Embed Size (px)
Citation preview

letters to nature
436 NATURE | VOL 415 | 24 JANUARY 2002 | www.nature.com
vernier. For the same reason, activity corresponding to four isolated(that is, unbound) contextual elements (Fig. 2a) might not bedecreased by neighbouring contextual elements and, thus, mightproduce high neural activity inhibiting the vernier target. For theextended standard grating, shine through is possible because littleneural activity occurs in the vernier's close neighbourhood. Analo-gously, extended contextual gratings might not interfere with thevernier because activity corresponding to their inner elements isweak. Therefore, context±context inhibition prevents context±target suppression of the vernier target.
Contextual elements might directly in¯uence the ®ring pattern ofcortical neurons that code the vernier. For example, the activity of aneuron that is responding vigorously to a vernier target shoulddiminish when isolated contextual elements are presented togetherwith the standard grating. By contrast, increasing the number ofcontextual elements should yield a rebound of activity. Becausevernier display times are short, we expect strongest modulationeffects in the transient response of the neuronsÐthat is, in theactivity immediately after the presentation of the target. M
Received 17 July; accepted 20 November 2001.
1. Knierim, J. J. & van Essen, D. C. Neuronal responses to static texture patterns in area V1 of the alert
macaque monkey. J. Neurophys. 67, 961±980 (1992).
2. Li, W., Thier, P. & Wehrhahn, C. Contextual in¯uence on orientation discrimination of humans and
responses of neurons in V1 of alert monkeys. J. Neurophys. 83, 941±954 (2000).
3. Nothdurft, H.-C., Gallant, J. L. & van Essen, D. C. Response modulation by texture surround in
primate area V1: Correlates of pop-out under anesthesia. Vis. Neurosci. 16, 15±34 (1999).
4. Herzog, M. H. & Koch, C. Seeing properties of an invisible element: feature inheritance and shine-
through. Proc. Natl Acad. Sci. USA 98, 4271±4275 (2001).
5. Herzog, M. H., Fahle, M. & Koch, C. Spatial aspects of object formation revealed by a new illusion,
shine-through. Vis. Res. 41, 2325±2335 (2001).
6. Mareschal, I., Sceniak, M. P. & Shapley, M. Contextual in¯uences on orientation discrimination:
binding local and global cues. Vis. Res. 41, 1915±1930 (2001).
7. Wehrhahn, C., Li, W. & Westheimer, G. Patterns that impair discrimination of orientation in human
vision. Perception 25, 1053±1064 (1996).
8. Stemmler, M., Usher, M. & Niebur, E. Lateral interactions in primary visual cortex: a model bridging
physiology and psychophysics. Science 269, 1877±1880 (1995).
9. Li, Z. Contextual in¯uences in V1 as a basis for pop out and asymmetry in visual search. Proc. Natl
Acad. Sci. USA 96, 10530±10535 (1999).
10. Wolfson, S. S. & Landy, M. S. Long range interactions between oriented texture elements. Vis. Res. 39,
933±945 (1999).
11. Nothdurft, H. C. Texture segmentation and pop-out from orientation contrast. Vis. Res. 31, 1073±
1078 (1991).
12. Cannon, M. W. & Fullenkamp, S. C. Spatial interactions in apparent contrast: inhibitory effects
among grating patterns of different spatial frequencies, spatial positions and orientations. Vis. Res. 31,
1985±1998 (1991).
13. Lamme, V. A. The neurophysiology of ®gure-ground segregation in primary visual cortex. J. Neurosci.
15, 1605±1615 (1995).
14. Zipser, K., Lamme, V. A. F. & Schiller, P. H. Contextual modulation in primary visual cortex.
J. Neurosci. 16, 7376±7389 (1996).
15. Caputo, G. The role of the background: Texture segregation and ®gure-ground segmentation. Vis. Res.
36, 2815±2826 (1996).
16. Sengpiel, F., Sen A. & Blakemore, C. Characteristics of surround inhibition in cat area 17. Exp. Brain
Res. 116, 216±228 (1997).
17. Sillito, A. M., Grieve, K. L., Jones, H. E., Cudeiro, J. & Davis, J. Visual cortical mechanisms detecting
focal orientation discontinuities. Nature 378, 492±496 (1995).
18. Macknik, S. L. & Livingstone, M. S. Neuronal correlates of visibility and invisibility in the primate
visual system. Nature Neurosci. 1, 144±149 (1998).
19. Polat, U. & Sagi, D. Lateral interactions between spatial channels: suppression and facilitation revealed
by lateral masking experiments. Vis. Res. 33, 993±999 (1993).
20. Kapadia, M. K., Ito, M., Gilbert, C. D. & Westheimer, G. Improvement in visual sensitivity by changes
in local context: parallel studies in human observers and in V1 of alert monkeys. Neuron 15, 843±856
(1995).
21. Levitt, J. B. & Lund, J. S. Contrast dependence of contextual effects in primate visual cortex. Nature
387, 73±76 (1997).
22. Bonneh, Y. & Sagi, D. Effects of spatial con®guration on contrast detection. Vis. Res. 38, 3541±53 (1998).
23. Yu, C. & Levi, D. Surround modulation in human vision unmasked by masking experiments. Nature
Neurosci. 3, 724±728 (2000).
24. Das, A. & Gilbert, C. D. Topography of contextual modulations mediated by short-range interactions
in primary visual cortex. Nature 399, 655±661 (1999).
25. Chubb, C., Sperling, G. & Solomon, J. A. Texture interactions determine perceived contrast. Proc. Natl
Acad. Sci. USA 86, 9631±9635 (1989).
26. Weisstein, N. & Harris, C. S. Visual detection of lines segments: an object-superiority effect. Science
186, 752±755 (1974).
27. Banks, W. P. & White, H. Lateral interference and perceptual grouping in visual detection. Percept.
Psychophys. 36, 285±295 (1984).
28. Kurylo, D. D. Time course of perceptual grouping. Percept. Psychophys. 59, 142±147 (1997).
29. Ernst, U. A., Pawelzik, K. R., Sahar-Pikielny, C. & Tsodyks, M. V. Intracortical origin of visual maps.
Nature Neurosci. 4, 431±436 (2001).
Acknowledgements
This work was supported by the Center of Excellence (SFB) `Neurocognition' of theGerman Research Council (Deutsche Forschungsgemeinschaft). We thank M. Repnow fortechnical support; and B. Gillam, C. Eurich, U. Ernst, A. Etzold, B. Zenger,K. Neumann, F. Kandil, J. Solomon, M. Morgan, J. Zanker and U. Schmonsees fordiscussions.
Competing interests statement
The authors declare that they have no competing ®nancial interests.
Correspondence and requests for materials should be addressed to M.H.H.(e-mail: [email protected]).
