Embed Size (px)
Citation preview

POLLINATION BIOLOGY OF DARLINGTONIA CALIFORNICA, THE CALIFORNIA
PITCHER PLANT
by
George A. Meindl
A Thesis
Presented to
The Faculty of Humboldt State University
In Partial Fulfillment
Of the Requirements for the Degree
Master of Arts
In Natural Resources: Biology
December 2009

POLLINATION BIOLOGY OF DARLINGTONIA CALIFORNICA, THE CALIFORNIA
PITCHER PLANT
by
George A. Meindl
We certify that we have read this study and that it conforms to acceptable standards of scholarly presentation and is fully acceptable, in scope and quality, as a thesis for the degree of Master of Arts. Michael R. Mesler, Major Professor Date John O. Reiss, Committee Member Date Erik S. Jules, Committee Member Date Terry W. Henkel, Committee Member Date Michael R. Mesler, Graduate Coordinator Date John Lyon, Dean Research & Graduate Studies Date

iii
ABSTRACT
Pollination biology of Darlingtonia californica, the California pitcher plant
George A. Meindl
The flowers of the California pitcher plant, Darlingtonia californica, are unusual
and have been the subject of much speculation. Despite the efforts of several workers,
the pollination ecology of this plant, including the identities of pollinators, remains
enigmatic. Along with determining visitation frequency, this study sought to describe the
relative contributions of two documented floral visitors, bees and spiders. Fruit and seed
production by emasculated flowers were used to estimate levels of cross-pollination in
natural populations of D. californica. Under the assumption that bees predominantly act
as cross-pollinators and spiders as self-pollinators, this treatment was used to infer which
organisms were responsible for pollen transfer. Levels of pollen-limitation were also
assessed at five study sites in northwest California. In order to identify floral visitors,
extensive pollinator surveys were conducted. A generalist solitary bee, Andrena
nigrihirta, was found to visit D. californica flowers near Scott Mountain and Mt. Eddy,
CA. Despite relatively low visitation rates, individual flowers at all study sites were
expected to receive at least one visit by A. nigrihirta. Other regular floral visitors
included thrips and several species of spiders. Fruit and seed production by emasculated
flowers indicated the occurrence of cross-pollination. Unmanipulated flowers produced
more fruits and seeds, on average, compared to emasculated blooms, suggesting that self-
pollination contributes to D. californica reproductive success as well. While bees were

iv
likely responsible for the majority of cross-pollination, both bees and spiders contributed
to autogamous pollen transfer. Following observations of floral visits by A. nigrihirta, it
was possible to interpret the functional morphology of D. californica’s flowers. The
shape of D. californica’s ovary and petals promote stigma contact both when pollinators
enter and exit a flower, contrary to previous thought. This system provides an excellent
example of the importance of identifying and observing pollinators in order to truly
understand the functional significance of a plant’s floral morphology.

v
ACKNOWLEDGMENTS
First and foremost, I would like to thank my adviser, Dr. Michael Mesler, whose
combination of patience and enthusiasm made this work possible. I am also grateful to
the professors who served on my committee: Dr. Erik Jules, Dr. Terry Henkel, and Dr.
John Reiss. Many thanks to Dave Franklin, whose general knowledge contributed much
to this thesis. Tim Buonaccorsi and Sasan Hariri-Moghadam were invaluable as research
assistants. Without their help I might still be counting Darlingtonia seeds, among other
tasks. Figures 2 and 3 were illustrated by Sasan Hariri-Moghadam. Will Goldenberg
assisted with video editing. As always, thank you to my family and friends.

vi
TABLE OF CONTENTS
Page
ABSTRACT ....................................................................................................................... iii
ACKNOWLEDGMENTS .................................................................................................. v
LIST OF TABLES ............................................................................................................ vii
INTRODUCTION .............................................................................................................. 1
METHODS AND MATERIALS ........................................................................................ 5
Study Species .......................................................................................................... 5
Study Sites .............................................................................................................. 6
Pollinator Surveys ................................................................................................... 9
Analysis of Pollen Loads ...................................................................................... 10
Pollination Treatments .......................................................................................... 11
Plant Community Context .................................................................................... 12
Statistical Analyses ............................................................................................... 13
RESULTS ......................................................................................................................... 14
Flower Visitors...................................................................................................... 14
Floral Mechanism/ Pollinator Behavior ................................................................ 18
Pollination Treatments .......................................................................................... 23
Plant/ Insect Community Context ......................................................................... 24
DISCUSSION ................................................................................................................... 29
Future Research .................................................................................................... 33
LITERATURE CITED ..................................................................................................... 35

vii
LIST OF TABLES
Table Page 1 Elevation and geographic coordinates of five study sites. ...................................... 7
2 Observed visits to Darlingtonia californica flowers. Aside from the one visit recorded by a European honeybee (Apis mellifera), Andrena nigrihirta was the only bee observed to visit the flowers. Numbers in parentheses indicate the number of visits to different flowers observed during a given time period(s). ..... 15
3 Floral visitation rates. The mean number of visits a flower was expected to receive per hour and over its lifetime is presented for each study site. Standard error values for mean visits/hour are given in parentheses for each study site. .... 16
4 Duration of visits to Darlingtonia californica flowers by Andrena nigrihirta. Average visit duration was 2 minutes 8 seconds (SE=12 seconds ....................... 17
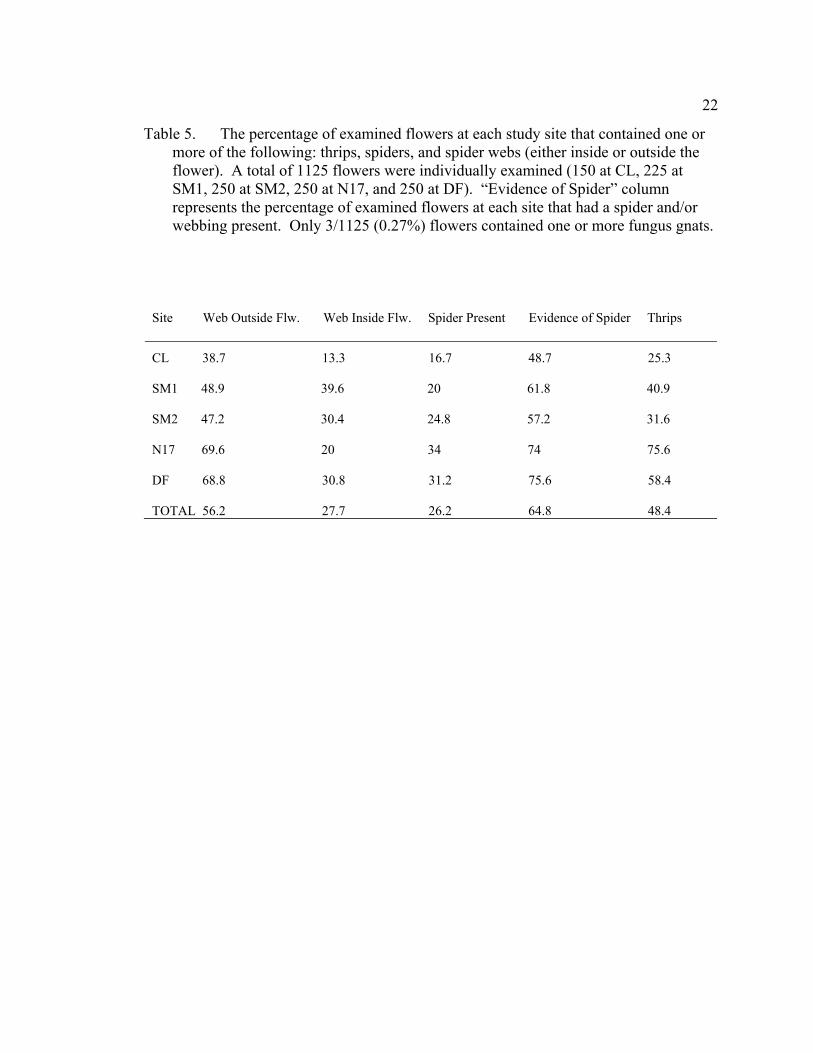
5 The percentage of examined flowers at each study site that contained one or more of the following: thrips, spiders, and spider webs (either inside or outside the flower). A total of 1125 flowers were individually examined (150 at CL, 225 at SM1, 250 at SM2, 250 at N17, and 250 at DF). “Evidence of Spider” column represents the percentage of examined flowers at each site that had a spider and/or webbing present. Only 3/1125 (0.27%) flowers contained one or more fungus gnats. ..................................................................................................................... 22
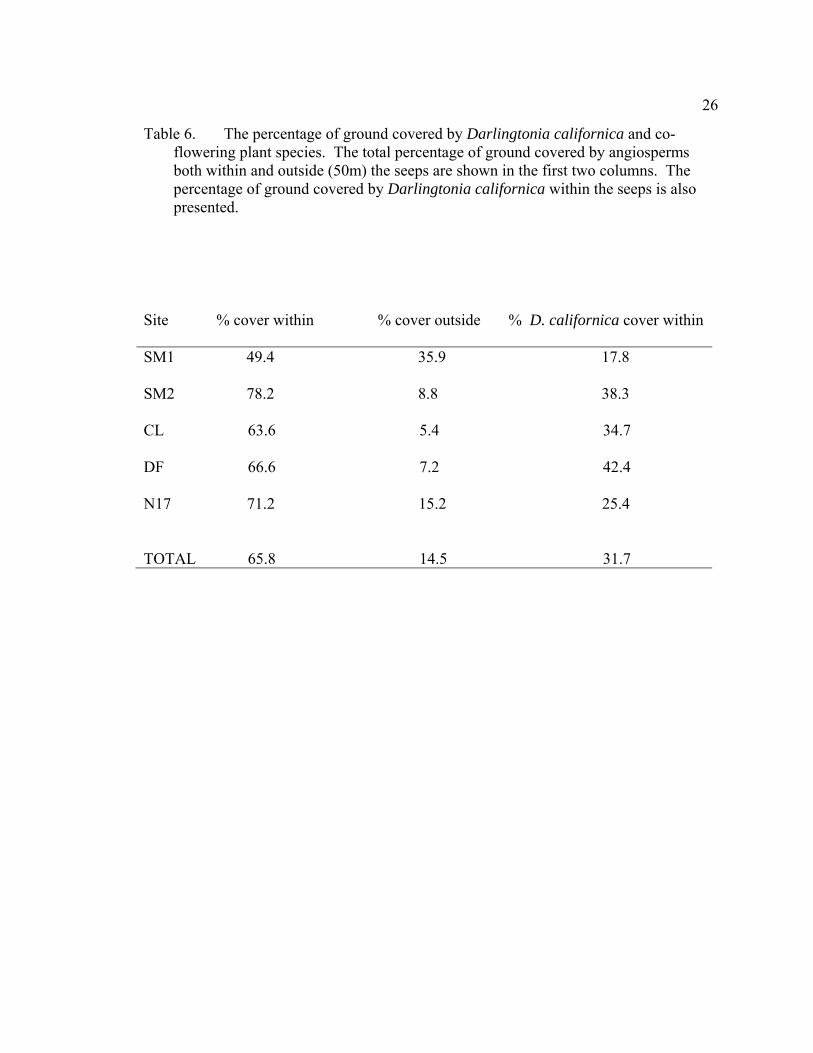
6 The percentage of ground covered by Darlingtonia californica and co-flowering plant species. The total percentage of ground covered by angiosperms both within and outside (50m) the seeps are shown in the first two columns. The percentage of ground covered by Darlingtonia californica within the seeps is also presented. .............................................................................................................. 26
7 Pollinators collected at or near study sites. Order and family is presented for all pollinators (and genus for Hymenopterans). Numbers in parentheses indicate the number of individual pollinators collected from each group ................................ 28

viii
LIST OF FIGURES
Figure Page
1 Map of study area. Triangles represent the five study sites. SM1, SM2, and CL were near the summit of Scott Mountain, while N17 and DF were near Mt. Eddy. The gray line separates Siskiyou County and Trinity County. .................... 8
2 Step by step foraging behavior of A. nigrihirta on a D. californica flower. The bee initially lands on the petals below the windows (a-c) and then enters a window and walks across stigmatic surfaces (d,e). The bee then utilizes the convex portion of one of the five petals to walk onto the ovary and up to collect pollen (f-i). Following pollen collection, the bee uses a petal convexity as before to walk down the ovary, across the stigmas again, and then out one of the windows (j-n). The flower is shown in d-m with one petal removed and half of the two lateral petals removed. F and k show the bee using the convex portion of the petal, which allows the bee to access the stamens. ......................................... 20
3 Interior view of a D. californica flower with the lower portion of petals removed. Arrows highlight the distance between the petals and the ovary both immediately above a window (shorter arrow) and in between two adjacent windows (larger arrow). More space is provided for A. nigrihirta in between the windows than above them, which encourages the bee to enter a window and then walk across the stigmatic surfaces. The bee then utilizes the convex portion of the petal opposite the window it entered en route to the flower’s stamens. ....................................... 21
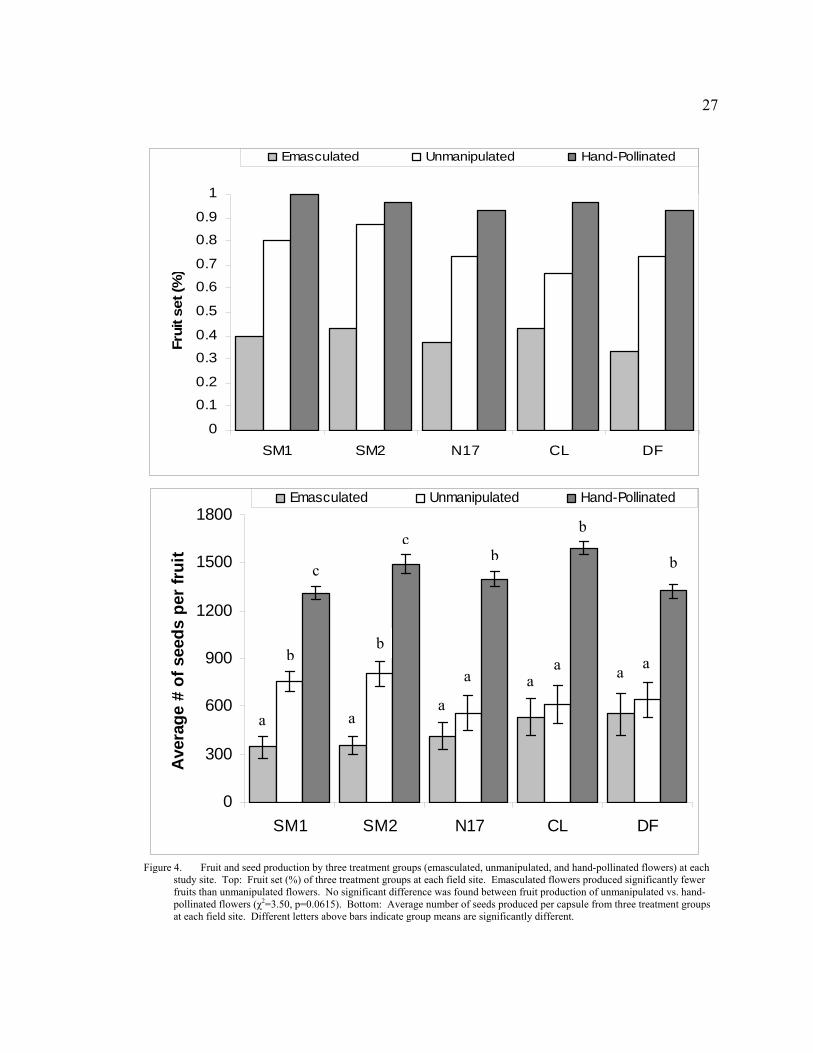
4 Fruit and seed production by three treatment groups (emasculated, unmanipulated, and hand-pollinated flowers) at each study site. Top: Fruit set (%) of three treatment groups at each field site. Emasculated flowers produced significantly fewer fruits than unmanipulated flowers. No significant difference was found between fruit production of unmanipulated vs. hand-pollinated flowers (χ2=3.50, p=0.0615). Bottom: Average number of seeds produced per capsule from three treatment groups at each field site. Different letters above bars indicate group means are significantly different. .......................................................................... 27

