Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions,research libraries, and research funders in the common goal of maximizing access to critical research.
Postnatal Growth and Age Estimation in Marshall's Horseshoe Bat,Rhinolophus marshalliAuthor(s): Longru Jin, Luo Bo, Keping Sun, Ying Liu, Jennifer Pan Ho and Jiang FengSource: Acta Chiropterologica, 14(1):105-110. 2012.Published By: Museum and Institute of Zoology, Polish Academy of SciencesDOI: http://dx.doi.org/10.3161/150811012X654312URL: http://www.bioone.org/doi/full/10.3161/150811012X654312
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological,and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and bookspublished by nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercialinquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.

INTRODUCTION
Most newborn bats are altricial and experience
a postnatal growth period in which they develop
appropriate sensory and locomotor skills to become
independent from their mothers (Baptista et al.,2000). Many researchers have studied the changes
in behavior, physiology, and ecology of this period
to investigate life history traits and to estimate ma-
ternal investment (Kunz et al., 2009).
There have been numerous studies on postnatal
growth of bats under both natural (Hoying and
Kunz, 1998; McLean and Speakman, 2000; Chaver -
ri and Kunz, 2006; Allen et al., 2009; Liu et al.,2009a; Jin et al., 2010) and captive conditions
(Boyd and Myhill, 1987; Rajan and Marimuthu,
1999; Elangovan et al., 2003, 2007; Raghuram and
Marimuthu, 2007). It has been shown that measure-
ments of body mass, forearm length and total length
of the epiphyseal gap of the fourth metacarpal-pha-
langeal joint (De Fanis and Jones, 1995; Hoying and
Kunz, 1998) can be used to estimate the age of bats.
Forearm length is the most accurately measured and
the most reliable characteristic for estimating age
during the early linear growth period of bats. How -
ever, the length of the epiphyseal gap is best for
estimating age at later stages of postnatal growth
(Kunz and Anthony, 1982; Krochmal and Sparks,
2007). Body mass is less reliable for estimating the
age of growing bats because it is highly sensitive to
variations in short term nutritional intake, energy
expenditure, and daily water flux (Stern and Kunz,
1998).
Accurate age determination is important for be-
havioral, physiological and ecological studies (Kunz
and Hood, 2000). In the absence of age estimates, it
is impossible to determine growth rates, the timing
of sexual maturity, the periodicity of reproduction,
the development of various behavioral repertoires,
or the longevity of an animal (Elangovan et al.,2003). In addition, patterns of growth and develop-
ment vary among species and families of bats.
Growth parameters derived from nonlinear models
(e.g., logistic, Gompertz, and von Bertalanffy) are
especially valuable for drawing interspecific com-
parisons because growth equations are independ-
ent of body size and duration of the postnatal
growth period (Kunz and Robson, 1995). However,
Acta Chiropterologica, 14(1): 105–110, 2012PL ISSN 1508-1109 © Museum and Institute of Zoology PAS
doi: 10.3161/150811012X654312
Postnatal growth and age estimation in Marshall’s horseshoe bat,
Rhinolophus marshalli
LONGRU JIN1, 2, LUO BO1, 2, KEPING SUN1, 2, YING LIU1, 2, JENNIFER PAN HO3, and JIANG FENG1, 2, 4
1Jilin Key Laboratory of Animal Resource Conservation and Utilization, Northeast Normal University, Changchun 130024, China2Key Laboratory for Wetland Ecology and Vegetation Restoration of National Environmental Protection,
Northeast Normal University, Changchun 130024, China3Department of Integrative Biology and Physiology, University of California, Los Angeles, USA
4Corresponding author: E-mail: [email protected]
Based on mark-recapture data, we studied the postnatal growth of Marshall’s horseshoe bat (Rhinolophus marshalli) in Hekou
County, Yunnan Province, China. Our results detected no significant differences both in body mass and in forearm length between
males and females at birth. On average, young bats were not agile fliers until 31 days of age. Body mass and forearm length of pups
followed a linear pattern of growth until day 13, with mean growth rates of 0.17 g/day and 1.22 mm/day, respectively, and thereafter
growth rates decreased. Length of the total epiphyseal gap of the fourth metacarpal-phalangeal joint showed a linear increase up to
10 days followed by a linear decrease until day 40 with a mean rate of 0.09 mm/day. Together, two equations permitted estimation
of the age of R. marshalli pups between 1 and 40 days of age. Of the three nonlinear growth models (logistic, Gompertz, and von
Bertalanffy), the logistic equation provided the best fit to the empirical curves for body mass and forearm length.
