.:
Involvement of poly(ADP-ribose) polymerase in base excision repair.
Biochemie. (1999) 81:69–75.
[14] Daya-Grosjean L., Dumaz N., Sarasin A.:The specificity of p53 mutation
spectra in sunlight induced human cancers. Journal of
Photochemistry and Photobiology (1995) 28:115-124.
[15] de Gruijl F. R., Forbes P. D.: UV-induced skin cancer in hairless mouse
model. Bio. Essays. (1995) 17:651-660.
[16] de Murcia G., Huletsky A., Lamarre D., Gaudreau A., Pouyet J., Daune
M., Poirier G. G.: Modulation of chromatin superstructure induced by
poly(ADP-ribose) synthesis and degradation. J. Biol. Chem. (1986)
261:7011–7017 .
[17] de Murcia G., Menissier de Murcia J.: Poly(ADP-ribose) polymerase: a
molecular nick-sensor. Trends. Biochem. Sci. (1994) 19:172-176.
[18] de Murcia J. M., Niedergang C., Trucco C., Ricoul M., Dutrillaux B.,
Mark M., Oliver F. J., Masson M., Dierich A., LeMeur M.:
Requirement of poly(ADP-ribose) polymerase in recovery from DNA

59
damage in mice and in cells. Proc. Natl. Acad. Sci. (1997) 94: 7303-
7307.
[19] Diffey B. L.: What is light? Photodermatol. Photoimmunol. Photomed.
(2002) 18:68-74.
[20] Drazen D. L., Bilu D., Edwards N., Nelson R. J.: Disruption of poly
(ADP-ribose) polymerase (PARP) protects against stress-evoked
immunocompromise. Mol. Med. (2001) 7:761-766.
[21] Durkacz B. W., Lunec J., Grindley H., Griffin S., Horner O., Simm A.:
Murine melanoma cell differentiation and melanogenesis induced by
poly (ADP-ribose) polymerase inhibitors. Exp. Cell. Res. (1992)
202:287-291.
[22] Durkacz B. W., Omidiji O., Gray D. A., Shall S.: (ADP-ribose)n
participates in DNA excision repair. Nature (1980) 283:593-596.
[23] Elmets C. A., Singh D., Tubesing K., Matsui M., Katiyar S., Mukhtar H.:
Cutaneous photoprotection from ultraviolet injury by green tea
polyphenols. J. Am. Acad. Dermatol. (2001) 44:425-432.
[24] Farkas B., Magyarlaki M., Csete B., Németh J., Rablóczky Gy., Bernáth
S., Literáti Nagy P., Sümegi B.: Reduction of acute photodamage in
skin by topical application of a novel PARP inhibitor Biochem.
Pharmacol. (2002) 63:921-932.
[25] Farkas B., Sümegi B., Rablóczky Gy., Csete B., Hodosi B., Magyarlaki
M., Bernáth S., Literáti Nagy P.: Protecting effect of PARP inhibition
on UV light-induced skin damage. In: Zhang J. (ed), PARP as a
therapeutic target. Pharmacology & Toxicology Series, Boca Raton,
London, New York, Washington, CRC Press, (2002) pp:257-276.
[26] Faro R., Toyoda Y., McCully J., Jagtap P., Szabo E., Virág L., Bianchi
C., Levitsky S., Szabo C.: Protective effect on regional myocardial
function and infarct size induced by PJ34: a novel poly(ADP-ribose)
synthetase inhibitor. Ann. Thorac. Surg. (2002) 73: 575-581.
[27] Fisher G. J., Datta S. C., Talwar H. S., Wang Z. Q., Varani J., Kang S.,
Voorhees J. J.: Molecular basis of sun-induced premature skin
ageing and retinoid antagonism. Nature (1996) 379:335-339.

60
[28] Freeman S. E., Hacham H., Gange R. W., Maytum D. J., Sutherland J.
C., Sutherland B. M.: Wavelength dependence of pyrimidine dimer
formation in DNA of human skin irradiated in situ with ultraviolet light,
Proc. Natl. Acad. Sci. (1989) 86:5605-5609.
[29] Germain M., Affar E. B., D’Amours D., Dixit V. M., Salvesen G. S.,
Poirier G. G.: Cleavage of automodified poly(ADP-ribose)
polymerase during apoptosis. Evidence for involvement of caspase-
7. J. Biol. Chem. (1999) 274:28379-28384.
[30] Gilchrest B. A.: A review of skin ageing and its medical therapy. Brit. J.
Dermatol. (1996) 135:867-875.
[31] Gilchrest B. A.: Skin aging and photoaging : an overview. J. Am. Acad.
Dermatol. (1989) 21:610-613.
[32] Hemminki K., Xu G., Le Curieux F.: Ultraviolet radiation-induced
photoproducts in human skin DNA as biomarkers of damage and its
repair, IARC Sci. Publ. (2001) 154:69-79.
[33] Hinshaw D. B., Lodhi I. J., Hurly L. L., Atkins K. B., Dabrowska M. I.:
Activation of poly(ADP-ribose) polymerase in endothelial cells and
keratinocytes: role in an in vitro model of sulfur mustard-mediated
vesication. Toxicol. Appl. Pharmacol. (1999) 156:17-29.
[34] Horkay I., Varga L., Tamasi P., Gundy S.: Repair of DNA damage in
light sensitive human skin diseases. Arch. Dermatol. Res. (1978)
263:307-15.
[35] Horkay I.: Sun protection, sunscreens. Bőr. Vener. Szle. (1998) 74:83-
85.
[36] Hönigsmann H.: Erythema and pigmentation. Photodermatol.
Photoimmunol. Photomed. (2002) 18:75-81.
[37] Kanai M., Uchida M., Hanai S., Uematsu N., Uchida K., Miwa M.:
Poly(ADP-ribose) polymerase localizes to the centrosomes and
chromosomes. Biochem. Biophys. Res. (2000) 278:385-389.
[38] Katiyar S. K., Matsui M. S., Mukhtar H.: Kinetics of UV light-induced
cyclobutane pyrimidine dimers in human skin in vivo: an
immunohistochemical analysis of both epidermis and dermis.
Photochem. Photobiol. (2000) 72:788.

61
[39] Kim J. W., Kim K., Kang K., Joe C. O.: Inhibition of homodimerization of
poly(ADP-ribose) polymerase by its C-terminal cleavage products
produced during apoptosis. J. Biol. Chem. (2000) 275:8121-8125.
[40] Kim J. W., Won J., Sohn S., Joe C. O.: DNA-binding activity of the N-
terminal cleavage product of poly(ADP-ribose) polymerase is
required for UV mediated apoptosis. J. Cell. Sci. (2000) 113:955-961.
[41] Kraemer K. H.: Sunlight and skin cancer: another link revealed. Proc.
Natl. Acad. Sci. U S A. (1997) 94:11-14.
[42] Kripke M. L., Cox P. A., Alas L. G., Yarosh D. B.: Pyrimidine dimers in
DNA initiate systemic immunosuppression in UV-irradiated mice.
Proc. Natl. Acad. Sci. U.S.A. (1992) 89:7516-7520.
[43] Kripke M. L.: Latency, histology, and antigenicity of tumors induced by
ultraviolet light in three inbred mouse strains. Cancer Res. (1977)
37:1395-1400.
[44] Krobock E., Rahbari H., Mehregan A. H.: Acid orcein and Giemsa stain.
Modification of a valuable stain for dermatological specimens. J.
Cutan. Pathol. (1978) 5: 37-38.
[45] Krutmann J., Croteau D. L., Bohr V. A.: Repair of oxidative damage to
nuclear and mitochondrial DNA in mammalian cells, J. Biol. Chem.
(1997) 272:25409.
[46] Krutmann J.: The role of UVA rays in skin aging, Eur. J. Dermatol.
(2001) 11:170-171.
[47] Kulms D., Schwarz T.: Molecular mechanisms of UV-induced apoptosis,
Photodermatol. Photoimmunol. Photomed. (2000) 16: 195-201.
[48] Lim H. W., Naylor M., Honigsmann H., Gilchrest B. A, Cooper K.,
Morison W., Deleo V. A., Scherschun L.: American Academy of
Dermatology Consensus Conference on UVA protection of
sunscreens: summary and recommendations, J. Am. Acad.
Dermatol. (2001) 44:505-508.
[49] Lindahl T., Satoh M. S., Poirier G. G., Klungland A.: Post-translational
modification of poly(ADP-ribose) polymerase induced by DNA strand
breaks. Trends. Biochem. Sci. (1995) 20:405-411.

62
[50] Lowe N. J., Dromgoole S. H., Sefton J.: Indoor and outdoor efficacy
testing of a broad spektrum sunscreen against ultraviolet A radiation
in psoralen-sensitized subjects. J. Am. Acad. Dermatol. (1987)
17:224-230.
[51] Lowe N. J.: Sun protection factor: Comparative techniques and
selection of UV sources. In: Sunscreens,development, evaluation
and regulatory aspects. eds. by Lowe N. J., Shaath N., Marcel D.,
New York, (1990) pp. 379-394.
[52] Luna L. G: Manual of histologic staining methods of the Armed Forces
Institute of Pathology. Third edition, McGraw-Hill New York, (1968).
[53] Maccarrone M., Catani M. V., Iraci S., Melino G., Agro A. F.: A survey of
reactive oxygen species and their role in dermatology, J. Eur. Acad.
Dermatol. Venereol. (1997) 8:185-202.
[54] Menisser-de Murcia J. M., Niedergang C., Trucco C., Ricoul M.,
Dutrillaux B., Mark M., Oliver F. J., Masson M., Dierich A., LeMeur
M., Walztinger C., Chambon P., de Murcia G.: Requirement of
poly(ADP-ribose) polymerase in recovery form DNA damage in mice
and in cells. Proc. Natl. Acad. Sci. (1997) 94:7303-7307.
[55] Mikel U. V.: Advanced laboratory methods in histology and pathology
Washington DC: Armed Forces Institute of Pathology and Armed
Registry of Pathology, (1994).
[56] Milam K. M., Cleaver I. E.: Inhibitors of poly(adenosine diphosphate-
ribose) synthesis: effect on other metabolic processes, Science.
(1984) 223:589-591.
[57] Mitchell D. L., Byrom M., Chiarello S., Lowery M. G.: Effects of chronic
exposure to ultraviolet B radiation on DNA repair in the dermis and
epidermis of the hairless mouse, J. Invest. Dermatol. (2001) 116:209-
215.
[58] Molinete M., Vermeulen W., Burkle A., Menissier de Murcia J., Kupper
J. H., Hoeijmakers J. H., de Murcia G.: Overproduction of the
poly(ADP-ribose) polymerase DNA-binding demain blocks alkylation-
induced DNA repair synthesis in mammalian cells. EMBO (Eur. Mol.
Biol. Organ.) J. (1993) 12:2109-2117.

63
[59] Oliver J. F., Menissier-de Murcia J., de Murcia G.: DNA Repair ’99
Poly(ADP-Ribose) Polymerase in the Cellular Response to DNA
Damage, Apoptosis, and Disease. Am. J. Hum. Genet. (1999)
64:1282-1288.
[60] Pacher P., Liaudet L., Soriano F. G., Mabley J., Szabo E., Szabo C.:
The Role of poly(ADP-ribose) polymerase in the development of
myocardial and endothelial dysfunction in diabetes mellitus.
Diabetes. (2002) 51: 514-521.
[61] Pacher P., Mabley J. G., Soriano F. G., Liaudet L., Komjati K., Szabo
C.: Endothelial dysfunction in aging animals: the role of poly(ADP-
ribose) polymerase activation. Br. J. Pharmacol. (2002) 135:1347-
1350.
[62] Pacher P., Mabley J. G., Soriano F. G., Liaudet L., Szabo C.: Activation
of poly(ADP-ribose) polymerase contributes to the endothelial
dysfunction associated with hypertension and aging. Int. J. Mol. Med.
(2002) 9:659-664.
[63] Pease, D. C.: Histological techniques for electron microscopy, 2nd Ed.,
Academic Press Inc, New York. (1964).
[64] Rigel D. S., Friedman R. J., Kopf A. W.: Lifetime risk for development of
skin cancer in the U.S. population: current estimate is now 1 in 5. J.
Am. Acad. Dermatol. (1996) 35:1012-1013.
[65] Rougier A.: Are UVA rays dangerous? In.:Protection of the skin
ultraviolet radiations. ed. by Rougier A. and Schaefer H.John Libbey
Eurotext, Montrouge, London, Rome, Paris, (1998) 1-9.
[66] Rünger T. M.: Role of UVA in the pathogenesis of melanoma and non-
melanoma skin cancer. Photodermatol. Photoimmunol. Photomed.
(1999) 15:212-216.
[67] Satoh M. S., Lindahl T.: Role of poly(ADP-ribose) formation in DNA
repair. Nature. (1992) 356:356-358.
[68] Satoh M. S., Poirier G. G., Lindahl T.: Dual function for poly(ADP-
ribose) synthesis in response to DNA strand breakage. Biochemistry.
(1994) 33:7099-7106.

64
[69] Satoh M. S., Poirier G. G., Lindahl T.: NAD(+)-dependent repair of
damaged DNA by human cell extracts. J. Biol. Chem. (1993)
268:5480-5487.
[70] Schwartz A., Stander S., Berneburg M., Bohm M., Kulms D., van Steeg
H., Grosse-Heitmeyer K., Krutmann J., Schwarz T.: Interleukin-12
suppresses ultraviolet radiation-induced apoptosis by inducing DNA
repair. Nat. Cell. Biol. (2002) 4:26-31.
[71] Shah G. M., Poirier D., Duchaine C., Brochu G., Desnoyers S., Lagueux
J., Verreault A., Hoflack J. C., Kirkland J. B., Poirier G. G.: Methods
for biochemical study of poly (ADP-ribose) metabolism in vitro and in
vivo. Anal. Biochem. (1995) 227:1-13.
[72] Shall S.: ADP-ribosylation reactions, Biochimie. (1995) 77:313-318.
[73] Sims J. L., Berger S. J., Berger N. A.: Poly-(ADP-ribose) polymerase
inhibitors preserve nicotinamide adenine dinucleotide and adenosine
5’-triphosphate pools in DNA-damaged cells: mechanism of
stimulation of unscheduled DNA synthesis. Biochemistry. (1983)
22:5188-5194.
[74] Smulson M., Istock N., Ding R., Cherney B.: Deletion mutants of
poly(ADP-ribose) polymerase support a model of cyclic association
and dissociation of enzyme from DNA ends during DNA repair.
Biochemistry. (1994) 33:6186-6196.
[75] Soriano F. G., Virag L., Jagtap P., Szabo E., Mabley J. G., Liaudet L.,
Marton A., Hoyt D. G., Murthy K. G., Salzman A. L.: Diabetic
endothelial dysfunction: the role of poly (ADP-ribose) polymerase
activation. Nat. Med. (2001) 7:108-113.
[76] Steenvoorden D. P. T., Beijersbergen G. M. J.: The use of endogenous
antioxidants to improve photoprotection, J. Photochem. Photobiol. B:
Biol. (1997) 41:1-10.
[77] Sundberg J. P.: Handbook of mouse mutations with skin and hair
abnormalities. CRC Press. Boca Raton, Ann Arbor, London Tokyo.
(1994) pp.:291-310.
[78] Szabados E., Literati-Nagy P., Farkas B., Sümegi B.: BGP-15, a
Nicotinic Amidoxime Derivate Protecting Heart from Ischaemia

65
Reperfusion Injury through Modulation of Poly (ADP-ribose)
Polymerase. Biochemical Pharmacology (2000) 59:937-945.
[79] Szabo C., Dawson V. L.: Role of poly(ADP-ribose) synthetase in
inflammation and ischeamia-reperfusion. Trends Pharmacol. Sci.
(1998) 19:287-298.
[80] Szabo C., Virag L., Cuzzocrea S., Scott G. S., Hake P., O’Connor M.
P., Zingarelli B., Salzman A., Kun E.: Protection against peroxynitrite-
induced fibroblast injury and arthritis development by inhibition of
poly(ADP-ribose) synthase. Proc. Natl. Acad. Sci. U.S.A. (1998)
95:3867-3872.
[81] Szabo C.: Potential role of the peroxynitrite-poly (ADP-ribose)
synthetase pathway in a rat model of severe hemorrhagic shock.
Shock (1998) 9:341-344.
[82] Szabo E., Virag L., Bakondi E., Gyure L., Hasko G., Bai P., Hunyadi J.,
Gergely P., Szabo C.: Peroxynitrite production, DNA breakage and
poly(ADP-ribose)polymerase activation in a mause model of
oxazolone-induced contact hypersensitivity. J. Invest. Dermatol.
(2001) 117:74-80.
[83] Vigh L., N. Literati P., Horvath I., Torok Zs., Balogh G., Glatz A., Kovacs
E., Boros I., Ferdinandy P., Farkas B., Jaszlits L., Jednakovits A.,
Koranyi L., Maresca B.: Bimoclomol: A nontoxic, hydroxylamine
derivative with stress protein-inducing activity and cytoprotective
effects. Nat. Med. (1997) 3:1150-1154.
[84] Virag L., Bai P., Bak I., Bakondi E., Szabo E., Liaudet L., Gergely P.,
Szabo C.: Inhibition of poly(ADP-ribose) polymerase attenuates
leukocyte migration in an ovalbumin-induced asthma model in
mice.(Abstract). FASEB J. (2002) 16: A674.
[85] Virag L., Salzman A. L., Szabo C.: Poly(ADP-ribose) synthetase
activation mediates mitochondrial injury during oxidant-induced cell
death. J. Immunol. (1998) 161:3753-3759.
[86] Virag L., Scott G. S., Cuzzocera S., Marmer D., Salzman A. L., Szabo
C.: Peroxynitrite-Induced Thymocyte Apoptosis: the Role of

66
Caspases and Poly (ADP-Ribose) Synthetase (PARS) Activation.
Immunology (1998) 94:345-355.
[87] Virag L., Szabo C.: The therapeutic potential of poly(ADP-Ribose)
polymerase inhibitors. Pharmacol. Rev. (2002) 4:375-429.
[88] Virag L., Szabo É., Bai P., Gergely P., Hunyadi J., Szabo C.: Nitric
oxide peroxynitrite-poly(ADP-ribose) polymerase pathway in the skin.
Exp. Dermatol. (2002) 11:189-202.
[89] Yarosh D., Klein J., O'Connor A., Hawk J., Rafal E., Wolf P.: Effect of
topically applied T4 endonuclease V in liposomes on skin cancer in
xeroderma pigmentosum: a randomised study. Lancet (2001)
357:926-929.
[90] Young A. R., Chadwick C. A., Harrison G. I., Nikaido O., Ramsden J.,
Potten C. S.: The similarity of action spectra for thymine dimers in
human epidermis and erythema uggest that DNA is the chromophore
for erythema. J. Invest. Dermatol. (1998) 111:982-988.
[91] Young A. R., Walker S. L.: Sunscreens: Photoprotection of non-
erythema endpoints relevant to skin cancer. Photodermatol.
Photoimmunol. Photomed. (1999) 15:221-225.
[92] Zdenko H., Wang Z. Q.: Functions of poly(ADP-ribose)polymerase
(PARP) in DNA repair, genomic integrity and cell death. Mut. Res.
(2001) 477:97-110.
[93] Zhang J., Li J. H.: Poly(ADP-ribose)polymerase inhibition by genetic
and pharmacological means. In: Szabó C (ed), Cell Death: the Role
of PARP. Pharmacology & Toxicology Series, Boca Raton, London,
New York, Washington, CRC Press, (2000) pp:209-230.
[94] Ziegler A., Jonason A. S., Leffell D. J., Simon J. A., Sharma H. W.,
Kimmelman J., Remington L., Jacks T., Brash D. E.: Sunburn and
p53 in the onset of skin cancer. Nature (1994) 372:773-776.
[95] Zingarelli B., Cuzzocrea S., Zsengeller Z., Salzman A. L., Szabo C.:
Protection against myocardial ischemia and reperfusion injury by 3-
aminobenzamide, an inhibitor of poly (ADP-ribose) synthetase.
Cardiovasc. Res. (1997) 36:205-215.