ix
LIST OF APPENDICES
Appendix Page
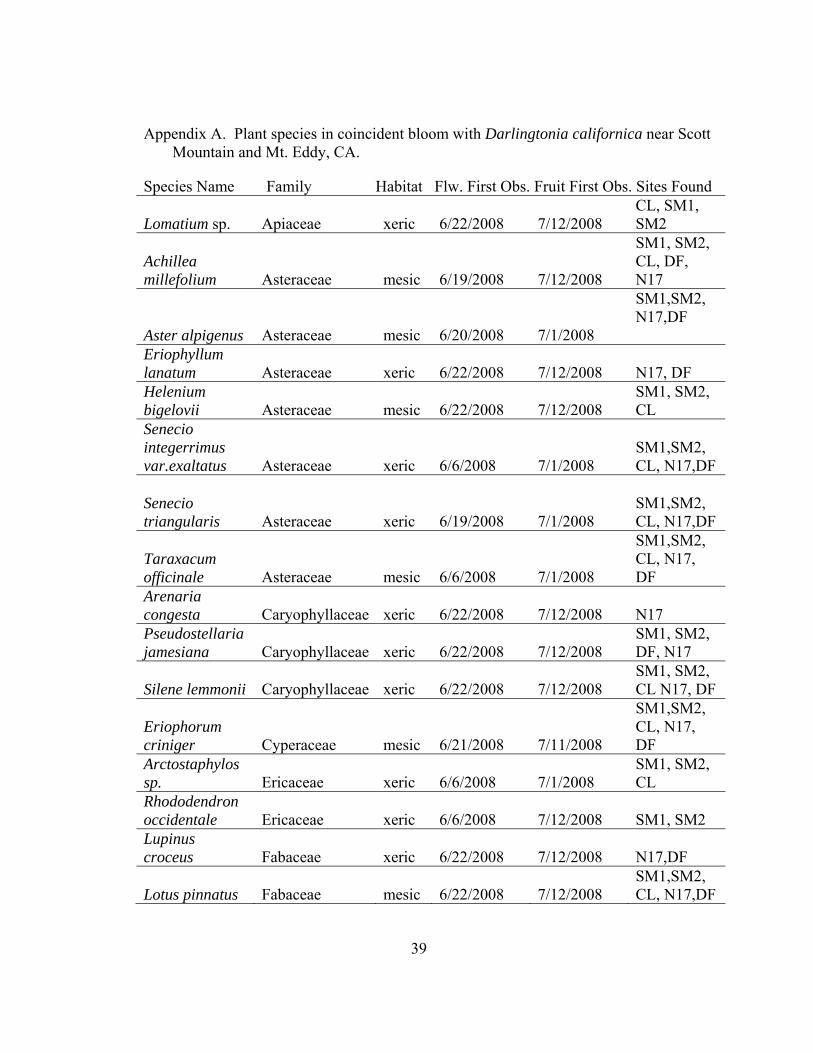
A Plant species in coincident bloom with Darlingtonia californica near Scott Mountain and Mt. Eddy, CA................................................................................. 39
B Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside seeps at all five study sites. .......................... 43

1
INTRODUCTION
The study of interactions between plants and their pollinators can provide
adaptive explanations for flower morphology (Fenster et al. 2004, Vogel 2006, Hu et al.
2008, Pauw et al. 2008, Crepet and Niklas 2009). Unless a plant’s effective pollinator is
determined, however, the adaptive significance of individual floral traits cannot be fully
understood. The flowers of the California pitcher plant, Darlingtonia californica, are
unusual and have been the subject of much speculation (Debuhr 1973, Schnell 1976).
For example, some have theorized that D. californica’s bell-shaped ovary serves to limit
self-pollination (Schnell 1976), but direct observations of flower handling by floral
visitors are lacking. Despite the efforts of several workers, the pollination ecology of this
plant, including the identities of pollinators, remains enigmatic (Austin 1875-1877, Elder
1997, Nyoka and Ferguson 1999, Nyoka 2000). Until D. californica’s pollinators are
identified and observed, we will not be in a position to interpret the functional
morphology of the flowers in a meaningful way.
Though pollinator sightings have been rare, fruit set in natural populations of D.
californica has been shown to be quite high, indicating an active pollinator community
since the flowers do not routinely self-pollinate without an animal pollen vector (Elder
1997, Nyoka 2000). Based on appearances, the flowers of D. californica seem adapted
for pollination by bees. They are large, showy, sweetly fragrant, and produce abundant
pollen (Debuhr 1973, Nyoka and Ferguson 1999) – all features commonly associated
with mellitophily. In addition, D. californica’s closest relatives, Sarracenia and

2
Heliamphora spp., are pollinated predominantly by bumble bees (Thomas and Cameron
1986, Renner 1989, Ne’eman et. al. 2006), suggesting that bee pollination may be
primitive for Sarraceniaceae. Nevertheless, bees have seldom been observed as visitors
to D. californica flowers (Austin 1875-1877, Elder 1997, Nyoka and Ferguson 1999,
Nyoka 2000). Spiders, in contrast, are relatively commonly observed on the flowers.
While pollination is generally not a service provided by spiders, recent studies suggest
that they may be significant pollinators of D. californica flowers (Nyoka and Ferguson
1999, Nyoka 2000). This interpretation seems problematic considering D. californica’s
floral morphology, but it is possible that spiders are currently responsible for the majority
of pollination even if flower morphology is the result of past selective pressures applied
by bees. If D. californica is pollinated exclusively by spiders, this system would provide
another example where floral morphology does not necessarily reflect contemporary
pollinator type (Fenster et al. 2004, Zhang et al. 2005, Valdivia and Niemeyer 2006).
Certain spider taxa, including members of the families Clubionidae, Salticidae,
and Theridiidae, use the flowers of D. californica as hunting grounds and may act as
pollen vectors as they move within flowers constructing webs and stalking prey (Nyoka
and Ferguson 1999, Nyoka 2000). By experimentally introducing spiders to bagged
flowers, Nyoka (2000) showed that spiders can cause autogamous pollen transfer. She
also observed pollen-dusted webbing constructed between adjacent flowers, indicating
the potential for xenogamous pollen transfer by spiders as well (Susan Nyoka, personal
communication).

3
The evidence for spider pollination, however, is far from conclusive. While
Nyoka (2000) demonstrated that spiders can potentially act as pollen vectors, fruit set by
open-pollinated flowers (96%, n=25) was much higher than that resulting from spider
introductions (41%, n=22) (Nyoka 2000). Of the flowers that matured fruits, far fewer
seeds were produced by spider-pollinated flowers (average=32 seeds/capsule) than open-
pollinated flowers (average=673 seeds/capsule) (Nyoka 2000). The discrepancy in
reproductive success between spider introduced and open-pollinated flowers suggests that
another, more effective pollinator is active among D. californica blooms. Though some
spiders supplement their diets with nectar and/or pollen (Smith and Mommsen 1984,
Vogelei and Greissl 1989), ultimately they rely on insect prey to sustain themselves. If
spiders feed on potentially effective pollinators, they may reduce seed production more
than they contribute to it.
This study sought to address the following questions regarding D. californica’s
reproductive biology at five study sites in northern California: (1) Do both bees and
spiders contribute to overall reproductive success? (2) Are past interpretations of the
functional morphology of floral traits correct, i.e., does the shape of D. californica’s
ovary limit self-pollination as has been suggested (Schnell 1976)? (3) Is natural
pollination sufficient for effective fruit and seed set, or are plants experiencing pollen-
limitation? (4) Do cross-pollination and self-pollination each contribute to natural
pollination? In order to identify floral visitors, extensive pollinator surveys were
conducted. Once a key floral visitor was identified detailed observations of its flower-
handling behavior were made to elucidate whether the shape of D. californica’s flowers

4
serves to limit self-pollination. Levels of pollen-limitation were assessed by comparing
fruit and seed production by hand-pollinated flowers with fruit and seed production by
unmanipulated flowers. The performance of emasculated flowers was used to estimate
levels of cross-pollination.

5
METHODS AND MATERIALS
Study Species
Darlingtonia californica is endemic to western Oregon and northern California,
and has a patchy distribution across its range due to its unique habitat requirements. The
species is considered an indicator of serpentine soil and is restricted to perennially wet
seeps (Whittaker 1954, Schnell 1976, Juniper et al. 1989). A long-lived perennial, D.
californica annually produces rosettes of carnivorous leaves from a creeping rhizome.
Plants often occur in dense patches, which are likely the result of clonal spread by
rhizomes and stolons (Schnell 1976).
The solitary flowers begin as upright buds, but become pendant when mature
(Debuhr 1973). The flower-bearing scape generally ranges from 40-60 cm, but may
grow as tall as 90 cm (Debuhr 1973). On average, each scape bears nine bracts, all of
which have nectar secreting glands (Macfarland 1908). Unlike some confamilial
Sarracenia spp. (Ne’eman et al. 2006), the flowers of D. californica produce no nectar
(Debuhr 1973). Abundant pollen is the only likely reward for pollinators, though a sugar-
rich stigmatic exudate may also attract visitors (Nyoka 2000). Five lanceolate-ovate,
yellow-green sepals hang loosely around five crimson petals. The five petals almost
completely enclose the reproductive whorls, except for windows formed by notches in
adjacent petals, which allow access to the flower’s interior. The windows are level with
the five stigmatic lobes, a feature that may promote the deposition of outcrossed pollen as

6
pollinators initially enter a flower. Twelve-fifteen stamens are located at the base of the
ovary. The bell-shaped ovary is flared towards the stigmas, which has been postulated to
function to guide pollinators away from the stigmas as they exit a flower and thus limit
self-pollination (Schnell 1976). Flowers mature into upright capsules capable of
producing around 2000 seeds (Debuhr 1973). The flowers of D. californica are self-
compatible, but do not self-pollinate autonomously (Elder 1997, Nyoka 2000).
Study Sites
Five seeps located near Scott Mountain and Mt. Eddy, CA were used in this study
(Figure 1, Table 1). The five study sites will hereafter be referred to as SM1, SM2, CL,
N17, and DF. Distance between sites ranged from ~0.1 to 14.5 km. Near the border of
Trinity and Siskiyou Counties, this portion of the Klamath Bioregion represents the
center of D. californica’s range (Debuhr 1973). Common woody associates adjacent to
seeps include Jeffrey pine (Pinus jeffreyi), huckleberry oak (Quercus vaccinifolia),
western azalea (Rhododendron occidentale), white fir (Abies concolor), red fir (Abies
magnifica) and incense cedar (Calocedrus decurrens). Within the seeps, common
associates include white rushlily (Hastingsia alba), California bog asphodel (Narthecium
californicum), Sierra shootingstar (Dodecatheon jeffreyi), marsh marigold (Caltha
leptosepala var. biflora), and Bigelow’s sneezeweed (Helenium bigelovii). Flowering
occurred at all study populations between June 12, 2008 and June 22, 2008, except for
CL where flowering started earlier (June 6, 2008). Study sites were chosen based on
accessibility and population size.
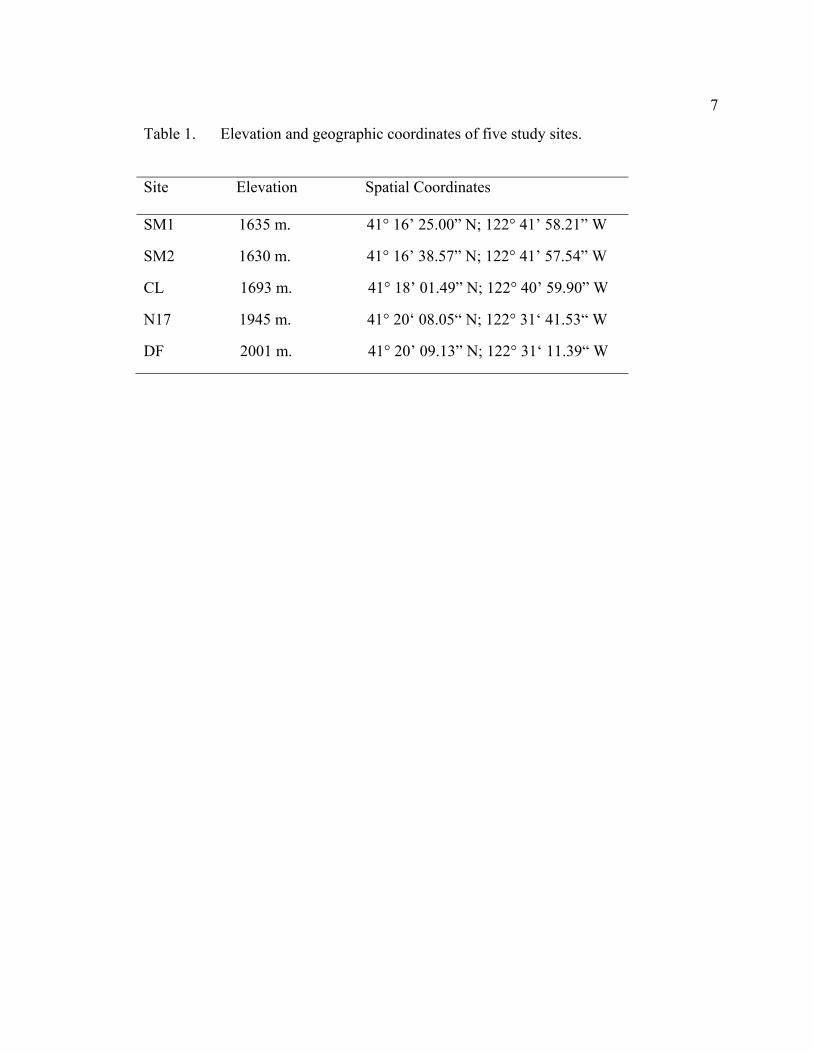
7
Table 1. Elevation and geographic coordinates of five study sites.
Site Elevation Spatial Coordinates
SM1 1635 m. 41° 16’ 25.00” N; 122° 41’ 58.21” W
SM2 1630 m. 41° 16’ 38.57” N; 122° 41’ 57.54” W
CL 1693 m. 41° 18’ 01.49” N; 122° 40’ 59.90” W
N17 1945 m. 41° 20‘ 08.05“ N; 122° 31‘ 41.53“ W
DF 2001 m. 41° 20’ 09.13” N; 122° 31‘ 11.39“ W

8
Figure 1. Map of study area. Triangles represent the five study sites. SM1, SM2,
and CL were near the summit of Scott Mountain, while N17 and DF were near Mt. Eddy. The gray line separates Siskiyou County and Trinity County.

