Embed Size (px)
Citation preview

Kidney International, Vol. 60 (2001), pp. 2164–2172
CELL BIOLOGY – IMMUNOLOGY – PATHOLOGY
Positive effect of the induction of p21WAF1/CIP1 on the course ofischemic acute renal failure
JUDIT MEGYESI, LUCIA ANDRADE, JOSE M. VIEIRA, JR., ROBERT L. SAFIRSTEIN,and PETER M. PRICE
Division of Nephrology, Department of Medicine, and Department of Physiology, University of Arkansas for Medical Sciencesand Department of Veterans Affairs Medical Center, Little Rock, Arkansas, USA
Positive effect of the induction of p21WAF1/CIP1 on the course of Experimental models of acute renal failure caused byischemic acute renal failure. disparate injuries, that is, ischemia, nephrotoxic drugs
Background. The p21 protein is found in the nucleus of most (such as cisplatin), and ureteral obstruction, result in se-cells where it modulates cell cycle activity. At low levels, p21vere and acute reduction in the glomerular filtration ratestabilizes interactions between D cyclins and their cyclin-and clearance of serum nitrogenous wastes, such as creat-dependent kinases (cdks), but at high levels after induction by
several different stress pathways, it causes cell cycle arrest. The inine and blood urea nitrogen (BUN). After ischemiap21 mRNA is induced in murine kidney after several types of and administration of many nephrotoxic drugs, there isacute renal failure, including cisplatin administration, ischemia- also varying degrees of kidney cell death, especially in-reperfusion, and ureteral obstruction. We reported that after
volving the S3 segment of the proximal tubules in thecisplatin injection, mice with a p21 gene deletion developeddeep cortex. We had previously demonstrated a rapidmuch more severe renal damage than wild-type mice. To disso-
ciate the effects of cisplatin-induced DNA damage and subse- and high level induction of a cell-cycle inhibitory protein,quent initiation of DNA damage-dependent cell death path- p21, in murine kidney after acute renal failure [1]. p21,ways from effects of acute renal failure, we have now examined also known as WAF1 [2] and CIP1 [3], is a protein ofmice after ischemia-reperfusion, a model of renal failure not 21 kD [4] and is also a member of a protein family con-associated with genotoxin-induced DNA damage early after
taining p27KIP1 [5] and p57KIP2 [6, 7]. The p21 protein hasthe injury.been shown to be overexpressed before terminal differen-Methods. Wild-type and p21(�/�) mice were made ischemic
by clamping both renal hila for 30 or 50 minutes. At various tiation [8–14], and in cultured cells undergoing senescencetimes after reflow, mortality and parameters of renal function [15]. In addition, the p21 gene is induced after DNAand morphology were quantified. Also, the nuclear proteins damage [2], and its induction is necessary for the p53-p21 and proliferating cell nuclear antigen (PCNA) were local-
dependent cell-cycle arrest after DNA damage [16–18].ized in kidney sections by immunohistochemistry.Induction of the p21 gene and overexpression of the p21Results. Kidney function was more impaired and mortality
increased significantly in p21(�/�) mice as compared with protein is associated with cell-cycle interruption and G1p21(�/�) mice. We found more cell cycle activity, indicated arrest [2, 3, 19–22], and p21 has been shown to inhibitby increased number of mitotic cells and nuclear PCNA-posi- one or more of the cyclin-cdk kinase activities [3, 19,tive cells, in kidney of p21(�/�) mice. 22]. The protein can also bind [23] and inhibit [24, 25]Conclusions. In this study, p21(�/�) mice were more sus-
proliferating cell nuclear antigen (PCNA), especially itsceptible to ischemia-induced acute renal failure, with similarlyrole in DNA replication rather than DNA repair [26].elevated levels of parameters of cell cycle activity. We propose
that the increased and inappropriate cell cycle activity in kidney The role of p21 in differentiation is probably redundant,cells is responsible for the increased kidney impairment and since mice lacking p21 develop normally [17]. We hadmortality. previously reported that the expression of p21 was bene-
ficial to kidney cells after cisplatin administration [27],whereas its expression in chronic renal failure enhancedprogression [28]. Its role in ischemic acute renal failureis the subject of this investigation.
Key words: cell cycle, cyclin-dependent kinase inhibitor, cisplatin, mi-totic cells, apoptosis, kidney impairment.
