Embed Size (px)
Citation preview

ix
Preface to Volume Two of the Second Edition ofBergey’s Manual� of Systematic Bacteriology
There is a long-standing tradition for the Editors of Bergey’s Man-ual to open their respective editions with the observation thatthe new edition is a departure from the earlier ones. As thisvolume goes to press, however, we recognize a need to deviatefrom this practice, by offering a separate preface to each volumewithin this edition. In part, this departure is necessary becausethe size and complexity of this edition far exceeded our expec-tations, as has the amount of time that has elapsed betweenpublication of the first volume of this edition and this volume.
Earlier, we noted that systematic procaryotic biology is a dy-namic field, driven by constant theoretical and methodologicaladvances that will ultimately lead to a more perfect and usefulclassification scheme. Clearly, the pace has been accelerating asevidenced in the super-linear rate at which new taxa are beingdescribed. Much of the increase can be attributed to rapid ad-vances in sequencing technology, which has brought about amajor shift in how we view the relationships among Bacteria andArchaea. While the possibility of a universally applicable naturalclassification was evident as the First Edition was in preparation,it is only recently that the sequence databases became largeenough, and the taxonomic coverage broad enough to makesuch an arrangement feasible. We have relied heavily upon thesedata in organizing the contents of this edition of Bergey’s Manualof Systematic Bacteriology, which will follow a phylogenetic frame-work based on analysis of the nucleotide sequence of the smallribosomal subunit RNA, rather than a phenotypic structure. Thisdeparts from the First Edition, as well as the Eighth and NinthEditions of the Determinative Manual. While the rationale for pre-senting the content of this edition in such a manner should beevident to most readers, they should bear in mind that this edi-tion, as in all preceding ones represents a progress report, ratherthan a final classification of procaryotes.
The Editors remind the readers that the Systematics Manual isa peer-reviewed collection of chapters, contributed by authorswho were invited by the Trust to share their knowledge andexpertise of specific taxa. Citation should refer to the author, thechapter title, and inclusive pages rather than to the Editors. TheTrust is indebted to all of the contributors and reviewers, withoutwhom this work would not be possible. The Editors are gratefulfor the time and effort that each expended on behalf of theentire scientific community. We also thank the authors for theirgood grace in accepting comments, criticisms, and editing of
their manuscripts. We would also like to thank Drs. Hans Truper,Brian Tindall, and Jean Euzeby for their assistance on mattersof nomenclature and etymology.
We would like to express our thanks to the Department ofMicrobiology and Molecular Genetics at Michigan State Univer-sity for housing our headquarters and editorial office and forproviding a congenial and supportive environment for microbialsystematics. We would also like to thank Connie Williams notonly for her expert secretarial assistance, but also for unflaggingdedication to the mission of Bergey’ s Manual Trust and Drs.Julia Bell and Denise Searles for their expert editorial assistanceand diligence in verifying countless pieces of critical informationand to Dr. Timothy G. Lilburn for constructing many of thephylogenetic trees used in this volume. We also extend our thanksto Alissa Wesche, Matt Chval and Kristen Johnson for their as-sistance in compilation of the bibliography.
A project such as the Systematics Manual also requires thestrong and continued support of a dedicated publisher, and wehave been most fortunate in this regard. We would also like toexpress our gratitude to Springer-Verlag for supporting our ef-forts and for the development of the Bergey’ s Document TypeDefinition (DTD). We would especially like to thank our Exec-utive Editor, Dr. William Curtis for his courage, patience, un-derstanding, and support; Catherine Lyons for her expertise indesigning and developing our DTD, and Jeri Lambert and LeslieGrossberg of Impressions Book and Journal Services for theirefforts during the pre-production and production phases. Wewould also like to acknowledge the support of ArborText, Inc.,for providing us with state-of-the-art SGML development and ed-iting tools at reduced cost. Lastly, I would like to express mypersonal thanks to my fellow trustees for providing me with theopportunity to participate in this effort, to Drs. Don Brenner,Noel Krieg, and James Staley for their enormous efforts as volumeeditors and to my wife, Nancy, and daughter, Jane, for their con-tinued patience, tolerance and support.
Comments on this edition are welcomed and should be di-rected to Bergey’s Manual Trust, Department of Microbiologyand Molecular Genetics, 6162 Biomedical and Physical SciencesBuilding, Michigan State University, East Lansing, MI, USA48824-4320. Email: [email protected]
George M. Garrity


xi
Preface to the First Edition of Bergey’s Manual� ofSystematic Bacteriology
Many microbiologists advised the Trust that a new edition of theManual was urgently needed. Of great concern to us was thesteadily increasing time interval between editions; this intervalreached a maximum of 17 years between the seventh and eightheditions. To be useful the Manual must reflect relatively recentinformation; a new edition is soon dated or obsolete in partsbecause of the nearly exponential rate at which new informationaccumulates. A new approach to publication was needed, andfrom this conviction came our plan to publish the Manual as asequence of four subvolumes concerned with systematic bacte-riology as it applies to taxonomy. The four subvolumes are di-vided roughly as follows: (a) the Gram-negatives of general, med-ical or industrial importance; (b) the Gram-positives other thanactinomycetes; (c) the archaeobacteria, cyanobacteria and re-maining Gram-negatives; and (d) the actinomycetes. The Trustbelieved that more attention and care could be given to prep-aration of the various descriptions within each subvolume, andalso that each subvolume could be prepared, published, andrevised as the area demanded, more rapidly than could be thecase if the Manual were to remain as a single, comprehensivevolume as in the past. Moreover, microbiologists would have theoption of purchasing only that particular subvolume containingthe organisms in which they were interested.
The Trust also believed that the scope of the Manual neededto be expanded to include more information of importance forsystematic bacteriology and bring together information dealingwith ecology, enrichment and isolation, descriptions of speciesand their determinative characters, maintenance and preserva-tion, all focused on the illumination of bacterial taxonomy. Toreflect this change in scope, the title of the Manual was changedand the primary publication becomes Bergey’ s Manual of SystematicBacteriology. This contains not only determinative material suchas diagnostic keys and tables useful for identification, but alsoall of the detailed descriptive information and taxonomic com-ments. Upon completion of each subvolume, the purely deter-minative information will be assembled for eventual incorpora-tion into a much smaller publication which will continue theoriginal name of the Manual, Bergey’ s Manual of DeterminativeBacteriology, which will be a similar but improved version of thepresent Shorter Bergey’ s Manual. So, in the end there will be twopublications, one systematic and one determinative in character.
An important task of the Trust was to decide which generashould be covered in the first and subsequent subvolumes. Wewere assisted in this decision by the recommendations of ourAdvisory Committees, composed of prominent taxonomic au-
thorities to whom we are most grateful. Authors were chosen onthe basis of constant surveillance of the literature of bacterialsystematics and by recommendations from our Advisory Com-mittees.
The activation of the 1976 Code had introduced some novelproblems. We decided to include not only those genera that hadbeen published in the Approved Lists of Bacterial Names in Jan-uary 1980 or that had been subsequently validly published, butalso certain genera whose names had no current standing innomenclature. We also decided to include descriptions of certainorganisms which had no formal taxonomic nomenclature, suchas the endosymbionts of insects. Our goal was to omit no im-portant group of cultivated bacteria and also to stimulate taxo-nomic research on “neglected” groups and on some groups ofundoubted bacteria that have not yet been cultivated and sub-jected to conventional studies.
The invited authors were provided with instructions and ex-emplary chapters in June 1980 and, although the intended dead-line for receipt of manuscripts was March 1981, all contributionswere assembled in January 1982 for the final preparations. TheManual was forwarded to the publisher in June 1982.
Some readers will note the consistent use of the stem -varinstead of -type in words such as biovar, serovar and pathovar.This is in keeping with the recommendations of the Bacterio-logical Code and was done against the wishes of some of theauthors.
We have deleted much of the synonymy of scientific nameswhich was contained in past editions. The adoption of the newstarting date of January 1, 1980 and publication of the ApprovedLists of Bacterial Names has made mention of past synonymyobsolete. We have included synonyms of a name only if theyhave been published since the new starting date, or if they werealso on the Approved Lists and, in rare cases with certain path-ogens, if the mention of an old name would help readers asso-ciate the organism with a clinical problem. If the reader is in-terested in tracing the history of a name we suggest he or sheconsult past editions of the Manual or the Index Bergeyana andits Supplement. In citations of names we have used the abbrevia-tion AL to denote the inclusion of the name on the ApprovedLists of Bacterial Names and VP to show the name has beenvalidly published.
In the matter of citation of the Manual in the scientific lit-erature we again stress the fact that the Manual is a collectionof authored chapters and the citation should refer to the author,the chapter title and its inclusive pages, not the Editor.

PREFACE TO THE FIRST EDITIONxii
To all contributors, the sincere thanks of the Trust is due; theEditor is especially grateful for the good grace with which theauthors accepted comments, criticisms and editing of their man-uscripts. It is only because of the voluntary and dedicated effortsof these authors that the Manual can continue to serve the sci-ence of bacteriology on an international basis.
A number of institutions and individuals deserve special ac-knowledgment from the Trust for their help in bringing aboutthe publication of this volume. We are grateful to the Departmentof Biology of the Virginia Polytechnic Institute and State Uni-versity for providing space, facilities and, above all, tolerance forthe diverted time taken by the Editor during the preparation ofthe book. The Department of Microbiology at Iowa State Uni-versity of Science and Technology continues to provide a wel-come home for the main editorial offices and archives of theTrust and we acknowledge their continued support. A grant(LM-03707) from the National Library of Medicine, National
Institutes of Health to assist in the preparation of this and thenext volume of the Manual is gratefully acknowledged.
A number of individuals deserve special mention and thanksfor their help. Professor Thomas O. McAdoo of the Departmentof Foreign Languages and Literatures at the Virginia PolytechnicInstitute and State University has given invaluable advice on theetymology and correctness of scientific names. Those assistingthe Editor in the Blacksburg office were R. Martin Roop II, DonD. Lee, Eileen C. Falk and Michael W. Friedman and their helpis sincerely appreciated. In the Ames office we were ably assistedby Gretchen Colletti and Diane Triggs during the early periodof preparation and by Cynthia Pease during the major portionof the editing process. Mrs. Pease has been responsible for theconstruction of the List of References and her willingness tohandle the cumbersome details of text editing on a big computeris gratefully acknowledged.
John G. Holt

xiii
Preface to the First Edition of Bergey’s Manual� ofDeterminative Bacteriology
The elaborate system of classification of the bacteria into families,tribes and genera by a Committee on Characterization and Clas-sification of the Society of American Bacteriologists (1911, 1920)has made it very desirable to be able to place in the hands ofstudents a more detailed key for the identification of species thanany that is available at present. The valuable book on “Deter-minative Bacteriology” by Professor F. D. Chester, published in1901, is now of very little assistance to the student, and all pre-vious classifications are of still less value, especially as earliersystems of classification were based entirely on morphologic char-acters.
It is hoped that this manual will serve to stimulate efforts toperfect the classification of bacteria, especially by emphasizingthe valuable features as well as the weaker points in the newsystem which the Committee of the Society of American Bacte-riologists has promulgated. The Committee does not regard theclassification of species offered here as in any sense final, butmerely a progress report leading to more satisfactory classifica-tion in the future.
The Committee desires to express its appreciation and thanksto those members of the society who gave valuable aid in thecompilation of material and the classification of certainspecies. . . .
The assistance of all bacteriologists is earnestly solicited in thecorrection of possible errors in the text; in the collection ofdescriptions of all bacteria that may have been omitted from thetext; in supplying more detailed descriptions of such organismsas are described incompletely; and in furnishing complete de-scriptions of new organisms that may be discovered, or in di-recting the attention of the Committee to publications of suchnewly described bacteria.
David H. Bergey, ChairmanFrancis C. HarrisonRobert S. BreedBernard W. HammerFrank M. HuntoonCommittee on Manual.August, 1923.


xv
Contents
Preface to Volume Two of the Second Edition of Bergey’s Manual� ofSystematic Bacteriology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ix
Preface to the First Edition of Bergey’s Manual� of SystematicBacteriology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi
Preface to the First Edition of Bergey’s Manual� of DeterminativeBacteriology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xiii
Contributors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xix
PHYLUM XIV.Proteobacteria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
Class III. Gammaproteobacteria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1Order I. Chromatiales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
Family I. Chromatiaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3Genus I. Chromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10Genus II. Allochromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12Genus III. Halochromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14Genus IV. Isochromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15Genus V. Lamprobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16Genus VI. Lamprocystis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18Genus VII. Marichromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20Genus VIII. Nitrosococcus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21Genus IX. Pfennigia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22Genus X. Rhabdochromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23Genus XI. Thermochromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Genus XII. Thioalkalicoccus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25Genus XIII. Thiocapsa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26Genus XIV. Thiococcus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28Genus XV. Thiocystis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29Genus XVI. Thiodictyon . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31Genus XVII. Thioflavicoccus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33Genus XVIII. Thiohalocapsa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34Genus XIX. Thiolamprovum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35Genus XX. Thiopedia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36Genus XXI. Thiorhodococcus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37Genus XXII. Thiorhodovibrio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38Genus XXIII. Thiospirillum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
Family II. Ectothiorhodospiraceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41Genus I. Ectothiorhodospira . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43Genus II. Arhodomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48Genus III. Halorhodospira . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49Genus IV. Nitrococcus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52Genus V. Thioalkalivibrio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56Genus VI. Thiorhodospira . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
Family III. Halothiobacillaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58Genus I. Halothiobacillus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58