Key words: Rhinolophus marshalli, postnatal growth, age estimation, nonlinear growth models

comparative studies among different taxa should be
based on the same model (Zullinger et al., 1984).
Most studies on postnatal growth in bats were
conducted on vespertilionids in temperate zones,
but only a few focused on rhinolophid bats (Sharifi,
2004a; Dietz et al., 2007; Funakoshi et al., 2010).
Marshall’s horseshoe bat (Rhinolophus marshalli) is a widespread species, ranging through Thailand,
Bur ma, Vietnam, Laos Peninsular Malaysia, and
more recently, China (Simmons, 2005; Zhang et al.,2009). However, little information is available on its
postnatal growth. Thus, our aims were to measure
the sizes of bat pups at birth, to derive age estima-
tion equations based on forearm length and the
length of the total epiphyseal gap, and to compare
the growth patterns of body mass and forearm length
using three non-linear equations.
MATERIALS AND METHODS
This study was conducted in Banshan Cave (22°36’N,
103°50’E, ca. 80 m long, 1.8 m wide, and 2 m high) in Hekou
County, Yunnan Province, China. The cave housed a mixed
colony of ca. 70 R. marshalli and 400 Hipposideros pomona.
The surrounding area was covered by dense forest of mostly
rubber tree (Hevea brasiliensis).
Prior investigation indicated that female R. marshalli un-
dergo parturition from late April to early May. Therefore, we
checked the cave daily from about 15 days before parturition
throughout the season (a total of 18 days). We hand-captured
neonates immediately following the nightly emergence of
adults. Neonates with an attached umbilical cord were assumed
to be 1 day old (Kunz and Robson, 1995). After the sex of each
pup was determined, a numbered aluminum alloy ring (Porzana
Ltd., United Kingdom) was placed on the forearm of each bat
for individual identification. In total, 31 neonates with umbilical
cords were marked.
Body mass was recorded to the nearest 0.01 g using an elec-
tronic balance (ProScale LC-50, United States). Forearm length
was measured to the nearest 0.01 mm with digital vernier
calipers (TESA-CAL IP67, Switzerland). The total length of the
epiphyseal gap was measured to the nearest 0.01 mm using
calipers while the wing of the bat was spread over a transparent
solid plastic sheet illuminated from below with a strong flash-
light to visualize the gap (Sharifi, 2004b). To minimize errors in
using vernier calipers, individual measurements were repeated
three times and the means were used in the analysis.
To limit disturbance and possible abandonment, marking of
young and measuring morphological characters of pups took
place when adults were away in the foraging areas and the pro-
cedure was completed within 1.5 hours. As soon as all young
had been measured and weighed, they were returned to
their sites, as close to their original locations in the cave as pos-
sible, before any females returned. The cave was visited every
three days (Table 1) and continued until the majority of pups
were volant and could no longer be captured, even with the aid
of mist-nets and a hand-net by the middle of June 2010.
Capturing of bats was performed with permission from the local
government.
