Embed Size (px)
Citation preview

doi:10.1152/japplphysiol.00095.2013 114:1743-1754, 2013. First published 11 April 2013;J Appl Physiol
Gregory C. HendersonPatrick M. Davitt, Shawn M. Arent, Marc A. Tuazon, Devon L. Golem andresistance exercisemetabolism in obese women after either endurance or Postprandial triglyceride and free fatty acid
You might find this additional info useful...
78 articles, 26 of which can be accessed free at:This article cites /content/114/12/1743.full.html#ref-list-1
2 other HighWire hosted articlesThis article has been cited by
[PDF] [Full Text] [Abstract]
, January 1, 2014; 116 (1): 67-75.J Appl PhysiolEric C. Freese, Nicholas H. Gist and Kirk J. CuretonEffect of prior exercise on postprandial lipemia: an updated quantitative review
[PDF] [Full Text] [Abstract], January 1, 2014; 116 (1): 95-103.J Appl Physiol
Gregory C. Henderson and Brandon L. AldermanDeterminants of resting lipid oxidation in response to a prior bout of endurance exercise
including high resolution figures, can be found at:Updated information and services /content/114/12/1743.full.html
can be found at:Journal of Applied Physiologyabout Additional material and information http://www.the-aps.org/publications/jappl
This information is current as of September 11, 2014.
ISSN: 0363-6143, ESSN: 1522-1563. Visit our website at http://www.the-aps.org/.Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2013 by the American Physiological Society.those papers emphasizing adaptive and integrative mechanisms. It is published 12 times a year (monthly) by the American
publishes original papers that deal with diverse areas of research in applied physiology, especiallyJournal of Applied Physiology
on Septem
ber 11, 2014D
ownloaded from
on Septem
ber 11, 2014D
ownloaded from

Postprandial triglyceride and free fatty acid metabolism in obese women aftereither endurance or resistance exercise
Patrick M. Davitt,1 Shawn M. Arent,1 Marc A. Tuazon,1,2 Devon L. Golem,1 and Gregory C. Henderson1,2
1Department of Exercise Science and Sport Studies, Rutgers University, New Brunswick, New Jersey; and 2Rutgers Center forLipid Research, Rutgers University, New Brunswick, New Jersey
Submitted 24 January 2013; accepted in final form 3 April 2013
Davitt PM, Arent SM, Tuazon MA, Golem DL, HendersonGC. Postprandial triglyceride and free fatty acid metabolism inobese women after either endurance or resistance exercise. J ApplPhysiol 114: 1743–1754, 2013. First published April 11, 2013;doi:10.1152/japplphysiol.00095.2013.—We investigated the effectsof two exercise modalities on postprandial triglyceride (TG) and freefatty acid (FFA) metabolism. Sedentary, obese women were studiedon three occasions in randomized order: endurance exercise for 60min at 60–65% aerobic capacity (E), �60 min high-intensity resistanceexercise (R), and a sedentary control trial (C). After exercise, a liquid-mixed meal containing [U-13C]palmitate was consumed, and subjectswere studied over 7 h. Isotopic enrichment (IE) of plasma TG, plasmaFFA, and breath carbon dioxide compared with meal IE indicated thecontribution of dietary fat to each pool. Total and endogenouslyderived plasma TG content was reduced significantly in both E and Rcompared with C (P � 0.05), with no effect of exercise on circulatingexogenous (meal-derived) TG content. Exogenous plasma FFA con-tent was increased significantly following both E and R comparedwith C (P � 0.05), whereas total and endogenous FFA concentrationswere elevated only in E (P � 0.05) compared with C. Fatty acid (FA)oxidation rates were increased significantly after E and R comparedwith C (P � 0.05), with no difference between exercise modalities.The present results indicate that E and R may be equally effective inreducing postprandial plasma TG concentration and enhancing lipidoxidation when the exercise sessions are matched for duration ratherthan for energy expenditure. Importantly, tracer results indicated thatthe reduction in postprandial lipemia after E and R exercise bouts isnot achieved by enhanced clearance of dietary fat but rather, isachieved by reduced abundance of endogenous FA in plasma TG.
physical activity; adiposity; indirect calorimetry; isotopes; fat oxida-tion
INADEQUATE PHYSICAL ACTIVITY level (PAL) is associated withthe accumulation of excess body fat (32). Obesity has beenlinked with many disease risks, including cardiovascular dis-ease (CVD) and type 2 diabetes (34, 62). Previously, it hasbeen reported that obese individuals exhibit a lower capacityfor lipid oxidation (Lox) compared with their lean counterparts(31, 38, 69). A reduced relative contribution of Lox is associ-ated with increased risk of gaining weight over time (81), andso, substrate partitioning, along with effects of positive energybalance, could potentially contribute to the propensity for gainand retention of body fat. Fed-state plasma triglyceride (TG),referred to as postprandial lipemia, is elevated in obesity (13,47) and is associated with increased CVD risk (4, 18, 46, 57,67). Exercise can potentially address these metabolic altera-tions by impacting the resting metabolism for many hours after
each session (26). People typically take a meal after exerciseand are in the postprandial state for a majority of the day, andstudies of the effects of exercise on postprandial metabolismare needed to improve understanding the integration of meta-bolic processes and for preclinical testing of exercise efficacy.
Benefits of chronic exercise represent the accumulation ofthe acute benefits from each individual exercise bout, andchronic exercise can reduce CVD risk (32). An individualexercise bout can increase subsequent resting Lox (21, 26, 28,61, 71, 72) and reduce postprandial plasma TG concentration(1, 16, 19, 21, 22, 28, 35, 36, 50, 52, 66, 71, 72, 77–80). Inaddition to the effect of the energy expenditure (EE) of exer-cise (EEE) (22, 78), there has been some indication thatexercise intensity could be important in the response of post-prandial lipemia (35, 70) and postprandial Lox (70), althoughthis intensity effect was not apparent in all cases (16, 71). Ifexercise intensity were a factor in determining the subsequentresponse of postprandial metabolism, one could expect that resis-tance exercise, because of its vigorous nature, could be useful inaltering subsequent lipid metabolism. Attempts have also beenmade to compare the modalities alongside one another in asingle study; in these studies, bouts of endurance exercise (E)and resistance exercise (R) were designed to match for EE.When doing so, the R bout was challenging (near the maxi-mum intensity possible during the bout), whereas the E boutwas quite easy, apparently, approximately one-half (or evenless) of the intensity that an individual could tolerate well (49,61). So, when matched for EEE, R appears to be superior to Ein its metabolic impacts on plasma TG metabolism and Lox(49, 61). E inherently entails a high EEE, whereas R, althoughphysiologically stressful in other ways, does not lead to highEEE when performed for a similar duration as E exercise.Matching EEE addresses an interesting biological question, butwe sought to compare E and R in the present study in a mannerthat could be particularly translational and clinically relevant.We studied E and R bouts that were typical of those that maybe attempted in an exercise program (matched for duration;each at a challenging intensity; each consistent with currentexercise recommendations stating that higher intensities ofexercise may carry additional benefits beyond those of lowerintensities). In such an experimental design, applying a morechallenging E bout could potentially clarify implications ofprevious results that had suggested inferiority of E vs. R.
Changes in postprandial lipid metabolism after exercisecould entail an alteration in metabolism of the fat from thepostexercise meal (exogenous fat), and alternatively, thechanges could be for metabolism of fat that was already withinthe body before meal ingestion (endogenous fat). In identifyingwhether the benefits of exercise on postprandial lipid metabo-lism involve altered handling of the meal’s fat content, insight
Address for reprint requests and other correspondence: G. C. Henderson,Rutgers Univ., 70 Lipman Dr., New Brunswick, NJ 08901 (e-mail: [email protected]).
J Appl Physiol 114: 1743–1754, 2013.First published April 11, 2013; doi:10.1152/japplphysiol.00095.2013.
8750-7587/13 Copyright © 2013 the American Physiological Societyhttp://www.jappl.org 1743
on Septem
ber 11, 2014D
ownloaded from

is provided into the physiological impacts of exercise. Instudies of postexercise metabolism, stable isotope tracers canbe used to label meal fatty acid (FA) content to track dietary fatand differentiate between exogenous (meal-derived) and en-dogenous FA contribution to the plasma TG excursion, plasmafree FA (FFA), and Lox (14, 24, 44, 45). In a small number ofstudies, FA tracers have been administered in postexercisemeals (21, 74–76), and a subset of this analytical potential wasrealized as exogenous FA oxidation was measured by isotopelabeling of carbon dioxide. However, in light of the potential toalso follow these tracers through other fates and pathways, weexpanded this methodological approach to study the relativelabeling of circulating TG and FFA as well. As obese individ-uals exhibit exaggerated responses of plasma TG concentrationto a meal (13, 47), we considered it important to study thispopulation when attempting to manipulate metabolism of di-etary fat with exercise. To our knowledge, this is the first studyto use an oral-fed tracer to track alterations in Lox, as well asplasma TG and FFA concentrations and labeling after exercise.
Interventions that lead to a favorable postprandial FA me-tabolism may reduce CVD risk compared with a sedentarystate and could also promote a lean phenotype if the achievednegative fat balances were maintained without compensation.Therefore, with the use of the methodology of stable isotopetracers, we studied the effects of a premeal exercise bout onpostprandial TG and FA metabolism over the course of apostprandial period, following time-matched exercise bouts ofdifferent modalities, and compared the results with a sedentarycontrol. We studied women, who are traditionally under-rep-resented as study participants in the exercise literature, andobese individuals because of a similar under-representation, aswell as a need to alter lipid metabolism in this group. We testedthe hypothesis that FA oxidation in both exercise groups wouldincrease during the postexercise period compared with thesedentary control. We also hypothesized that either exerciseapproach would decrease plasma TG concentrations during thepostexercise period by a reduction in exogenous and endoge-nous fat content in circulation, whereas the concentration ofplasma FFA from both sources would increase after exercise.