67
RÖVIDÍTÉSEK
8-MOP: 8-metoxipsoralen ABC: avidin-biotin complex ADP: adenozin monofoszfát ATP: adenozin trifoszfát AMD: automodifikációs domén BRCT: breast cancer susceptibility protein C terminus CPD: ciklobután-pirimidin-dimerek CTL: citotoxikus T limfocita DBD: DNS kötő domén DEVD: kaszpáz hasító hely DNS: dezoxiribonukleinsav ECL: enhanced chemiluminescence system He: hematoxillin-eozin IL-10: interleukin-10 MED: minimális eritéma dózis NAD+: nikotinsavamid NK sejtek: természetes ölősejtek NLS: nukleáris lokalizációs szignál PARG: poli(ADP-ribóz) glükohidroláz PARP: poli(ADP-ribóz) polimeráz RNS: ribonukleinsav ROS: reaktív oxigén gyök SIS: skin immune system SPF 30: sun protection factor TNFα: tumor nekrózis faktor-α UV: ultraibolya

68
Köszönetnyilvánítás
Ezúton szeretném köszönetemet és hálámat kifejezni mindazoknak, akik e
munka elvégzésében, Ph.D. értekezésem megírásában szakmai és baráti
tanácsaikkal segítettek.
Prof. Dr. Sümegi Balázsnak és a PTE ÁOK Biokémiai Intézet összes
dolgozójának, akik kutatói pályámon elindítottak és munkámat mindvégig
segítették.
Prof. Dr. Farkas Beatrixnak, a PTE Bőr-, Nemikórtani és Onkodermatológiai
Klinika tanszékvezető egyetemi tanárának, aki Ph.D. munkám megírására
ösztönzött és akinek vezetése alatt végeztem tudományos munkámat.
Dr. Magyarlaki Márta és Dr Zombai Erzsébet adjunktusnőknek a szövettani,
immunhisztológiai és elektronmikroszkópos felvételek elkészítéséért és
értékeléséért.
Stein Anikónak és Kavas Máriának a szőrtelen egerekkel végzett
kísérleteimben nyújtott segítségükért.
Hálámat szeretném kifejezni családomnak támogatásukért, megértésükért és
szeretetükért.
A kísérletekhez megvalósításához támogatást nyújtottak: az OTKA, az ETT,
a DAAD-MÖB, valamint az N-Gene Research Laboratórium.

69
A szerző tudományos közleményei:
Petykó Z., Lénárd L., Sümegi B., Hajnal A., Csete B., Faludi
B., Jandó G.: Learning disturbances in offspring of
Zidovudine (AZT) treated rats. Neurobiology 5:83, 1997.
Szabados E., Fischer G.,Toth K., Csete B., Nemeti B.,
Trombitás K., Habon T., Endrei D., Sumegi B.: Role of
reactive oxygen species and poly-ADP-ribose polymerase in
the development of AZT-induced cardiomyopathy in rat. Free
Radical Biology and Medicine 26:309, 1999.
Farkas B., Sümegi B., Csete B., Szekeres Gy.: Novel
poly(ADP)-ribose polymerase inhibitor with photoprotective
activity. J Invest Dermatol 113:438, 1999.
Farkas B., Sümegi B., Rablóczky Gy., Csete B., Hodosi B.,
Magyarlaki M., Bernáth S., Literáti Nagy P.: Protecting
utilities of PARP inhibitors. Chapter 13, CRC Press, Boca
Raton, 2001.
Farkas B., Magyarlaki M., Csete B., Németh J., Rabloczky
Gy., Bernáth S., Literáti Nagy P., Sümegi B.: Reduction of
acute photodamage in skin by topical application of a novel
PARP inhibitor. Biochem. Pharmacology, 63: 1-12, 2002.
B. Farkas, B. Sümegi, Gy. Rabloczky, B. Csete, B. Hodosi,
M. Magyarlaki, S. Bernáth, P. Literáti Nagy: Protecting Effect
of PARP Inhibition on Ultraviolet Light-Induced Skin Damage.
Pp.: 257-271, In: Edit.: Jie Zhang: PARP as a therapeutic
Target. CRC Press, London, New York, Washington, 2002

70
Csete B., Zombai E., Farkas B.: Human papilloma virus
infection leading to social exclusion. Ann. Dermatol.
Vénéréol. 129: 681, 2002.
Hodosi B., Csete B., Zombai E., Farkas B.: Chronic actinic
dermatosis combined with allergy against occupational
factors. Ann. Dermatol. Vénéréol. 129: 1724, 2002.
Csete B., Németh J., Farkas B.: Giant multiplex condiloma
acuminata, acanthosis nigricans, and insulin resistant
diabetes mellitus Ann. Dermatol. Vénéréol. 129: 842, 2002.
Farkas B., Csete B., Magyarlaki M., Bernáth S., Sümegi B.:
Topical Poly(ADP-Ribose) Polymerase (PARP) regulator and
its prospects for use. Ann. Dermatol. Vénéréol. 129: 91, 2002.
Csete B., Magyarlaki M., Rablóczky Gy., Sümegi B., Farkas
B.,: Novel Poly(ADP-Ribose) Polymerase (PARP) regulator in
sun protection J. Eur Acad Derm. Vener. 17 (Suppl. 1.) :64,
2003.
Csete B., Magyarlaki M., Farkas B.: Effect of PARP-regulator
in chronic UVA-damage JDDG Supplement 1: 16, 2003.

71
A szerző kongresszusi előadásainak jegyzéke: Csete B., Skuta G., Fischer G., Sümegi B.,: AZT induced
cardiomyopathy in rats. XIII. International Student
Conference, Istanbul, 1997.
Csete B., Farkas B., Sümegi B.: AZT induced mitochondrial
DNA fragmentation. Congress of Hungarian Dermatologycal
Society, Budapest, 1997.
Csete B., Péter I., Farkas B. : PCR-Chlamydia diagnosis by
psoriatic and autoimmun patients. Annual meeting of
Hungarian Dermatologycal Society, Lillafüred, 1998.
Csete B., Fábos B., Nagy Gy., Farkas B.: Therapyresistent
pyoderma gangrenosum treated with Sandimmun NeoralR.
Congress of Young Dermatologist, Kecskemét, 2000.
Csete Béla, Szabados Eszter, Farkas Betarix, Sümegi
Balázs: Role of PARP in the development of AZT induced
tissue damage. Annual meeting of Hungarian
Dermatologycal Society, Lillafüred, 2001.
Csete B., Zombai E., Farkas B.: Human papilloma virus
infection leading social exclusion. Annual meeting of
Hungarian Dermatologycal Society, Debrecen, 2002.
Csete B., Zombai E., Farkas B.: Humán papilloma virus
infection leading to social exclusion. XX. World Congress of
dermatology, 2002. Paris

72
Csete B., Zombai E., Farkas B.: Florid cutaneous and mucosal
papillomatosis. DUDG 2002. Düsseldorf
Csete Béla, Zombai Erzsébet, Kádár Zsolt, Farkas Beatrix:
Calcinosis cutis Congress of Hungarian Dermatologycal
Society, 2002. Budapest
Csete B., Magyarlaki M., Farkas B.: Effect of PARP-regulator
in chronic UVA-damage 42. Tagung der Deutschen
Dermatologischen Gesellschaft 2003. Berlin
Csete B., Zombai E., Farkas B. Társadalmi
kirekesztettséghez vezető florid cutan papillomatosis Annual
Meeting of Hungarian Dermatologycal Society, 2003. Szeged
Csete B., Magyarlaki M., Farkas B.: Effect of PARP-regulator
on UV light induced photodamage. Tagung der Deutschen-
Ungarischen Dermatologischen Gesellschaft 2004. Pécs
Csete B., Lengyel Zs., Kádár Zs., Battyáni Z.: Poly(Adenosine
Diphosphate-Ribose) Polymerase-1 expression in cutaneous
malignant melanomas as a new molekular marker of agressive
tumors. Pathol Ocol Res 2008. Aug. 28. ( Epub ahead of print)
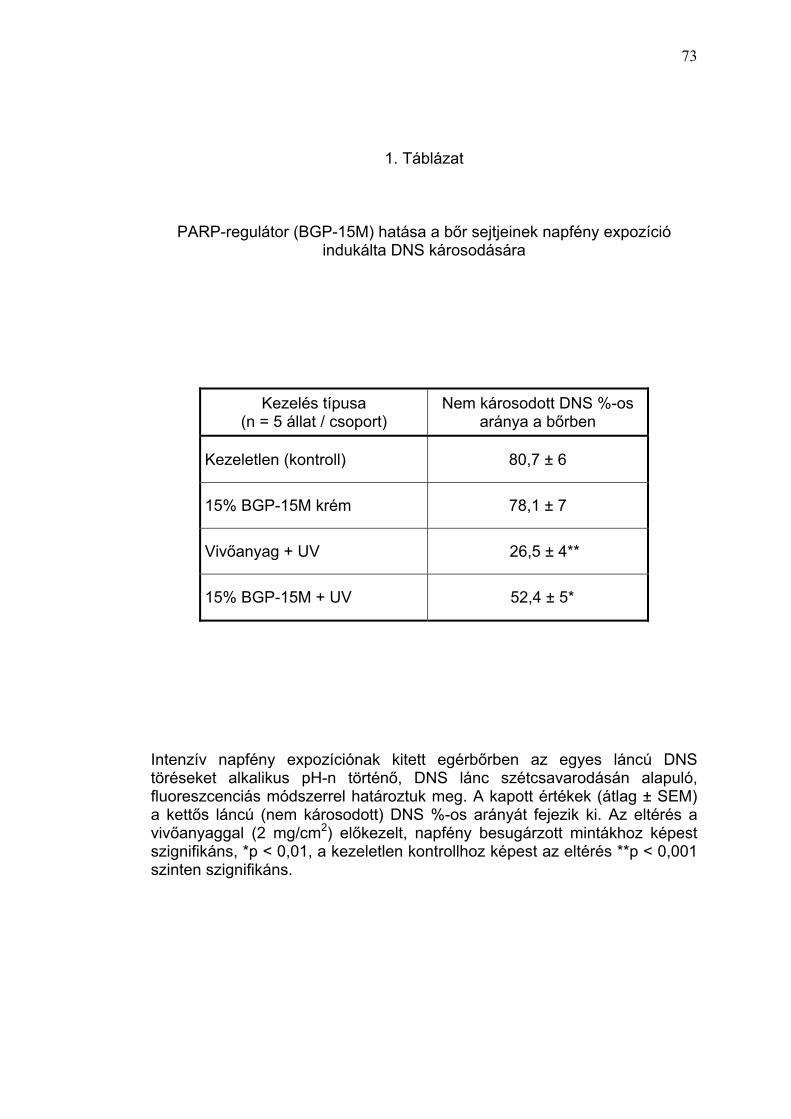
73
1. Táblázat
PARP-regulátor (BGP-15M) hatása a bőr sejtjeinek napfény expozíció
indukálta DNS károsodására
Kezelés típusa (n = 5 állat / csoport)
Nem károsodott DNS %-os aránya a bőrben
Kezeletlen (kontroll) 80,7 ± 6
15% BGP-15M krém 78,1 ± 7
Vivőanyag + UV 26,5 ± 4**
15% BGP-15M + UV 52,4 ± 5*
Intenzív napfény expozíciónak kitett egérbőrben az egyes láncú DNS töréseket alkalikus pH-n történő, DNS lánc szétcsavarodásán alapuló, fluoreszcenciás módszerrel határoztuk meg. A kapott értékek (átlag ± SEM) a kettős láncú (nem károsodott) DNS %-os arányát fejezik ki. Az eltérés a vivőanyaggal (2 mg/cm2) előkezelt, napfény besugárzott mintákhoz képest szignifikáns, *p < 0,01, a kezeletlen kontrollhoz képest az eltérés **p < 0,001 szinten szignifikáns.
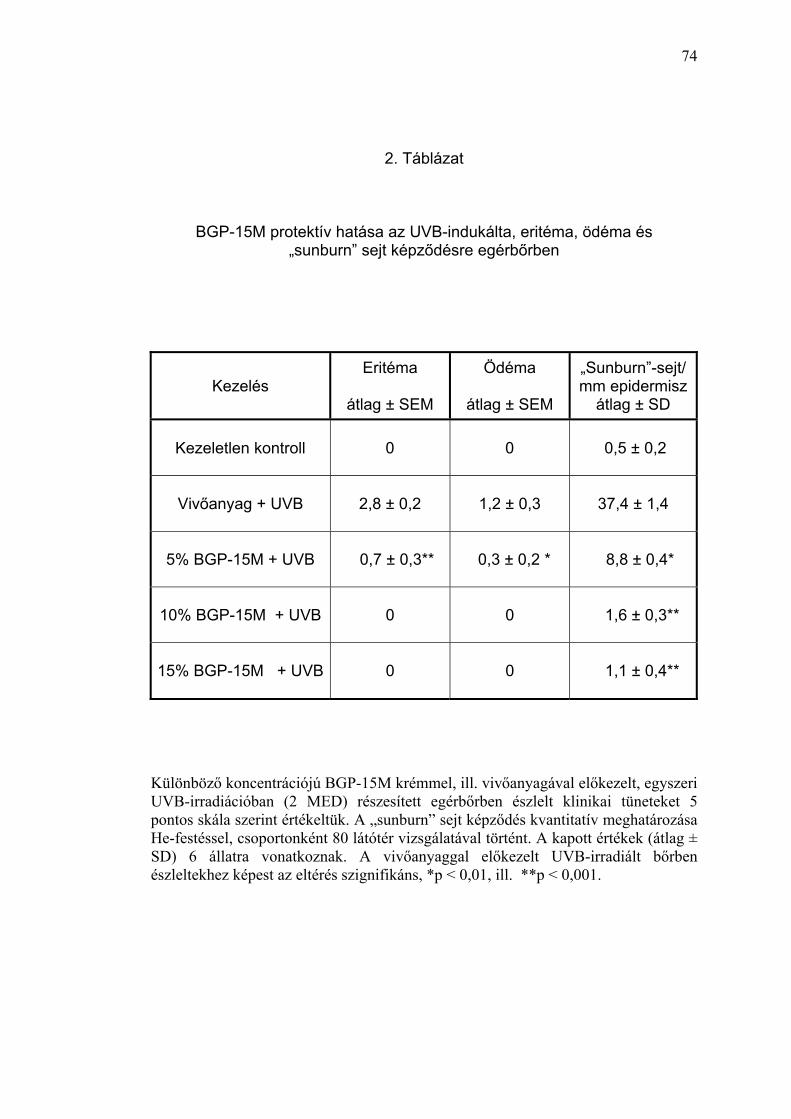
74
2. Táblázat
BGP-15M protektív hatása az UVB-indukálta, eritéma, ödéma és „sunburn” sejt képződésre egérbőrben
Különböző koncentrációjú BGP-15M krémmel, ill. vivőanyagával előkezelt, egyszeri UVB-irradiációban (2 MED) részesített egérbőrben észlelt klinikai tüneteket 5 pontos skála szerint értékeltük. A „sunburn” sejt képződés kvantitatív meghatározása He-festéssel, csoportonként 80 látótér vizsgálatával történt. A kapott értékek (átlag ± SD) 6 állatra vonatkoznak. A vivőanyaggal előkezelt UVB-irradiált bőrben észleltekhez képest az eltérés szignifikáns, *p < 0,01, ill. **p < 0,001.
Kezelés Eritéma
átlag ± SEM
Ödéma
átlag ± SEM
„Sunburn”-sejt/ mm epidermisz
átlag ± SD
Kezeletlen kontroll 0 0 0,5 ± 0,2
Vivőanyag + UVB 2,8 ± 0,2 1,2 ± 0,3 37,4 ± 1,4
5% BGP-15M + UVB 0,7 ± 0,3** 0,3 ± 0,2 * 8,8 ± 0,4*
10% BGP-15M + UVB 0 0 1,6 ± 0,3**
15% BGP-15M + UVB 0 0 1,1 ± 0,4**