An independent sample t-test was used to compare forearm
length and body mass of males and females at birth. Linear re-
gression equations were derived to predict age from pooled data
for forearm length (1–13 days) and total epiphyseal gap (10–40
days). To derive an age-predictive equation from the measure-
ments, the axes on the growth curve were reversed and age for
the specific periods were considered as the dependent variable
(Kunz and Anthony, 1982). Ninety-five percent confidence and
prediction intervals were plotted for the regression equations for
forearm length and total gap. In addition, growth data of body
mass and forearm length in young bats were fitted to the
three models: logistic, Gompertz and von Bertalanffy equations
(Zul linger et al., 1984). The equations were as follows:
logistic: W = A (1 + exp (-K (t - I)))-1
Gompertz: W = A exp (-exp (-K (t - I)))
von Bertalanffy: W = A (1 -(1 / 3) exp (-K (t - I)))3
where: A is the asymptotic value (g), W is the body mass (g) at
age t (days), K is the growth rate constant (day-1), and I is the
age at the inflection point (days). The parameters A and K in
each model were estimated for the growth of mass in the
neonate population. Similar equations were used for forearm
length. The Levenberg-Marquardt algorithm was used to derive
the best fit to the three nonlinear equations. Results from the
three models were compared by the goodness of fit obtained
from each model (Zullinger et al., 1984). All statistical analyses
were conducted using SPSS ver. 15.0 (SPSS Inc., Chicago, IL,
USA) and data were described using mean ± SD unless stated
otherwise.
RESULTS
Each adult observed females produced a single
offspring. For 31 newborns with umbilical cords, no
significant difference was found in forearm length
between males and females (t = -1.16, d.f. = 29,
P > 0.05). Similarly, body mass of male and female
did not differ significantly (t = -0.78, d.f. = 29,
P > 0.05). The mean values of forearm length and
body mass of newborns were 17.05 ± 0.41 mm and
2.85 ± 0.14 g, and they were 37.4% and 36.3% of
that of adult females, respectively.
At birth, young R. marshalli were naked and pink
with closed eyes, folded ears, and deciduous teeth.
106 L. Jin, L. Bo, K. Sun, Y. Liu, and J. Pan Ho
TABLE 1. Number of individuals captured in Banshan Cave
ParameterDay of sampling
1 4 7 10 13 16 19 22 25 28 31 34 37 40
Number of re-captured bats 31 28 26 27 24 25 23 21 18 19 15 13 8 6
Marked bats re-captured (%) – 90 84 87 77 81 74 68 59 61 48 42 26 19
Seven days after birth, the short, fine, and soft hair
of the pups was distinguishable. Sixteen days after