METHODS
Study participants. Twelve sedentary, premenopausal, obese [bodymass index (BMI) � 30] women were recruited from Rutgers Uni-versity (New Brunswick campus) and its surrounding community byposted notice. Participants were required to have partaken in �1 h/wkof physical activity at moderate or higher intensity during the prior 3mo. Nine subjects were administered an isotope tracer, and three wereprovided unlabeled meals and solely included in indirect calorimetry(IDC) analysis. Due to a technical error in labeled meal preparationfor one participant, the total sample size was reduced to eight for thetracer analyses and remained at 12 for IDC. Potential study partici-pants underwent subsequent screening tests if they were disease free,as determined by a health history questionnaire, and were not takingmedications known to affect energy metabolism. Study participantsreported regular menstrual cycles (24–32 days); if taking oral contra-ceptives (OC), they were required to have been on them for at least 6mo and were instructed to continue taking the OC for the duration ofthe study. Effects of menstrual-cycle phase and OC were expected tobe subtle compared with effects of exercise per se (12, 68), and therandomization of trial order was meant to balance any minor effects ofmenstrual cycle and OC phase on substrate metabolism. The proce-dures and risks were explained thoroughly to the study participants,
and their written, informed consent was obtained. Rutgers UniversityInstitutional Review Board approved the study protocol (no. 11-030R).
Screening tests. Fasting blood glucose was tested to confirm that allstudy participants were nondiabetic (glucose � 125 mg/dl). Studyparticipants underwent a progressive exercise test to assess VO2peak
before beginning the study. The test and experimental trials wereperformed on a graded high-speed treadmill (TrackMaster, Newton,KS), and the oxygen consumption (VO2) and carbon dioxide produc-tion (VCO2) measurements were made using a metabolic cart(TrueOne 2400; ParvoMedics, Sandy, UT). A continual, progressiveprotocol was used to determine VO2peak with an increase in workloadat 3-min intervals until volitional exhaustion (2). Body compositionwas determined by measuring body volume via air-displacementplethysmography using the BOD POD (Life Measurement Instru-ments, Concord, CA), as described in previous literature (15). Bodycomposition and VO2peak were tested on the same day. On twoseparate occasions, 1) each subject was familiarized with the entireresistance-training protocol and 2) administered a 10-repetition max-imum (10 RM) test for each exercise (3).
Experimental design. With at least 1 wk between trials, participantswere studied under each of three conditions, each on separate occa-sions, assigned in a random order. Subjects were given a test meal, 30min after 1) 60 min of E at 60–65% VO2peak, 2) 60 min of R, and 3) atime-matched resting condition (C), and studied for a 400-min post-prandial period. A study design schematic is shown in Fig. 1.
Study participants were instructed to maintain their current dietaryand activity habits throughout the entire duration of the study. The daybefore each trial, study participants were instructed to consume solelytheir standardized diet and water ad libitum and to abstain fromstructured physical exercise sessions but to continue typical activitiesof daily living and were fed for a PAL of 1.4, according to the currentdietary reference intake guidelines of the Institutes of Medicine fordaily estimated energy requirement (EER) (32). Each standardizeddiet was provided to the study participant 2 days before the upcomingtrial and was consumed by the participant outside of the laboratory onthe day before and morning of each trial at the instructed times.Dietary energy intake was individualized for each study participant(2,294.0 � 81.2 kcal/day), and macronutrient composition was madesimilar among individuals for carbohydrate (CHO; 54.3 � 0.7%),lipid (25.4 � 0.5%), and protein (20.3 � 0.4%). On the day of trials,study participants consumed a provided bagel at 7:30 AM (81.9%CHO, 3.6% fat, 14.5% protein, 249 kcal) and arrived at the laboratoryat 8:30 AM. Upon arrival at the laboratory, participants confirmed thatthey had consumed the standardized diets. We chose to feed our studyparticipants 1.5 h before exercise to increase tolerance of the exerciseprotocol (pre-exercise meal described above).
-
Catheter into hand vein
8:30 amArrive at lab
MealExercise
Draw blood sample
Draw breath sample
Breakfast
nim 08nim 04
(Minutes)
5:30 pmDepart lab
Fig. 1. Experimental design. Each study participant completed 3 differentexperimental trials in randomized order, 1 involving endurance exercise (E), 1involving resistance exercise (R), and 1 with no exercise (C). Minutes � timeelapsed since the study participant commenced meal consumption. E or Rbouts were performed from �90 to �30 min or semisupine rest during thistime period in the C trial.
1744 Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from

Duration of E was exactly 60 min, and the R bout was designed tobe �60 min. The intensity for E was 60–65% VO2peak, which in ourexperience, approximates the maximum intensity that can be com-pleted over 1 h consistently by sedentary people. IDC was used duringthe E bout to achieve the desired intensity. The R trial was time-of-daymatched and duration matched and consisted of eight whole-bodyexercises, with three sets and 10 repetitions of each exercise (benchpress, lat pulldown, shoulder press, squat, leg curls, triceps pushdown,biceps curl, lunges) at 90% 10 RM. If 10 repetitions in a given setcould not be completed, then the weight was reduced to complete 10repetitions on the subsequent set. IDC was not used during the Rsession. Prior to exercise and at the same time of day in the sedentarycontrol, a single-use needle was used for the first blood draw. Aftereach exercise session (or again, same time in C), study participants satin a chair where venous catheters were placed (described below). Thetest meal (also described below) was given 30 min postexercise, afterwhich, participants remained seated for the remaining 400 min of thepostprandial period, quietly reading or watching movies. On the dayof each trial, at the same time of day, a catheter was placed in a handvein to obtain arterialized blood using the heated hand-vein technique.In a previous study (29), blood was drawn simultaneously from theradial artery and from a heated hand vein, and there were no differ-ences among blood-sampling sites for glucose, glycerol, or palmitateisotopic enrichment (IE) during tracer administration. The catheterwas kept patent throughout the study day, using a 0.9% salinesolution. Water was consumed ad libitum during recovery, but asidefrom the test meal, study participants consumed no other beveragesand no food until the 400-min postprandial measurement periodended. Study participants were transported in a wheelchair for trips tothe restroom. During each trial, urine was collected, and aliquots weretaken and stored at �80°C for subsequent analysis of creatinine andurinary urea nitrogen (UUN).
To study the partitioning of dietary FA postexercise, [U-13C]palmi-tate (Cambridge Isotope Laboratories, Andover, MA) was adminis-tered in a liquid test meal. Thirty minutes after the termination ofexercise, the postexercise blood draw and first IDC measurementswere taken. Blood samples for the analysis of TG and FA concentra-tion and palmitate IE were collected in tubes containing EDTA. Ateach breath-sampling time point, pulmonary gas exchange was deter-mined for assessment of metabolic rate and energy substrate parti-tioning, and an aliquot of expired breath was collected in evacuatedExetainer tubes for subsequent determination of 13CO2 IE by isotoperatio mass spectrometry (IRMS).
Test meal. The liquid test meal consisted of Boost Plus (NestléHealthCare Nutrition, Florham Park, NJ) and evaporated milk andwas administered immediately following the postexercise breathdraw, which was 30 min after cessation of exercise. The test mealprovided 20 kcal/kg fat-free mass (FFM; 46.2 � 1.1% of EER). The[U-13C]palmitate isotopic tracer dose of 5 mg/kg FFM was added tothe test meal, which was heated to 85–90°C, using a hot plate withconstant stirring, using a magnetic stir bar. The temperature of themeal was monitored with a thermometer; this procedure ensuressuccessful dissolution and delivery of the tracer, as reported previ-ously (14). The IE of meal palmitate was 5.7 � 0.2 mol percent excess(MPE), which we determined by liquid chromatography/mass spec-trometry (LC/MS) of meal aliquots, as described below. The macro-nutrient composition of the drink was 47.8% CHO, 36.1% fat, and16.1% protein (1,076.4 � 62.7 kcal), and the average volume was599.5 � 34.9 ml. To maintain consistency, participants were asked toconsume the entire drink within 20 min. The sides of the containerwere scraped and that small remaining quantity also consumed.
Urine collection. Upon completion of exercise, subjects werewheeled to the restroom to void their bladder. This first urinary voidwas discarded, such that UUN would be from the postexercise period,and all subsequent voids were collected for the final analysis ofnitrogen excretion and creatinine concentration. All urine collectionsfrom a trial over the 400-min postprandial period were pooled. At the
end of the collection period, the urine volume was assessed and analiquot saved for subsequent analysis.
Calculation of energy substrate oxidation. The last 5 min of each10-min breath measurement were used from pulmonary gas exchangeto calculate rates of EE, CHO oxidation (CHOox), and Lox (17). Inthe substrate oxidation calculations, we used nonprotein respiratory-quotient values for each time point that we derived from the respira-tory exchange ratio (RER) and timed UUN excretion, assuming thatthe percentage of the resting metabolic rate fueled by protein wasconsistent across the entire postmeal period. We assumed that eachgram of UUN represents oxidation of 6.25 g protein (17). EE, CHOox,and Lox were calculated as kcal/min or kcal/kg FFM/min, and resultsfrom statistical analyses were similar between these two manners ofexpressing the data. In addition to data expressed at each individualtime point, postexercise averages (and those in the C trial at thecorresponding time of day) were calculated as area under the curve,multiplied by duration (i.e., time-weighted average), and incrementalconcentrations were calculated as time-weighted average minus pre-exercise baseline values. Incremental concentration changes werecalculated in the same manner for other variables, such as plasmalipids.
Laboratory analyses. Glucose concentration was analyzed enzy-matically (Sigma-Aldrich, St. Louis, MO) in plasma. Urine wasanalyzed for UUN content using an enzymatic urea nitrogen kit(Stanbio Laboratory, Boerne, TX). Timed UUN output was calculatedas (nitrogen content � urine volume)/time interval, reported as g/min,and was used to correct substrate oxidation for protein oxidation.Creatinine concentration in urine was analyzed using a calorimetricassay (Oxford Biomedical Research, Oxford, MI). With the assump-tion that creatinine excretion is relatively constant throughout the day,the UUN results were normalized to urinary creatinine concentrationto derive a proxy index of protein oxidation.
Isolation of lipids. Following addition of known amounts of tri-heptadecanoin and heptadecanoic acid as internal standards, 0.5 mlplasma for TG and FFA analysis was extracted with 4 ml 30:70heptane/isopropanol (v/v) and subsequently mixed with 2 ml 0.003 Msulfuric acid. The organic layer was removed and then dried undernitrogen gas, and the TG and FFA were isolated by TLC, as describedpreviously (26, 73). Standard lanes in TLC contained oleic acid andtriolein and were visualized with iodine. The TG and FFA spots werescraped and extracted and dried under nitrogen gas. FA were resus-pended in 90% acetonitrile with 0.5 mM ammonium acetate. TGsunderwent saponification using a 0.25 M KOH in 90% ethanolsolution and heated for 45 min at 80°C. Next, to extract the releasedFA, we added 0.6 ml 1 M HCl and 4 ml hexane, mixed thoroughly,and then centrifuged to separate phases and subsequently transferredthe top layer (hexane). The sample was then dried under nitrogen gas,and the remaining lipid was resuspended in 90% acetonitrile with 0.5mM ammonium acetate. Samples were then analyzed by LC/MS(Varian, Walnut Creek, CA). For analysis of the liquid test meal, 10�l of the test-meal shake was combined with a known amount ofheptadecanoic acid (internal standard). The mixture underwent sapon-ification, as described above, and the organic phase of the subsequenthexane extraction was dried under nitrogen gas. Samples were thenresuspended in 90% acetonitrile with 0.5 mM ammonium acetate andanalyzed by LC/MS by the same method as used for plasma samples.