CONTENTSxvi
Order II. Acidithiobacillales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60Family I. Acidithiobacillaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
Genus I. Acidithiobacillus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60Family II. Thermithiobacillaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
Genus I. Thermithiobacillus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62Order III. Xanthomonadales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
Family I. Xanthomonadaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63Genus I. Xanthomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63Genus II. Frateuria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91Genus III. Luteimonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 93Genus IV. Lysobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95Genus V. Nevskia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 101Genus VI. Pseudoxanthomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . 105Genus VII. Rhodanobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 106Genus VIII. Schineria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 106Genus IX. Stenotrophomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107Genus X. Thermomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 116Genus XI. Xylella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
Order IV. Cardiobacteriales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123Family I. Cardiobacteriaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123
Genus I. Cardiobacterium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123Genus II. Dichelobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 126Genus III. Suttonella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 130
Order V. Thiotrichales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131Family I. Thiotrichaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131
Genus I. Thiothrix . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131Genus II. Achromatium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142Genus III. Beggiatoa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 148Genus IV. Leucothrix . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162Genus V. Thiobacterium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169Genus VI. Thiomargarita . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 169Genus VII. Thioploca . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 171Genus VIII. Thiospira . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 178
Family II. Piscirickettsiaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180Genus I. Piscirickettsia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 180Genus II. Cycloclasticus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184Genus III. Hydrogenovibrio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 188Genus IV. Methylophaga . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 190Genus V. Thioalkalimicrobium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 193Genus VI. Thiomicrospira . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 193
Family III. Francisellaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199Genus I. Francisella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 200
Order VI. Legionellales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 210Family I. Legionellaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 210
Genus I. Legionella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 212Family II. Coxiellaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237
Genus I. Coxiella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237Genus II. Rickettsiella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 241
Order VII. Methylococcales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 248Family I. Methylococcaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 252
Genus I. Methylococcus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 254Genus II. Methylobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 258Genus III. Methylocaldum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261Genus IV. Methylomicrobium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 262Genus V. Methylomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 265Genus VI. Methylosarcina . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 268Genus VII. Methylosphaera . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 269

CONTENTS xvii
Order VIII. Oceanospirillales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 270Family I. Oceanospirillaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 271
Genus I. Oceanospirillum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 271Genus II. Balneatrix . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 282Genus III. Marinomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 284Genus IV. Marinospirillum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290Genus V. Neptunomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 292
Family II. Alcanivoraceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295Genus I. Alcanivorax . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 295
Family III. Hahellaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 299Genus I. Hahella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 300
Family IV. Halomonadaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 300Genus I. Halomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 300Genus II. Carnimonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 313Genus III. Chromohalobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 316Genus IV. Zymobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 319
Order IX. Pseudomonadales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 323Family I. Pseudomonadaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 323
Genus I. Pseudomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 323Genus II. Azomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 379Genus III. Azotobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 384Genus IV. Cellvibrio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 402Genus V. Mesophilobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 403Genus VI. Rhizobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 404Genus VII. Rugamonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 407Genus VIII. Serpens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 408
Family II. Moraxellaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 411Genus I. Moraxella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 417Genus II. Acinetobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 425Genus III. Psychrobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 437
Family III. Incertae Sedis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 441Genus I. Enhydrobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 441
Order X. Alteromonadales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 443Family I. Alteromonadaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 443
Genus I. Alteromonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 444Genus II. Alishewanella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 447Genus III. Colwellia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 447Genus IV. Ferrimonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 454Genus V. Glaciecola . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 456Genus VI. Idiomarina . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 458Genus VII. Marinobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 459Genus VIII. Marinobacterium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 464Genus IX. Microbulbifer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 464Genus X. Moritella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 465Genus XI. Pseudoalteromonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 467Genus XII. Psychromonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 478Genus XIII. Shewanella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 480
Order XI. “Vibrionales” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 491Family I. Vibrionaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 491
Genus I. Vibrio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 494Genus II. Photobacterium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 546Genus III. Salinivibrio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 552
Order XII. Aeromonadales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 556Family I. Aeromonadaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 556
Genus I. Aeromonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 557Genus II. Oceanimonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 578Genus Incertae Sedis III. Tolumonas . . . . . . . . . . . . . . . . . . . . . . . . . 579

CONTENTSxviii
Family II. Succinivibrionaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 581Genus I. Succinivibrio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 581Genus II. Anaerobiospirillum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 582Genus III. Ruminobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 584Genus IV. Succinimonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 586
Order XIII. “Enterobacteriales” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 587Family I. Enterobacteriaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 587
Genus I. Escherichia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 607Genus II. Alterococcus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 625Genus III. Arsenophonus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 626Genus IV. Brenneria . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 628Genus V. Buchnera . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 633Genus VI. Budvicia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 639Genus VII. Buttiauxella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 641Genus VIII. Calymmatobacterium . . . . . . . . . . . . . . . . . . . . . . . . . . . 645Genus IX. Cedecea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 648Genus X. Citrobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 651Genus XI. Edwardsiella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 657Genus XII. Enterobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 661Genus XIII. Erwinia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 670Genus XIV. Ewingella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 679Genus XV. Hafnia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 681Genus XVI. Klebsiella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 685Genus XVII. Kluyvera . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 694Genus XVIII. Leclercia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 698Genus XIX. Leminorella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 702Genus XX. Moellerella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 705Genus XXI. Morganella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 707Genus XXII. Obesumbacterium . . . . . . . . . . . . . . . . . . . . . . . . . . . . 710Genus XXIII. Pantoea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 713Genus XXIV. Pectobacterium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 721Genus XXV. “Candidatus Phlomobacter” . . . . . . . . . . . . . . . . . . . . . . 730Genus XXVI. Photorhabdus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 732Genus XXVII. Plesiomonas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 740Genus XXVIII. Pragia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 744Genus XXIX. Proteus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 745Genus XXX. Providencia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 753Genus XXXI. Rahnella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 759Genus XXXII. Saccharobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 762Genus XXXIII. Salmonella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 764Genus XXXIV. Serratia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 799Genus XXXV. Shigella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 811Genus XXXVI. Sodalis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 823Genus XXXVII. Tatumella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 825Genus XXXVIII. Trabulsiella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 827Genus XXXIX. Wigglesworthia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 828Genus XL. Xenorhabdus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 831Genus XLI. Yersinia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 838Genus XLII. Yokenella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 848
Order XIV. Pasteurellales . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 850Family I. Pasteurellaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 851
Genus I. Pasteurella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 857Genus II. Actinobacillus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 866Genus III. Haemophilus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 883Genus IV. Lonepinella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 904Genus V. Mannheimia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 907Genus VI. Phocoenobacter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 912
Bibliography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 913Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1085

FAMILY I. OCEANOSPIRILLACEAE 271
Family I. Oceanospirillaceae fam. nov.GEORGE M. GARRITY, JULIA A. BELL AND TIMOTHY LILBURN
O.ce.an.o.spi.ril.la!ce.ae. M.L. neut. n. Oceanospirillum type genus of the family; -aceae endingto denote family; M.L. fem. pl. n. Oceanospirillaceae the Oceanospirillum family.
The family Oceanospirillaceae was circumscribed for this volumeon the basis of phylogenetic analysis of 16S rDNA sequences; thefamily contains the genera Oceanospirillum (type genus), Balnea-trix, Marinomonas, Marinospirillum, Neptunomonas, Oceanobacter,Oleispira, and Pseudospirillum. Oceanobacter, Pseudospirillum andOleispira were proposed after the cut-off date for inclusion in thisvolume ( June 30, 2001) and are not described here (see Satomiet al. (2002) and Yakimov et al. (2003a), respectively).
Motile by polar flagella. Aerobic; strictly respiratory exceptfor Neptunomonas, which gives weak fermentation reactions.Aquatic; Balneatrix is found in fresh water, whereas other generaare marine.
Type genus: Oceanospirillum Hylemon, Wells, Krieg and Jan-nasch 1973, 361AL.
Genus I. Oceanospirillum Hylemon, Wells, Krieg and Jannasch 1973, 361AL*
BRUNO POT AND MONIQUE GILLIS
O.ce.an.o.spi.ril!lum. M.L. n. oceanus ocean; Gr. n. spira a spiral; M.L. dim. neut. n. spirillum spirillum asmall spiral; Oceanospirillum a small spiral (organism) from the ocean (seawater).
Rigid, helical cells 0.4–1.2 lm in diameter. Motile by bipolar tuftsof flagella. A polar membrane underlies the cytoplasmic mem-brane at the cell poles in all species so far examined by electronmicroscopy. Intracellular poly-b-hydroxybutyrate is formed. Allspecies form thin-walled coccoid bodies that predominate in oldcultures. Gram negative. Aerobic, having a strictly respiratorytype of metabolism with oxygen as the terminal electron acceptor.Nitrate respiration does not occur. Nitrate can be reduced tonitrite in all oceanospirilla. Optimum temperature for growth,25–32!C. Oxidase positive. Indole and aryl sulfatase negative.Casein, starch, hippurate, and esculin are not hydrolyzed. Sea-water is required for growth. Carbohydrates are neither oxidizednor fermented. Amino acids or the salts of organic acids serveas carbon sources. Growth factors are not usually required. Iso-lated from coastal seawater, decaying seaweed, and putrid infu-sions of marine mussels.
The mol% G " C of the DNA is: 45–50.Type species: Oceanospirillum linum (Williams and Rittenberg
1957) Hylemon, Wells, Krieg and Jannasch 1973, 374 (Spirillumlinum Williams and Rittenberg 1957, 82.)
FURTHER DESCRIPTIVE INFORMATION
All species of Oceanospirillum consist of helical cells; however,variants having less curvature may arise after prolonged transfer.For example, the type strain of O. japonicum consisted initially oflong, helical cells with several turns (Watanabe, 1959), but nowconsists of slightly curved or S-shaped cells. The cells have aconstant and characteristic type of clockwise (right-handed) he-lix. Only one phylogenetically unrelated species (O. pusillum) has
*Editorial Note: The genus Oceanospirillum has recently undergone taxonomic re-evaluation. O. minutulum has been transferred to the genus Marinospirillum as Ma-rinospirillum minutulum (Satomi et al., 1998); see the Taxonomic Comments sectionin this chapter. O. commune and O. vagum are homotypic synonyms of Marinomonascommunis and Marinomonas vaga, respectively. After the cut-off date for inclusion oftaxonomic changes in this volume of the Systematics, Satomi et al. (2002) emendedthe description of Oceanospirillum and proposed the transfer of O. jannaschii to thegenus Marinobacterium as Marinobacterium jannaschii, the transfer of O. japonicum tothe new genus Pseudospirillum as Pseudospirillum japonicum, the transfer of O. kriegiito the new genus Oceanobacter as Oceanobacter kriegii, and the transfer of O. pusillumto the new genus Terasakiella as Terasakiella pusilla.
a counterclockwise (left-handed) helix (Terasaki, 1972). Photo-graphs showing the size and shape of various species of ocean-ospirilla and O. minutulum (now Marinospirillum minutulum) arepresented in Fig. BXII.c.104.
An unusual elaboration of the plasma membrane, the “polarmembrane” , occurs in all of the species so far examined (Bev-eridge and Murray, unpublished results). It is attached to theinside of the plasma membrane by bar-like links and is located,most commonly, in the region surrounding the polar flagella(Murray and Birch-Andersen, 1963). Such a membrane has beenfound mainly in genera of helical bacteria, such as Spirillum,Campylobacter, Aquaspirillum, Ectothiorhodospira, and Rhodospirillum.
All species have intracellular poly-b-hydroxybutyrate, but gran-ules may not be evident in cells having a small diameter andchemical analysis may be required to demonstrate the polymer.
All species have bipolar tufts of flagella and all species showextensive formation of coccoid bodies (sometimes termed “mi-crocysts”) in old cultures. These bodies have thin walls and re-semble spheroplasts; however, they are resistant to lysis in distilledwater (Kelly, 1959). Whether coccoid bodies are resistant to des-iccation is not known. Three main modes of formation of coccoidbodies were described by Williams and Rittenberg (1957), asfollows: (a) two cells may entwine and apparently fuse. The cellsbecome shorter and thicker and a protuberance develops at thepoint of fusion. This gradually enlarges and absorbs the organ-isms to form the coccoid body. More than one coccoid body maydevelop from a pair of entwined spirilla; (b) a spirillum maybecome shorter and thicker and a protuberance arises from thecenter of the cell or from each end of the cell. The protuberancesenlarge and eventually merge into a single coccoid body as thehelical cell is absorbed; (c) a spirillum may undergo a gradualshortening and rounding to form a coccoid body. The majorityof coccoid bodies present in old cultures appears to be viableand can “germinate” when placed into a fresh medium (Williamsand Rittenberg, 1956). Germination is by unipolar or bipolargrowth of a helical cell from the coccoid body, with the latterbeing absorbed into the developing helical cell.
Seawater is required for the growth of all species. Media pre-pared with natural seawater or with 2.75% NaCl have been usedfor enrichment and isolation (Williams and Rittenberg, 1957;Terasaki, 1963, 1970, 1980). Commonly used culture media for
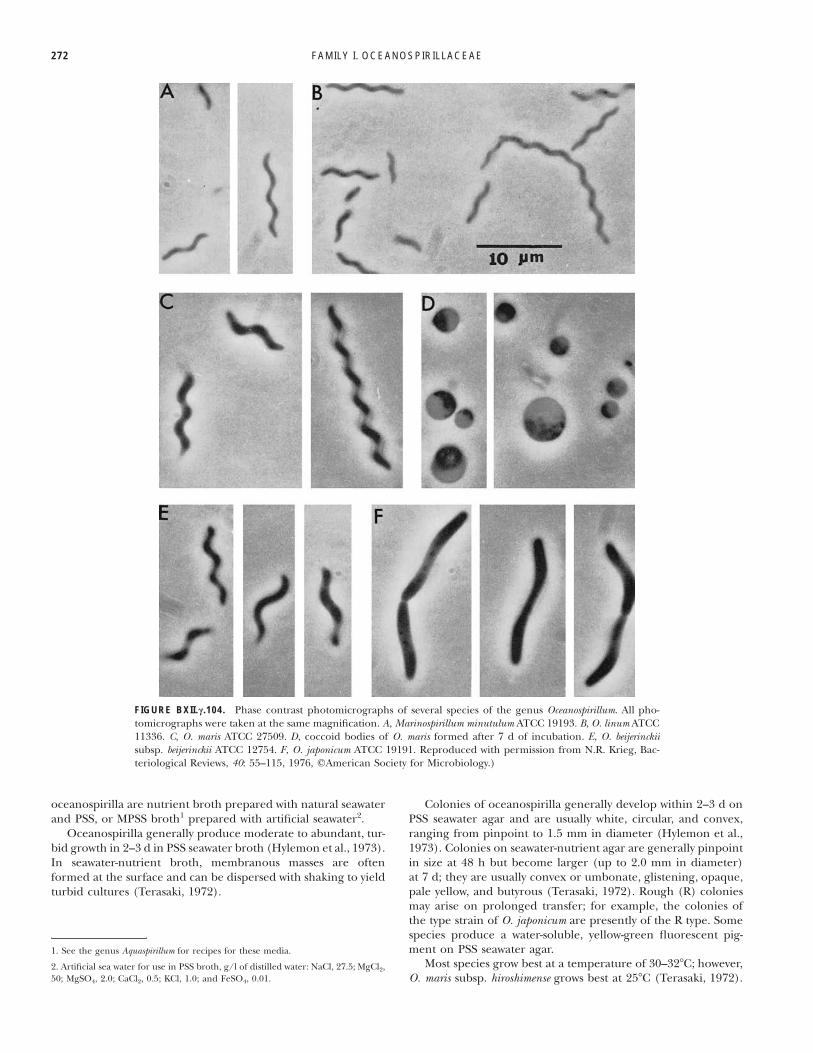
FAMILY I. OCEANOSPIRILLACEAE272
FIGURE BXII.c.104. Phase contrast photomicrographs of several species of the genus Oceanospirillum. All pho-tomicrographs were taken at the same magnification. A, Marinospirillum minutulum ATCC 19193. B, O. linum ATCC11336. C, O. maris ATCC 27509. D, coccoid bodies of O. maris formed after 7 d of incubation. E, O. beijerinckiisubsp. beijerinckii ATCC 12754. F, O. japonicum ATCC 19191. Reproduced with permission from N.R. Krieg, Bac-teriological Reviews, 40: 55–115, 1976, #American Society for Microbiology.)
oceanospirilla are nutrient broth prepared with natural seawaterand PSS, or MPSS broth1 prepared with artificial seawater2.
Oceanospirilla generally produce moderate to abundant, tur-bid growth in 2–3 d in PSS seawater broth (Hylemon et al., 1973).In seawater-nutrient broth, membranous masses are oftenformed at the surface and can be dispersed with shaking to yieldturbid cultures (Terasaki, 1972).
1. See the genus Aquaspirillum for recipes for these media.
2. Artificial sea water for use in PSS broth, g/l of distilled water: NaCl, 27.5; MgCl2,50; MgSO4, 2.0; CaCl2, 0.5; KCl, 1.0; and FeSO4, 0.01.
Colonies of oceanospirilla generally develop within 2–3 d onPSS seawater agar and are usually white, circular, and convex,ranging from pinpoint to 1.5 mm in diameter (Hylemon et al.,1973). Colonies on seawater-nutrient agar are generally pinpointin size at 48 h but become larger (up to 2.0 mm in diameter)at 7 d; they are usually convex or umbonate, glistening, opaque,pale yellow, and butyrous (Terasaki, 1972). Rough (R) coloniesmay arise on prolonged transfer; for example, the colonies ofthe type strain of O. japonicum are presently of the R type. Somespecies produce a water-soluble, yellow-green fluorescent pig-ment on PSS seawater agar.
Most species grow best at a temperature of 30–32!C; however,O. maris subsp. hiroshimense grows best at 25!C (Terasaki, 1972).