.................................................................Prediction of central nervous systemembryonal tumour outcome basedon gene expressionScott L. Pomeroy*, Pablo Tamayo², Michelle Gaasenbeek²,Lisa M. Sturla*, Michael Angelo², Margaret E. McLaughlin³,John Y. H. Kim*§, Liliana C. Goumnerovak, Peter M. Blackk, Ching Lau¶,Jeffrey C. Allen#, David ZagzagI, James M. Olson**, Tom Curran²²,Cynthia Wetmore²², Jaclyn A. Biegel³³, Tomaso Poggio§§,Shayan Mukherjee§§, Ryan Rifkin§§, Andrea Califanokk,Gustavo Stolovitzkykk, David N. Louis¶¶, Jill P. Mesirov²,Eric S. Lander²## & Todd R. Golub²§II
* Division of Neuroscience, Department of Neurology; ³ Department of Pathology;kDepartment of Neurosurgery; II Department of Medicine, Children's Hospital;
§ Department of Pediatric Oncology, Dana-Farber Cancer Institute; and
¶¶ Department of Pathology and Neurosurgical Service, Massachusetts GeneralHospital, Harvard Medical School, Boston, Massachusetts 02115, USA² Whitehead Institute/MIT Center for Genome Research, and §§ McGovern
Institute, Center for Biological and Computational Learning, AI Lab,
Massachusetts Institute of Technology, Cambridge, Massachusetts 02139, USA¶ Division of Pediatric Oncology, Baylor College of Medicine, Houston, Texas
77030, USA
# Beth Israel Medical Center, New York 10128, USAI Department of Pathology, New York University School of Medicine, New York10016, USA
** Clinical Research Division, Fred Hutchinson Cancer Research Center, Seattle,
Washington 98109, USA²² Department of Developmental Neurobiology, St Jude Children's Research
Hospital, Memphis, Tennessee 38105, USA³³ Division of Human Genetics, The Children's Hospital of Philadelphia,
Department of Pediatrics, University of Pennsylvania School of Medicine,Philadelphia, Pennsylvania 19104, USA
## Department of Biology, Massachusetts Institute of Technology, Cambridge,
Massachusetts 02139, USA
kk IBM Watson Research Center, Yorktown Heights, New York 10598, USA
..............................................................................................................................................
Embryonal tumours of the central nervous system (CNS) repre-sent a heterogeneous group of tumours about which little isknown biologically, and whose diagnosis, on the basis of mor-phologic appearance alone, is controversial. Medulloblastomas,for example, are the most common malignant brain tumour ofchildhood, but their pathogenesis is unknown, their relationshipto other embryonal CNS tumours is debated1,2, and patients'response to therapy is dif®cult to predict3. We approached theseproblems by developing a classi®cation system based on DNAmicroarray gene expression data derived from 99 patient samples.Here we demonstrate that medulloblastomas are molecularlydistinct from other brain tumours including primitive neuro-ectodermal tumours (PNETs), atypical teratoid/rhabdoid tumours
© 2002 Macmillan Magazines Ltd
letters to nature
NATURE | VOL 415 | 24 JANUARY 2002 | www.nature.com 437
MD Mglio Rhab Ncer PNETM93119 INSM1 insulinoma-associated 1M30448 Casein kinase II beta subunit S82240 RhoED80004 KIAA0182 geneD76435 ZIC proteinX83543 APXL apical proteinX62534 HMG2 high-mobility groupM96739 NSCL1 U26726 11 beta-hydroxysteroid dehydrogenase type IIHG311-HT311 Ribosomal protein L30X86693 High endothelial venuleM93426 PTPRZ protein tyrosine phosphataseU48705 DDR geneX86809 Major astrocytic phosphoprotein PEA15U45955 Neuronal membrane glycoprotein M6bU53204 Plectin (PLEC1) X13916 LDL-receptor related proteinD87258 Serin protease with IGF-binding motifZ31560 SOX2 SRY (sex-determining region Y)-box 2M3288 6SRI sorcinJ04164 RPS3 ribosomal protein S3M12125 Skeletal beta-tropomyosinD29958 KIAA0116 geneD17400 PTS 6-pyruvoyltetrahydropterin synthaseD83174 CBP1 collagen-binding protein 1D83735 Adult heart mRNA for neutral calponinD84454 UDP-galactose translocatorL38969 Thrombospondin 3 (THBS3) U12465 RPS11 ribosomal protein S11 D80005 KIAA0183 geneD87463 KIAA0273 geneU90902 Clone 23612 mRNA sequenceD26070 Type 1 inositol 1,4,5-trisphosphate receptorX63578 ParvalbuminZ15108 PRKCZ protein kinase C, zetaL35592 Germline mRNA sequenceL10338 SCN1B sodium channelL33243 PKD1 polycystic kidney disease protein 1L77864 Stat-like protein (Fe65) J04469 Mitochondrial creatine kinase (CKMT)M80397 POLD1 polymerase (DNA directed), delta 1X14830 CHRNB1 cholinergic receptor, nicotinic, beta polypeptide 1U97018 Echinoderm microtubule-associated protein homologue HuEMAPHG4178-HT4448 Af-17K02882 IGHD gene X52228 MUC1 mucin 1, transmembraneU22314 Neural-restrictive silencer factorD29675 Inducible nitric oxide synthase geneS82471 SSX3M54951 Human atrial natriuretic factor gene
AT/RT CNS
Ncer
MDMglio
PNET
AT/RT renal/extrarenal
13.00
–13.0
20.0
10.00
–10.0 –12.00
Comp 2
Com
p 3
Comp 1
Comp 2
Com
p 3
Comp 1
12.024.0
4.0
0
–4.06.0
3.00
–3.0–6.0 –6.0
–3.00
3.0
A
D
E
B
C
a
c d e
b
–3σ –2σ –1σ 0 +1σ +2σ +3σ
n
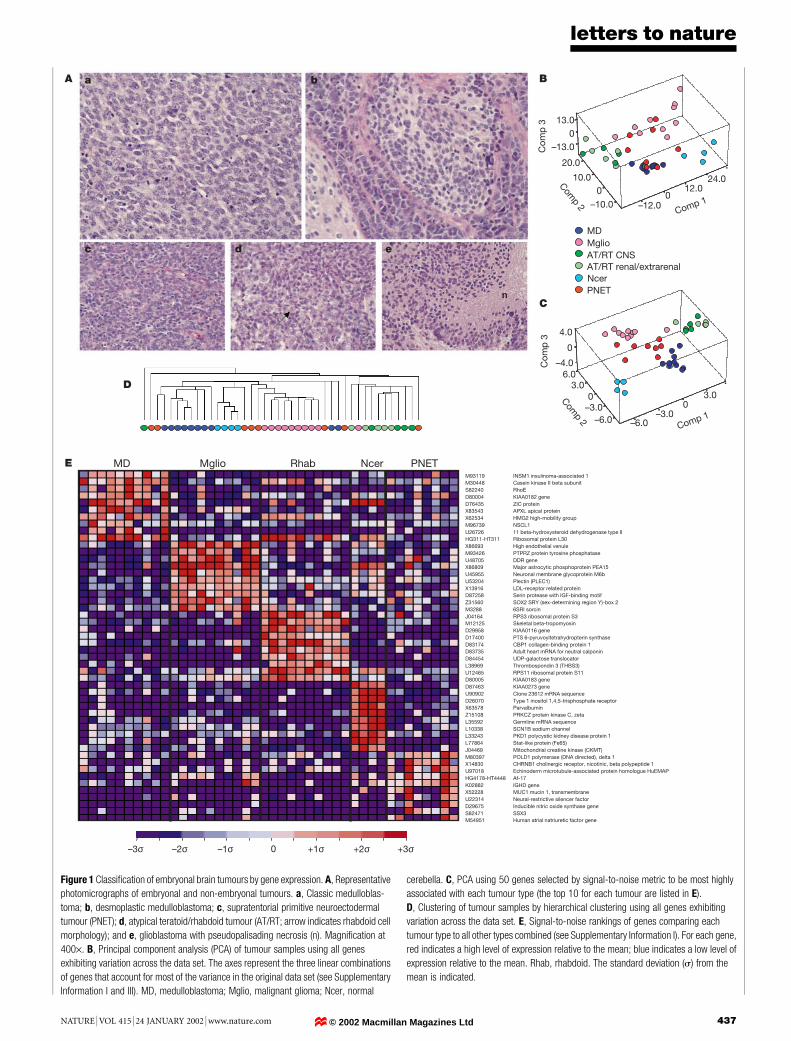
Figure 1 Classi®cation of embryonal brain tumours by gene expression. A, Representative
photomicrographs of embryonal and non-embryonal tumours. a, Classic medulloblas-
toma; b, desmoplastic medulloblastoma; c, supratentorial primitive neuroectodermal
tumour (PNET); d, atypical teratoid/rhabdoid tumour (AT/RT; arrow indicates rhabdoid cell
morphology); and e, glioblastoma with pseudopalisading necrosis (n). Magni®cation at
400´. B, Principal component analysis (PCA) of tumour samples using all genes
exhibiting variation across the data set. The axes represent the three linear combinations
of genes that account for most of the variance in the original data set (see Supplementary
Information I and III). MD, medulloblastoma; Mglio, malignant glioma; Ncer, normal
cerebella. C, PCA using 50 genes selected by signal-to-noise metric to be most highly
associated with each tumour type (the top 10 for each tumour are listed in E).