METHODSReceived for publication March 29, 2001 p21(�/�) mice and induction of acute renal failureand in revised form June 13, 2001Accepted for publication July 5, 2001 Mice carrying a deletion of a large portion of the p21
gene in which neither p21 mRNA nor p21 protein is 2001 by the International Society of Nephrology
2164

Megyesi et al: p21 affects ischemic ARF 2165
expressed were obtained from Dr. Philip Leder (Harvard extensive necrosis of all areas of the cortex and in whichMedical School) [17]. Mice homozygous for the p21 dele- necrotic tubules were present near the surface of thetion were selected from the offspring of heterozygous kidney. Each parameter was determined on at least fivematings using Southern blotting of tail DNA as described different animals. Statistical significance was assessed by[17]. Similarly, wild-type homozygous litter-mates were the two-sided Student t test for independent samples,selected as controls. Male mice from 8 to 15 weeks of and was indicated with asterisks (***) if the P valueage were studied as described. The animals used in these was 0.05 or less. To determine the overall morphologicstudies were housed at the Veterinary Medical Unit at assessment, data from the seven criteria of kidney dam-the John L. McClellan Memorial Veterans Hospital (Lit- age [brush border loss, casts, tubule dilatation, red bloodtle Rock, Arkansas, USA). When appropriate, animals cell (RBC) extravasation, inflammation, distal nephronwere painlessly sacrificed in accordance with methods of damage, and necrosis] was pooled and assessed.euthanasia approved by the Panel on Euthanasia of the
Apoptosis detection and quantificationAmerican Veterinary Medical Association. Ischemia wasinduced in anesthetized (sodium pentobarbital, 50 mg/kg, The TUNEL technique (ApopTag kit; Intergen, Inc.,IP) male mice by exposure of the kidneys under sterile Purchase, NY, USA) was used to detect apoptotic cellsconditions through a midline incision. The kidneys were in situ [29].decapsulated and both renal hila were clamped with smallvascular clamps for either 30 or 50 minutes and released. ImmunohistochemistryDuring the operation, the mice received approximately Nuclear antigens (p21 and PCNA) were determined1 mL of saline IP. Sham operations were also performed on paraffin embedded tissue sections using a microwavein which the kidneys were manipulated as described, antigen retrieval technique (Antigen Unmasking Solution;without induction of ischemia. After surgery, the animals Vector Laboratories, Burlingame, CA, USA). PCNA waswere returned to their cages and allowed free access to detected using a mouse monoclonal antibody (Calbio-food and water. Blood was drawn by retro-orbital bleed- chem, La Jolla, CA, USA) and a rabbit polyclonal anti-ing at the time of sacrifice for determining blood urea body (Calbiochem) was used to detect mouse p21. Bothnitrogen (BUN; Sigma No. 535; Sigma Chemical Co., St. antigens were visualized using the ABC Elite VectastainLouis, MO, USA) and creatinine concentration in serum. Kit (Vector Laboratories).Differences between mouse genotypes were assessed bythe two-sided Student t test for independent samples, Survival after ischemiaand was indicated in the P value was 0.05 or less.
The comparison of survival of mice from the two geno-types was by the Kaplan and Meier method [30]. Statisti-Morphologic assessmentcal significance was assessed by the log-rank score.At various times after ischemia, kidneys were removed,
immersed in 4% neutral-buffered formaldehyde and fixedfor 48 to 72 hours. The tissues were paraffin embedded RESULTSand processed for light microscopy. Sections of 5 �m Localization of p21 expression after renal ischemiawere stained with hematoxylin-eosin or periodic acid
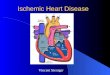
Nuclear p21 was localized by immunohistochemistrySchiff (PAS) and the histologic criteria determined. Theto tubules of both the distal and proximal nephron andfollowing parameters were chosen as indicative of mor-to regenerating tubules (Fig. 1). Staining was detected inphological damage to the kidney after ischemia: brushthick ascending limbs, distal convoluted tubules, proxi-border loss, tubular cast formation, tubular dilatation,mal tubules, and collecting ducts. There was no apparentred blood cell extravasation, inflammation, and distalstaining of cells in the glomerulus.nephron damage. These parameters were evaluated on
a scale of 0 to 4, which ranged from not present (0), mildKaplan-Meier survival curves(1), moderate (2), severe (3), to very severe (4). Tubular
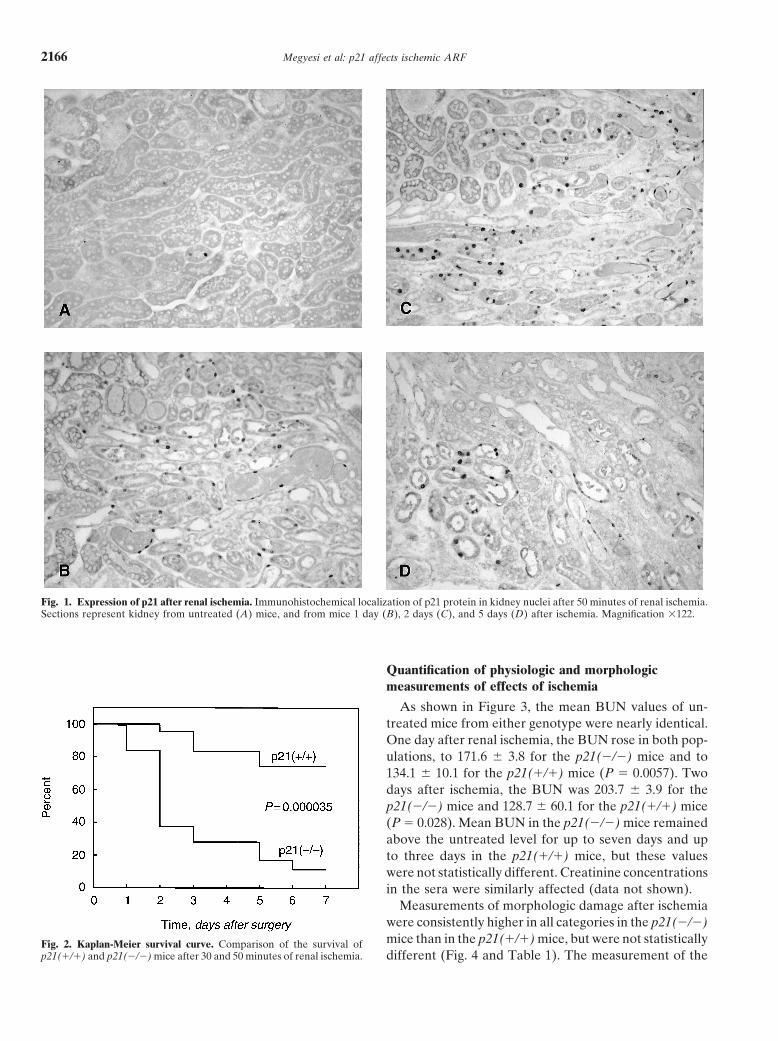
After renal ischemia, the survival rates of p21(�/�)necrosis was graded from none (0) to confluent (4). Theand p21(�/�) mice were compared (Fig. 2). The highestnumber of apoptotic (TUNEL-positive) cells per mm2
fatality rate in the p21(�/�) mice occurred between 24kidney tissue section was evaluated. The kidneys wereto 48 hours after the operation. After two days, thealso graded from 0 to 4 for relative numbers of mitoticpercentage surviving for the p21(�/�) and p21(�/�)cells and signs of tubular regeneration, that is, cells relin-populations was 95.5% and 37.6%, respectively. Aftering damaged tubules. The extent of the (cortical) necrosisseven days, the percentage surviving for the p21(�/�)was graded from 0 to 3 in which 0 � no necrosis, 1 �and p21(�/�) populations was 74.2% and 11.3%, respec-necrosis confined to the inner 1/3 of the cortex, primarilytively (P � 0.000035). There was no mortality in thethe S3 segment of the proximal tubules, 2 � necrosis
extending into the upper 2/3 of the cortex, and 3 � sham operated animals (data not shown).
