Embed Size (px)
Citation preview

Journal of Ecology 1994, 82, 933-950
ESSAY REVIEW
Predicting dispersal spectra: a minimal set of hypotheses based on plant attributes LESLEY HUGHES, MICHAEL DUNLOP, KRISTINE FRENCH,* MICHELLE R. LEISHMAN,t BARBARA RICE, LOUISE RODGERSON and MARK WESTOBY School of Biological Sciences, Macquarie University, North Ryde, NSW 2109, Australia
Summary
1 The dispersal mode adopted by a plant species is frequently associated with other attributes of the plant and its habitat. In this paper we review these associations and present a set of hypotheses which, when considered together, make a probabilistic prediction of the dispersal mode adopted by a plant species. When applied to a species list, the hypotheses can be used to generate a prediction of its dispersal spectrum, i.e. the percentages of different dispersal modes that have been adopted. 2 The formulation of such a set of hypotheses has several purposes: (i) to summarize existing knowledge about dispersal adaptations and their interrelations with other attributes of plants and their habitats; (ii) to couch that knowledge in such a way that falsifiable predictions can be made; (iii) to arrive at provisional conclusions about which factors are the most important in shaping the evolution of dispersal mode in different plants or different environments. 3 The review of relationships between dispersal mode and other attributes of plants and their habitats lead to the following provisional conclusions; (i) seeds larger than 100 mg tend to be adapted for dispersal by vertebrates while those smaller than 0.1 mg tend to be unassisted; most seeds, however, are between 0.1 and 100 mg, and in this range all of the dispersal modes are feasible; (ii) plant growth form and stature (sometimes in relation to the canopy height of the vegetation) seem to exclude certain dispersal modes; (iii) the availability of specific dispersal vectors seems rarely to be an important determinant of dispersal mode; (iv) attributes of the physical environment also seem rarely to be important, except indirectly through their influence on plant stature and seed size.
Keywords: dispersal correlates, dispersal mode, dispersal spectrum, plant attributes, seed dispersal
Journal of Ecology (1994) 82, 933-950
Introduction
The diaspores of many plant species have charac- teristic morphological structures that enhance their probability of being dispersed away from the mother plant. Dispersal of seeds is presumed to have fitness advantages for plants, such as increasing the chances of founding a lineage in a new locality, or reducing
density-dependent mortality. The advantages of dis- persal have been considered theoretically (e.g. Ham- ilton & May 1977; Motro 1982; Venable & Brown 1993), and the empirical evidence for advantages in particular species has been extensively reviewed (e.g. Harper 1977; Howe & Smallwood 1982; Willson 1983, 1992).
In this paper we are concerned with the question of why a plant 'chooses' one dispersal mode over another, and why certain vegetation types have characteristic proportions of plant species with different dispersal modes (dispersal spectra). Dis- persal spectra vary from place to place (Willson et al. 1990), presumably because the relative merits of different dispersal modes are affected by attributes of
Correspondence: Lesley Hughes, Present address: Museum of Comparative Zoology, Harvard University, 26 Oxford St, Cambridge, MA 02138, USA.
Present addresses: *Biology Department University of Wollongong, Wollongong, NSW 2500, Australia and tlmperial College at Silwood Park, Ascot, Berks. SL5 7PY, UK 933
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

934 Predicting dispersal spectra from plant attributes
the plant in question (e.g. seed size, height) and by the environmental circumstances under which seedlings establish or fail (e.g. after fire from soil seed bank, in deep shade, in dry soil conditions).
We outline a series of hypotheses about associ- ations between dispersal mode and plant attributes that, when considered together, form a compound hypothesis that seeks to explain the distribution of dispersal modes within vegetation. In the first section, seven hypotheses are described that lead to the identi- fication of particular dispersal modes that seem rarely to be adopted by plants with certain attributes. These can be termed 'exclusion hypotheses', for convenience. Many of the ideas expressed in these hypotheses have been suggested in different forms before, and some are well supported by empirical evidence. Others seem to be demanded by logic, and yet others seem to be required by known patterns of dispersal spectra, even though there is little direct evidence as to why they have arisen. We have attempted to formulate a minimal set of such hypoth- eses, and have not included every feature of a plant or an environment that might conceivably be important. For example, we have not included successional status or possible phylogenetic constraints in the list, although we consider the possible importance of these factors in the Discussion. During the review of evi- dence for each hypothesis we note that for most there are at least one or two plant species that are obvious exceptions. These exceptions reinforce the fact that these hypotheses are preliminary, and will require test- ing and refinement. In the second section of the paper we present a set of probability distributions for different dispersal modes as a function of seed size. These distributions have been obtained from a com- pilation of data from four temperate floras. If a species list for a particular area of vegetation is available, these probabilities may be applied to each species, for dispersal modes not already excluded. This process leads to an assessment as to which dispersal mode, or modes, are most likely to be adopted by a seed of the given size. The whole set of hypotheses can be used to make a probabilistic prediction as to what dispersal mode a particular plant species might be expected to have adopted over evolutionary time, or a prediction as to the percentages of species adopting different dispersal modes in a vegetation type.
This formulation of a set of hypotheses about dis- persal spectra has two main aims. First, it provides a framework within which existing knowledge can be organized and highlights the weaknesses as well as the strengths of the available evidence. Secondly, it provides testable predictions allowing a-priori tests to be devised and interpreted. Such tests in turn should lead to continuing refinement of the set of hypotheses, by acceptance or rejection of particular hypotheses, and by recognition of the need for further ones.
We have restricted the hypotheses to those needed to explain the distribution of the six major types of
morphological adaptations for dispersal considered by Willson et al. (1990) in their review of dispersal spectra.
1 Wind-dispersed: indicated by the presence of wings, plumes or hairs that increase air resistance and slow the rate of descent. 2 Vertebrate-dispersed (internal): indicated by the presence of fleshy pulp or aril that is ingested; the seeds are then either regurgitated or defaecated. 3 Ant-dispersed: indicated by the presence of an elaio- some, an appendage on seeds that is attractive to ants. 4 Ballistically dispersed: the diaspore is ejected explosively from a seed-pod. 5 Adhesively dispersed: indicated by the presence of hooks, barbs or sticky substances that cling to fur or feathers. 6 Unassisted: no obvious dispersal adaptation.
EXCLUSION HYPOTHESES
The hypotheses are summarized in Table 1, and the logic/evidence for each is discussed below.
Species below a dense canopy are unlikely to use wind
Wind speed decreases sharply from the level of the upper canopy down to the level of maximum leaf area and winds in the lower canopy are very weak (Oke 1987). For example, wind speed measured by Oliver (1971) in a mixed pine forest declined from -0.95m s-', measured 0.5m above the canopy, to less than 0.2 m s-' at the canopy base. Within the canopy, as a general rule, wind speed is inversely related to the density of the foliage (Grace 1977).
Diaspores with adaptations for wind dispersal such as wings or plumes do not travel far in still air. For example, the lateral movement in still air of 27 wind- dispersed species measured by Matlack (1987) ranged from 0.04-0.87 m, with only two species travelling further than 0.3 m. The estimated lateral distance these species would have achieved in a 2.8 m s-' (10 km
Table 1 Summary of exclusion hypotheses
Dispersal modes Plant attribute to be excluded
1 Species below a dense wind canopy
2 Canopy width ) 4 m ants, ballistic 3 Presence of serotinous seed vertebrate, ants,
bank which survives fire adhesion, ballistic 4 Presence of soil seed bank all modes except ant
which survives fire unless seeds/diaspores < 3mg
5 Epiphytes ballistic, adhesion
6 Stem parasites wind, ants, unassisted, ballistic, adhesion
7 Height over 2 m adhesion
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

935 L. Hughes et al.
h-') breeze ranged from 1.47-111.6 m (81% over 1O m, 41% over 40m), and all species, except for the two with the heaviest seeds, would be able to 'escape' the immediate vicinity of the parent.
Several studies have compared the percentage of wind-dispersed species in the canopy and the sub- canopy or middlestorey of tropical and temperate forests (summarized in the Appendix, TableAl; see also Willson 1991). Wind-dispersed species are con- sistently found in higher proportion in canopy species, with the proportion of other dispersal modes, such as fleshy fruits for vertebrate dispersal, increasing towards the lower strata. although this generalization, like several others to be described below, does have some known exceptions. Wind dispersed species are not completely absent from the understorey, e.g. Sol- idago spp. in north temperate woodlands (P.J. Grubb, personal communication), but the dispersal success of such species is not known.
A slight variant on this hypothesis may be more appropriate for species that only release seeds after fire. For these species the density of the canopy in the immediate post-fire period, rather than just before the next fire, will be the most relevant.
Species with a canopy diameter wider than 4 m are unlikely to use ant or ballistic dispersal
The dispersal distances achieved by different dispersal modes vary a great deal (Appendix Tables A2-5; see also Willson 1993). Distances that seeds are carried by ants, limited by ant foraging distances, are typically 1-2 m and rarely exceed 5 m although the occasional exceptional distance has been reported (Table A2). Distances achieved by ballistic propulsion are con- strained by the physical mechanism itself (Willson 1993) and, like ant-dispersal distances, rarely exceed a few metres (Table A3).
Plant stature may be an important constraint on the particular dispersal mode 'chosen' by a plant. It is generally accepted that escaping competition in the immediate vicinity of the parent plant is an important advantage of seed dispersal (Howe & Smallwood 1982). The restricted distances that seeds are dispersed by ants or by ballistic mechanisms mean that effective escape will only be achieved if the parent is small.
In order that this hypothesis be applied to an actual database of plant attributes we have chosen 4 m as an arbitrary canopy diameter above which we consider dispersal by ants or ballistic mechanisms unlikely to have been adopted. This cut-off point is fairly con- servative as the vast majority of ant- and ballistically dispersed plants have canopy diameters much less than this. In the northern hemisphere, ant-dispersed plants are mostly herbs in the understorey of mesic forests (e.g. Handel et al. 1981; Beattie & Culver 1981). In the southern hemisphere they are typically sclerophyllous shrubs (Westoby et al. 1982; Milewski & Bond 1982), the majority of which have canopy
diameters less than 2 m (e.g. Westoby et al. 1990). Ballistically dispersed plants also tend to be of small stature (Ridley 1930). With only two exceptions (see below), all the ballistic species listed in Table A3 are shrubs or understorey herbs. Air resistance results in initial velocity having a greater influence on dispersal distance than the height of release (Beer & Swaine 1977; Stamp 1989a) so tall plants do not have a par- ticular advantage due to their height.
The most notable exceptions to this hypothesis are the ballistically dispersed tropical trees, Hura cre- pitans and Bauhinia purpurea. Both these species have extraordinarily powerful ballistic mechanisms which can propel seeds many metres (Stebbins 1971; Swaine & Beer 1977).
In contrast to dispersal by ants and ballistic pro- pulsion, seeds dispersed by wind and vertebrates can potentially be much more widely dispersed even if the greater distances are achieved by only by a small proportion of the seed crop; these dispersal modes may therefore be more likely to be adopted by larger plants. Wind dispersal produces a strongly leptokurtic distribution (Harper 1977). Many seeds are found around the base of the parent plant but a small pro- portion of seeds may travel considerable distances (examples in TableA4), depending on the speed and turbulence of the wind as well as the height of release and the size, shape and aerodynamic properties of the seed. Most estimates of distances achieved by wind dispersal have been made by estimating the fall vel- ocities of seeds in still air, and then extrapolating for different wind speeds (e.g. Sheldon & Burrows 1973; Matlack 1987). Field measurements of dispersal dis- tances have been made using seed traps and in these studies, the maximum distances measured are of course constrained by the maximum distances from the plant at which the traps are placed (e.g. Cremer 1966; Gashwiler 1969). The more anecdotal accounts of wind dispersal distances frequently stem from observations of plants that have colonized oceanic islands (e.g. Fridriksson 1975).
Frequency distributions of dispersal distances for seeds ingested by vertebrates share some of the char- acteristics of those for wind dispersed seeds (Table A5), although the direction of seed movement and the local density of seed deposition will be influenced by the behaviour of the particular frugivore (Stiles 1992). Dispersal distances for ingested seeds are lim- ited by gut passage rates which vary from a few minutes in frugivorous birds to several months in ungulates (Janzen 1984, and see review by Willson 1989). The few field measurements of dispersal dis- tances that have been made, like those for wind-dis- persed seeds, are usually limited by the maximum number and distance of traps from the seed source (e.g. Smith 1975; McDonnell & Stiles 1983; Hoppes 1988).
Seeds dispersed by nonfrugivorous vertebrates may also have the potential to travel considerable
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