GENUS I. OCEANOSPIRILLUM 273
The nutrition of oceanospirilla is generally simple. Most spe-cies grow in simple defined media with amino acids or the saltsof organic acids as carbon sources and ammonium ions as thenitrogen source. However, O. linum is specifically stimulated bymethionine in a medium containing succinate and malate ascarbon sources, and O. maris subsp. williamsae has a growth factorrequirement that has not yet been identified. A listing of thecarbon sources for oceanospirilla is given below in TableBXII.c.77. Some apparent contradictions occur between the re-sults obtained from different laboratories, although the resultswithin each laboratory are reproducible. These differences arelikely attributable to differences in definitions of what constitutesa positive growth response, and in some cases to the use of dif-ferent strains.
The use of antisera in agglutination tests with a limited num-ber of strains has indicated that the species of Oceanospirillumcan be distinguished serologically (McElroy and Krieg, 1972).The antisera were prepared against whole cells and adsorbedwith heated cells, leaving only antibodies against thermolabileantigens.
Oceanospirilla have been isolated from coastal seawater (Wil-liams and Rittenberg, 1957), decaying seaweed ( Jannasch, 1963),and putrid infusions of marine mussels (Terasaki, 1963, 1970,1980). By direct microscopic counts of the bacteria present inclear and turbid seawaters near Port Aransas, Texas, Oppenhei-mer and Jannasch (1962) found that spirilla comprised only 0.1–2.5% of the total bacteria present. Whether oceanospirilla occurin the open sea is not known. Based on chemostat experiments,Jannasch (1963) suggested that the growth of oceanospirillamight be restricted to environments of higher nutrient concen-tration than is found in ordinary seawater, such as in zones sur-rounding decaying particulate matter. With regard to occurrenceof oceanospirilla in putrid infusions of marine mussels, thesource is most likely marine mud adherent to the mussels (Ter-asaki, 1970).
ENRICHMENT AND ISOLATION PROCEDURES
The enrichment and isolation method used by Williams and Rit-tenberg (1957) is as follows. A seawater sample is mixed with anequal volume of Giesberger’ s base medium (NH4Cl, 0.1%;K2HPO4, 0.05%; MgSO4, 0.05%) plus 1.0% calcium lactate. Afterincubation and appearance of spirilla, a portion of the initialculture is sterilized and mixed with an equal volume of sterileGiesberger’ s medium lacking NH4Cl. This mixture is then in-oculated from the unsterilized portion of the initial culture. Oneto three subcultures done in this manner are sufficient to estab-lish the spirilla as the predominant type. For isolation, the en-richment is diluted 1:100 to 1:100,000 with sterile seawater. Thedilution bottles are shaken vigorously and allowed to stand atroom temperature for 20 min to allow migration of spirilla tothe surface of the dilution. Isolation is then accomplished bystreaking the surface water onto a suitable agar medium such asnutrient agar prepared with seawater and containing 0.3% yeastautolysate. Plates are incubated at 30!C and after 24 h examinedfor distinctive, granular, umbonate or pulvinate colonies with aground-glass appearance.
The method of Terasaki (1970) has yielded excellent resultsfor the isolation of oceanospirilla from putrid infusions. Marinemussels are smashed with a hammer and placed in a Petri dishwith a teaspoon of marine mud. Sterilized seawater is pouredinto the dish until the mussels sink completely in the solution.The infusion is incubated at 27–28!C and examined for the de-
velopment of spirilla after 1, 2, 4, and 7 d. Isolation is accom-plished by streaking dilutions onto suitable agar media.
For enrichment by use of continuous cultures, see Jannasch(1967).
MAINTENANCE PROCEDURES
Oceanospirilla may be maintained in semisolid PSS seawater me-dium (containing 0.15% agar to give a jelly-like consistency) at30!C with weekly transfer (Hylemon et al., 1973). Cultures mayalso be maintained as stabs in seawater-nutrient agar at roomtemperature with monthly transfer (Terasaki, 1972).
Preservation is most easily accomplished by suspending adense concentration of cells in seawater-nutrient broth contain-ing 10% (v/v) dimethylsulfoxide, with subsequent freezing inliquid nitrogen. A method for freeze-drying oceanospirilla hasbeen reported by Terasaki (1975).
PROCEDURES FOR TESTING SPECIAL CHARACTERS
Characterization methods for oceanospirilla have been describedin detail by Terasaki (1972, 1979) and Hylemon et al. (1973).The comments given in this Manual for the genus Aquaspirillumalso apply to the genus Oceanospirillum, except that media con-taining natural or artificial seawater must be used for all char-acterization tests.
DIFFERENTIATION OF THE GENUS OCEANOSPIRILLUM FROM
OTHER GENERA
See the genus Aquaspirillum, in Volume 2 Part C in this Manual,for characteristics of Oceanospirillum that distinguish the genusfrom other morphologically or physiologically similar genera.
TAXONOMIC COMMENTS
In the eighth edition of Bergey’ s Manual of Determinative Bacteri-ology (Krieg, 1974), a single genus, Spirillum, contained all of thevarious aerobic and microaerophilic spirilla, including freshwaterand marine species. However, the DNA base composition for thegenus ranged from 38 to 65 mol% G " C and appeared to beunusually broad for a bacterial genus. Moreover, three groupswere evident within the genus: (a) the aerobic, freshwater spirillathat could not tolerate 3% NaCl (mol% G " C 50–65); (b) theaerobic, marine spirilla that required seawater for growth (mol%G " C $ 42–48); and (c) the large, microaerophilic spirilla thatbelong to the species S. volutans (mol% G " C $ 38). Accord-ingly, Hylemon et al. (1973) divided the genus into the threegenera Spirillum, Aquaspirillum, and Oceanospirillum, with the ma-rine organisms comprising the latter genus. This subdivision wasused in the first edition of Bergey’ s Manual of Systematic Bacteriology(Krieg, 1984a). Although this scheme proved useful for practicalpurposes, it was only gradually that the phylogenetic aspects ofthe three subdivisions were revealed.
In an analysis of the 16S rRNA oligonucleotide catalogs of thespecies O. minutulum (now Marinospirillum minutulum) and Oce-anospirillum maris, Woese et al. (1982) found that both organismsbelonged to group III of the phototrophic bacteria as definedby Gibson et al. (1979), but they were not closely related to eachother. Later, Woese et al. (1985) studied three additional speciesof Oceanospirillum—O. japonicum, O. linum, and O. beijerinckii.These species, together with the families Enterobacteriaceae andVibrionaceae, constituted the core of ‘subgroup 3’ of the Gam-maproteobacteria (Stackebrandt et al., 1988).
An organism known as “Spirillum lunatum” (Williams and Rit-
FAMILY I. OCEANOSPIRILLACEAE274
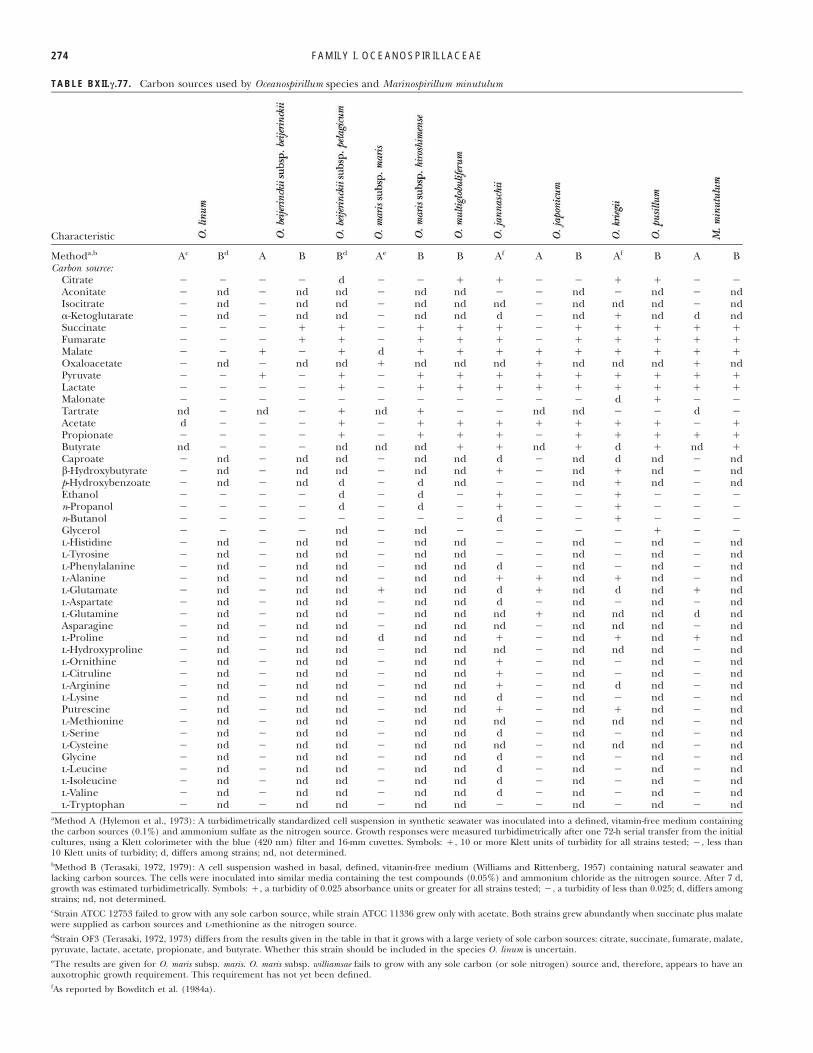
TABLE BXII.c.77. Carbon sources used by Oceanospirillum species and Marinospirillum minutulum
Characteristic
Methoda,b Ac Bd A B Bd Ae B B Af A B Af B A BCarbon source:
Citrate % % % % d % % " " % % " " % %Aconitate % nd % nd nd % nd nd % % nd % nd % ndIsocitrate % nd % nd nd % nd nd nd % nd nd nd % nd!-Ketoglutarate % nd % nd nd % nd nd d % nd " nd d ndSuccinate % % % " " % " " " % " " " " "Fumarate % % % " " % " " " % " " " " "Malate % % " % " d " " " " " " " " "Oxaloacetate % nd % nd nd " nd nd nd " nd nd nd " ndPyruvate % % " % " % " " " " " " " " "Lactate % % % % " % " " " " " " " " "Malonate % % % % % % % % % % % d " % %Tartrate nd % nd % " nd " % % nd nd % % d %Acetate d % % % " % " " " " " " " % "Propionate % % % % " % " " " % " " " " "Butyrate nd % % % nd nd nd " " nd " d " nd "Caproate % nd % nd nd % nd nd d % nd d nd % ndb-Hydroxybutyrate % nd % nd nd % nd nd " % nd " nd % ndp-Hydroxybenzoate % nd % nd d % d nd % % nd " nd % ndEthanol % % % % d % d % " % % " % % %n-Propanol % % % % d % d % " % % " % % %n-Butanol % % % % % % % % d % % " % % %Glycerol % % % % nd % nd % % % % % " % %l-Histidine % nd % nd nd % nd nd % % nd % nd % ndl-Tyrosine % nd % nd nd % nd nd % % nd % nd % ndl-Phenylalanine % nd % nd nd % nd nd d % nd % nd % ndl-Alanine % nd % nd nd % nd nd " " nd " nd % ndl-Glutamate % nd % nd nd " nd nd d " nd d nd " ndl-Aspartate % nd % nd nd % nd nd d % nd % nd % ndl-Glutamine % nd % nd nd % nd nd nd " nd nd nd d ndAsparagine % nd % nd nd % nd nd nd % nd nd nd % ndl-Proline % nd % nd nd d nd nd " % nd " nd " ndl-Hydroxyproline % nd % nd nd % nd nd nd % nd nd nd % ndl-Ornithine % nd % nd nd % nd nd " % nd % nd % ndl-Citruline % nd % nd nd % nd nd " % nd % nd % ndl-Arginine % nd % nd nd % nd nd " % nd d nd % ndl-Lysine % nd % nd nd % nd nd d % nd % nd % ndPutrescine % nd % nd nd % nd nd " % nd " nd % ndl-Methionine % nd % nd nd % nd nd nd % nd nd nd % ndl-Serine % nd % nd nd % nd nd d % nd % nd % ndl-Cysteine % nd % nd nd % nd nd nd % nd nd nd % ndGlycine % nd % nd nd % nd nd d % nd % nd % ndl-Leucine % nd % nd nd % nd nd d % nd % nd % ndl-Isoleucine % nd % nd nd % nd nd d % nd % nd % ndl-Valine % nd % nd nd % nd nd d % nd % nd % ndl-Tryptophan % nd % nd nd % nd nd % % nd % nd % nd
aMethod A (Hylemon et al., 1973): A turbidimetrically standardized cell suspension in synthetic seawater was inoculated into a defined, vitamin-free medium containingthe carbon sources (0.1%) and ammonium sulfate as the nitrogen source. Growth responses were measured turbidimetrically after one 72-h serial transfer from the initialcultures, using a Klett colorimeter with the blue (420 nm) filter and 16-mm cuvettes. Symbols: ", 10 or more Klett units of turbidity for all strains tested; %, less than10 Klett units of turbidity; d, differs among strains; nd, not determined.bMethod B (Terasaki, 1972, 1979): A cell suspension washed in basal, defined, vitamin-free medium (Williams and Rittenberg, 1957) containing natural seawater andlacking carbon sources. The cells were inoculated into similar media containing the test compounds (0.05%) and ammonium chloride as the nitrogen source. After 7 d,growth was estimated turbidimetrically. Symbols: ", a turbidity of 0.025 absorbance units or greater for all strains tested; %, a turbidity of less than 0.025; d, differs amongstrains; nd, not determined.cStrain ATCC 12753 failed to grow with any sole carbon source, while strain ATCC 11336 grew only with acetate. Both strains grew abundantly when succinate plus malatewere supplied as carbon sources and l-methionine as the nitrogen source.dStrain OF3 (Terasaki, 1972, 1973) differs from the results given in the table in that it grows with a large veriety of sole carbon sources: citrate, succinate, fumarate, malate,pyruvate, lactate, acetate, propionate, and butyrate. Whether this strain should be included in the species O. linum is uncertain.eThe results are given for O. maris subsp. maris. O. maris subsp. williamsae fails to grow with any sole carbon (or sole nitrogen) source and, therefore, appears to have anauxotrophic growth requirement. This requirement has not yet been defined.fAs reported by Bowditch et al. (1984a).

