Embed Size (px)
Citation preview

EXPERIMENTAL NEUROLOGY 69, 61-73 (1980)
Pontine-Geniculate-Occipital Spikes in the Albino Rat: Evidence for the Presence of the Pontine Component
as Revealed by Cerebellar Lesions
PETER B. REINER AND ADRIAN R. MORRISON’
Laboratories of Anatomy. School of Veterinary Medicine, University of Pennsylvania, Philadelphia, Pennsylvania 19104
Received July 25, 1979; revision received December 7, 1979
The presence of pontine-geniculate-occipital (PGO) spikes in the albino rat has been questioned in recent years. Because cats with cerebellar cortical lesions exhibit either flexor or extensor limb jerks in association with PGO spikes at the transition to paradoxical sleep (PS), cerebellar cortical lesions were made in albino rats to see if similar behavioral changes, i.e., limb jerks at the transition to PS, would ensue. Limb jerks were seen in the rat at the transition to PS although at a lower frequency than that seen in the cat. Limb jerks could also be elicited by novel, external stimuli in much the same manner as in the cat. Spontaneous and elicited jerks are presumably due to activation by the pontine reticular formation of neurons in the deep cerebellar nuclei, which are no longer modulated by the inhibitory Purkinje cells that have been ablated. We conclude that pontine neural activity similar to that underlying the PGO spike in the cat exists in the albino rat.
INTRODUCTION
Paradoxical sleep (PS) is characterized by low-voltage, high-frequency waves in the electroencephalogram (EEG), tonic muscle atonia, and rapid eye movements. PS normally follows an initial phase of synchronized sleep during which the EEG exhibits high-amplitude, low-frequency waves; those two states then continue to occur alternately during any given sleep period (19). A great deal of effort has been devoted toward understanding and explaining the various electrophysiological events which occur during these sleep states. Much attention has focused on a rather intriguing
Abbreviations: PGO-pontine-geniculate-occipital, PS-paradoxical sleep. r Supported by National Institutes of Health grants NS-13 110 and GM-07170. We thank Ms.
Graziella Mann for her expert technical assistance.
61
0014-4886/80/070061-13$02.00/0 Copyright 0 1980 by Academic Press, Inc. All rights of reproduction in any form reserved.

62 REINER AND MORRISON
phenomenon of PS, the pontine-geniculate-occipital (PGO) spike. This large-amplitude waveform has been studied extensively in the cat (4) and to some extent in the monkey (1). PGO spikes derive their name from the fact that they may be recorded with macroelectrodes from the pontine reticular formation, the lateral geniculate nucleus, and the occipital cortex. They occur just prior to and throughout each episode of PS; they are seen singly as well as in clusters in association with the rapid eye movements of PS.
In spite of the fact that the presence of PGO spikes is well established in two common laboratory mammals, their existence in the albino rat has been subject to some debate. Thus pontine spikes were reported during PS in the “parasagittal pons” (15), although treatment with reset-pine produced no release of spike activity into other states as was seen in the cat (20). The neural activity underlying the PGO spike is thought to arise in the pons and from there spread to the lateral geniculate body and occipital cortex (5). However, workers have reported the absence of spike activity in these structures in the chronically prepared albino rat (32) although geniculate spikes were recorded in the acute, rat treated with serotonin antagonists (7,8). Because of these discrepancies between the cat and rat, we decided to use an indirect approach to test for the presence of neural activity underlying the PGO spike in the albino rat.
Morrison and Bowker (26, 27) demonstrated in the cat that vermal and paravermal lesions of the cerebellar cortex resulted in extensor and flexor limb jerks, respectively, at the transition to PS and that these jerks corresponded 1:l with PGO spikes. Both could easily be elicited during synchronized sleep (3). This led to the hypothesis that PGO spikes are associated with a neural alerting response to various stimuli, either external, as in the case of tone-induced PGO spikes, or internal, as those occurring spontaneously during PS, when brain stem neurons are disinhibited from serotonergic control (31). Those authors reasoned that the jerks are the result of excitation by pontine tegmental cells of fastigial and interpositus neurons, which themselves are hyperactive after their release from the inhibitory control of Purkinje axons by cortical lesions (35) or cooling (30, 35).