75
3. Táblázat
BGP-15M krém kezelés hatása a bőr PARP-enzim poli ADP-ribozilációjára a bőrben
Kezelés formája
Jelintenzitás mértéke (átlag ± SEM)
Kezeletlen kontroll
2 ± 1
Vivőanyag + UV
51 ± 6**
10% BGP-15M krém + UV
23 ± 2*
Vivőanyag + UV
48 ± 4**
20% BGP-15M krém + UV
21 ± 3*
Vivőanyag + UV
47 ± 5**
PARP (pozitív kontroll)
52 ± 4
Különböző koncentrációjú BGP-15M krémmel, ill. vivőanyagával előkezelt (2mg/cm2), UV-besugárzott egérbőrben (n=5 állat/csoport) az ADP-riboziláció mértékét a Western-bloton kapott jelek intenzitásának kvantitatív meghatározásával („Image Tool image processing” program) végeztük. Az értékeket átlag ± SEM formájában fejeztük ki. Az eltérés a vivőanyaggal előkezelt, UV-besugárzott mintákhoz képest szignifikiáns, *p < 0,01. A kezeletlen kontrollhoz képest az eltérés **p < 0,001 szinten szignifikáns.
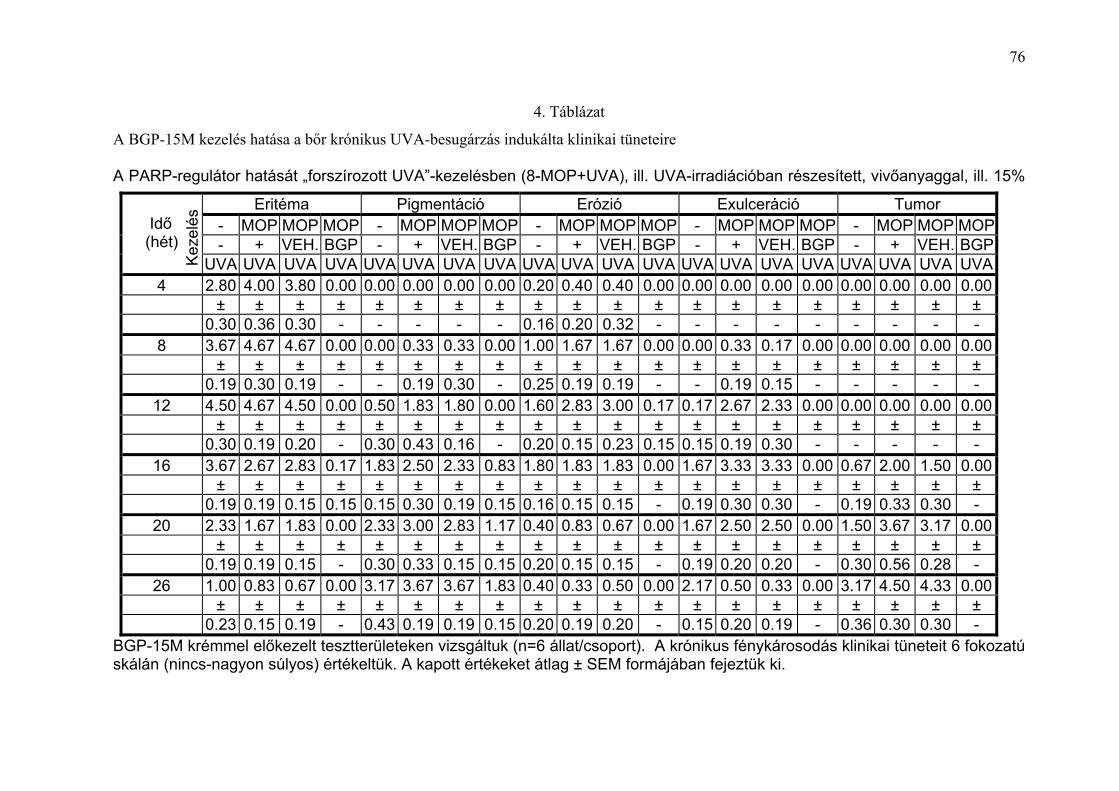
76
4. Táblázat
A BGP-15M kezelés hatása a bőr krónikus UVA-besugárzás indukálta klinikai tüneteire A PARP-regulátor hatását „forszírozott UVA”-kezelésben (8-MOP+UVA), ill. UVA-irradiációban részesített, vivőanyaggal, ill. 15%
BGP-15M krémmel előkezelt tesztterületeken vizsgáltuk (n=6 állat/csoport). A krónikus fénykárosodás klinikai tüneteit 6 fokozatú skálán (nincs-nagyon súlyos) értékeltük. A kapott értékeket átlag ± SEM formájában fejeztük ki.
Eritéma Pigmentáció Erózió Exulceráció Tumor - MOP MOP MOP - MOP MOP MOP - MOP MOP MOP - MOP MOP MOP - MOP MOP MOP - + VEH. BGP - + VEH. BGP - + VEH. BGP - + VEH. BGP - + VEH. BGP
Idő (hét)
UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA UVA 4 2.80 4.00 3.80 0.00 0.00 0.00 0.00 0.00 0.20 0.40 0.40 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± 0.30 0.36 0.30 - - - - - 0.16 0.20 0.32 - - - - - - - - - 8 3.67 4.67 4.67 0.00 0.00 0.33 0.33 0.00 1.00 1.67 1.67 0.00 0.00 0.33 0.17 0.00 0.00 0.00 0.00 0.00 ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± 0.19 0.30 0.19 - - 0.19 0.30 - 0.25 0.19 0.19 - - 0.19 0.15 - - - - -
12 4.50 4.67 4.50 0.00 0.50 1.83 1.80 0.00 1.60 2.83 3.00 0.17 0.17 2.67 2.33 0.00 0.00 0.00 0.00 0.00 ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± 0.30 0.19 0.20 - 0.30 0.43 0.16 - 0.20 0.15 0.23 0.15 0.15 0.19 0.30 - - - - -
16 3.67 2.67 2.83 0.17 1.83 2.50 2.33 0.83 1.80 1.83 1.83 0.00 1.67 3.33 3.33 0.00 0.67 2.00 1.50 0.00 ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± 0.19 0.19 0.15 0.15 0.15 0.30 0.19 0.15 0.16 0.15 0.15 - 0.19 0.30 0.30 - 0.19 0.33 0.30 -
20 2.33 1.67 1.83 0.00 2.33 3.00 2.83 1.17 0.40 0.83 0.67 0.00 1.67 2.50 2.50 0.00 1.50 3.67 3.17 0.00 ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± 0.19 0.19 0.15 - 0.30 0.33 0.15 0.15 0.20 0.15 0.15 - 0.19 0.20 0.20 - 0.30 0.56 0.28 -
26 1.00 0.83 0.67 0.00 3.17 3.67 3.67 1.83 0.40 0.33 0.50 0.00 2.17 0.50 0.33 0.00 3.17 4.50 4.33 0.00 ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± ± 0.23 0.15 0.19 - 0.43 0.19 0.19 0.15 0.20 0.19 0.20 - 0.15 0.20 0.19 - 0.36 0.30 0.30 -
Kez
elés
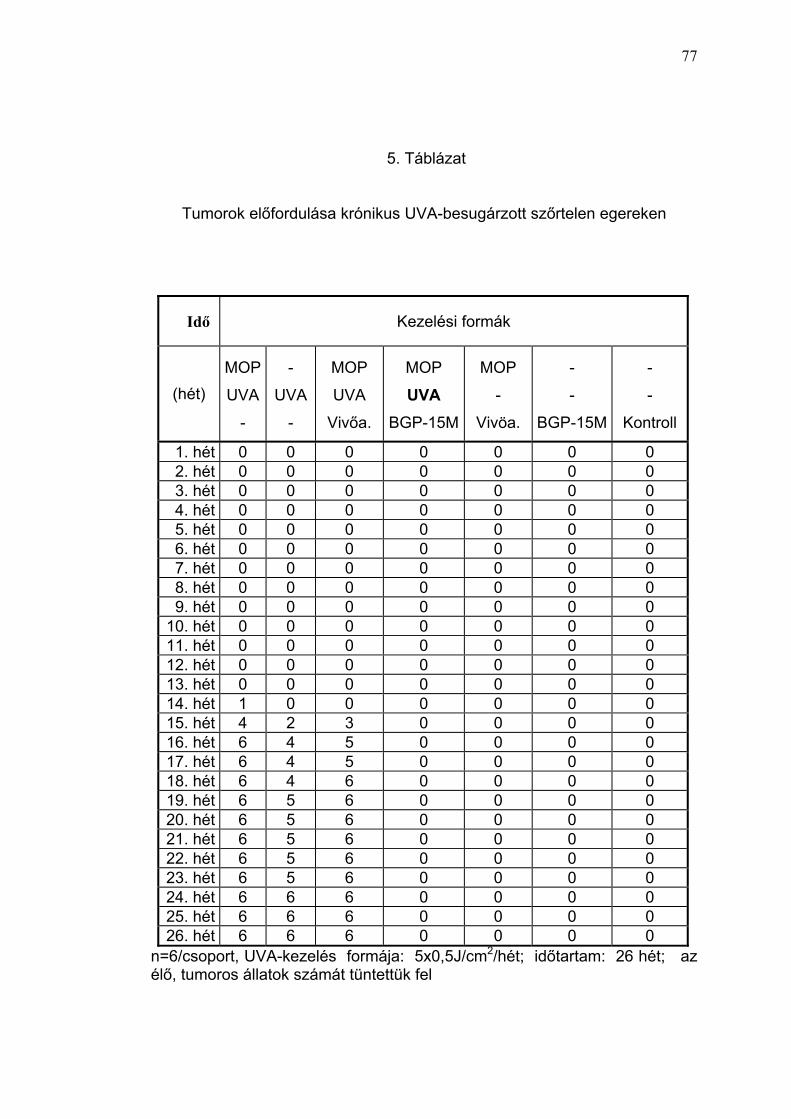
77
5. Táblázat
Tumorok előfordulása krónikus UVA-besugárzott szőrtelen egereken
n=6/csoport, UVA-kezelés formája: 5x0,5J/cm2/hét; időtartam: 26 hét; az élő, tumoros állatok számát tüntettük fel
Idő Kezelési formák
(hét)
MOP
UVA
-
-
UVA
-
MOP
UVA
Vivőa.
MOP
UVA BGP-15M
MOP
-
Vivöa.
-
-
BGP-15M
-
-
Kontroll
1. hét 0 0 0 0 0 0 0 2. hét 0 0 0 0 0 0 0 3. hét 0 0 0 0 0 0 0 4. hét 0 0 0 0 0 0 0 5. hét 0 0 0 0 0 0 0 6. hét 0 0 0 0 0 0 0 7. hét 0 0 0 0 0 0 0 8. hét 0 0 0 0 0 0 0 9. hét 0 0 0 0 0 0 0
10. hét 0 0 0 0 0 0 0 11. hét 0 0 0 0 0 0 0 12. hét 0 0 0 0 0 0 0 13. hét 0 0 0 0 0 0 0 14. hét 1 0 0 0 0 0 0 15. hét 4 2 3 0 0 0 0 16. hét 6 4 5 0 0 0 0 17. hét 6 4 5 0 0 0 0 18. hét 6 4 6 0 0 0 0 19. hét 6 5 6 0 0 0 0 20. hét 6 5 6 0 0 0 0 21. hét 6 5 6 0 0 0 0 22. hét 6 5 6 0 0 0 0 23. hét 6 5 6 0 0 0 0 24. hét 6 6 6 0 0 0 0 25. hét 6 6 6 0 0 0 0 26. hét 6 6 6 0 0 0 0
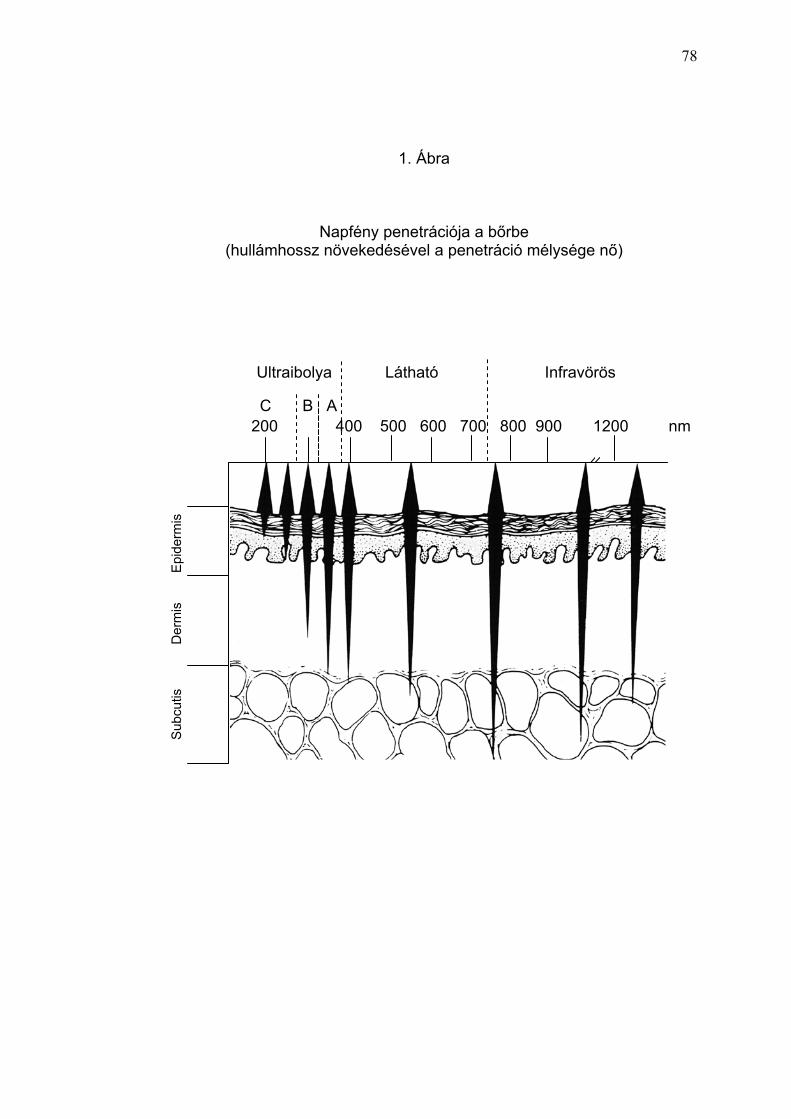
78
1. Ábra
Napfény penetrációja a bőrbe (hullámhossz növekedésével a penetráció mélysége nő)
800400200 500 600 700 900 1200
Ultraviolet Visible InfraredC B A
200 400 500 600 700 800 900 1200 nm C B A
S
ubcu
tis
D
erm
is
E
pide
rmis
Ultraibolya Látható Infravörös
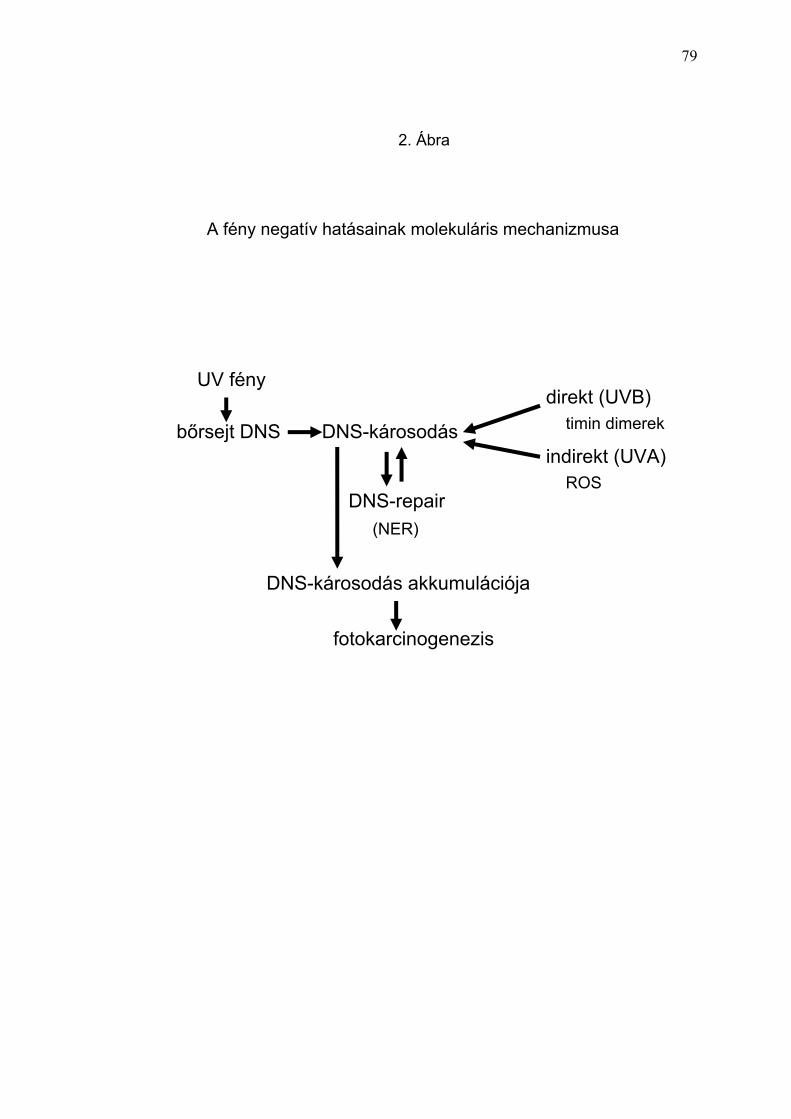
79
2. Ábra
UV fény
DNS-károsodás
direkt (UVB) timin dimerek
indirekt (UVA)ROS
bőrsejt DNS
DNS-repair (NER)
DNS-károsodás akkumulációja
fotokarcinogenezis
A fény negatív hatásainak molekuláris mechanizmusa