Mass spectrometry. With minor modification, we used the LC/MSmethod of Persson et al. (60). With the use of an Agilent 1200 HPLCsystem, Ascentis C18 2.1 � 150 mm column (Sigma-Aldrich), andVarian 1200L quadrupole mass spectrometer with electrospray ion-ization, FA concentrations and palmitate IE were determined. Mobilephase A (mp-A) was 80% acetonitrile with 0.5 mM ammoniumacetate, and mp-B was 100% acetonitrile with 0.5 mM ammoniumacetate. The flow rate was 0.4 ml/min, and FA were eluted isocrati-cally with 45% mp-A and 55% mp-B, followed by a column wash athigher organic strength. FA were identified by retention time andmass-to-charge ratio (m/z). The following ions were selectively mon-
1745Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from

itored in negative mode: myristate (m/z 227), palmitoleate (m/z 253),palmitate (m/z 255), heptadecanoate (m/z 269), [U-13C]palmitate (m/z271), -linolenate (m/z 277), linoleate (m/z 279), oleate (m/z 281),stearate (m/z 283), and arachidonate (m/z 303). Selected ion abun-dances were compared against external standard curves for calculationof concentration and IE. Aliquots of breath samples were sent toMetabolic Solutions (Nashua, NH) for IRMS measurements of 13CO2.
Calculations for tracer data. For each individual time point,percent contribution of dietary fat to plasma TG and FFA wascalculated as follows: IEplasma/IEmeal � 100, where IEplasma representsIE of palmitate in either plasma TG or FFA, and IEmeal represents IEof meal palmitate. Absolute contents of exogenous (dietary) andendogenous TG and FFA were calculated from this derived percent-age and the total concentration of TG and FFA in plasma. In additionto calculations for each individual time point, average trial values andincremental change of this average value were calculated as describedabove for energy substrate oxidation. The exogenously derived fattyacyl chains would be from the standardized liquid meal and wouldpresumably first label chylomicrons but would also extensively labelother pools in circulation. The endogenously derived fatty acyl chainswould be from lipids residing within the body before meal ingestion,including adipose TG, circulating FFA, VLDL-TG, and other lipiddepots. During the postprandial period, each specific pool could becomprised of both exogenous and endogenous FA. IEs were correctedfor premeal baseline IE in each trial. The 13CO2 excretion rates werecorrected using a bicarbonate retention factor (27). Bicarbonate re-tention returns to baseline soon after exercise (27) and is not alteredby the level of adiposity (33). Oxidation of meal fat (exogenous Lox)was calculated as
Exogenous Lox � [VCO2 � (IECO2/100) � (meal fat dose)]/[(tracer dose) � 16 � k]where IECO2 is the IE (units of MPE) of breath 13CO2 corrected forbackground, the “meal fat dose” is the total fat content of the testmeal, tracer dose is the molar quantity of [U-13C]palmitate in themeal, the factor of 16 extrapolates palmitate tracer dose to the totaldose of 13carbon, and k is the bicarbonate correction factor (0.83).
Statistical analyses. Data are presented as mean � SE. Two-waycomparisons among trials and across time points were made byANOVA with repeated measures (RM-ANOVA) with post hoc com-parisons using Fisher’s protected least significant difference test.Additionally, one-way RM-ANOVA was used to compare the averagevalues among trials. For incremental concentration change compari-son, analysis of covariance was used with baseline concentration asthe covariate. Statistical analyses were performed using JMP 10.0software (SAS Institute, Cary, NC). Statistical significance was set at � 0.05.
RESULTS
Characteristics of study participants and exercise bouts.Subject characteristics can be found in Table 1. All subjectswere obese (BMI, 30.1–58.0). E was time matched with R and
entailed 60 min of walking at an intensity of 62.5 � 1.4%VO2peak, with a gross EEE of 430.8 � 22.4 kcal. In the R trial,all three sets were completed by each study participant forevery exercise at an intensity of 88.9 � 2.3% of 10 RM, withan average number of 9.9 � 0.03 completed repetitions/set.Habitual exercise participation was 0.5 � 0.1 h/wk, indicatingsedentary lifestyle status.
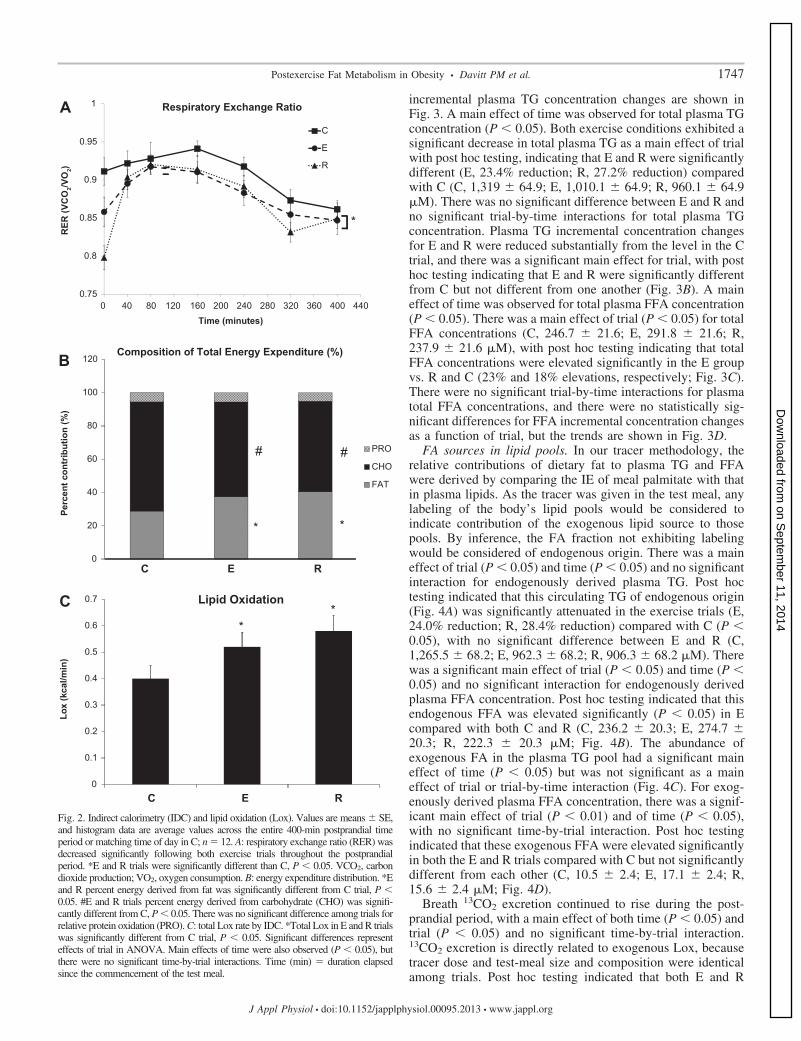
Metabolic rate and substrate partitioning. Average postmealvalues are reported. A main effect of time was detected for allmetabolic rate and substrate partitioning values (P � 0.05);however, there were no significant trial-by-time interactions.There was a significant main effect of trial for VO2 (P � 0.05),with post hoc testing indicating the R trial VO2 was elevatedsignificantly above C but not statistically different from E(Table 2). There was no significant main effect of trial forVCO2. In the postexercise recovery period, there was a maineffect of trial (P � 0.05) for RER with post hoc testingindicating significantly decreased RER in E and R comparedwith C (Table 2 and Fig. 2A). RER value contributions towardenergy substrate partition are addressed below. EE was notsignificantly different among trials when averaged throughoutthe postexercise time period. There was a significant maineffect of trial (P � 0.05) for percentage of energy from fat andfrom CHO. E and R groups derived a significantly higherpercentage of energy from fat compared with C (P � 0.05),with no significant difference between E and R (C, 28.76 �3.0; E, 37.39 � 3.0; R, 40.39 � 3.0%; Fig. 2B). E and R groupsderived a significantly lower percentage of energy from CHOcompared with C (P � 0.05), and there was no significantdifference between E and R (C, 65.65 � 2.1; E, 56.99 � 2.1;R, 54.52 � 2.1%; Fig. 2B). There were no significant maineffects of trial for energy derived from protein based on totalUUN (C, 1,241 � 333; E, 1,304 � 375; R, 1,187 � 407 mg)or urea-to-creatinine ratio in urine (C, 19.2 � 4.5; E, 22.5 �5.4; R, 17.3 � 5.6). When considered across time during eachtrial, there was a significant main effect of trial for absoluteLox (P � 0.001); post hoc testing indicated significantlyincreased Lox (Fig. 2C) in E and R compared with C, with nodifference between E and R (C, 0.40 � 0.05; E, 0.52 � 0.05;R, 0.58 � 0.05 kcal/min).