936 Predicting dispersal spectra from plant attributes
distances, although few field measurements have been made. Virtually the only published data on dispersal distances of adhesive diaspores are from Bullock & Primack (1977), who estimated dispersal distances of adhesive species by walking through vegetation with a cloth board to which the diaspores adhered. The mean dispersal distances obtained ranged from 109 to 250 m. In principle, however, adhesive fruits may be carried by a dispersal agent indefinitely, depending on how quickly the fruit is detected and removed by the host. Adhesive fruits on inaccessible body parts of long-lived animals may remain attached for long periods of time (Liddle & Elgar 1984; Shmida & Ellner 1983). For example, the dispersal of seeds over at least 650 km to Macquarie Island is assumed to have occurred by adhesion to sea birds (Taylor 1954). Many weed species may have been introduced to North America from Mexico through long distance dispersal of adhesive fruits attached to sheep (reviewed by Sorensen 1986).
In summary, plant stature may place constraints on which dispersal mode a species might adopt. Seeds adapted for dispersal by wind, vertebrates and adhesion have the potential for far greater dispersal distances than those dispersed by ants or ballistic propulsion. The relatively short dispersal distances achieved by ants and ballistic dispersal may therefore be more likely to be associated with small plants.
Species with a serotinous seed bank that survives fire are unlikely to use vertebrate, ant, ballistic or adhesive dispersal
Many species in the fire-prone sclerophyllous shrub- lands and woodlands of Australia and South Africa and in the coniferous forests of the Northern Hemi- sphere are serotinous, retaining their seeds in per- sistent cones or fruits within the canopy. These seeds may be stored for many years before being released either after a fire, or when the plant dies. Most sero- tinous species have hairs or wings, adapting them for dispersal by wind. Bond (1985) noted, for example, that within the South African genus Leucadendron, the serotinous species are generally adapted for wind dispersal whereas many of the non-serotinous species are dispersed by ants. Serotinous species in the family Myrtaceae are exceptions to this generalization, hav- ing seeds with no obvious dispersal adaptations (Lamont et al. 1991).
Wind dispersal may be enhanced in the immediate post-fire period because fire frees the canopy and ground of many obstacles to the passage of wind and seeds. For example, Bond (1988) showed that following fire, fruits of some Protea and Leucadendron species may be tumbled by wind over 50Gm. In addition, the heat generated by the blackened soil surface following fire creates up-draughts and small whirlwinds which may also carry and disperse seeds.
(Le Maitre, personal observation cited in Lamont etal. 1991).
Dispersal structures such as fleshy fruit for ver- tebrate dispersal, or elaiosomes for ant dispersal, may not be physically compatible with long-term storage of seeds in the canopy. Prolonged storage of simple carbohydrates (as pulp) or lipids (as elaiosomes) between fires may increase the probability of des- iccation or of attack by micro-organisms and insects. Excessive heating could also reduce the attractiveness and palatability of the diaspore. It is not so intuitively obvious why hooks or barbs for adhesive dispersal, or ballistic mechanisms, are not found on serotinous seeds. A possible explanation for the lack of adhesive dispersal is that the general availability of vertebrate vectors directly after a fire is limited.
Species with a soil seed bank that survives fire are unlikely to use dispersal modes other than ants unless seeds and/or diaspores are small (less than - 3 mg) and therefore able to filter down through the soil
During a fire, seeds on the soil surface or in the litter may be exposed to temperatures in excess of 300?C (Floyd 1966; Scotter 1970; Dunn & DeBano 1977; Stott 1986). As temperatures greater than 140?C are lethal for most seeds even for short durations (e.g. Auld 1986; Auld & O'Connell 1991; French & Westoby, unpublished), mortality of seeds on the sur- face and in the top 1-3 cm of soil during a fire can be considerable (Floyd 1966; Ewel et al. 1981; Cheplick & Quinn 1987). A dispersal mechanism that results in some form of seed burial will clearly be advantageous for species developing soil seed banks in fire-prone habitats.
There is probably no optimum burial depth for seeds as temperatures reached even during any single fire vary from place to place within a site and variation between fires is also considerable (Christensen & Kimber 1975; Stott 1986; Hobbs & Atkins 1988; Brad- stock et al. 1992; Hughes & Westoby 1992). In addition, soil temperatures during fires decrease rap- idly with depth such that seeds at one depth may be exposed to a temperature several times greater than those buried only 1 or 2cm deeper (e.g. Floyd 1966; Auld 1986). As a result, during any single fire, there will be only a narrow range of depths at any particular place where the soil temperature will be suitable to stimulate germination without being lethally hot. The depth at which this temperature 'window' will be reached varies considerably, even over a few metres or less. A dispersal mode which results in the burial of seeds at a range of depths might therefore be an appropriate form of risk-spreading in such an environment.
Dispersal modes other than ant dispersal result in seeds being initially deposited on the soil surface. Such seeds may become buried shallowly by surface runoff or falling litter, but the range of depths will be narrow.
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