We believed that evidence of homologous brain stem activity leading to both PGO spikes and jerks could be revealed by cerebellar lesions in rats. It was reported (14) that the rodent cerebellum, like that of the feline (lo), is functionally organized into longitudinal vermal and paravermal zones. Thus, rats with cerebellar cortical lesions should undergo behavioral changes similar to those seen in the cat, and the presence of jerks at times when PGO-associated jerks appear in cats should be reasonably interpreted as evidence of the existence of the basic pontine activity underlying PGO activity in the albino rat. The present study reports the results of such

PGO-RELATED PHENOMENA IN THE ALBINO RAT 63
vermal and paravermal cerebellar cortical lesions and their association with sleep phenomena in the albino rat.
METHODS
Thirteen adult, male, albino rats, weighing 350 to 550 g were used. All surgical procedures were carried out under an anesthetic mixture of chloral hydrate, magnesium sulfate, and pentobarbital or halothane anesthesia. The animals were placed in a stereotaxic device (David Kopf Instruments), the occipital bone was removed from the nuchal crest caudally, the dura reflected, and indicated portions of the vermal (N = 8) or paravermal (N = 5) cerebellar cortex ablated by aspiration. The vermal lesions were bilateral and the paravermal ones were ipsilateral only. Initially (N = 5), in order to ascertain the existence of the jerks and the appropriate size of the lesion, the animals were without electrodes and were monitored only behaviorally. Later, the animals (N = 8) were chronically implanted, at least 10 days prior to lesions, with stainless-steel skull screws for monitoring the EEG. Stainless-steel, Teflon-insulated wires were sutured into the nuchal musculature for recording of electromyograms. The wires were then attached to a miniature plug and embedded in dental acrylic upon the animal’s skull, allowing for convenient connection to the polygraph (Beckman, type R). Control recordings of sleep episodes were made prior to the lesion to ensure that the sleep patterns were essentially normal (24). The animals were housed and studied in a 30 x 30 x 35 cm Plexiglas cage.
Neurological examinations, including the vestibular drop and tactile placing tests, were routinely made both pre- and postoperatively. The vestibular drop test involves holding an animal and suddenly dropping it 15 to 30 cm, thereby releasing labyrinthine reflexes which normally produce extension of the limbs with some toe fanning (11). The tactile placing test is done by holding the animal and slowly bringing the dorsal (or lateral) aspect of the limb into contact with the edge of a table or other suitable surface. The normal response is a coordinated stepping motion onto the table. These two responses may be altered by cerebellar lesions (10).
Three weeks to two months after the lesion, the animals were killed by an overdose of anesthetic agent and perfused intracardially with warm (37°C) saline followed by formalin. The brains were removed and embedded in celloidin, sliced into 30-pm sections, and stained alternately with a cresyl violet cell stain and Weil’s stain for myelin sheaths.
RESULTS
Animals with lesions exhibited behavioral changes similar to those seen in cats (26), detectable by neurological examinations, polygraphic

64 REINER AND MORRISON
recordings, and behavioral observations. The most salient characteristic of the sleep behavior of the animals with lesions was the occurrence of limb jerks during synchronized sleep and, more markedly, at the transition to PS. The jerks were readily divisible into two groups: vermal lesions resulted in extensor jerks whereas paravermal lesions resulted in flexor jerks. A further difference was noted between animals receiving superficial vermal lesions and those in which the ablation was more extensive. Superficial lesions (N = 4) produced virtually no behavioral deficits during waking and no jerks during sleep or other obvious changes in the sleep pattern. Animals with deeper vermal lesions (N = 4), often extending down to the deep cerebellar nuclei (Fig. IA), exhibited extensor jerks of axial musculature and forelimbs, with dorsiflexion of the spine during sleep and wakefulness. The behavioral manifestation of the paravermal lesions depended on the placement of the lesion. When the lesion involved lobule V (Fig. lB), which is primarily involved with mediation of forelimb coordination, flexion jerks of the shoulder and elbow joints occurred during sleep and in association with auditory stimuli (N = 3); when the lesion was lateral (N = 1) or small (N = 1), such behavioral changes were not seen. Lesions capable of producing jerks during sleep tended to be much deeper in animals with vermal lesions than in those with paravermal ablations.