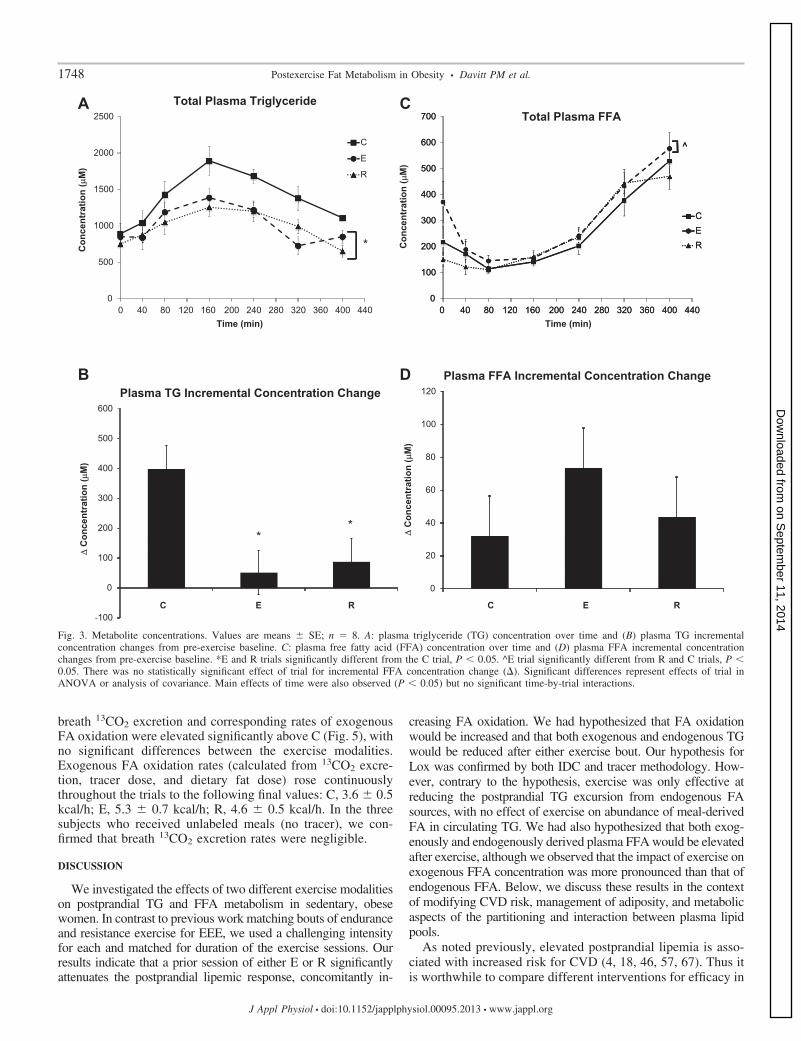
Metabolite concentrations. The pre-exercise values for allblood measurements were not statistically different amongtrials, and this time point was used for normalizing incrementalconcentration changes across trials. A main effect of time (P �0.05) was observed for plasma glucose concentration but withno main effect of trial or trial-by-time interactions (averagepostexercise values: C, 105.7 � 5.5; E, 106.4 � 5.7; R,102.6 � 5.7 mg/dl; P � 0.05). The total plasma TG and
Table 1. Characteristics of study participants
Age, yr 23.8 � 1.6Height, cm 162.6 � 2.3Weight, kg 98.9 � 6.8BMI, kg/m2 37.3 � 2.3Body fat, % 45.4 � 1.2FFM, kg 53.6 � 3.1Fat mass, kg 45.3 � 4.0VO2peak, l/min 2.4 � 0.1VO2peak, ml · kg�1 · min�1 25.2 � 1.4VO2peak, ml · kg FFM�1 · min�1 46.0 � 2.3
Values are means � SE; n � 12. BMI, body mass index; FFM, fat-freemass; VO2peak, peak oxygen consumption.
Table 2. Metabolic rate
C E R
VO2, l/min 0.26 � 0.01 0.27 � 0.01 0.28 � 0.01*VCO2, l/min 0.24 � 0.01 0.24 � 0.01 0.24 � 0.01RER 0.91 � 0.01 0.88 � 0.01* 0.87 � 0.01*EE, kcal/min 1.37 � 0.5 1.40 � 0.5 1.43 � 0.5
Values are means � SE; n � 12. Postexercise metabolic rate, average of 400min. C, no exercise (control) trial; E, endurance exercise trial; R, resistanceexercise trial; VO2, oxygen consumption; VCO2, carbon dioxide production;RER, respiratory exchange ratio; EE, energy expenditure. *Significantly dif-ferent from C trial in ANOVA, P � 0.05.
1746 Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from
incremental plasma TG concentration changes are shown inFig. 3. A main effect of time was observed for total plasma TGconcentration (P � 0.05). Both exercise conditions exhibited asignificant decrease in total plasma TG as a main effect of trialwith post hoc testing, indicating that E and R were significantlydifferent (E, 23.4% reduction; R, 27.2% reduction) comparedwith C (C, 1,319 � 64.9; E, 1,010.1 � 64.9; R, 960.1 � 64.9�M). There was no significant difference between E and R andno significant trial-by-time interactions for total plasma TGconcentration. Plasma TG incremental concentration changesfor E and R were reduced substantially from the level in the Ctrial, and there was a significant main effect for trial, with posthoc testing indicating that E and R were significantly differentfrom C but not different from one another (Fig. 3B). A maineffect of time was observed for total plasma FFA concentration(P � 0.05). There was a main effect of trial (P � 0.05) for totalFFA concentrations (C, 246.7 � 21.6; E, 291.8 � 21.6; R,237.9 � 21.6 �M), with post hoc testing indicating that totalFFA concentrations were elevated significantly in the E groupvs. R and C (23% and 18% elevations, respectively; Fig. 3C).There were no significant trial-by-time interactions for plasmatotal FFA concentrations, and there were no statistically sig-nificant differences for FFA incremental concentration changesas a function of trial, but the trends are shown in Fig. 3D.
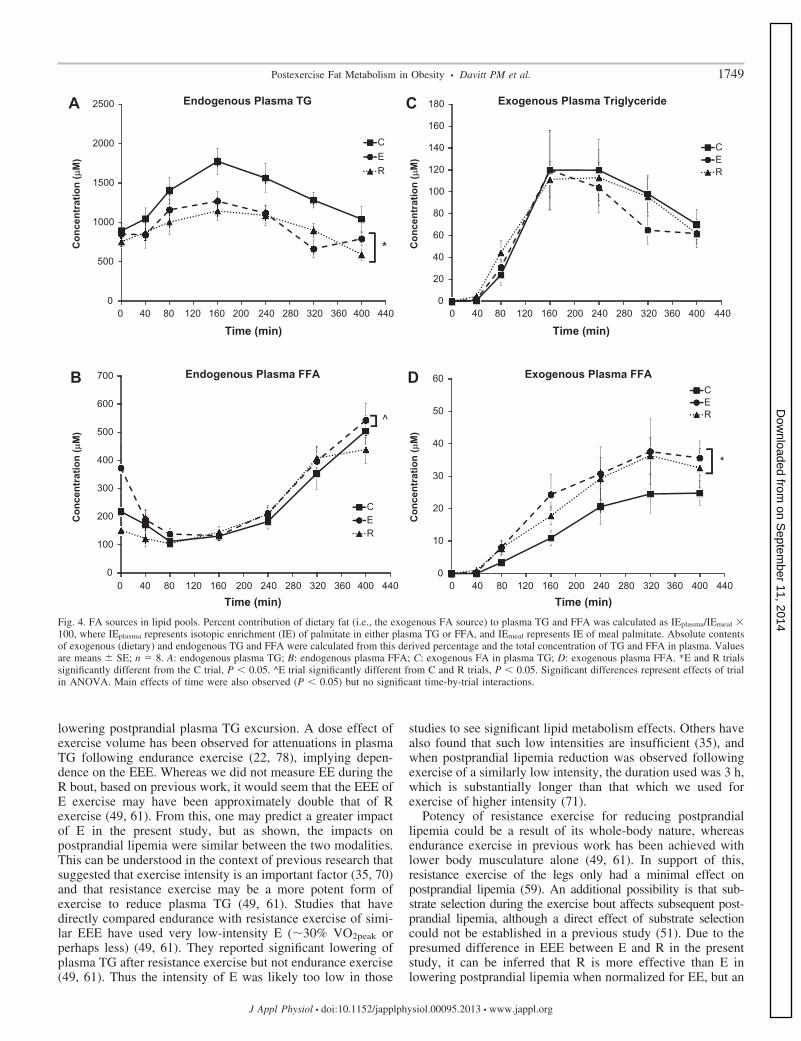
FA sources in lipid pools. In our tracer methodology, therelative contributions of dietary fat to plasma TG and FFAwere derived by comparing the IE of meal palmitate with thatin plasma lipids. As the tracer was given in the test meal, anylabeling of the body’s lipid pools would be considered toindicate contribution of the exogenous lipid source to thosepools. By inference, the FA fraction not exhibiting labelingwould be considered of endogenous origin. There was a maineffect of trial (P � 0.05) and time (P � 0.05) and no significantinteraction for endogenously derived plasma TG. Post hoctesting indicated that this circulating TG of endogenous origin(Fig. 4A) was significantly attenuated in the exercise trials (E,24.0% reduction; R, 28.4% reduction) compared with C (P �0.05), with no significant difference between E and R (C,1,265.5 � 68.2; E, 962.3 � 68.2; R, 906.3 � 68.2 �M). Therewas a significant main effect of trial (P � 0.05) and time (P �0.05) and no significant interaction for endogenously derivedplasma FFA concentration. Post hoc testing indicated that thisendogenous FFA was elevated significantly (P � 0.05) in Ecompared with both C and R (C, 236.2 � 20.3; E, 274.7 �20.3; R, 222.3 � 20.3 �M; Fig. 4B). The abundance ofexogenous FA in the plasma TG pool had a significant maineffect of time (P � 0.05) but was not significant as a maineffect of trial or trial-by-time interaction (Fig. 4C). For exog-enously derived plasma FFA concentration, there was a signif-icant main effect of trial (P � 0.01) and of time (P � 0.05),with no significant time-by-trial interaction. Post hoc testingindicated that these exogenous FFA were elevated significantlyin both the E and R trials compared with C but not significantlydifferent from each other (C, 10.5 � 2.4; E, 17.1 � 2.4; R,15.6 � 2.4 �M; Fig. 4D).