D, Clustering of tumour samples by hierarchical clustering using all genes exhibiting
variation across the data set. E, Signal-to-noise rankings of genes comparing each
tumour type to all other types combined (see Supplementary Information I). For each gene,
red indicates a high level of expression relative to the mean; blue indicates a low level of
expression relative to the mean. Rhab, rhabdoid. The standard deviation (s) from the
mean is indicated.
© 2002 Macmillan Magazines Ltd

letters to nature
438 NATURE | VOL 415 | 24 JANUARY 2002 | www.nature.com
Classic DesmoplasticHG1980-HT2023 Tubulin, beta 2U63842 NeuroD3 geneX67951 PAGA proliferation-associated gene AX64330 ATP-citrate lyaseJ03241 TGFB3 transforming growth factor, beta 3U44839 Putative ubiquitin C-terminal hydrolase (UHX1)Z27113 H. sapiens gene for RNA polymerase II subunitX12447 ALDOA aldolase AU73328 DLX7 distal-less homeobox 7U59913 SMAD5Z75190 Apolipoprotein E receptor 2X15183 Human mRNA for 90K heat-shock proteinU61263 Acetolactate synthase homologueL07515 Human heterochromatin protein homologue (HP1)M34677 Human nested gene protein gene (coag. factorVIII)U40391 Serotonin N-acetyltransferase geneD16611 CPO coproporphyrinogen oxidaseL37127 RNA polymerase IIU33839 No description availableX81817 BAP31X51804 Human PMI gene for a putative receptor proteinY09305 Protein kinase, Dyrk4X14885 Transforming growth factor-beta 3 (TGF-beta 3)X57398 NME1 non-metastatic cells 1 (NM23A)X02152 LDHA lactate dehydrogenase AX64364 BSG basiginU04806 FLT3/FLK2 ligand mRNAU52191 SMCY (H-Y) mRNAM35296 Tyrosine kinase arg geneD50840 Ceramide glucosyltransferaseS69189 Peroxisomal acyl-coenzyme A oxidaseL03785 MYL5 myosin, light polypeptide 5M82919 GABRB3 gamma-aminobutyric acid A receptor, beta 3U65092 Melanocyte-specific gene 1 (MSG1)S49592 Transcription factor E2F-like proteinD79994 KIAA0172 geneX95586 PSMB5 proteasome subunit, beta type, 5U79299 Human neuronal olfactomedin-related ER localized proteinX71973 GPX4 phospholipid hydroperoxide glutathione peroxidaseU31342 Human nucleobindin geneU48437 Amyloid precursor-like protein 1D86957 KIAA0202 geneHG2279-HT2375 Triosephosphate isomeraseZ19585 THBS4 thrombospondin 4M16405 Human m4 muscarinic acetylcholine receptor geneU32315 Syntaxin 3U11701 LIM-homeobox domain protein (hLH-2)Y11251 Novel member of serine-arginine domain protein, SRrp129U68018 Mad protein homologue (hMAD-2)M81181 ATP1B2 ATPase, Na+/K+ transporting, beta 2 polypeptideU58334 Bcl2, p53 binding protein BBP/53BP2L38490 ARF4L ADP-ribosylation factor 4-likeM31328 GNB3 guanine nucleotide binding proteinM60626 FPR1 formyl peptide receptor 1M97815 CRABP2 cellular retinoic acid-binding protein 2J00116 COL2A1 collagen, type II, alpha 1U01824 Glutamate transporterY00282 RPN2 ribophorin IIM65254 PPP2R1B protein phosphatase 2X64877 HFL1 H factor (complement)-like 1D31885 KIAA0069 geneD87434 KIAA0247 geneZ35093 SURF1 surfeit 1Y08374 GP-39 cartilage protein geneZ54367 PlectinZ46632 PDE4C phosphodiesterase 4CHG3517-HT3711 Alpha-1-antitrypsinL42354 Clone 48ES4 mRNA fragmentX15376 GABRG2 gamma-aminobutyric acid A receptor, gamma 2X89059 Unknown protein expressed in macrophages
HG3543-HT3739 Insulin-like growth factor 2X53331 MGP matrix protein GlaX65724 NDP norrie disease proteinD14530 40S ribosomal protein S23Y00757 SGNE1 secretory granule, neuroendocrine protein 1U25789 Ribosomal protein L21L27560 Insulin-like growth factor binding protein 5 (IGFBP5)X83543 APXL apical protein (Xenopus laevis-like)X52966 RPL35A ribosomal protein L35aL06797 Human (clone L5) orphan G protein-coupled receptorM14745 BCL2 BHG3431-HT3616 DecorinD79205 Ribosomal protein L39D82345 NB thymosin betaD38549 KIAA0068 geneU14972 Ribosomal protein S10X59841 PBX3J03242 IGF2 insulin-like growth factor 2 (somatomedin A)HG3214-HT3391 Metallopanstimulin 1X06617 RPS11 ribosomal protein S11J02611 APOD apolipoprotein DX16064 Human mRNA for translationally controlled tumour proteinZ74616 COL1A2 collagen, type I, alpha-2L40386 DP2 (Humdp2)X04741 Human mRNA for protein gene product (PGP)M55210 LAMC1 laminin, gamma 1M96739 NSCL1U73304 CB1 cannabinoid receptor (CNR1) geneM65292 HFL1 H factor (complement)-like 1HG311-HT311 Ribosomal protein L30M83233 TCF12 transcription factor 12U07919 ALDH6 aldehyde dehydrogenase 6X57959 RPL17 ribosomal protein L7J04080 C1S complement component 1, s subcomponentX76029 Neuromedin UU14973 40S ribosomal protein S29U24576 Breast tumour autoantigenL41066 NFAT3X60489 Elongation factor-1-betaM62843 Paraneoplastic encephalomyelitis antigen HUDHG662-HT662 Epstein–Barr virus small RNA-associated proteinHG613-HT613 Ribosomal protein S12L42379 Quiescin (Q6) mRNAM13241 N-MYCU12404 HSPB1 heat shock 27K protein 1M55998 Alpha-1 collagen type I geneS82240 RhoEU78027 L44L geneX06700 COL3A1 alpha-1 type 3 collagenD13413 Tumour-associated 120K nuclear protein p120M74719 SEF2-1A protein (SEF2-1A)M93119 INSM1 insulinoma-associated 1M92287 CCND3 cyclin D3HG33-HT33 Ribosomal protein S4U16306 CSPG2 chondroitin sulphate proteoglycan 2Z37976 LTBP2 latent transforming growth factor beta binding protein 2X69150 Ribosomal protein S18HG4542-HT4947 Ribosomal protein L10L41607 GCNT2 glucosaminyl (N-acetyl) transferase 2U43148 PTCH Patched homologueM30269 NID nidogen (enactin)X07384 GLI glioma-associated oncogene homologueL38941 RPL37 ribosomal protein L37U09953 RPL9 ribosomal protein L9D87464 KIAA0274 geneM18000 40S ribosomal protein S17M91196 ICSBP1 interferon consensus sequence binding protein 1L27559 IGFBP5 insulin-like growth factor binding protein 5S76475 NTRK3 neurotrophic tyrosine kinase, receptor, type 3 (TrkC)L41067 Transcription factor NFATx
Classic Desmoplastic
IGF2
N-MYC
PTCH
GLI
–3σ –2σ –1σ 0 +1σ +2σ +3σ
Figure 2 Differential expression of genes in classic versus desmoplastic medullo-
blastomas. Genes were ranked by the signal-to-noise metric according to their correlation
with the classic versus desmoplastic distinction. Genes shown are those more highly
correlated with the distinction than 99% of permutations of the class labels (P , 0.01; see
Supplementary Information III). GenBank accession numbers and gene descriptions are
shown. Genes regulated by SHH are shown at the right. Normalized level of expression of
selected SHH-regulated genes is shown at right. Each bar represents a different tumour.