During synchronized sleep, spontaneous jerks were seen at a rate of one to fourjerks per minute. Often, but not invariably, these jerks were elicited during quiet wakefulness or synchronized sleep by auditory stimuli (snapping of fingers; light pencil tap on cage), which were mild enough not to arouse the animal behaviorally. In some animals, the jerks themselves were of such violent nature that the animal was aroused to a varying extent, as noted both polygraphically and behaviorally. The arousing effects were most prominent in those animals with bilateral vermal lesions in which extensor jerks of axial musculature served to raise the entire animal off the floor of the cage; with unilateral paravermal lesions, only the ipsilateral forelimb responded with a flexor response, and this was not a strong arousing stimulus. When the animals were in synchronized sleep, the arousing effect of the extensorjerk was often a transient desynchronization of the EEG with a resumption of slow-wave activity within 60 s and often as soon as 10 s.
Spontaneous jerks occurred at the transition to PS at a rate of four to six per minute, often resulting in aborted PS episodes (Fig. 2). During these episodes, muscle tone diminished (at least 33%) in much the same manner as during a normal transition to PS; however, one or more jerks would interrupt the smooth transition, and one would observe an increased degree of muscle tone as well as a synchronized EEG. Another interrupted
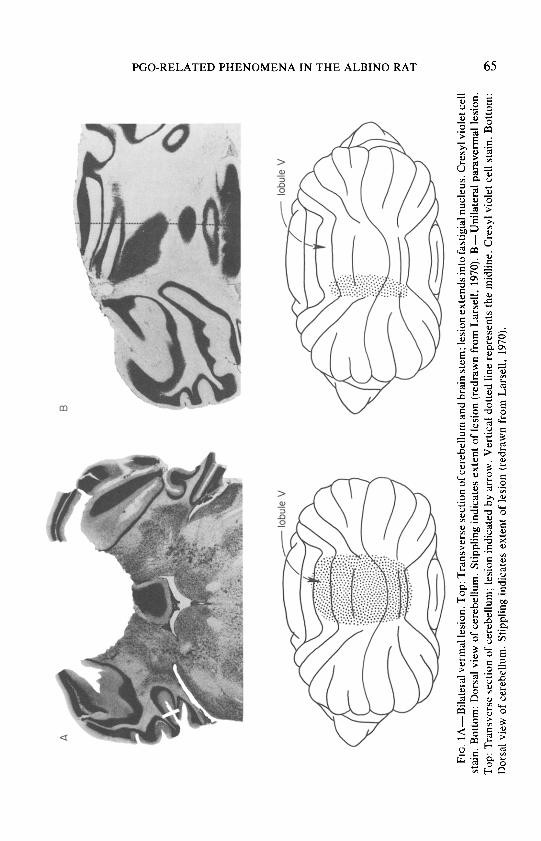
FIG.
1 A
-Bila
tera
l ve
rmal
lesio
n.
Top:
Tr
ansv
erse
se
ctio
n of
ce
rebe
llum
an
d br
ain
stem
; le
sion
exte
nds
into
fa
stig
ial
nucle
us.
Cres
yl vio
let
cell
stai
n.
Botto
m:
Dors
al vie
w of
ce
rebe
llum
. St
ippl
ing
indi
cate
s ex
tent
of
le
sion
(redr
awn
from
La
rsel
l, 19
70).
B-Un
ilate
ral
para
verm
al le
sion.
To
p:
Tran
sver
se
sect
ion
of
cere
bellu
m;
lesio
n in
dica
ted
by
arro
w.
Verti
cal
dotte
d lin
e re
pres
ents
the
mid
line.