80
3. Ábra
Hairless egér
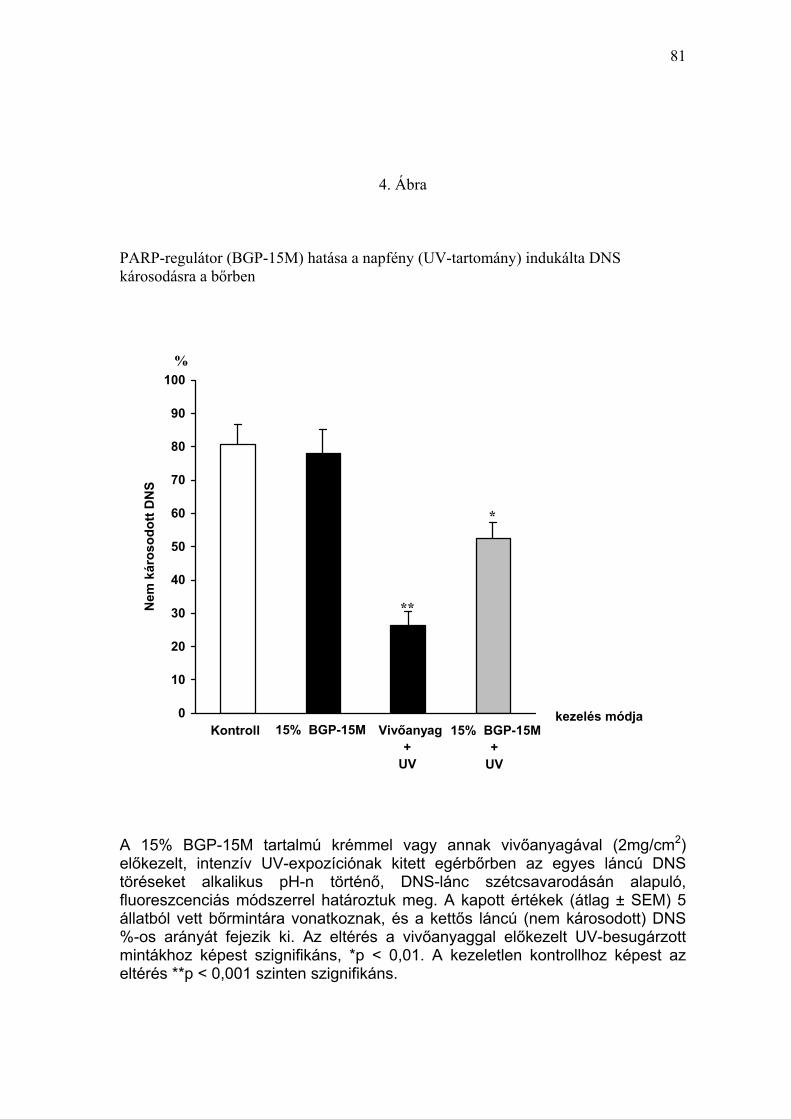
81
4. Ábra PARP-regulátor (BGP-15M) hatása a napfény (UV-tartomány) indukálta DNS károsodásra a bőrben A 15% BGP-15M tartalmú krémmel vagy annak vivőanyagával (2mg/cm2) előkezelt, intenzív UV-expozíciónak kitett egérbőrben az egyes láncú DNS töréseket alkalikus pH-n történő, DNS-lánc szétcsavarodásán alapuló, fluoreszcenciás módszerrel határoztuk meg. A kapott értékek (átlag ± SEM) 5 állatból vett bőrmintára vonatkoznak, és a kettős láncú (nem károsodott) DNS %-os arányát fejezik ki. Az eltérés a vivőanyaggal előkezelt UV-besugárzott mintákhoz képest szignifikáns, *p < 0,01. A kezeletlen kontrollhoz képest az eltérés **p < 0,001 szinten szignifikáns.
0
10
20
30
40
50
60
70
80
90
100
kezelés módja
Nem
kár
osod
ott D
NS
Kontroll 15% BGP-15M 15% BGP-15M+
UV
Vivőanyag+
UV
**
*
%
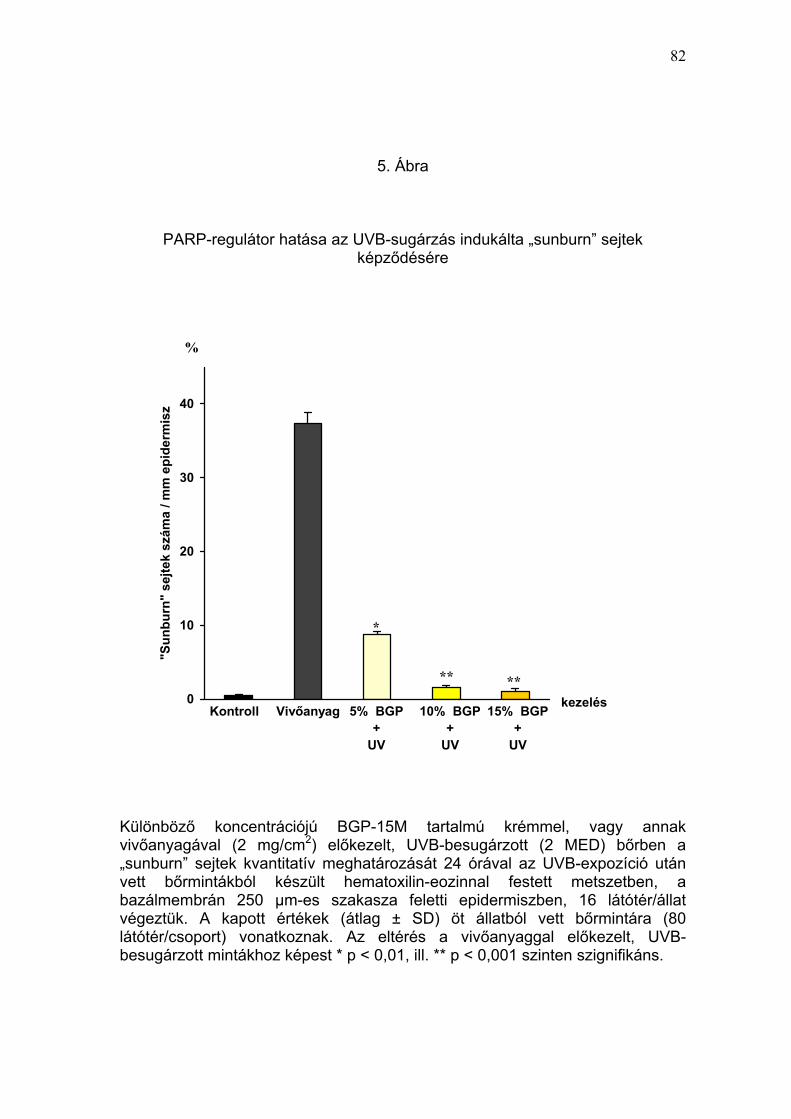
82
5. Ábra
PARP-regulátor hatása az UVB-sugárzás indukálta „sunburn” sejtek képződésére
Különböző koncentrációjú BGP-15M tartalmú krémmel, vagy annak vivőanyagával (2 mg/cm2) előkezelt, UVB-besugárzott (2 MED) bőrben a „sunburn” sejtek kvantitatív meghatározását 24 órával az UVB-expozíció után vett bőrmintákból készült hematoxilin-eozinnal festett metszetben, a bazálmembrán 250 µm-es szakasza feletti epidermiszben, 16 látótér/állat végeztük. A kapott értékek (átlag ± SD) öt állatból vett bőrmintára (80 látótér/csoport) vonatkoznak. Az eltérés a vivőanyaggal előkezelt, UVB-besugárzott mintákhoz képest * p < 0,01, ill. ** p < 0,001 szinten szignifikáns.
0
10
20
30
40
kezelés
"Sun
burn
" se
jtek
szám
a / m
m e
pide
rmis
z
Kontroll 5% BGP+
UV
Vivőanyag
**
*
%
**
10% BGP+
UV
15% BGP+
UV
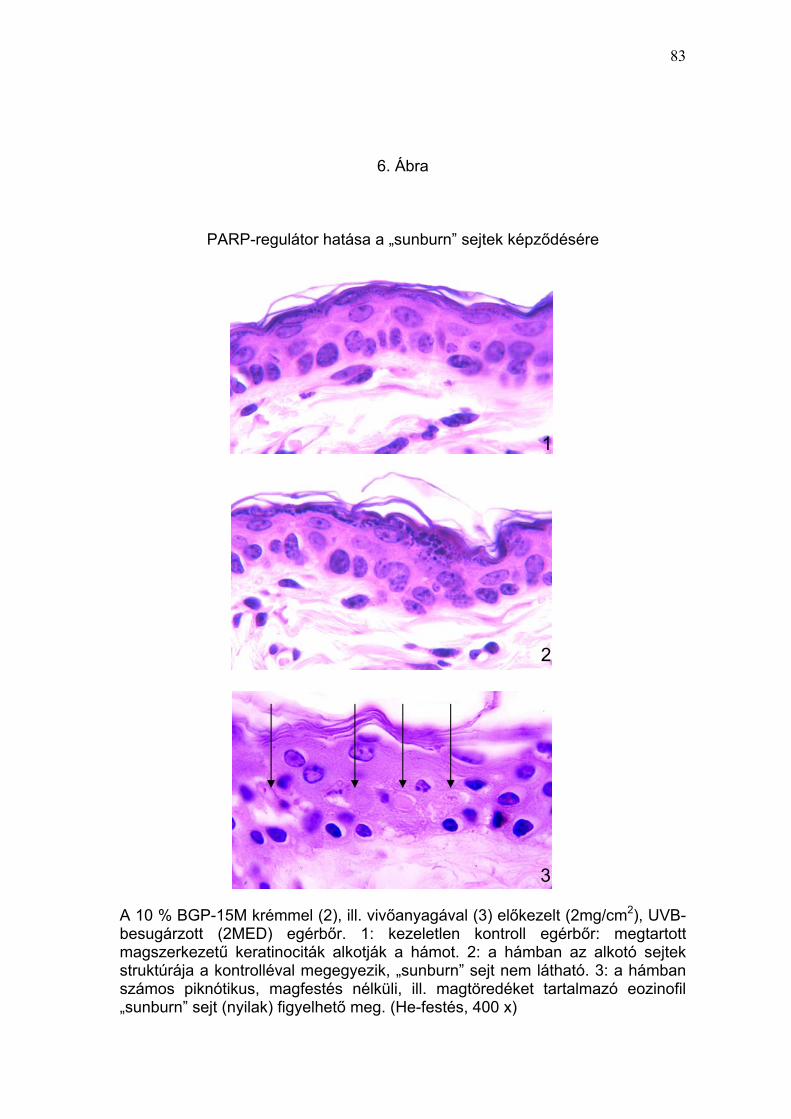
83
6. Ábra
PARP-regulátor hatása a „sunburn” sejtek képződésére A 10 % BGP-15M krémmel (2), ill. vivőanyagával (3) előkezelt (2mg/cm2), UVB-besugárzott (2MED) egérbőr. 1: kezeletlen kontroll egérbőr: megtartott magszerkezetű keratinociták alkotják a hámot. 2: a hámban az alkotó sejtek struktúrája a kontrolléval megegyezik, „sunburn” sejt nem látható. 3: a hámban számos piknótikus, magfestés nélküli, ill. magtöredéket tartalmazó eozinofil „sunburn” sejt (nyilak) figyelhető meg. (He-festés, 400 x)
1
2
3
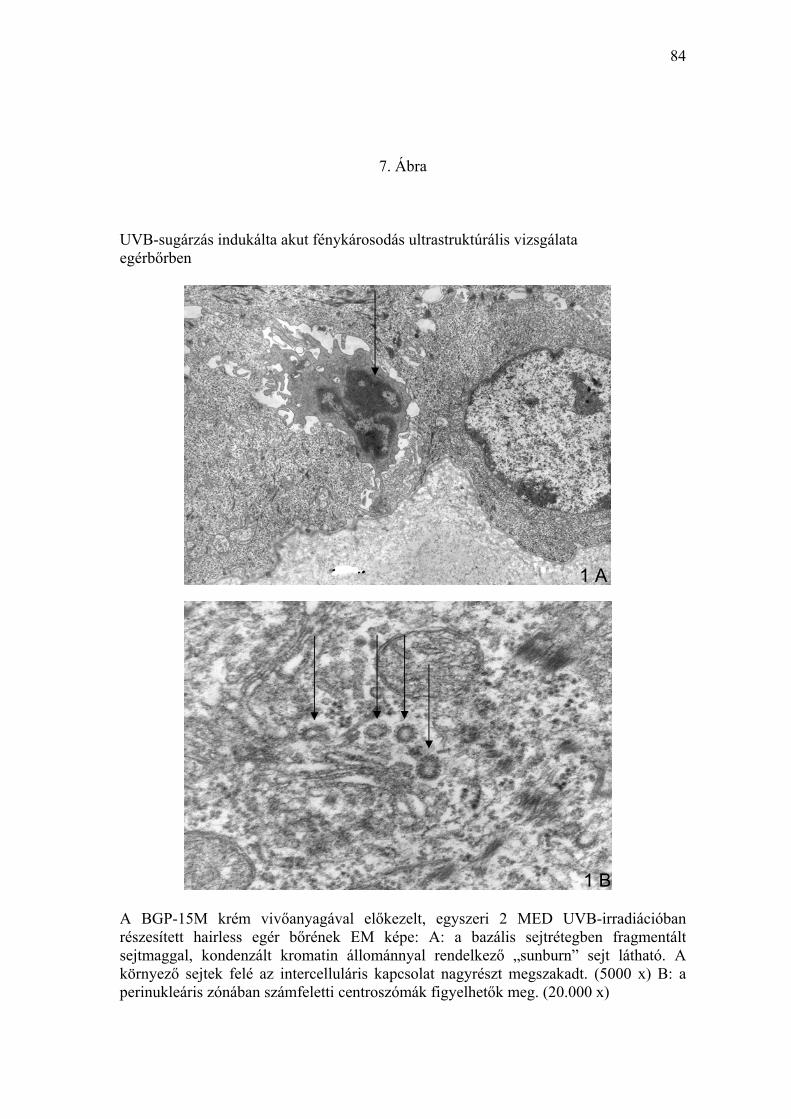
84
7. Ábra UVB-sugárzás indukálta akut fénykárosodás ultrastruktúrális vizsgálata egérbőrben A BGP-15M krém vivőanyagával előkezelt, egyszeri 2 MED UVB-irradiációban részesített hairless egér bőrének EM képe: A: a bazális sejtrétegben fragmentált sejtmaggal, kondenzált kromatin állománnyal rendelkező „sunburn” sejt látható. A környező sejtek felé az intercelluláris kapcsolat nagyrészt megszakadt. (5000 x) B: a perinukleáris zónában számfeletti centroszómák figyelhetők meg. (20.000 x)
1 A
1 B
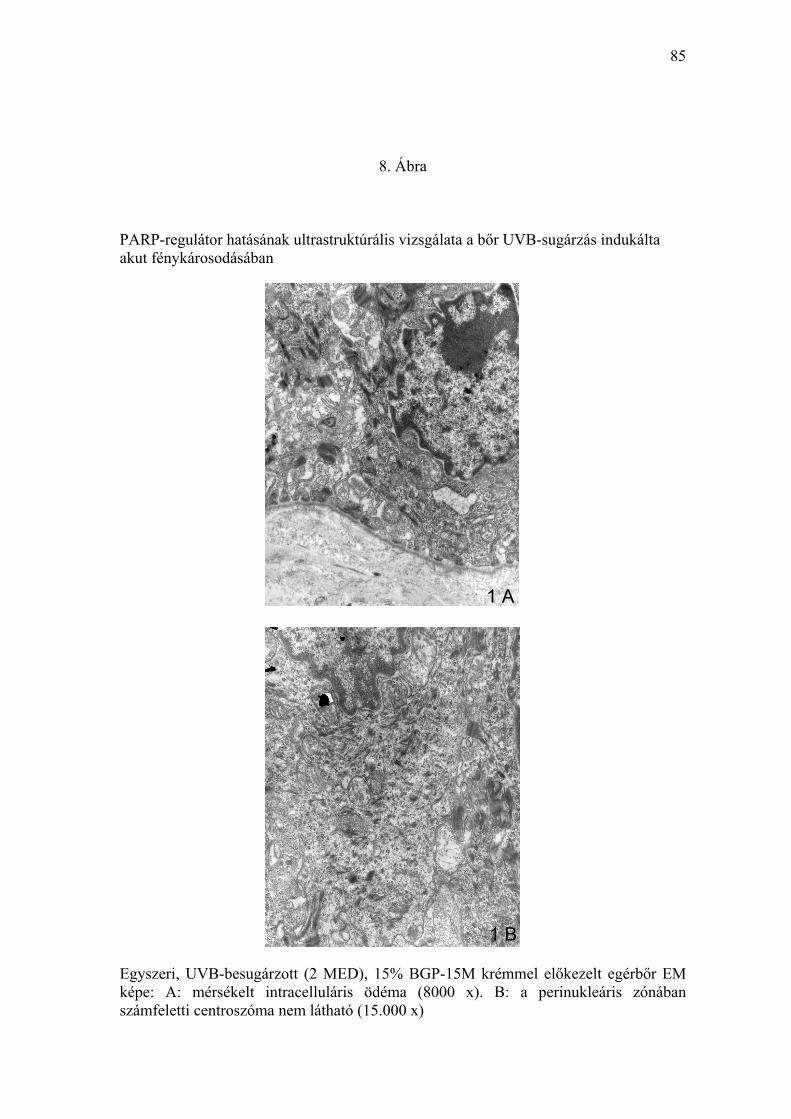
85
8. Ábra PARP-regulátor hatásának ultrastruktúrális vizsgálata a bőr UVB-sugárzás indukálta akut fénykárosodásában Egyszeri, UVB-besugárzott (2 MED), 15% BGP-15M krémmel előkezelt egérbőr EM képe: A: mérsékelt intracelluláris ödéma (8000 x). B: a perinukleáris zónában számfeletti centroszóma nem látható (15.000 x)
1 A
1 B

86
9. Ábra
A PARP ADP-ribozilációjának meghatározása bőrben Western blot technikával
A PARP enzim ADP-ribozilációját a BGP-15M tartalmú krémmel, vagy annak vivőanyagával (2mg/cm2) előkezelt, napfény-expozíciónak kitett bőrből, közvetlenül a besugárzás után vett mintákban Western-blot analízissel határoztuk meg. Az ADP-riboziláció mértékét a Western-bloton kapott jelek intenzitása fejezi ki: kezeletlen, nem besugárzott (natív) egérbőr (1. oszlop); pozitív kontroll: ADP-ribozilált PARP (2. oszlop); vivőanyaggal előkezelt, UV-besugárzott egérbőr (3. és 5. oszlop); 10% BGP-15M krémmel előkezelt és UV-besugárzott (4. oszlop), valamint 20% BGP-15M krémmel előkezelt és UV-besugárzott bőr (6. oszlop).

87
10. Ábra
A PARP-regulátor hatása az egyszeri UVB-expozíció klinika tüneteire
BGP-15M tartalmú krémmel (5%, 10%, 15% és 20%) (A), ill. vivőanyagával (B) előkezelt (2mg/cm2), UVB-irradiált (2 MED) akut fénykárosodás klinikai tünetei az expozíció után 24 órával: 1-2 A: dermatitis solaris klinikai képe, 1B: enyhe eritéma, ödéma nélkül, 2B: tünetmentes bőr
1
A
B
2
A
B
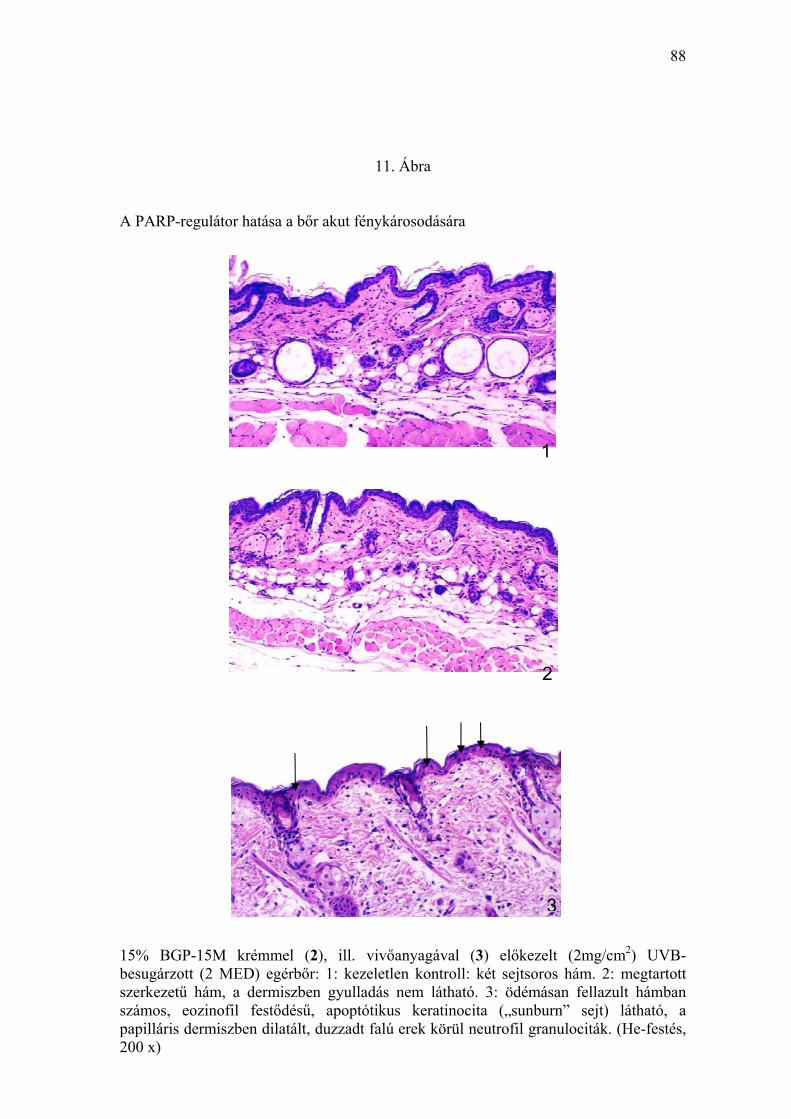
88
11. Ábra A PARP-regulátor hatása a bőr akut fénykárosodására 15% BGP-15M krémmel (2), ill. vivőanyagával (3) előkezelt (2mg/cm2) UVB-besugárzott (2 MED) egérbőr: 1: kezeletlen kontroll: két sejtsoros hám. 2: megtartott szerkezetű hám, a dermiszben gyulladás nem látható. 3: ödémásan fellazult hámban számos, eozinofil festődésű, apoptótikus keratinocita („sunburn” sejt) látható, a papilláris dermiszben dilatált, duzzadt falú erek körül neutrofil granulociták. (He-festés, 200 x)
1
3
2