© 2002 Macmillan Magazines Ltd

letters to nature
NATURE | VOL 415 | 24 JANUARY 2002 | www.nature.com 439
(AT/RTs) and malignant gliomas. Previously unrecognizedevidence supporting the derivation of medulloblastomas fromcerebellar granule cells through activation of the Sonic Hedgehog(SHH) pathway was also revealed. We show further that theclinical outcome of children with medulloblastomas is highlypredictable on the basis of the gene expression pro®les of theirtumours at diagnosis.
We ®rst addressed the problem of distinguishing different embry-onal CNS tumours from each other. This is important as theclassi®cation of these tumours based on histopathological appear-ance is debated (Fig. 1A). There are two hypotheses regarding theclassi®cation of medulloblastomas: the ®rst is that they are part of alarger class of PNETs arising from a common cell type in thesubventricular germinal matrix1, the second is that they arise fromcerebellar granule cell progenitors2. To begin to generate a moleculartaxonomy of CNS embryonal tumours, we analysed the geneexpression pro®les of 42 patient samples (data set A: 10 medullo-blastomas, 5 CNS AT/RTs, 5 renal and extrarenal rhabdoid tumours,and 8 supratentorial PNETs, as well as 10 non-embryonal braintumours (malignant glioma) and 4 normal human cerebella). RNAextracted from frozen specimens was analysed with oligonucleotidemicroarrays containing probes for 6,817 genes. The gene expressiondata are available as Supplementary Information II (see also http://www.genome.wi.mit.edu/MPR/CNS).
To determine whether the different types of tumours could bemolecularly distinguished, we used a method of data reductioncalled principal component analysis in which the high dimension-ality of the data was reduced to three viewable dimensions repre-senting linear combinations of variables (genes) that account formost of the variance in the original data set (Fig. 1B)4. Normal brainwas easily separable from the brain tumours, and the differenttumour types were similarly separable. Separation of tumour typeswas also seen using hierarchical clustering (Fig. 1D)5. A moreappropriate strategy for distinguishing known tumour types, how-ever, is to use supervised learning methods to identify the genesmost highly correlated with the tumour type distinctions (Fig. 1C,E). Analysis of 1,000 random permutations of the data failed to yielda separation of tumour classes to the extent observed in Fig. 1C,indicating that the observed gene expression patterns could not beexplained by chance (Supplementary Information III). The robust-ness of these markers for classi®cation was further investigatedusing a weighted-voting algorithm and evaluated by cross validationtesting6. Correct classi®cation of the tumours could be achievedwith accuracy (35 out of 42 correct classi®cations, P , 10-10
compared with random classi®cation; see Supplementary Informa-tion III).
As expected, malignant gliomas were clearly separable frommedulloblastomas, re¯ecting the derivation of gliomas from cellsof non-neuronal origin. Consistent with this, the gliomas expressed
genes typical of the astrocytic and oligodendrocytic lineage (PEA15,SOX2, PMP2, Olig-2, TrkB kinase-negative splice variant, S100,GFAP), genes related to metabolism (fructose 2,6-bisphosphatase,glutamate dehydrogenase), and genes involved in cell differentiation(ID2, GDF1, TYK2; Fig. 1E and Supplementary Information III).The medulloblastomas form a cluster that is also separate from thePNETs (Fig. 1C), supporting the hypothesis that these two classes ofembryonal tumours are molecularly distinct. Among the genes mosthighly correlated with the medulloblastoma class were ZIC andNSCL1, encoding transcription factors that are speci®c for cere-bellar granule cells (Fig. 1E)7,8. This result suggests that medullo-blastomas, but not PNETs, arise from cerebellar granule cells, oralternatively, have activated the transcriptional programme ofcerebellar granule cells.
We next analysed the AT/RT tumours, which have only recentlybeen distinguished from medulloblastoma9. Accurate identi®cationof AT/RT is particularly important because patients with thesetumours have an extremely poor prognosis. AT/RT tumours arisein the CNS or in other organs such as the kidney, where they arereferred to as rhabdoid tumours. Most tumours harbour hSNF5/INI1 mutations, but it is unknown whether AT/RTs arising indifferent anatomical locations are molecularly distinct9±11. Asshown in Fig. 1C, the AT/RTs and rhabdoid tumours were easilydistinguishable from the other tumour types in the study. Of note,the CNS AT/RTs and abdominal rhabdoid tumours were molecu-larly similar despite having arisen in different anatomical locations.This ®nding supports the idea that they arise from a similar cell oforigin. Alternatively, a common mechanism of transformation mayyield similar transcriptional programmes in cells of distinct origin.Markers of the distinction between AT/RT and rhabdoid tumoursinclude genes speci®cally expressed during myogenesis, includingskeletal b-tropomyosin, neutral calponin, NFAT3, and myosinregulatory light chain (Fig. 1E and Supplementary InformationIII). This ®nding is consistent with the hypothesis that the tumourshave a mesenchymal origin.
We next focused on molecular heterogeneity within a singletumour type, medulloblastoma. The principal histological subclassof medulloblastoma is desmoplastic medulloblastoma, although itsdiagnosis is highly subjective (Fig. 1A). Desmoplastic medulloblas-toma is of interest because it is seen with high frequency in patientswith Gorlin's syndrome, a rare autosomal dominant disorderresulting from mutation of the SHH receptor PTCH12,13. It is unclearwhether dysregulation of the SHH pathway, known to be mitogenicfor cerebellar granule cells, is also involved in the pathogenesis ofsporadic desmoplastic medulloblastoma14±18.