Cr
esyl
viole
t ce
ll st
ain.
Bo
ttom
: Do
rsal
view
of
cere
bellu
m.
Stip
plin
g in
dica
tes
exte
nt
of
lesio
n (re
draw
n fro
m
Lars
ell,
1970
).

EMG r-
----
-.*
. 7
-~-p
r
- --
.

PGO-RELATED PHENOMENA IN THE ALBINO RAT 67
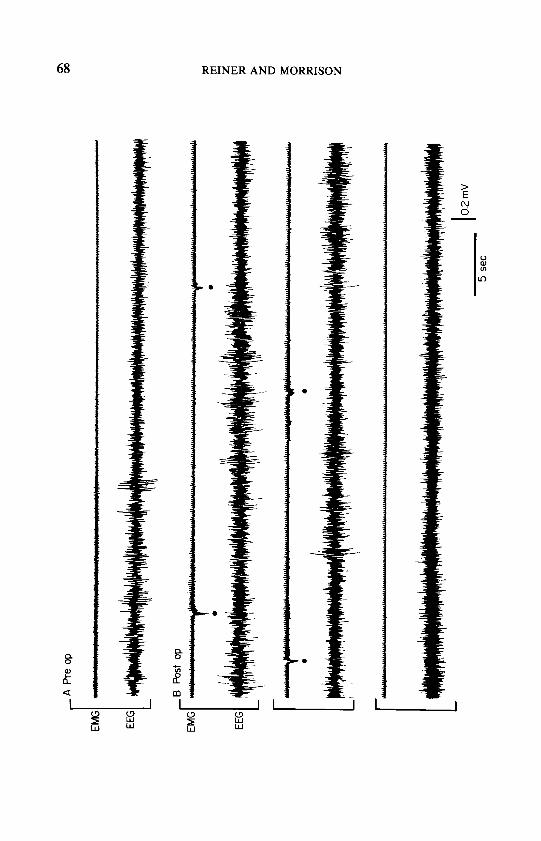
transition would follow, usually within 2 min, and the pattern would repeat (Fig. 2) for as long as 10 min. Termination of these aborted episodes occurred when the animal was awakened briefly by the jerks, at which point the rat would almost always shift its position and return to synchronized sleep. Some animals underwent periods as long as 12 h during which only synchronized sleep and aborted PS episodes occurred. Eventually, the animals would enter PS, either with a mild jerk at the transition (Fig. 3) or, after some degree of recovery from the effects of the lesion had occurred, without a jerk; the ensuing PS episodes appeared normal.
Recovery from the effect of the lesions was generally rapid. The strongest jerks were seen during the first 36 h postoperatively, and they disappeared completely by 96 h. Results of neurological examination generally paralleled the observations of sleep behavior. Animals exhibiting extensor jerks after vermal lesions showed hyperextension and fanning of digits, especially of the forelimbs, during the vestibular drop test. Animals in which paravermal lesions resulted in flexor jerks showed hypermetria of the ipsilateral forelimb during locomotion. No change was noted in tactile placing test performance in animals with lesions. The neurological signs diminished as the jerks subsided with time but remained to varying extents even after the animal was no longer displaying jerks during sleep.
DISCUSSION
These results further substantiate the hypothesis that the basic neuronal activity underlying the PGO spike in the cat also exists in the albino rat. The subsequent discussion will include a brief review of the relevant data regarding the PGO phenomenon in the cat, an overview of those aspects of cerebellar anatomy and physiology thought to be responsible for PGO-related jerks, and an examination of the present status of PGO spikes in the albino rat.
The PGO spike has been extensively studied in the cat. The neural activity underlying PGO spikes is thought to arise in the pontine reticular formation (5). PGO activity is strongly modulated by serotonergic fibers of the midline raphe (3 l), a finding which is supported by studies which alter central nervous system serotonin (20) and recordings of raphe neurons during PS (9,25,34). Recent work indicates that PGO spikes are associated with a form of alerting in that external stimuli to which an animal orients
FIG. 2. Aborted paradoxical sleep episodes; 6-min record with four aborted episodes. The electromyogram (EMG) diminishes as during normal transition to paradoxical sleep (see Fig. 3), but the extensor jerks result in aborted episodes. This record is taken from the middle of an II-min bout of aborted paradoxical sleep episodes. The animal awakened at the end of the bout.