GENUS I. OCEANOSPIRILLUM 275
tenberg, 1957) was included in the genus Oceanospirillum by Hy-lemon et al. (1973), but this posed taxonomic problems. Thecharacteristics of the type strain (ATCC 11337 or NCMB 54) didnot fit the original description of the species, and Linn and Krieg(1978) found that NCMB strain 54 consisted of a mixture of twodissimilar organisms. The first type was a short, vibrioid rod thatpossessed a single polar flagellum, grew in either the presenceor absence of seawater, catabolized sugars, did not form coccoidbodies, and had a mol% G " C of 63–64. The second type wasa larger, helical organism that possessed bipolar flagellar tufts,required seawater, failed to attack sugars, formed coccoid bodies,and had a mol% G " C of 45. The smaller organism did notappear to belong to either Oceanospirillum or Aquaspirillum andit remains unclassified. The larger organism had characteristicsmore in accord with the original description of “S. lunatum” butdiffered in certain respects; it has been classified as a new sub-species of O. maris: O. maris subsp. williamsae.
Bowditch et al. (1984a, b) added four species to the genusOceanospirillum, mainly based on immunological relationships.These species were Oceanospirillum commune, for the organismpreviously named Marinomonas communis (Van Landschoot andDe Ley, 1983, 1984), Oceanospirillum vagum for Marinomonas vaga(Van Landschoot and De Ley, 1983, 1984), and two species Oce-anospirillum jannaschii and Oceanospirillum kriegii for two groupsof unnamed marine bacteria I-1 and H-1, respectively. As a result,the genus definition of Oceanospirillum needed to be changeddrastically, with the unfortunate loss of most of the readily de-terminable phenotypic features from the genus definition (Krieg,1984a) and the extension of the upper mol% G " C limit forthe genus from 51 to 57. By this extension, a considerable overlapof mol% G " C range was introduced between the genera Aqua-spirillum (49–65 mol% G " C) and Oceanospirillum (42–51 mol%G " C). In this way, one of the most reliable genotypic featuresdiscriminating both genera was lost. Phylogenetic data (Pot etal. 1989, Pot, 1996; Satomi et al., 1998), however, have sinceshown that all four species cannot be regarded as members ofthe genus Oceanospirillum.
On the basis of a polyphasic approach including DNA–DNAand DNA–rRNA hybridizations, Pot et al. (1989) showed thatonly five species, including the type species, constituted a sepa-rate rRNA branch in the Gammaproteobacteria and redefined thegenus Oceanospirillum to contain O. linum, O. maris, O. beijerinckii,O. multiglobuliferum, and, more distantly, O. japonicum. Based on
DNA–DNA hybridizations (as suggested by Krieg, 1984a) andnumerical comparison of whole-cell proteins, O. maris subsp. hi-roshimense and O. beijerinckii subsp. pelagicum were created for theformer species O. hiroshimense and O. pelagicum. O. pusillum wasshown to belong to the Alphaproteobacteria, and O. commune andO. vagum were relegated to their original generic positions asMarinomonas communis and Marinomonas vaga, respectively. Thetwo species O. jannaschii and O. kriegii were shown to be phylo-genetically too remote to be considered members of the genusOceanospirillum, and, together with O. minutulum, they constitutedseparate rRNA branches in the Gammaproteobacteria.
Subsequently, this phylogenetic heterogeneity was confirmedby studies of fatty acid, quinone, and polyamine compositions(Hamana et al., 1994; Sakane and Yokota, 1994; Bertone et al.,1996). All species, except O. pusillum, contained ubiquinone-8(Q-8) as a major respiratory quinone (Table BXII.c.78). Likeother spirilla from the Alphaproteobacteria (see the genus Aqua-spirillum in this book), O. pusillum contained over 90% Q-10. Thethirteen strains of Oceanospirillum that have been investigated fortheir fatty acid composition by Sakane and Yokota were dividedinto three groups (Table BXII.c.79 and BXII.c.80). Group I in-cluded the 10 strains belonging to O. linum, O. maris subsp. hi-roshimense, O. maris subsp. williamsae, O. beijerinckii subsp. beijer-inckii, O. beijerinckii subsp. pelagicum, O. multiglobuliferum, and O.japonicum, all of which have a low mol% G " C (42.5–48.4).Group II included the two type strains of O. jannaschii and O.kriegii and had a high mol% G " C content (54.8–54.9). GroupIII included only O. pusillum and could be clearly distinguishedfrom other marine spirilla in having C14:0 3OH as the major 3-hydroxy fatty acid, besides Q-10 (Table BXII.c.80). Bertone et al.(1996) confirmed the separate position of O. japonicum, O. jan-naschii, and O. kriegii.
All Oceanospirillum species including O. jannaschii and O. kriegiicontain both putrescine and spermidine. The relative content(Table BXII.c.81) of putrescine is very small when comparedwith the level found in members of the Alphaproteobacteria. Therelative concentration of putrescine for O. pusillum correspondswith that of other members of the Alphaproteobacteria. The absenceof 2-hydroxy putrescine and homospermidine is a unifying char-acter for the Gammaproteobacteria. The polyamine profile of Oce-anospirillum I and II is not different, nor are their fatty acidprofiles.
Later, 16S rDNA sequence analysis of all Oceanospirillum spe-
TABLE BXII.c.78. Cellular quinone systems in Oceanospirillum species and Marinospirillum minutuluma
Speciesb Strain GroupQuinone system
Q-6 Q-7 Q-8 Q-9 Q-10
O. linum IFO 15448T Ic 3 4 91 2IFO 15449 Ic 1 2 96 1
O. beijerinckii subsp. beijerinckii IFO 15445T Id 1 12 83 4O. beijerinckii subsp. pelagicum IFO 13612T Id 2 7 91 1O. maris subsp. hiroshimense IFO 13616T Ic 1 4 94 1O. maris subsp. williamsae IFO 15468T Ic 12 7 80 1O. multiglobuliferum IFO 13614T Ic 1 4 94 1O. jannaschii IFO 15466T IIb 3 7 89 1O. japonicum IFO 15446T Ib 1 14 84 1
IFO 15447 Ib 9 88 3O. kriegii IFO 15467T IIa 2 5 89 4O. pusillum IFO 13613T III 1 6 93Marinospirillum minutulum IFO 15450T Ia 1 1 97 1aAfter Sakane and Yokota (1994).bOceanospirillum maris subsp. maris has not been investigated.
FAMILY I. OCEANOSPIRILLACEAE276
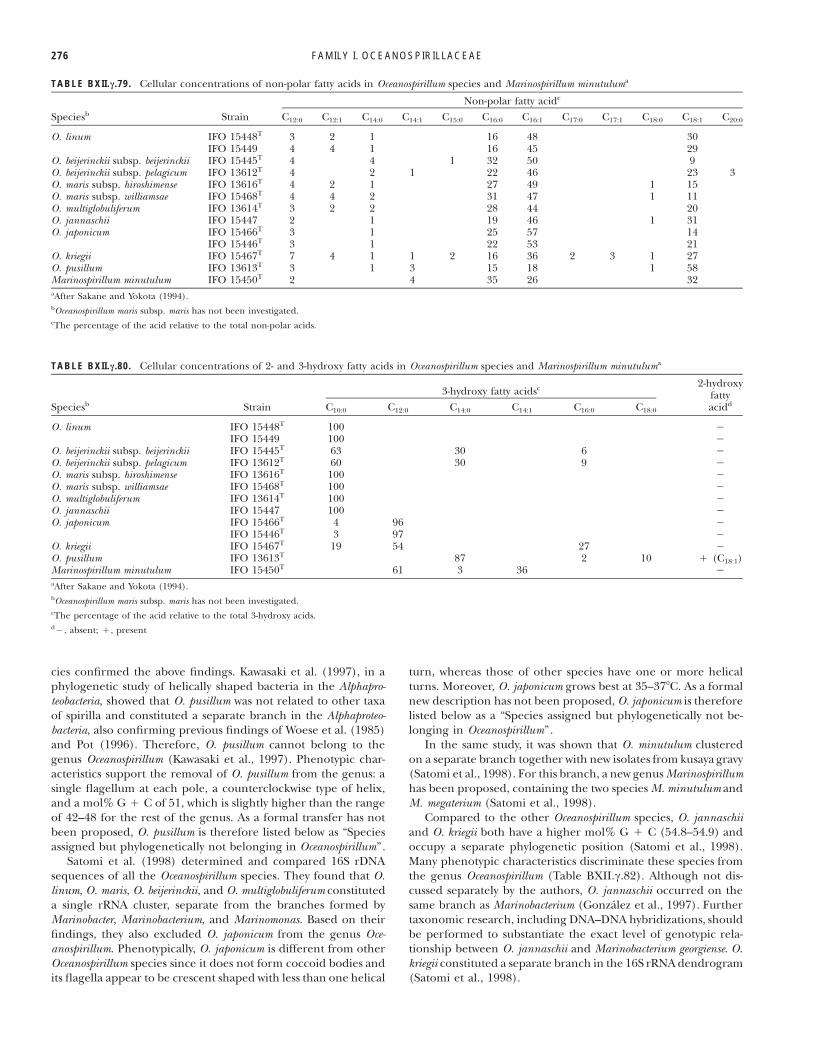
TABLE BXII.c.79. Cellular concentrations of non-polar fatty acids in Oceanospirillum species and Marinospirillum minutuluma
Speciesb StrainNon-polar fatty acidc
C12:0 C12:1 C14:0 C14:1 C15:0 C16:0 C16:1 C17:0 C17:1 C18:0 C18:1 C20:0
O. linum IFO 15448T 3 2 1 16 48 30IFO 15449 4 4 1 16 45 29
O. beijerinckii subsp. beijerinckii IFO 15445T 4 4 1 32 50 9O. beijerinckii subsp. pelagicum IFO 13612T 4 2 1 22 46 23 3O. maris subsp. hiroshimense IFO 13616T 4 2 1 27 49 1 15O. maris subsp. williamsae IFO 15468T 4 4 2 31 47 1 11O. multiglobuliferum IFO 13614T 3 2 2 28 44 20O. jannaschii IFO 15447 2 1 19 46 1 31O. japonicum IFO 15466T 3 1 25 57 14
IFO 15446T 3 1 22 53 21O. kriegii IFO 15467T 7 4 1 1 2 16 36 2 3 1 27O. pusillum IFO 13613T 3 1 3 15 18 1 58Marinospirillum minutulum IFO 15450T 2 4 35 26 32aAfter Sakane and Yokota (1994).bOceanospirillum maris subsp. maris has not been investigated.cThe percentage of the acid relative to the total non-polar acids.
TABLE BXII.c.80. Cellular concentrations of 2- and 3-hydroxy fatty acids in Oceanospirillum species and Marinospirillum minutuluma
Speciesb Strain3-hydroxy fatty acidsc 2-hydroxy
fattyaciddC10:0 C12:0 C14:0 C14:1 C16:0 C18:0
O. linum IFO 15448T 100 %IFO 15449 100 %
O. beijerinckii subsp. beijerinckii IFO 15445T 63 30 6 %O. beijerinckii subsp. pelagicum IFO 13612T 60 30 9 %O. maris subsp. hiroshimense IFO 13616T 100 %O. maris subsp. williamsae IFO 15468T 100 %O. multiglobuliferum IFO 13614T 100 %O. jannaschii IFO 15447 100 %O. japonicum IFO 15466T 4 96 %
IFO 15446T 3 97 %O. kriegii IFO 15467T 19 54 27 %O. pusillum IFO 13613T 87 2 10 " (C18:1)Marinospirillum minutulum IFO 15450T 61 3 36 %aAfter Sakane and Yokota (1994).bOceanospirillum maris subsp. maris has not been investigated.cThe percentage of the acid relative to the total 3-hydroxy acids.d%, absent; ", present
cies confirmed the above findings. Kawasaki et al. (1997), in aphylogenetic study of helically shaped bacteria in the Alphapro-teobacteria, showed that O. pusillum was not related to other taxaof spirilla and constituted a separate branch in the Alphaproteo-bacteria, also confirming previous findings of Woese et al. (1985)and Pot (1996). Therefore, O. pusillum cannot belong to thegenus Oceanospirillum (Kawasaki et al., 1997). Phenotypic char-acteristics support the removal of O. pusillum from the genus: asingle flagellum at each pole, a counterclockwise type of helix,and a mol% G " C of 51, which is slightly higher than the rangeof 42–48 for the rest of the genus. As a formal transfer has notbeen proposed, O. pusillum is therefore listed below as “Speciesassigned but phylogenetically not belonging in Oceanospirillum” .
Satomi et al. (1998) determined and compared 16S rDNAsequences of all the Oceanospirillum species. They found that O.linum, O. maris, O. beijerinckii, and O. multiglobuliferum constituteda single rRNA cluster, separate from the branches formed byMarinobacter, Marinobacterium, and Marinomonas. Based on theirfindings, they also excluded O. japonicum from the genus Oce-anospirillum. Phenotypically, O. japonicum is different from otherOceanospirillum species since it does not form coccoid bodies andits flagella appear to be crescent shaped with less than one helical
turn, whereas those of other species have one or more helicalturns. Moreover, O. japonicum grows best at 35–37!C. As a formalnew description has not been proposed, O. japonicum is thereforelisted below as a “Species assigned but phylogenetically not be-longing in Oceanospirillum” .
In the same study, it was shown that O. minutulum clusteredon a separate branch together with new isolates from kusaya gravy(Satomi et al., 1998). For this branch, a new genus Marinospirillumhas been proposed, containing the two species M. minutulum andM. megaterium (Satomi et al., 1998).
Compared to the other Oceanospirillum species, O. jannaschiiand O. kriegii both have a higher mol% G " C (54.8–54.9) andoccupy a separate phylogenetic position (Satomi et al., 1998).Many phenotypic characteristics discriminate these species fromthe genus Oceanospirillum (Table BXII.c.82). Although not dis-cussed separately by the authors, O. jannaschii occurred on thesame branch as Marinobacterium (Gonzalez et al., 1997). Furthertaxonomic research, including DNA–DNA hybridizations, shouldbe performed to substantiate the exact level of genotypic rela-tionship between O. jannaschii and Marinobacterium georgiense. O.kriegii constituted a separate branch in the 16S rRNA dendrogram(Satomi et al., 1998).
GENUS I. OCEANOSPIRILLUM 277
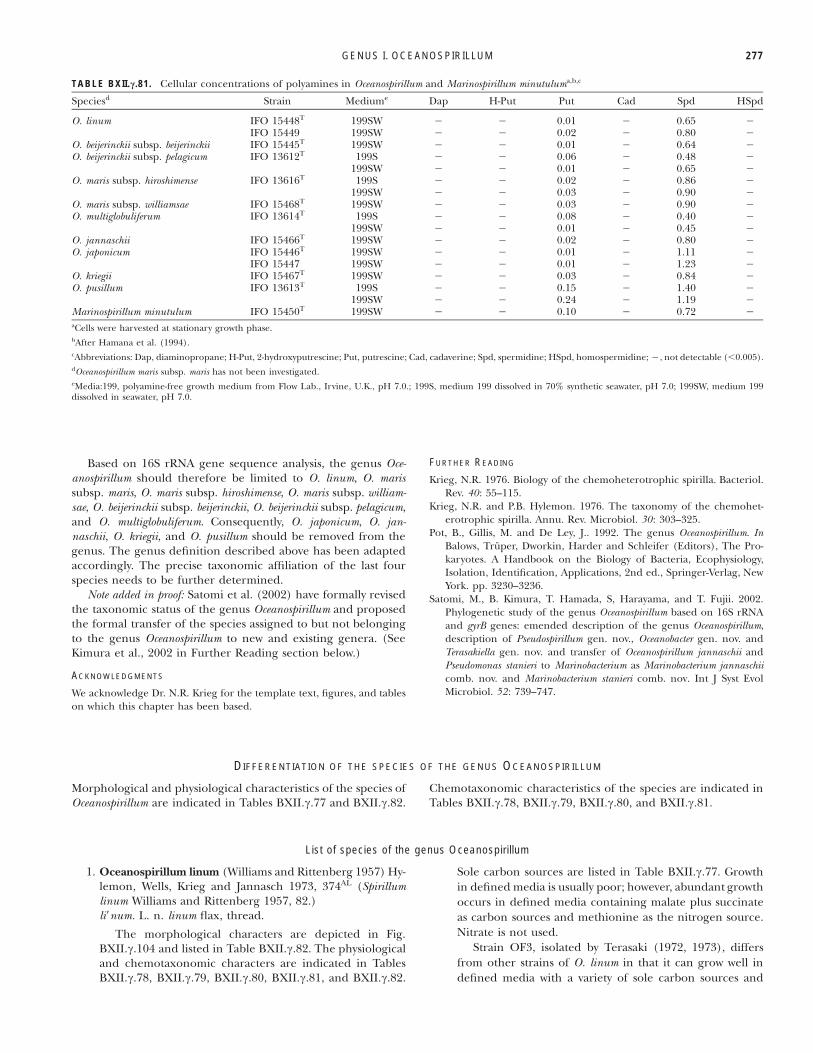
TABLE BXII.c.81. Cellular concentrations of polyamines in Oceanospirillum and Marinospirillum minutuluma,b,c
Speciesd Strain Mediume Dap H-Put Put Cad Spd HSpd
O. linum IFO 15448T 199SW % % 0.01 % 0.65 %IFO 15449 199SW % % 0.02 % 0.80 %
O. beijerinckii subsp. beijerinckii IFO 15445T 199SW % % 0.01 % 0.64 %O. beijerinckii subsp. pelagicum IFO 13612T 199S % % 0.06 % 0.48 %
199SW % % 0.01 % 0.65 %O. maris subsp. hiroshimense IFO 13616T 199S % % 0.02 % 0.86 %
199SW % % 0.03 % 0.90 %O. maris subsp. williamsae IFO 15468T 199SW % % 0.03 % 0.90 %O. multiglobuliferum IFO 13614T 199S % % 0.08 % 0.40 %
199SW % % 0.01 % 0.45 %O. jannaschii IFO 15466T 199SW % % 0.02 % 0.80 %O. japonicum IFO 15446T 199SW % % 0.01 % 1.11 %
IFO 15447 199SW % % 0.01 % 1.23 %O. kriegii IFO 15467T 199SW % % 0.03 % 0.84 %O. pusillum IFO 13613T 199S % % 0.15 % 1.40 %
199SW % % 0.24 % 1.19 %Marinospirillum minutulum IFO 15450T 199SW % % 0.10 % 0.72 %aCells were harvested at stationary growth phase.bAfter Hamana et al. (1994).cAbbreviations: Dap, diaminopropane; H-Put, 2-hydroxyputrescine; Put, putrescine; Cad, cadaverine; Spd, spermidine; HSpd, homospermidine; %, not detectable ("0.005).dOceanospirillum maris subsp. maris has not been investigated.eMedia:199, polyamine-free growth medium from Flow Lab., Irvine, U.K., pH 7.0.; 199S, medium 199 dissolved in 70% synthetic seawater, pH 7.0; 199SW, medium 199dissolved in seawater, pH 7.0.
Based on 16S rRNA gene sequence analysis, the genus Oce-anospirillum should therefore be limited to O. linum, O. marissubsp. maris, O. maris subsp. hiroshimense, O. maris subsp. william-sae, O. beijerinckii subsp. beijerinckii, O. beijerinckii subsp. pelagicum,and O. multiglobuliferum. Consequently, O. japonicum, O. jan-naschii, O. kriegii, and O. pusillum should be removed from thegenus. The genus definition described above has been adaptedaccordingly. The precise taxonomic affiliation of the last fourspecies needs to be further determined.
Note added in proof: Satomi et al. (2002) have formally revisedthe taxonomic status of the genus Oceanospirillum and proposedthe formal transfer of the species assigned to but not belongingto the genus Oceanospirillum to new and existing genera. (SeeKimura et al., 2002 in Further Reading section below.)
ACKNOWLEDGMENTS
We acknowledge Dr. N.R. Krieg for the template text, figures, and tableson which this chapter has been based.
FURTHER READING
Krieg, N.R. 1976. Biology of the chemoheterotrophic spirilla. Bacteriol.Rev. 40: 55–115.
Krieg, N.R. and P.B. Hylemon. 1976. The taxonomy of the chemohet-erotrophic spirilla. Annu. Rev. Microbiol. 30: 303–325.
Pot, B., Gillis, M. and De Ley, J.. 1992. The genus Oceanospirillum. InBalows, Truper, Dworkin, Harder and Schleifer (Editors), The Pro-karyotes. A Handbook on the Biology of Bacteria, Ecophysiology,Isolation, Identification, Applications, 2nd ed., Springer-Verlag, NewYork. pp. 3230–3236.
Satomi, M., B. Kimura, T. Hamada, S, Harayama, and T. Fujii. 2002.Phylogenetic study of the genus Oceanospirillum based on 16S rRNAand gyrB genes: emended description of the genus Oceanospirillum,description of Pseudospirillum gen. nov., Oceanobacter gen. nov. andTerasakiella gen. nov. and transfer of Oceanospirillum jannaschii andPseudomonas stanieri to Marinobacterium as Marinobacterium jannaschiicomb. nov. and Marinobacterium stanieri comb. nov. Int J Syst EvolMicrobiol. 52: 739–747.
DIFFERENTIATION OF THE SPECIES OF THE GENUS OCEANOSPIRILLUM
Morphological and physiological characteristics of the species ofOceanospirillum are indicated in Tables BXII.c.77 and BXII.c.82.
Chemotaxonomic characteristics of the species are indicated inTables BXII.c.78, BXII.c.79, BXII.c.80, and BXII.c.81.
List of species of the genus Oceanospirillum
1. Oceanospirillum linum (Williams and Rittenberg 1957) Hy-lemon, Wells, Krieg and Jannasch 1973, 374AL (Spirillumlinum Williams and Rittenberg 1957, 82.)li!num. L. n. linum flax, thread.
The morphological characters are depicted in Fig.BXII.c.104 and listed in Table BXII.c.82. The physiologicaland chemotaxonomic characters are indicated in TablesBXII.c.78, BXII.c.79, BXII.c.80, BXII.c.81, and BXII.c.82.
Sole carbon sources are listed in Table BXII.c.77. Growthin defined media is usually poor; however, abundant growthoccurs in defined media containing malate plus succinateas carbon sources and methionine as the nitrogen source.Nitrate is not used.
Strain OF3, isolated by Terasaki (1972, 1973), differsfrom other strains of O. linum in that it can grow well indefined media with a variety of sole carbon sources and