To determine whether desmoplastic and classic medulloblastomaare distinguishable by gene expression, we analysed 34 medullo-blastoma samples (data set B) whose histology was scored usingWorld Health Organization (WHO) criteria19. As shown in Fig. 2, asharp and statistically signi®cant gene expression signature ofdesmoplastic histology was evident, and this signature was suf®cientfor correct classi®cation of 33 out of 34 tumours (P = 8.6 ´ 10-7compared with random classi®cation; see Supplementary Informa-tion III). Notably, among the genes most highly correlated withdesmoplastic medulloblastoma was PTCH (itself a transcriptionaltarget of SHH), as well as two other SHH downstream targets: GLI20
and N-MYC (A. Kenney and D. Rowitch, personal communication).Furthermore, insulin-like growth factor II expression was correlatedwith desmoplastic histology, and its expression is essential for SHH-mediated tumorigenesis in mice21. Taken together, the transcrip-tional pro®ling indicates that sporadic desmoplastic medulloblas-tomas, like tumours associated with Gorlin's syndrome, arecharacterized by activation of the SHH signalling pathway, furthersupporting the proposal that SHH dysregulation may be importantin the pathogenesis of medulloblastoma.
A clinical challenge concerning medulloblastoma is the highlyvariable response of patients to therapy. Whereas some patients are
Figure 3 Representative electron micrographs showing medulloblastomas with low
ribosome (a) and high ribosome (b) content. Each panel shows a portion of a single cell
with a portion of the nucleus (n) (arrows designate ribosomes). Scale bars, 0.5 mm.
© 2002 Macmillan Magazines Ltd
letters to nature
440 NATURE | VOL 415 | 24 JANUARY 2002 | www.nature.com
Failures Survivors
C1 C0 C1 C0
P = 0.009
Ove
rall
surv
ival
Months
00
0.2
0.4
0.6
0.8
1.0
20 40 1208060 100
L17131 High mobility group protein (HMG-I(Y))X13482 Human mRNA for U2 snRNP-specific A’ proteinL12711 TKT transketolaseL19711 Dystroglycan (DAG1)X04741 U2 small nuclear ribonucleoprotein A’U12404 HSPB1 heat shock 27K protein 1U15008 SnRNP core protein Sm D2U81375 Placental equilibrative nucleoside transporter 1 (hENT1)X13794 Lactate dehydrogenase B geneZ49148 Enhancer of rudimentary homologueU39318 AF-4X67247 RpS8 gene for ribosomal protein S8U14968 Ribosomal protein L27aHG613-HT613 Ribosomal protein S12D63880 KIAA0159 geneY07604 Nucleoside-diphosphate kinaseJ04823 Cytochrome c oxidase subunit VIII (COX8)M13934 RPS14 geneU30872 CENP-F kinetochore proteinM81757 40S ribosomal protein S19L07515 Human heterochromatin protein homologue (HP1)M14328 ENO1 enolase 1, (alpha)D82348 5-aminoimidazole-4-carboxamide-1-beta-D-ribonucleotide transformylase/inosinicaseD78586 Human CAD mRNA for multifunctional protein CADM32886 SRI sorcinU31556 E2F5 E2F transcription factor 5, p130-bindingX94910 ERp31 proteinY10313 Nerve growth factor-inducible PC4 homologueS78187 M-phase inducer phosphatase 2HG2479-HT2575 Helix-loop-helix protein Sef2-1dU12595 Tumour necrosis factor type 1 receptor associated protein (TRAP1)L36720 BystinHG3214-HT3391 Metallopanstimulin 1HG4542-HT4947 Ribosomal protein L10D29805 GGTB2 glycoprotein-4-beta-galactosyltransferase 2X52966 RPL35A ribosomal protein L35aM64716 RPS25 ribosomal protein S25M64347 FGFR3 fibroblast growth factor receptor 3U09770 Cysteine-rich heart protein (hCRHP)D28473 IARS isoleucine-tRNA synthetaseX69908 P2 gene for c subunit of mitochondrial ATP synthase geneU76638 BRCA1-associated RING domain protein (BARD1)X79234 Ribosomal protein L11X15376 GABRG2 gamma-aminobutyric acid A receptor, gamma 2M14199 LAMR1 laminin receptor (2H5 epitope)
U28963 Gps2 (GPS2)X69636 H. sapiens mRNA sequence (15q11-13)U18018 ETV4 Ets variant gene 4 (E1A enhancer-binding protein, E1AF)M97287 SATB1 special (A+T)-rich sequence binding protein 1U78180 Sodium channel 2 (hBNaC2) mRNA, alternatively splicedS76475 NTRK3 neurotrophic tyrosine kinase, receptor, type 3 (TrkC)D28124 Unknown productU70867 Prostaglandin transporter hPGTM17733 Thymosin beta-4L10333 Neuroendocrine-specific protein A (NSP)D14686 AMT glycine cleavage system protein T (aminomethyltransferase)S66541 B-50 neural phosphoproteinAC0020 A-589H1.2 from H. sapiens chromosome 16 BAC clone CIT987-SKA-589H1M96739 NSCL1D86963 PTB ribosomal protein L26U40271 PTK7 protein-tyrosine kinase 7L09229 FACL1 long chain fatty acid acyl-coA ligaseD78012 CRMP1 collapsin response mediator protein 1M74715 IDUA iduronidase, alpha-L-HG2525-HT2621 Helix-loop-helix protein delta max, alt. splice 1L32164 Zinc finger proteinL04731 Translocation T(4:11) of ALL-1 gene to chromosome 4M22919 MLC gene (non-muscle myosin light chain)X15882 COL6A2 collagen, type VI, alpha 2U20657 Ubiquitin protease (Unph) proto-oncogeneL17327 Pre-T/NK cell associated protein (3B3)J05412 REG1A regenerating islet-derived 1 alpha (pancreatic stone protein)D43682 Very-long-chain acyl-CoA dehydrogenase (VLCAD)X58521 Nuclear pore glycoprotein P62M21142 Guanine nucleotide-binding protein G-s-alpha-3 geneX52896 RNA for dermal fibroblast elastinD50663 CW-1U35139 NECDIN related proteinU16660 Peroxisomal enoyl-CoA hydratase-like protein (HPXEL)U04241 Homologue of Drosophila enhancer of split m9/m10Y07847 RRP22 proteinU78521 Immunophilin homologue ARA9X93511 Telomeric repeat binding factor (TRF1)D30715 Exon2a from human PAP (pancreatitis-associated protein) geneU51920 SRP54 signal recognition particle 54K proteinU02619 TFIIIC box B-binding subunitU14417 Ral guanine nucleotide dissociation stimulatorM73547 Human polyposis locus (DP1 gene)U09820 Helicase II (RAD54L)X13461 Calmodulin-related protein NB-1Z56281 Interferon regulatory factor 3