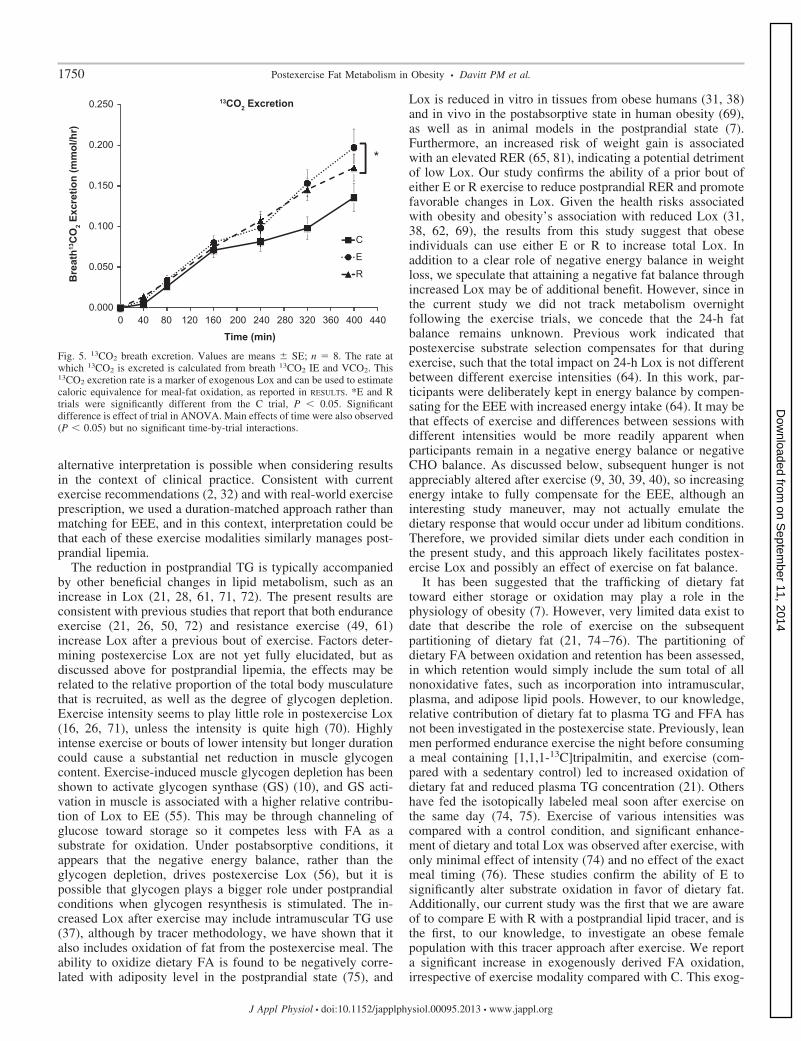
Breath 13CO2 excretion continued to rise during the post-prandial period, with a main effect of both time (P � 0.05) andtrial (P � 0.05) and no significant time-by-trial interaction.13CO2 excretion is directly related to exogenous Lox, becausetracer dose and test-meal size and composition were identicalamong trials. Post hoc testing indicated that both E and R
0.75
0.8
0.85
0.9
0.95
A
B
C
1
RER
(VC
O2/V
O2)
Time (minutes)
Respiratory Exchange Ratio
C
E
R
*
0
20
40
60
80
100
120
Perc
ent c
ontr
ibut
ion
(%)
Composition of Total Energy Expenditure (%)
PRO
CHO
FAT
*
# #
*
C E R
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
C E R
Lox
(kca
l/min
)
Lipid Oxidation
**
Fig. 2. Indirect calorimetry (IDC) and lipid oxidation (Lox). Values are means � SE,and histogram data are average values across the entire 400-min postprandial timeperiod or matching time of day in C; n � 12. A: respiratory exchange ratio (RER) wasdecreased significantly following both exercise trials throughout the postprandialperiod. *E and R trials were significantly different than C, P � 0.05. VCO2, carbondioxide production; VO2, oxygen consumption. B: energy expenditure distribution. *Eand R percent energy derived from fat was significantly different from C trial, P �0.05. #E and R trials percent energy derived from carbohydrate (CHO) was signifi-cantly different from C, P � 0.05. There was no significant difference among trials forrelative protein oxidation (PRO). C: total Lox rate by IDC. *Total Lox in E and R trialswas significantly different from C trial, P � 0.05. Significant differences representeffects of trial in ANOVA. Main effects of time were also observed (P � 0.05), butthere were no significant time-by-trial interactions. Time (min) � duration elapsedsince the commencement of the test meal.
1747Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from
breath 13CO2 excretion and corresponding rates of exogenousFA oxidation were elevated significantly above C (Fig. 5), withno significant differences between the exercise modalities.Exogenous FA oxidation rates (calculated from 13CO2 excre-tion, tracer dose, and dietary fat dose) rose continuouslythroughout the trials to the following final values: C, 3.6 � 0.5kcal/h; E, 5.3 � 0.7 kcal/h; R, 4.6 � 0.5 kcal/h. In the threesubjects who received unlabeled meals (no tracer), we con-firmed that breath 13CO2 excretion rates were negligible.
DISCUSSION
We investigated the effects of two different exercise modalitieson postprandial TG and FFA metabolism in sedentary, obesewomen. In contrast to previous work matching bouts of enduranceand resistance exercise for EEE, we used a challenging intensityfor each and matched for duration of the exercise sessions. Ourresults indicate that a prior session of either E or R significantlyattenuates the postprandial lipemic response, concomitantly in-
creasing FA oxidation. We had hypothesized that FA oxidationwould be increased and that both exogenous and endogenous TGwould be reduced after either exercise bout. Our hypothesis forLox was confirmed by both IDC and tracer methodology. How-ever, contrary to the hypothesis, exercise was only effective atreducing the postprandial TG excursion from endogenous FAsources, with no effect of exercise on abundance of meal-derivedFA in circulating TG. We had also hypothesized that both exog-enously and endogenously derived plasma FFA would be elevatedafter exercise, although we observed that the impact of exercise onexogenous FFA concentration was more pronounced than that ofendogenous FFA. Below, we discuss these results in the contextof modifying CVD risk, management of adiposity, and metabolicaspects of the partitioning and interaction between plasma lipidpools.
As noted previously, elevated postprandial lipemia is asso-ciated with increased risk for CVD (4, 18, 46, 57, 67). Thus itis worthwhile to compare different interventions for efficacy in
0
500
1000
1500
2000
2500
Time (min)
Total Plasma TriglycerideA C
B D
C
E
R
*
-100
0
100
200
300
400
500
600Plasma TG Incremental Concentration Change
**
0
100
200
300
400
500
600
700
CER
^
0
100
200
300
400
500
600
700
Con
cent
ratio
n (µ
M)
Time (min)
Total Plasma FFA
CER
^
0
20
40
60
80
100
120Plasma FFA Incremental Concentration Change
Con
cent
ratio
n (µ
M)
∆ C
once
ntra
tion
(µM
)
∆ C
once
ntra
tion
(µM
)
Fig. 3. Metabolite concentrations. Values are means � SE; n � 8. A: plasma triglyceride (TG) concentration over time and (B) plasma TG incrementalconcentration changes from pre-exercise baseline. C: plasma free fatty acid (FFA) concentration over time and (D) plasma FFA incremental concentrationchanges from pre-exercise baseline. *E and R trials significantly different from the C trial, P � 0.05. ^E trial significantly different from R and C trials, P �0.05. There was no statistically significant effect of trial for incremental FFA concentration change (). Significant differences represent effects of trial inANOVA or analysis of covariance. Main effects of time were also observed (P � 0.05) but no significant time-by-trial interactions.
1748 Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from
lowering postprandial plasma TG excursion. A dose effect ofexercise volume has been observed for attenuations in plasmaTG following endurance exercise (22, 78), implying depen-dence on the EEE. Whereas we did not measure EE during theR bout, based on previous work, it would seem that the EEE ofE exercise may have been approximately double that of Rexercise (49, 61). From this, one may predict a greater impactof E in the present study, but as shown, the impacts onpostprandial lipemia were similar between the two modalities.This can be understood in the context of previous research thatsuggested that exercise intensity is an important factor (35, 70)and that resistance exercise may be a more potent form ofexercise to reduce plasma TG (49, 61). Studies that havedirectly compared endurance with resistance exercise of simi-lar EEE have used very low-intensity E (�30% VO2peak orperhaps less) (49, 61). They reported significant lowering ofplasma TG after resistance exercise but not endurance exercise(49, 61). Thus the intensity of E was likely too low in those
studies to see significant lipid metabolism effects. Others havealso found that such low intensities are insufficient (35), andwhen postprandial lipemia reduction was observed followingexercise of a similarly low intensity, the duration used was 3 h,which is substantially longer than that which we used forexercise of higher intensity (71).
Potency of resistance exercise for reducing postprandiallipemia could be a result of its whole-body nature, whereasendurance exercise in previous work has been achieved withlower body musculature alone (49, 61). In support of this,resistance exercise of the legs only had a minimal effect onpostprandial lipemia (59). An additional possibility is that sub-strate selection during the exercise bout affects subsequent post-prandial lipemia, although a direct effect of substrate selectioncould not be established in a previous study (51). Due to thepresumed difference in EEE between E and R in the presentstudy, it can be inferred that R is more effective than E inlowering postprandial lipemia when normalized for EE, but an
0
500
1000
1500
2000
2500
Time (min)
Endogenous Plasma TGA C
CER
*
0
100
200
300
400
500
600
700
Time (min)
Endogenous Plasma FFA
CER
^
0
20
40
60
80
100
120
140
160
180
Time (min)
Exogenous Plasma Triglyceride
CER
0
10
20
30
40
50
60
Time (min)
Exogenous Plasma FFACER
*
Con
cent
ratio
n (µ
M)
Con
cent
ratio
n (µ
M)
Con
cent
ratio
n (µ
M)
Con
cent
ratio
n (µ
M)
B D
Fig. 4. FA sources in lipid pools. Percent contribution of dietary fat (i.e., the exogenous FA source) to plasma TG and FFA was calculated as IEplasma/IEmeal �100, where IEplasma represents isotopic enrichment (IE) of palmitate in either plasma TG or FFA, and IEmeal represents IE of meal palmitate. Absolute contentsof exogenous (dietary) and endogenous TG and FFA were calculated from this derived percentage and the total concentration of TG and FFA in plasma. Valuesare means � SE; n � 8. A: endogenous plasma TG; B: endogenous plasma FFA; C: exogenous FA in plasma TG; D: exogenous plasma FFA. *E and R trialssignificantly different from the C trial, P � 0.05. ^E trial significantly different from C and R trials, P � 0.05. Significant differences represent effects of trialin ANOVA. Main effects of time were also observed (P � 0.05) but no significant time-by-trial interactions.
1749Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from
alternative interpretation is possible when considering resultsin the context of clinical practice. Consistent with currentexercise recommendations (2, 32) and with real-world exerciseprescription, we used a duration-matched approach rather thanmatching for EEE, and in this context, interpretation could bethat each of these exercise modalities similarly manages post-prandial lipemia.