birth, their ears were erect and some pups’ eyes were
completely open. After 22 days, some young were
able to flutter and glide when they were released by
hand. After 31 days the mean body mass and fore-
arm length were 80.3% and 96.6% of adult values,
respectively. Additionally, most young bats were
observed to fly freely with gentle turns in the cave.
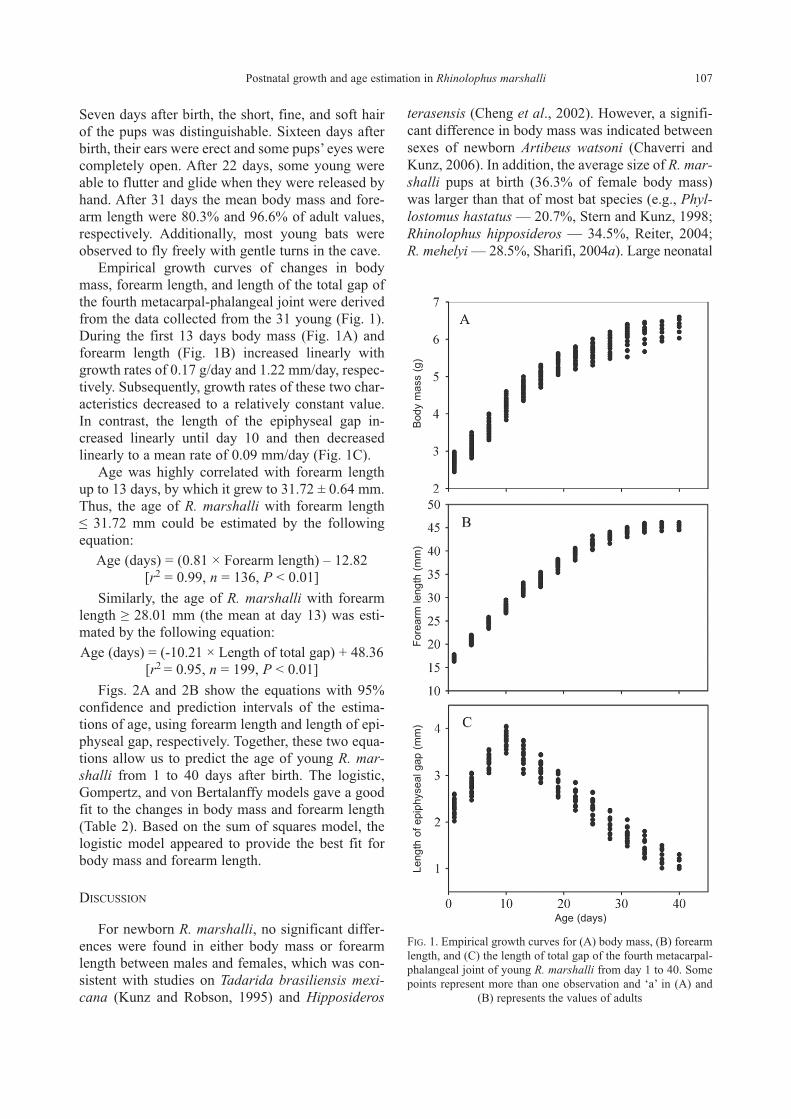
Empirical growth curves of changes in body
mass, forearm length, and length of the total gap of
the fourth metacarpal-phalangeal joint were derived
from the data collected from the 31 young (Fig. 1).
During the first 13 days body mass (Fig. 1A) and
forearm length (Fig. 1B) increased linearly with
growth rates of 0.17 g/day and 1.22 mm/day, respec-
tively. Subsequently, growth rates of these two char-
acteristics decreased to a relatively constant value.
In contrast, the length of the epiphyseal gap in-
creased linearly until day 10 and then decreased
linearly to a mean rate of 0.09 mm/day (Fig. 1C).
Age was highly correlated with forearm length
up to 13 days, by which it grew to 31.72 ± 0.64 mm.
Thus, the age of R. marshalli with forearm length
≤ 31.72 mm could be estimated by the following
equation:
Age (days) = (0.81 × Forearm length) – 12.82
[r2 = 0.99, n = 136, P < 0.01]
Similarly, the age of R. marshalli with forearm
length ≥ 28.01 mm (the mean at day 13) was esti-
mated by the following equation:
Age (days) = (-10.21 × Length of total gap) + 48.36
[r2 = 0.95, n = 199, P < 0.01]
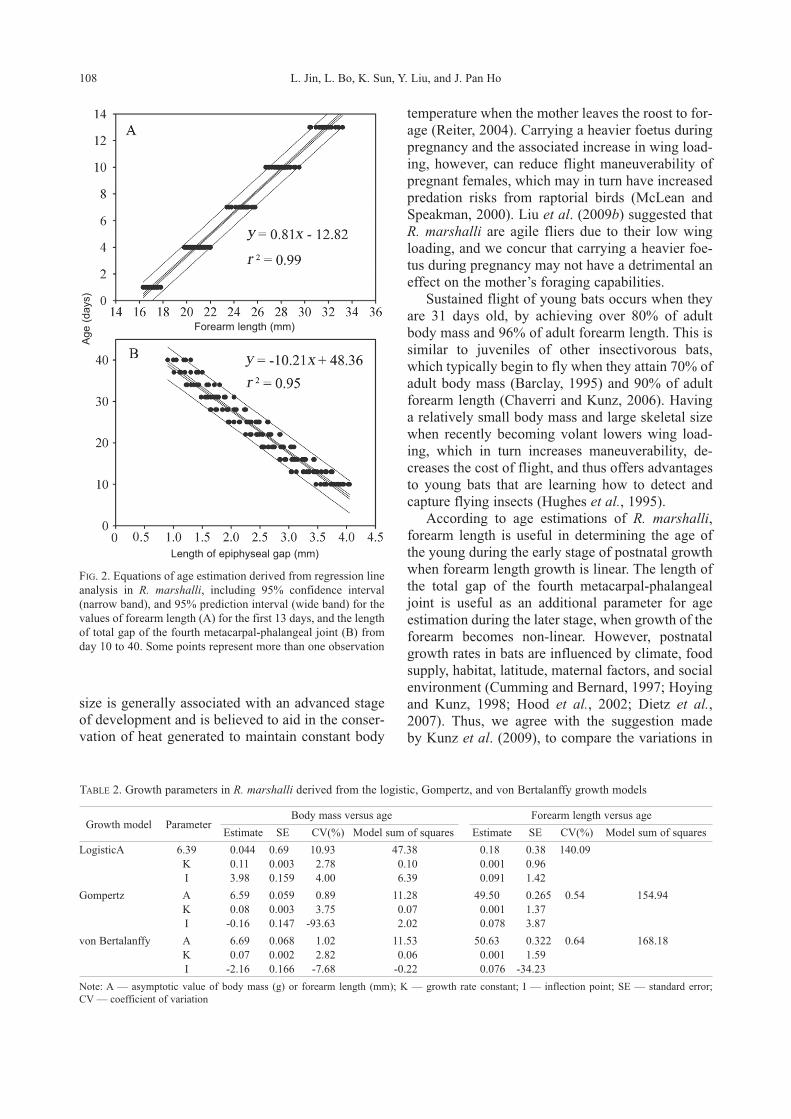
Figs. 2A and 2B show the equations with 95%
confidence and prediction intervals of the estima-
tions of age, using forearm length and length of epi-
physeal gap, respectively. Together, these two equa-
tions allow us to predict the age of young R. mar-shalli from 1 to 40 days after birth. The logistic,
Gom pertz, and von Bertalanffy models gave a good
fit to the changes in body mass and forearm length
(Table 2). Based on the sum of squares model, the
logistic model appeared to provide the best fit for
body mass and forearm length.
DISCUSSION
For newborn R. marshalli, no significant differ-
ences were found in either body mass or forearm
length between males and females, which was con-
sistent with studies on Tadarida brasiliensis mexi-cana (Kunz and Robson, 1995) and Hipposide ros
Postnatal growth and age estimation in Rhinolophus marshalli 107
FIG. 1. Empirical growth curves for (A) body mass, (B) forearm
length, and (C) the length of total gap of the fourth metacarpal-
phalangeal joint of young R. marshalli from day 1 to 40. Some
points represent more than one observation and ‘a’ in (A) and