Megyesi et al: p21 affects ischemic ARF2166
Fig. 1. Expression of p21 after renal ischemia. Immunohistochemical localization of p21 protein in kidney nuclei after 50 minutes of renal ischemia.Sections represent kidney from untreated (A) mice, and from mice 1 day (B), 2 days (C), and 5 days (D) after ischemia. Magnification �122.
Quantification of physiologic and morphologicmeasurements of effects of ischemia
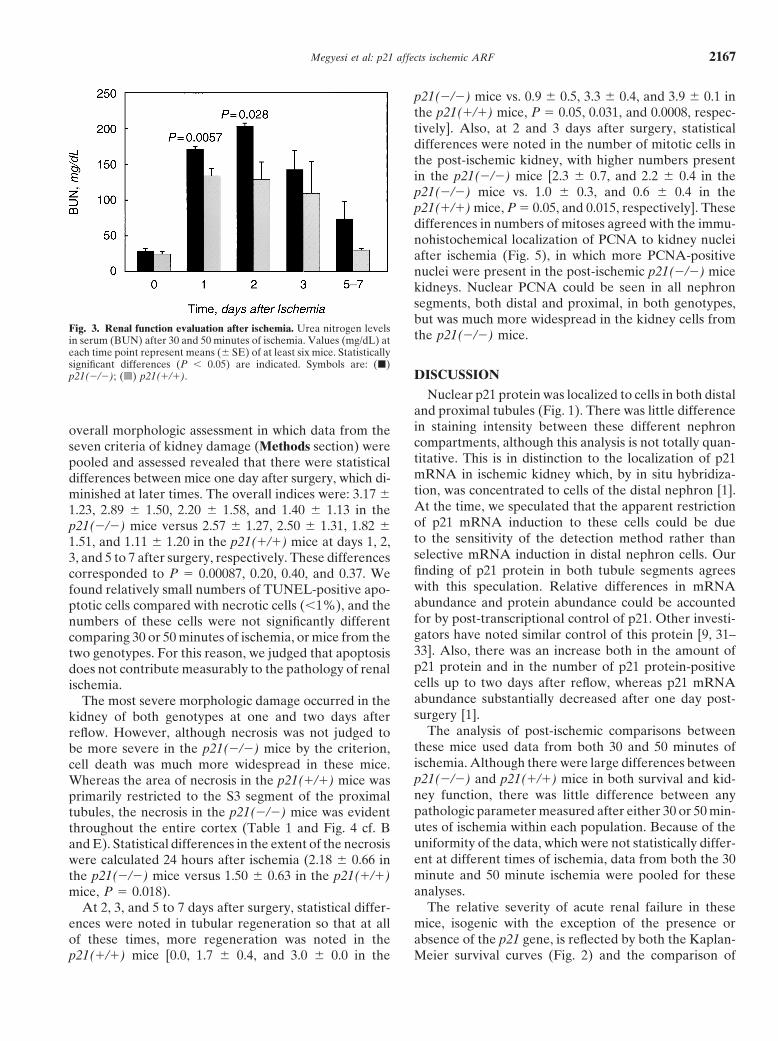
As shown in Figure 3, the mean BUN values of un-treated mice from either genotype were nearly identical.One day after renal ischemia, the BUN rose in both pop-ulations, to 171.6 � 3.8 for the p21(�/�) mice and to134.1 � 10.1 for the p21(�/�) mice (P � 0.0057). Twodays after ischemia, the BUN was 203.7 � 3.9 for thep21(�/�) mice and 128.7 � 60.1 for the p21(�/�) mice(P � 0.028). Mean BUN in the p21(�/�) mice remainedabove the untreated level for up to seven days and upto three days in the p21(�/�) mice, but these valueswere not statistically different. Creatinine concentrationsin the sera were similarly affected (data not shown).
Measurements of morphologic damage after ischemiawere consistently higher in all categories in the p21(�/�)mice than in the p21(�/�) mice, but were not statisticallyFig. 2. Kaplan-Meier survival curve. Comparison of the survival of
p21(�/�) and p21(�/�) mice after 30 and 50 minutes of renal ischemia. different (Fig. 4 and Table 1). The measurement of the
Megyesi et al: p21 affects ischemic ARF 2167
p21(�/�) mice vs. 0.9 � 0.5, 3.3 � 0.4, and 3.9 � 0.1 inthe p21(�/�) mice, P � 0.05, 0.031, and 0.0008, respec-tively]. Also, at 2 and 3 days after surgery, statisticaldifferences were noted in the number of mitotic cells inthe post-ischemic kidney, with higher numbers presentin the p21(�/�) mice [2.3 � 0.7, and 2.2 � 0.4 in thep21(�/�) mice vs. 1.0 � 0.3, and 0.6 � 0.4 in thep21(�/�) mice, P � 0.05, and 0.015, respectively]. Thesedifferences in numbers of mitoses agreed with the immu-nohistochemical localization of PCNA to kidney nucleiafter ischemia (Fig. 5), in which more PCNA-positivenuclei were present in the post-ischemic p21(�/�) micekidneys. Nuclear PCNA could be seen in all nephronsegments, both distal and proximal, in both genotypes,but was much more widespread in the kidney cells from
Fig. 3. Renal function evaluation after ischemia. Urea nitrogen levels the p21(�/�) mice.in serum (BUN) after 30 and 50 minutes of ischemia. Values (mg/dL) ateach time point represent means (� SE) of at least six mice. Statisticallysignificant differences (P � 0.05) are indicated. Symbols are: (�)
DISCUSSIONp21(�/�); ( ) p21(�/�).