FAMILY I. OCEANOSPIRILLACEAE278
TAB
LEB
XII.c.
82.
Mor
phol
ogic
alan
dph
ysio
logi
calc
hara
cter
istic
sof
the
spec
ies
ofO
cean
ospi
rillu
man
dof
Mar
inos
piril
lum
min
utul
uma
Cha
ract
eris
tics
Typ
eof
helix
bC
CC
CSR
CSR
CC
CW
avel
engt
hof
helix
,lm
1.8–
4.0
6.3–
7.2
3.5–
7.0
3.5–
5.0
nd7.
0–20
.0nd
1.7–
2.0
2.0–
2.8
Hel
ixdi
amet
er,l
m0.
8–1.
41.
5–3.
01.
4–2.
81.
0–2.
0nd
2.0–
5.0
nd1.
0–1.
20.
6–1.
5Le
ngth
ofhe
lix,l
m4.
0–30
.02.
0–15
.52.
5–40
.02.
0–10
.0nd
5.0–
75.0
nd1.
2–4.
03.
0–8.
0C
elld
iam
eter
,lm
0.4–
0.6
0.6–
1.2
0.6–
1.1
0.5–
0.9
nd0.
8–1.
4nd
0.3–
0.5
0.3–
0.4
Flag
ella
rar
rang
emen
tcB
TB
TB
TB
TM
1-2
BT
MS
BS
BT
Pola
rm
embr
ane
pres
ent
nd"
d"
dnd
nd"
ndnd
"In
trac
ellu
lar
poly
-b-h
ydro
xybu
tyra
tefo
rmed
""
""
""
""
"e
Cocc
oid
bodi
espr
edom
inan
t:A
fter
3–4
wee
ks"
""
"nd
%nd
""
Aft
er24
–48
h%
%%
"nd
%nd
%%
Ran
geof
NaC
l(%
)fo
rgr
owth
inpe
pton
ew
ater
afte
r7
df0.
5–8.
00.
5–8.
0nd
0.5–
4.0
nd0.
5–8.
0nd
0.5–
8.0
0.5–
8.0
Gro
wth
inPS
S-se
awat
erbr
oth
cont
aini
ng:g
9.75
%to
talN
aCl
""
"nd
ndnd
ndnd
"12
.75%
tota
lNaC
l%
%%
ndnd
ndnd
ndd
Tem
pera
ture
rang
efo
rgr
owth
(!C
)h11
–39
8–41
2–35
6–37
10–4
36–
4011
–37
Opt
imum
grow
thte
mpe
ratu
re25
!Cra
ther
than
30–3
2!C
%%
di%
"%
"%
%Ph
osph
atas
e"
"di
"nd
wnd
w%
Nitr
ate
redu
ced
toni
trite
%%
%%
"%
%"
"O
xida
se"
""
""
""
""
Cat
alas
e"
orw
"or
wdi
"nd
wor
%nd
wor
%%
Ana
erob
icgr
owth
with
nitr
ate
%%
%%
nd%
nd%
%D
enitr
ifica
tion
%%
%%
%%
%%
%A
cid
prod
uced
from
suga
rs%
%%
%nd
%nd
%%
Hyd
roly
sis
ofes
culin
,hip
pura
te,s
tarc
h,or
case
in%
%%
%nd
%nd
%%
Indo
lete
st%
%%
%nd
%nd
%%
Sulfa
tase
(0.1
%ph
enol
phth
alei
ndi
sulfa
te)g
%%
%nd
ndnd
ndnd
%G
elatin
lique
fact
ion
at20
!Caf
ter:f,
j
7d
%d
%%
%d
%%
%28
d"
d%
%%
d%
%%
42d
"d
%%
%d
%%
%
(con
tinue
d)