K03189 Chorionic gonadotropin (HCG) beta subunit
–3σ –2σ –1σ 0 +1σ +2σ +3σ
Markers of treatment failureX69150 Ribosomal protein S18
U14972 Ribosomal protein S10
M36072 RPL7A ribosomal protein L7aX13293 MYBL2 V-mmyb avian myeloblastosis viral oncogene homologue-like 2
L06419 PLOD lysyl hydroxylaseJ02611 APOD apolipoprotein DD86974 KIAA0220 geneU37673 Beta-NAP
Markers of survival
a
b
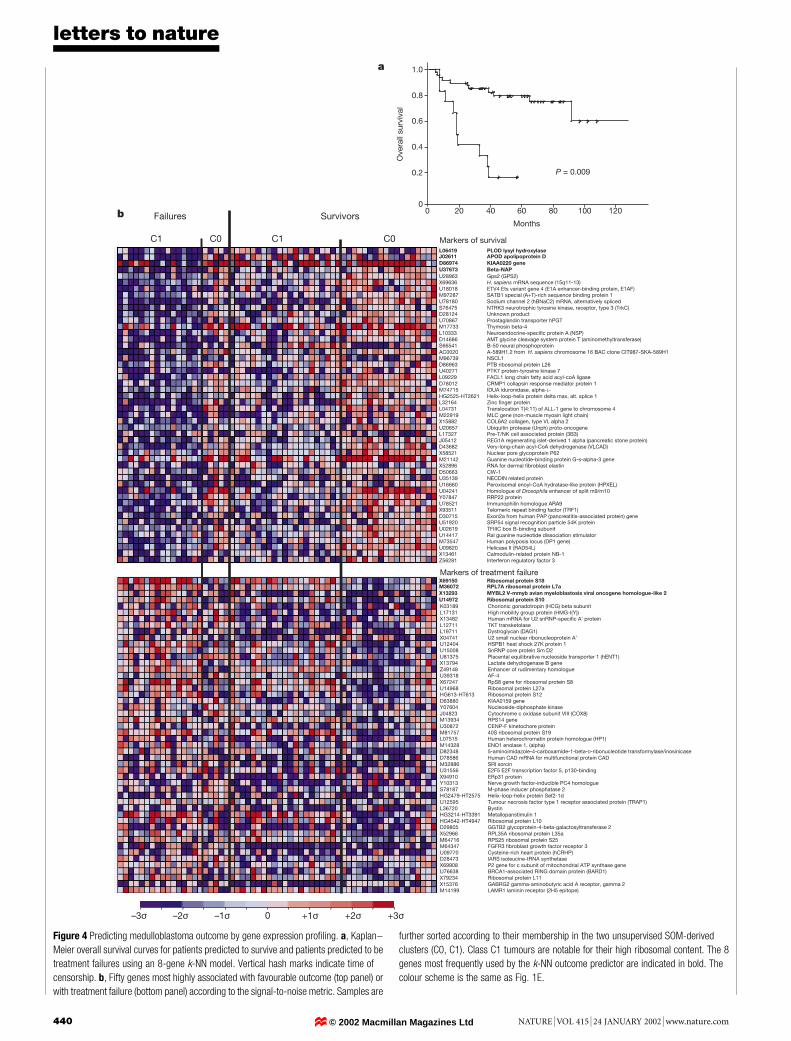
Figure 4 Predicting medulloblastoma outcome by gene expression pro®ling. a, Kaplan±
Meier overall survival curves for patients predicted to survive and patients predicted to be
treatment failures using an 8-gene k-NN model. Vertical hash marks indicate time of
censorship. b, Fifty genes most highly associated with favourable outcome (top panel) or
with treatment failure (bottom panel) according to the signal-to-noise metric. Samples are
further sorted according to their membership in the two unsupervised SOM-derived
clusters (C0, C1). Class C1 tumours are notable for their high ribosomal content. The 8
genes most frequently used by the k-NN outcome predictor are indicated in bold. The
colour scheme is the same as Fig. 1E.
© 2002 Macmillan Magazines Ltd

letters to nature
NATURE | VOL 415 | 24 JANUARY 2002 | www.nature.com 441
cured by chemotherapy and radiation, others have progressivedisease3. Currently, the only prognostic factor used in clinical practiceis tumour staging; a re¯ection of postoperative tumour size and thepresence of metastases. Unfortunately, staging-based prognosticationis imperfect in that many patients with low-stage disease stillsuccumb to their disease. There are currently no molecular markersof outcome used in clinical practice for any brain tumour. Highlevels of expression of the neurotrophin-3 receptor (TRKC), how-ever, have been reported to correlate with a favourable medullo-blastoma outcome22,23, suggesting a molecular basis for thevariability of medulloblastoma outcome. Molecular correlates ofmedulloblastoma metastasis have also been recently reported24.
To explore the heterogeneity in response to treatment of medullo-blastomas, we expanded our analysis to include 60 similarly treatedpatients from whom biopsies were obtained before receiving treat-ment, and for whom clinical follow-up was available (data set C).We ®rst investigated whether clustering methods would identifybiologically distinct subsets of the tumours. The tumours wereclustered into two groups using self-organizing maps (SOM); anunsupervised algorithm that groups samples into a predeterminednumber of clusters on the basis of their gene expression patterns6,25.The genes most highly correlated with the SOM clusters wereprimarily ribosomal protein-encoding genes (Supplementary Infor-mation III), suggesting differences in ribosome biogenesis. Blindedelectron microscopic examination of 9 samples by 3 observerscon®rmed that tumours falling into the cluster characterized byhigh expression of ribosomal protein genes contained highernumbers of ribosomes (P = 0.03, Fisher's exact test; Fig. 3). Wenext investigated whether the SOM-derived clusters were correlatedwith patient survival. No statistically signi®cant difference in theproportion of survivors compared with treatment failures in eachcluster was observed (Fisher's exact test, P = 0.1; see SupplementaryInformation III). Because unsupervised methods are generally notthe most appropriate analytical approach to predicting knowndistinctions such as outcome, we developed a supervised learningoutcome predictor based on gene expression in which the classi®er`learns' the distinction between patients who are alive after treat-ment (`survivors') compared with those who succumbed to theirdisease (`failures', minimum follow-up of 24 months for survivingpatients, overall median 41.5 months).
We used a k-nearest neighbours (k-NN) algorithm26 that com-putes the distance of a test sample to each of the training set samples,each of which has an associated class (in this case, survivor orfailure), and then predicts the class of the test sample to be that ofthe majority of the k-closest samples. The k-NN classi®er wasevaluated by cross-validation, whereby one sample is randomlywithheld, a model is trained on the remaining samples, and themodel is then used to predict the class of the withheld sample. Theprocess is repeated until all of the samples are tested.
Outcome predictions based on gene expression were statisticallysigni®cant for k-NN models ranging from 2 to 21 genes, withoptimal predictions made by an 8-gene model that made only 13/60classi®cation errors (Fisher's exact test, P = 0.0002). Shown mostclearly by a Kaplan±Meier survival analysis in Fig. 4a, patients thatwere predicted to be survivors had a 5 year overall survival of 80%compared with 17% for patients predicted to have a poor outcome(P = 0.000003, log rank test). A more conservative method ofassessing statistical signi®cance is to attempt to optimize classi®ersof random permutations of the survivor/failure class labels. Weperformed 1,000 such permutations and found only 9 for whichprediction accuracy matched or exceeded our observed result(Supplementary Information III), indicating that the result isunlikely to be achieved by chance (P = 0.009). We subsequentlytested several other classi®cation algorithms including weightedvoting6,27, support vector machines28,29 and IBM SPLASH30, all ofwhich performed with similarly high accuracy (SupplementaryInformation I and III).