937 L. Hughes et al.
For example, experiments performed by van Tooren (1988) using 'seed-sized' glass beads showed that only 4% placed on the surface became buried at depths greater than 1 cm in 6 months. Such a narrow, and shallow, range of burial depths will increase the prob- ability of large-scale mortality during a hot fire. Small seeds, however, have a greater chance of filtering down through the soil. The only data available for the relationship between seed size and the likelihood of passive burial are somewhat indirect. Thompson et al. (1993) showed that plant species with persistent seed banks (seeds staying for more than 5 years in the soil) generally have compact diaspores weighing less than 3 mg. Therefore to apply this hypothesis to a species list, we have chosen 3 mg as an arbitrary cut-off point, above which we cons'ider seeds unlikely to survive fires in a seed bank unless actively buried by ants.
Dispersal by ants, unlike other dispersal modes, may result in seeds being buried down to depths of 20 cm, although the majority of seeds in ant nests are found in the upper 8cm (Shea et al. 1979; Bond & Slingsby 1983; Bond & Stock 1989; Hughes & Wes- toby 1992). Burial of seeds by ants over a range of depths may have been a selective pressure leading to the dominance of ant-dispersed plants in sclero- phyllous vegetation in Australia and South Africa where fires are frequent (e.g. mean interval between fires in south-eastern Australia is about 5-12 years; Walker 1981). In fire-prone vegetation types with longer intervals between fires such as the Californian chaparral (20-30 years; Keeley & Zedler 1978) ant- dispersed plants are relatively uncommon (Berg 1966; Bullock 1974); one explanation is that longer fire intervals may increase the probability that seeds will be passively buried at a greater range of depths with- out the active participation of ants.
In the chaparral vegetation of California, Keeley (1991) identified two distinct seed germination syn- dromes. Seeds of 'fire-resister' species do not form persistent seed banks and are not stimulated to ger- minate by fire. These species are almost exclusively fleshy-fruited and dispersed by vertebrates. In contrast, seeds of 'fire-recruiter' species survive fire in persistent seed banks and are stimulated to germinate immediately after fire. Consistent with the hypothesis outlined above, most of these species have no obvious dispersal adaptation but generally have smaller seeds than those of the fire-resisters,.when similar growth forms are compared. Two genera of fire-recruiters, Fremontodendron and Dendromecon, are ant- dispersed. Ceanothus, the one genus that is bal- listically dispersed, is an exception to this hypothesis.
Epiphytes are unlikely to use ballistic and adhesive dispersal
Epiphytes have very specific site requirements for establishment, i.e. the limbs of plants. As such safe sites may be few and patchily distributed, two alter-
native dispersal strategies may be favoured by these plants. First, by producing large quantities of small, light seeds (either dust seeds or with structures for wind dispersal) epiphytes can not only increase the probability that a few by chance will reach a suitable site for establishment but also that some seeds will land on branches above, as well as below, the release point. Alternatively, they may adopt a dispersal strat- egy that results in some degree of targeting toward specific sites. This can be achieved by producing seeds with fleshy fruits attractive to certain vertebrates, especially birds, which are then deposited directly onto branches when the vector perches. Dispersal by ants may also result in seeds being placed in suitable sites for establishment, such as nest carton. Dispersal by ballistic explosion, or by adhesion, may be unsuit- able for epiphytes because both dispersal mechanisms result in untargeted dispersal, unlike dispersal by cer- tain frugivores or ants.
The dispersal mechanisms for the estimated 28 000 species of epiphytes have been summarized by Madi- son (1977) and the results show that the vast majority have adopted one of the two strategies outlined above. Over 20 000 species (from 605 genera) have dust-seeds or spores which lack appendages (strategy 1). These diaspores are very small (most are less than 1 mm in length) and are dispersed by wind and/or rain splash. The majority of these species are orchids but diaspores of this type are also found in the Melastomataceae, Begoniaceae, Lentibulariaceae, Rapateaceae and the ferns. Winged or plumed seeds are found in 1190 species (37 genera) and are characteristic of many Bromeliaceae, Asclepiadaceae, Gesneriaceae and Rubiaceae. Fleshy fruits (strategy 2) are found in 4400 species (191 genera). Many of these fleshy fruits in fact contain numerous minute seeds which, when removed from the fruit, are very similar to the dust-seeds of capsular fruited epiphytes; these species can be thought of as combining strategies 1 and 2. A few species of epiphytes have elaiosomes (Janzen 1974; Weir & Kiew 1986) and are removed by ants. Some species without elaiosomes are also removed by ants which are attracted by the fleshy pulp (e.g. Roberts & Heithaus 1986). Several of these epiphyte species are restricted to arboreal ant nest gardens (Kleinfeldt 1978; Madison 1979; Davidson 1988; Benzing & Clements 1991). A further small group of species have fruit structures that suggest adaptation for dispersal by both vertebrates and ants (Kleinfeldt 1978, 1986; Madison 1979; Davidson 1988; Kaufmann etal. 1991).
The only exception to this hypothesis that we know of is the epiphyte Peperomiafernandeziana which has sticky diaspores (Armesto & Rozzi 1989).
Stem parasites are unlikely to use dispersal mechanisms other than vertebrates
Like epiphytes, stem parasites need to be dispersed to the limbs of suitable host plants. Unlike epiphytes
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions
938 Predicting dispersal spectra from plant attributes
however, they also produce a haustorium which pen- etrates the host during establishment. This charac- teristic may set a lower limit on seed size. The tar- geting of larger seeds is presumably achieved best by vertebrate dispersal.
The majority of stem parasites are in the Loran- thaceae (mistletoes). Mistletoe diaspores are generally fleshy, with sticky seeds that adhere to branches after being defaecated by birds (Liddy 1983; Willson 1983; Davidar 1987; Benzing 1989). Stem parasites in other families, such as Cassytha (Cassythaceae) and Cuscuta (Convolvulaceae), also have fleshy fruits. An exception to this hypothesis is the dwarf mistletoe (Arceu- thobium) in North America which may be dispersed ballistically (Hinds, Hawksworth & McGinnies 1968), or by adhesion to fur or feathers (Willson 1983).
Plants over 2 m tall are unlikely to use adhesion
Several studies have shown that adhesive dispersal modes tend to be confined to low-growing species. In a survey of species with adhesive fruits in 10 floras, Sorensen (1986) found that nearly 75% were less than 1 m in height and 95% were less than 2 m. Eighty-five per cent of the adhesive Asteraceae surveyed in 18 regional floras by Venable and Levin (1983) were low- growing herbaceous plants and 6.6% were shrubs; none were trees. Thirteen out of 14 adhesive species in the Brazilian cerrado surveyed by Gottsberger & Silberbauer-Gottsberger (1983, cited in Sorensen 1986) occurred in the ground layer. Adhesive seeds are consistently restricted to herbs in Andean paramo vegetation (Frantzen & Bouman 1989). Milton, Sieg- fried & Dean (1990) noted that few species with adhesive seeds were tall, and in a survey of adhesive species in African Karoo savannah and grassland, found these species to be exclusively confined to the grassland. In central Australian arid vegetation, adhesive species are confined to low-growing graminoids and forbs (Jurado et al. 1991).
The reason for adhesive dispersal mechanisms being confined to plants of low stature is presumably related to the availability of dispersal vectors at a suitable height (Guitian & Sanchez 1992). Arboreal mammals may not be suitable because they tend to be more agile and therefore more likely to find and remove diaspores quickly while grooming (Willson et al. 1990).
RELATIONSHIP OF DISPERSAL MODE AND
SEED SIZE
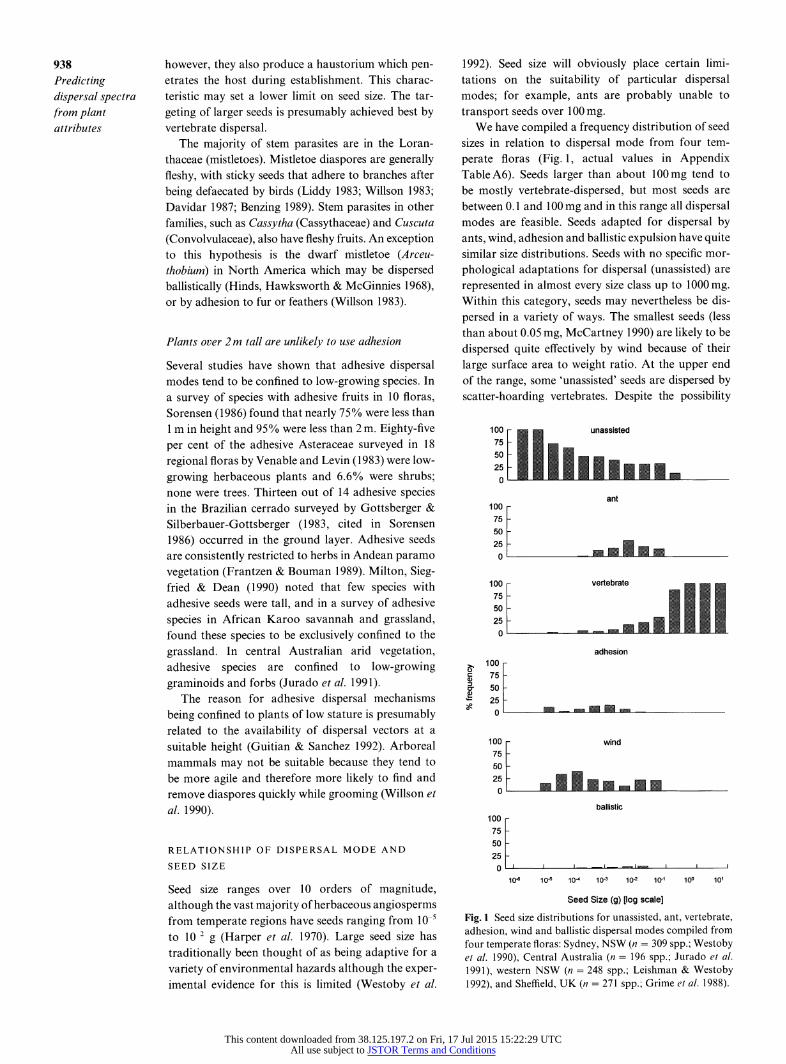
Seed size ranges over 10 orders of magnitude, although the vast majority of herbaceous angiosperms from temperate regions have seeds ranging from 10-5 to 10 2 g (Harper et al. 1970). Large seed size has traditionally been thought of as being adaptive for a variety of environmental hazards although the exper- imental evidence for this is limited (Westoby et al.
1992). Seed size will obviously place certain limi- tations on the suitability of particular dispersal modes; for example, ants are probably unable to transport seeds over 1 00 mg.
We have compiled a frequency distribution of seed sizes in relation to dispersal mode from four tem- perate floras (Fig. 1, actual values in Appendix Table A6). Seeds larger than about 100mg tend to be mostly vertebrate-dispersed, but most seeds are between 0. 1 and 1 O0 mg and in this range all dispersal modes are feasible. Seeds adapted for dispersal by ants, wind, adhesion and ballistic expulsion have quite similar size distributions. Seeds with no specific mor- phological adaptations for dispersal (unassisted) are represented in almost every size class up to 1000 mg. Within this category, seeds may nevertheless be dis- persed in a variety of ways. The smallest seeds (less than about 0.05 mg, McCartney 1990) are likely to be dispersed quite effectively by wind because of their large surface area to weight ratio. At the upper end of the range, some 'unassisted' seeds are dispersed by scatter-hoarding vertebrates. Despite the possibility
100 _ unassisted 75
50 25 -
ant 100 v 75 - 50 E 25 -| 0
r0 vertebrateXXX 75 -
50 50 25 -
0
adhesion 100 w
<,75 -
v50 - 4- 25-
F 100 rwind 75 50 25 m 0
ballistic 100 - 75 - 50 -
25-
10-6 10-5 10-4 10-3 10-2 10-1 10? 101
Seed Size (g) [log scale]
Fig. I Seed size distributions for unassisted, ant, vertebrate, adhesion, wind and ballistic dispersal modes compiled from four temperate floras: Sydney, NSW (n = 309 spp.; Westoby et al. 1990), Central Australia (n = 196 spp.; Jurado et al. 1991), western NSW (n = 248 spp.; Leishman & Westoby 1992), and Sheffield, UK (n = 271 spp.; Grime et al. 1988).
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions
939 L. Hughes et al.
that unassisted seeds will actually be dispersed by a variety of vectors we have followed Willson et al. (1990) in not subdividing this category for two main reasons. First, it is difficult to assign an exact seed weight below which seeds should be regarded as dispersed by wind, and it seems unlikely that small seed size, in itself, is a specific adaptation for dispersal. Secondly, reliable information as to which seeds are dispersed by scatter- hoarders is not available for these floras.
PREDICTING DISPERSAL SPECTRA
To illustrate how the hypotheses outlined above might be applied to a particular flora we provide a worked example using data compiled by Westoby et al. (1990) for four sites near Sydney, NSW. The sites consist of fire-prone, dry sclerophyll vegetation on infertile, sandstone-derived soils. As many ideas expressed in this paper have been influenced by our knowledge of the vegetation at these sites, and we have used this data in the compilation of the seed-size frequency distribution, we present this exercise as an illustration of the process rather than an independent test of the set of hypotheses.
The following data are needed for each species: canopy width, habit (particularly whether the species is epiphytic or parasitic), height, presence or absence of either a serotinous or soil seed bank that survives fire, and seed size. The seven exclusion hypotheses are applied to each species, resulting in up to five of the possible six dispersal modes being ruled as unlikely to be adopted. The seed size of the species can then be compared to the frequency distributions of seeds for those dispersal modes not already excluded. A set of probabilities for each dispersal mode, for each species, is then generated (examples outlined below). Once all the species within the flora have been 'processed', the probabilities for each dispersal mode are added to create a hypothetical dispersal spectrum. A com- parison of this spectrum to the actual spectrum then
serves to indicate how adequately the set of hypoth- eses explain the observed pattern.
Table 2 shows examples of the predictions of dis- persal mode for six of the species from the sites; these species display a range of habits, heights, seed sizes and seed bank characteristics. The table also gives the actual dispersal mode for each species. A brief description of each of these six species and the way in which the various hypotheses were applied is described below:
1 Banksia ericifolia is a large shrub (to -4 m) with serotinous seeds i.e. the seeds are stored on the plant in woody follicles until after fire. If seed size alone were used as a criterion to predict the dispersal mode for this species, the histogram in Fig. 1 would give the following probabilities: unassisted 31%, ant 20%, vertebrate 22%, adhesion 1%, wind 22%, and bal- listic 4%. However, the application of hypothesis 3 results in vertebrate, ant, ballistic and adhesion dis- persal modes being excluded (Table 2). The prob- abilities are therefore adjusted, resulting in a pre- diction of the seeds having a 58% probability of being unassisted and a 42% probability of being wind- dispersed. 2 Dillwynia rudis is a sclerophyll shrub (1 m) with hard seeds (3.4 mg) that germinate from a soil seed bank after fire. As this type of seed needs to be buried to survive surface fire temperatures we would exclude all dispersal modes except ants (hypothesis 4). 3 Eucalyptus piperita is a large tree (- 20 m) with a canopy width of at least 6 m when mature. Following application of hypotheses 2 and 7, the dispersal cat- egories of ants, ballistic and adhesion are excluded. The seed size histogram then generates a prediction that this species has a 59% probability of being unas- sisted, 11% probability of being vertebrate-dispersed and 30% probability of being wind-dispersed. 4 Pterostylis daintreana is a small (- 30cm) orchid with very small seeds (0.0008 mg). From Fig. 1, all seeds in this size category are unassisted. 5 Cassytha pubescens is a twining stem parasite. Fol-
Table 2 Attributes and predicted dispersal modes of six selected species from the database of Westoby, Rice and Howell (1990) Dispersal modes have been predicted by the application of the 7 exclusion hypotheses and the seed size histogram (Fig. 1). U = unassisted, A = ant, V = vertebrate, Ad = adhesion, W = wind, B = ballistic, X = dispersal mode excluded
Canopy Seed Seed bank Predicted dispersal mode (%) Actual Height width size in relation dispersal
Species Habit (m) (m) (mg) to fire U A V Ad W B mode
Banksia ericifolia shrub 4 2-3 25.5 serotinous 58 X X X 42 X W Dillwynia rudis shrub 1 1 3.4 survives X 100 X X X X A
fire in soil Eucalyptus piperita tree 20 > 6 1.03 - 59 X 11 X 30 X U Pterostylis pubescens herb 0.03 0.05 0.0008 - 100 0 0 0 0 0 U Cassytha pubescens twining stem - - 92.17 - X X 100 X X X V
parasite Stylidium lineare perennial 0.20 0.04 0.12 - 47 1 5 7 39 1 U
herb
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