A Pr
e op
EMG
EEG
EMG
EEG
rti r*
/ 14
, I I 0.
2 m
V
5 se
t

PGO-RELATED PHENOMENA IN THE ALBINO RAT 69
will induce a PGO spike during all states, and this response will habituate (3, 27). This work led to the hypothesis that the same set of brainstem neurons is activated during spontaneous PGO spikes in PS, eye movement potentials during wakefulness, and PGO spikes induced by novel stimuli. It is a similar brain stem neural assembly which we will argue exists in the albino rat, based upon the results of behavioral changes after cerebellar lesions.
We must then consider the question as to why cerebellar cortical lesions should allow a unique behavioral expression of central activity during the transition to PS. Transcortical recordings from the cerebellar vermis reveal the existence of phasic slow waves coincident with PGO spikes (28). Thus some form of cerebellar activation occurs during each PGO spike. The paravermal cortical Purkinje cells project to and inhibit the nucleus interpositus (17,18) which provides excitatory input to the red nucleus (33). The vermal Purkinje cells are also inhibitory and ultimately exert their effects upon the lateral vestibular nucleus, either directly or indirectly, via the fastigial nucleus (16, 17) (Fig. 4). Afferent fibers to the cerebellar cortex give off collaterals to the deep cerebellar nuclei which provide the excitatory background which is then modulated by the inhibitory Purkinje cells (12). Paravermal cortical ablation or cooling allows unregulated activity in the interpositus nucleus; behaviorally this results in exaggerated flexor responses via the interpositus-rubrospinal pathway (35,36). Because the vermal zone has a similar corticonuclear relationship (18), but is functionally related to axial muscular systems (lo), we think it reasonable to assume that removal of the vermal cortex results in exaggerated extensor responses via the midline cerebellar-vestibular-spinal path.
When the neuronal activity which arises in the pontine reticular formation during PGO spikes reaches the cortically ablated cerebellum, collaterals still excite the deep cerebellar nuclei, but there are many fewer Purkinje cells which project back to the relevant deep nuclear zone. Thus nuclear activity goes unchecked and jerks occur, either extensor or flexor, whenever PGO spikes occur in an animal with some degree of muscle tone. This situation arises during the occasional PGO spikes of synchronized sleep, and, more markedly, at the transition to PS. During wakefulness, auditory stimuli induce limb jerks similar to those seen in sleep in association with PGO spikes in cats with cerebellar cortical lesions (26). We showed that jerks can also be induced by auditory stimuli during wakefulness in rats with similar lesions. The mechanism is thought to be due to the fact that the same set of brain stem neurons is activated during both
FIG. 3. Transition to paradoxical sleep. A-Preop: normal transition to paradoxical sleep prior to lesion. B-Postop: transition to paradoxical sleep after lesion with jerks [seen as
downward deflection on the electromyogram (EMG)] during the transition.

70 REINER AND MORRISON
-
T NUChS
interpositus -
Lateral vestibular nucleus of Deiters
Pontine
+
El
reticular + formation
T Extensor FlWX
motOneurOnS motoneurons
FIG. 4. Schematic representation of neuronal circuitry involved in pontine-geniculate- occipital (PGO)-associated limb jerks. Removal of the cerebellar cortical Purkinje cells in either the vermal or paravermal zones results in abolition of their inhibitory influence upon the deep cerebellar (and Deiters’) nuclei. Subsequent excitation by pontine reticular neurons (as during PGO spikes) has its effect only upon the unregulated deep cerebellar nuclei, thereby resulting in the limb jerks seen at the transition to paradoxical sleep.