89
12. Ábra A PARP-regulátor antieritematogén hatásának összehasonlító vizsgálata ismert fényvédő faktorú készítménnyel PARP-regulátor antieritematogén hatásának összehasonlító vizsgálatát ismert fényvédő faktorú (SPF: 30, UVB/UVA), „AS” jelzésű készítménnyel végeztük (n=6 állat). 10% BGP-15M krémmel (B), ill. „AS” fényvédővel (A) előkezelt (2mg/cm2) és UVB-besugárzott (2 MED), valamint előkezelés nélkül UVB-irradiált (U) hairless egérbőr. 2-3 U: dermatitis solaris klinikai tünetei. 1-3 A,B: UVB-expozíció klinikai tünetet nem okozott. 1A-1B: 10% BGP-15M krém fotoprotektív hatása megegyezik az „AS” jelzésű, 30-as fényvédő faktorú készítményével.
1 2
B
A U
A
3
U
B 10% BGP-15M
+ UVB
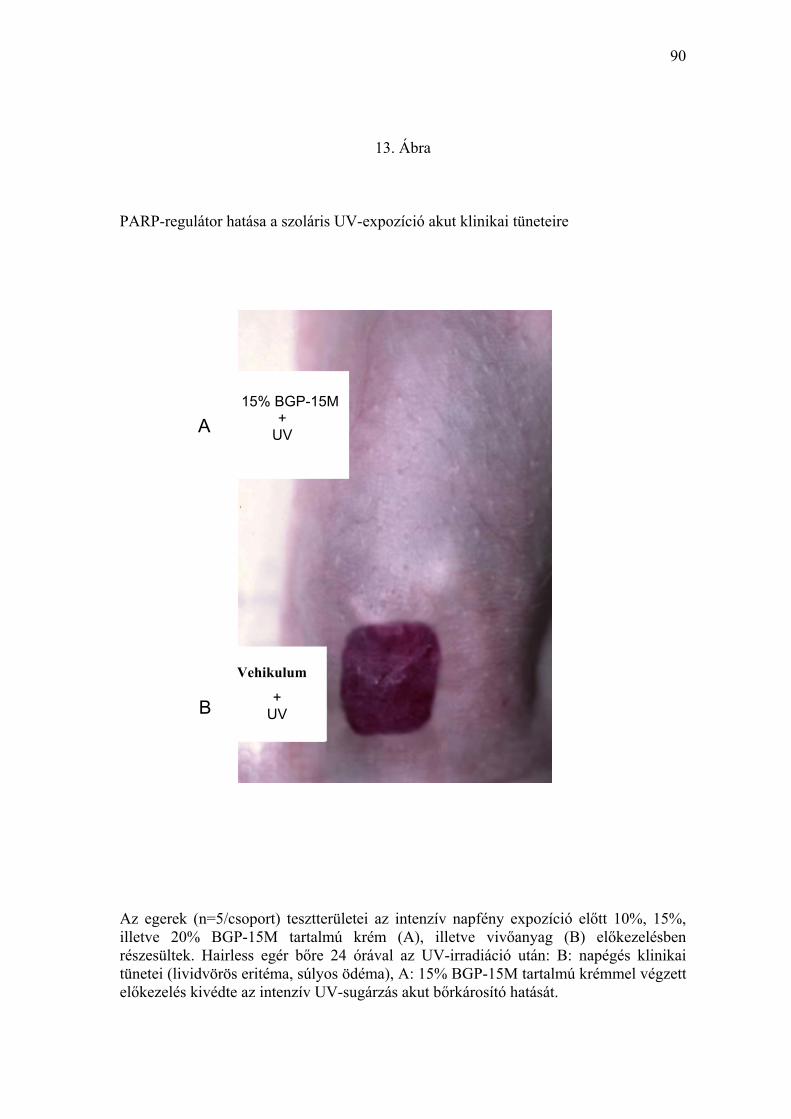
90
13. Ábra
PARP-regulátor hatása a szoláris UV-expozíció akut klinikai tüneteire Az egerek (n=5/csoport) tesztterületei az intenzív napfény expozíció előtt 10%, 15%, illetve 20% BGP-15M tartalmú krém (A), illetve vivőanyag (B) előkezelésben részesültek. Hairless egér bőre 24 órával az UV-irradiáció után: B: napégés klinikai tünetei (lividvörös eritéma, súlyos ödéma), A: 15% BGP-15M tartalmú krémmel végzett előkezelés kivédte az intenzív UV-sugárzás akut bőrkárosító hatását.
A
B
15% BGP-15M
+ UV
Vehikulum
+ UV
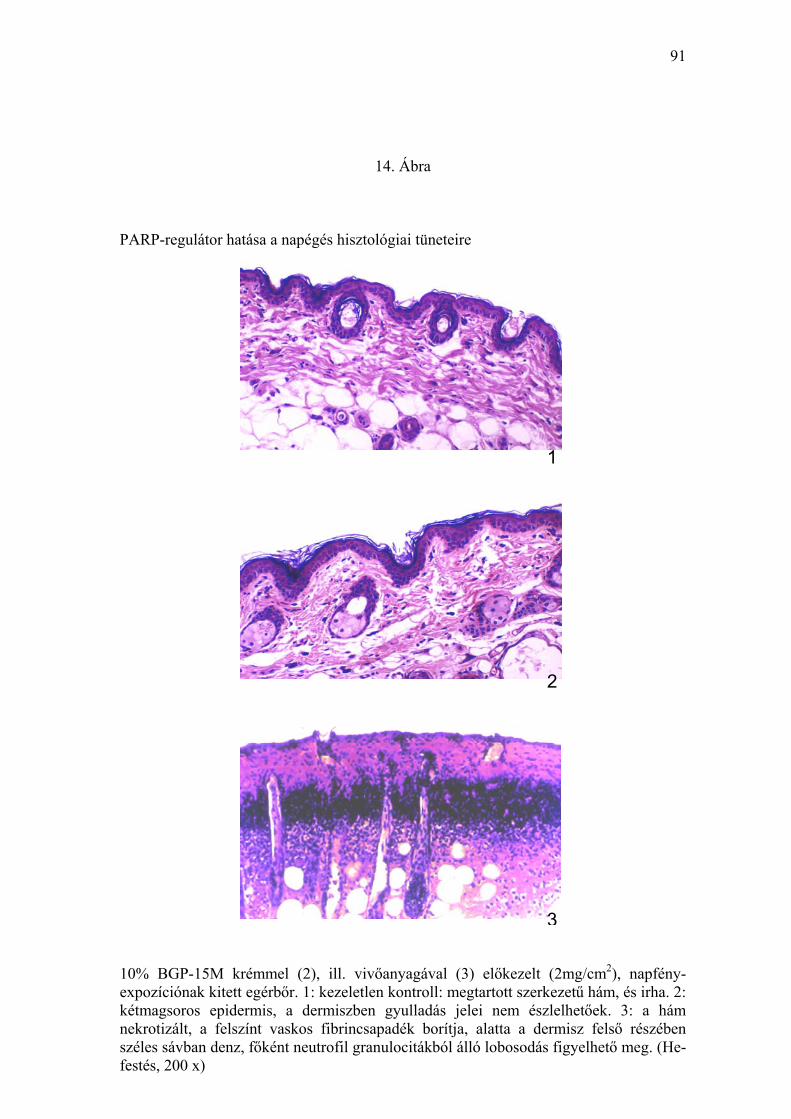
91
14. Ábra PARP-regulátor hatása a napégés hisztológiai tüneteire 10% BGP-15M krémmel (2), ill. vivőanyagával (3) előkezelt (2mg/cm2), napfény-expozíciónak kitett egérbőr. 1: kezeletlen kontroll: megtartott szerkezetű hám, és irha. 2: kétmagsoros epidermis, a dermiszben gyulladás jelei nem észlelhetőek. 3: a hám nekrotizált, a felszínt vaskos fibrincsapadék borítja, alatta a dermisz felső részében széles sávban denz, főként neutrofil granulocitákból álló lobosodás figyelhető meg. (He-festés, 200 x)
1
2
3
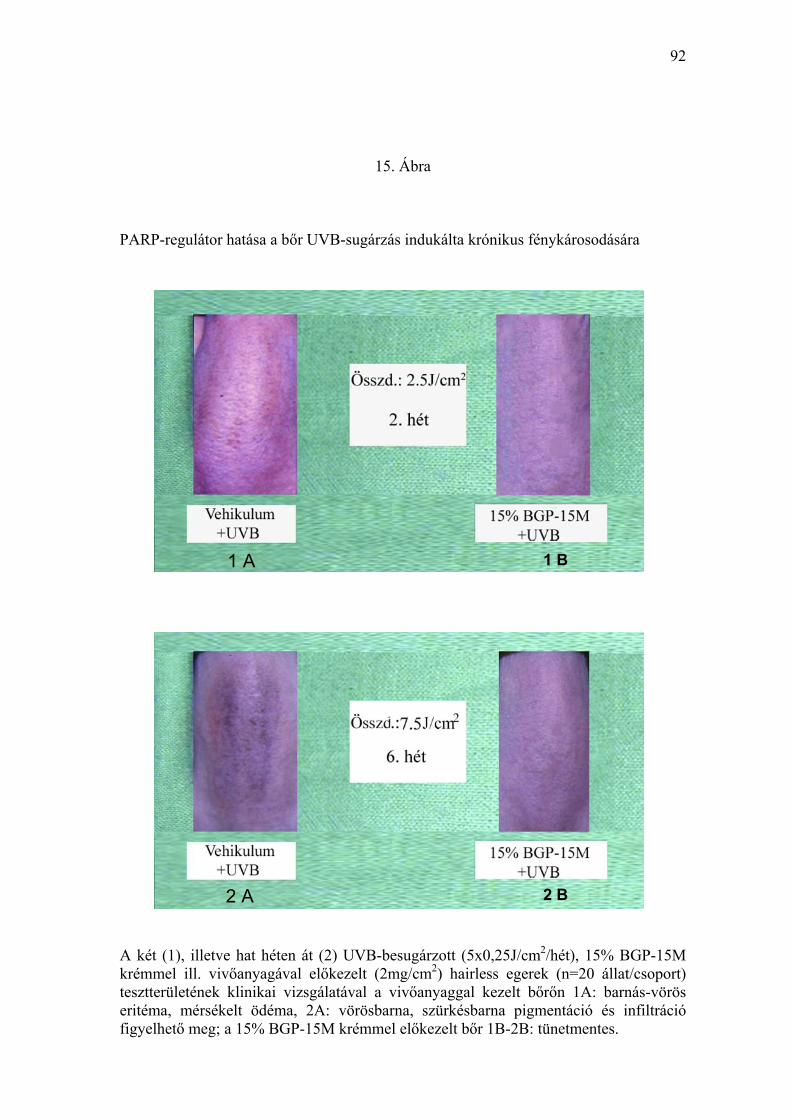
92
15. Ábra PARP-regulátor hatása a bőr UVB-sugárzás indukálta krónikus fénykárosodására A két (1), illetve hat héten át (2) UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel ill. vivőanyagával előkezelt (2mg/cm2) hairless egerek (n=20 állat/csoport) tesztterületének klinikai vizsgálatával a vivőanyaggal kezelt bőrőn 1A: barnás-vörös eritéma, mérsékelt ödéma, 2A: vörösbarna, szürkésbarna pigmentáció és infiltráció figyelhető meg; a 15% BGP-15M krémmel előkezelt bőr 1B-2B: tünetmentes.
1 A 1 B
2 A 2 B
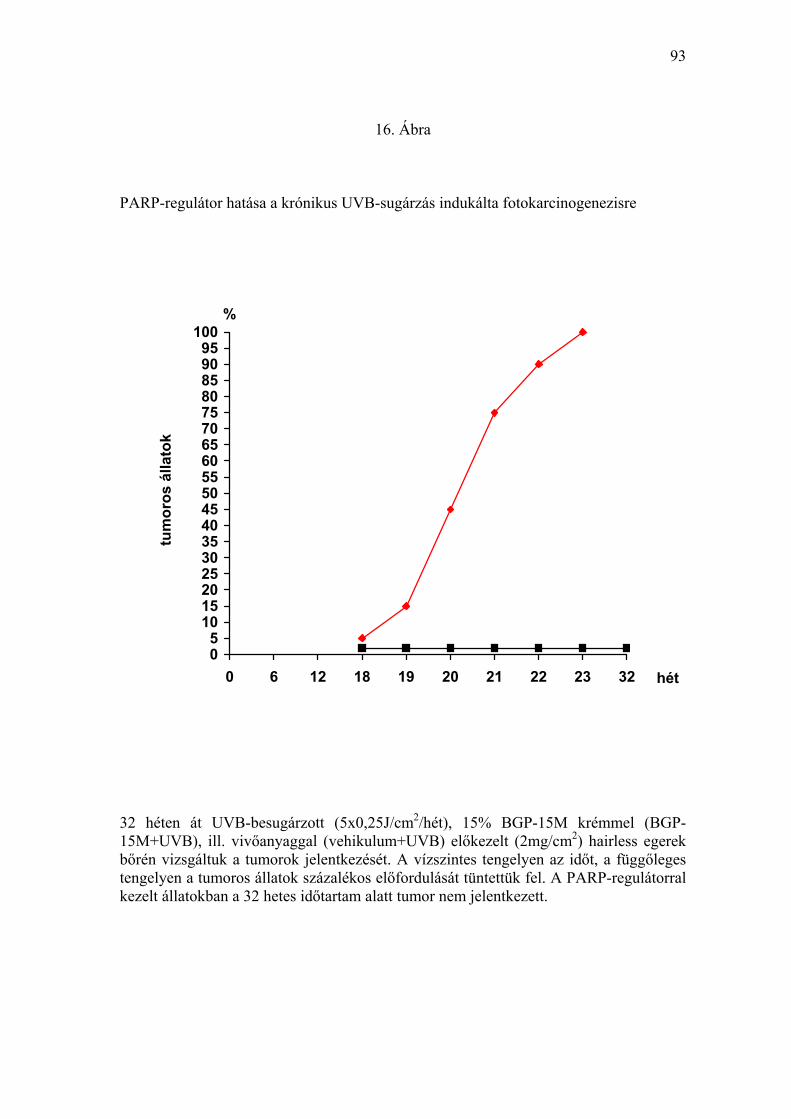
93
16. Ábra PARP-regulátor hatása a krónikus UVB-sugárzás indukálta fotokarcinogenezisre
32 héten át UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel (BGP-15M+UVB), ill. vivőanyaggal (vehikulum+UVB) előkezelt (2mg/cm2) hairless egerek bőrén vizsgáltuk a tumorok jelentkezését. A vízszintes tengelyen az időt, a függőleges tengelyen a tumoros állatok százalékos előfordulását tüntettük fel. A PARP-regulátorral kezelt állatokban a 32 hetes időtartam alatt tumor nem jelentkezett.
05
101520253035404550556065707580859095
100
0 6 12 18 19 20 21 22 23 32 hét
tum
oros
álla
tok
%
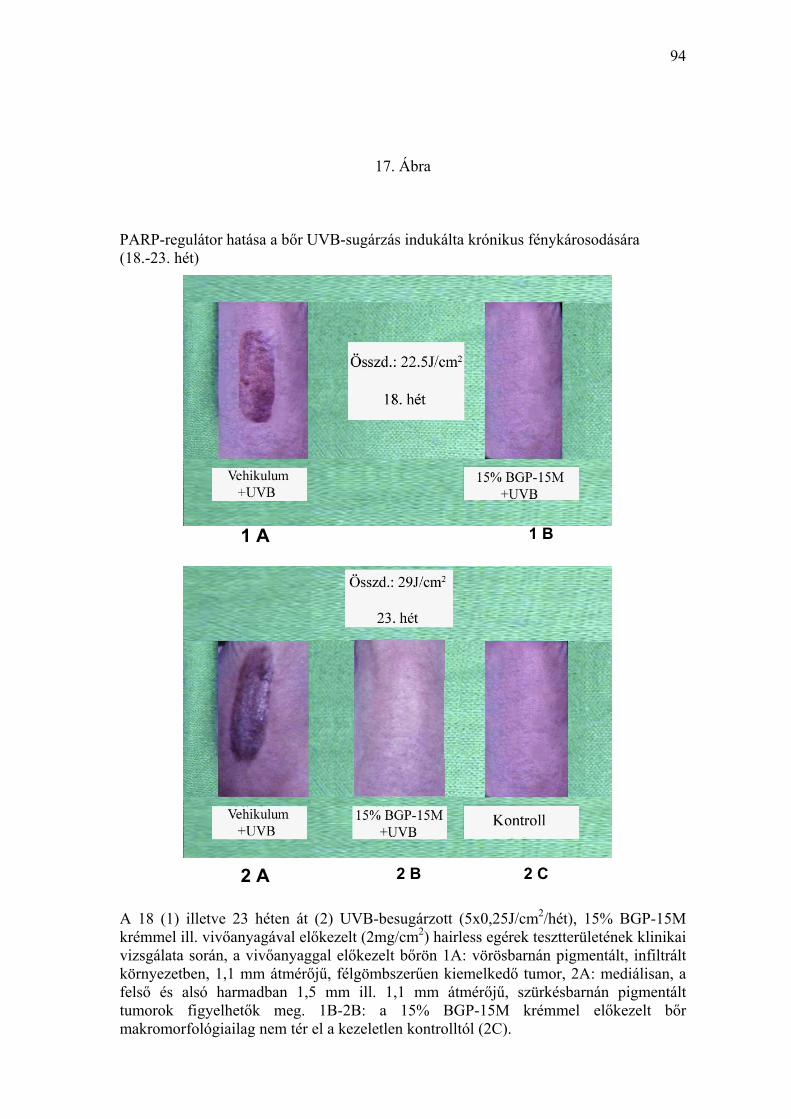
94
17. Ábra PARP-regulátor hatása a bőr UVB-sugárzás indukálta krónikus fénykárosodására (18.-23. hét) A 18 (1) illetve 23 héten át (2) UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel ill. vivőanyagával előkezelt (2mg/cm2) hairless egérek tesztterületének klinikai vizsgálata során, a vivőanyaggal előkezelt bőrön 1A: vörösbarnán pigmentált, infiltrált környezetben, 1,1 mm átmérőjű, félgömbszerűen kiemelkedő tumor, 2A: mediálisan, a felső és alsó harmadban 1,5 mm ill. 1,1 mm átmérőjű, szürkésbarnán pigmentált tumorok figyelhetők meg. 1B-2B: a 15% BGP-15M krémmel előkezelt bőr makromorfológiailag nem tér el a kezeletlen kontrolltól (2C).
1 A 1 B
2 A 2 C 2 B
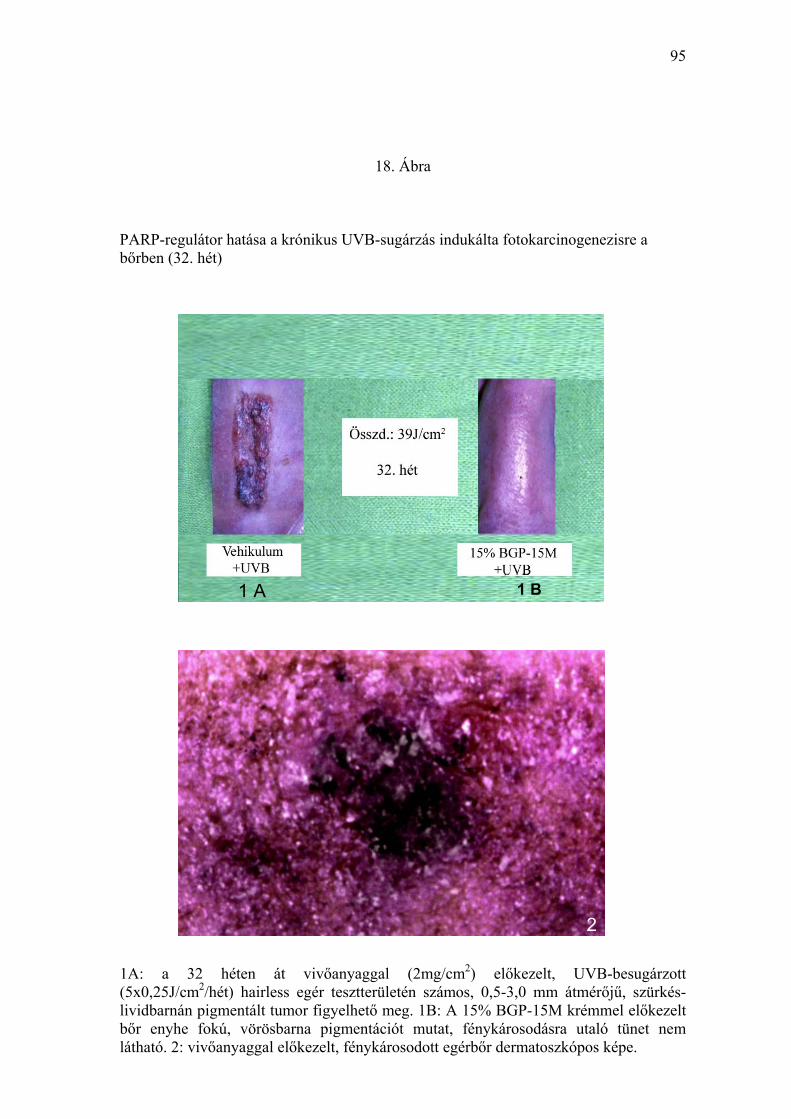
95
18. Ábra PARP-regulátor hatása a krónikus UVB-sugárzás indukálta fotokarcinogenezisre a bőrben (32. hét) 1A: a 32 héten át vivőanyaggal (2mg/cm2) előkezelt, UVB-besugárzott (5x0,25J/cm2/hét) hairless egér tesztterületén számos, 0,5-3,0 mm átmérőjű, szürkés-lividbarnán pigmentált tumor figyelhető meg. 1B: A 15% BGP-15M krémmel előkezelt bőr enyhe fokú, vörösbarna pigmentációt mutat, fénykárosodásra utaló tünet nem látható. 2: vivőanyaggal előkezelt, fénykárosodott egérbőr dermatoszkópos képe.
1 A
2
1 B
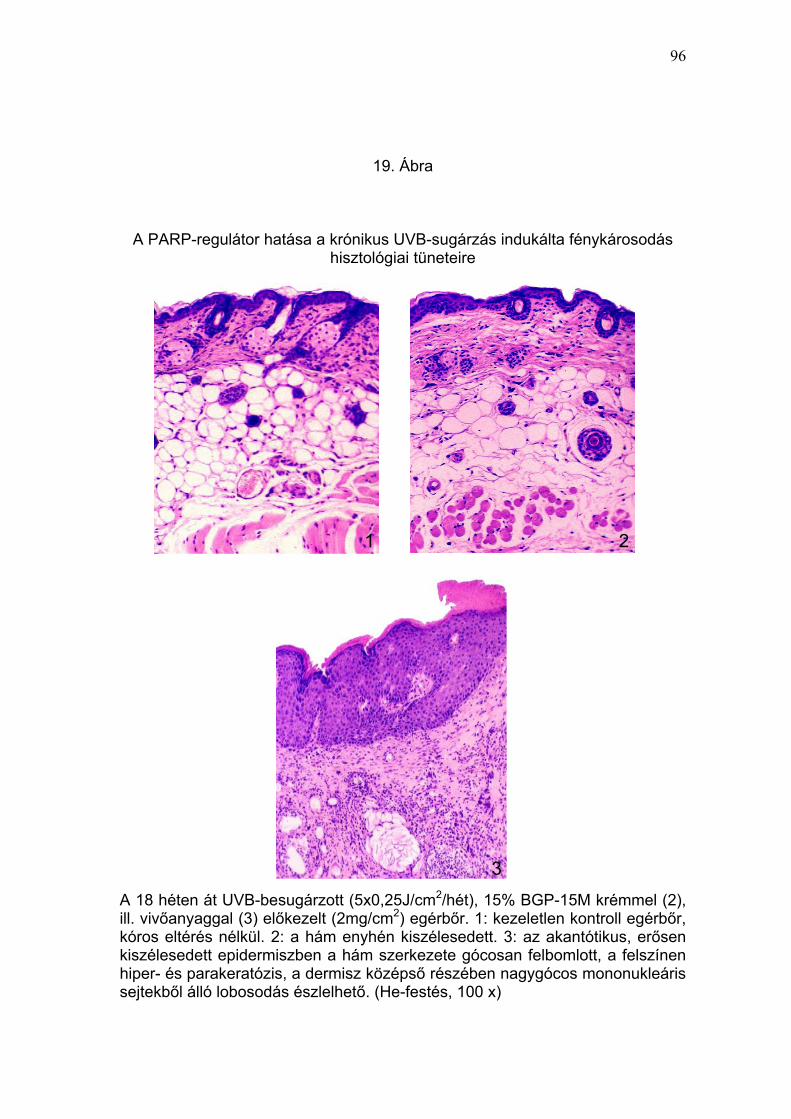
96
19. Ábra
A PARP-regulátor hatása a krónikus UVB-sugárzás indukálta fénykárosodás hisztológiai tüneteire
A 18 héten át UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel (2), ill. vivőanyaggal (3) előkezelt (2mg/cm2) egérbőr. 1: kezeletlen kontroll egérbőr, kóros eltérés nélkül. 2: a hám enyhén kiszélesedett. 3: az akantótikus, erősen kiszélesedett epidermiszben a hám szerkezete gócosan felbomlott, a felszínen hiper- és parakeratózis, a dermisz középső részében nagygócos mononukleáris sejtekből álló lobosodás észlelhető. (He-festés, 100 x)