940 Predicting dispersal spectra from plant attributes
lowing application of hypothesis 6, all dispersal modes except vertebrate are excluded for this species. 6 Stylidium lineare is a small perennial herb that grows in open heath and woodland. None of the hypotheses, as they currently stand, result in the exclusion of dispersal modes for this species. The pre- diction of its dispersal mode must therefore be gen- erated solely from the seed-size histogram (Fig. 1).
When the remaining 155 species at the sites are treated in a similar fashion, the following predicted dispersal spectrum is obtained by summing the prob- abilities for each of the six dispersal modes: unassisted 34%, ant 41%, vertebrate 6%, adhesive 4%, wind 14% and ballistic 1%. The actual dispersal spectrum, obtained from an examination of the diaspores of each species (Westoby et al. 1990), is as follows: unas- sisted 38%, ant 45%, vertebrate 7%, adhesive 0%, wind 9% and ballistic 1%. Eight of the 161 species (4.1 %) had their correct dispersal modes wrongfully excluded using the hypotheses. The close match between the predicted and actual dispersal spectra is not surprising for the reasons outlined above and we must again stress that this is an illustrative exercise rather than a test of the set of hypotheses. The most important point to note is that the prediction for each species is probabilistic, but for the dispersal spectrum is absolute. We will return to this point during the discussion in the section regarding phylogeny.
Discussion
SOME PROSPECTS FOR DISPERSAL ECOLOGY
A great deal is known about the ecology of seed dis- persal. Many observations have accumulated about distances and seed shadows achieved and there has been some quantification of dispersal spectra both for different vegetation types (Willson et al. 1990), and in relation to plant height and seed size (this paper). There has also been considerable experimentation on the merits of different dispersal modes.
Because of this firm basis of accumulated knowl- edge, it is now realistic to set the following target for future research: to establish a well-tested set of hypotheses, applicable world-wide, relating dispersal mode to attributes of the plants and the environment. Our purpose has been to take a step toward this target, by reviewing existing knowledge and formulating a draft set of hypotheses amenable to future falsification and refinement.
An example of a prospective falsification of the hypotheses would be as follows: Several studies have found that the incidence of fleshy-fruited species adapted for dispersal by vertebrates increases with increasing rainfall, while that of wind-dispersed spe- cies decreases (e.g. Gentry 1982; Howe & Smallwood 1982; Hoffmann et al. 1989; Willson et al. 1990; Jor- dano 1992). If the hypotheses proposed here are
sufficient, this pattern should prove to be interpretable as a secondary correlate of changes in plant height, canopy density, habit, and seed size. Alternatively, if the generalizations are not sufficient, it will prove necessary to add a further hypothesis invoking rainfall or some other correlate of rainfall.
There are certain groups of plants for which dis- persal modes will not be predicted with any degree of accuracy by the hypotheses as they stand at present. For example the dipterocarps, an important group of tropical trees, have very large seeds which are winged and thus adapted for dispersal by wind (e.g. Diptero- carpus grandiflorus seeds are 6 cm long with a wing 14 cm long; Ridley 1930). Emergent species of Shorea are predominantly winged while species within the canopy are not (Ashton 1989), which is consistent with our hypothesis 1. Nevertheless, our system of hypotheses would give dipterocarps a very low prob- ability of being wind-dispersed, due to their large seed size. It is possible that dipterocarps are specifically adapted for dispersal in tropical cyclones; Webber (1934) noted that Shorea seeds may be carried several hundred metres in strong winds. For the present, how- ever, there is not enough evidence to justify a hypoth- esis predicting large-seeded species to be adapted for wind-dispersal in any cyclone-prone environment. This example, as well as the other exceptions to the hypotheses already noted, illustrate that our hypoth- eses will certainly need elaboration before they can be considered fully satisfactory. At present they are likely to prove more useful in certain vegetation types, such as temperate woodlands, than in others, such as grass- lands.
In the remainder of this discussion we first consider some possible relationships which have not been included as prospective hypotheses, a particular case of which is the relationship of dispersal mode with phylogeny. We then discuss some biological impli- cations of the hypotheses, if indeed they prove to be sufficient to explain a significant proportion of patterns in dispersal spectra.
RELATIONSHIPS NOT INCLUDED AMONG THE
PROPOSED HYPOTHESES
Some factors have been quite extensively discussed in relation to dispersal mode, but nevertheless do not appear in the hypotheses described above. For exam- ple, wind dispersal has been described as common in early successional or disturbed habitats and animal- mediated dispersal as more common in later suc- cessional or more stable environments (e.g. Houssard et al. 1980; Fenner 1987; Hodgson & Grime 1990; Willson 1991; Guitian & Sanchez 1992; but see Ven- able & Levin 1983). In effect, we are hypothesizing that dispersal spectra can be understood in terms of plant attributes such as height and seed size, without direct reference to successional status.
Another factor we have not explicitly considered is
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

941 L. Hughes et al.
the availability of specific dispersal vectors. Fossil records of angiosperms indicate that dispersal syn- dromes are maintained in lineages even though the nature of the disperser fauna changes (Tiffney 1986; Bremer & Eriksson 1992). Tightly coevolved relation- ships between plants and their animal dispersers appear to be rare, with most plants being dispersed by several animal species and most seed-dispersing animals taking the diaspores of several plant species (Wheelright & Orians 1982). Exceptions to this gen- eralization are known, such as the dispersal of mistle- toe seeds by mistletoe birds (Reid 1991), and dispersal of certain species of Pinus by corvids (reviewed by Tomback & Linhart 1990; for other examples see Chapman et al. 1992)). Following the policy of for- mulating only a minimal set of hypotheses, we con- sidered that the devising of special hypotheses to cover these cases was unnecessary at this stage. Mistletoes would in any event be predicted as vertebrate-dis- persed by our hypothesis 6, and scatterhoarded Pinus seeds would not be treated as possessing adaptations for dispersal by vertebrates.
The set of hypotheses predicting the dispersal mode a species might adopt over evolutionary time, as well as the mix of dispersal modes found in a particular area of vegetation, despite being implicitly about evol- utionary processes, do not invoke phylogeny as a factor shaping dispersal mode. Our thinking on this subject is as follows: It is certainly true that whole genera and families often share a common mode of dispersal. To quite a large extent, such patterns would be generated by the hypotheses, since whole genera and families will also often tend to be tall rather than short plants, large- rather than small-seeded, and so forth. Such an effect, whereby the whole of a genus might be adapted for dispersal by vertebrates because the genus has large seeds and a physiology adapted to forest understorey, can be labelled 'phylogenetic niche conservatism' (Harvey & Pagel 1991). Under phylogenetic niche conservatism, relating dispersal mode to seed size and plant height is complementary to relating it to phylogeny, rather than an alternative interpretation.
Might dispersal mode be constrained by the devel- opmental pathway governing the structure of fruits (which in turn would be related to phylogeny)? Many families and some genera include species that have modified fruit structure sufficiently to have different dispersal modes (Ridley 1930; Corner 1949; Harper et al. 1970; Stebbins 1971; van der Pijl 1982; Bond & Slingsby 1983; Bremer & Eriksson 1992); for example, ant, bird, mammal and wind dispersal are all found among Mimosaceae (Willson 1992). Furthermore, the structures achieving dispersal by a given vector are often quite varied. Elaiosomes have developed in different species from completely different plant struc- tures (Roth 1977). Vertebrate dispersal can be achieved by arils or by pulp developed in several different ways. Epidermal hairs and wing-like out-
growths to increase air resistance can be found on various structures without any profound modi- fications of their architecture or development pattern (Stebbins 1971). Thus the potential for different dis- persal syndromes to arise within a lineage, over macro- evolutionary periods of millions of years, is con- siderable. Nevertheless, new fruit structures arise by modification of genes governing the developmental pathway, and it can hardly be doubted that a par- ticular sort of genetic pathway may predispose a lin- eage to evolving some sorts of structures more readily than others. Janson (1992), for example, showed that some transitions among adaptive syndromes are indeed more likely than others.
The hypotheses put forward in this paper do not, in fact, make a definite prediction of dispersal mode for each species. Rather, they generate probabilities, or options for dispersal modes that might be con- ceivable. These probabilities, when taken in aggregate for the list of species in a vegetation type, generate a definite prediction of a dispersal spectrum, and it is this predicted dispersal spectrum that is best suited to falsification. In a sense, then, the predictions from the present set of hypotheses are not about species from known lineages, but about cases of species occurring in a vegetation type and adopting different dispersal modes. The list of species assembled at a location is drawn over time from the whole of a regional flora, and this, we believe, makes it possible for a prediction about the dispersal spectrum to be made without ref- erence to constraints in the developmental pathway for fruit structures.
In order to increase the power of the set of hypoth- eses so that a definite prediction of dispersal mode could be made for each named species, it will probably be necessary to add further exclusion rules invoking developmental patterns of the fruit. Formal phylo- genetic analysis will be useful to identify situations where developmental constraints might be important (e.g. Bremer & Eriksson 1992; Herrera 1992), as well as traits such as seed size and plant stature.
PROVISIONAL CONCLUSIONS ABOUT THE
EVOLUTION OF DISPERSAL MODE
We hope that the set of hypotheses outlined above capture some of the important patterns relating dis- persal mode to other aspects of a plant's ecology. If we suppose for the sake of argument that these are indeed the most important patterns, the following biological conclusions emerge.
First, the availability of vectors rarely seems to limit dispersal modes adopted, at least when dispersal modes are aggregated at a broad level such as 'ver- tebrates'. Vectors such as wind are nearly ubiquitous. Fruit-eating birds probably become available in response to the supply of fruits, rather than vice versa (French & Westoby 1992). Seeds with elaiosomes are removed by many ant species, even in environments
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