9
Pollinator Surveys
Three observation points were established in each seep in order to monitor
pollinator activity. At these points a series of timed (15 minute) surveys were conducted,
focusing on 13-17 flowers. I sat quietly and motionless while observing the flowers in an
effort not to disturb active pollinators or alter their behavior. Ten 15-minute surveys (2.5
hours total) were conducted during each trip to a field site. Most surveys were made
between 10:00 a.m. and 6:00 p.m., but several were made in the early morning and late
evening. In total, 57.5 hours were devoted to surveys between June 6, 2008 and July 3,
2008. A subset of pollinator visits was timed and filmed with a digital camera. Timing
began when a pollinator entered one of the windows, and ended when it exited a window.
Videos were used to provide detailed accounts of insect visitation.
Mean flower visitation rates (visits/flower/hour) were calculated for each study
site. The expected number of visits a flower received over its lifetime was estimated by
multiplying the flowering period (in days) by the number of hours in a day pollinators
were active (six hrs.) by the visits/hour calculated for each site. D. californica pollinators
were considered to be active for six hours a day because all visits occurred between 10:30
a.m. and 4:30 p.m. Flower lifespan was determined by monitoring the development of 30
tagged buds at each study site. The flowers were synchronously bisexual (i.e., stigma
receptivity and anther dehiscence occurred simultaneously). Flowers were deemed
mature when stigmas were receptive (exudates present) and anthers were dehiscing

10
pollen and were considered past maturity when stigmas withered and ovaries began to
swell and turn upright. By July 1, 2008, flowers were developing fruits at all sites.
Tagged flowers were considered receptive at CL between June 6, 2008 and June 22, 2008
(17 days), while tagged flowers were receptive at SM1, SM2, N17, and DF between June
12, 2008 and June 22, 2008 (11 days). Visitation rates were calculated for each site
independently.
Following each 15 minute census period, five flowers were carefully examined by
spreading apart the sepals and petals to check for animals already present within the
flowers. A total of 1125 flowers were inspected in this way for spiders, spider webs
(either inside or outside the flower), fungus gnats, and thrips. Insects were captured by
aerial netting or by hand, and identified.
Pollinator surveys were also conducted on concurrently flowering plant species.
Approximately 10 hours were devoted to collecting and describing the local pollinator
community. Surveys were conducted by walking in and around seeps and stopping
periodically to observe and collect pollinators on concurrently blooming plants.
Analysis of Pollen Loads
Each insect collected during surveys that carried a visible pollen load was
systematically dabbed with a small cube of glycerin jelly containing basic fuchsin stain
(Kearns and Inouye 1993). Following pollen removal, the jelly was placed on a
microscope slide, melted and covered with a cover slip for analysis. Pollen grains were
identified by comparing them to a series of slides prepared from flowers at each site.

11
Pollination Treatments
In order to test the assumption that fruit and seed set in natural populations of D.
californica are high, 30 unmanipulated flowers at each site were monitored through their
development. Because flowers do not autonomously self-pollinate (Elder 1997, Nyoka
2000), pollinator activity can be estimated based on the reproductive success of
unmanipulated flowers. Fruit and seed set resulting from unmanipulated flowers were
also compared against that of an equal number of hand-pollinated flowers to determine if
plants were experiencing pollen-limitation. If there is no difference in fruit and seed set
between these two treatment groups then we can conclude that natural pollination is
sufficient, i.e., plants were not pollen-limited. Following the appearance of stigmatic
exudates, supplemental pollen was applied twice (separated by one week) to 30 flowers at
each site by rubbing two-three mature anthers directly against stigmatic surfaces. Pollen
used for hand-pollinations was collected from flowers at least five m away in the same
population.
To gauge the relative importance of spiders versus bees as pollinators, 30 flowers
in each seep were emasculated prior to maturity. Bees, which tend to visit multiple
flowers on any given foraging bout, would likely promote xenogamous pollen transfer for
D. californica (though depending on their foraging behavior and flower handling, bees
may cause some level of autogamy as well). If past interpretations of the functional
morphology of flowers hold true (i.e., if ovary shape limits insect mediated self-
pollination) then bees may contribute minimally to autogamous pollen transfer. Spiders,
however, tend to remain resident within single flowers for longer periods of time and

12
would likely contribute primarily to autogamous pollen transfer. Any seeds produced in
the emasculated treatment group must be the result of cross-pollination, thus levels of
cross-pollination and self-pollination can be estimated by comparing the seed set of the
emasculated flowers with the seed set of unmanipulated flowers. Under the assumption
that bees would act mainly as cross-pollinators and spiders as self-pollinators, this
comparison was also intended to show what organisms were likely responsible for pollen
transfer.
A total of 450 flowers were used for fruit and seed set experiments, with 150
flowers in each of the three treatments: hand-pollinated, emasculated, and unmanipulated.
These treatments were spread equally across the five study sites (i.e., 90 flowers at each
site in three treatment groups of 30). Once fruit maturation began, all treatment flowers
were bagged with Reemay® to ensure seeds were not lost when capsules began to
dehisce. This proved to be an unnecessary precaution since all fruits were collected prior
to dehiscence. Fruit set was determined for each site, as well as the number of seeds
produced by each capsule.
Plant Community Context
To characterize floral resources available to pollinators that were active during the
blooming period of D. californica, all concurrently blooming plant species were
identified in and around the field sites (within 150 m). Due to the close proximity of
SM1 and SM2, all coincidentally blooming plants discovered at either site were

13
considered to be present at both sites. In order to quantify floral resources provided by
these plants, percent ground cover was determined in eight m x eight m plots both within
the seeps, as well as 50 m outside each seep in the surrounding uplands. Five eight m
transects, separated by two m from each other, were laid down to establish the plots.
Plots within the seeps were chosen haphazardly, while the corresponding upland plots
were placed 50 m outside each seep at a right angle to the center of the plot within the
seep. Within each eight m x eight m plot, 25 0.5 m x 0.5 m subplots were used to
describe percent ground cover of all coincidentally blooming species. A frame
constructed of PVC pipe with a wire grid (100 squares) was laid over the vegetation and
used to estimate ground cover on a scale of 0-100% for each species in bloom.
Statistical Analyses
Log linear analysis was used to compare the fruit set of the three experimental
treatment groups, with treatment, site and the interaction term included in the model. A
two-way ANOVA was used to compare seed set across all sites, with treatment and site
as the independent variables. Due to a significant interaction term from the two-way
ANOVA (p=0.041), separate one-way ANOVAs were run for each site independently
using treatment as the independent variable. A multiple comparison of means was also
completed, using a Bonferroni adjustment. A Kruskal-Wallis one-way analysis of
variance was used to compare visitation rates across all sites. All statistical analyses
were performed using NCSS (Hintze 2004).

14
RESULTS
Flower Visitors
Over 57.5 hours of surveys, only 38 visits by flying pollinators were observed: 37
by a solitary bee, Andrena nigrihirta, and one by a European honeybee (Apis mellifera)
(Table 2). Estimated visit rates varied from 0.077 (SE=0.025) visits/flower/hour at N17
to 0.016 (SE=0.029) visits/flower/hour at CL, which correspond to 5.08 and 1.60
visits/flower’s lifetime, respectively (Table 3). Visit rates, however, were not
significantly different across all five sites (Kruskal-Wallis χ2=5.72, p=0.22). A. nigrihirta
stayed in individual flowers for extended time periods. Out of 14 timed visits, A.
nigrihirta foraged on a single D. californica flower for an average of approximately two
minutes and eight seconds (128 seconds +/- 12 seconds; Table 4). Eight individuals were
collected immediately following visits, and all carried D. californica pollen on their
scopae. Of these, six carried D. californica pollen exclusively while two carried
heterospecific pollen as well (Asteraceae). A. nigrihirta was the only pollinator collected
found to carry the pollen of D. californica.
Spiders were common on flowers at all five study sites, and were active at all
hours of the day. Whereas 26.2% of all flowers examined contained a spider, 64.8%
showed evidence of spider occupancy (webbing and/or spider present). Thrips were also
present in large numbers at all five sites: 48.4% of flowers contained thrips actively
foraging for pollen (Table 5). Fungus gnats, while frequently encountered in the seeps,
were only observed within D. californica flowers three times.
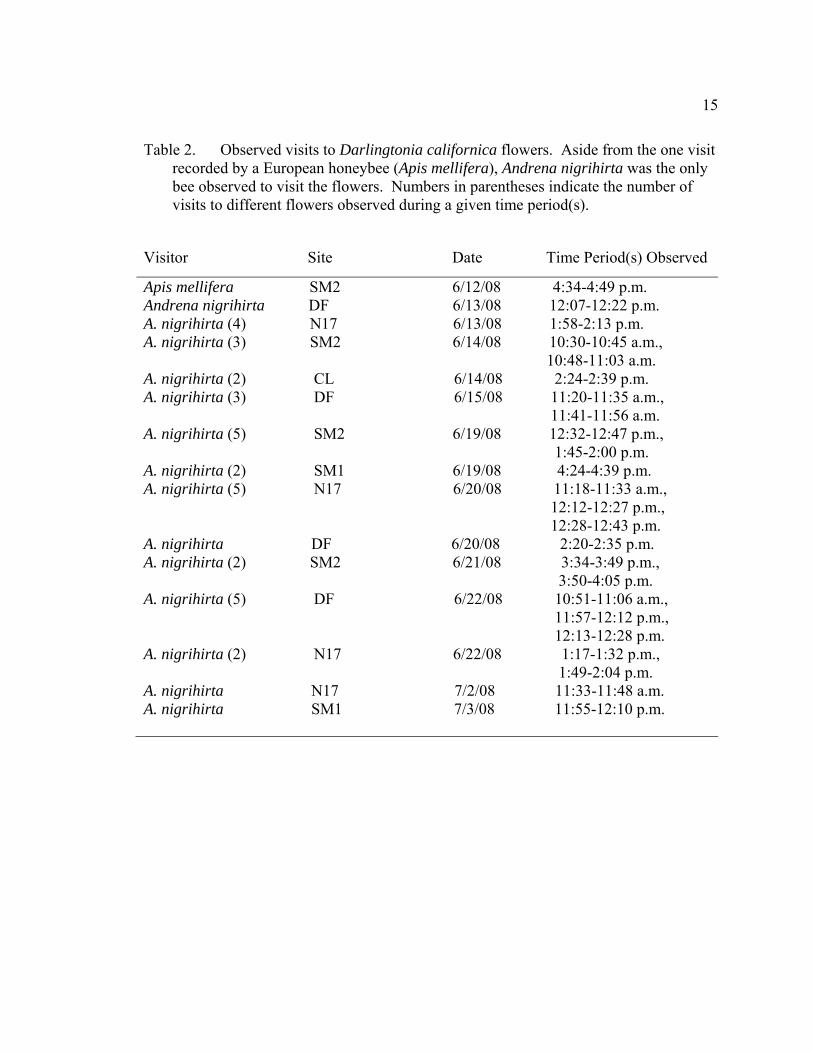
15
Table 2. Observed visits to Darlingtonia californica flowers. Aside from the one visit recorded by a European honeybee (Apis mellifera), Andrena nigrihirta was the only bee observed to visit the flowers. Numbers in parentheses indicate the number of visits to different flowers observed during a given time period(s).
Visitor Site Date Time Period(s) Observed
Apis mellifera SM2 6/12/08 4:34-4:49 p.m. Andrena nigrihirta DF 6/13/08 12:07-12:22 p.m. A. nigrihirta (4) N17 6/13/08 1:58-2:13 p.m. A. nigrihirta (3) SM2 6/14/08 10:30-10:45 a.m., 10:48-11:03 a.m. A. nigrihirta (2) CL 6/14/08 2:24-2:39 p.m. A. nigrihirta (3) DF 6/15/08 11:20-11:35 a.m., 11:41-11:56 a.m. A. nigrihirta (5) SM2 6/19/08 12:32-12:47 p.m., 1:45-2:00 p.m. A. nigrihirta (2) SM1 6/19/08 4:24-4:39 p.m. A. nigrihirta (5) N17 6/20/08 11:18-11:33 a.m., 12:12-12:27 p.m., 12:28-12:43 p.m. A. nigrihirta DF 6/20/08 2:20-2:35 p.m. A. nigrihirta (2) SM2 6/21/08 3:34-3:49 p.m., 3:50-4:05 p.m. A. nigrihirta (5) DF 6/22/08 10:51-11:06 a.m., 11:57-12:12 p.m., 12:13-12:28 p.m. A. nigrihirta (2) N17 6/22/08 1:17-1:32 p.m., 1:49-2:04 p.m. A. nigrihirta N17 7/2/08 11:33-11:48 a.m. A. nigrihirta SM1 7/3/08 11:55-12:10 p.m.
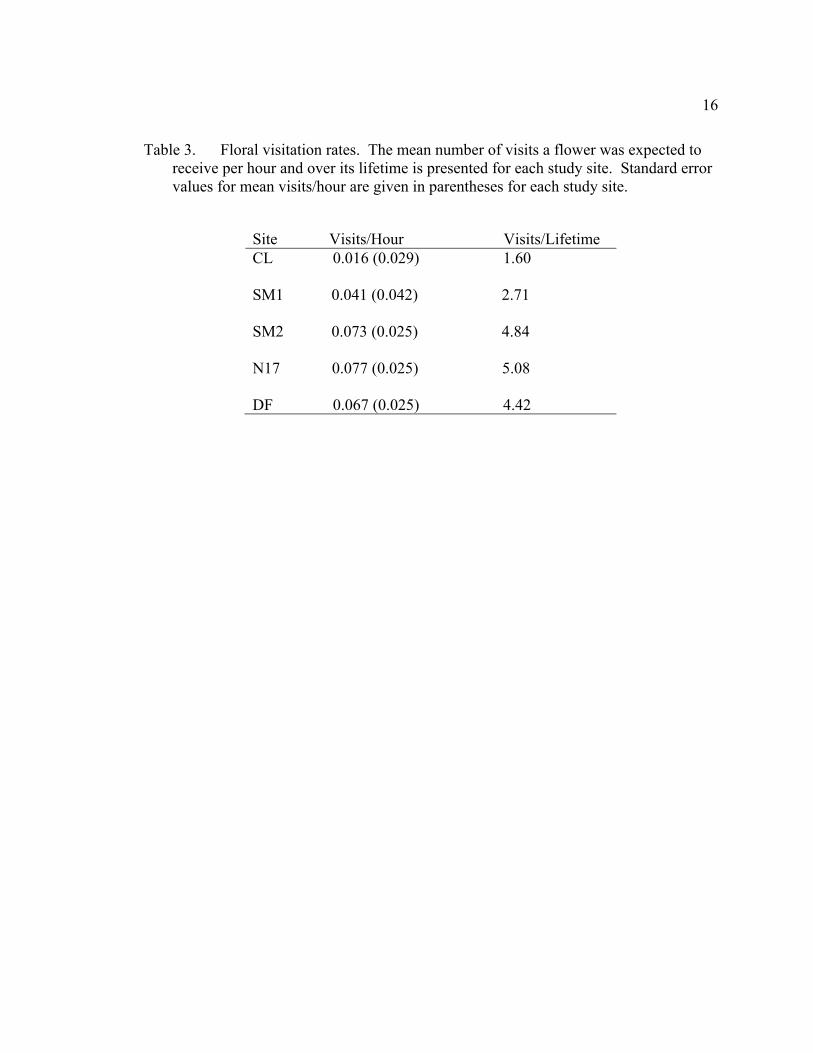
16
Table 3. Floral visitation rates. The mean number of visits a flower was expected to receive per hour and over its lifetime is presented for each study site. Standard error values for mean visits/hour are given in parentheses for each study site.
Site Visits/Hour Visits/Lifetime CL 0.016 (0.029) 1.60 SM1 0.041 (0.042) 2.71 SM2 0.073 (0.025) 4.84 N17 0.077 (0.025) 5.08 DF 0.067 (0.025) 4.42
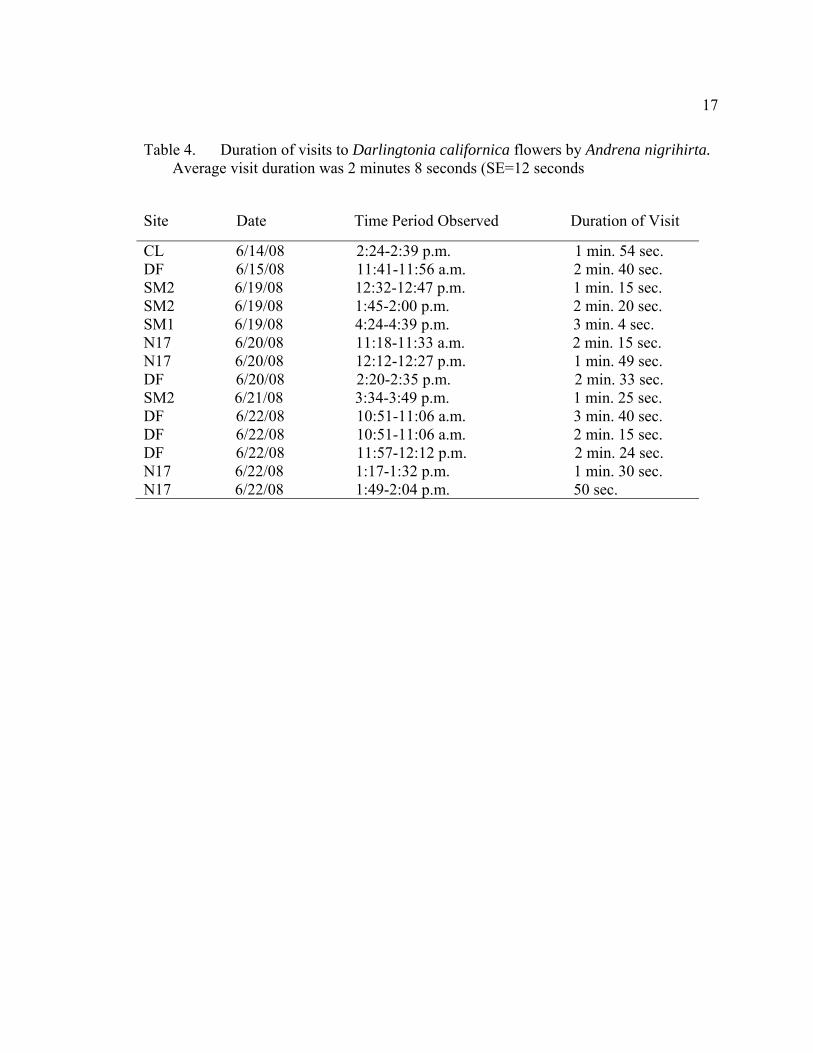
17
Table 4. Duration of visits to Darlingtonia californica flowers by Andrena nigrihirta. Average visit duration was 2 minutes 8 seconds (SE=12 seconds
Site Date Time Period Observed Duration of Visit
CL 6/14/08 2:24-2:39 p.m. 1 min. 54 sec. DF 6/15/08 11:41-11:56 a.m. 2 min. 40 sec. SM2 6/19/08 12:32-12:47 p.m. 1 min. 15 sec. SM2 6/19/08 1:45-2:00 p.m. 2 min. 20 sec. SM1 6/19/08 4:24-4:39 p.m. 3 min. 4 sec. N17 6/20/08 11:18-11:33 a.m. 2 min. 15 sec. N17 6/20/08 12:12-12:27 p.m. 1 min. 49 sec. DF 6/20/08 2:20-2:35 p.m. 2 min. 33 sec. SM2 6/21/08 3:34-3:49 p.m. 1 min. 25 sec. DF 6/22/08 10:51-11:06 a.m. 3 min. 40 sec. DF 6/22/08 10:51-11:06 a.m. 2 min. 15 sec. DF 6/22/08 11:57-12:12 p.m. 2 min. 24 sec. N17 6/22/08 1:17-1:32 p.m. 1 min. 30 sec. N17 6/22/08 1:49-2:04 p.m. 50 sec.