(B) represents the values of adults
terasensis (Cheng et al., 2002). However, a signifi-
cant difference in body mass was indicated between
sexes of newborn Artibeus watsoni (Chaverri and
Kunz, 2006). In addition, the average size of R. mar-shalli pups at birth (36.3% of female body mass)
was larger than that of most bat species (e.g., Phyl-lo stomus hastatus — 20.7%, Stern and Kunz, 1998;
Rhi nolophus hipposideros — 34.5%, Reiter, 2004;
R. mehelyi — 28.5%, Sharifi, 2004a). Large neonatal
Length
of
epip
hyseal gap (
mm
)F
ore
arm
length
(m
m)
Body m
ass (
g)
Age (days)
Body mass versus age Forearm length versus ageGrowth model Parameter
Estimate SE CV(%) Model sum of squares Estimate SE CV(%) Model sum of squares
LogisticA 6.39 0.044 0.69 10.93 47.38 0.18 0.38 140.09
K 0.11 0.003 2.78 0.10 0.001 0.96
I 3.98 0.159 4.00 6.39 0.091 1.42
Gompertz A 6.59 0.059 0.89 11.28 49.50 0.265 0.54 154.94
K 0.08 0.003 3.75 0.07 0.001 1.37
I -0.16 0.147 -93.63 2.02 0.078 3.87
von Bertalanffy A 6.69 0.068 1.02 11.53 50.63 0.322 0.64 168.18
K 0.07 0.002 2.82 0.06 0.001 1.59
I -2.16 0.166 -7.68 -0.22 0.076 -34.23
Note: A — asymptotic value of body mass (g) or forearm length (mm); K — growth rate constant; I — inflection point; SE — standard error;
CV — coefficient of variation
size is generally associated with an advanced stage
of development and is believed to aid in the conser-
vation of heat generated to maintain constant body
108 L. Jin, L. Bo, K. Sun, Y. Liu, and J. Pan Ho
TABLE 2. Growth parameters in R. marshalli derived from the logistic, Gompertz, and von Bertalanffy growth models
FIG. 2. Equations of age estimation derived from regression line
analysis in R. marshalli, including 95% confidence interval
(narrow band), and 95% prediction interval (wide band) for the
values of forearm length (A) for the first 13 days, and the length
of total gap of the fourth metacarpal-phalangeal joint (B) from
day 10 to 40. Some points represent more than one observation
Length of epiphyseal gap (mm)
Forearm length (mm)
Age (
days)
temperature when the mother leaves the roost to for-
age (Reiter, 2004). Carrying a heavier foetus during
pregnancy and the associated increase in wing load-
ing, however, can reduce flight maneuverability of
pregnant females, which may in turn have increased
predation risks from raptorial birds (McLean and
Speakman, 2000). Liu et al. (2009b) suggested that
R. marshalli are agile fliers due to their low wing
loading, and we concur that carrying a heavier foe-
tus during pregnancy may not have a detrimental an
effect on the mother’s foraging capabilities.