Nuclear p21 protein was localized to cells in both distaland proximal tubules (Fig. 1). There was little differencein staining intensity between these different nephronoverall morphologic assessment in which data from thecompartments, although this analysis is not totally quan-seven criteria of kidney damage (Methods section) weretitative. This is in distinction to the localization of p21pooled and assessed revealed that there were statisticalmRNA in ischemic kidney which, by in situ hybridiza-differences between mice one day after surgery, which di-tion, was concentrated to cells of the distal nephron [1].minished at later times. The overall indices were: 3.17 �At the time, we speculated that the apparent restriction1.23, 2.89 � 1.50, 2.20 � 1.58, and 1.40 � 1.13 in theof p21 mRNA induction to these cells could be duep21(�/�) mice versus 2.57 � 1.27, 2.50 � 1.31, 1.82 �to the sensitivity of the detection method rather than1.51, and 1.11 � 1.20 in the p21(�/�) mice at days 1, 2,selective mRNA induction in distal nephron cells. Our3, and 5 to 7 after surgery, respectively. These differencesfinding of p21 protein in both tubule segments agreescorresponded to P � 0.00087, 0.20, 0.40, and 0.37. Wewith this speculation. Relative differences in mRNAfound relatively small numbers of TUNEL-positive apo-abundance and protein abundance could be accountedptotic cells compared with necrotic cells (�1%), and thefor by post-transcriptional control of p21. Other investi-numbers of these cells were not significantly differentgators have noted similar control of this protein [9, 31–comparing 30 or 50 minutes of ischemia, or mice from the33]. Also, there was an increase both in the amount oftwo genotypes. For this reason, we judged that apoptosisp21 protein and in the number of p21 protein-positivedoes not contribute measurably to the pathology of renalcells up to two days after reflow, whereas p21 mRNAischemia.abundance substantially decreased after one day post-The most severe morphologic damage occurred in thesurgery [1].kidney of both genotypes at one and two days after
The analysis of post-ischemic comparisons betweenreflow. However, although necrosis was not judged tothese mice used data from both 30 and 50 minutes ofbe more severe in the p21(�/�) mice by the criterion,ischemia. Although there were large differences betweencell death was much more widespread in these mice.p21(�/�) and p21(�/�) mice in both survival and kid-Whereas the area of necrosis in the p21(�/�) mice wasney function, there was little difference between anyprimarily restricted to the S3 segment of the proximalpathologic parameter measured after either 30 or 50 min-tubules, the necrosis in the p21(�/�) mice was evidentutes of ischemia within each population. Because of thethroughout the entire cortex (Table 1 and Fig. 4 cf. Buniformity of the data, which were not statistically differ-and E). Statistical differences in the extent of the necrosisent at different times of ischemia, data from both the 30were calculated 24 hours after ischemia (2.18 � 0.66 inminute and 50 minute ischemia were pooled for thesethe p21(�/�) mice versus 1.50 � 0.63 in the p21(�/�)analyses.mice, P � 0.018).
The relative severity of acute renal failure in theseAt 2, 3, and 5 to 7 days after surgery, statistical differ-mice, isogenic with the exception of the presence orences were noted in tubular regeneration so that at allabsence of the p21 gene, is reflected by both the Kaplan-of these times, more regeneration was noted in the
p21(�/�) mice [0.0, 1.7 � 0.4, and 3.0 � 0.0 in the Meier survival curves (Fig. 2) and the comparison of
Megyesi et al: p21 affects ischemic ARF2168
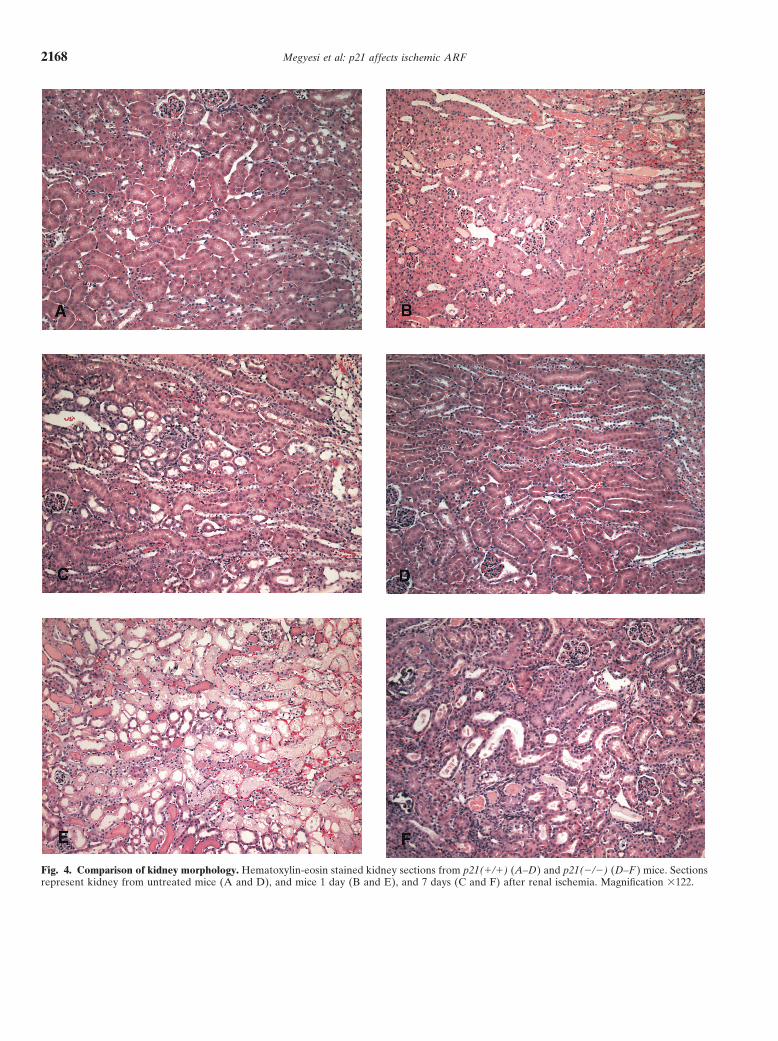
Fig. 4. Comparison of kidney morphology. Hematoxylin-eosin stained kidney sections from p21(�/�) (A–D) and p21(�/�) (D–F ) mice. Sectionsrepresent kidney from untreated mice (A and D), and mice 1 day (B and E), and 7 days (C and F) after renal ischemia. Magnification �122.