GENUS I. OCEANOSPIRILLUM 279
TAB
LEB
XII.c.
82.
(con
t.)
Cha
ract
eris
tics
Gela
tinhy
drol
ysis
at30
!Caf
ter:
4dd
,g%
%%
nd%
%%
nd%
Red
uctio
nof
0.3%
H2S
eO3g
"%
%nd
ndnd
ndnd
"W
ater
-solu
ble
brow
npi
gmen
tfor
med
inth
epr
esen
ceof
:g
0.1%
tyro
sine
""
dnd
ndnd
ndnd
%0.
1%ph
enyl
alan
ine
%%
%nd
ndnd
ndnd
%0.
1%tr
ypto
phan
d%
dnd
ndnd
ndnd
dG
row
thin
the
pres
ence
of:g
0.1%
oxga
ll"
""
ndnd
ndnd
nd"
0.1%
glyc
ine
"%
"nd
ndnd
ndnd
"G
row
thon
:g
Eosi
nm
ethy
lene
blue
agar
%%
dnd
ndnd
ndnd
%M
acC
onke
yag
ar%
%%
ndnd
ndnd
nd%
Trip
le-S
ugar
iron
agar
"%
"nd
ndnd
ndnd
"Se
llers
agar
%%
%nd
ndnd
ndnd
%M
ethy
lred
-Vog
es–P
rosk
auer
brot
h"
%"
ndnd
ndnd
nd%
Wat
er-so
lubl
eye
llow
-gre
enflu
ores
cent
pigm
ent
form
edg
"%
dnd
%nd
%nd
%D
eoxy
ribo
nucl
ease
g%
"%
ndnd
ndnd
nd%
Rib
onuc
leas
egd
"%
ndnd
ndnd
nd%
Ure
aseg
%%
%nd
ndnd
ndnd
%A
uxot
roph
icgr
owth
requ
irem
ent
"k
%dk
%nd
%nd
%%
Mol
%(G
"C
)of
DN
A48
–50
47–4
945
–47
4656
–57
4554
–56
5142
–44
a Phen
otyp
icda
taar
efr
omK
rieg
(198
4a),
Bau
man
net
al.(
1972
),an
dB
owdi
tch
etal
.(19
84a)
;",p
rese
ntin
all
stra
ins;
%,l
acki
ngin
all
stra
ins;
d,de
pend
ing
onth
est
rain
;w,w
eak
reac
tion;
nd,n
otde
term
ined
.b C
,clo
ckw
ise
helix
;CC
,cou
nter
cloc
kwis
ehe
lix(d
eter
min
edby
focu
sing
onth
ebo
ttom
ofth
ece
lls.
c SR,s
trai
ght
rod;
BT,
bipo
lar
tuft
s,B
S,bi
pola
rsi
ngle
;M1-
2,1–
2fla
gella
aton
epo
le;M
S,m
onop
olar
sing
le.
dC
hara
cter
istic
has
not
been
test
edin
O.
mar
issu
bsp.
hiro
shim
ense
and
O.
beije
rinck
iisu
bsp.
pela
gicu
m.
e No
gran
ules
are
visi
ble
mic
rosc
opic
ally
inM
arin
ospi
rillu
mm
inut
ulum
but
chem
ical
anal
ysis
indi
cate
spr
esen
ceof
the
poly
mer
.f D
ata
from
Tera
saki
(197
2,19
79).
g Dat
afr
omH
ylem
onet
al.(
1973
).hO
cean
ospi
rillu
mkr
iegii
grow
sat
35!C
but
not
at40
!C;O
cean
ospi
rillu
mja
nnas
chii
does
not
grow
atei
ther
35!C
or40
!C.
i Oce
anos
piril
lum
mar
issu
bsp.
hiro
shim
ense
has
anop
timal
grow
thte
mpe
ratu
reof
25!C
;ca
tala
sere
actio
nca
nbe
nega
tive,
wea
k,or
posi
tive;
phos
phat
ase
reac
tion
can
bepo
sitiv
eor
nega
tive.
For
deta
iled
info
rmat
ion:
see
Hyl
emon
etal
.(19
73),
Tera
saki
(197
2,19
79),
and
Tabl
eB
XII
.c.8
3.j G
elat
inliq
uefa
ctio
nw
asno
tte
sted
inO
.m
aris
subs
p.m
aris.
Itw
asne
gativ
ein
O.
mar
issu
bsp.
hiro
shim
ense
,pos
itive
inO
.be
ijerin
ckii
subs
p.be
ijerin
ckii,
and
diff
ered
depe
ndin
gon
the
stra
inin
O.
beije
rinck
iisu
bsp.
pela
gicu
m.
k Oce
anos
piril
lum
linum
grow
spo
orly
orno
tat
all
ina
defin
edm
ediu
mw
itha
sing
leca
rbon
sour
cean
dam
mon
ium
ions
asth
eni
trog
enso
urce
;how
ever
,abu
ndan
tgr
owth
ocur
sin
ade
fined
med
ium
cont
aini
ngsu
ccin
ate
plus
mal
ate
asca
rbon
sour
cean
dm
ethi
onin
eas
the
nitr
ogen
sour
ce.
(See
also
foot
note
din
Tabl
eB
XII
.c.8
2.)
Oce
anos
piril
lum
mar
issu
bsp.
will
iam
sae
ATC
C29
54T
does
not
grow
invi
tam
in-fr
eem
ediu
man
dre
quir
esan
unid
entifi
edgr
owth
fact
or.

FAMILY I. OCEANOSPIRILLACEAE280
TABLE BXII.c.83. Differentiating characteristics for the subspecies ofOceanospirillum beijerinckiia
Characteristic
O. beijerinckiisubsp.
beijerinckii
O. beijerinckiisubsp.
pelagicum
Temperature range for growth (!C) 14–37 8–41Growth with:Lactate, citrate, malate, pyruvate,
acetate, propionate, tartrate% "
p-Hydroxybenzoate, ethanol,n-propanol
% d
Gelatin liquified at 20!C " dGrowth after 7 d in peptone water with:0.5–6.0% NaCl " %0.5–8.0% NaCl % "Mol% G " C of DNA 47 49aPhenotypic data from Terasaki (1972, 1979); ", present in all strains; %, lackingin all strains; d, depending on the strain
ammonium ions as the nitrogen source (see footnote d,Table BXII.c.77). Other characteristics of this strain aresimilar to those of the type strain (Terasaki, 1973), butwhether it should be included in this species is uncertain.
The species includes organisms previously assigned tothe two species Spirillum linum and Spirillum atlanticum byWilliams and Rittenberg (1957). The two species were com-bined into the single species O. linum by Hylemon et al.(1973), based on a high degree of similarity in phenotypiccharacters and in DNA base composition.
Isolated from coastal seawater.The mol% G " C of the DNA is: 48–50 (Tm).Type strain: ATCC 11336, DSM 6292, NCMB 56.
2. Oceanospirillum beijerinckii (Williams and Rittenberg1957) Hylemon, Wells, Krieg and Jannasch 1973, 374AL (Spi-rillum beijerinckii Williams and Rittenberg 1957, 90.)bei.jer.inck!i.i. M.L. gen. n. beijerinckii of Beijerinck; namedafter Prof M.W. Beijerinck of Delft, Holland.
The morphological characters are depicted in Fig.BXII.c.104 and listed in Table BXII.c.82. The physiologicaland chemotaxonomic characters are indicated in TablesBXII.c.78, BXII.c.79, BXII.c.80, BXII.c.81, and BXII.c.82.Sole carbon sources are listed in Table BXII.c.77. Ammo-nium ions can serve as a sole nitrogen source; nitrate is notused.
Isolated from coastal seawater.The mol% G " C of the DNA is: 47 (Tm).Type strain: ATCC 12754, DSM 7166, NCMB 52.
a. Oceanospirillum beijerinckii subsp. beijerinckii (Wil-liams and Rittenberg 1957) Hylemon, Wells, Krieg andJannasch 1973, 375AL (Spirillum beijerinckii Williams andRittenberg 1957, 90.)
Differs from O. beijerinckii subsp. pelagicum as indi-cated in Table BXII.c.83.
The mol% G " C of the DNA is: 47 (Tm).Type strain: ATCC 12754 , DSM 7166, NCMB 52.
b. Oceanospirillum beijerinckii subsp. pelagicum (Terasaki1973) Pot, Gillis, Hoste, Van de Velde, Bekaert, Kerstersand De Ley 1989, 32VP (Spirillum pelagicum Terasaki 1973,65.)pe.la!gi.cum. L. neut. adj. pelagicum belonging to the sea.
Differs from the O. beijerinckii subsp. beijerinckii as in-dicated in Table BXII.c.83.
Isolated from putrid infusions of marine mussels.The mol% G " C of the DNA is: 49 (Tm).Type strain: ATCC 33337, DSM 6288, IFO 13612,
NCMB 2228.GenBank accession number (16S rRNA): AB006761.
3. Oceanospirillum maris Hylemon, Wells, Krieg and Jannasch1973, 376AL
ma!ris. L. n. mare the sea; L. gen. n. maris of the sea.The morphological characters are depicted in Fig.
BXII.c.104 and listed in Table BXII.c.82. The physiologicaland chemotaxonomic characters are indicated in TablesBXII.c.78, BXII.c.79, BXII.c.80, BXII.c.81, and BXII.c.82.Sole carbon sources are listed in Table BXII.c.77.
Isolated from coastal seawater ( Jannasch, 1967).The mol% G " C of the DNA is: 45–46 (Tm).Type strain: ATCC 27509, DSM 6286, LMG 5213.GenBank accession number (16S rRNA): AB006771.
c. Oceanospirillum maris subsp. maris Hylemon, Wells,Krieg and Jannasch 1973, 376AL
Differs from the O. maris subsp. williamsae and the O.maris subsp. hiroshimense as indicated in Table BXII.c.84.
The mol% G " C of the DNA is: 46 (Tm).Type strain: ATCC 27509, DSM 6286, LMG 5213.GenBank accession number (16S rRNA): AB006771.
d. Oceanospirillum maris subsp. hiroshimense (Terasaki1973) Pot, Gillis, Hoste, Van de Velde, Bekaert, Kerstersand De Ley 1989, 33VP (Spirillum hiroshimense Terasaki1973, 62.)hi.ro.shi.men!se. M.L. neut. adj. hiroshimense pertaining toHiroshima. Japan.
Characters are as described for the species. Differsfrom the O. maris subsp. maris and the O. maris subsp.williamsae as indicated in Table BXII.c.84.
Isolated from putrid infusions of marine mussels.The mol% G " C of the DNA is: 47 (Tm).Type strain: IFO 13616, DSM 9524.
e. Oceanospirillum maris subsp. williamsae Linn and Krieg1984, 355VP
will!iam.sae. M.L. gen. n. williamsae of Williams; namedafter Marion A. Williams, who was the first to describespecies of marine spirilla.
Characters are as described for the species. Differsfrom the O. maris subsp. maris and the O. maris subsp.hiroshimense as indicated in Table BXII.c.84.
Isolated from a mixture of organisms comprisingNCMB strain 54 by Linn and Krieg (1978).
The mol% G " C of the DNA is: 45 (Tm).Type strain: ATCC 29547, IFO 15468.GenBank accession number (16S rRNA): AB006763.
4. Oceanospirillum multiglobuliferum (Terasaki 1973) Tera-saki 1979, 143AL (Spirillum multiglobuliferum Terasaki 1973,69.)mul.ti.glo.bu.li!fe.rum. L. adj. multus much, many; L. dim. n.globulus a small sphere, globule; L. v. fero to bear, carry; M.L.neut. adj. multiglobuliferum bearing many globules.
The morphological characters are listed in TableBXII.c.82. The physiological and chemotaxonomic char-acters are indicated in Tables BXII.c.78, BXII.c.79,BXII.c.80, BXII.c.81, and BXII.c.82. Sole carbon sources