Sustained flight of young bats occurs when they
are 31 days old, by achieving over 80% of adult
body mass and 96% of adult forearm length. This is
similar to juveniles of other insectivorous bats,
which typically begin to fly when they attain 70% of
adult body mass (Barclay, 1995) and 90% of adult
forearm length (Chaverri and Kunz, 2006). Having
a relatively small body mass and large skeletal size
when recently becoming volant lowers wing load-
ing, which in turn increases maneuverability, de-
creases the cost of flight, and thus offers advantages
to young bats that are learning how to detect and
capture flying insects (Hughes et al., 1995).
According to age estimations of R. marshalli,forearm length is useful in determining the age of
the young during the early stage of postnatal growth
when forearm length growth is linear. The length of
the total gap of the fourth metacarpal-phalangeal
joint is useful as an additional parameter for age
estimation during the later stage, when growth of the
forearm becomes non-linear. However, postnatal
growth rates in bats are influenced by climate, food
supply, habitat, latitude, maternal factors, and social
environment (Cumming and Bernard, 1997; Hoying
and Kunz, 1998; Hood et al., 2002; Dietz et al.,2007). Thus, we agree with the suggestion made
by Kunz et al. (2009), to compare the variations in

growth in different years and to generate geographi-
cally specific age equations whenever possible.
Growth curves as empirical models usually have
few parameters and are meant to reflect the nature
and dynamics of the underlying biological process-
es (Karkach, 2006). In our analysis, growth data for
forearm length and body mass were best described
by the logistic equation. The logistic model reflects
the rapid attainment of adult forearm length and
body mass, is computationally simple, and has bio-
logical relevance (Kunz and Robson, 1995). Similar
conclusions were made for Pipistrellus pipistrellus(Hughes et al., 1995), Plecotus auritus (De Fanis
and Jones, 1995) and T. b. mexicana (Kunz and
Robson, 1995). Given the relatively few studies that
have compared different models, the general signif-
icance of these differences remains unclear. In ad-
dition, on the basis of reviews by Kunz and Hood
(2000), growth constant rates of body mass of all
tropical microchiropterans were 0.04–0.11 (n = 11)
and the rates of most temperate microchiropteran
species were 0.12–0.25 (n = 13). In our study, the
growth constant rate of body mass was 0.10, which
embodied the regional features, i.e., a transition zone
between subtropical and tropical environments.
ACKNOWLEDGEMENTS
We thank Zheng Liu and his family for their invaluable field
assistance. This study was financed by the National Natural
Science Foundation of China (Grant Nos. 31030011, 31100305,
and 30900132) and China Postdoctoral Science Foundation
(Grant Nos. 20100481044 and 201104520).
LITERATURE CITED
ALLEN, L. C., C. S. RICHARDSON, G. F. MCCRACKEN, and T. H.
KUNZ. 2009. Birth size and postnatal growth in cave- and
bridge-roosting Brazilian free-tailed bats. Journal of Zool -
ogy (London), 280: 8–16.