The reduction in postprandial TG is typically accompaniedby other beneficial changes in lipid metabolism, such as anincrease in Lox (21, 28, 61, 71, 72). The present results areconsistent with previous studies that report that both enduranceexercise (21, 26, 50, 72) and resistance exercise (49, 61)increase Lox after a previous bout of exercise. Factors deter-mining postexercise Lox are not yet fully elucidated, but asdiscussed above for postprandial lipemia, the effects may berelated to the relative proportion of the total body musculaturethat is recruited, as well as the degree of glycogen depletion.Exercise intensity seems to play little role in postexercise Lox(16, 26, 71), unless the intensity is quite high (70). Highlyintense exercise or bouts of lower intensity but longer durationcould cause a substantial net reduction in muscle glycogencontent. Exercise-induced muscle glycogen depletion has beenshown to activate glycogen synthase (GS) (10), and GS acti-vation in muscle is associated with a higher relative contribu-tion of Lox to EE (55). This may be through channeling ofglucose toward storage so it competes less with FA as asubstrate for oxidation. Under postabsorptive conditions, itappears that the negative energy balance, rather than theglycogen depletion, drives postexercise Lox (56), but it ispossible that glycogen plays a bigger role under postprandialconditions when glycogen resynthesis is stimulated. The in-creased Lox after exercise may include intramuscular TG use(37), although by tracer methodology, we have shown that italso includes oxidation of fat from the postexercise meal. Theability to oxidize dietary FA is found to be negatively corre-lated with adiposity level in the postprandial state (75), and
Lox is reduced in vitro in tissues from obese humans (31, 38)and in vivo in the postabsorptive state in human obesity (69),as well as in animal models in the postprandial state (7).Furthermore, an increased risk of weight gain is associatedwith an elevated RER (65, 81), indicating a potential detrimentof low Lox. Our study confirms the ability of a prior bout ofeither E or R exercise to reduce postprandial RER and promotefavorable changes in Lox. Given the health risks associatedwith obesity and obesity’s association with reduced Lox (31,38, 62, 69), the results from this study suggest that obeseindividuals can use either E or R to increase total Lox. Inaddition to a clear role of negative energy balance in weightloss, we speculate that attaining a negative fat balance throughincreased Lox may be of additional benefit. However, since inthe current study we did not track metabolism overnightfollowing the exercise trials, we concede that the 24-h fatbalance remains unknown. Previous work indicated thatpostexercise substrate selection compensates for that duringexercise, such that the total impact on 24-h Lox is not differentbetween different exercise intensities (64). In this work, par-ticipants were deliberately kept in energy balance by compen-sating for the EEE with increased energy intake (64). It may bethat effects of exercise and differences between sessions withdifferent intensities would be more readily apparent whenparticipants remain in a negative energy balance or negativeCHO balance. As discussed below, subsequent hunger is notappreciably altered after exercise (9, 30, 39, 40), so increasingenergy intake to fully compensate for the EEE, although aninteresting study maneuver, may not actually emulate thedietary response that would occur under ad libitum conditions.Therefore, we provided similar diets under each condition inthe present study, and this approach likely facilitates postex-ercise Lox and possibly an effect of exercise on fat balance.
It has been suggested that the trafficking of dietary fattoward either storage or oxidation may play a role in thephysiology of obesity (7). However, very limited data exist todate that describe the role of exercise on the subsequentpartitioning of dietary fat (21, 74–76). The partitioning ofdietary FA between oxidation and retention has been assessed,in which retention would simply include the sum total of allnonoxidative fates, such as incorporation into intramuscular,plasma, and adipose lipid pools. However, to our knowledge,relative contribution of dietary fat to plasma TG and FFA hasnot been investigated in the postexercise state. Previously, leanmen performed endurance exercise the night before consuminga meal containing [1,1,1-13C]tripalmitin, and exercise (com-pared with a sedentary control) led to increased oxidation ofdietary fat and reduced plasma TG concentration (21). Othershave fed the isotopically labeled meal soon after exercise onthe same day (74, 75). Exercise of various intensities wascompared with a control condition, and significant enhance-ment of dietary and total Lox was observed after exercise, withonly minimal effect of intensity (74) and no effect of the exactmeal timing (76). These studies confirm the ability of E tosignificantly alter substrate oxidation in favor of dietary fat.Additionally, our current study was the first that we are awareof to compare E with R with a postprandial lipid tracer, and isthe first, to our knowledge, to investigate an obese femalepopulation with this tracer approach after exercise. We reporta significant increase in exogenously derived FA oxidation,irrespective of exercise modality compared with C. This exog-
*
0.000
0.050
0.100
0.150
0.200
0.250
Time (min)
C
E
RBre
ath13
CO
2 Exc
retio
n (m
mol
/hr)
13CO2 Excretion
Fig. 5. 13CO2 breath excretion. Values are means � SE; n � 8. The rate atwhich 13CO2 is excreted is calculated from breath 13CO2 IE and VCO2. This13CO2 excretion rate is a marker of exogenous Lox and can be used to estimatecaloric equivalence for meal-fat oxidation, as reported in RESULTS. *E and Rtrials were significantly different from the C trial, P � 0.05. Significantdifference is effect of trial in ANOVA. Main effects of time were also observed(P � 0.05) but no significant time-by-trial interactions.
1750 Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from

enous FA oxidation after E confirms previous literature butexpands the finding to a new population and adds recognitionof R as an alternate means of deriving a similar outcome.Whereas other studies have not measured exogenous FA oxi-dation after R specifically, they have reported increased totalLox by IDC after R (49, 61), and our tracer findings show thatexogenous fat contributes to this increase in Lox after exercise.
A prior exercise bout can reduce both VLDL and chylomi-cron TG concentrations (20, 21), but still, without assessingincorporation of a dietary FA tracer into the unfractionatedplasma TG pool, it is not yet definitively known if the reduc-tion in postprandial lipemia was due to a reduction in bothexogenously and endogenously derived FA in plasma TG. Inthe current study, we differentiated between exogenous andendogenous sources of plasma TG and FFA, and we discov-ered that the significant attenuation in total plasma TG camefrom endogenous TG only, not from meal-derived TG (Fig. 4).This could be a result of a reduction in hepatic VLDL-TGsecretion, enhanced VLDL-TG clearance without enhancedchylomicron clearance, or increased relative contribution ofrecycled dietary FA from chylomicrons into VLDL. In supportof this speculation, recently, it was reported that women exhibitreduced VLDL-TG secretion after a bout of endurance exercisein the postabsorptive state (6); it is possible that this physio-logical impact is also present in the postprandial state.
Rather than focus on specific lipoprotein particles per se, ourquestions were with regard to the role of the meal fat, and sowe assessed whether the changes in plasma TG and FFAconcentrations were because of altered abundance of the in-gested fat in circulation. The inter-relationships between lipidpools dynamically change over time during meal absorption, someasuring lipid concentrations cannot sufficiently answerquestions about the sources (dietary vs. endogenous). Wheninvestigators are interested in addressing contributions of ex-ogenous and endogenous lipid to circulating TG, there may bea tendency to conceptualize the FA sources for TG content oflipid pools as solely exogenous for chylomicrons and solelyendogenous for VLDL, but this is not entirely accurate. Fur-thermore, when plasma TG rises after a meal, it may beassumed by some scientists to be primarily the result of risingchylomicron levels, but this is also not accurate. During thecourse of absorbing an isotopically labeled meal, the IE ofchylomicron TG changes over time (44, 63), possibly indicat-ing impurity of the isolated fraction but also likely indicatingchanging relative contribution of meal fat and contribution offat previously residing in intestinal cells. After a meal, plasmaVLDL-TG can rise to a similar or even greater absolute extentthan chylomicron-TG (20, 21), and FA from a meal rapidlylabel the VLDL pool, in addition the chylomicron pool (24, 44,63), as chylomicron remnants, and dietary-derived plasma FFAcan deliver FA to VLDL (5, 24, 25). As we were interested inthe relative role of the actual meal fat in the TG and FFAexcursions, we derived an approach and a simple calculation tocompare the meal IE with IE of plasma TG and FFA, and thisdirectly assessed the contribution of the meal to the plasma TGexcursion and generated surprising and novel data about post-prandial lipid metabolism after exercise.
Similar to previous research showing an elevation in totalFFA concentration postexercise using E (21, 26, 50, 72), wehave seen similar total FFA concentration increases after E butno change in total plasma FFA after R. Further analysis
revealed a significant rise in exogenous FFA in both exercisegroups but a significant rise in endogenous plasma FFA con-centration after E only (Fig. 4). In the E trial, the significantrise in endogenous FFA concentration was likely a result oflipolysis and mobilization of FFA during the prior E bout,which is related to hormonal changes during exercise, such asincreased catecholamines, growth hormone, and cortisol (26);removal of the most immediate postexercise time point elimi-nated the statistical significance for elevation of FFA concen-tration across the recovery period. The rise in plasma FFA ofexogenous origin after either exercise bout indicates a poten-tially enhanced spillover of dietary FA, as exogenous FFA inplasma have been considered a surrogate marker for systemicdietary spillover (53, 54, 58). This process of spillover de-scribes FA released by intravascular lipase activities (lipopro-tein lipase and/or hepatic lipase) that were not channeledefficiently to direct uptake at the site of lipolysis and are thusreleased into venous circulation (53). In the process of spill-over, FFA become systemically available and can ultimately beused by tissues after recirculation through the arterial vascula-ture. Although we consider the increased exogenously derivedFFA concentration to be spillover, without arterial-venoussampling across tissues, this mechanism cannot be confirmedwith absolute certainty; we note that an alternative explanationfor this observation could be that dietary FA were brieflyesterified intracellularly and then reliberated by lipolysis intothe plasma FFA pool. Nonetheless, we observed the increasedapparent postexercise spillover alongside increased oxidationof exogenous FA. It is possible that this process promotedoxidation by making labeled FA available to specific tissuesthat exhibit high rates of FA oxidation, such as the liver orothers, and this increased FFA availability may also haveincreased nonoxidative disposal rates in specific tissues. An-other major contributor to postexercise Lox could be partition-ing of the endogenous lipid toward oxidation through changesin intracellular signaling related to glycogen concentration andcellular energy charge.
In each obese individual, positive fat balance has led to thestorage of excess fat over time (23). Unless dietary compen-sation were to occur, enhancing the ability to oxidize lipidcould help rectify this situation and could lead to a negative fatbalance, which can ultimately lead to a reduction in body fat(23). Elevations in fat oxidation have been demonstrated in thecurrent study after both E and R, with no significant differencesbetween the exercise conditions. The increased Lox in con-junction with reductions in postprandial lipemia has implica-tions for management of adiposity and of CVD risk. A criticalquestion to ask is whether it would have been more appropriateto increase the size of the postexercise meals in the E and Rtrials compared with the C trial in the present study to preventthe negative energy balance resulting from exercise. Our goalwas to mimic nonlaboratory conditions, and the commonfinding has been that an acute bout of exercise does notincrease subsequent hunger or ad libitum energy intakethroughout the rest of the day (8, 9, 11, 30, 39–43, 48). Todescribe this phenomenon, previously, the association betweenEE and energy intake has been described as being only a loosecoupling (8). For this reason, we provided the same caloriccontent in the test meal for each condition rather than deliber-ately encouraging dietary compensation. As we focused onfemale subjects, we cannot comment on comparisons with an
1751Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from

obese male population, and such a comparison could be animportant future direction to be explored. Our results provideobese women with more than one means of increasing Lox andattenuating the postprandial lipemic response.