1 2
3
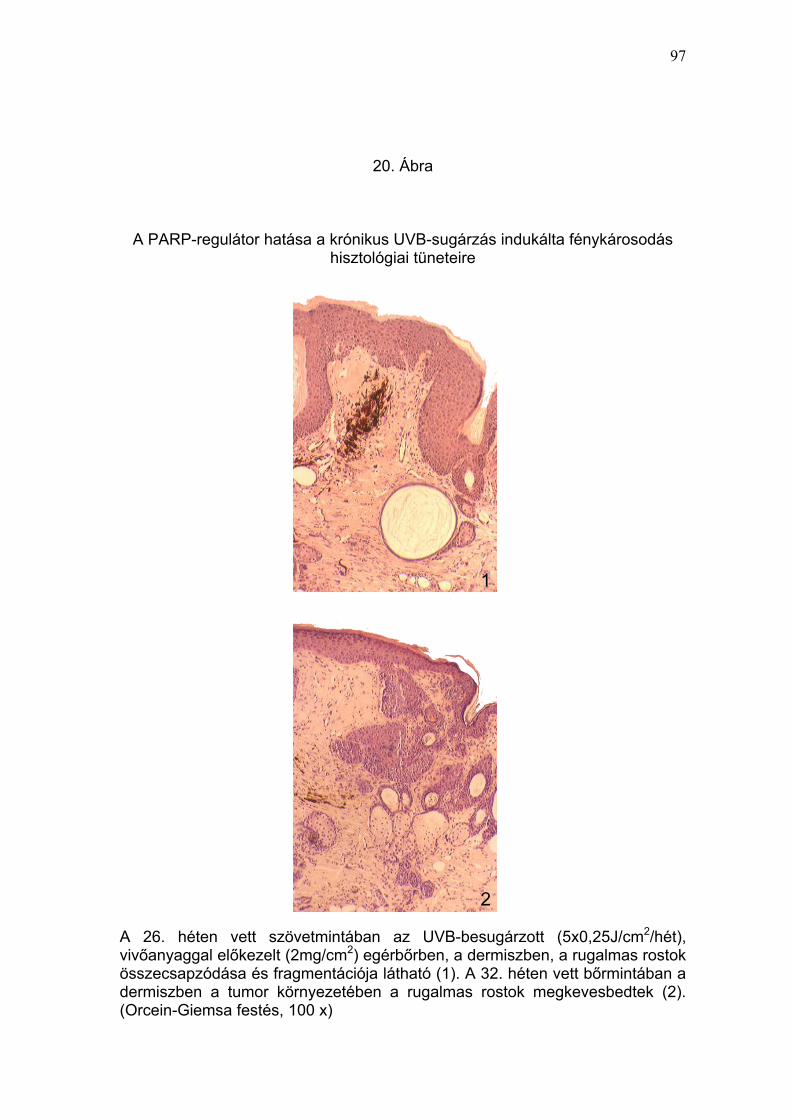
97
20. Ábra
A PARP-regulátor hatása a krónikus UVB-sugárzás indukálta fénykárosodás
hisztológiai tüneteire A 26. héten vett szövetmintában az UVB-besugárzott (5x0,25J/cm2/hét), vivőanyaggal előkezelt (2mg/cm2) egérbőrben, a dermiszben, a rugalmas rostok összecsapzódása és fragmentációja látható (1). A 32. héten vett bőrmintában a dermiszben a tumor környezetében a rugalmas rostok megkevesbedtek (2). (Orcein-Giemsa festés, 100 x)
2
1
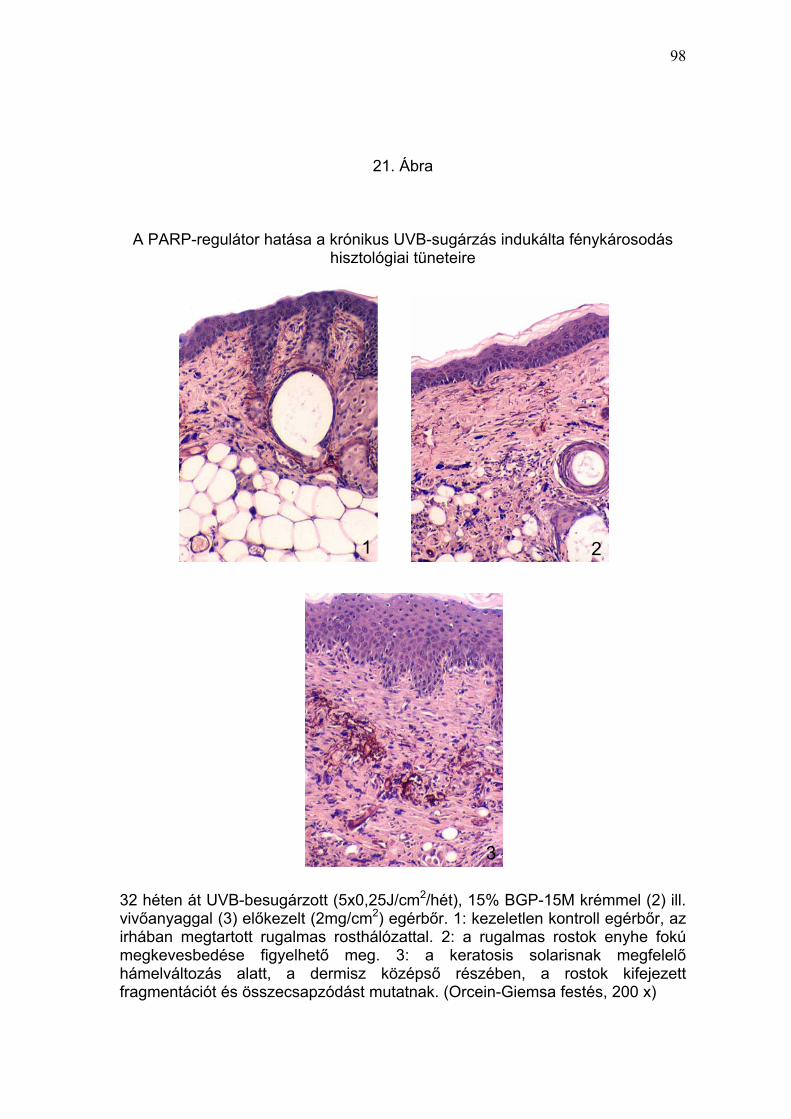
98
21. Ábra
A PARP-regulátor hatása a krónikus UVB-sugárzás indukálta fénykárosodás
hisztológiai tüneteire
32 héten át UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel (2) ill. vivőanyaggal (3) előkezelt (2mg/cm2) egérbőr. 1: kezeletlen kontroll egérbőr, az irhában megtartott rugalmas rosthálózattal. 2: a rugalmas rostok enyhe fokú megkevesbedése figyelhető meg. 3: a keratosis solarisnak megfelelő hámelváltozás alatt, a dermisz középső részében, a rostok kifejezett fragmentációt és összecsapzódást mutatnak. (Orcein-Giemsa festés, 200 x)
3
2 1
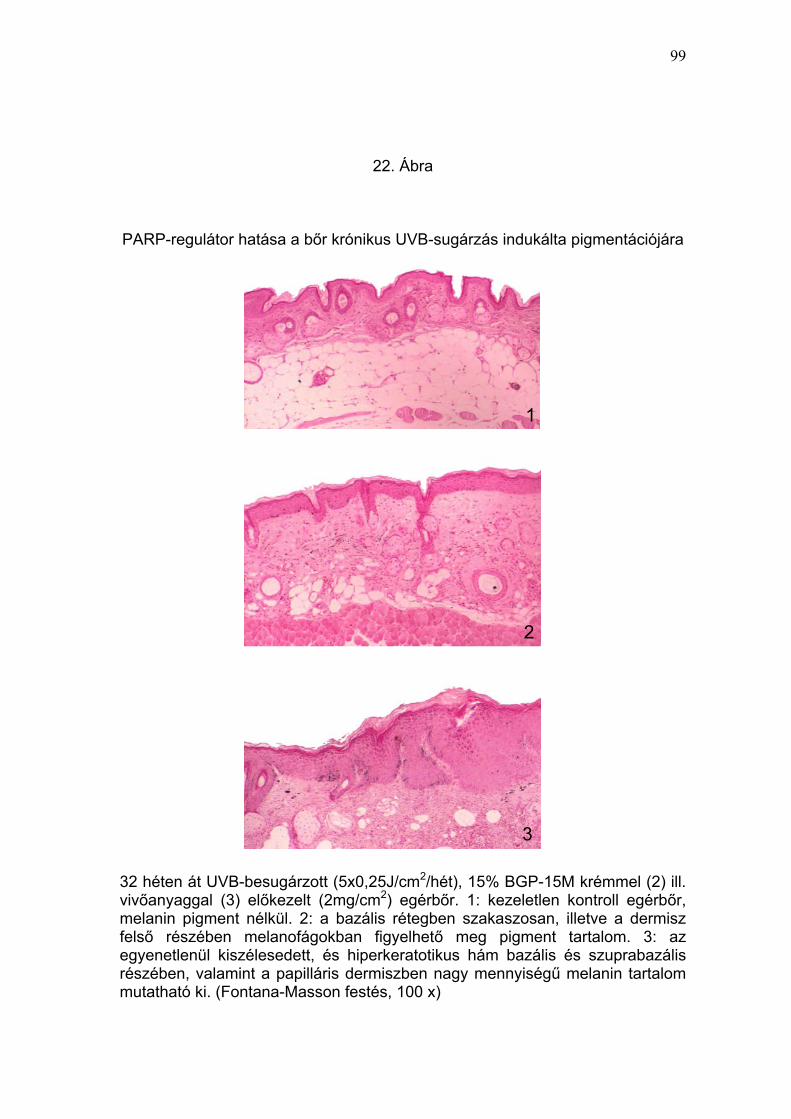
99
22. Ábra
PARP-regulátor hatása a bőr krónikus UVB-sugárzás indukálta pigmentációjára 32 héten át UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel (2) ill. vivőanyaggal (3) előkezelt (2mg/cm2) egérbőr. 1: kezeletlen kontroll egérbőr, melanin pigment nélkül. 2: a bazális rétegben szakaszosan, illetve a dermisz felső részében melanofágokban figyelhető meg pigment tartalom. 3: az egyenetlenül kiszélesedett, és hiperkeratotikus hám bazális és szuprabazális részében, valamint a papilláris dermiszben nagy mennyiségű melanin tartalom mutatható ki. (Fontana-Masson festés, 100 x)
1
2
3
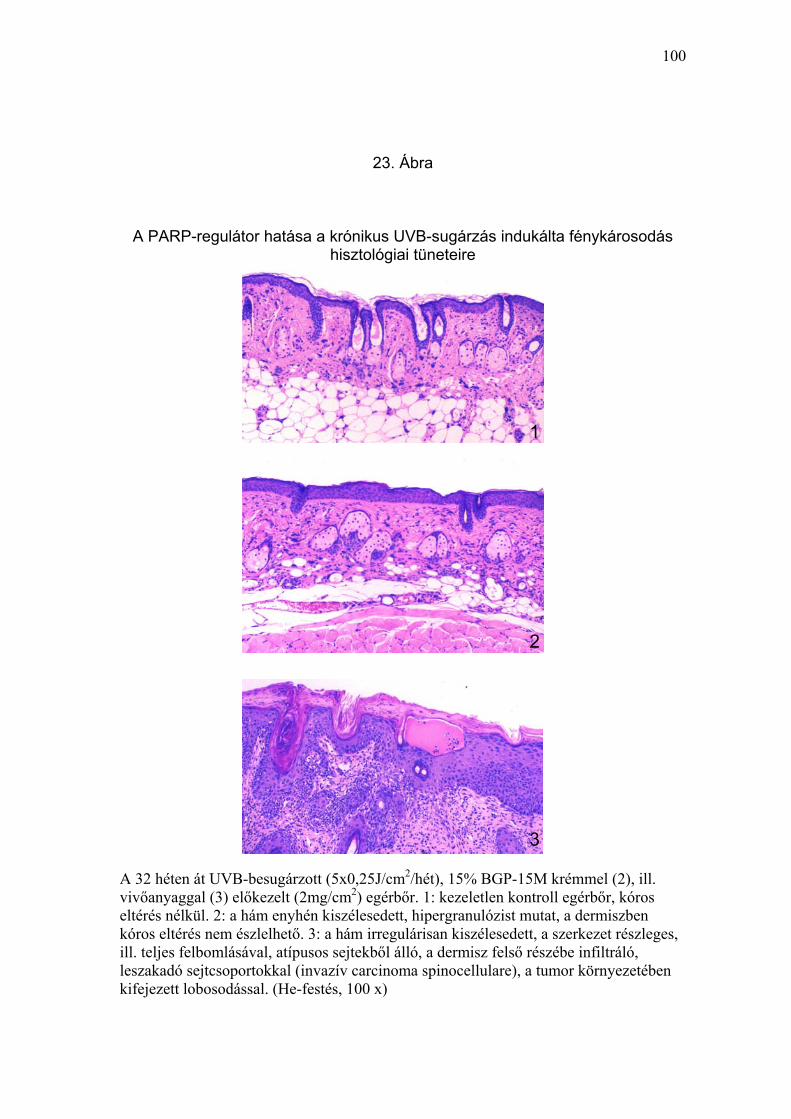
100
23. Ábra
A PARP-regulátor hatása a krónikus UVB-sugárzás indukálta fénykárosodás hisztológiai tüneteire
A 32 héten át UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel (2), ill. vivőanyaggal (3) előkezelt (2mg/cm2) egérbőr. 1: kezeletlen kontroll egérbőr, kóros eltérés nélkül. 2: a hám enyhén kiszélesedett, hipergranulózist mutat, a dermiszben kóros eltérés nem észlelhető. 3: a hám irregulárisan kiszélesedett, a szerkezet részleges, ill. teljes felbomlásával, atípusos sejtekből álló, a dermisz felső részébe infiltráló, leszakadó sejtcsoportokkal (invazív carcinoma spinocellulare), a tumor környezetében kifejezett lobosodással. (He-festés, 100 x)
1
2
3
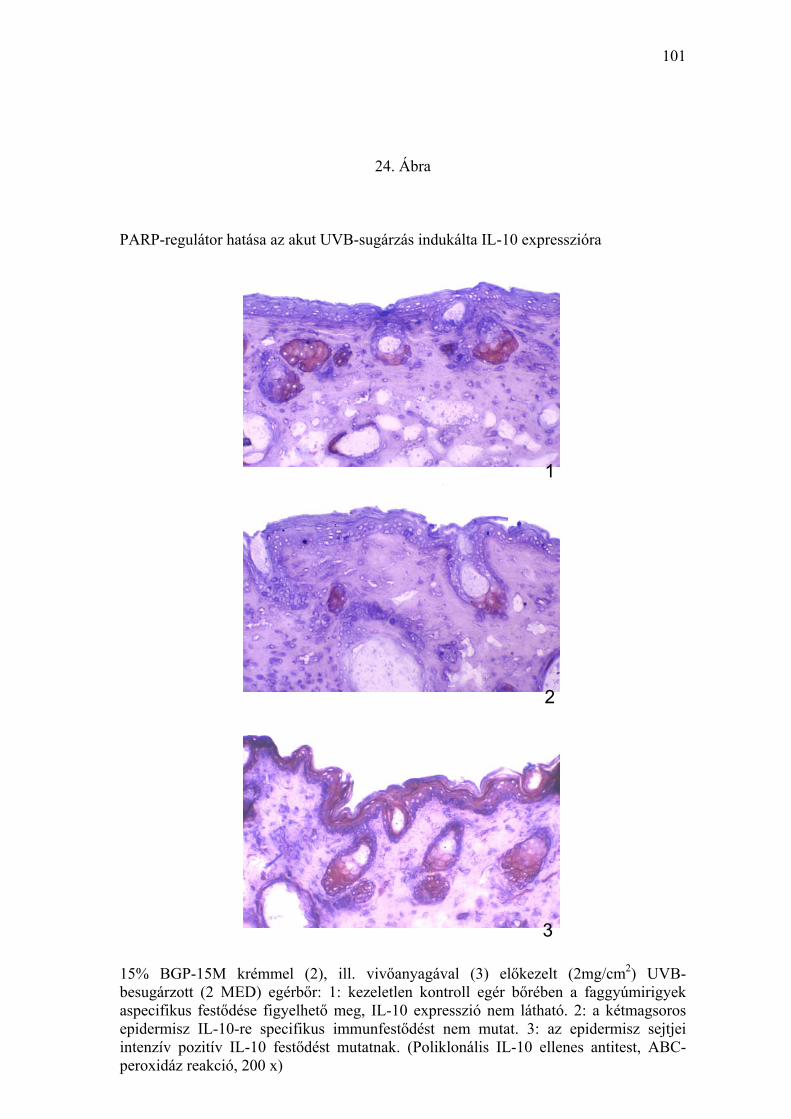
101
24. Ábra PARP-regulátor hatása az akut UVB-sugárzás indukálta IL-10 expresszióra 15% BGP-15M krémmel (2), ill. vivőanyagával (3) előkezelt (2mg/cm2) UVB-besugárzott (2 MED) egérbőr: 1: kezeletlen kontroll egér bőrében a faggyúmirigyek aspecifikus festődése figyelhető meg, IL-10 expresszió nem látható. 2: a kétmagsoros epidermisz IL-10-re specifikus immunfestődést nem mutat. 3: az epidermisz sejtjei intenzív pozitív IL-10 festődést mutatnak. (Poliklonális IL-10 ellenes antitest, ABC-peroxidáz reakció, 200 x)
1
2
3
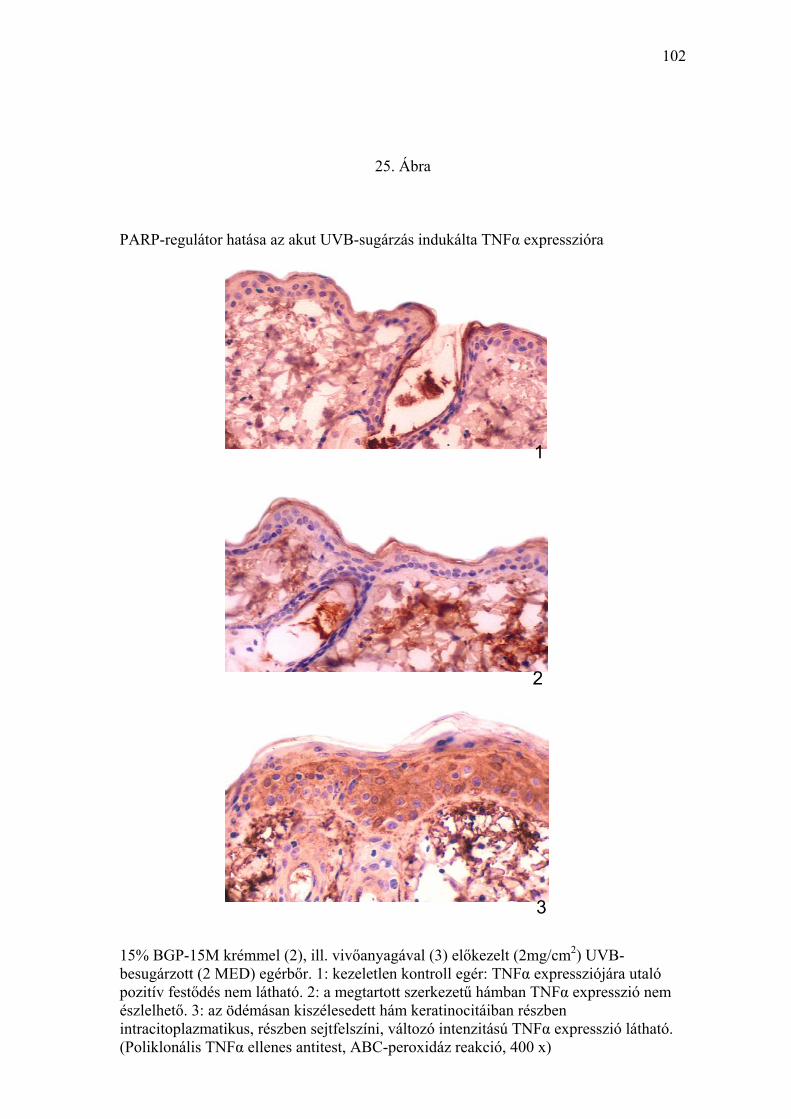
102
25. Ábra PARP-regulátor hatása az akut UVB-sugárzás indukálta TNFα expresszióra 15% BGP-15M krémmel (2), ill. vivőanyagával (3) előkezelt (2mg/cm2) UVB-besugárzott (2 MED) egérbőr. 1: kezeletlen kontroll egér: TNFα expressziójára utaló pozitív festődés nem látható. 2: a megtartott szerkezetű hámban TNFα expresszió nem észlelhető. 3: az ödémásan kiszélesedett hám keratinocitáiban részben intracitoplazmatikus, részben sejtfelszíni, változó intenzitású TNFα expresszió látható. (Poliklonális TNFα ellenes antitest, ABC-peroxidáz reakció, 400 x)
1
2
3
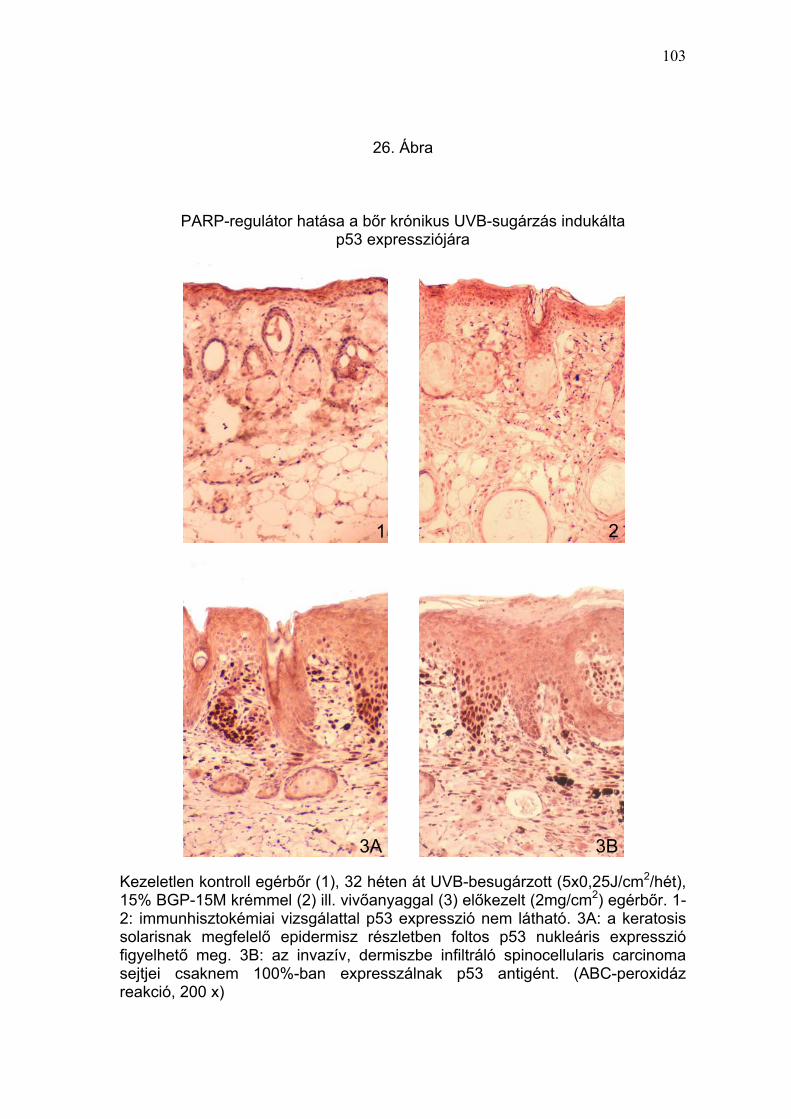
103
26. Ábra
PARP-regulátor hatása a bőr krónikus UVB-sugárzás indukálta p53 expressziójára
Kezeletlen kontroll egérbőr (1), 32 héten át UVB-besugárzott (5x0,25J/cm2/hét), 15% BGP-15M krémmel (2) ill. vivőanyaggal (3) előkezelt (2mg/cm2) egérbőr. 1-2: immunhisztokémiai vizsgálattal p53 expresszió nem látható. 3A: a keratosis solarisnak megfelelő epidermisz részletben foltos p53 nukleáris expresszió figyelhető meg. 3B: az invazív, dermiszbe infiltráló spinocellularis carcinoma sejtjei csaknem 100%-ban expresszálnak p53 antigént. (ABC-peroxidáz reakció, 200 x)
3A 3B
2 1
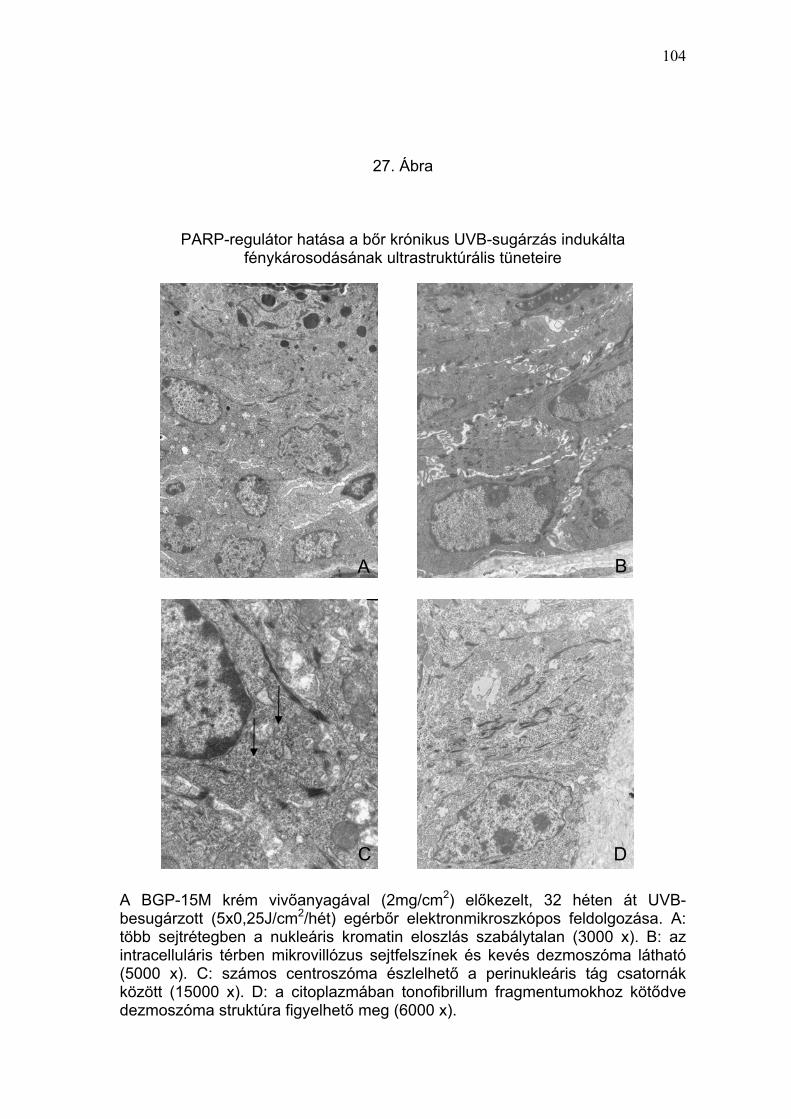
104
27. Ábra
PARP-regulátor hatása a bőr krónikus UVB-sugárzás indukálta fénykárosodásának ultrastruktúrális tüneteire
A BGP-15M krém vivőanyagával (2mg/cm2) előkezelt, 32 héten át UVB-besugárzott (5x0,25J/cm2/hét) egérbőr elektronmikroszkópos feldolgozása. A: több sejtrétegben a nukleáris kromatin eloszlás szabálytalan (3000 x). B: az intracelluláris térben mikrovillózus sejtfelszínek és kevés dezmoszóma látható (5000 x). C: számos centroszóma észlelhető a perinukleáris tág csatornák között (15000 x). D: a citoplazmában tonofibrillum fragmentumokhoz kötődve dezmoszóma struktúra figyelhető meg (6000 x).