18
Floral Mechanism/ Pollinator Behavior
Detailed observations of visits by A. nigrihirta revealed that the ovary shape of D.
californica promotes stigma contact by bees both as they enter and exit a flower (Figure
2). Immediately above the windows (towards the morphological base of the pendant
flower), the petals overlap and the underlying petal is appressed to the flared portion of
the ovary, which limits the ability of a pollinator the size of A. nigrihirta to enter a
window and crawl directly up onto the ovary on its way to collect pollen (Figure 3). In
between the windows, however, the petals bulge outward (Figure 3), and it is this space
that A. nigrihirta utilized to ascend the ovary to reach the stamens. This convex portion
of each of the five petals is located directly opposite each of the five windows, such that a
pollinator enters a window and walks in a straight line across the stigmas and then onto
the ovary (directed by the convex portion of the petal). The shape of D. californica’s
ovary has previously been thought to guide an insect pollinator away from the receptive
stigmatic surfaces as it exits the flower, thus preventing self-pollination. However, in
exiting the flower, the pollinator was observed to leave in the same fashion as it entered
(guided by petal convexities across the stigmas and out one of the windows, thus likely
effecting autogamy). This behavior was exhibited by multiple individuals, and was
consistent at all sites.

19
a b c d e f g h i j k l m n o p

20
Figure 2. Step by step foraging behavior of A. nigrihirta on a D. californica flower. The bee initially lands on the petals below the windows (a-c) and then enters a window and walks across stigmatic surfaces (d,e). The bee then utilizes the convex portion of one of the five petals to walk onto the ovary and up to collect pollen (f-i). Following pollen collection, the bee uses a petal convexity as before to walk down the ovary, across the stigmas again, and then out one of the windows (j-n). The flower is shown in d-m with one petal removed and half of the two lateral petals removed. F and k show the bee using the convex portion of the petal, which allows the bee to access the stamens.

21
Figure 3. Interior view of a D. californica flower with the lower portion of petals removed. Arrows highlight the distance between the petals and the ovary both immediately above a window (shorter arrow) and in between two adjacent windows (larger arrow). More space is provided for A. nigrihirta in between the windows than above them, which encourages the bee to enter a window and then walk across the stigmatic surfaces. The bee then utilizes the convex portion of the petal opposite the window it entered en route to the flower’s stamens.
22
Table 5. The percentage of examined flowers at each study site that contained one or more of the following: thrips, spiders, and spider webs (either inside or outside the flower). A total of 1125 flowers were individually examined (150 at CL, 225 at SM1, 250 at SM2, 250 at N17, and 250 at DF). “Evidence of Spider” column represents the percentage of examined flowers at each site that had a spider and/or webbing present. Only 3/1125 (0.27%) flowers contained one or more fungus gnats.
Site Web Outside Flw. Web Inside Flw. Spider Present Evidence of Spider Thrips
CL 38.7 13.3 16.7 48.7 25.3 SM1 48.9 39.6 20 61.8 40.9 SM2 47.2 30.4 24.8 57.2 31.6 N17 69.6 20 34 74 75.6 DF 68.8 30.8 31.2 75.6 58.4 TOTAL 56.2 27.7 26.2 64.8 48.4

23
Pollination Treatments
Both fruit and seed production varied depending on treatment. Fruit set was
significantly affected by floral treatment (χ2=37.69, p<<0.01) and treatment effects were
similar across all sites (χ2=0.85, p=0.99). Differences in mean seed production between
the three treatment groups were also evident at all five sites (SM1: F2, 63=65.35, p<<0.01;
SM2: F2, 65=53.76, p<<0.01; CL: F2, 59=52.49, p<<0.01; N17: F2, 58=46.31, p<<0.01; DF:
F2, 57=24.19, p<<0.01; Fig. 3).
Fruit set of D. californica populations near Scott Mountain and Mt. Eddy, CA was
high. Fruit production by unmanipulated flowers (76%) was marginally comparable to
that of hand-pollinated flowers (96%) (χ2=3.50, p=0.0615). Though fruit set by
unmanipulated flowers did not indicate significant pollen-limitation, seed production did.
Hand-pollinated flowers produced significantly more seeds than unmanipulated flowers
at each of the five study sites (Fig. 3), indicating that plants in all five populations
experienced pollen-limitation. On average, unmanipulated flowers produced 674
(n=114) seeds per capsule, versus 1422 (n=144) by hand-pollinated flowers and 439
(n=59) by emasculated flowers.
Self- and cross-pollination both contribute to D. californica reproductive success.
Emasculated flowers produced fruit and seed at all five sites, indicating that cross-
pollination occurred. However, overall fruit set of emasculated flowers (39%) was
significantly lower than that of unmanipulated flowers (76%) (χ2=17.79, p<0.001),
highlighting the importance of autogamous pollen transfer for fruit production.

24
Unmanipulated flowers produced significantly more seeds, on average, than emasculated
flowers at SM1 and SM2, but there was no significant difference found between these
two treatment groups at the remaining three sites (Fig. 3). Average seed production by
unmanipulated flowers was always higher than that of emasculated flowers, regardless of
statistical significance, suggesting that cross-pollination cannot account for all of the
seeds that were produced by unmanipulated flowers. Therefore, fruit and seed production
of naturally pollinated flowers were the result of both autogamous and xenogamous
pollen transfer.
Plant/ Insect Community Context
Plant communities in and around D. californica seeps were diverse. A total of 51
angiosperm species, all with blooming periods that at least partially overlapped that of D.
californica, were found at the study sites (Appendix 1). The plant communities at all five
sites were similar, with approximately 1/3 of all observed taxa found at every site. 42
concurrently blooming species were found at SM1 and SM2 combined, 35 at DF, 32 at
N17, and 27 at CL.
Species richness and percent ground cover were greater in the mesic areas within
the seeps compared to adjacent xeric habitats (Table 6, Appendix 2). In the seeps, D.
californica covered 31.7% of the ground and concurrently blooming plants covered
34.1% of the ground. In the more xeric areas surrounding the seeps, an average of 14.5%
of the ground was occupied by coflowering species. As judged by cover values, more
floral resources were available within the seeps than outside them.

25
Eighty eight pollinators (including the eight A. nigrihirta individuals collected
after visits to D. californica flowers) were collected from the community, representing
four orders of insects (Coleoptera, Diptera, Hymenoptera, and Lepidoptera) (Table 8).
Hymenopterans were the most abundant insect floral visitors, representing 62% of all
collected insect pollinators (n=49). Mason bees (Osmia spp.) were the most frequently
collected pollinator (n=23), followed by andrenid bees (Andrena spp., n=17) and
bumblebees (Bombus spp., n=9). A. nigrihirta was not collected on any plant other than
D. californica, but one individual was collected in flight (i.e., not on a flower) that carried
D. californica pollen.
26
Table 6. The percentage of ground covered by Darlingtonia californica and co-flowering plant species. The total percentage of ground covered by angiosperms both within and outside (50m) the seeps are shown in the first two columns. The percentage of ground covered by Darlingtonia californica within the seeps is also presented.
Site % cover within % cover outside % D. californica cover within
SM1 49.4 35.9 17.8
SM2 78.2 8.8 38.3
CL 63.6 5.4 34.7
DF 66.6 7.2 42.4
N17 71.2 15.2 25.4
TOTAL 65.8 14.5 31.7
27
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
SM1 SM2 N17 CL DF
Frui
t set
(%)
Emasculated Unmanipulated Hand-Pollinated
0
300
600
900
1200
1500
1800
SM1 SM2 N17 CL DF
Ave
rage
# o
f see
ds p
er fr
uit
Emasculated Unmanipulated Hand-Pollinated
Figure 4. Fruit and seed production by three treatment groups (emasculated, unmanipulated, and hand-pollinated flowers) at each
study site. Top: Fruit set (%) of three treatment groups at each field site. Emasculated flowers produced significantly fewer fruits than unmanipulated flowers. No significant difference was found between fruit production of unmanipulated vs. hand-pollinated flowers (χ2=3.50, p=0.0615). Bottom: Average number of seeds produced per capsule from three treatment groups at each field site. Different letters above bars indicate group means are significantly different.
a
b
c
a
b
c
a
a
b
aa
b
a a
b

28
Table 7. Pollinators collected at or near study sites. Order and family is presented for all pollinators (and genus for Hymenopterans). Numbers in parentheses indicate the number of individual pollinators collected from each group
Diptera (18) Anthomyiidae (1) Asilidae (1) Bombyliidae (6) Callophoridae (1) Muscidae (1) Syrphidae (8) Colleoptera (5) Hymenoptera (58) Andrenidae Andrena nigrihirta (9) Other Andrena spp. (7) Apidae Apis (1) Bombus (9) Colletidae Hylaeus (1) Halictidae Agapostemon (2) Lasioglossum (4) Megachilidae Osmia (23) Vespidae (2) Lepidoptera (7)

29
DISCUSSION
Near the summits of Scott Mountain and Mount Eddy, CA, populations of
Darlingtonia californica are pollinated by the solitary bee Andrena nigrihirta, with
additional pollination likely provided by spiders and small insects. Direct observations of
floral visits and the results of pollination treatments suggest pollination by both bees and
spiders. The success of emasculated flowers indicated that cross-pollination occurred at
all sites, implicating bees as pollen vectors. However, considering that average fruit and
seed production was always higher in unmanipulated versus emasculated flowers, some
level of autogamy must have occurred at all sites as well. Based on observations of visits
by bees and spiders, both organisms have the potential to effect self-pollination. Because
both self-pollination and cross-pollination contributed to fruit and seed production, both
spiders and bees likely contributed to D. californica reproductive success.
Both fruit and seed production by emasculated flowers implicate bees as pollen
vectors. However, because bees contact stigmatic surfaces when they exit a flower, they
have the ability to effect self-pollination and likely contributed to autogamous pollen
transfer as well. In light of this, the great majority of pollination may be mediated
exclusively by A. nigrihirta. Spiders, which were highly abundant on flowers and have
already been shown to effect autogamy in D. californica (Nyoka 2000), likely further
contributed to fruit and seed production via self-pollination. Spiders were constantly
seen patrolling and building webs on flowers, and in several instances webs were

30
constructed within flowers linking anthers and stigmas and were completely dusted with
pollen, indicating spider facilitated pollen movement within flowers. Thrips, which also
tend to remain resident within single flowers, were also present in large numbers and
were observed within D. californica flowers at all sites. Though fungus gnats do not
appear to be major pollinators near Scott Mountain and Mt. Eddy, CA, Nyoka and
Ferguson (1999) collected fungus gnats in southwestern Oregon that carried the pollen of
D. californica. Repeated floral visits by small insects like thrips and fungus gnats have
been shown to contribute substantially to fruit set in other angiosperms (Mesler et al.
1980, Zamora 1999), and may contribute to seed set in D. californica as well.
The lifespan of D. californica flowers is much longer than most temperate zone
flowering plants, and near sea level D. californica populations may bloom for as long as
48 days (Debuhr 1973). Assuming an active pollinator community, extended flower
longevity will result in an increase in pollinator visits (Primack 1985, Ashman and
Schoen 1994, Rathke 2003). The estimated visitation rates from this study show that
visits to D. californica flowers by its main insect pollinator, A. nigrihirta, are relatively
few. Despite low visit rates, flowers are still likely to receive at least one visit by A.
nigrihirta due to the flowers’ prolonged lifespans. A long blooming period not only
promotes visits by effective cross pollinators like bees, but also allows for repeated visits
by less effective pollinators like spiders and thrips. The longevity of D. californica
flowers may be an adaptation to deal with an unpredictable pollinator community and
favors pollination by several groups of organisms.