942 Predicting dispersal spectra from plant attributes
where the plant species does not naturally occur (e.g. Pemberton 1988). Situations where vectors do limit dispersal mode are reflected in our hypothesis 1 (wind dispersal limited beneath a dense canopy), and hypothesis 7 (adhesive dispersal limited in species taller than 2 m).
Secondly, it was not found necessary to directly invoke attributes of the physical environment, with the exception of fire regime, in formulating the hypotheses. Most aspects of the physical environment are treated by the hypotheses as influencing plant stature and seed size, which in turn shape dispersal modes. If this is true, it suggests that different dis- persal modes are differentiated mainly by the dis- tances they achieve, rather than by their capacity to position seeds in particular microsites (with the excep- tion of the desirability of being buried, in environ- ments with a high fire frequency).
Thirdly, seed size is associated with dispersal mode, but the relationship is quite loose. Most seeds above c. 100mg are adapted for dispersal by vertebrates, presumably because other dispersal modes are less effective for these seeds, and because the high costs of pulp or arils represent a smaller proportional expense for larger seeds. Most seeds below c. 0.1 mg are unas- sisted, presumably because in this size range they are quite effectively dispersed by wind even without pos- sessing any special structures to increase air-resist- ance, and because the costs of dispersal adaptations would be high as a proportion of investment in the seed itself. However, in the central range of 0.1- 100mg occupied by most seeds (at least in the tem- perate zone), any dispersal mode is feasible.
Fourth, the repeated association of certain dis- persal modes with plant stature suggests that in some plants, there may be strong selection to be dispersed at least a distance in the order of 1-2 canopy diam- eters, with diminishing further advantage at greater distances. The fact that dispersal modes such as bal- listic and ants exist at all is evidence that for some plants dispersing 1-2 canopy diameters is adequate to pass genes on successfully. Theoretical models have established clearly that dispersal can provide a variety of fitness benefits even when a more favourable physi- cal environment is not reached (reviewed by Johnson & Gaines 1990). Seedlings will often be strongly dis- advantaged if they are close enough to suffer com- petition from the parent. Even if the parent is no longer alive at the time of seedling establishment, strong selection to achieve 1-2 canopy diameters could well be generated by competition among maternal-sib seedlings. We draw the tentative con- clusion that for many plants, the most important out- come achieved by having a particular dispersal struc- ture is a distance of 1-2 canopy diameters from the parent plant. The finding of Portnoy & Willson (1993) that there is little association between dispersal mode and the shape of the seed shadow tail supports the idea that if there are further advantages to be obtained
at distances greater than 1-2 canopy diameters, or in particular microsites, the chances of obtaining these other advantages are not greatly affected by the dis- persal mode adopted.
Finally, it is interesting to consider selection to achieve distances of 1-2 canopy diameters in relation to the high incidence of species with no apparent adaptation for dispersal at all; in most of the dispersal spectra surveyed by Willson et al. (1990) 20-50% of species had no morphological structures for dispersal and in some spectra this figure was 80-90%. Long seed shadows are generally considered to be advan- tageous for finding of safe sites in which to establish and for colonizing new habitats (e.g. Howe & Small- wood 1982; Green 1983). Why then do so many spec- ies apparently have no means to achieve any dispersal at all, let alone long distance dispersal? Willson (1993) discusses evidence for several arguments as to why so many species have no mechanism for long range seed dispersal. She concludes there is little evidence that phylogenetic constraints offer a likely explanation, and that there is no strong correlation between the distribution of safe sites and dispersal mode. Other explanations discussed by Willson include that, for some species, dispersal in time may be more important than dispersal in space, and that the development of dispersal structures may be too costly for some spec- ies, necessitating trade-offs with other components of fitness. This last argument is perhaps the most plaus- ible explanation. The abundance of species with no obvious dispersal mechanism implies that the benefits of adopting a structure assisting dispersal often does not outweigh the costs of foregone seed production. For these species, producing a larger number of seeds may have just as beneficial an effect on the numbers of seeds travelling further than a canopy diameter away, as would producing a smaller number of seeds each equipped with a structure assisting dispersal. The concept that the major fitness benefits of dispersal are achieved at rather modest distances, can be seen as complementary to the observation of Wheelwright & Orians (1982) that a plant does not have the means available to determine the fate of its seeds; even elab- orate dispersal structures have, at best, a modest and statistical influence on seed fate.
Acknowledgements
We are grateful to Peter Grubb, Rosemary Lott and Mary Willson for their comments on the manuscript. This research was been supported by the Australian Research Council, by an A.R.C Postdoctoral Research Fellowship (to L.H.) and by Australian Postgraduate Research Awards (to M.D. and M.R.L.).
References
Alvarez-Buylla, E.R. & Martinez-Ramos, M. (1990) Seed bank versus seed rain in the regeneration of a tropical pioneer tree. Oecologia, 84, 314-325.
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

943 L. Hughes et al.
Andersen, A.N. (1988) Dispersal distance as a benefit of myrmecochory. Qecologia, 75, 507-511.
Armbruster, W.S. (1982) Seed production and dispersal in Dalechampia (Euphorbiaceae): divergent patterns and ecological consequences. American Journal of Botany, 69, 1429-1440.
Armesto, J.J. & Rozzi, R. (1989) Seed dispersal syndromes in the rainforest of Chiloe: evidence for the importance of biotic dispersal in a temperate rainforest. Journal of Biogeography, 16, 219-226.
Ashton, P.S. (1989) Dipterocarp reproductive biology. Eco- systems of the world Vol. 14B. Tropical rainforest eco- systems: biogeographical and ecological studies (eds H. Lieth & M.J.A. Werger), pp. 219-240. Elsevier, Amster- dam.
Augspurger, C.K. (1986) Morphology and dispersal poten- tial of wind-dispersed diaspores of neotropical trees. American Journal of Botany, 73, 353-363.
Augspurger, C.K. & Hogan, K.P. (1983) Wind dispersal of fruits with variable seed number in a tropical tree (Lonchocarpus pentaphyllus: Leguminosae). American Journal of Botany, 70, 1031-1037.
Auld, T.D. (1986) Population dynamics of the shrub Acacia suaveolens (Sm.) Willd: fire and the transition to seed- lings. Australian Journal of Ecology, 11, 373-385.
Auld, T.D. & O'Connell, M.A. (1991) Predicting patterns of post-fire germination in 35 eastern Australian Fabaceae. Australian Journal of Ecology, 16, 53-70.
Ballardie, R.T. & Whelan, R.J. (1986) Masting, seed dis- persal and seed predation in the cycad Macrozamia com- munis. Oecologia, 70, 100-105.
Beattie, A.J. & Culver, D.C. (1981) The guild of myr- mecochores in the herbaceous flora of West Virginia forests. Ecology, 62, 107-115.
Beattie, A.J. & Lyons, N. (1975) Seed dispersal in Viola (Violaceae): adaptations and strategies. American Jour- nal of Botany, 62, 714-722.
Beer, T. & Swaine, M.D. (1977) On the theory of explosively dispersed seeds. New Phytologist, 78, 681-694.
Benzing, D.H. (1989) Vascular epiphytism in America. Eco- systems of the World, Vol. 14B. Tropical Rainforest Eco- systems. Biogeographical and Ecological Studies (eds H. Lieth & M.J.A. Werger), pp. 133-154. Elsevier, Amster- dam.
Benzing, D.H. & Clements, M.A. (1991) Dispersal of the orchid Dendrobium insigne by the ant Iridomyrmex cord- atus in Papua New Guinea. Biotropica, 23, 604-607.
Berg, R.Y. (1966) Seed dispersal of Dendromecon: its ecol- ogic, evolutionary and taxonomic significance. American Journal of Botany, 53, 61-73.
Berg, R.Y. (1975) Myrmecochorous plants in Australia and their dispersal by ants. Australian Journal of Botany, 23, 475-508.
Bond, W.J. (1985) Canopy-stored seed reserves (serotiny) in Cape Proteaceae. South African Journal of Botany, 51, 181-186.
Bond, W.J. (1988) Proteas as 'tumbleseeds': wind dispersal through air and over soil. South African Journal of Botany, 54, 455-460.
Bond, W.J. & Slingsby, P. (1983) Seed dispersal by ants in shrublands of the Cape Province and its evolutionary implications. South African Journal of Science, 79, 231- 233.
Bond, W.J. & Stock, W.D. (1989) The costs of leaving home: ants disperse myrmecochorous seeds to low nutrient sites. Qecologia, 81, 412-417.
Boyer, W.D. (1958) Longleaf pine seed dispersal in South Alabama. Journal of Forestry, 56, 265-268.
Bradstock, R.A. Auld, T.D. Ellis, M.E. & Cohn, J.S. (1992) Soil temperature during bushfires in semi-arid, mallee shrublands. Australian Journal of Ecology, 17, 433-440
Bremer, B. & Eriksson, 0. (1992) Evolution of fruit charac- ters and dispersal modes in the tropical family Rubi- aceae. Biological Journal of the Linnean Society, 47, 79- 95.
Bullock, S.H. (1974) Seed dispersal of Dendromnecon by the seed predator Pogonomyrmex. Madrono, 22, 378-379.
Bullock, S.H. & Primack, R.B. (1977) Comparative exper- imental study of seed dispersal on animals. Ecology, 58, 681-686.
Bulow-Olsen, A. (1984) Diplochory in Viola: a possible relation between seed dispersal and soil seed bank. American Midland Naturalist, 112, 251-260.
Chapman, L.J. Chapman, C.A. & Wrangham, R.W. (1992) Balanites wilsoniana: elephant dependent dispersal? Journal of Tropical Ecology, 8, 275-283.
Cheplick, G.P. & Quinn, J.A. (1987) The role of seed depth, litter, and fire in the seedling establishment of amphi- carpic peanutgrass (Amphicarpum purshii). Oecologia, 73, 459-464.
Christensen, P.E. & Kimber, P.C. (1975) Effect of prescribed burning on the flora and fauna of south-west Australian forests. Proceedings of the Ecological Society of Australia, 9, 85-106.
Coates-Estrada, R. & Estrada, A. (1988) Frugivory and seed dispersal in Cymbopetalum bailloni (Annonaceae) at Los Tuxtlas, Mexico. Journal of Tropical Ecology, 4, 157-172.
Corner, E.J.H. (1949) The durian theory or the origin of the modern tree. Annals of Botany, 13, 367-414.
Cremer, K.W. (1966) Dissemination of seed from Eucalyptus regnans. Australian Forestry, 30, 33-37.
Cremer, K.W. (1977) Distance of seed dispersal in Eucalypts estimated from seed weights. Australian Forest Research, 7, 225-228.
Culver, D.C. & Beattie, A.J. (1978) Myrmecochory in Viola: dynamics of seed-ant interactions in some west Virginia species. Journal of Ecology, 66, 53-72.
Davidar, P. (1987) Fruit structure in two neotropical mistle- toes and its consequences for seed dispersal. Biotropica, 19, 137-139.
Davidson, D.W. (1988) Ecological studies of neotropical ant gardens. Ecology, 69, 1138-1152.
Davidson, D.W. & Morton, S.R. (1981) Myrmecochory in some plants (F. Chenopodiaceae) of the Australian arid zone. Oecologia, 50, 357-366.
Dunn, P.H. & DeBano, L.F. (1977) Fire's effect on biological and chemical properties of chaparral soil. Proceedings of the Symposium on the Environmental Consequences of Fire and Fuel Management in Mediterranean Ecosystems (eds H.A. Mooney & C.E. Conrad), pp. 75-94. USDA Forest Service Technical Report WO-3.
Estrada, A. & Coates-Estrada, R. (1986) Frugivory in how- ling monkeys (Alouattapalliata) at Los Tuxtlas, Mexico: dispersal and fate of seeds. Frugivores and Seed Dispersal (eds A. Estrada & T.H. Fleming), pp. 93-104. Dr W. Junk, Dordrecht.
Evans, R.A. Biswell, H.H. & Palmquist, D.E. (1987) Seed dispersal in Ceanothus cuneatus and C. leucodermis in a sierran oak-woodland savanna. Madrono, 34, 283-293.
Ewel, J. Berish, C. Brown, B. Price, N. & Raich, J. (1981) Slash and burn impacts on a Costa Rican wet forest site. Ecology, 62, 816-829.
Fenner, M. (1987) Seed characteristics in relation to succession. Colonization, Succession and Stability (eds A.J. Gray, M.J. Crawley & P.J. Edwards), pp. 103-114. Blackwell Scientific Publications, Oxford.
Fleming, T.H. (1981) Fecundity, fruiting pattern, and seed dispersal in Piper amalago (Piperaceae), a bat-dispersed tropical shrub. Qecologia, 51, 42-46.
Floyd, A.G. (1966) Effect of fire upon weed seeds in the
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