Megyesi et al: p21 affects ischemic ARF 2169
BUN values (Fig. 3) after surgery. Statistical differencesin BUNs occurred at days 1 and 2 after surgery, andcoincided with the greatest death rates of the p21(�/�)mice. Although BUNs in the p21(�/�) mice were higherthan those in the p21(�/�) mice from days 3 to 7, thesignificantly higher mortality in the p21(�/�) group,which eliminated the most severely affected animalsfrom the population [62.4% of the p21(�/�) mice butonly 4.5% of the p21(�/�) mice died by day 2] probablycontributed to the lack of statistical difference in BUNvalues between the two genotypes after day 2.
Interestingly, even with the difference in BUN valuesand the even larger difference in survival between thetwo genotypes, there was little statistical difference notedbetween many of the morphologic assessment compari-sons (Table 1). The best statistical difference (P �0.00087) that could be obtained was when the sevenmorphologic criteria were pooled, when it was notedthere was more damage in the p21(�/�) kidney one dayafter ischemia. Similar to the lack of statistical differ-ences noted in BUN values at later time points, we thinkthe lack of apparent differences in the morphologic dam-age between the two genotypes at later times is becauseof the very high mortality in the p21(�/�) mice popula-tion starting day 1 after surgery, effectively eliminatingthe most affected mice from the study. The most appar-ent difference noted here was in the extent of necrosiswithin the cortex and outer medulla. Necrosis was evi-dent throughout wide areas of the kidney cortex inp21(�/�) mice after ischemia (Fig. 4E), whereas in thep21(�/�) mice, necrosis, although confluent, was pri-marily confined to the S3 segment (Fig. 4B). The greaterextent of cell death and necrosis throughout the kidneycortex in the p21(�/�) population and the increasedBUN values were the most likely causes of the increasedmortality in this group.
After ischemia-reperfusion injury, the most suscepti-ble section of the kidney to cell death, especially in wild-type animals, is the S3 segment of the cortex. The proxi-mal tubules within this area are the most sensitive andcommonly undergo necrosis. Distal nephron segmentsusually survive the injury, but the small percentage thatdie usually die from apoptosis. However, during the re-generation following ischemic injury, larger numbers ofapoptotic cells are known to occur [34–36]. We foundthat apoptosis is a minor event in the first few days afterischemia, and have seen few TUNEL-positive nuclei inkidneys of mice of either genotype. Even in some casesin which distal nephron damage was found to occur,most cell death was necrotic and few apoptotic cells wereobserved. Forbes et al found that TUNEL-positive cellsin the kidney one day after ischemia did not have charac-teristic morphologic changes associated with apoptoticcells, and concluded that maximum apoptosis occurredabout a week after surgery [36]. This agrees with Zager
Tab
le1.
His
tolo
gic
eval
uati
onof
kidn
eyaf
ter
rena
lis
chem
ia
Bru
shbo
rder
Tub
ule
RB
CD
ista
lne
phro
nT
ubul
eE
xten
tof
loss
Cas
tsdi
lata
tion
extr
avas
atio
nIn
flam
mat
ion
dam
age
Nec
rosi
sre
gene
rati
onM
itos
esne
cros
isD
ays
afte
rsu
rger
y�
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/��
/�
13.
8�
0.5
3.9
�0.
33.
8�
0.4
3.8
�0.
32.
2�
1.1
2.8
�1.
43.
1�
0.8
3.4
�1.
01.
4�
0.9
1.8
�1.
32.
1�
1.3
2.4
�1.
63.
6�
0.5
3.9
�0.
30.
0�
0.0
0.0
�0.
00.
1�
0.1
0.1
�0.
11.
5�
0.6
2.2
�0.
7a
23.
7�
0.4
4.0
�0.
13.
6�
0.5
4.0
�0.
02.
1�
1.0a
3.3
�0.
6a2.
6�
0.8
2.7
�1.
31.
6�
0.7
1.3
�1.
40.
4�
0.6
1.0
�1.
53.
4�
0.7
4.0
�0.
00.
9�
0.5a
0.0
�0.
0a1.
0�
0.3a
2.3
�0.
7aN
DN
D3
3.1
�0.