In conclusion, we have demonstrated that a prior bout ofeither 60 min of E or R attenuated postprandial lipemia byreducing the circulating abundance of endogenously derivedplasma TG and that prior exercise also increased both total FAand exogenous FA oxidation in obese women. Our findingsindicate that when matched for duration, R provides an at leastequivalent effect on postprandial lipemia and fat oxidation as Ein obese women. These results may be distinct from those thatwould have been obtained if bouts were matched for EE. Theresults exemplify the complex relationship between lipid poolsand the ability of lifestyle changes to alter these relationships.Additional preclinical studies comparing potential exerciseprescriptions on postprandial lipemia and the trafficking ofdietary fat toward oxidation are needed in various populationswith both acute and chronic application of the exercise inter-ventions.
ACKNOWLEDGMENTS
We thank Julie Cameron Gervasi, Donna Collins, Donna Anzalone, andDiane Smith for clinical support. We thank Karla Schacher, Derek Gieschen,Mark Siehl, Altamash Rajaa, and Pat Sarcone for assistance in conductingisotope trials. We thank Sebastian Roque for assistance with isotope trials andfor laboratory support.
GRANTS
Support for this work was provided by the Charles and Johanna BuschBiomedical Foundation and by an instrumentation gift from Bay BioanalyticalLaboratory.
DISCLOSURES
The authors declare no conflicts of interest.
AUTHOR CONTRIBUTIONS
Author contributions: S.M.A. and G.C.H. conception and design of re-search; P.M.D., S.M.A., M.A.T., D.L.G., and G.C.H. performed experiments;P.M.D., M.A.T., and G.C.H. analyzed data; P.M.D., S.M.A., and G.C.H.interpreted results of experiments; P.M.D. prepared figures; P.M.D. draftedmanuscript; P.M.D., S.M.A., and G.C.H. edited and revised manuscript;P.M.D., S.M.A., M.A.T., D.L.G., and G.C.H. approved final version ofmanuscript.
REFERENCES
1. Aldred HE, Perry IC, Hardman AE. The effect of a single bout of briskwalking on postprandial lipemia in normolipidemic young adults. Metab-olism 43: 836–841, 1994.
2. American College of Sports Medicine, Thompson WR, Gordon NF,Pescatello LS. ACSM’s Guidelines for Exercise Testing and Prescription.Philadelphia: Lippincott Williams & Wilkins, 2010, p. xxi, 380.
3. Baechle TR, Earle RW, National Strength and Conditioning Associ-ation. Essentials of Strength Training and Conditioning. Champaign, IL:Human Kinetics, 2008, p. xiv, 641.
4. Bansal S, Buring JE, Rifai N, Mora S, Sacks FM, Ridker PM. Fastingcompared with nonfasting triglycerides and risk of cardiovascular eventsin women. JAMA 298: 309–316, 2007.
5. Barrows BR, Parks EJ. Contributions of different fatty acid sources tovery low-density lipoprotein-triacylglycerol in the fasted and fed states. JClin Endocrinol Metab 91: 1446–1452, 2006.
6. Bellou E, Siopi A, Galani M, Maraki M, Tsekouras YE, PanagiotakosDB, Kavouras SA, Magkos F, Sidossis LS. Acute effects of exercise andcalorie restriction on triglyceride metabolism in women. Med Sci SportsExerc 45: 455–461, 2013.
7. Bessesen DH, Rupp CL, Eckel RH. Dietary fat is shunted away fromoxidation, toward storage in obese Zucker rats. Obes Res 3: 179–189,1995.
8. Blundell JE, King NA. Physical activity and regulation of food intake:current evidence. Med Sci Sports Exerc 31: S573–S583, 1999.
9. Blundell JE, Stubbs RJ, Hughes DA, Whybrow S, King NA. Cross talkbetween physical activity and appetite control: does physical activitystimulate appetite? Proc Nutr Soc 62: 651–661, 2003.
10. Bogardus C, Thuillez P, Ravussin E, Vasquez B, Narimiga M, AzharS. Effect of muscle glycogen depletion on in vivo insulin action in man.J Clin Invest 72: 1605–1610, 1983.
11. Broom DR, Batterham RL, King JA, Stensel DJ. Influence of resistanceand aerobic exercise on hunger, circulating levels of acylated ghrelin, andpeptide YY in healthy males. Am J Physiol Regul Integr Comp Physiol296: R29–R35, 2009.
12. Casazza GA, Jacobs KA, Suh SH, Miller BF, Horning MA, BrooksGA. Menstrual cycle phase and oral contraceptive effects on triglyceridemobilization during exercise. J Appl Physiol 97: 302–309, 2004.
13. Dallongeville J, Gruson E, Dallinga-Thie G, Pigeyre M, Gomila S,Romon M. Effect of weight loss on the postprandial response to high-fatand high-carbohydrate meals in obese women. Eur J Clin Nutr 61:711–718, 2007.
14. DeLany JP, Windhauser MM, Champagne CM, Bray GA. Differentialoxidation of individual dietary fatty acids in humans. Am J Clin Nutr 72:905–911, 2000.
15. Dempster P, Aitkens S. A new air displacement method for the determi-nation of human body composition. Med Sci Sports Exerc 27: 1692–1697,1995.
16. Ferreira AP, Ferreira CB, Souza VC, Cordova CO, Silva GC, No-brega Ode T, Franca NM. The influence of intense intermittent versusmoderate continuous exercise on postprandial lipemia. Clinics (Sao Paulo)66: 535–541, 2011.
17. Frayn KN. Calculation of substrate oxidation rates in vivo from gaseousexchange. J Appl Physiol 55: 628–634, 1983.
18. Freiberg JJ, Tybjaerg-Hansen A, Jensen JS, Nordestgaard BG. Non-fasting triglycerides and risk of ischemic stroke in the general population.JAMA 300: 2142–2152, 2008.
19. Gill JM, Al-Mamari A, Ferrell WR, Cleland SJ, Packard CJ, SattarN, Petrie JR, Caslake MJ. Effects of prior moderate exercise on post-prandial metabolism and vascular function in lean and centrally obesemen. J Am Coll Cardiol 44: 2375–2382, 2004.
20. Gill JM, Al-Mamari A, Ferrell WR, Cleland SJ, Perry CG, Sattar N,Packard CJ, Caslake MJ, Petrie JR. Effect of prior moderate exerciseon postprandial metabolism in men with type 2 diabetes: heterogeneity ofresponses. Atherosclerosis 194: 134–143, 2007.
21. Gill JM, Frayn KN, Wootton SA, Miller GJ, Hardman AE. Effects ofprior moderate exercise on exogenous and endogenous lipid metabolismand plasma factor VII activity. Clin Sci (Lond) 100: 517–527, 2001.
22. Gill JM, Herd SL, Hardman AE. Moderate exercise and post-prandialmetabolism: issues of dose-response. J Sports Sci 20: 961–967, 2002.
23. Hansen K, Shriver T, Schoeller D. The effects of exercise on the storageand oxidation of dietary fat. Sports Med 35: 363–373, 2005.
24. Heath RB, Karpe F, Milne RW, Burdge GC, Wootton SA, Frayn KN.Dietary fatty acids make a rapid and substantial contribution to VLDL-triacylglycerol in the fed state. Am J Physiol Endocrinol Metab 292:E732–E739, 2007.
25. Heath RB, Karpe F, Milne RW, Burdge GC, Wootton SA, Frayn KN.Selective partitioning of dietary fatty acids into the VLDL TG pool in theearly postprandial period. J Lipid Res 44: 2065–2072, 2003.
26. Henderson GC, Fattor JA, Horning MA, Faghihnia N, Johnson ML,Mau TL, Luke-Zeitoun M, Brooks GA. Lipolysis and fatty acid metab-olism in men and women during the postexercise recovery period. JPhysiol 584: 963–981, 2007.
27. Henderson GC, Fattor JA, Horning MA, Faghihnia N, Luke-ZeitounM, Brooks GA. Retention of intravenously infused [13C]bicarbonate istransiently increased during recovery from hard exercise. J Appl Physiol103: 1604–1612, 2007.
28. Herd SL, Kiens B, Boobis LH, Hardman AE. Moderate exercise,postprandial lipemia, and skeletal muscle lipoprotein lipase activity. Me-tabolism 50: 756–762, 2001.
29. Horning MA, Friedlander AL, Casazza GA, Huie MJ, Brooks GA.Arterial and “arterialized” sampling sites do not change isotopic enrich-ment using [6,6-D-glucose] and [1,1,2,3,3-D-glycerol]. FASEB J 12: A854,1998.
1752 Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from

30. Hubert P, King NA, Blundell JE. Uncoupling the effects of energyexpenditure and energy intake: appetite response to short-term energydeficit induced by meal omission and physical activity. Appetite 31: 9–19,1998.
31. Hulver MW, Berggren JR, Cortright RN, Dudek RW, Thompson RP,Pories WJ, MacDonald KG, Cline GW, Shulman GI, Dohm GL,Houmard JA. Skeletal muscle lipid metabolism with obesity. Am JPhysiol Endocrinol Metab 284: E741–E747, 2003.
32. Institute of Medicine. Dietary Reference Intakes for Energy, Carbohy-drate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids.Washington, DC: The National Academies Press, 2002.
33. Issekutz B Jr, Paul P, Miller HI, Bortz WM. Oxidation of plasma FFAin lean and obese humans. Metabolism 17: 62–73, 1968.
34. James PT, Leach R, Kalamara E, Shayeghi M. The worldwide obesityepidemic. Obes Res 9 Suppl 4: 228S–233S, 2001.
35. Katsanos CS, Grandjean PW, Moffatt RJ. Effects of low and moderateexercise intensity on postprandial lipemia and postheparin plasma lipo-protein lipase activity in physically active men. J Appl Physiol 96:181–188, 2004.
36. Katsanos CS, Moffatt RJ. Acute effects of premeal versus postmealexercise on postprandial hypertriglyceridemia. Clin J Sport Med 14:33–39, 2004.
37. Kiens B, Richter EA. Utilization of skeletal muscle triacylglycerol duringpostexercise recovery in humans. Am J Physiol 275: E332–E337, 1998.