944 Predicting dispersal spectra from plant attributes
wet sclerophyll forests of northern N.S.W. Australian Journal of Botany, 14, 243-256.
Frankie, G.W. Baker, H.G. & Opler, P.A. (1974) Com- parative phenological studies of trees in tropical wet and dry forests in the lowlands of Costa Rica. Journal of Ecology, 62, 881-919.
Frantzen, N.M.L.H.F. & Bouman, F. (1989) Dispersal and growth form patterns of some zonal paramo vegetation types. Acta Botanica Neerlandica, 38, 449-465.
French, K. & Westoby, M. (1992) Removal of vertebrate- dispersed fruits in vegetation on fertile and infertile soils. Oecologia, 91, 447-454.
Fridriksson, S. (1975) Surtsey. Evolution of Life on a Volcanic Island. Butterworths, London.
Gashwiler, J.S. (1969) Seed fall of three conifers in west- central Oregon. Forest Science, 15, 290-295.
Gentry, A.H. (1982) Patterns of neotropical plant species diversity. Evolutionary Biology, 15, 1-84.
Gill, A.M. (1985) Acacia cyclops G. Don (Leguminosae- Mimosaceae) in Australia: distribution and dispersal. Journal of the Royal Society of Western Australia, 67, 59-65.
Grace, J. (1977) Plant Response to Wind. Academic Press, London.
Green, D.S. (1983) The efficacy of dispersal in relation to safe site density. Oecologia, 56, 356-358.
Greene, D.F. & Johnson, E.A. (1989) A model of wind dispersal of winged or plumed seeds. Ecology, 70, 339- 347.
Grime, J.P. Hodgson, J.G. & Hunt, R. (1988) Comparative Plant Ecology. a Functional Approach to Common British Species. Unwin Hyman, London.
Guitian, J. & Sanchez, J.M. (1992) Seed dispersal spectra of plant communities in the Iberian Peninsula. Vegetatio, 98, 157-164.
Hamilton, W.D. & May, R.M. (1977) Dispersal in stable habitats. Nature, 269, 578-581.
Handel, S.N. Fisch, S.B. & Schatz, G.E. (1981) Ants disperse a majority of herbs in a mesic forest community in New York State. Bulletin of the Torrey Botanical Club, 108, 130-137.
Harper, J.L. (1977) The Population Biology of Plants. Aca- demic Press, London.
Harper, J.L. Lovell, P.H. & Moore, K.G. (1970) The shapes and sizes of seeds. Annual Review of Ecology and Sys- tematics, 1, 327-357.
Harvey, P.H. & Pagel, M.D. (1991) The Comparative Method in Evolutionary Biology. Oxford University Press, Oxford.
Hawksworth, F.G. (1959) Ballistics of dwarf mistletoe seeds. Science, 130, 504.
Herrera, C.M. (1992) Interspecific variation in fruit shape: allometry, phylogeny, and adaptation to dispersal agents. Ecology, 73, 1832-1841.
Herrera, J. (1991) Herbivory, seed dispersal, and the dis- tribution of a ruderal plant living in a natural habitat. Oikos, 62, 209-215.
Higashi, S. Tsuyuzaki, S. Ohara, M. & Ito, F. (1989) Adapt- ive advantages of ant-dispersed seeds in the myr- mecochorous plant Trillium tschonoskii (Liliaceae). Oikos, 54, 389-394.
Hinds, T.E. Hawksworth, F.G. & McGinnies, W.J. (1968) Seed discharge in Arceuthobium: a photographic study. Science, 160, 1236-1238.
Hobbs, R.J. & Atkins, L. (1988) Spatial variability of exper- imental fires in south-west Western Australia. Australian Journal of Ec ology, 13, 295-299.
Hodgson, J.G. & Grime, J.P. (1990) The role of dispersal mechanisms, regenerative strategies and seed banks in the vegetation dynamics of the British landscape. Spec ies
Dispersal in Agricultural Habitats (eds R.G.H. Bunce & D.C. Howard), pp. 65-81. Belhaven Press, London.
Hoffmann, A.J. Teillier, S. & Fuentes, E.R. (1989) Fruit and seed characteristics of woody species in mediterranean- type regions of Chile and California. Revista Chilena de Historia Natural, 62, 43-60.
Honig, M.A. Cowling, R.M. & Richardson, D.M. (1992) The invasive potential of Australian banksias in South African fynbos: a comparison of the reproductive poten- tial of Banksia ericifolia and Leucodendron laureolum. Australian Journal of Ecology, 17, 2305-314.
Hoppes, W.G. (1988) Seedfall patterns of several species of bird-dispersed plants in an Illinois woodland. Ecology, 69, 320-329.
Horvitz, C.C. & Beattie, A.J. (1980) Ant dispersal of Cal- athea (Marantaceae) seeds by carnivorous ponerines (Formicidae) in a tropical rainforest. American Journal of Botany, 67, 321-326.
Horvitz, C.C. & Schemske, D.W. (1986) Seed dispersal of a neotropical myrmecochore: variation in removal rates and dispersal distance. Biotropica, 18, 319-323.
Houssard, C. Escarre, J. & Romane, F. (1980) Development of species diversity in some Mediterranean plant com- munities. Vegetatio, 43, 59-72.
Howe, H.F. & Smallwood, J. (1982) Ecology of seed disper- sal. Annual Review of Ecology and Systematics, 13, 201- 228.
Hughes, L. & Westoby, M. (1992) Fate of seeds adapted for dispersal by ants in Australian sclerophyll vegetation. Ecology, 73, 1285-1299.
Janson, C.H. (1992) Measuring evolutionary constraints: a Markov model for phylogenetic transitions among seed dispersal syndromes. Evolution, 46, 136-158.
Janzen, D.J. (1974) Epiphytic myrmecophytes in Sarawak; mutualism through feeding of plants by ants. Biotropica, 6, 69-103.
Janzen, D. (1984) Dispersal of small seeds by big herbivores: foliage is the fruit. American Naturalist, 123, 338-353.
Johnson, M.L. & Gaines, M.S. (1990) Evolution of dispersal: theoretical models and empirical tests using birds and mammals. Annual Review of Ecology and Systematics, 21, 449-480.
Jones, E.W. (1956) Ecological studies on the rainforest of southern Nigeria. Journal of Ecology, 44, 83-117.
Jordano, P. (1992) Fruits and frugivory. Seeds. the Ecology of Regeneration in Plant Communities (ed. M. Fenner), pp. 105-156.C.A.B. International, Wallingford.
Jurado, E. Westoby, M. & Nelson, D. (1991) Diaspore weight, dispersal, growth form and perenniality of cen- tral Australian plants. Journal of Ecology, 79, 811-830.
Kaufmann, S. McKey, D.B. Hossaert-McKey, M. & Horv- itz, C.C. (1991) Adaptations for a two-phase seed dis- persal system involving vertebrates and ants in a hem- iepiphytic fig (Ficus microcarpa: Moraceae). American Journal of Botany, 78, 971-977.
Keay, R.W.J. (1957) Wind-dispersed species in a Nigerian forest. Journal of Ecology, 45, 471-478.
Keeley, J.E. (1991) Seed germination and life history syn- dromes in the California chaparral. Botanical Review, 57, 81-116.
Keeley, J.E. & Zedler, P.H. (1978) Reproduction of chap- arral shrubs after fire: a comparison of sprouting and seeding strategies. American Midland Naturalist, 99, 142-161.
Kjellsson, G. (1985) Seed fate in a population of CarexY
pilulifera L. I Seed dispersal and ant-seed mutualism. Qecologia, 67, 416-423.
Kleinfeldt, S.E. (1978) Ant-gardens: the interaction of Codonanthe c rassifolia (Gesneriaceae) and Cre- matogaster longispina (Formicidae). Ecology, 59, 449- 456.
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

945 L. Hughes et al.
Kleinfeldt, S. (1986) Ant-gardens: mutual exploitation. Insects and the Plant Surface (eds B. Juniper & T.R.E. Southwood), pp. 283-294. Edward Arnold, London.
Klinkhamer, P.G.L., De Jong, T.J. & van der Meijden, E. (1988) Production, dispersal and predation of seeds in the biennial Cirsium vulgare. Journal of Ecology, 76, 403-414.
Kohlerman, V.L. (1950) Untersuchungen uber die Wind- verbreitung der Fruchte und Samen mitteleuropaisher Waldbaume. Forstwissenschaftliches Zentralblatt, 69, 606-624.
Lamont, B. (1985) Dispersal of the winged fruits of Nuytsia floribunda (Loranthaceae). Australian Journal of Ecol- ogy, 10, 187-193.
Lamont, B.B. Le Maitre, D.C. Cowling, R.M. & Enright, N.J. (1991) Canopy seed storage in woody plants. Bot- anical Review, 57, 277-317.
Leishman, M.R. & Westoby, M. (1992) Classifying plants into groups on the basis of association of individual traits - evidence from Australian semi-arid woodlands. Journal of Ecology, 80, 417-424.
Levin, D.A. & Kerster, H.W. (1968) Local gene dispersal in Phlox. Evolution, 22, 130-139.
Liddle, M.J. & Elgar, M.A. (1984) Multiple pathways in diaspore dispersal, exemplified by studies of Noogoora Burr (Xanthium occidentale Bertol. Compositae). Bot- anical Journal of the Linnean Society, 88, 303-315.
Liddy, J. (1983) Dispersal of Australian mistletoes: the Cowiebank study. The Biology of Mistletoes (eds M. Calder & P. Bernhardt), pp. 101-116. Academic Press, Sydney.
Madison, M. (1977) Vascular epiphytes: their systematic occurrence and salient features. Selbyana, 2, 1-13.
Madison, M. (1979) Additional observations on ant-gardens in Amazonas. Selbyana, 5, 107-115.
Matlack, G.R. (1987) Diaspore size, shape, and fall behavior in wind-dispersed plant species. American Journal of Bot- any, 74,1150-1160.
McCartney, H.A. (1990) Dispersal mechanisms through the air. Species Dispersal in Agricultural Habitats (eds R.G.H. Bunce & D.C. Howard), pp. 133-158. Belhaven Press, London.
McClanahan, T.R. & Wolfe, R.W. (1987) Dispersal of orni- thochorous seeds from forest edges in central Florida. Vegetatio, 71, 107-112.
McDonnell, M.J. & Stiles, E.W. (1983) The structural com- plexity of old field vegetation and the recruitment of bird-dispersed plant species. Oecologia, 56, 109-116.
Mesler, M.R. & Lu, K.L. (1983) Seed dispersal of Trillium ovatum (Liliaceae) in second-growth redwood forests. American Journal of Botany, 70, 1460-1467.
Milewski, A.V. & Bond, W.J. (1982) Convergence of myr- mecochory in mediterranean Australia and South Africa. Ant-Plant Interactions in Australia (ed. R.C. Buckley), pp. 89-98. Dr W. Junk, The Hague.
Milton, S.J. Siegfried, W.R. & Dean, W.R.J. (1990) The distribution of epizoochoric plant species: a clue to the prehistoric use of arid Karoo rangelands by large her- bivores. Journal of Biogeography, 17, 25-34.
Motro, U. (1982) Optimal rates of dispersal II. Diploid populations. Theoretical Population Biology, 21, 412- 429.
Murray, K.G. (1988) Avian seed dispersal of three neo- tropical gap-dependent plants. Ecological Monographs, 58, 271-298.
O'Dowd, D.J. & Hay, M.E. (1980) Mutualism between har- vester ants and a desert ephemeral: seed escape from rodents. Ecology, 61, 531-540.
Oke, T. R. (1987) Boundary Layer Climates, 2nd edn. Methuen, London.
Oliver, H.R. (1971) Wind profiles in and above a forest
canopy. Quarterly Journal of the Royal Meteorological Society, 97, 548-553.
Pemberton, R.W. (1988) Myrmecochory in the introduced range weed, leafy spurge (Euphorbia esula L. ). American Midland Naturalist, 119, 431-435.
van der Pijl, L. (1982) Principles of dispersal in higher plants. Springer-Verlag, Berlin.
Plitmann, U. (1986) Alternative modes in dispersal stra- tegies, with an emphasis on herbaceous plants of the Middle East. Proceedings of the Royal Society of Edin- burgh, 89B, 193-202.
Portnoy, S. & Willson, M.F. (1993) Seed dispersal curves: behavior of the tail of the distribution. Evolutionary Ecology, 7, 25-44.
Pudlo, R.J. Beattie, A.J. & Culver, D.C. (1980) Population consequences of changes in an ant-seed mutualism in Sanguinaria canadensis. Oecologia, 146, 32-37.
Reid, N. (1991) Coevolution of mistletoes and frugivorous birds? Australian Journal of Ecology, 16, 457-470.
Ridley, H.N. (1930) The Dispersal of Plants Throughout the World. Ashford, Reeve.
Roberts, J.T. & Heithaus, E.R. (1986) Ants rearrange the vertebrate-generated seed shadow of a Neotropical figtree. Ecology, 67, 1046-1051.
Roth, I. (1977) The fruits of Angiosperms. Encyclopedia of Plant Anatomy, pp. 572-579. Gebruder Borntraeger, Berlin.
Sacchi, C.F. (1987) Variability in dispersal ability of com- mon milkweed Asclepias syriaca. Oikos, 49, 191-198.
Schemske, D.W. (1978) Evolution and reproductive charac- teristics in Impatiens (Balsaminaceae): the significance of cleistogamy and chasmogamy. Ecology, 59, 596-613.
Scotter, D.R. (1970) Soil temperatures under grass fires. Australian Journal of Forest Research, 8, 273-279.
Shea, S.R. McCormick, J. & Portlock, C.C. (1979) The effect of fires on regeneration of leguminous species in the northern jarrah (Eucalyptus marginata Sm.) forest of Western Australia. Australian Journal of Ecology, 4, 195-205.
Sheldon, J.C. & Burrows, F.M. (1973) The dispersal effec- tiveness of the achene-pappus units of selected Com- positae in steady winds with convection. New Phytol- ogist, 72, 665-675.
Shmida, A. & Ellner, S. (1983) Seed dispersal on pastoral grazers in open Mediterranean chaparral, Israel. Israeli Journal of Botany, 32, 147-159.
Slingsby, P. & Bond, W.J. (1985) The influence of ants on the dispersal distance and seedling recruitment of Leu- cospermum conocarpodendron (L.) Buek (Proteaceae). South African Journal of Botany, 51, 30-34.
Smith, A.J. (1975) Invasion and ecesis of bird-disseminated woody plants in a temperate forest sere. Ecology, 56, 19-34.
Smith, J.M.B. (1989) An example of ant-assisted plant invasion. Australian Journal of Ecology, 14, 247-250.
Sorensen, A.E. (1986) Seed dispersal by adhesion. Annual Review of Ecology and Systematics, 17, 443-463.
Stamp, N.E. (1989a) Efficacy of explosive vs hygroscopic seed dispersal by an annual grass species. American Jour- nal of Botany, 76, 555-561.
Stamp, N.E. (1989b) Seed dispersal of four sympatric grass- land annual species of Erodium. Journal of Ecology, 77, 1005-1020.
Stamp, N.E. & Lucas, J.R. (1983) Ecological correlates of explosive seed dispersal. Oecologia, 59, 272-278.
Stamp, N.E. & Lucas, J.R. (1990) Spatial patterns and dis- persal distances of explosively dispersing plants in Flo- rida sandhill vegetation. Journal of Ecology, 78, 589- 600.
Stebbins, G.L. (1971) Adaptive radiation of reproductive
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