31
Plant species with overlapping flowering periods may either facilitate each other’s
pollination (Gross et al. 2000, Bruno et al. 2003, Moeller 2004) or inhibit it through
competition for pollinator services (Vamosi et al. 2006). Darlingtonia californica plants
at all five sites showed evidence of pollen-limitation of seed production. Due to the
abundance of heterospecific taxa with overlapping flowering times and the scarcity of
large bodied pollinators found to carry the pollen of D. californica, it is likely that these
populations are experiencing a reduction in seed set due to competition for floral visitors.
Other than spiders and thrips, A. nigrihirta was the only pollinator seen to consistently
visit D. californica flowers, and was the only insect taxon collected found to carry the
pollen of D. californica.
How close is the relationship between D. californica and A. nigrihirta? Though
many species of Andrena are specialists (Larsson 2005, Diamond et al. 2006), only
utilizing the pollen from particular groups of angiosperms, many are also generalist
foragers using a wide range of pollen hosts (Nyoka 2000). Across its range, which spans
North America and greatly exceeds that of D. californica, A. nigrihirta is a generalist that
has been observed to visit flowers from a diverse array of plants, including members of
Portulacaceae (Motten et al. 1982), Fabaceae (Tepedino et al. 1995), and Ericaceae
(Barry Rice, personal communication), along with D. californica. Three individuals of A.
nigrihirta were collected that carried both D. californica and Asteraceae pollen,
indicating that A. nigrihirta is utilizing floral resources from multiple species of
flowering plants. As the general biology and life history of A. nigrihirta is still poorly
described, future studies are needed to identify what floral resources are used by A.

32
nigrihirta and how much these bees rely on resources provided by D. californica in the
Pacific Northwest. Though D. californica is not considered threatened or endangered by
state or federal government, the California Native Plant Society has placed it on their
watch list of species with limited distributions (Skinner and Pavlik 1994). Because D.
californica may rely on A. nigrihirta for reproductive success, both organisms should be
considered in management efforts aimed at conserving present D. californica
populations.
Pollinator communities have been known to fluctuate in composition and
abundance from year to year (Wolfe 1988, Price et al. 2005), and past researchers trying
to determine the pollinators of D. californica may have inadvertently chosen time periods
when bees were largely absent. Others have witnessed visitation of D. californica by A.
nigrihirta as well, but only on a single day over the course of nine years observing the
flowers (Barry Rice, personal communication). Whether there is synchronicity between
the emergence of A. nigrihirta and the flowering period of D. californica throughout its
range needs to be determined. For D. californica populations near sea level, A. nigrihirta
may only be active for a fraction of the time D. californica is in bloom. Also, there may
be geographic regions where D. californica populations exist, but those of A. nigrihirta
do not. Either of these possibilities could potentially hinder efforts to witness floral
visitation. Alternatively, the foraging behavior of A. nigrihirta on D. californica flowers
may have limited prior pollinator sightings. After entering a D. californica flower the
bee is out of sight until emerging up to four minutes later. Therefore, if the initial

33
approach of the bee is not witnessed, an observer may never realize a flower had been
entered.
The bell-shaped ovary of D. californica was previously thought to prevent self-
pollination from occurring by guiding an insect pollinator away from stigmatic surfaces
as it exits a flower. However, following observations of visits by A. nigrihirta, it was
determined that the shape of the ovary actually promotes stigma contact both when bees
enter and exit flowers. This system provides an excellent example of the importance of
identifying and observing pollinators in order to truly understand the functional
significance of a plant’s floral morphology.
Future Research
While the present study sought to estimate the relative contributions of spiders vs.
other pollinators to D. californica seed set, these contributions were not directly
quantified. To better understand the importance of spiders as pollinators, future studies
should develop spider exclusion treatments that would prevent spiders from colonizing
flowers. By comparing the seed set of open-pollinated flowers with those that exclude
the presence of spiders, the contribution of spiders to seed and fruit production would
become evident. Spider exclusions may prove to be difficult, however. Not only can
some spiders jump relatively great distances (e.g. salticid spiders), but smaller spiders can
also “fly” by releasing a silken strand into the wind, which may then carry them to a new
location (potentially a D. californica flower). Therefore, simply coating the scape with a

34
sticky substance to limit cursorial colonization of flowers may not provide an adequate
barrier to all spiders. Further development of this method is needed.
There are a number of interesting ecological questions that have yet to be
addressed regarding D. californica pollination. For instance, how do spiders occupying
D. californica flowers interact with bees? Does the presence of spiders within a flower
deter visitation by bees, or do bees frequently fall victim to lurking spiders, and what
bearing does this have on D. californica reproductive success? Over the course of floral
observations conducted in this study, A. nigrihirta was seen “buzzing” flowers, i.e.
approaching flowers but not entering them, more frequently than entering flowers (37
flowers visited, 50 flowers buzzed). This behavior could be the result of floral marking
by bees, which may be done to alert future visitors of resource availability (Schmitt and
Bertsch 1990, Goulson et al. 2001), but may also be the result of altered foraging
behavior due to the presence of flower-occupying spiders (Bruce et al. 2005, Goncalves-
Souza 2008). Also, do the same floral traits that promote pollen deposition on stigmatic
surfaces by bees (shape of ovary, etc.) also promote pollen deposition by spiders, or
should we expect divergence of floral morphology in D. californica populations that
occur in areas where A. nigrihirta is absent over time? As we seek to explain the
adaptive significance of D. californica’s floral traits, we need to understand, in greater
detail, the effects of these multi-species interactions on trait selection.

35
LITERATURE CITED
Ashman, T. L. and D. J. Schoen. 1994. How long do flowers live? Nature 371: 788-791.
Austin, R. M. 1875-1877. Letters to W. M. Canby on Darlingtonia. The Society of Natural History of Delaware, Wilmington, DE: Wilmington Institute Free Library.
Bruce, M. J., A. M. Heiling, and M. E. Herberstein. 2005. Spider signals: are web decorations visible to birds and bees? Biology Letters 1: 299-302.
Bruno, J. F., J. J. Stachowicz, and M. D. Bertness. 2003. Inclusion of facilitation into ecological theory. Trends in Ecology & Evolution 18: 119-125.
Crepet, W. L., and K. J. Niklas. 2009. Darwin’s second “abominable mystery”: why are there so many angiosperm species? American Journal of Botany 96: 366-381.
Debuhr, L. E. 1973. Distribution and reproductive biology of Darlingtonia californica. Master’s thesis. Claremont Graduate School, Claremont, CA.
Diamond, A. R., D. R. Folkerts, and R. S. Boyd. 2006. Pollination biology, seed dispersal, and recruitment in Rudbeckia auriculata (Perdue) Kral, a rare southeastern endemic. Castanea 71: 226-238.
Elder, C. L. 1994. Reproductive biology of the California pitcher plant (Darlingtonia californica). Fremontia 22: 29-30.
Elder, C. L. 1997. Reproductive biology of Darlingtonia californica. Master’s thesis. Humboldt State University, Arcata, CA.
Fenster, C. B., W. S. Armbruster, and P. Wilson. 2004. Pollination syndromes and floral specialization. Annual Review of Ecology, Evolution and Systematics 35: 375-403.
Forrest, J. and J. D. Thomson. 2009. Pollinator experience, neophobia and the evolution of flowering time. Proceedings of the Royal Society Biological Sciences Series B 276: 935-943.
Goncalves-Souza, T., P. M. Omena, J. C. Souza, and G. Q. Romero. 2008. Trait-mediated effects on flowers: artificial spiders deceive pollinators and decrease plant fitness. Ecology 89: 2407-2413.

36
Goulson, D. 1999. Foraging strategies of insects for gathering nectar and pollen, and implications for plant ecology and evolution. Perspectives in Plant Ecology Evolution and Systematics 2: 185-209.
Goulson, D., J. W. Chapman, W. O. H. Hughes. 2001. Discrimination of unrewarding flowers by bees; direct detection of rewards and use of repellent scent marks. Journal of Insect Behavior 14: 669-678.
Gross, C. L., D. A. Mackay, and M. A. Whalen. 2000. Aggregated flowering phenologies among three sympatric legumes: the degree of non-randomness and the effect of overlap on fruit set. Plant Ecology 148: 13-21.
Hintze, J. 2004. NCSS and PASS. Number Cruncher Statistical Systems. Kaysville, Utah.
Hu, S., D. L. Dilcher, D. M. Jarzen, and D. W. Taylor. 2008. Early steps of angiosperm-pollinator coevolution. Proceedings of the National Academy of Sciences of the United States of America 105: 240-245.
Juniper, B. E., R .J. Robbins and D. M. Joel. 1989. The carnivorous plants. Academic Press, London. 353 pp.
Kearns, C. A. and D. W. Inouye. 1993. Techniques for pollination biologists. University Press of Colorado, Niwot. 583 pp.
Larsson, M. 2005. Higher pollinator effectiveness by specialist than generalist flower-visitors of unspecialized Knautia arvensis (Dipsacaceae). Oecologia 146: 394-403.
Macfarlane, J. M. 1908. Sarraceniaceae. In A. Engler [ed.], Das Pflanzenreich. Regni vegetabilis conspectus, vol. IV, pp. 1–39. Verlag von WilhelmEngelmann, Leipzig, Germany.
Mesler, M. R., J. D. Ackerman and K. L. Lu. 1980. The effectiveness of fungus gnats as pollinators. American Journal of Botany 67: 564-567.
Moeller, D. A. 2004. Facilitative interactions among plants via shared pollinators. Ecology 85: 3289‐3301.
Motten, A. F., D. R. Campbell, D. E. Alexander, and H. L. Miller. 1981. Pollination effectiveness of specialist and generalist visitors to a North Carolina population of Claytonia virginica. Ecology 62: 1278-1287.

37
Ne’eman, C., G. Ne’eman and A. M. Ellison. 2006. Limits to reproductive success of Sarracenia purpurea (Sarraceniaceae). American Journal of Botany 93: 1660-1666.
Nyoka, S. E., and C. Ferguson. 1999. Pollinators of Darlingtonia californica Torr., the California pitcher plant. Natural Areas Journal 19: 386-391.
Nyoka, S.E. 2000. The spider and the fly: a proposed pollination scenario for Darlingtonia californica, the California pitcher plant. Field research grant report for the Native Plant Society of Oregon.
Pauw, A., J. Stofberg, R. J. Waterman. 2008. Flies and flowers in Darwin’s race. Evolution 63: 268-279.
Price, M.V., N. M. Waser, R. E. Irwin, D. R. Campbell, and A. K. Brody. 2005. Temporal and spatial variation in pollination of a montane herb: a seven-year study. Ecology 86: 2106-2116.
Primack, R. B. 1985. The longevity of individual flowers. Annual Review of Ecology and Sytematics 16: 15-37.
Rathcke, B. J. 2003. Floral longevity and reproductive assurance: seasonal patterns and an experimental test with Kalmia latifolia (Ericaceae). American Journal of Botany 90: 1328-1332.
Renner, S. S. 1989. Floral biological observations on Heliamphora tatei (Sarraceniaceae) and other plants from Cerro de la Neblina in Venezuela. Plant Systematics and Evolution 163: 21-29.
Schmitt, U., and A. Bertsch. 1990. Do foraging bumblebees scent-mark food sources and does it matter? Oecologia 82: 137-144.
Schnell, D. E. 1976. Carnivorous plants of the U. S. and Canada. John F. Blair, Winston- Salem. 125 pp.
Skinner, M. W. and B. M. Pavlik. 1994. California Native Plant Society’s inventory of rare and endangered vascular plants of California. Special publication No. 1. Fifth Edition. California Native Plant Society, Sacramento, CA. 338 pp.
Tepedino, V. J., T. L. Griswold, and W. R. Bowlin. 1995. Pollinator sharing by three sympatric milkvetches including the endangered species Astragalus montii. Great Basin Naturalist 55: 19-25.

38
Thomas, K. and D. M. Cameron. 1986. Pollination and fertilization in the pitcher plant Sarracenia purpurea. American Journal of Botany 73: 678.
Valdivia, C. E., and H. M. Niemeyer. 2006. Do floral syndromes predict specialization in plant pollination systems? Assessment of diurnal and nocturnal pollination of Escallonia myrtoidea. New Zealand Journal of Botany 44: 135-141.
Vamosi, J. C., T. M. Knight, J. A. Steets, S. J. Mazer, M. Burd, and T. L. Ashman. 2006. Pollination decays in biodiversity hotspots. Proceedings of the National Academy of Sciences of the United States of America 103: 956-961.
Vogel, S. 2006. Floral syndromes: empiricism versus typology. Botanische Jahrbuecher fuer Systematik Pflanzengeschichte und Pflanzengeographie 127: 5-11.
Vogelei, A., and R. Greissl. 1989. Survival strategies of the crab spider Thomisus onustus Walckenaer 1806 (Chelicerata, Arachnida, Thomisidae). Oecologia 80: 513-515.
Whittaker, R. H. 1954. The ecology of serpentine soils IV. The vegetational response to serpentine soils. Ecology 35: 275-288.
Wolfe, L. M., S. C. H. Barrett. 1988. Temporal changes in the pollinator fauna of tristylous Pontederia cordata, an aquatic plant. Canadian Journal of Zoology 66: 1421-1424.
Zamora, R. 1999. Conditional outcomes of interactions: the pollinator-prey conflict of an insectivorous plant. Ecology 80: 786-795.
Zhang, L., S. C. H. Barrett, J. Y. Gao, J. Chen, W. W. Cole, Y. Liu, Z. L. Bai, and Q. J. Li. 2005. Predicting mating patterns from pollination syndromes: the case of ‘sapromyophily’ in Tacca chantrieri (Taccaceae). American Journal of Botany 92: 517-524.
39
Appendix A. Plant species in coincident bloom with Darlingtonia californica near Scott Mountain and Mt. Eddy, CA.
Species Name Family Habitat Flw. First Obs. Fruit First Obs. Sites Found
Lomatium sp. Apiaceae xeric
6/22/2008
7/12/2008
CL, SM1, SM2
Achillea millefolium Asteraceae
mesic
6/19/2008
7/12/2008
SM1, SM2, CL, DF, N17
Aster alpigenus
Asteraceae
mesic 6/20/2008 7/1/2008
SM1,SM2, N17,DF
Eriophyllum lanatum
Asteraceae
xeric
6/22/2008
7/12/2008
N17, DF
Helenium bigelovii
Asteraceae
mesic
6/22/2008
7/12/2008
SM1, SM2, CL
Senecio integerrimus var.exaltatus
Asteraceae
xeric
6/6/2008
7/1/2008
SM1,SM2, CL, N17,DF
Senecio triangularis
Asteraceae
xeric
6/19/2008
7/1/2008
SM1,SM2, CL, N17,DF
Taraxacum officinale
Asteraceae
mesic
6/6/2008
7/1/2008
SM1,SM2, CL, N17, DF
Arenaria congesta
Caryophyllaceae
xeric
6/22/2008
7/12/2008 N17
Pseudostellaria jamesiana
Caryophyllaceae
xeric
6/22/2008
7/12/2008
SM1, SM2, DF, N17
Silene lemmonii
Caryophyllaceae
xeric
6/22/2008
7/12/2008
SM1, SM2, CL N17, DF
Eriophorum criniger
Cyperaceae
mesic
6/21/2008
7/11/2008
SM1,SM2, CL, N17, DF
Arctostaphylos sp.
Ericaceae
xeric
6/6/2008
7/1/2008
SM1, SM2, CL
Rhododendron occidentale
Ericaceae
xeric
6/6/2008
7/12/2008 SM1, SM2
Lupinus croceus
Fabaceae
xeric
6/22/2008
7/12/2008 N17,DF
Lotus pinnatus
Fabaceae
mesic
6/22/2008
7/12/2008
SM1,SM2, CL, N17,DF