GENUS I. OCEANOSPIRILLUM 281
TABLE BXII.c.84. Differentiating characteristics for the subspecies of Oceanospirillum marisa
Characteristic O. maris subsp. maris O. maris subsp. hiroshimense O. maris subsp. williamsae
Optimal growth temperature (!C) 30–32 25 30–32Catalase reaction strongly " d weakly "Phosphatase activity % " %DNase activity % "RNase activity % "Growth with 1% glycine " %Growth in vitamin-free, defined growth medium " %Growth with:
l-Glutamate, oxaloacetate " %Succinate, pyruvate, lactate, tartrate, acetate, propionate % " %
Mol% G " C of DNA 46 47 45aPhenotypic data from Hylemon et al. (1973); ", present in all strains; %, lacking in all strains; d, depending on the strain
are listed in Table BXII.c.77. Differs from other species byforming unusually large numbers of coccoid bodies evenin 24- to 48-h-old broth cultures. Ammonium ions can serveas a sole nitrogen source; nitrate is not used.
Isolated from putrid infusions of marine mussels.The mol% G " C of the DNA is: 46 (Tm).Type strain: IFO 13614.GenBank accession number (16S rRNA): AB006764.
Species assigned but phylogenetically not belonging inOceanospirillum
1. Oceanospirillum jannaschii Bowditch, Baumann and Bau-man 1984b, 503VP (Effective publication: Bowditch, Bau-mann and Bauman 1984a, 227.)jan.nasch!i.i. M.L. gen. n. jannaschii of Jannasch; namedafter H.W. Jannasch.
Straight rods. Some morphological, physiological, andchemotaxonomic characters are listed in Tables BXII.c.78,BXII.c.79, BXII.c.80, BXII.c.81, and BXII.c.82 (see alsoBaumann et al., 1972). Sole carbon sources are listed inTable BXII.c.77. Utilize 39–46 organic compounds includ-ing fatty acids, tricarboxylic acid cycle intermediates, alco-hols, amino acids, and amines. Utilizes c-aminovalerate andhistamine (Baumann et al., 1972).
Isolated from seawater after enrichment (Baumann etal., 1972).
The mol% G " C of the DNA is: 56–57 (Tm).Type strain: ATCC 27135, DSM 6295, IFO 15466.GenBank accession number (16S rRNA): AB006765.
2. Oceanospirillum japonicum (Watanabe 1959) Hylemon,Wells, Krieg and Jannasch 1973, 375AL (Spirillum japonicumWatanabe 1959, 78.)ja.pon!i.cum. M.L. neut. adj. japonicum pertaining to Japan.
The morphological characters are depicted in Fig.BXII.c.104 and listed in Table BXII.c.82; however, the typestrain presently has morphological features that differ fromthe original description, in that the cells are no longerhelical with several waves but instead are curved, straight,or S-shaped; moreover, colonies of this strain are presentlyof the R (rough) type. Therefore, it is likely that the typestrain has undergone alteration since its isolation in 1959.Three reference strains isolated by Terasaki (1972, 1973)have morphological features that more nearly correspond
to those given in the original description (strains IF4, IF8,and UF3), and also they form colonies of the S (smooth)type.
The physiological and chemotaxonomic characters areindicated in Tables BXII.c.78, BXII.c.79, BXII.c.80,BXII.c.81, and BXII.c.82. Sole carbon sources are listed inTable BXII.c.77.
Isolated from putrid infusions of marine mussels.The mol% G " C of the DNA is: 45 (Tm).Type strain: ATCC 19191, DSM 7165.
3. Oceanospirillum kriegii Bowditch, Baumann and Bauman1984b, 503VP (Effective publication: Bowditch, Baumannand Bauman 1984a, 227.)krie!gi.i. M.L. gen. n. kriegii of Krieg; named after N.R. Krieg.
Straight rods. Some morphological and physiologicalcharacters are listed in Tables BXII.c.78, BXII.c.79,BXII.c.80, BXII.c.81, and BXII.c.82 (see also Baumann etal., 1972). Sole carbon sources are listed in Table BXII.c.77.Utilize 29–33 organic compounds including d-glucose andd-fructose but no other pentose, hexose, or disaccharide;also utilize tricarboxylic acid cycle intermediates, alcohols,amino acids, and amines. Produces an extracellular lipase(Baumann et al., 1972).
Isolated from see water after enrichment (Baumann etal., 1972).
The mol% G " C of the DNA is: 54–56 (Tm).Type strain: ATCC 27133, DSM 6294, IFO 15467, NCMB
2042.GenBank accession number (16S rRNA): AB006767.
4. Oceanospirillum pusillum (Terasaki 1973) Terasaki 1979,142AL (Spirillum pusillum Terasaki 1973, 67.)pu.sil!lum. L. dim. neut. adj. pusillum very small.
The morphological characters are listed in TableBXII.c.82. The physiological and chemotaxonomic char-acters are indicated in Tables BXII.c.78, BXII.c.79,BXII.c.80, BXII.c.81, and BXII.c.82. Sole carbon sourcesare listed in Table BXII.c.77. Ammonium ions can serve asa sole nitrogen source; nitrate is not used.
Isolated from putrid infusions of marine mussels.Belongs to the Alphaproteobacteria.The mol% G " C of the DNA is: 51 (Tm).Type strain: ATCC 33338, DSM 6293, IAM 14442, IFO
13613, NCMB 2229.GenBank accession number (16S rRNA): AB006768.