946 Predic ting dispersal spectra from plant attributes
characteristics in Angiosperms, II. Seeds and seedlings. Annual Review of Ecology and Svstematics, 2, 237-260.
Stiles, E.W. (1992) Animals as seed dispersers. Seeds: the Ecology of Regeneration in Plant Communities (ed. M. Fenner), pp. 87-104. C.A.B. International, Wallingford.
Stott, P. (1986) The spatial pattern of dry season fires in the savanna forests of Thailand. Journal of Biogeography, 13, 345-358.
Swaine, M.D. & Beer, T. (1977) Explosive seed dispersal in Hura crepitans L. (Euphorbiaceae). New Phytologist, 78, 695-708.
Taylor, B.W. (1954) An example of long distance dispersal. Ecology, 35, 569-572.
Thomas, D.W. Cloutier, D. & Provencher, M. (1988) The shape of bird- and bat-generated seed shadows around a tropical fruiting tree. Biotropica, 20, 347-348.
Thompson, K. Band, S.R. & Hodgson, J.G. (1993) Seed size and shape predict persistence in soil. Functional Ecology, 7, 236-241.
Tiffney, B.H. (1986) Evolution of seed dispersal syndromes according to the fossil record. Seed Dispersal (ed. D.R. Murray), pp. 273-305. Academic Press, Sydney.
Tomback, D.F. & Linhart, Y.B. (1990) The evolution of bird-dispersed plants. Evolutionary Ecology, 4, 185-219.
van Tooren, B.F. (1988) The fate of seeds after dispersal in chalk grassland: the role of the bryophyte layer. Oikos, 53, 41-48.
Venable, D.L. & Brown, J.S. (1993) The population- dynamic functions of seed dispersal. Vegetatio, 107/108, 31-55.
Venable, D.L. & Levin, D.A. (1983) Morphological dispersal structures in relation to growth habit in the Compositae. Plant Systematics and Evolution, 143, 1-16.
Walker, J. (1981) Fuel dynamics in Australian vegetation. Fire and the Australian Biota (eds A.M. Gill, R.H. Groves & I.R. Noble), pp. 101-127. Australian Acad- emy of Sciences, Canberra.
Webber, M.L. (1934) Fruit dispersal. Malayan Forester, 3, 18-19.
Weir, J.S. & Kiew, R. (1986) A reassessment of the relations in Malaysia between ants (Crematogaster) on trees (Lep-
tospernmuni and Dacryidiumn) and epiphytes of the genus Dischidia (Asclepiadaceae). Biological Journal oft/ie Lin- nean Societ ', 27, 1.
Westoby, M. Jurado, E. & Leishman, M. (1992) Com- parative evolutionary ecology of seed size. Trends in Ecology and Evolution, 7, 368-372.
Westoby, M. Rice, B. & Howell, J. (1990) Seed size and plant growth form as factors in dispersal spectra. Ecology, 71, 1307-1315.
Westoby, M. Rice, B. Shelley, J.M. Haig, D. & Kohen, J.L. (1982) Plants' use of ants for dispersal at West Head, New South Wales. Ant-plant interactions in Australia (ed. R.C. Buckley), pp. 75-87. Dr W. Junk, The Hague.
Wheelwright, N.T. & Orians, G.H. (1982) Seed dispersal by animals: contrasts with pollen dispersal, problems of terminology, and constraints on evolution. American Naturalist, 119, 402-413.
Wikander, T. (1984) Mecanismos de Dispersion de diasporus de una Selva Decidua en Venezuela. Biotropica, 16, 276- 283.
Willson, M.F. (1983) Plant Reproductive Ecology. John Wiley and Sons, New York.
Willson, M.F. (1989) Gut retention times of experimental pseudoseeds by emus. Biotropica, 21, 210-213.
Willson, M.F. (1991) Dispersal of seeds by frugivorous ani- mals in temperate forests. Revista Chilena de Historia Natural, 64, 537-554.
Willson, M.F. (1992) The ecology of seed dispersal. Seeds: the Ecology of Regeneration in Plant Communities (ed. M. Fenner), pp. 61-85. C.A.B. International, Wall- ingford.
Willson, M.F. Dispersal mode, seed shadows, and colon- ization patterns. Vegetatio, 107/108, 261-280.
Willson, M.F. Rice, B.L. & Westoby, M. (1990) Seed dis- persal spectra: a comparison of temperate plant com- munities. Journal of Vegetation Science, 1, 547-562.
Woodall, S.L. (1982) Seed dispersal in Melaleuca quin- quenervia. Florida Scientist, 45, 81-93.
Received 28 April 1993 Revised version accepted 20 May 1994
Appendix
Table Al Distribution of wind-dispersed species in relation to growth form and vegetation strata. Table is in part derived from Table 2, Howe & Smallwood (1982)
% wind dispersed species Forest type & Location in different strata/growth forms Reference
Deciduous forest, Indiana trees 29%, shrubs 0% Potzger et al. (1942 cited in Howe & Smallwood 1982)
Deciduous forest, Indiana trees 38%, shrubs 0% Keller (1946 cited in Howe & Smallwood 1982)
Rainforest, southern Nigeria canopy trees 46%, lower strata 7% Jones (1956) Wet forest, Nigeria emergent trees 56%, canopy trees 25%, Keay (1957)
subcanopy trees 2%, shrubs 0%; canopy lianas 48%, subcanopy lianas 0%
Wet forest, Costa Rica canopy trees 11 %, subcanopy trees 2% Frankie et al. (1974) Moist forest, Barro Colorado Is, canopy trees 16%, subcanopy trees 5% Gentry (1982)
Panama Dry forest, Costa Rica canopy trees 29%, subcanopy trees 8% Gentry (1982) Deciduous forest, Venezuela upper strata (3-10 m) 52%, middle strata (3 m) Wikander (1984)
15%, lower strata (0.3 m) 0% Rainforest, Chiloe, Chile emergent trees 86%, subcanopy trees 8%, Armesto & Rozzi (1989)
shrubs 14% Wet sclerophyll forest, Sydney, trees 25%, shrubs 10% Westoby et al. (1990)
Australia
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

947 L. Hughes et al.
Table A2 Distances that seeds are dispersed by ants
Species Mean distance (except where stated) Reference
Acacia cyclops up to 2m Gill (1985) A. pulchella 1.9 m (Iridomyrmex sp.) Shea et al. (1979) A. suaveolens 2.0 m (Rhytidoponera tasmaniensis, n = 87); 6.5 m Andersen (1988)
(Camponotus intrepidus, n = 3); 1.2 m (Iridomyrmex 'bicknelli', n = 6); overall mean 2.1 m, max. < 11 m
A. suaveolens 2.2m ('Pheidole sp. A', n = 40); 1 m (Pheidole sp.B, Auld (1986) n = 130)
1.3 m (Iridomyrmex sp. n = 20, max. 5 m) A. terminalis 1.1 m (n = 64, various spp, range 0.2-1.9m) Hughes & Westoby (1992) Calathea ovandensis 0.8 m (Pachycondyla harpax, max. 2.5 m); 9.3 m (F. Horvitz & Schemske (1986)
apicalis max. 20.5 m); 0.08 m (Solenopsis geminata, max. 0.4 m); 0.04 m ( Wasmannia auropunctata, max. 0.07 m), 51% seed moved > 0.5m, the average radius of the plant canopy)
C. ovandensis range 0.1-10.4m (Pachycondyla harpax, Horvitz & Beattie (1980) Odontomachus laticeps, 0. minutus, n = 55)
Carex pilulifera up to 2 m (Myrmica ruginodis) Kjellsson (1985) Datura discolor 2-3 m, up to 39 m (Pogonomyrmex californicus and O'Dowd & Hay (1980)
Veromessor pergandei) Dillwyniajuniperina 1.1 m (various spp. n = 133, range 0.06-11 m) Hughes & Westoby (1992) Euphorbia esula 2-3 m (n = 28), 6-8 m (n = 15), 25 m (Formica Pemberton (1988)
obscuripes,n = 1) Hovea rosmarinifolia rarely more than 1 m (Pheidole spp.) Berg (1975) Leucospermum 2.5 m between seed depots and emerging seedlings, Slingsby & Bond (1985)
conocarpodendron range 0.3-9.8 m (Pheidole capensis & Anoplolepis steingroeveri)
Leucospermum spp. rarely exceeded 2-3 m, max. 17 m Bond & Slingsby (1983) Sanguinaria canadense 0.17-3.1 m depending on site (Formica subsericea, Pudlo et al. (1980)
Aphaenogaster rudis and Lasius alienus) Sclerolaena diacantha max: 4m (Rhytidoponera sp.B); 4m (Pheidole sp.B) Davidson & Morton (1981)
77 m (Iridomyrmex sp. D) Trillium ovatum seldom more than 1 m (Lasius pallitarsus) Mesler & Lu (1983) T. tschonoskii 0.64m (up to 2.7m) (Myrmica ruginodis and Higashi et al. (1989)
Aphaenogasterjaponica) Viola odorata and up to 40-50 m (Iridomyrmex purpureus) Smith (1989)
Grevillea rosmarinifolia Viola spp. 0.35-0.72m (max. 1.5m) Culver & Beattie (1978)
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