A B
C D
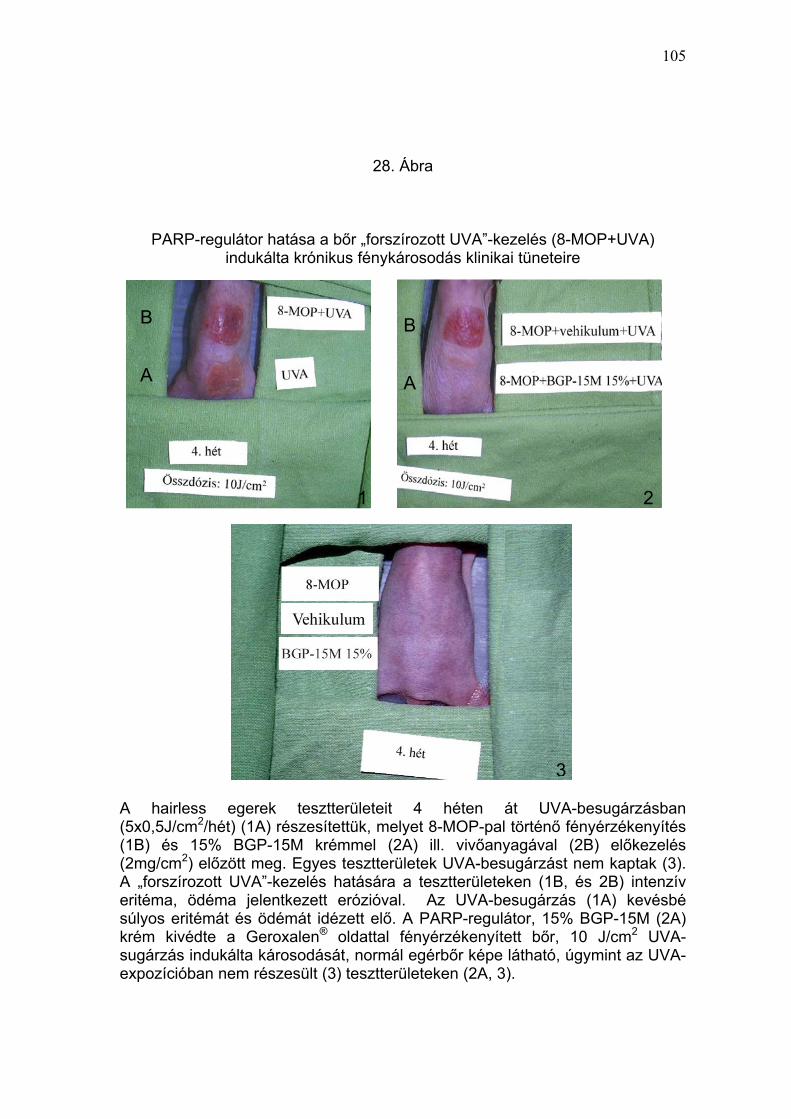
105
28. Ábra
PARP-regulátor hatása a bőr „forszírozott UVA”-kezelés (8-MOP+UVA) indukálta krónikus fénykárosodás klinikai tüneteire
A hairless egerek tesztterületeit 4 héten át UVA-besugárzásban (5x0,5J/cm2/hét) (1A) részesítettük, melyet 8-MOP-pal történő fényérzékenyítés (1B) és 15% BGP-15M krémmel (2A) ill. vivőanyagával (2B) előkezelés (2mg/cm2) előzött meg. Egyes tesztterületek UVA-besugárzást nem kaptak (3). A „forszírozott UVA”-kezelés hatására a tesztterületeken (1B, és 2B) intenzív eritéma, ödéma jelentkezett erózióval. Az UVA-besugárzás (1A) kevésbé súlyos eritémát és ödémát idézett elő. A PARP-regulátor, 15% BGP-15M (2A) krém kivédte a Geroxalen® oldattal fényérzékenyített bőr, 10 J/cm2 UVA-sugárzás indukálta károsodását, normál egérbőr képe látható, úgymint az UVA-expozícióban nem részesült (3) tesztterületeken (2A, 3).
1 2
3
B
A
B
A
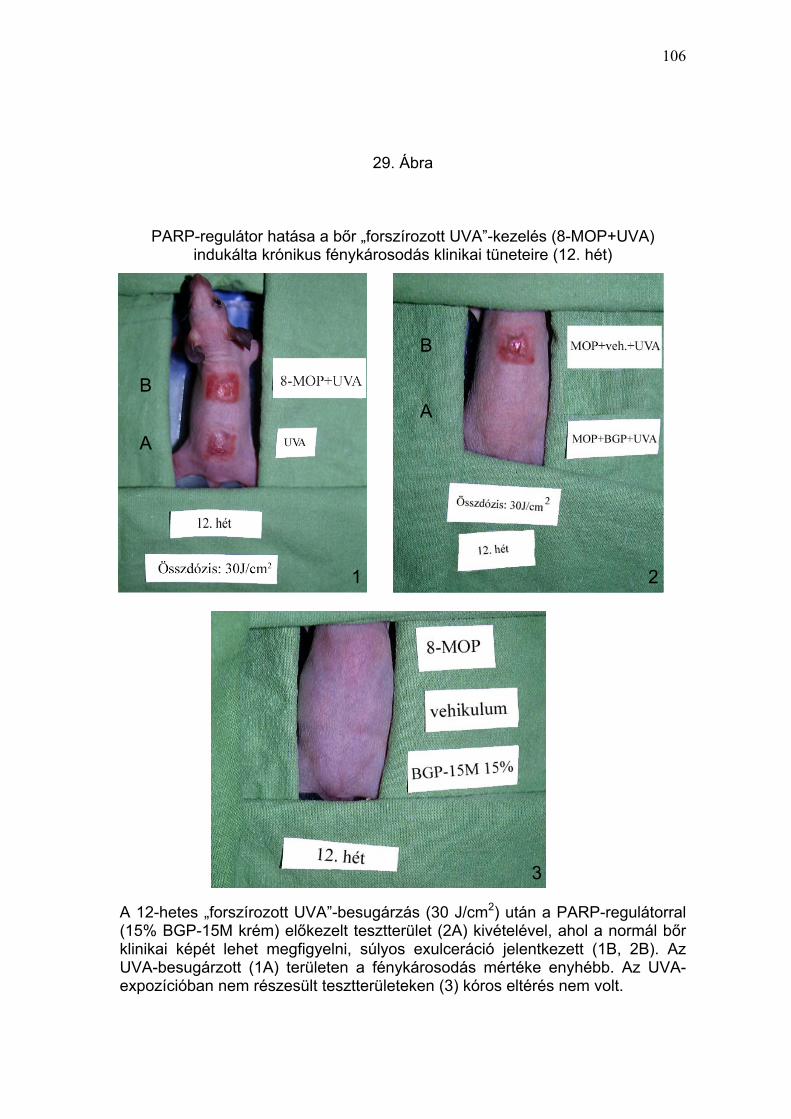
106
29. Ábra
PARP-regulátor hatása a bőr „forszírozott UVA”-kezelés (8-MOP+UVA)
indukálta krónikus fénykárosodás klinikai tüneteire (12. hét) A 12-hetes „forszírozott UVA”-besugárzás (30 J/cm2) után a PARP-regulátorral (15% BGP-15M krém) előkezelt tesztterület (2A) kivételével, ahol a normál bőr klinikai képét lehet megfigyelni, súlyos exulceráció jelentkezett (1B, 2B). Az UVA-besugárzott (1A) területen a fénykárosodás mértéke enyhébb. Az UVA-expozícióban nem részesült tesztterületeken (3) kóros eltérés nem volt.
1 2
3
B
A
B
A
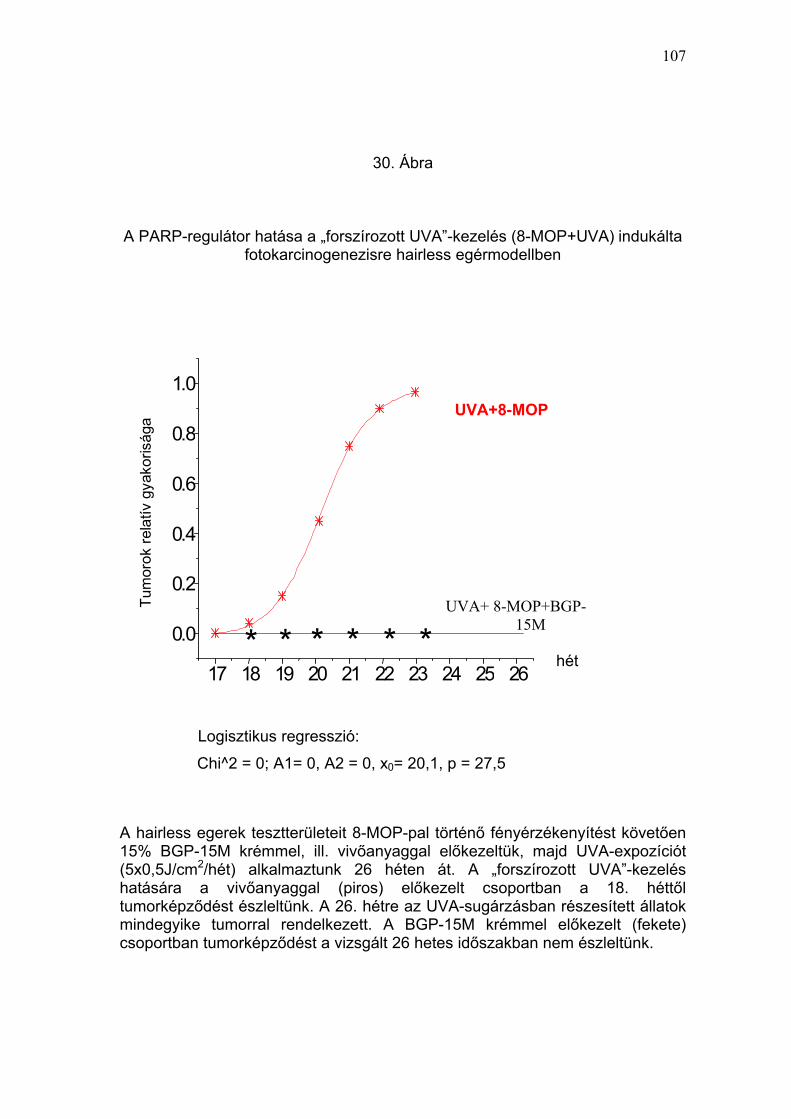
107
30. Ábra
A PARP-regulátor hatása a „forszírozott UVA”-kezelés (8-MOP+UVA) indukálta fotokarcinogenezisre hairless egérmodellben
A hairless egerek tesztterületeit 8-MOP-pal történő fényérzékenyítést követően 15% BGP-15M krémmel, ill. vivőanyaggal előkezeltük, majd UVA-expozíciót (5x0,5J/cm2/hét) alkalmaztunk 26 héten át. A „forszírozott UVA”-kezelés hatására a vivőanyaggal (piros) előkezelt csoportban a 18. héttől tumorképződést észleltünk. A 26. hétre az UVA-sugárzásban részesített állatok mindegyike tumorral rendelkezett. A BGP-15M krémmel előkezelt (fekete) csoportban tumorképződést a vizsgált 26 hetes időszakban nem észleltünk.
17 18 19 20 21 22 23 24 25 26
0.0
0.2
0.4
0.6
0.8
1.0
*** * * *
Tum
orok
rela
tív g
yako
riság
a
hét
Chi^2 = 0; A1= 0, A2 = 0, x0= 20,1, p = 27,5
Logisztikus regresszió:
UVA+8-MOP
UVA+ 8-MOP+BGP-15M
hét
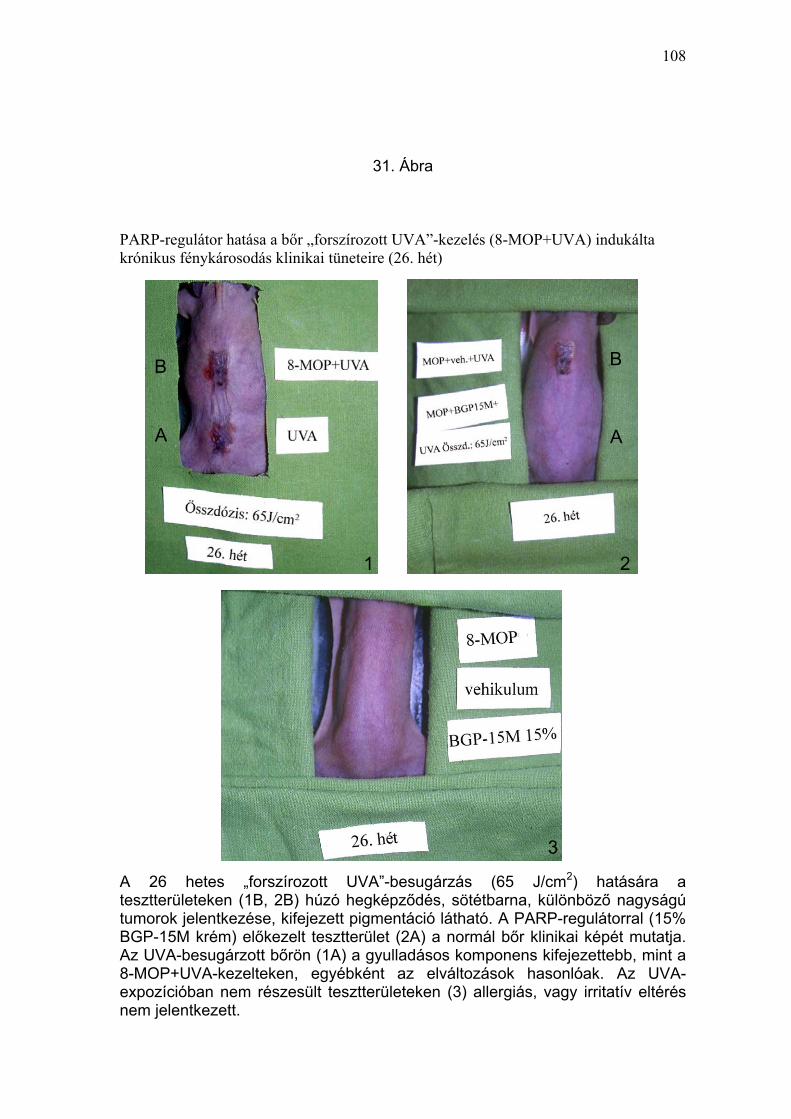
108
31. Ábra PARP-regulátor hatása a bőr „forszírozott UVA”-kezelés (8-MOP+UVA) indukálta krónikus fénykárosodás klinikai tüneteire (26. hét) A 26 hetes „forszírozott UVA”-besugárzás (65 J/cm2) hatására a tesztterületeken (1B, 2B) húzó hegképződés, sötétbarna, különböző nagyságú tumorok jelentkezése, kifejezett pigmentáció látható. A PARP-regulátorral (15% BGP-15M krém) előkezelt tesztterület (2A) a normál bőr klinikai képét mutatja. Az UVA-besugárzott bőrön (1A) a gyulladásos komponens kifejezettebb, mint a 8-MOP+UVA-kezelteken, egyébként az elváltozások hasonlóak. Az UVA-expozícióban nem részesült tesztterületeken (3) allergiás, vagy irritatív eltérés nem jelentkezett.
1 2
3
B
A
B
A
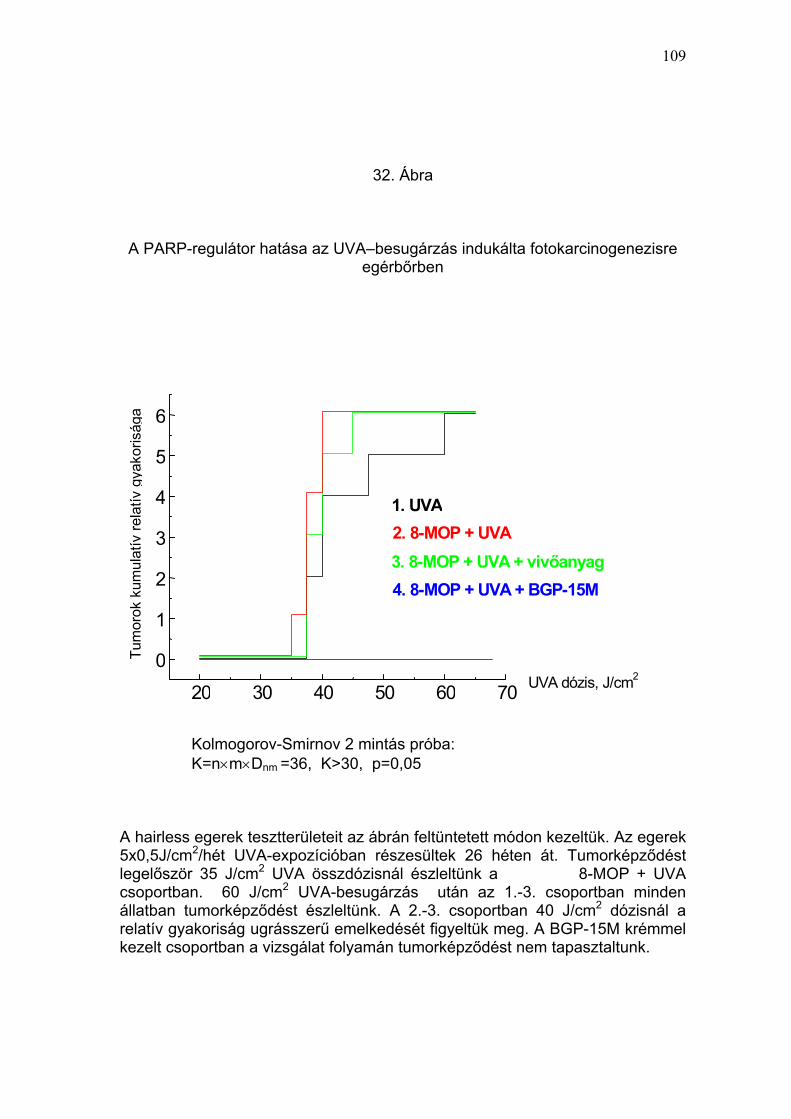
109
32. Ábra
A PARP-regulátor hatása az UVA–besugárzás indukálta fotokarcinogenezisre
egérbőrben
Kolmogorov-Smirnov 2 mintás próba: K=n×m×Dnm =36, K>30, p=0,05
A hairless egerek tesztterületeit az ábrán feltüntetett módon kezeltük. Az egerek 5x0,5J/cm2/hét UVA-expozícióban részesültek 26 héten át. Tumorképződést legelőször 35 J/cm2 UVA összdózisnál észleltünk a 8-MOP + UVA csoportban. 60 J/cm2 UVA-besugárzás után az 1.-3. csoportban minden állatban tumorképződést észleltünk. A 2.-3. csoportban 40 J/cm2 dózisnál a relatív gyakoriság ugrásszerű emelkedését figyeltük meg. A BGP-15M krémmel kezelt csoportban a vizsgálat folyamán tumorképződést nem tapasztaltunk.
20 30 40 50 60 70
0
1
2
3
4
5
6
4. 8-MOP + UVA + BGP-15M 3. 8-MOP + UVA + vivőanyag 2. 8-MOP + UVA 1. UVA
UVA dózis, J/cm2
Tum
orok
kum
ulat
ív re
latív
gya
koris
ága
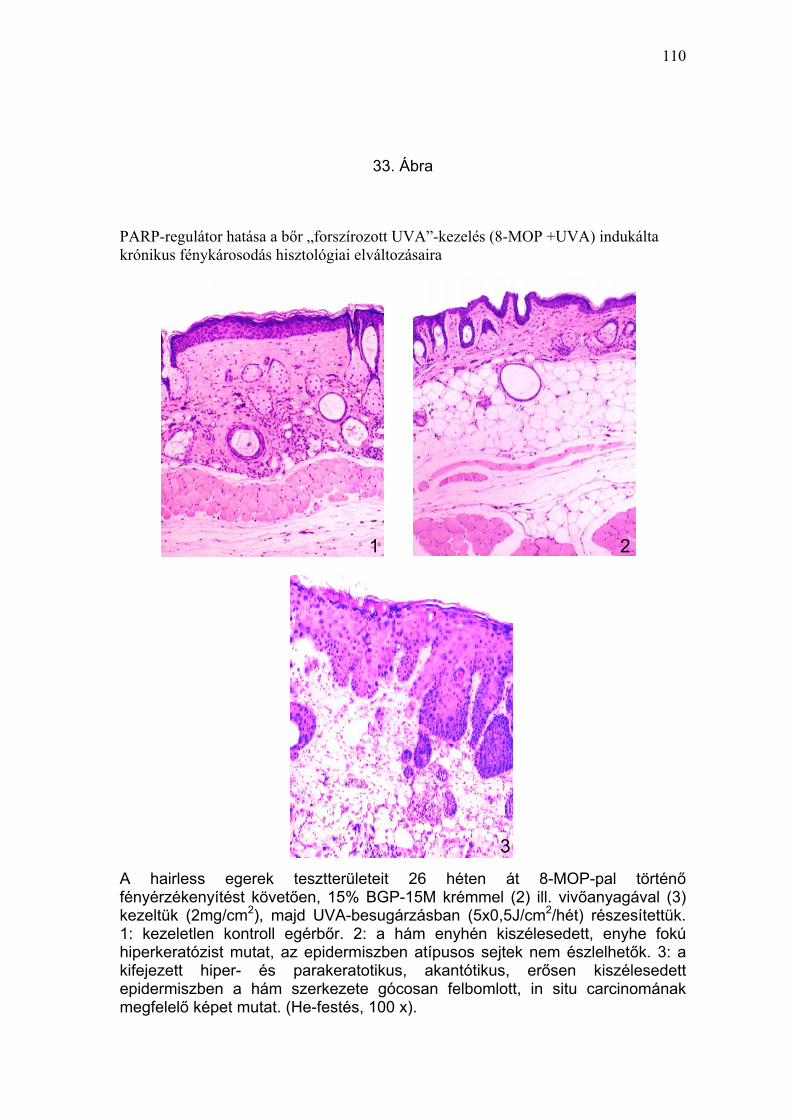
110
33. Ábra PARP-regulátor hatása a bőr „forszírozott UVA”-kezelés (8-MOP +UVA) indukálta krónikus fénykárosodás hisztológiai elváltozásaira A hairless egerek tesztterületeit 26 héten át 8-MOP-pal történő fényérzékenyítést követően, 15% BGP-15M krémmel (2) ill. vivőanyagával (3) kezeltük (2mg/cm2), majd UVA-besugárzásban (5x0,5J/cm2/hét) részesítettük. 1: kezeletlen kontroll egérbőr. 2: a hám enyhén kiszélesedett, enyhe fokú hiperkeratózist mutat, az epidermiszben atípusos sejtek nem észlelhetők. 3: a kifejezett hiper- és parakeratotikus, akantótikus, erősen kiszélesedett epidermiszben a hám szerkezete gócosan felbomlott, in situ carcinomának megfelelő képet mutat. (He-festés, 100 x).
1 2
3