PGO spikes and the neuronal response to novel stimuli; indeed, auditory stimuli induce PGO spikes without jerks in normal cats (3). Thus lesions of the vermal and paravermal cerebellar cortices result in unregulated excitation of the fastigial and interposed nuclei, respectively, ultimately resulting in hyperreflexia in wakefulness and jerks during sleep.
The crux of the matter is, however, whether or not PGO spikes exist in the albino rat. PGO spikes are thought of less and less as being restricted to the domain of sleep; indeed recent evidence points to their similarity to eye movement potentials during wakefulness (2, 6). Their association with behavioral alerting (3), an activity whose function is so crucial to the survival of the animal, argues strongly for the hypothesis that the brain stem neural activity underlying this phenomenon exists in all mammals exhibiting behavioral alerting and PS. As such, it would be quite remarkable for this activity to arise, de nova, in such animals as the cat without any related phenomena existing in other animals with complex behavioral repertoires. Our work as well as that of others (13,21,23) clearly shows that the pontine generator of PGO spikes exists in the albino rat. Activation of this pontine region causes unregulated excitation of the cerebellar nuclear cells and results in ajerk during sleep in the same manner as described for the cat.
Although similarities in sleep behavior of cats and rats with cerebellar cortical lesions argue for the same basic phenomena existing in both

PGO-RELATED PHENOMENA IN THE ALBINO RAT 71
species, some differences are seen. Limb jerks occur at a rate of four per minute in the rat with a cerebellar cortical lesion, whereas in the cat they are seen at a rate of 30 per minute (26). We were baffled by this until Farber et al. (13), independent of and concurrent with the preliminary discussion of the data presented herein (29), reported that pontine spikes, recordable in the rat from “the area of the locus coeruleus,” occurred during the transition to PS at a rate comparable to the rate of limb jerks we found in the rat with a cerebellar cortical lesion. These results have since been confirmed and amplified (21, 23). This difference in the frequency of both spikes and jerks between the rat and the cat may indicate that the neuronal network which generates spontaneous PGO spikes is not identical in every detail in these two species. However, the fact that in both species, the frequency of limb jerks, a behavioral event, corresponds so well to that of the pontine spike, an electrophysiological event, implicates homologous brain stem neural activity underlying both events in both species.
The specific topographic site of the lesion was closely correlated with the behavioral manifestation of the jerks. In both vermal and paravermal lesions, ablation of lobule V appeared to be crucial to the presence of the jerks. Lobule V primarily mediates forelimb coordination (22). The depth of the lesion was also an important factor, especially in the vermal lesions. Although the jerks were produced with extremely superficial damage to the paravermal cortex, only in animals in which the vermal lesions encroached upon the fastigial nucleus did the jerks appear. In the cat, such extensive vermal lesions are not required to produce the jerks (26). An explanation for this difference might be the fact that the rat, with its body virtually hugging the ground, is less debilitated by lesions of cortical regions regulating axial muscular activity than the relatively long-legged feline, and it is not until virtually all the Purkinje cells are ablated that the behavior manifests itself.
Thus the present status of PGO spikes in the albino rat is as follows: pontine spikes have been recorded (13, 21, 23); the existence of lateral geniculate and occipital cortical spikes has not yet been demonstrated (7,8, 15, 32); and we have demonstrated that cerebellar lesions confirm the existence of the pontine component of PGO spikes through behavioral changes. Recent work indicates that the PGO phenomenon is a more generalized one than workers have previously thought it to be. Although emphasis was originally placed on their occurrence in paradoxical sleep, they have been shown to occur spontaneously during slow-wave sleep in the cat (19), and quite recently in the rat pons (13, 21, 23); evidence is accumulating which suggests that they result from activation of a set of brain stem neurons associated with alerting, functional throughout all states of consciousness (2). As such, one might expect to find them in some

72 REINER AND MORRISON
form in all mammals exhibiting PS. We suggest that the understanding of this phenomenon will become clearer as we begin to view it within the framework of the animal’s behavior as a whole, rather than as an isolated event occurring only during sleep.