BAPTISTA, T. L., C. S. RICHARDSON, and T. H. KUNZ. 2000.
Postnatal growth and age estimation in free-ranging bats:
a comparison of longitudinal and cross-sectional sampling
methods. Journal of Mammalogy, 81: 709–718.
BARCLAY, R. M. R. 1995. Does energy or calcium availability
constrain reproduction by bats? Symposia of the Zoological
Society of London, 67: 245–258.
BOYD, I. L., and D. G. MYHILL. 1987. Variations in the post-
natal growth of pipistrelle bats (Pipistrellus pipistrellus).
Jour nal of Zoology (London), 213: 750–755.
CHAVERRI, G., and T. H. KUNZ. 2006. Reproductive biology
and postnatal development in the tent-making bat Artibeuswatsoni (Chiroptera: Phyllostomidae). Journal of Zoology
(Lon d on), 270: 650–656.
CHENG, H. C, L. L. LEE, and W. L. GANNON. 2002. Postnatal
growth, age estimation, and sexual maturity in the For mo -
san leaf-nosed bat (Hipposideros terasensis). Journal of
Mam malogy, 83: 785–793.
CUMMING, G. S., and R. T. F. BERNARD. 1997. Rainfall, food
abundance and timing of parturition in African bats. Oecol -
ogia, 111: 309–317.
DE FANIS, E., and G. JONES. 1995. Postnatal growth, mother-
infant interactions and development of vocalizations in the
vespertilionid bat Plecotus auritus. Journal of Zoology
(Lon don), 235: 85–97.
DIETZ, C., I. DIETZ, and B. M. SIEMERS. 2007. Growth of horse-
shoe bats (Chiroptera: Rhinolophidae) in temperate conti-
nental conditions and the influence of climate. Mammalian
Biology, 72: 129–144.
ELANGOVAN, V., E. Y. S. PRIYA, H. RAGHURAM and G. MARI -
MUTHU. 2003. Postnatal development in the Indian short-
nosed fruit bat Cynopterus sphinx: growth rate and age esti-
mation. Acta Chiropterologica, 5: 107–116.
ELANGOVAN, V., E. Y. S. PRIYA, H. RAGHURAM and G. MARI -
MUTHU. 2007. Wing morphology and flight development in
the short-nosed fruit bat Cynopterus sphinx. Zoology, 110:
189–196.
FUNAKOSHI, K., E. NOMURA, M. MATSUKUBO, and Y. WAKITA.
2010. Postnatal growth and vocalization development of the
lesser horseshoe bat, Rhinolophus cornutus, in the Kyushu
district, Japan. Mammal Study, 35: 65–78.
HOOD, W. R., J. BLOSS, and T. H. KUNZ. 2002. Intrinsic and ex-
trinsic sources of variation in size at birth and rates of post-
natal growth in the big brown bat Eptesicus fuscus (Chiro -
ptera: Vespertilionidae). Journal of Zoology (London), 258:
355–363.
HOYING, K. M., and T. H. KUNZ. 1998. Variation in size at birth
and postnatal growth in the insectivorous bat Pipistrellussubflavus (Chiroptera: Vespertilionidae). Journal of Zoology
(London), 245: 15–27.
HUGHES, P. M., J. M. V. RAYNER, and G. JONES. 1995. Ontogeny
of ‘true’ flight and other aspects of growth in the bat Pi -pistrellus pipistrellus. Journal of Zoology (London), 236:
291–318.
JIN, L. R., A. Q. LIN, K. P. SUN, Y. LIU, and J. FENG. 2010.
Postnatal growth and age estimation in the ashy leaf-nosed
bat, Hipposideros cineraceus. Acta Chiropterologica, 12:
155–160.
KARKACH, A. 2006. Trajectories and models of individual
growth. Demographic Research, 15: 347–400.
KROCHMAL, A. R., and D. W. SPARKS. 2007. Timing of birth
and estimation of age of juvenile Myotis septentrionalis and
My otis lucifugus in west-central Indiana. Journal of Mam -
mal ogy, 88: 649–656.