111
34. Ábra
A PARP-regulátor hatása a krónikus „forszírozott UVA”-kezelés indukálta rugalmas rost károsodásra egérbőrben
A hairless egerek tesztterületeit 26 héten át 8-MOP–pal történő fényérzékenyítést követően, 15% BGP-15M krémmel (2) ill. vivőanyagával (3) előkezeltük (2mg/cm2) és UVA-besugárzásban (5x0,5J/cm2/hét) részesítettük. 1: kezeletlen kontroll egérbőr: kronológiai öregedésnek megfelelően a rugalmas rostok megkevesbedése és enyhe feltöredezése, valamint a dermisz középső, alsó részében ciszták kialakulása látható. 2: a hám 5-8 sorosra kiszélesedett, a rugalmas rost hálózat enyhe fokú megkevesbedése figyelhető meg. 3: az in situ carcinoma környezetében rugalmas rost fragmentációt észleltünk. (Orcein-Giemsa festés, 100 x)
1 2
3
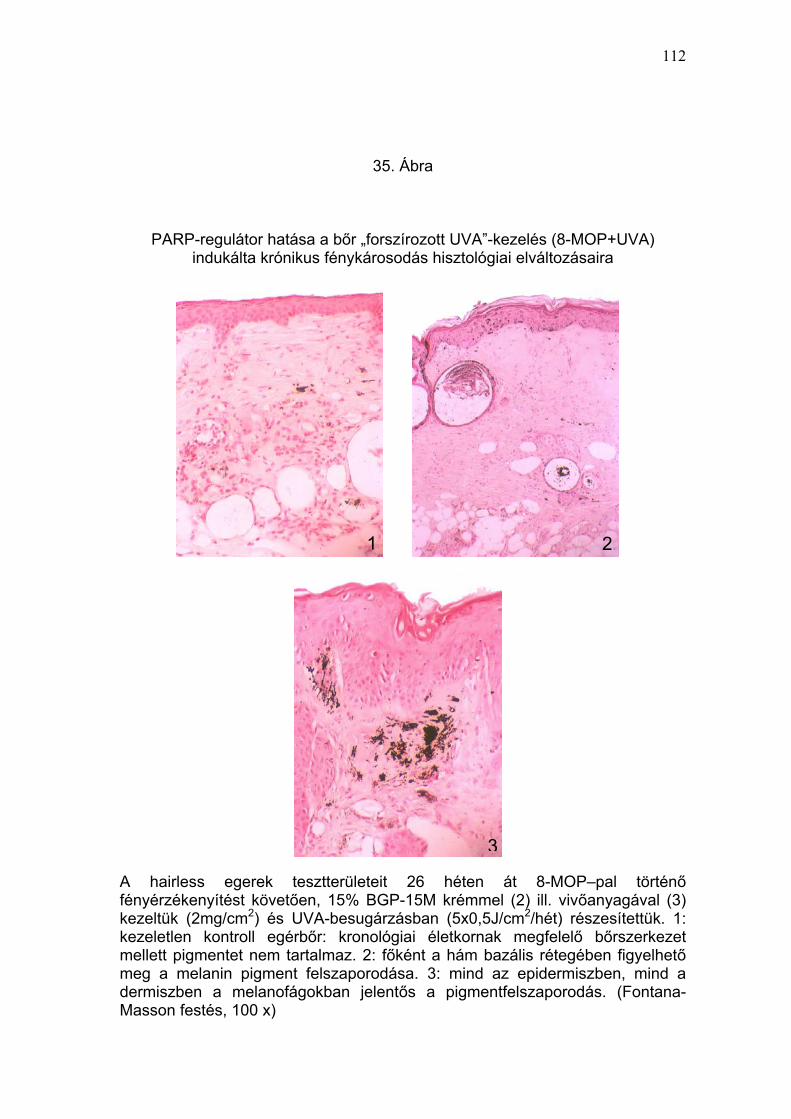
112
35. Ábra
PARP-regulátor hatása a bőr „forszírozott UVA”-kezelés (8-MOP+UVA) indukálta krónikus fénykárosodás hisztológiai elváltozásaira
A hairless egerek tesztterületeit 26 héten át 8-MOP–pal történő fényérzékenyítést követően, 15% BGP-15M krémmel (2) ill. vivőanyagával (3) kezeltük (2mg/cm2) és UVA-besugárzásban (5x0,5J/cm2/hét) részesítettük. 1: kezeletlen kontroll egérbőr: kronológiai életkornak megfelelő bőrszerkezet mellett pigmentet nem tartalmaz. 2: főként a hám bazális rétegében figyelhető meg a melanin pigment felszaporodása. 3: mind az epidermiszben, mind a dermiszben a melanofágokban jelentős a pigmentfelszaporodás. (Fontana-Masson festés, 100 x)
1
3
2
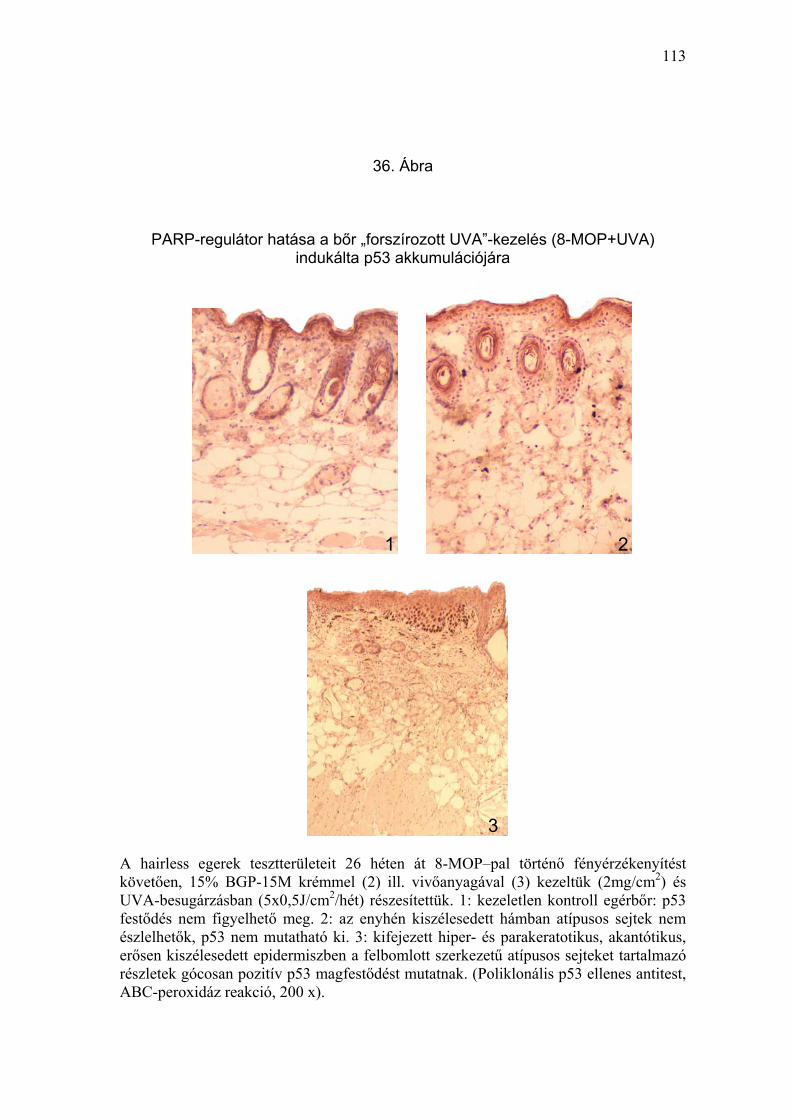
113
36. Ábra
PARP-regulátor hatása a bőr „forszírozott UVA”-kezelés (8-MOP+UVA)
indukálta p53 akkumulációjára
A hairless egerek tesztterületeit 26 héten át 8-MOP–pal történő fényérzékenyítést követően, 15% BGP-15M krémmel (2) ill. vivőanyagával (3) kezeltük (2mg/cm2) és UVA-besugárzásban (5x0,5J/cm2/hét) részesítettük. 1: kezeletlen kontroll egérbőr: p53 festődés nem figyelhető meg. 2: az enyhén kiszélesedett hámban atípusos sejtek nem észlelhetők, p53 nem mutatható ki. 3: kifejezett hiper- és parakeratotikus, akantótikus, erősen kiszélesedett epidermiszben a felbomlott szerkezetű atípusos sejteket tartalmazó részletek gócosan pozitív p53 magfestődést mutatnak. (Poliklonális p53 ellenes antitest, ABC-peroxidáz reakció, 200 x).
1
3
2
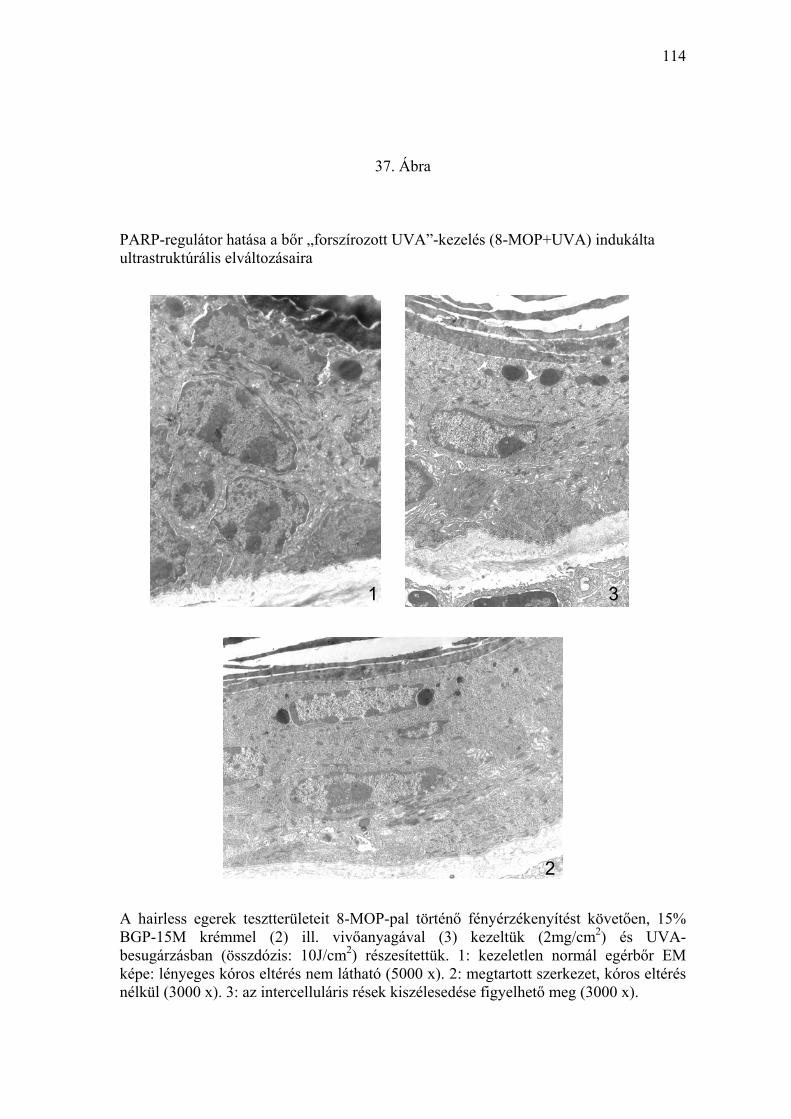
114
37. Ábra PARP-regulátor hatása a bőr „forszírozott UVA”-kezelés (8-MOP+UVA) indukálta ultrastruktúrális elváltozásaira
A hairless egerek tesztterületeit 8-MOP-pal történő fényérzékenyítést követően, 15% BGP-15M krémmel (2) ill. vivőanyagával (3) kezeltük (2mg/cm2) és UVA-besugárzásban (összdózis: 10J/cm2) részesítettük. 1: kezeletlen normál egérbőr EM képe: lényeges kóros eltérés nem látható (5000 x). 2: megtartott szerkezet, kóros eltérés nélkül (3000 x). 3: az intercelluláris rések kiszélesedése figyelhető meg (3000 x).
1 3
2
![Flyer PARP FAmily NP V01 - biolinks k.k.1].pdf · Tomorrow’s Reagents Manufactured Today® International Edition The PARP Family T highlight PRODUCT FLYER The PARP Family](https://img.pdfslide.net/doc/110x75/5cb9946788c993f37c8c0cfc/flyer-parp-family-np-v01-biolinks-kk-1pdf-tomorrows-reagents-manufactured.jpg)




