93.
5�
0.6
3.4
�0.
93.
5�
0.6
2.6
�1.
43.
6�
0.4
1.8
�0.
51.
6�
0.5
0.1
�0.
30.
3�
0.4
0.4
�0.
80.
0�
0.0
1.9
�1.
82.
9�
1.2
3.3
�0.
4a1.
7�
0.4a
0.6
�0.
4a2.
2�
0.4a
ND
ND
5–7
2.5
�0.
52.
3�
0.6
1.9
�1.
22.
3�
1.2
2.0
�1.
21.
8�
0.8
1.3
�0.
71.
7�
0.6
0.0
�0.
00.
0�
0.0
0.0
�0.
00.
0�
0.0
0.1
�0.
11.
7�
1.2
3.9
�0.
1a3.
0�
0.0a
0.1
�0.
10.
2�
0.1
ND
ND
Qua
ntit
ativ
eev
alua
tion
ofm
orph
olog
icki
dney
dam
age
1,2,
3,an
d5–
7da
ysof
repe
rfus
ion
afte
r30
or50
min
utes
ofre
nal
isch
emia
expr
esse
dre
lati
vely
acco
rdin
gto
the
Met
hods
sect
ion.
Val
ues
repr
esen
tm
eans
(N�
SD)
ofki
dney
sect
ions
from
atle
ast
five
mic
efo
rea
chti
me
poin
tan
dge
noty
pe.R
epre
sent
ativ
eph
otom
icro
grap
hsfr
omth
ese
tim
epo
ints
and
geno
type
sar
ein
Fig
ure
4.M
orph
olog
yw
assc
ored
acco
rdin
gto
prox
imal
tubu
lebr
ush
bord
erlo
ss,c
ast
form
atio
nw
ithi
ntu
bule
s(C
asts
),tu
bule
dila
tati
on,r
edbl
ood
cell
extr
avas
atio
n,in
flam
mat
ion,
dist
alne
phro
nda
mag
e,tu
bule
necr
osis
,tub
ule
rege
nera
tion
,and
mit
oses
.ND
isno
tde
term
ined
aSt
atis
tica
llysi
gnif
ican
tdi
ffer
ence
s,P
�0.
05
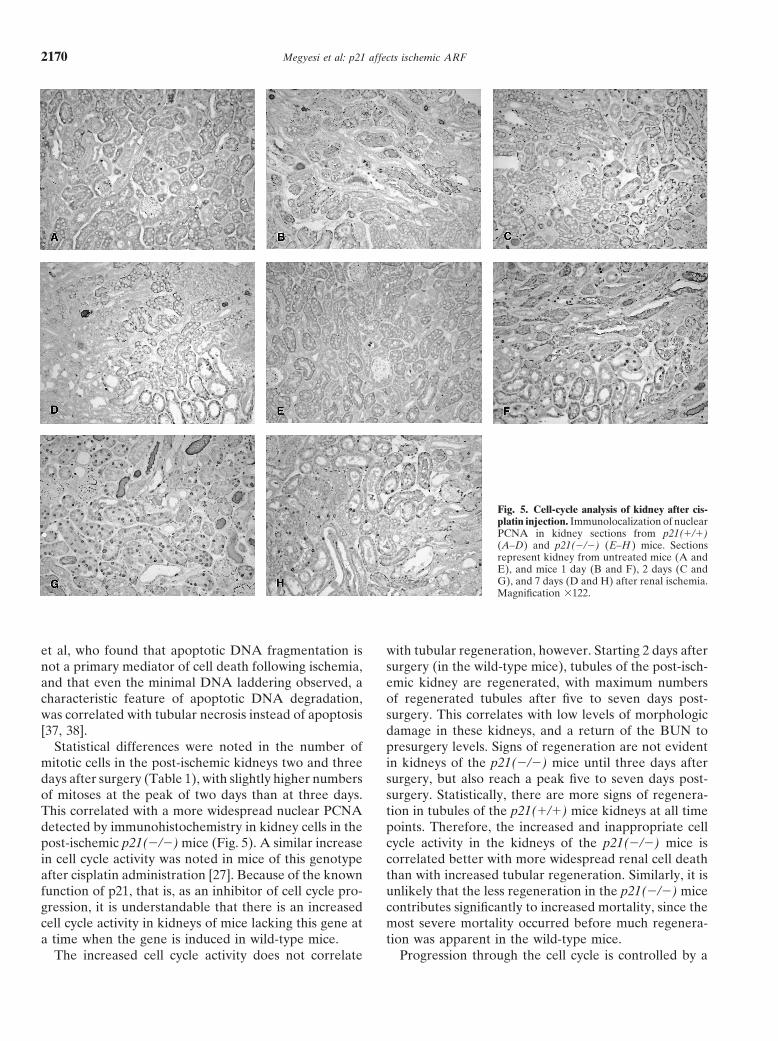
Megyesi et al: p21 affects ischemic ARF2170
Fig. 5. Cell-cycle analysis of kidney after cis-platin injection. Immunolocalization of nuclearPCNA in kidney sections from p21(�/�)(A–D) and p21(�/�) (E–H ) mice. Sectionsrepresent kidney from untreated mice (A andE), and mice 1 day (B and F), 2 days (C andG), and 7 days (D and H) after renal ischemia.Magnification �122.
et al, who found that apoptotic DNA fragmentation is with tubular regeneration, however. Starting 2 days aftersurgery (in the wild-type mice), tubules of the post-isch-not a primary mediator of cell death following ischemia,
and that even the minimal DNA laddering observed, a emic kidney are regenerated, with maximum numbersof regenerated tubules after five to seven days post-characteristic feature of apoptotic DNA degradation,
was correlated with tubular necrosis instead of apoptosis surgery. This correlates with low levels of morphologicdamage in these kidneys, and a return of the BUN to[37, 38].