REFERENCES 1. BALZAMO, E., AND J. BERT. 1975. Sleep in papio annubis: its organization and lateral
geniculate spikes. Page 138 in M. CHASE, W. C. STERN, AND P. L. WALTER, Eds., Sleep Research, Vol. 4. Brain Information Service/Brain Research Institute, Univ. of California, Los Angeles.
2. BOWKER, R. M. 1979. The Biological Significance of the Ponto-Geniculo-Occipital Wave and the Eye Movement Potential in the Cat, Ph.D. Thesis. University of Pennsylvania, Philadelphia.
3. BOWKER, R. M., AND A. R. MORRISON. 1976. The startle reflex and PGO spikes. Brain Res. 102: 185-190.
4. BROOKS, D. C. 1973. Commentary on: “Brainstem electrical activity during sleep.” Pages 72-83 in W. B. WEBB, Ed., Sleep: An Active Process, Scott, Foresman, Glenview, Illinois.
5. BROOKS,D. C.,ANDE. BIZZI. 1963. Brainstemelectricalactivityduringdeepsleep.Arch. Ital. Biol. 101: 648-665.
6. BROOKS, D. C., AND M. D. GERSHON. 1971. Eye movement potentials in the oculomotor and visual systems of the cat: a comparison of reserpine induced waves and those present during wakefulness and REM sleep. Brain Res. 27: 223-239.
7. CESPUGLIO, R., J. M. CALVO, AND H. DEPOORTERE. 1975. Phasic electrical activity in the rat. Page 53 in M. H. CHASE, W. C. STERN, AND P. L. WALTER, Eds., Sleep Research, Vol. 4. Brain Information Service/Brain Research Institute, Univ. of Calfomia, Los Angeles.
8. CESPUGLIO, R., J. M. CALVO, R. MUSOLINO, AND J. L. VALATX. 1977. Activite phasique chez le rat. Physiol. Behav. 19: 589-596.
9. CESPUGLIO, R., M. E. GOMEZ, E. WALKER, AND M. JOUVET. 1977. Singleunitrecordings of the nuclei raphe dorsalis and magnus during sleep-waking cycle. Page 26 in M. H. CHASE, M. M. MITLER, AND P. L. WALTER, Eds., Sleep Research, Vol. 7. Brain Information Service/Brain Research Institute, Univ. of California, Los Angeles.
10. CHAMBERS, W. W., AND J. M. SPRAGUE. 1955. Functional localization in the cerebellum. I. Organization in longitudinal cortico-nuclear zones and their contribution to the control of posture, both extrapyramidal and pyramidal. J. Comp. Neurol. 103: 105- 129.
11. DEKLEIJN, A. 1924. Experimental physiology of the labyrinth. Proc. R. Sot. Med. Sect. Otol. 17: 6-23.
12. ECCLES, J. C., M. ITO, AND J. SZENTAGOTHAI. 1967. The Cerebellum as a Neuronal Machine, p. 274. Springer-Verlag, New York.
13. FARBER, J., G. MARKS, C. BERWISE, AND H. ROFFWARG. 1976. Pontine sharp waves during REM sleep in the albino rat. Page 21 in M. H. CHASE, M. M. MITLER, AND P. L. WALTER, Eds., Sleep Research, Vol. 5. Brain Information Service/Brain Research Institute, Univ. of California, Los Angeles.
14. GOODMAN, D. C., AND J. T. SIMPSON. 1961. Functional localization in the cerebellum of the albino rat. Exp. Neuroi. 3: 174- 188.
15. GOTTESMANN, C. 1969. Etude sur les activites tlectrophysiologiques phasiques chez le rat. Physiol. Behav. 4: 495-504.

PGO-RELATED PHENOMENA IN THE ALBINO RAT 73
16. Iro, M., AND M. YOSHIDA. 1966. The origin of cerebellar induced inhibition of Deiters neurons. I. Monosynaptic initiation of the inhibitory postsynaptic potentials. Exp. Bruin Res. 2: 330-349.
17. ITO, M., M. YOSHIDA, K. OBATA, N. KAWAI, AND M. UDO. 1970. Inhibitory control of intracerebellar nuclei by the Purkinje cell axons. Exp. Bruin Res. 10: 64-80.