40
Appendix A. Plant species in coincident bloom with Darlingtonia californica near Scott
Mountain and Mt. Eddy, CA. Species Name Family Habitat Flw. First Obs. Fruit First Obs. Sites Found Trifolium longipes
Fabaceae
mesic
6/19/2008
7/1/2008
SM1,SM2, CL, N17,DF
Ribes roezlii
Grossulariaceae
xeric
6/19/2008
7/1/2008
SM1,SM2, N17
Sisyrinchium idahoense ssp.occidentale
Iridaceae
mesic
6/6/2008
7/4/1998
SM1,SM2, CL, N17, DF
Prunella vulgaris var. lanceolata
Lamiaceae
mesic
6/22/2008
7/12/2008
SM1, SM2, DF
Allium validum Liliaceae mesic 6/22/2008 7/12/2008 N17 Calochortus nudus
Liliaceae
xeric
6/19/2008
7/11/2008
SM1,SM2,CL, N17, DF
Calochortus tolmiei
Liliaceae
xeric
6/19/2008
7/11/2008
SM1, SM2, CL, N17, DF
Dichelostemma multiflorum
Liliaceae
xeric
6/22/2008
7/12/2008 SM1, SM2
Hastingsia alba
Liliaceae
mesic
6/22/2008
7/12/2008
SM1,SM2,CL,N17, DF
Lilium pardalinum ssp. shastense
Liliaceae
xeric
6/22/2008
SM1,SM2, CL, N17, DF
Narthecium californicum Liliaceae
mesic
6/22/2008
7/12/2008 SM1, SM2
Xeropyhylum tenax
Liliaceae
xeric
6/19/2008
7/1/2008 SM1, SM2
Sidalcea glaucescens Malvaceae
xeric
6/22/2008
7/12/2008
SM1, SM2, N17
Sidalcea oregana
Malvaceae
mesic
6/22/2008
7/12/2008
SM1, SM2, N17, DF
Epilobium ciliatum ssp. ciliatum Onagraceae
mesic
6/22/2008
7/12/2008
SM1, SM2, DF
Corallorhiza maculata Orchidaceae
xeric
6/22/2008
SM1, SM2
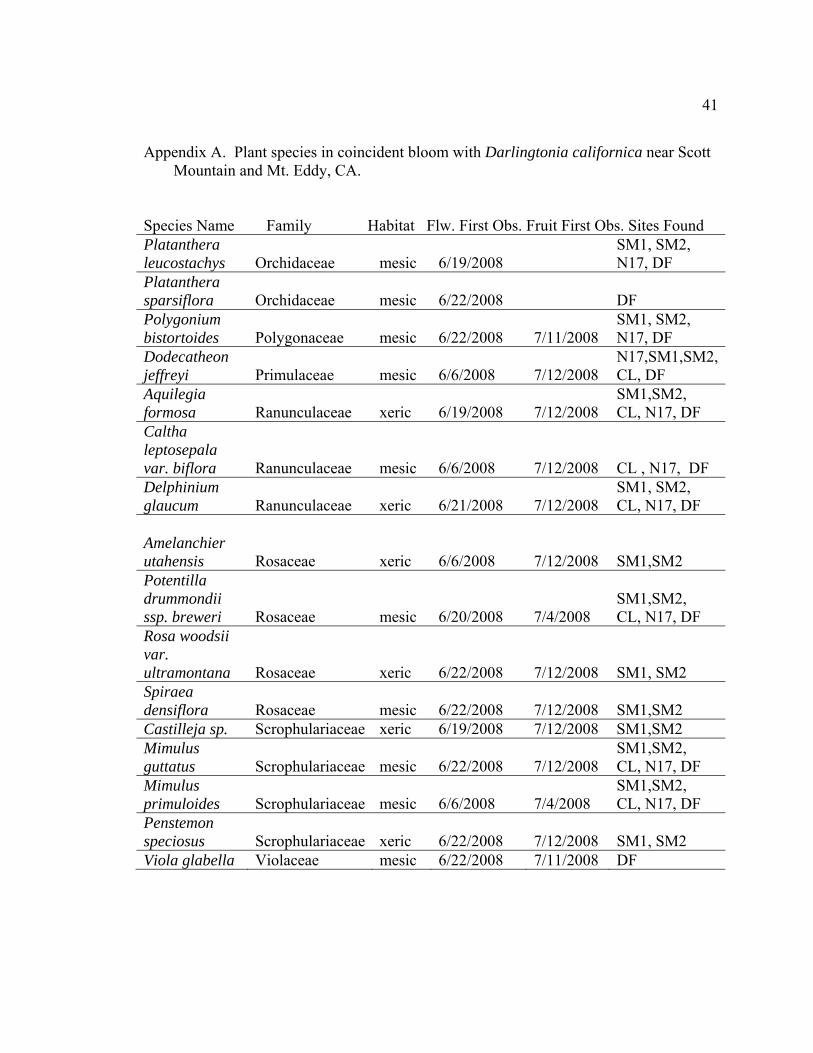
41
Appendix A. Plant species in coincident bloom with Darlingtonia californica near Scott
Mountain and Mt. Eddy, CA. Species Name Family Habitat Flw. First Obs. Fruit First Obs. Sites Found Platanthera leucostachys Orchidaceae
mesic
6/19/2008
SM1, SM2, N17, DF
Platanthera sparsiflora Orchidaceae
mesic
6/22/2008
DF
Polygonium bistortoides Polygonaceae
mesic
6/22/2008
7/11/2008
SM1, SM2, N17, DF
Dodecatheon jeffreyi Primulaceae
mesic
6/6/2008
7/12/2008
N17,SM1,SM2,CL, DF
Aquilegia formosa Ranunculaceae
xeric
6/19/2008
7/12/2008
SM1,SM2, CL, N17, DF
Caltha leptosepala var. biflora Ranunculaceae
mesic
6/6/2008
7/12/2008 CL , N17, DF
Delphinium glaucum Ranunculaceae
xeric
6/21/2008
7/12/2008
SM1, SM2, CL, N17, DF
Amelanchier utahensis Rosaceae
xeric
6/6/2008 7/12/2008 SM1,SM2
Potentilla drummondii ssp. breweri Rosaceae mesic
6/20/2008
7/4/2008
SM1,SM2, CL, N17, DF
Rosa woodsii var. ultramontana Rosaceae
xeric
6/22/2008
7/12/2008 SM1, SM2
Spiraea densiflora Rosaceae
mesic
6/22/2008
7/12/2008 SM1,SM2
Castilleja sp. Scrophulariaceae xeric 6/19/2008 7/12/2008 SM1,SM2 Mimulus guttatus Scrophulariaceae
mesic
6/22/2008
7/12/2008
SM1,SM2, CL, N17, DF
Mimulus primuloides Scrophulariaceae
mesic
6/6/2008
7/4/2008
SM1,SM2, CL, N17, DF
Penstemon speciosus Scrophulariaceae
xeric
6/22/2008
7/12/2008 SM1, SM2
Viola glabella Violaceae mesic 6/22/2008 7/11/2008 DF
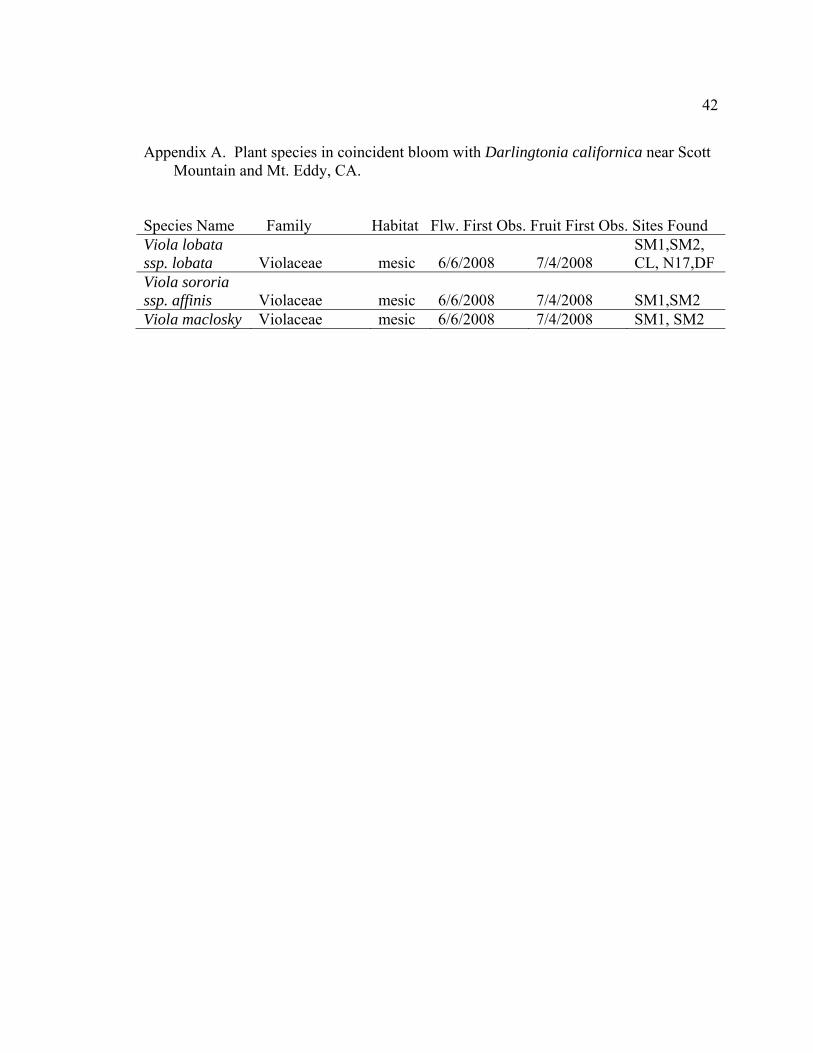
42
Appendix A. Plant species in coincident bloom with Darlingtonia californica near Scott
Mountain and Mt. Eddy, CA. Species Name Family Habitat Flw. First Obs. Fruit First Obs. Sites Found Viola lobata ssp. lobata Violaceae
mesic
6/6/2008
7/4/2008
SM1,SM2, CL, N17,DF
Viola sororia ssp. affinis Violaceae
mesic
6/6/2008
7/4/2008
SM1,SM2
Viola maclosky Violaceae mesic 6/6/2008 7/4/2008 SM1, SM2
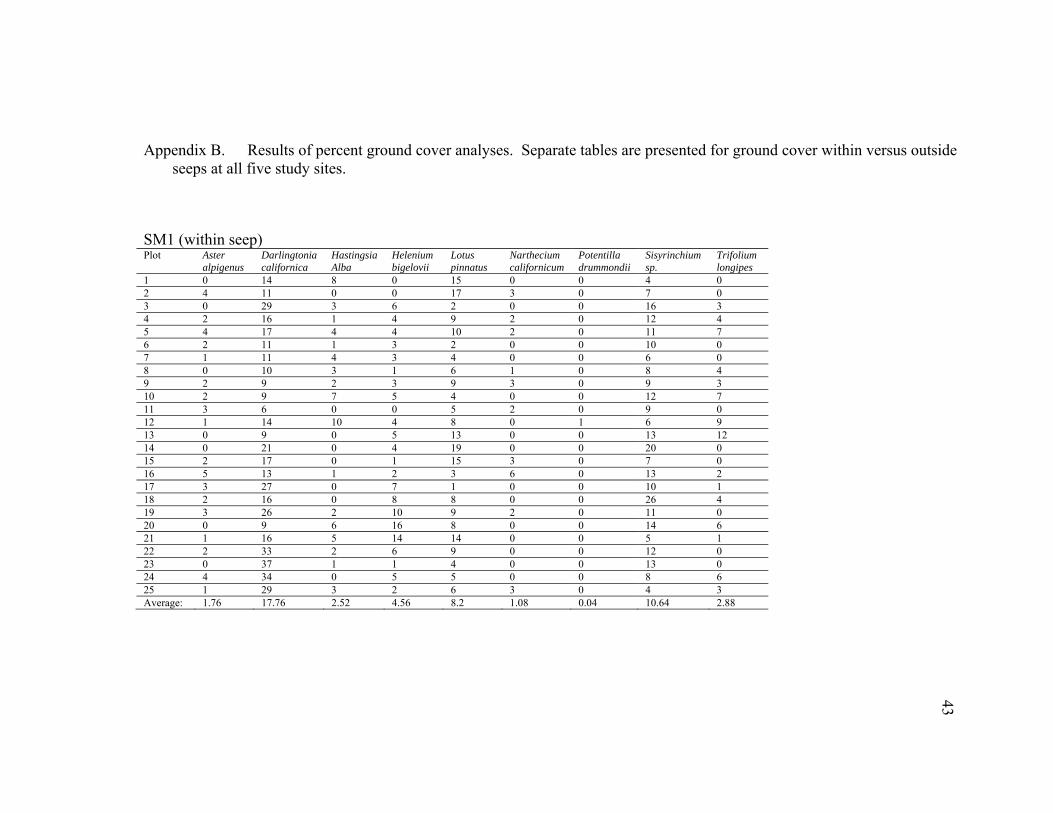
43
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside seeps at all five study sites.
SM1 (within seep) Plot Aster
alpigenus Darlingtonia californica
Hastingsia Alba
Helenium bigelovii
Lotus pinnatus
Narthecium californicum
Potentilla drummondii
Sisyrinchium sp.
Trifolium longipes
1 0 14 8 0 15 0 0 4 0 2 4 11 0 0 17 3 0 7 0 3 0 29 3 6 2 0 0 16 3 4 2 16 1 4 9 2 0 12 4 5 4 17 4 4 10 2 0 11 7 6 2 11 1 3 2 0 0 10 0 7 1 11 4 3 4 0 0 6 0 8 0 10 3 1 6 1 0 8 4 9 2 9 2 3 9 3 0 9 3 10 2 9 7 5 4 0 0 12 7 11 3 6 0 0 5 2 0 9 0 12 1 14 10 4 8 0 1 6 9 13 0 9 0 5 13 0 0 13 12 14 0 21 0 4 19 0 0 20 0 15 2 17 0 1 15 3 0 7 0 16 5 13 1 2 3 6 0 13 2 17 3 27 0 7 1 0 0 10 1 18 2 16 0 8 8 0 0 26 4 19 3 26 2 10 9 2 0 11 0 20 0 9 6 16 8 0 0 14 6 21 1 16 5 14 14 0 0 5 1 22 2 33 2 6 9 0 0 12 0 23 0 37 1 1 4 0 0 13 0 24 4 34 0 5 5 0 0 8 6 25 1 29 3 2 6 3 0 4 3 Average: 1.76 17.76 2.52 4.56 8.2 1.08 0.04 10.64 2.88
44
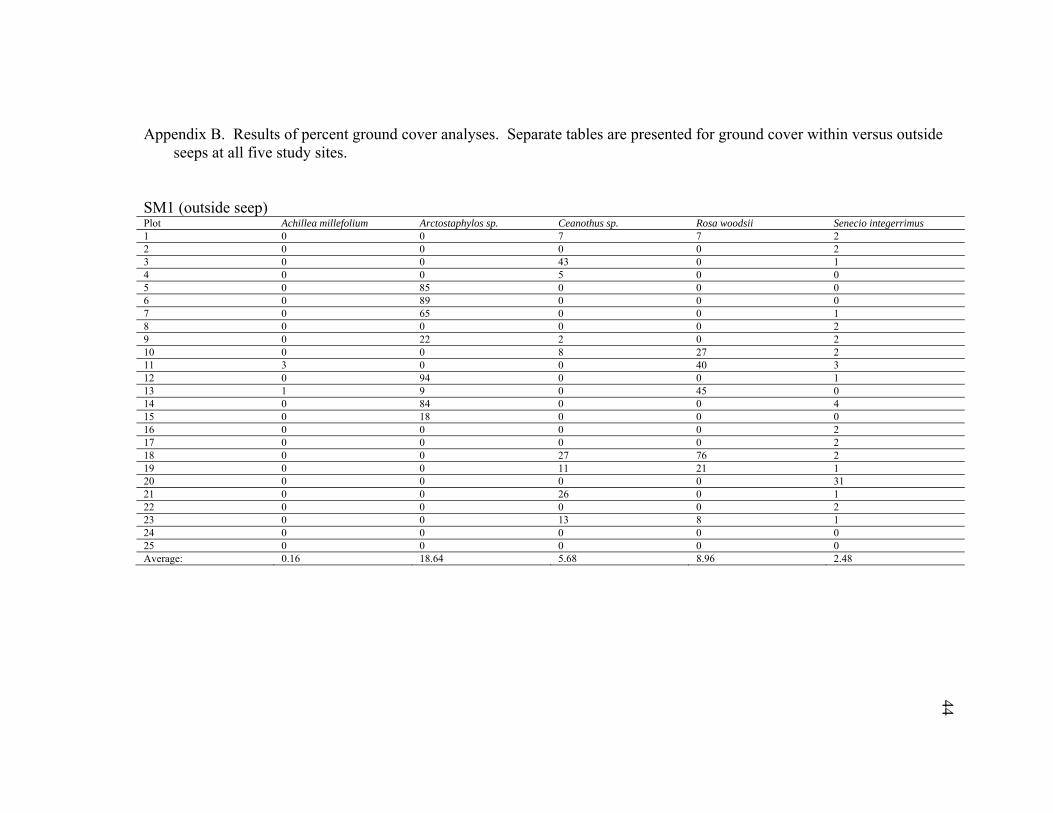
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside seeps at all five study sites.
SM1 (outside seep) Plot Achillea millefolium Arctostaphylos sp. Ceanothus sp. Rosa woodsii Senecio integerrimus 1 0 0 7 7 2 2 0 0 0 0 2 3 0 0 43 0 1 4 0 0 5 0 0 5 0 85 0 0 0 6 0 89 0 0 0 7 0 65 0 0 1 8 0 0 0 0 2 9 0 22 2 0 2 10 0 0 8 27 2 11 3 0 0 40 3 12 0 94 0 0 1 13 1 9 0 45 0 14 0 84 0 0 4 15 0 18 0 0 0 16 0 0 0 0 2 17 0 0 0 0 2 18 0 0 27 76 2 19 0 0 11 21 1 20 0 0 0 0 31 21 0 0 26 0 1 22 0 0 0 0 2 23 0 0 13 8 1 24 0 0 0 0 0 25 0 0 0 0 0 Average: 0.16 18.64 5.68 8.96 2.48
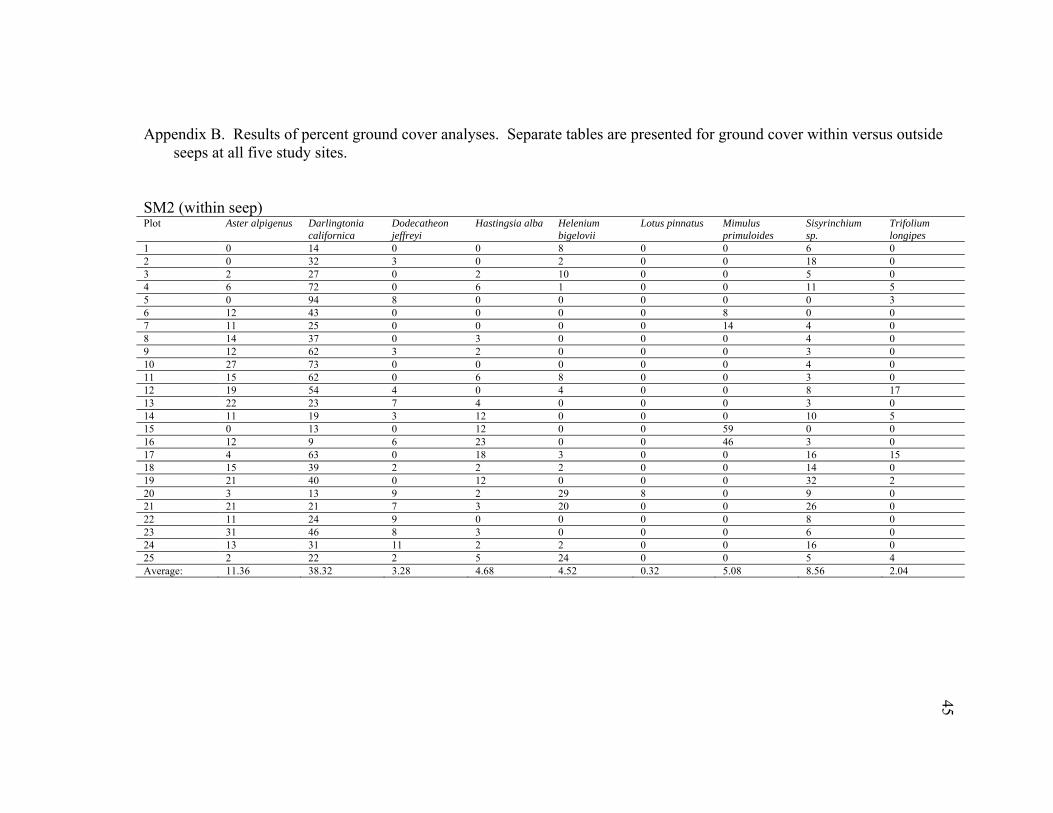
45
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside seeps at all five study sites.
SM2 (within seep) Plot Aster alpigenus Darlingtonia
californica Dodecatheon jeffreyi
Hastingsia alba Helenium bigelovii
Lotus pinnatus Mimulus primuloides
Sisyrinchium sp.
Trifolium longipes
1 0 14 0 0 8 0 0 6 0 2 0 32 3 0 2 0 0 18 0 3 2 27 0 2 10 0 0 5 0 4 6 72 0 6 1 0 0 11 5 5 0 94 8 0 0 0 0 0 3 6 12 43 0 0 0 0 8 0 0 7 11 25 0 0 0 0 14 4 0 8 14 37 0 3 0 0 0 4 0 9 12 62 3 2 0 0 0 3 0 10 27 73 0 0 0 0 0 4 0 11 15 62 0 6 8 0 0 3 0 12 19 54 4 0 4 0 0 8 17 13 22 23 7 4 0 0 0 3 0 14 11 19 3 12 0 0 0 10 5 15 0 13 0 12 0 0 59 0 0 16 12 9 6 23 0 0 46 3 0 17 4 63 0 18 3 0 0 16 15 18 15 39 2 2 2 0 0 14 0 19 21 40 0 12 0 0 0 32 2 20 3 13 9 2 29 8 0 9 0 21 21 21 7 3 20 0 0 26 0 22 11 24 9 0 0 0 0 8 0 23 31 46 8 3 0 0 0 6 0 24 13 31 11 2 2 0 0 16 0 25 2 22 2 5 24 0 0 5 4 Average: 11.36 38.32 3.28 4.68 4.52 0.32 5.08 8.56 2.04