Statistical differences were noted in the number of presurgery levels. Signs of regeneration are not evidentin kidneys of the p21(�/�) mice until three days aftermitotic cells in the post-ischemic kidneys two and three
days after surgery (Table 1), with slightly higher numbers surgery, but also reach a peak five to seven days post-surgery. Statistically, there are more signs of regenera-of mitoses at the peak of two days than at three days.
This correlated with a more widespread nuclear PCNA tion in tubules of the p21(�/�) mice kidneys at all timepoints. Therefore, the increased and inappropriate celldetected by immunohistochemistry in kidney cells in the
post-ischemic p21(�/�) mice (Fig. 5). A similar increase cycle activity in the kidneys of the p21(�/�) mice iscorrelated better with more widespread renal cell deathin cell cycle activity was noted in mice of this genotype
after cisplatin administration [27]. Because of the known than with increased tubular regeneration. Similarly, it isunlikely that the less regeneration in the p21(�/�) micefunction of p21, that is, as an inhibitor of cell cycle pro-
gression, it is understandable that there is an increased contributes significantly to increased mortality, since themost severe mortality occurred before much regenera-cell cycle activity in kidneys of mice lacking this gene at
a time when the gene is induced in wild-type mice. tion was apparent in the wild-type mice.Progression through the cell cycle is controlled by aThe increased cell cycle activity does not correlate

Megyesi et al: p21 affects ischemic ARF 2171
ple protein kinases and the DNA replication and repair factorcoordinated series of events, including activation andPCNA. Cell 71:505–514, 1992
repression, synthesis and degradation, and protein com- 5. Polyak K, Kato J, Solomon MJ, et al: p27Kip1, a cyclin-Cdk inhibi-tor, links transforming growth factor-B and contact inhibition topartmentalization. At two major transition points, pre-cell cycle arrest. Genes Dev 8:9–22, 1994ceding DNA synthesis and preceding mitosis, the cycle
6. Lee M-H, Reynisdottir I, Massague J: Cloning of p57KIP2, a cyclin-can be arrested by an over-abundance of proteins capa- dependent kinase inhibitor with unique domain structure and tissue
distribution. Genes Dev 9:639–649, 1995ble of inhibiting the activity of cyclin-dependent kinases.7. Matsuoka S, Edwards MC, Bai C, et al: p57KIP2, a structurallyA prominent member of this group of proteins is p21,
distinct member of the p21CIP1 Cdk inhibitor family, is a candidatealso known as WAF1 (2; wild-type p53-activated frag- tumor suppressor gene. Genes Dev 9:650–662, 1995
8. Halevy O, Novitch BG, Spicer DB, et al: Correlation of terminalment 1) and CIP1 (3; cyclin-dependent kinase interactingcell cycle arrest of skeletal muscle with induction of p21 by myoD.protein 1). We had found that p21 is transcriptionally Science 267:1018–1021, 1995
activated in kidney by p53-independent stress pathways 9. Macleod KF, Sherry N, Hannon G, et al: p53-Dependent andindependent expression of p21 during cell growth, differentiation,after the induction of acute renal failure by a variety ofand DNA damage. Genes Dev 9:935–944, 1995techniques [1]. In the case of acute renal failure induced 10. Parker SB, Eichele G, Zhang P, et al: p53-Independent expres-
by the DNA-damaging drug, cisplatin, it also is activated sion of p21Cip1 in muscle and other terminally differentiating cells.Science 267:1024–1027, 1995by the p53 transcription factor. In that model we had
11. Skapek SX, Rhee J, Spicer DB, Lassar AB: Inhibition of myogenicfound that the absence of the p21 gene is associated with differentiation in proliferating myoblasts by cyclin D1-dependent
kinase. Science 267:1022–1024, 1995an increased nephrotoxicity of the drug [27].12. Steinman RA, Hoffman B, Iro A, et al: Induction of p21 (WAF1/To evaluate whether increased protection also is af-
CIP1) during differentiation. Oncogene 9:3389–3396, 1994forded by over expression of the p21 gene when it is not 13. Zhang W, Grasso L, McClain CD, et al: p53-independent induc-
tion of WAF1/CIP1 in human leukemia cells is correlated withdependent on its activation by p53, we explored thegrowth arrest accompanying monocyte/macrophage differentiation.ischemic model of renal failure in which our previousCancer Res 55:668–674, 1995
study results were unable to demonstrate either nuclear 14. Jiang H, Lin J, Su ZZ, et al: Induction of differentiation in humanpromyelocytic HL-60 leukemia cells activates p21, WAF1/CIP1,p53 or p53-dependent gene activation [1]. It is apparentexpression in the absence of p53. Oncogene 9:3397–3406, 1994that in renal ischemia, as was seen after cisplatin, the 15. Noda AF, Ning Y, Venable S, et al: Cloning of senescent cell-
increased mortality caused by the absence of the p21 derived inhibitors of DNA synthesis using an expression screen.Exp Cell Res 211:90–98, 1994gene is correlated by inappropriate cell cycle activity in
16. Brugarolas J, Chandrasekaran C, Gordon JI, et al: Radiation-the post-ischemic kidney. However, most genes activated induced cell cycle arrest compromised by p21 deficiency. Natureby p53-dependent mechanisms also can be activated by 377:552–557, 1995
17. Deng C, Zhang P, Harper JW, et al: Mice lacking p21CIP1/WAF1other factors. It therefore remains to be seen whetherundergo normal development, but are defective in G1 checkpoint
stress pathways that are responsive to p53, but which control. Cell 82:675–684, 199518. Waldman T, Kinzler KW, Vogelstein B: p21 is necessary forcan be activated independently of DNA damage and
the p53-mediated G1 arrest in human cancer cells. Cancer Res 55:p53 stabilization, could be responsible for the increased5187–5190, 1995
sensitivity of the p21(�/�) mice to renal injury. 19. Dulic V, Kaufmann WK, Wilson SJ, et al: p53-dependent inhibi-tion of cyclin-dependent kinase activities in human fibroblasts dur-ing radiation-induced G1 arrest. Cell 76:1013–1023, 1994ACKNOWLEDGMENTS
20. El-Deiry WS, Harper JW, O’Conner PM, et al: WAF1/CIP1 isinduced in p53-mediated G1 arrest and apoptosis. Cancer Res 54:This work was supported by National Institute of Diabetes and1169–1174, 1994Digestive and Kidney Diseases grant 5R01DK54471 and with resources
21. Gu Y, Turck CW, Morgan DO: Inhibition of cdk2 activity in vivoand the use of facilities at the John L. McClennan Memorial Veterans’by an associated 20K regulatory subunit. Nature 366:707–710, 1993Hospital, Little Rock, AR, USA. We thank Dr. Philip Leder (Harvard
22. Xiong Y, Hannon GJ, Zhang H, et al: p21 is a universal inhibitorMedical School) for providing several heterozygous mice carrying theof cyclin kinases. Nature 366:701–704, 1993p21 gene deletion, and for providing a probe for screening.