KUNZ, T. H., and E. L. P. ANTHONY. 1982. Age estimation and
post natal growth in the bat Myotis lucifugus. Journal of
Mam malogy, 63: 23–32.
KUNZ, T. H., R. A. ADAMS, and W. R. HOOD. 2009. Methods
for assessing postnatal growth and development of bats.
Pp. 273–324, in Ecological and behavioral methods for the
study of bats, 2nd edition (T. H. KUNZ and S. PARSONS, eds.).
The Johns Hopkins University Press, Baltimore, 901 pp.
KUNZ, T. H., and W. R. HOOD. 2000. Parental care and postna-
tal growth in the Chiroptera. Pp. 415–468, in Reproductive
biology of bats (E. G. CRICHTON and P. H. KRUTZSCH, eds.).
Academic Press, London, 510 pp.
KUNZ, T. H., and S. K. ROBSON. 1995. Postnatal growth and
development in the Mexican free-tailed bat (Tadarida bra -siliensis mexicana): birth size, growth rates, and age estima-
tion. Journal of Mammalogy, 76: 769–783.
LIU, Y., L. R. JIN, W. METZNER and J. FENG. 2009a. Postnatal
Postnatal growth and age estimation in Rhinolophus marshalli 109

growth and age estimation in big-footed myotis, Myotismacrodactylus. Acta Chiropterologica, 11: 105–111.
LIU, Y., T. L. JIANG, S. BERQUIST, and J. FENG. 2009b. Vocal
characters and wing morphology of Rhinolophus marshallifrom Tiantang Cave, Guangxi Province, China. Mammalia,
73: 373–376.
MCLEAN, J. A., and J. R. SPEAKMAN. 2000. Morphological
changes during postnatal growth and reproduction in the
brown long-eared bat Plecotus auritus: implications for
wing loading and predicted flight performance. Journal of
Natural History, 34: 773–791.
RAGHURAM, H., and G. MARIMUTHU. 2007. Maternal feeding of
offspring with vertebrate prey in captive Indian false vam-
pire bat, Megaderma lyra. Acta Chiropterologica, 9: 437–443.
RAJAN, K. E., and G. MARIMUTHU. 1999. Postnatal growth and
age estimation in the Indian false vampire bat (Megadermalyra). Journal of Zoology (London), 248: 529–534.
REITER, G. 2004. Postnatal growth and reproductive biology of
Rhinolophus hipposideros (Chiroptera: Rhinolophidae).
Jour nal of Zoology (London), 262: 231–241.
SHARIFI, M. 2004a. Postnatal growth and age estimation in the
Mehelys horseshoe bat (Rhinolophus mehelyi). Acta Chiro -
pterologica, 6: 155–161.
SHARIFI, M. 2004b. Postnatal growth in Myotis blythii (Chiro -
ptera, Vespertilionidae). Mammalia, 68: 283–289.
SIMMONS, N. B. 2005. Order Chiroptera. Pp. 312–529, inMam mal species of the World: a taxonomic and geograph-
ic reference, 3rd edition (D. E. WILSON and D. M. REEDER,
eds.). The Johns Hopkins University Press, Baltimore,
2142 pp.
STERN, A. A., and T. H. KUNZ. 1998. Intraspecific variation in
postnatal growth in the greater spear-nosed bat. Journal of
Mammalogy, 79: 755–763.
ZHANG, L. B., G. JONES, J. S. ZHANG, G. J. ZHU, S. PARSONS, S.
J. ROSSITER, and S. Y. ZHANG. 2009. Recent surveys of bats
(Mammalia: Chiroptera) from China. I. Rhinolophidae and
Hipposideridae. Acta Chiropterologica, 11: 71–88.
ZULLINGER, E. M., R. E. RICKLEFS, K. H. REDFORD and G. M.
MACE. 1984. Fitting sigmoidal equations to mammalian
growth curves. Journal of Mammalogy, 65: 607–636.
110 L. Jin, L. Bo, K. Sun, Y. Liu, and J. Pan Ho
Received 05 November 2010, accepted 11 April 2011





