46
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside
seeps at all five study sites. SM2 (outside seep) Plot Arctostaphlyos sp. Castilleja sp. Rosa woodsii Silene lemmonii 1 0 0 34 0 2 0 0 27 0 3 0 0 0 0 4 0 0 0 0 5 0 0 0 0 6 0 0 0 0 7 0 0 11 0 8 0 0 0 0 9 0 0 4 0 10 0 0 1 0 11 0 0 5 2 12 0 0 0 0 13 0 0 5 0 14 0 0 0 0 15 0 0 18 0 16 0 0 0 0 17 0 0 2 0 18 0 0 0 4 19 0 0 31 1 20 0 0 3 0 21 33 0 0 0 22 16 3 0 0 23 0 0 0 3 24 0 0 3 4 25 0 0 0 11 Average: 1.96 0.12 5.76 1
47
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside
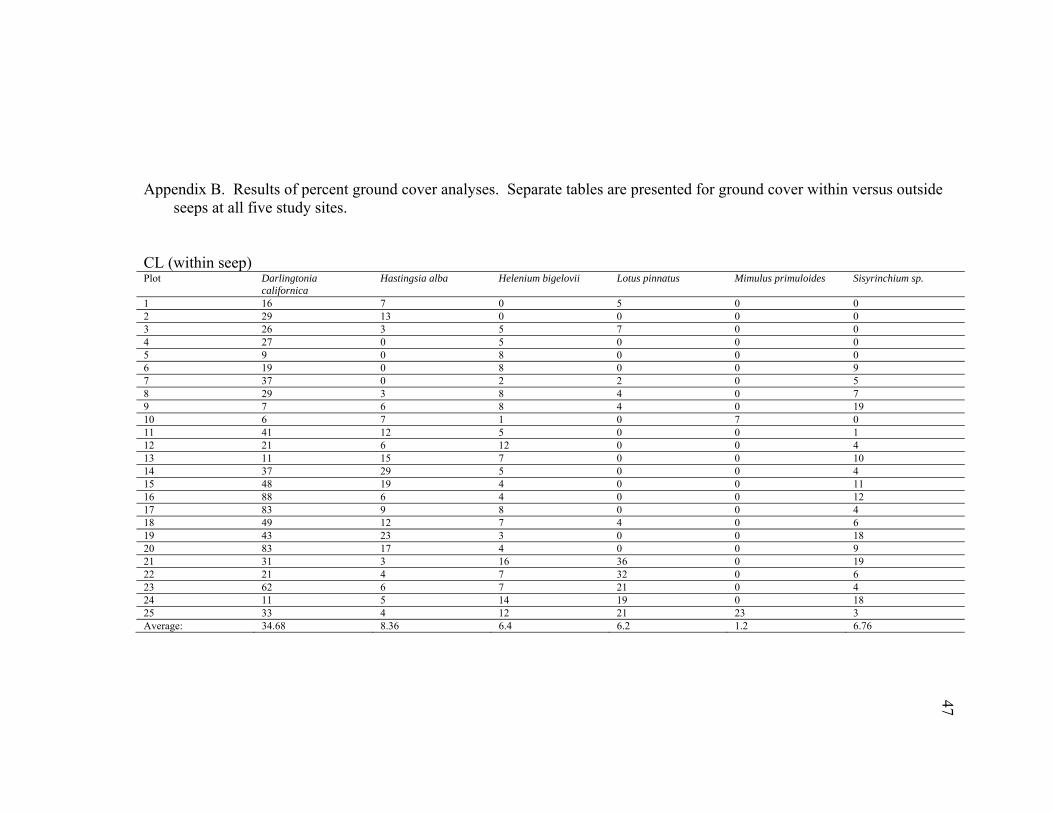
seeps at all five study sites. CL (within seep) Plot Darlingtonia
californica Hastingsia alba Helenium bigelovii Lotus pinnatus Mimulus primuloides Sisyrinchium sp.
1 16 7 0 5 0 0 2 29 13 0 0 0 0 3 26 3 5 7 0 0 4 27 0 5 0 0 0 5 9 0 8 0 0 0 6 19 0 8 0 0 9 7 37 0 2 2 0 5 8 29 3 8 4 0 7 9 7 6 8 4 0 19 10 6 7 1 0 7 0 11 41 12 5 0 0 1 12 21 6 12 0 0 4 13 11 15 7 0 0 10 14 37 29 5 0 0 4 15 48 19 4 0 0 11 16 88 6 4 0 0 12 17 83 9 8 0 0 4 18 49 12 7 4 0 6 19 43 23 3 0 0 18 20 83 17 4 0 0 9 21 31 3 16 36 0 19 22 21 4 7 32 0 6 23 62 6 7 21 0 4 24 11 5 14 19 0 18 25 33 4 12 21 23 3 Average: 34.68 8.36 6.4 6.2 1.2 6.76
48
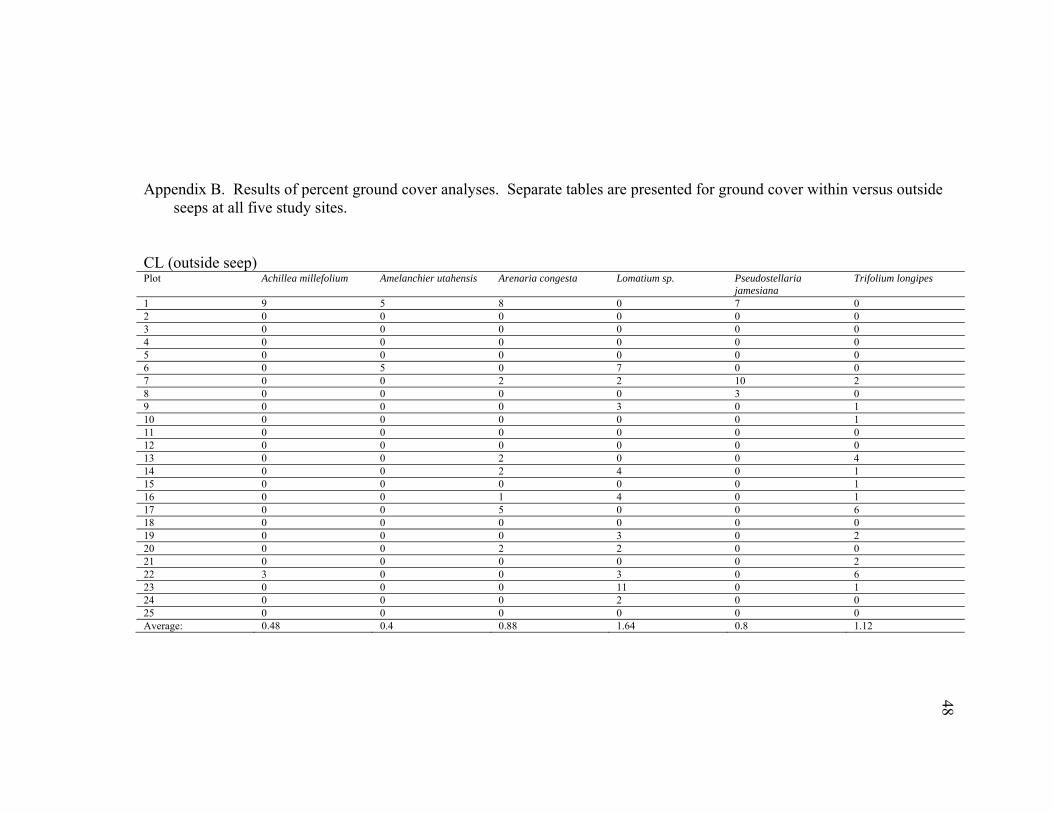
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside
seeps at all five study sites. CL (outside seep) Plot Achillea millefolium Amelanchier utahensis Arenaria congesta Lomatium sp. Pseudostellaria
jamesiana Trifolium longipes
1 9 5 8 0 7 0 2 0 0 0 0 0 0 3 0 0 0 0 0 0 4 0 0 0 0 0 0 5 0 0 0 0 0 0 6 0 5 0 7 0 0 7 0 0 2 2 10 2 8 0 0 0 0 3 0 9 0 0 0 3 0 1 10 0 0 0 0 0 1 11 0 0 0 0 0 0 12 0 0 0 0 0 0 13 0 0 2 0 0 4 14 0 0 2 4 0 1 15 0 0 0 0 0 1 16 0 0 1 4 0 1 17 0 0 5 0 0 6 18 0 0 0 0 0 0 19 0 0 0 3 0 2 20 0 0 2 2 0 0 21 0 0 0 0 0 2 22 3 0 0 3 0 6 23 0 0 0 11 0 1 24 0 0 0 2 0 0 25 0 0 0 0 0 0 Average: 0.48 0.4 0.88 1.64 0.8 1.12
49
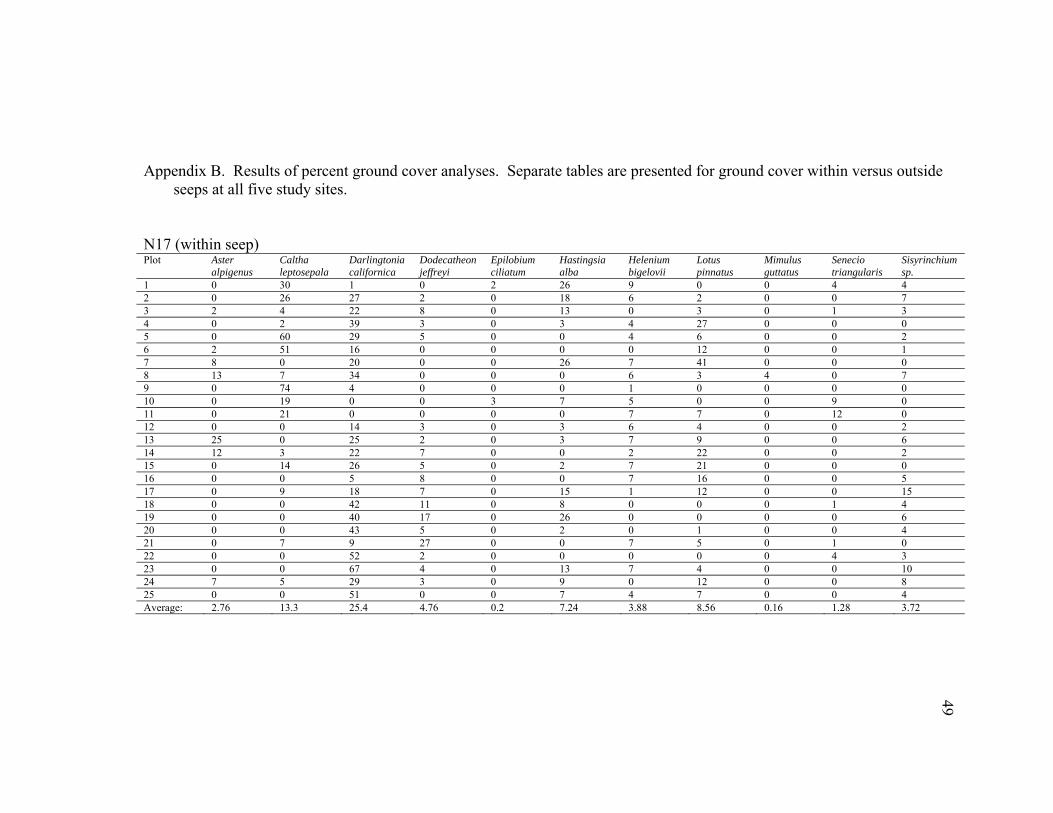
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside
seeps at all five study sites. N17 (within seep) Plot Aster
alpigenus Caltha leptosepala
Darlingtonia californica
Dodecatheon jeffreyi
Epilobium ciliatum
Hastingsia alba
Helenium bigelovii
Lotus pinnatus
Mimulus guttatus
Senecio triangularis
Sisyrinchium sp.
1 0 30 1 0 2 26 9 0 0 4 4 2 0 26 27 2 0 18 6 2 0 0 7 3 2 4 22 8 0 13 0 3 0 1 3 4 0 2 39 3 0 3 4 27 0 0 0 5 0 60 29 5 0 0 4 6 0 0 2 6 2 51 16 0 0 0 0 12 0 0 1 7 8 0 20 0 0 26 7 41 0 0 0 8 13 7 34 0 0 0 6 3 4 0 7 9 0 74 4 0 0 0 1 0 0 0 0 10 0 19 0 0 3 7 5 0 0 9 0 11 0 21 0 0 0 0 7 7 0 12 0 12 0 0 14 3 0 3 6 4 0 0 2 13 25 0 25 2 0 3 7 9 0 0 6 14 12 3 22 7 0 0 2 22 0 0 2 15 0 14 26 5 0 2 7 21 0 0 0 16 0 0 5 8 0 0 7 16 0 0 5 17 0 9 18 7 0 15 1 12 0 0 15 18 0 0 42 11 0 8 0 0 0 1 4 19 0 0 40 17 0 26 0 0 0 0 6 20 0 0 43 5 0 2 0 1 0 0 4 21 0 7 9 27 0 0 7 5 0 1 0 22 0 0 52 2 0 0 0 0 0 4 3 23 0 0 67 4 0 13 7 4 0 0 10 24 7 5 29 3 0 9 0 12 0 0 8 25 0 0 51 0 0 7 4 7 0 0 4 Average: 2.76 13.3 25.4 4.76 0.2 7.24 3.88 8.56 0.16 1.28 3.72