23. Xiong Y, Zhang H, Beach D: Subunit rearrangement of the cyclin-dependent kinases is associated with cellular transformation. GenesReprint requests to Peter M. Price, Ph.D., Division of Nephrology,Dev 7:1572–1583, 1993Department of Internal Medicine, University of Arkansas for Medical
24. Flores-Rozas H, Kelman Z, Dean FB, et al: Cdk-interactingSciences, 4300 West 7th Street, mail route 151, Little Rock, Arkansasprotein 1 directly binds with proliferating cell nuclear antigen and72205, USA.inhibits DNA replication catalyzed by the DNA polymerase deltaE-mail: [email protected]. Proc Natl Acad Sci USA 91:8655–8659, 1994
25. Waga S, Hannon GJ, Beach D, Stillman B: The p21 inhibitor ofcyclin-dependent kinases controls DNA replication by interactionREFERENCESwith PCNA. Nature 369:574–578, 1994
1. Megyesi J, Udvarhelyi N, Safirstein RL, Price PM: The p53- 26. Li R, Waga S, Hannon GJ, et al: Differential effects by the p21independent activation of transcription of p21WAF1/CIP1/SDI1 after acute CDK inhibitor on PCNA-dependent DNA replication and repair.renal failure. Am J Physiol 271:F1211–F1216, 1996 Nature 371:534–537, 1994
2. El-Deiry WS, Tokino T, Velculescu VE, et al: WAF-1, a potential 27. Megyesi J, Safirstein RL, Price PM: Induction of p21WAF1/CIP1/SDI1
mediator of p53 tumor suppression. Cell 75:817–825, 1993 in kidney tubule cells affects the course of cisplatin-induced acute3. Harper JW, Adami GR, Wei N, et al: The p21 cdk-interacting renal failure. J Clin Invest 101:777–782, 1998
protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. 28. Megyesi J, Price PM, Tamayo E, Safirstein RL: The lack of aCell 75:805–816, 1993 functinal p21WAF1/CIP1 gene ameliorates progression to chronic renal
failure. Proc Natl Acad Sci USA 96:10830–10835, 19994. Xiong Y, Zhang H, Beach D: D type cyclins associate with multi-

Megyesi et al: p21 affects ischemic ARF2172
29. Gavrieli Y, Sherman Y, Ben-Sasson SA: Identification of pro- 34. Hammerman MR: Regulation of cell survival during renal develop-grammed cell death in situ via specific labeling of nuclear DNA ment. Pediatr Nephrol 12:596–602, 1998fragmentation. J Cell Biol 119:493–501, 1992 35. Shimizu A, Yamanaka N: Apoptosis and cell desquamation in
30. Peto R, Pike MC, Armitage P, et al: Design and analysis of random- repair process of ischemic tubular necrosis. Virchows Arch B 64:ized clinical trials requiring prolonged observation of each patient. 171–180, 1993II Analysis and examples. Br J Cancer 35:1–39, 1977 36. Forbes JM, Hewitson TD, Becker BJ, Jones CL: Ischemic acute
31. Timchenko NA, Harris TE, Wilde M, et al: CCAAT/enhancer renal failure: Long-term histology of cell and matrix changes inbinding protein alpha regulates p21 protein and hepatocyte prolif- the rat. Kidney Int 57:2375–2385, 2000eration in newborn mice. Mol Cell Biol 17:7353–7361, 1997
37. Zager RA, Fuerstenberg SM, Baehr PH, et al: An evaluation32. Kivinen L, Tsubari M, Haapajarvi T, et al: Ras induces p21Cip1/of antioxidant effects on recovery from postischemic acute renalWaf1 cyclin kinase inhibitor transcriptionally through Sp1-bindingfailure. J Am Soc Nephrol 4:1588–1597, 1994sites. Oncogene 18:6252–6261, 1999
38. Iwata M, Myerson D, Torok-Storb B, Zager RA: An evaluation33. Mitra J, Dai CY, Somasundaram K, et al: Induction of p21(WAF1/of renal tubular DNA laddering in response to oxygen deprivationCIP1) and inhibition of Cdk2 mediated by the tumor suppressor
p16(INK4a). Mol Cell Biol 19:3916–3928, 1999 and oxidant injury. J Am Soc Nephrol 5:1307–1313, 1994