18. JANSEN, J., AND A. BRODAL. 1940. Experimental studies on the intrinsic fibers of the cerebellum II. The cortico-nuclear projection. J. Comp. Neurol. 73: 267-321.
19. JOUVET, M. 1967. Neurophysiology of the states of sleep. Physiol. Rev. 47: 117-177. 20. JOUVET, M. 1969. Biogenic amines and the states of sleep. Science 163: 32-41. 21. KAUFMAN, L. S., AND A. R. MORRISON. 1978. PGO spikes in rats: a component of the
alerting response. Sot. Neurosci. Abstr. 4: 541. 22. LARSELL, 0. 1970. Pages 31-58 in J. JANSEN, Ed., The Comparative Anatomy and
Histology of the Cerebelium From Monotremes through Apes. Univ. of Minnesota Press, Minneapolis.
23. MARKS. G. 1978. Central Phasic Activity Associated w*ith REM Sleep in the Albino Rat: the Homologue of the PC0 Spike, Ph.D. Thesis, City University of New York.
24. MCGINTY, D. J. 1966. Neural Control of Sleep Patterns in the Rat, Ph.D. Thesis. University of Pennsylvania, Philadelphia.
25. MCGINTY, D. J., AND R. M. HARPER. 1976. Dorsal raphe neurons: depression of firing during sleep in cats. Brain Res. 101: 569-575.
26. MORRISON, A. R., AND R. M. BOWKER. 1973. Cerebellar and spinal contributions to the regulation of muscle tone and movement during sleep. Pages 270-277 in U. J. JOVANOVIC, Ed., The Nature of Sleep. Fischer, Stuttgart.
27. MORRISON, A. R., AND R. M. BOWKER. 1975. The biological significance of PGO spikes. Acta Neurobiol. Exp. 35: 821-840.
28. PELLET, J., M. -F. TARDY, S. DUBROCARD, AND F. HARLAY. 1974. Etude de l’activite Clectrique phasique du cortex du cervelet au tours des etats de veille et de sommeil. Arch. Ital. Biol. 112: 163-195.
29. REINER, P. B., AND A. R. MORRISON. 1976. Phasic phenomena during sleep in the albino rat. Page 32 in M. H. CHASE, M. M. MITLER, AND P. L. WALTER, Eds., Sleep Research, Vol. 5. Brain Information Service/Brain Research Institute, Univ. of California, Los Angeles.
30. ROSEN, I., AND P. SCHEID. 1972. Cerebellar surface cooling influencing evoked activity in cortex and interpositus nucleus. Brain Res. 45: 580-584.
31. SIMON, R. P., M. D. GERSHON, AND D. C. BROOKS. 1973. The role ofthe raphe nuclei in the regulation of PGO wave activity. Brain Res. 58: 313-330.
32. STERN, W. C., W. B. FORBES, AND P. J. MORGANE. 1974. Absence of pontogeniculo- occipital (PGO) spikes in rats. Physiol. Behav. 12: 293-295.
33. TOYAMA, K., N. TSUKAHARA. AND M. UDO. 1968. Nature of cerebellar influences upon the red nucleus neurons. Exp. Brain Res. 4: 292-309.
34. TRULSON, M. E., AND B. L. JACOBS. 1979. Raphe unit activity in freely moving cats: correlation with level of behavioral arousal. Bruin Res. 163: 135-150.
35. Yu, J. 1972. The pathway mediating ipsilateral limb hyperflexion after cerebellar paravermal cortical ablation or cooling in cats. Exp. Neurol. 36: 549-562.
36. Yu, J.. R. TARNECKI, W. W. CHAMBERS, C. N. Lru, AND J. KONORSKI. 1973. Mechanisms mediating ipsilateral limb hyperflexion after cerebellar paravermal cortical ablation or cooling. Exp. Neural. 38: 144- 156.





