We explored further the clinical value of the predictor by con-sidering existing prognostic factors for medulloblastoma outcome.Patients with localized disease (M0) had a more favourable outcomecompared with patients with involvement of the cerebrospinal ¯uidor with distant metastases (M+) (P = 0.03 comparing M0 withM+ by Kaplan±Meier analysis), although not all M0 patientssurvived. When our outcome predictor was applied only to the42 M0 patients, the prediction of outcome remained signi®cant(P = 0.002), indicating that the expression-based predictor sub-stantially improved staging-based prognostication. Similarly, pre-diction based on TRKC expression was imperfect in this series inthat not all patients in the unfavourable (TRKC-low) category died.When our gene expression-based predictor was applied to the 33TRKC-low patients, the surviving patients could be signi®cantlyseparated from those who succumbed to their disease (P = 0.01,Supplementary Information III). Of note, not all patients in thisstudy received identical therapy. However, restricting our analysis tothe 35 patients that received surgery, vincristine, cisplatin andcyclophosphamide, the predictor continued to yield a signi®cantKaplan±Meier survival distinction (P = 0.0012). Taken together,these results demonstrate that the outcome predictor based on geneexpression exceeds other approaches to prognosis determination.
A number of genes not previously associated with clinical out-come were identi®ed (Fig. 4b). Those correlated with favourableoutcome included many genes characteristic of cerebellar differ-entiation (vesicle coat protein b-NAP, NSCL1, TRKC, sodiumchannels), and genes encoding extracellular matrix proteins(PLOD lysyl hydroxylase, collagen type V aI, elastin). As expected,TRKC expression was correlated with a favourable outcome,consistent with previous reports of this association22,23. In contrast,genes related to cerebellar differentiation were underexpressed inpoor prognosis tumours, which were dominated by the expressionof genes related to cell proliferation and metabolism (MYBL2,enolase 1, LDH, HMG1(Y), cytochrome C oxidase) and multidrugresistance (sorcin). Genes correlated with poor outcome included anumber of the ribosomal protein-encoding genes identi®ed by theSOM clustering experiments (Fig. 4b). This indicates that whereasthis ribosomal signature is correlated with poor outcome, optimaloutcome prediction requires not only these genes, but also genescorrelated with a favourable outcome that were not identi®ed by theunsupervised clustering analysis.
The routine clinical implementation of genomics-based outcomepredictors must await con®rmation in independent data sets, andthe models may need to be modi®ed as treatment regimens evolve.For patients predicted to have a favourable outcome, efforts tominimize toxicity of therapy might be indicated, whereas for thosepredicted not to respond to standard therapy, earlier treatment withexperimental regimens might be considered. This work illustrateshow genomic technologies have the potential to advance treatmentplanning beyond the empiric, towards a more molecularly de®ned,individualized approach to medicine. M
MethodsPatient samples
Patients included 60 children with medulloblastomas, 10 young adults with malignantgliomas (WHO grades III and IV), 5 children with AT/RTs, 5 with renal/extrarenalrhabdoid tumours, and 8 children with supratentorial PNETs (see SupplementaryInformation I). Medulloblastoma patients were treated with craniospinal irradiation to2,400±3,600 centiGray (cGy) with a tumour dose of 5,300±7,200 cGy. All patients withmedulloblastoma were treated with chemotherapy consisting of cisplatin and vincristine,plus combinations of carboplatin, etoposide, cyclophosphamide or lumustine (1-(2-chloroethyl)-3-cyclohexyl-1-nitrosourea, CCNU) (details in Supplementary InformationII). Samples were snap frozen in liquid nitrogen and stored at -80 8C. Studies were donewith approval of the Committee for Clinical Investigation of Boston Children's Hospital.The data were organized into three sets: data set A (42 samples containing 10 medullo-blastomas, 10 malignant gliomas, 10 AT/RTs, 8 PNETs and 4 normal cerebella); data set B(34 samples containing 9 desmoplastic medulloblastomas and 25 classic medulloblasto-mas); and data set C (60 samples containing 39 medulloblastoma survivors and 21treatment failures). The clinical attributes of each of the patients in the study are available
© 2002 Macmillan Magazines Ltd

letters to nature
442 NATURE | VOL 415 | 24 JANUARY 2002 | www.nature.com
in Supplementary Information II. Tissues were homogenized in guanidinium isothio-cyanate and RNA was isolated by centrifugation over a CsCl gradient. RNA integrity wasassessed either by northern blotting or by gel electrophoresis. Ten±twelve microgramstotal RNA was used to generate biotinlylated antisense RNAs, which were hybridizedovernight to HuGeneFL arrays containing 5,920 known genes and 897 expressed sequencetags, as described previously6. Arrays were scanned on Affymetrix scanners and theexpression value for each gene was calculated using GENECHIP software (Affymetrix,Santa Clara, California). Minor differences in microarray intensity were corrected using alinear scaling method as detailed in Supplementary Information I. Scans were rejected ifthe scaling factor exceeded 3, fewer than 1,000 genes received `present' calls, or microarrayartefacts were visible.
Preprocessing and clustering
The gene expression data were subjected to a variation ®lter that excluded genes showingminimal variation across the samples being analysed, as detailed in SupplementaryInformation I.
The data were ®rst normalized by standardizing each column (sample) to mean 0 andvariance 1. SOMs were performed using our GeneCluster clustering package (http://www.genome.wi.mit.edu/MPR/software). Hierarchical clustering was performed usingCluster and TreeView software5. Principal component analysis (PCA) was performed bycomputing and then plotting the three principal components using the S-Plus statisticalsoftware package (www.insightful.com/products/desktop.asp) using default settings.
Supervised learning
Genes correlated with particular class distinctions (for example, classic versus desmo-plastic medulloblastoma) were identi®ed by sorting all of the genes on the array accordingthe signal-to-noise statistic (m0 - m1)/(j0 + j1), where m and s represent the median andstandard deviation of expression, respectively, for each class. Similar results were obtainedusing a standard t-statistic as the metric ��m0 2 m1�=Î�j2
0=N0 � j21=N1��, where N repre-
sents the number of samples in each class (see Supplementary Information). Permutationof the column (sample) labels was performed to compare these correlations to what wouldbe expected by chance in 99% of the permutations. For classi®cation, we developed amodi®cation of the k-NN algorithm26 that predicts the class of a new data point bycalculating the euclidean distance (d) of the new sample to the k nearest samples (for theseexperiments we set k = 5) in the training set using normalized gene expression data, andselecting the class to be that of most of the k samples. The weight given to each neighbourwas 1/d. The k-NN models were evaluated by 60-fold `leave-one-out' cross-validation,whereby a training set of 59 samples was used to predict the class of a randomly withheldsample, and the cumulative error rate was recorded. We tested models with variablenumbers of genes (1±200, selected according to their correlation with the survivor versustreatment failure distinction in the training set) in this manner. An 8-gene k-NN outcomeprediction model yielded the lowest error rate, and was therefore used to generate Kaplan±Meier survival plots using S-Plus. Predictors using metastatic staging or TRKC expressionwere constructed by ®nding the decision boundary half way between the classes, (mclass0+ mclass1)/2 using either the staging values 0 versus 1, 2, 3, 4 or the continuous TRKCmicroarray gene expression levels, and then predicting the unknown sample according toits location with respect to that boundary.
Received 18 July; accepted 13 November 2001.
1. Rorke, L. B. The cerebellar medulloblastoma and its relationship to primitive neuroectodermal
tumors. J. Neuropathol. Exp. Neurol. 42, 1±15 (1983).
2. Kadin, M. E., Rubenstein, L. J. & Nelson, J. S. Neonatal cerebellar medulloblastoma originating from
the fetal external granular layer. J. Neuropath. Exp. Neurol. 29, 583±600 (1970).
3. Packer, R. J. et al. Treatment of children with medulloblastomas with reduced-dose craniospinal
radiation therapy and adjuvant chemotherapy: a children's cancer group study. J. Clin. Oncol. 17,
2127±2136 (1999).