948 Predicting dispersal spectra firom plant attributes
Table A3 Distances seeds are dispersed by ballistic expulsion
Species Mean distance (except where stated) Reference
Amphicarpaea bracteata 31% aerial chasmogamous seeds and 25% Trapp (1988) cleistogamous seeds > 2 m (estimated from initial velocity, pod height and firing angle)
Arceuthobium vaginalum 5.3 m (max. 12.8 m) Hawksworth (1959) ssp. cryptopodium
Bauhinia purpurea up to 15 m Stebbins (1971) Ceanothus spp. less than 2% > 9m Evans et al. (1987) Cnidoscolus stimulosus 0.63 m (max. 1.15 m) Stamp & Lucas (1990) Crotalaria rotundifolia 0.94m (max. 2.4m) Stamp & Lucas (1990) Dalechampia (15 spp.) range 1-6 m Armbruster (1982) Dendromecon harfordii 3.6m (max. 5.1 m, n = 14) Berg (1966) D. rigida 1.3 m (max. 2.9 m, seeds placed 15-30cm high, Berg (1966)
n = 64) 2.7 m (max. 4.8 m, 90 cm high, n = 88) Ecballium elaterium range 1-16 m Pitmann (1986) Erodium moschatum 56cm (range 0-86cm, n = 181) Stamp (1989a) Erodium (4 spp.) 0.54-0.76 m (n = 58-60 depending on species, Stamp (1989b)
measured in lab. 0.34m in presence of neighbours) Geranium purpureum range 0-160cm (50% < 44cm, 35% within canopy of Herrera (1991)
parent) Hura crepitans max. 45 m Swaine & Beer (1977) Lupinus texensis 0.6m (n = 263, max. 3.4m) Schaal (1980) Impatiens biflora and I. up to 2m Schemske (1978)
pallida I. capensis 1.5-2 m Waller (1980) Papaver somniferum up to 15 m Ulbrich (1928); in van der Pijl
(1972) Phlox pilosa 1.12m (n = 395, max. 3.6m) Levin & Kerster (1968) Stillingia sylvatica 1.08 m (max. 3.0 m) Stamp & Lucas (1990) Thlaspi perfoliatum 0.8 m Muller (1955); in van der Pijl
(1972) Viola blanda, V. /anceolata 2-3 m (in glasshouse) Bulow-Olsen (1984)
and V. fimbriatula Viola spp. up to 5.4m (in glasshouse) Culver & Beattie (1978) Viola spp. 0.02-5.1 m Beattie & Lyons (1975) herbs, 6 spp. mode 0.2-3.4m (few > 4m) Stamp & Lucas (1983)
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

949 L. Hughes et al.
Table A4 Distances seeds are dispersed by wind
Species Mean distance (except where stated) Reference
Abies alba 72m (from 14m height in 5.9ms-' wind, n = 200) Kohlermann (1950) Abies sp. up to 7 km Muller (1955); in van der Pijl
(1972) Abietinae spp. 40-800 m van der Pijl (1972) Acer platanoides, 83 m (from 14 m height in 5.9 m s-' wind, n = 200) Kohlermann (1950) A. pseudoplatanus 69 m (from 14 m height in 5.9 m s-' wind, n = 200) Kohlermann (1950) Acer sp. up to 4km Muller (1955); in van der Pijl
(1972) A. syriaca 9.8 m (10.4 km h-' wind); 15.4 m (16.4 km h-' wind) Sacchi (1987) Aster tripolium up to 5 km Feekes (1936); in van der Pijl
(1972) Betula sp. up to 1.6 km Muller (1955); in van der Pijl
(1972) Carpinus betulus 73 m (from 14 m height in 5.9 m s-' wind, n = 200) Kohlermann (1950) Cirsium vulgare 50% < 1 m, 66% < 2m, 10% > 32m (n = 256) Klinkhamer et al. (1988) Compositae (18 spp.) estimates: 0.5-3.8m (in 5.5 kmh-' wind), 1.0-7.6m Sheldon & Burrows (1973)
(all < 90cm tall) (10.9 km h-'), 1.5-11.6 (16.4km/h), calculated from fall velocity in still air
Eucalyptus (15 spp.) estimates: 20.1-52.9 m (calculated from fall velocity Cremer (1977) using 40 m release height and 10 km h-' wind)
Fraxinus excelsior up to 0.5 km Muller (1955); in van der Pijl (1972)
Fraxinus excelsior 53 m (from 14 m height in 5.9 m s-' wind, n = 200) Kohlerman (1950) Larix decidua 29 m (from 14 m height in 1.7 m s-' wind, n = 50) Kohlerman (1950) Nuytsiafloribunda mode 5-7.5 m (93% beyond canopy, 63% 2.5-1 Om, Lamont (1985)
max. 37.5 m, n = 990); hand released: 9.9 m (5 m s-' wind), 5.9 m (9.5 m s-' wind, n = 25)
Orchidae spp. blown to Florida from West Indies van der Pijl (1972) Picea excelsa 35 m (from 14 m height in 1.7 m s-' wind, n = 50) Kohlerman (1950) Pinus contorta mode 0.75-1.83 m Greene & Johnson (1989) P. palustris 66% under canopy, 23% < 20m, 98% < 90m Boyer (1958) P. sylvestris 39m (from 14m height in 1.7ms-' wind, n = 50) Kohlerman (1950) P. strobus 46m (from 14m height in 2.7ms-' wind, n = 50) Kohlerman (1950) Platypodium elegans 45% 0-10m, 34% 10-20m, 21% 50-60m (n = 6550, Augspurger (1986)
two trees, double and single seeds combined) Populus sp. up to 30 km van der Pijl (1972) Protea repens max. 31 m (primary dispersal); max. 500m (blowing Bond (1988)
along ground, 50% > 20m, 20% > l00m) Pseudotsuga menziesii 61% 15-30 m, 26% 61-76 m, 13 % 107-122 m Gashwiler (1969) Senecio congestus up to 200 km van der Pijl (1972) S.jacobea 60% at base of parent, 0.4% > 4.6 m, Poole & Cairns (1940); in Harper
0.1% > 9 m, 0.02% > 18 m, 0.0005% > 36 m (1977) Thuja plicata 79% 15-30 m, 17% 61-76 m, 4% 107-122 m Gashwiler (1969) Tilia grandifolia 58 m (from 14m height in 5.9 m s-' wind, n = 200) Kohlerman (1950) T. parvifolia 64 m (from 14 m height in 5.9 m s-' wind, n = 200) Kohlerman (1950) Tsuga heterophylla 67% 15-30 m, 22% 61-76 m, 11 % 107-122 m Gashwiler (1969) Ulmus campestris 60m (from 14m height in 2.7ms-' wind, n = 50) Kohlerman (1950) 38 spp. herbs, estimate: 0.75-112 m in I0 km h-' wind Matlack (1987) 34 spp. tropical trees on estimate: 22-194m from fall velocity in still air Augspurger (1986)
Barro Colorado Is.
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions

950 Predicting dispersal spectra from plant attributes
Table A5 Distances seeds with fleshy fruits are dispersed by vertebrates
Species Distance Reference
Andira inermis max. 207m (under bat roosts) Janzen et al. (1976) Cecropia obtusifolia max. 86 m Alvarez-Buylla & Martinez-Ramos
(1990) Coprosma pumila 650 km (to Macquarie Is) Taylor (1954) Cymbopetalum baillonii 31% dropped under canopy, 8% 10-19 m, 27% 20- Coates-Estrada & Estrada (1988)
29m, 9% > 30m (n = 1O trees) Macrozamia communis 3.5-9.6 m (depending on pop. Trichosurus vulpecula, Ballardie & Whelan (1986)
max. 25 m = limit of search) M. riedlei 28% < 0.5m, 6% > 5m, 1.5% > lOm, up to 24m, Burbidge & Whelan (1982)
(most by Trichosurus vulpecula) Muntingia calabura max. 15 m ( = max. trap distance) Thomas et al. (1988) Piper amalago more than 90% > 50m, some > 300m (under bat Fleming (1981)
roosts, estimated from GPR and behavioural observations)
Phytolacca rivinoides, median 35-60 m, 12-36% < 30 m, up to 510 m, Murray (1988) Witheringia solanaceae, (estimated from GPR and observations of bird and W. coccoloboides movement distances
Prunus serotina max. 25 m Smith (1975) various spp. East USA old
field near forest max. at least 160m (most near forest edge) McDonnell & Stiles (1983)
7 spp. in Illinois max. 9-33 m (33 m = max. trap distance) Hoppes (1988) various spp. of wetland max. 120 m (= max. trap distance) McClanahan & Wolfe (1987)
forest plants various spp. on Surtsey, over 1000 km (birds such as the snow bunting) Fridriksson (1975); in Moore
volanic island, off Iceland (1987) various spp. means 94-262 m (range 10-811 m, estimated from Estrada & Coates-Estrada (1986)
GPR and observations of movements of howling monkeys)
TableA6 Seed size distributions for six dispersal modes compiled from four temperate floras: Sydney, NSW (n = 307 spp.; Westoby et al. 1990), Central Australia (n = 196 spp.; Jurado et al. 1991), western NSW (n = 248 spp.; Leishman & Westoby 1992), and Sheffield, UK (n = 271 spp.; Grime et al. 1988). Data from which Fig. 1 is drawn
Seed size class (mg) U A V AD W B
<0.003 100 0 0 0 0 0 0.003-0.009 100 0 0 0 0 0 0.010-0.031 72 0 2 11 15 0 0.032-0.099 64 0 0 2 34 0 0.100-0.315 47 1 5 7 39 1 0.316-0.999 46 12 4 13 23 2 1.000-3.161 39 17 7 15 20 2 3.161-9.999 31 32 17 7 10 3 10.00-31.611 31 20 22 1 22 4
31.612-99.999 32 15 32 0 21 0 100.00-316.108 13 0 87 0 0 0
316.109-999.99 0 0 100 0 0 0 1000.00-3161 0 0 100 0 0 0
>3161 0 0 100 0 0 0
This content downloaded from 38.125.197.2 on Fri, 17 Jul 2015 15:22:29 UTCAll use subject to JSTOR Terms and Conditions





