38. Kim JY, Hickner RC, Cortright RL, Dohm GL, Houmard JA. Lipidoxidation is reduced in obese human skeletal muscle. Am J PhysiolEndocrinol Metab 279: E1039–E1044, 2000.
39. King JA, Miyashita M, Wasse LK, Stensel DJ. Influence of prolongedtreadmill running on appetite, energy intake and circulating concentrationsof acylated ghrelin. Appetite 54: 492–498, 2010.
40. King JA, Wasse LK, Broom DR, Stensel DJ. Influence of brisk walkingon appetite, energy intake, and plasma acylated ghrelin. Med Sci SportsExerc 42: 485–492, 2010.
41. King NA, Burley VJ, Blundell JE. Exercise-induced suppression ofappetite: effects on food intake and implications for energy balance. EurJ Clin Nutr 48: 715–724, 1994.
42. King NA, Lluch A, Stubbs RJ, Blundell JE. High dose exercise does notincrease hunger or energy intake in free living males. Eur J Clin Nutr 51:478–483, 1997.
43. King NA, Snell L, Smith RD, Blundell JE. Effects of short-term exerciseon appetite responses in unrestrained females. Eur J Clin Nutr 50:663–667, 1996.
44. Knuth ND, Horowitz JF. The elevation of ingested lipids within plasmachylomicrons is prolonged in men compared with women. J Nutr 136:1498–1503, 2006.
45. Knuth ND, Remias DB, Horowitz JF. Adding carbohydrate to a high-fatmeal blunts postprandial lipemia in women and reduces meal-derived fattyacids in systemic circulation. Appl Physiol Nutr Metab 33: 315–325, 2008.
46. Langsted A, Freiberg JJ, Nordestgaard BG. Fasting and nonfastinglipid levels: influence of normal food intake on lipids, lipoproteins,apolipoproteins, and cardiovascular risk prediction. Circulation 118:2047–2056, 2008.
47. Lewis GF, O’Meara NM, Soltys PA, Blackman JD, Iverius PH,Druetzler AF, Getz GS, Polonsky KS. Postprandial lipoprotein metab-olism in normal and obese subjects: comparison after the vitamin Afat-loading test. J Clin Endocrinol Metab 71: 1041–1050, 1990.
48. Lluch A, King NA, Blundell JE. Exercise in dietary restrained women:no effect on energy intake but change in hedonic ratings. Eur J Clin Nutr52: 300–307, 1998.
49. Magkos F, Tsekouras YE, Prentzas KI, Basioukas KN, Matsama SG,Yanni AE, Kavouras SA, Sidossis LS. Acute exercise-induced changesin basal VLDL-triglyceride kinetics leading to hypotriglyceridemia man-ifest more readily after resistance than endurance exercise. J Appl Physiol105: 1228–1236, 2008.
50. Magkos F, Wright DC, Patterson BW, Mohammed BS, MittendorferB. Lipid metabolism response to a single, prolonged bout of enduranceexercise in healthy young men. Am J Physiol Endocrinol Metab 290:E355–E362, 2006.
51. Malkova D, Hardman AE, Bowness RJ, Macdonald IA. The reductionin postprandial lipemia after exercise is independent of the relativecontributions of fat and carbohydrate to energy metabolism during exer-cise. Metabolism 48: 245–251, 1999.
52. Maraki M, Magkos F, Christodoulou N, Aggelopoulou N, SkenderiKP, Panagiotakos D, Kavouras SA, Sidossis LS. One day of moderate
energy deficit reduces fasting and postprandial triacylglycerolemia inwomen: the role of calorie restriction and exercise. Clin Nutr 29: 459–463,2010.
53. Miles JM, Nelson RH. Contribution of triglyceride-rich lipoproteins toplasma free fatty acids. Horm Metab Res 39: 726–729, 2007.
54. Miles JM, Park YS, Walewicz D, Russell-Lopez C, Windsor S, IsleyWL, Coppack SW, Harris WS. Systemic and forearm triglyceridemetabolism: fate of lipoprotein lipase-generated glycerol and free fattyacids. Diabetes 53: 521–527, 2004.
55. Mott DM, Pratley RE, Bogardus C. Postabsorptive respiratory quotientand insulin-stimulated glucose storage rate in nondiabetic Pima Indians arerelated to glycogen synthase fractional activity in cultured myoblasts. JClin Invest 101: 2251–2256, 1998.
56. Newsom SA, Schenk S, Thomas KM, Harber MP, Knuth ND, Gold-enberg N, Horowitz JF. Energy deficit after exercise augments lipidmobilization but does not contribute to the exercise-induced increase ininsulin sensitivity. J Appl Physiol 108: 554–560, 2010.
57. Nordestgaard BG, Benn M, Schnohr P, Tybjaerg-Hansen A. Nonfast-ing triglycerides and risk of myocardial infarction, ischemic heart disease,and death in men and women. JAMA 298: 299–308, 2007.
58. Olivecrona T. The metabolism of 1-C14-palmitic acid labeled chylomi-crons in rats. Acta Physiol Scand 55: 170–176, 1962.
59. Pafili ZK, Bogdanis GC, Tsetsonis NV, Maridaki M. Postprandiallipemia 16 and 40 hours after low-volume eccentric resistance exercise.Med Sci Sports Exerc 41: 375–382, 2009.
60. Persson XM, Blachnio-Zabielska AU, Jensen MD. Rapid measurementof plasma free fatty acid concentration and isotopic enrichment usingLC/MS. J Lipid Res 51: 2761–2765, 2010.
61. Petitt DS, Arngrimsson SA, Cureton KJ. Effect of resistance exerciseon postprandial lipemia. J Appl Physiol 94: 694–700, 2003.
62. Poirier P, Eckel RH. Obesity and cardiovascular disease. Curr Athero-scler Rep 4: 448–453, 2002.
63. Puga GM, Meyer C, Mandarino LJ, Katsanos CS. Postprandial spill-over of dietary lipid into plasma is increased with moderate amounts ofingested fat and is inversely related to adiposity in healthy older men. JNutr 142: 1806–1811, 2012.
64. Saris WH, Schrauwen P. Substrate oxidation differences between high-and low-intensity exercise are compensated over 24 hours in obese men.Int J Obes Relat Metab Disord 28: 759–765, 2004.
65. Seidell JC, Muller DC, Sorkin JD, Andres R. Fasting respiratoryexchange ratio and resting metabolic rate as predictors of weight gain: theBaltimore Longitudinal Study on Aging. Int J Obes Relat Metab Disord16: 667–674, 1992.
66. Singhal A, Trilk JL, Jenkins NT, Bigelman KA, Cureton KJ. Effect ofintensity of resistance exercise on postprandial lipemia. J Appl Physiol106: 823–829, 2009.
67. Stampfer MJ, Krauss RM, Ma J, Blanche PJ, Holl LG, Sacks FM,Hennekens CH. A prospective study of triglyceride level, low-densitylipoprotein particle diameter, and risk of myocardial infarction. JAMA276: 882–888, 1996.
68. Suh SH, Casazza GA, Horning MA, Miller BF, Brooks GA. Effects oforal contraceptives on glucose flux and substrate oxidation rates duringrest and exercise. J Appl Physiol 94: 285–294, 2003.
69. Thyfault JP, Kraus RM, Hickner RC, Howell AW, Wolfe RR, DohmGL. Impaired plasma fatty acid oxidation in extremely obese women. AmJ Physiol Endocrinol Metab 287: E1076–E1081, 2004.
70. Trombold JR, Christmas KM, Machin DR, Kim IY, Coyle EF. Acutehigh-intensity endurance exercise is more effective than moderate-inten-sity exercise for attenuation of postprandial triglyceride elevation. J ApplPhysiol 114: 792–800, 2013.
71. Tsetsonis NV, Hardman AE. Reduction in postprandial lipemia afterwalking: influence of exercise intensity. Med Sci Sports Exerc 28: 1235–1242, 1996.
72. Tsetsonis NV, Hardman AE, Mastana SS. Acute effects of exercise onpostprandial lipemia: a comparative study in trained and untrained middle-aged women. Am J Clin Nutr 65: 525–533, 1997.
73. Tuazon MA, Henderson GC. Fatty acid profile of skeletal musclephospholipid is altered in mdx mice and is predictive of disease markers.Metabolism 61: 801–811, 2012.
74. Votruba SB, Atkinson RL, Hirvonen MD, Schoeller DA. Prior exerciseincreases subsequent utilization of dietary fat. Med Sci Sports Exerc 34:1757–1765, 2002.
1753Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from

75. Votruba SB, Atkinson RL, Schoeller DA. Prior exercise increasesdietary oleate, but not palmitate oxidation. Obes Res 11: 1509–1518,2003.
76. Votruba SB, Atkinson RL, Schoeller DA. Sustained increase in dietaryoleic acid oxidation following morning exercise. Int J Obes (Lond) 29:100–107, 2005.
77. Zafeiridis A, Goloi E, Petridou A, Dipla K, Mougios V, Kellis S.Effects of low- and high-volume resistance exercise on postprandiallipaemia. Br J Nutr 97: 471–477, 2007.
78. Zhang JQ, Ji LL, Fogt DL, Fretwell VS. Effect of exercise duration onpostprandial hypertriglyceridemia in men with metabolic syndrome. JAppl Physiol 103: 1339–1345, 2007.
79. Zhang JQ, Ji LL, Nunez G, Feathers S, Hart CL, Yao WX. Effect ofexercise timing on postprandial lipemia in hypertriglyceridemic men. CanJ Appl Physiol 29: 590–603, 2004.
80. Zotou E, Magkos F, Koutsari C, Fragopoulou E, Nomikos T, SidossisLS, Antonopoulou S. Acute resistance exercise attenuates fasting andpostprandial triglyceridemia in women by reducing triglyceride concen-trations in triglyceride-rich lipoproteins. Eur J Appl Physiol 110: 869–874, 2010.
81. Zurlo F, Lillioja S, Esposito-Del Puente A, Nyomba BL, Raz I, SaadMF, Swinburn BA, Knowler WC, Bogardus C, Ravussin E. Low ratioof fat to carbohydrate oxidation as predictor of weight gain: study of 24-hRQ. Am J Physiol 259: E650–E657, 1990.
1754 Postexercise Fat Metabolism in Obesity • Davitt PM et al.
J Appl Physiol • doi:10.1152/japplphysiol.00095.2013 • www.jappl.org
on Septem
ber 11, 2014D
ownloaded from





