4. Mardia, K. V., Kent, J. T. & Bibby, J. M. Multivariate Analysis (Academic, London, 1979).
5. Eisen, M. B., Spellman, P. T., Brown, P. O. & Botstein, D. Cluster analysis and display of genome-wide
expression patterns. Proc. Natl Acad. Sci. USA 95, 14863±14868 (1998).
6. Golub, T. R. et al. Molecular classi®cation of cancer: class discovery and class prediction by gene
expression monitoring. Science 286, 531±537 (1999).
7. Aruga, J. et al. A novel zinc ®nger protein, Zic, is involved in neurogenesis, especially in the cell lineage
of cerebellar granule cells. J. Neurochem. 63, 1880±1890 (1994).
8. Yokota, N. et al. Predominant expression of human Zic in cerebellar granule cell lineage and
medulloblastoma. Cancer Res. 56, 377±383 (1996).
9. Rorke, L. B., Packer, R. J. & Biegel, J. A. Central nervous system atypical teratoid/rhabdoid tumors of
infancy and childhood: de®nition of an entity. J. Neurosurg. 85, 56±65 (1996).
10. Biegel, J. A. et al. Germ-line and acquired mutations of INI1 in atypical teratoid and rhabdoid tumors.
Cancer Res. 59, 74±79 (1999).
11. Versteege, I. et al. Truncating mutations of hSNF5/INI1 in aggressive paediatric cancer. Nature 394,
203±206 (1998).
12. Hahn, H. et al. Mutations of the human homolog of Drosophila patched in the nevoid basal cell
carcinoma syndrome. Cell 85, 841±851 (1996).
13. Johnson, R. L. et al. Human homolog of patched, a candidate gene for the basal cell nevus syndrome.
Science 272, 1668±1671 (1996).
14. Pietsch, T. et al. Medulloblastomas of the desmoplastic variant carry mutations of the human
homologue of Drosophila patched. Cancer Res. 57, 2085±2088 (1997).
15. Raffel, C. et al. Sporadic medulloblastomas contain PTCH mutations. Cancer Res. 57, 842±845 (1997).
16. Xie, J. et al. Mutations of the PATCHED gene in several types of sporadic extracutaneous tumors.
Cancer Res. 57, 2369±2372 (1997).
17. Wechsler-Reya, R. J. & Scott, M. P. Control of neuronal precursor proliferation in the cerebellum by
Sonic Hedgehog. Neuron 22, 103±114 (1999).
18. Wetmore, C., Eberhart, D. E. & Curran, T. The normal patched allele is expressed in medulloblastomas
from mice with heterozygous germ-line mutation of patched. Cancer Res. 60, 2239±2246 (2000).
19. Giangaspero, F. et al. in World Health Organization Histological Classi®cation of Tumours of the Nervous
System (eds Kleihues, P. & Cavenee, W. K.) 129±137 (International Agency for Research on Cancer,
Lyon, 2000).
20. Murone, M., Rosenthal, A. & deSauvage, F. J. Sonic hedgehog signaling by the patched-smoothened
receptor complex. Curr. Biol. 28, 76±84 (1999).
21. Hahn, H. et al. Patched target IGF2 is indispensable for the formation of medulloblastoma and
rhabdomyosarcoma. J. Biol. Chem. 275, 28341±28344 (2000).
22. Segal, R. A., Goumnerova, L. C., Kwon, Y. K., Stiles, C. D. & Pomeroy, S. L. Expression of the
neurotrophin receptor TRKC is linked to a favorable outcome in medulloblastoma. Proc. Natl Acad.
Sci. USA 91, 12867±12871 (1994).
23. Kim, J. Y. H. et al. Activation of neurotrophin-3 receptor TRKC induces apoptosis in medulloblas-
tomas. Cancer Res. 59, 711±719 (1999).
24. MacDonald, T. J. et al. Expression pro®ling of medulloblastoma: PDGFRA and the RAS/MAPK
pathway as therapeutic targets for metastatic disease. Nature Genet. 29, 143±152 (2001).
25. Tamayo, P. et al. Interpreting patterns of gene expression with self-organizing maps: Methods and
application to hematopoietic differentiation. Proc. Natl Acad. Sci. USA 96, 2907±2912 (1999).
26. Dasarathy, V. B. (ed). Nearest Neighbor (NN) Norms: NN Pattern Classi®cation Techniques (IEEE
Computer Society Press, Los Alamitos, California, 1991).
27. Slonim, D. K. et al. in Proc. 4th Annu. Int. Conf. Computational Mol. Biol. 263±272 (ACM Press, New
York, 2000).
28. Mukherjee, S. et al. Support vector machine classi®cation of microarray data. CBCL paper 182/AI
memo 1676 (Massachusetts Institute of Technology, Cambridge, Massachusetts, 1999); also at http://
www.ai.mit.edu/projects/cbcl/publications/ps/cancer.ps.
29. Brown, M. P. S. et al. Knowledge-based analysis of microarray gene expression data by using support
vector machines. Proc. Natl Acad. Sci. USA 97, 262±267 (2000).
30. Califano, et al. in Proc. 8th Int. Conf. Intel. Syst. Mol. Biol. (eds Bourne, P. et al.) 75±85 (AAAI Press,
Menlo Park, CA, 2000.
Supplementary Information accompanies the paper on Nature's website(http://www.nature.com).
Acknowledgements
We thank members of the Whitehead/MIT Center for Genome Research, Program inCancer Genomics, and J. Volpe for discussions and comments on the manuscript. Thiswork was supported in part by Millennium Pharmaceuticals, Affymetrix and Bristol-Myers Squibb (E.S.L.); NIH grants (S.L.P. and T.C.); NIH-supported Mental RetardationResearch Center (S.L.P.) and Cancer Center Support CORE (T.C.); the American LebaneseSyrian Associated Charities (ALSAC); and the Kyle Mullarkey Medulloblastoma ResearchFund. We acknowledge the Cooperative Human Tissue Network and the Children'sOncology Group for contributing tumour samples.
Competing interests statement
The authors declare competing ®nancial interests: details accompany the paper onNature's website (http://www.nature.com).
Correspondence and requests for materials should be addressed to S.L.P.(e-mail: [email protected]) or T.R.G. (e-mail: [email protected]).
.................................................................A calcium sensor in the sodiumchannel modulates cardiacexcitabilityHanno L. Tan*², Sabina Kupershmidt³, Rong Zhang³§,Svetlana Stepanovic*, Dan M. Roden³§, Arthur A. M. Wilde²,Mark E. Anderson³§ & Jeffrey R. Balser*³
Departments of * Anesthesiology, § Medicine, and ³ Pharmacology,
Vanderbilt University School of Medicine, Nashville 37232, Tennessee, USA² The Experimental and Molecular Cardiology Group, Academic Medical Center,
University of Amsterdam, 1105 AZ, Amsterdam, the Netherlands
..............................................................................................................................................
Sodium channels are principal molecular determinants respon-sible for myocardial conduction and maintenance of the cardiacrhythm. Calcium ions (Ca2+) have a fundamental role in thecoupling of cardiac myocyte excitation and contraction, yetmechanisms whereby intracellular Ca2+ may directly modulateNa channel function have yet to be identi®ed. Here we show thatcalmodulin (CaM), a ubiquitous Ca2+-sensing protein, binds to
© 2002 Macmillan Magazines Ltd





