50
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside seeps at all five study sites.
N17 (outside seep) Plot Achillea
millefolium Arenaria congesta
Eriophyllum lanatum
Lomatium sp. Pseudostellaria jamesiana
Senecio integerrimus
Taraxacum officinale
Viola sororia
1 7 3 0 0 0 3 0 3 2 5 0 0 1 0 0 2 2 3 0 0 3 0 4 0 1 2 4 0 0 0 3 0 0 0 1 5 0 0 0 0 6 0 0 5 6 6 0 0 0 0 1 0 0 7 5 0 9 0 3 2 0 0 8 7 0 3 0 4 0 0 1 9 8 14 5 0 0 0 0 3 10 3 0 0 0 0 2 0 0 11 20 7 0 0 0 4 1 0 12 2 0 0 0 5 5 0 1 13 0 0 2 0 3 2 1 0 14 2 0 4 0 4 0 0 0 15 1 0 5 0 2 0 0 0 16 0 0 0 0 4 0 0 0 17 0 0 7 3 0 7 0 2 18 6 2 13 2 0 2 0 0 19 4 0 0 0 4 0 1 0 20 6 0 2 0 1 0 0 0 21 3 1 10 0 2 5 0 0 22 6 0 11 0 0 7 0 0 23 3 11 0 0 6 2 0 2 24 5 0 0 0 6 0 0 0 25 0 0 3 0 1 0 0 0 Average: 3.96 1.52 3.08 0.36 2.2 1.68 0.24 0.88
51
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside seeps at all five study sites.
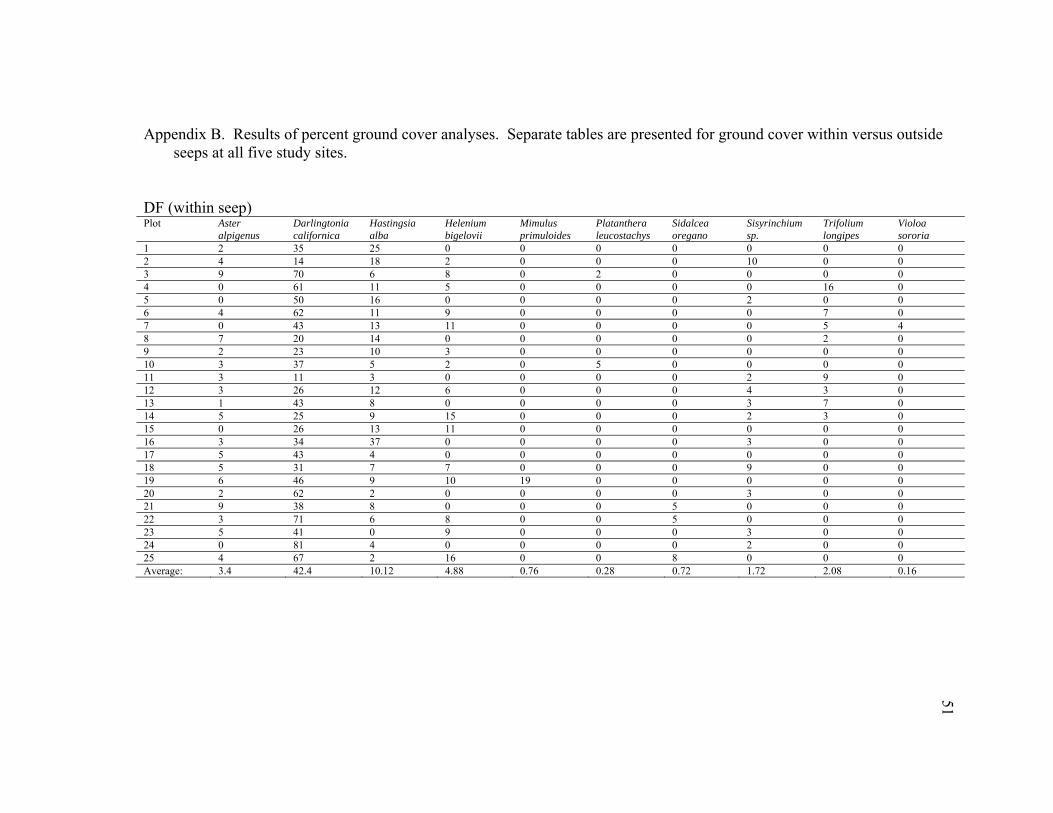
DF (within seep) Plot Aster
alpigenus Darlingtonia californica
Hastingsia alba
Helenium bigelovii
Mimulus primuloides
Platanthera leucostachys
Sidalcea oregano
Sisyrinchium sp.
Trifolium longipes
Violoa sororia
1 2 35 25 0 0 0 0 0 0 0 2 4 14 18 2 0 0 0 10 0 0 3 9 70 6 8 0 2 0 0 0 0 4 0 61 11 5 0 0 0 0 16 0 5 0 50 16 0 0 0 0 2 0 0 6 4 62 11 9 0 0 0 0 7 0 7 0 43 13 11 0 0 0 0 5 4 8 7 20 14 0 0 0 0 0 2 0 9 2 23 10 3 0 0 0 0 0 0 10 3 37 5 2 0 5 0 0 0 0 11 3 11 3 0 0 0 0 2 9 0 12 3 26 12 6 0 0 0 4 3 0 13 1 43 8 0 0 0 0 3 7 0 14 5 25 9 15 0 0 0 2 3 0 15 0 26 13 11 0 0 0 0 0 0 16 3 34 37 0 0 0 0 3 0 0 17 5 43 4 0 0 0 0 0 0 0 18 5 31 7 7 0 0 0 9 0 0 19 6 46 9 10 19 0 0 0 0 0 20 2 62 2 0 0 0 0 3 0 0 21 9 38 8 0 0 0 5 0 0 0 22 3 71 6 8 0 0 5 0 0 0 23 5 41 0 9 0 0 0 3 0 0 24 0 81 4 0 0 0 0 2 0 0 25 4 67 2 16 0 0 8 0 0 0 Average: 3.4 42.4 10.12 4.88 0.76 0.28 0.72 1.72 2.08 0.16
52
Appendix B. Results of percent ground cover analyses. Separate tables are presented for ground cover within versus outside seeps at all five study sites.
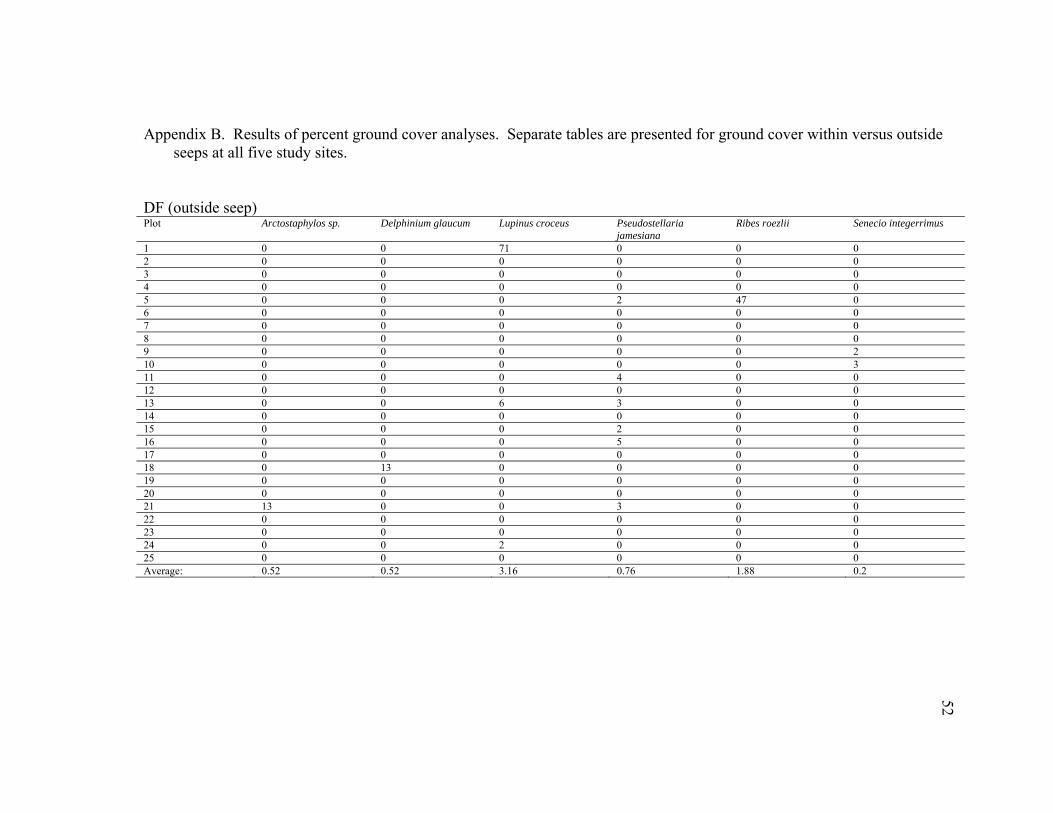
DF (outside seep) Plot Arctostaphylos sp. Delphinium glaucum Lupinus croceus Pseudostellaria
jamesiana Ribes roezlii Senecio integerrimus
1 0 0 71 0 0 0 2 0 0 0 0 0 0 3 0 0 0 0 0 0 4 0 0 0 0 0 0 5 0 0 0 2 47 0 6 0 0 0 0 0 0 7 0 0 0 0 0 0 8 0 0 0 0 0 0 9 0 0 0 0 0 2 10 0 0 0 0 0 3 11 0 0 0 4 0 0 12 0 0 0 0 0 0 13 0 0 6 3 0 0 14 0 0 0 0 0 0 15 0 0 0 2 0 0 16 0 0 0 5 0 0 17 0 0 0 0 0 0 18 0 13 0 0 0 0 19 0 0 0 0 0 0 20 0 0 0 0 0 0 21 13 0 0 3 0 0 22 0 0 0 0 0 0 23 0 0 0 0 0 0 24 0 0 2 0 0 0 25 0 0 0 0 0 0 Average: 0.52 0.52 3.16 0.76 1.88 0.2

53





