FAMILY I. ENTEROBACTERIACEAE 607
kazakii in most biochemical reactions, including production of ayellow pigment. However, they were only 43% related to E. sa-kazakii by DNA–DNA hybridization. Recent DNA relatedness data
by Kosako et al. (1996) indicated that Enterobacter kobei is theclosest relative of Enteric Group 69.
Genus I. Escherichia Castellani and Chalmers 1919, 941TAL
FLEMMING SCHEUTZ AND NANCY A. STROCKBINE
Esch.er.i!chi.a. M.L. fem. n. Escherichia named after Theodor Escherich, who isolated the type speciesof the genus.
Straight cylindrical rods, 1.1–1.5 " 2.0–6.0 lm, occurring singlyor in pairs. Conform to the general definition of the family En-terobacteriaceae. Gram negative. Motile by peritrichous flagella ornonmotile. Aerobic and facultatively anaerobic having both arespiratory and a fermentative type of metabolism, but anaero-genic biotypes occur. Oxidase negative. Chemoorganotrophic.Both acid and gas are formed from most fermentable carbo-hydrates, but i-inositol is not utilized and d-adonitol is utilizedonly by Escherichia fergusonii. Lactose is fermented by most strainsof Escherichia coli, but fermentation may be delayed or absentin Escherichia blattae, Escherichia hermannii, Escherichia fergusonii,and Escherichia vulneris. Do not grow in KCN (with the exceptionof E. hermannii and a small proportion of E. vulneris). Usuallydo not produce H2S. E. coli occur naturally in the lower part ofthe intestine of warm-blooded animals, E. blattae in the hind-gutof cockroaches, and E. fergusonii, E. hermannii, and E. vulneris arefound in the intestine, as well as extraintestinal sites of warm-blooded animals. Seven copies of the rrn operon with genescoding for 16S, 23S, and 5S rRNA are present on the chromo-some of E. coli. Comparative sequence analysis between the genesfor 16S rRNA of E. coli, E. vulneris, and E. hermannii and ho-mologous genes from all eubacteria places E. coli and E. vulneristogether in a tightly related cluster with shigellae, and E. her-mannii between Salmonella spp. and Citrobacter freundii (Cilia etal., 1996). Based on 16S rRNA sequencing, escherichiae belongin the Gammaproteobacteria.
The mol% G # C of the DNA is: 48–59.Type species: Escherichia coli (Migula 1895) Castellani and
Chalmers 1919, 941 (Bacillus coli Migula 1895, 27.)
FURTHER DESCRIPTIVE INFORMATION
Cell morphology Escherichiae are straight, cylindrical,Gram-negative rods with rounded ends that are 1.1–1.5 lm indiameter and 2.0–6.0 lm in length. They occur singly or in pairsand can be motile by peritrichous flagella or nonmotile. Fig.BXII.c.191 shows negatively stained preparations of each of theEscherichia species. See Nanninga (1985) for a comprehensivetreatment of the ultrastructure of E. coli.
Phylogenetic and systematic treatment The genus consists offive species: E. coli, E. hermannii, E. fergusonii, E. vulneris, and E.blattae. Biochemical reactions that will help in differentiating be-tween the species of Escherichia are listed in Table BXII.c.197.
Biochemical reactions Escherichiae produce strong acids andusually gas from the fermentation of d-glucose (positive in theMethyl Red test) and do not produce acetyl-methyl carbinol (ace-toin) (negative in the Voges–Proskauer test). Sodium acetate isfrequently used as a sole carbon source, except by E. blattae anda majority of E. vulneris strains. Citrate (Simmons’ citrate agar)
cannot be used by E. coli and E. vulneris, whereas a smaller pro-portion of E. fergusonii and E. hermannii exhibit immediate ordelayed use of this substrate. Growth on Simmons’ citrate agarby E. blattae is variable and probably strain specific.
Lysine is decarboxylated by the majority of strains. Exceptionsinclude “metabolically inactive” E. coli strains, the majority ofenteroinvasive E. coli strains (EIEC), and E. hermannii. Ornithineis decarboxylated by all species except by E. vulneris and a littleless than half of E. coli strains. Indole is produced by all speciesexcept E. blattae and E. vulneris.
Other tests that help in differentiating between the speciesof Escherichia include growth in potassium cyanide, malonate util-ization, and acid production from d-adonitol, d-arabitol, cello-biose, dulcitol, lactose, d-mannitol, melibiose, d-sorbitol, and mu-cate. See Table BXII.c.197.
Molecular data Findings from the comparison of 16S rDNAsequences performed with strains of E. coli, Salmonella spp., andC. freundii have shown a close phylogenetic relatedness betweenthese bacteria (Ahmad et al., 1990; Cilia et al., 1996; Chang etal., 1997). Analysis of 16S rDNA sequences separates the twoSalmonella species (S. enterica and S. bongori) from the complexof E. coli and Shigella, and shows that E. hermannii is more closelyrelated to Salmonella enterica and C. freundii (Christensen et al.,1998). This analysis is unable to separate inter-operon variationof E. coli from strains of E. coli and from Shigella. Patterns ofsequence heterogeneity in strains from the E. coli Collection ofReference (ECOR) (Ochman and Selander, 1984) have beenlocated at regions V1 and V6 of cloned 16S rRNA genes (Mar-tinez-Murcia et al., 1999).
Average DNA relatedness assessed by DNA–DNA hybridizationamong Escherichia species ranges from 29% to 94% (TableBXII.c.198). DNAs from different strains of E. coli are closelyrelated (average, 84%; Table BXII.c.198). With the exception ofS. boydii serotype 13, the DNAs of E. coli and the four Shigellaspecies show such a high degree of relatedness (average 80–87%,except S. boydii type 13, which is about 65%) that these speciesshould be considered as a single species (Brenner et al., 1973a).The distinction between these bacteria prevails, however, for rea-sons of historical/medical precedent and to avoid confusion inthe literature and with existing surveillance systems.
Based on complete sequencing of the K-12 strain MG1655(Blattner et al., 1997), it is estimated that the E. coli lineagediverged from the Salmonella lineage some 100 million years ago(Lawrence and Ochman, 1998). This is a little less than the 120–160 million years estimated by calibration of the rate of 16S rRNAevolution in bacteria (Ochman and Wilson, 1987). The chro-mosome of E. coli strain MG1655 is 4,639,221 bp and contains4,288 open reading frames (ORFs). Approximately 18% of theseORFs represent genes that have been acquired and have persisted
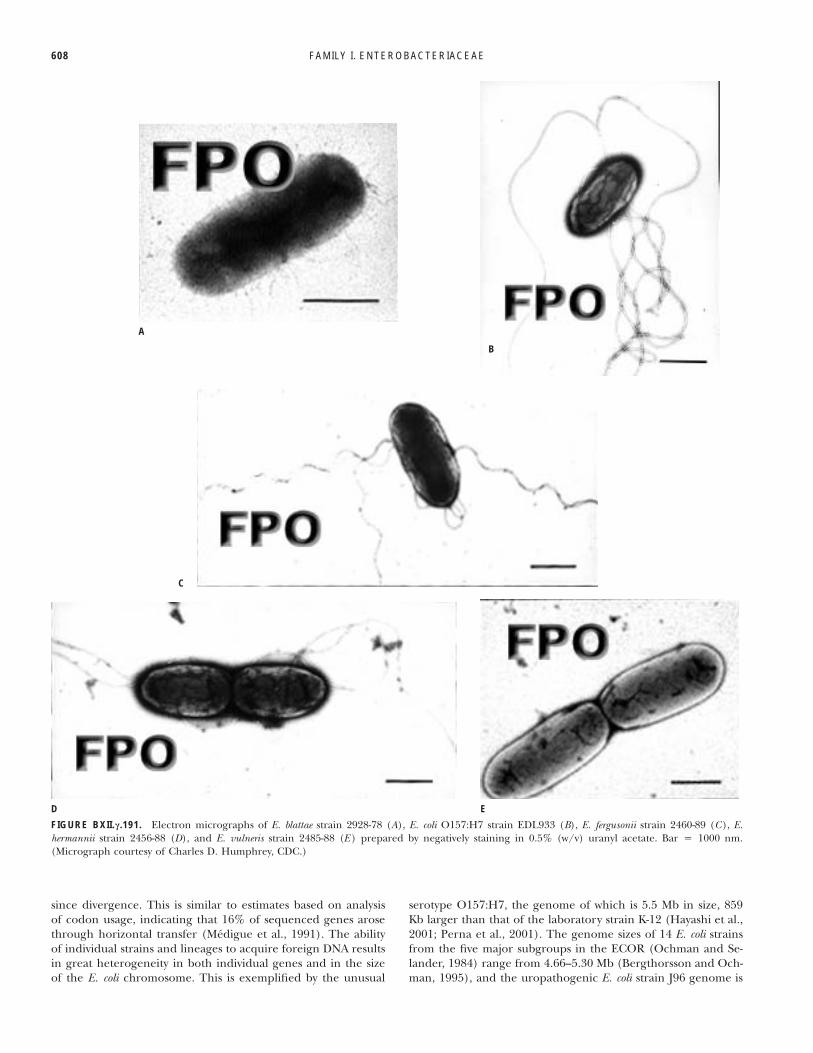
FAMILY I. ENTEROBACTERIACEAE608
A
B
C
D E
FIGURE BXII.c.191. Electron micrographs of E. blattae strain 2928-78 (A), E. coli O157:H7 strain EDL933 (B), E. fergusonii strain 2460-89 (C), E.hermannii strain 2456-88 (D), and E. vulneris strain 2485-88 (E) prepared by negatively staining in 0.5% (w/v) uranyl acetate. Bar $ 1000 nm.(Micrograph courtesy of Charles D. Humphrey, CDC.)
since divergence. This is similar to estimates based on analysisof codon usage, indicating that 16% of sequenced genes arosethrough horizontal transfer (Medigue et al., 1991). The abilityof individual strains and lineages to acquire foreign DNA resultsin great heterogeneity in both individual genes and in the sizeof the E. coli chromosome. This is exemplified by the unusual
serotype O157:H7, the genome of which is 5.5 Mb in size, 859Kb larger than that of the laboratory strain K-12 (Hayashi et al.,2001; Perna et al., 2001). The genome sizes of 14 E. coli strainsfrom the five major subgroups in the ECOR (Ochman and Se-lander, 1984) range from 4.66–5.30 Mb (Bergthorsson and Och-man, 1995), and the uropathogenic E. coli strain J96 genome is
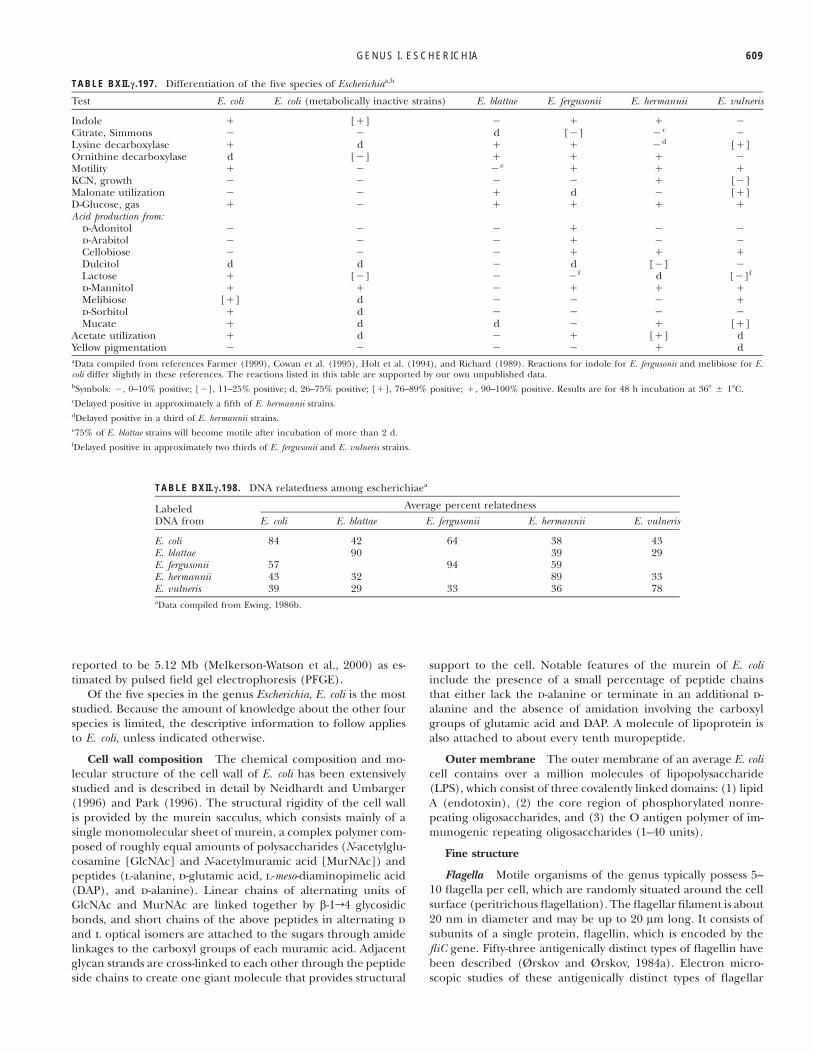
GENUS I. ESCHERICHIA 609
TABLE BXII.c.197. Differentiation of the five species of Escherichiaa,b
Test E. coli E. coli (metabolically inactive strains) E. blattae E. fergusonii E. hermannii E. vulneris
Indole # [#] % # # %Citrate, Simmons % % d [%] %c %Lysine decarboxylase # d # # %d [#]Ornithine decarboxylase d [%] # # # %Motility # % %e # # #KCN, growth % % % % # [%]Malonate utilization % % # d % [#]D-Glucose, gas # % # # # #Acid production from:
d-Adonitol % % % # % %d-Arabitol % % % # % %Cellobiose % % % # # #Dulcitol d d % d [%] %Lactose # [%] % %f d [%]f
d-Mannitol # # % # # #Melibiose [#] d % % % #d-Sorbitol # d % % % %Mucate # d d % # [#]
Acetate utilization # d % # [#] dYellow pigmentation % % % % # daData compiled from references Farmer (1999), Cowan et al. (1995), Holt et al. (1994), and Richard (1989). Reactions for indole for E. fergusonii and melibiose for E.coli differ slightly in these references. The reactions listed in this table are supported by our own unpublished data.bSymbols: %, 0–10% positive; [%], 11–25% positive; d, 26–75% positive; [#], 76–89% positive; #, 90–100% positive. Results are for 48 h incubation at 36! & 1!C.cDelayed positive in approximately a fifth of E. hermannii strains.dDelayed positive in a third of E. hermannii strains.e75% of E. blattae strains will become motile after incubation of more than 2 d.fDelayed positive in approximately two thirds of E. fergusonii and E. vulneris strains.
TABLE BXII.c.198. DNA relatedness among escherichiaea
LabeledDNA from
Average percent relatednessE. coli E. blattae E. fergusonii E. hermannii E. vulneris
E. coli 84 42 64 38 43E. blattae 90 39 29E. fergusonii 57 94 59E. hermannii 43 32 89 33E. vulneris 39 29 33 36 78aData compiled from Ewing, 1986b.
reported to be 5.12 Mb (Melkerson-Watson et al., 2000) as es-timated by pulsed field gel electrophoresis (PFGE).
Of the five species in the genus Escherichia, E. coli is the moststudied. Because the amount of knowledge about the other fourspecies is limited, the descriptive information to follow appliesto E. coli, unless indicated otherwise.
Cell wall composition The chemical composition and mo-lecular structure of the cell wall of E. coli has been extensivelystudied and is described in detail by Neidhardt and Umbarger(1996) and Park (1996). The structural rigidity of the cell wallis provided by the murein sacculus, which consists mainly of asingle monomolecular sheet of murein, a complex polymer com-posed of roughly equal amounts of polysaccharides (N-acetylglu-cosamine [GlcNAc] and N-acetylmuramic acid [MurNAc]) andpeptides (l-alanine, d-glutamic acid, l-meso-diaminopimelic acid(DAP), and d-alanine). Linear chains of alternating units ofGlcNAc and MurNAc are linked together by b-1r4 glycosidicbonds, and short chains of the above peptides in alternating dand l optical isomers are attached to the sugars through amidelinkages to the carboxyl groups of each muramic acid. Adjacentglycan strands are cross-linked to each other through the peptideside chains to create one giant molecule that provides structural
support to the cell. Notable features of the murein of E. coliinclude the presence of a small percentage of peptide chainsthat either lack the d-alanine or terminate in an additional d-alanine and the absence of amidation involving the carboxylgroups of glutamic acid and DAP. A molecule of lipoprotein isalso attached to about every tenth muropeptide.
Outer membrane The outer membrane of an average E. colicell contains over a million molecules of lipopolysaccharide(LPS), which consist of three covalently linked domains: (1) lipidA (endotoxin), (2) the core region of phosphorylated nonre-peating oligosaccharides, and (3) the O antigen polymer of im-munogenic repeating oligosaccharides (1–40 units).
Fine structure
Flagella Motile organisms of the genus typically possess 5–10 flagella per cell, which are randomly situated around the cellsurface (peritrichous flagellation). The flagellar filament is about20 nm in diameter and may be up to 20 lm long. It consists ofsubunits of a single protein, flagellin, which is encoded by thefliC gene. Fifty-three antigenically distinct types of flagellin havebeen described (Ørskov and Ørskov, 1984a). Electron micro-scopic studies of these antigenically distinct types of flagellar

FAMILY I. ENTEROBACTERIACEAE610
filaments revealed differences in surface structure; six uniqueflagellar morphotypes have been described (Lawn et al., 1977).Unlike Salmonella, most E. coli strains have only one flagellin geneand do not undergo phase variation. Exceptions have been de-scribed (Ratiner, 1967, 1982, 1999). See Macnab (1996) for acomplete description of the structure and genes involved in mo-tility.
Fimbriae In addition to the proteinaceous flagella, moststrains have fimbriae (pili) or fibrillar proteins often extendingin great numbers from the bacterial surface and far out into thesurrounding medium. A typical E. coli K-12 cell contains 100–500 type 1 fimbriae arranged peritrichously, each with a diameterof approximately 7 nm and a length of 0.2–2.0 lm. More than30 different fimbriae have been described in E. coli, which com-monly expresses more than one type at a time. The character-istics, biogenesis, and classification schemes of fimbriae are re-viewed by Low et al. (1996). Fimbriae have historically been clas-sified by phenotypic properties. One widely used scheme clas-sifies fimbriae according to their adhesive properties for redblood cells from different host species in the presence of man-nosides. By this method, two main types of fimbriae are recog-nized: mannose-sensitive (MS) fimbriae, which are unable to ag-glutinate red blood cells in the presence of !-d-mannose, andmannose-resistant (MR) fimbriae, which are able to agglutinatered blood cells in the presence of this sugar. MS fimbriae, whichinclude the so-called type 1 fimbriae (pili), are found in themajority of E. coli strains and comprise a group of more or lessserologically related antigens. Because they are expressed bypathogens as well as commensal organisms, their role in virulencehas been difficult to establish. Evidence for the role of type 1fimbriae in virulence is reviewed by Abraham and Jaiswal (1997).Type 1 fimbriae mediate avid bacterial attachment to mucosalsurfaces, to noncellular host constituents, and to various inflam-matory cells. They bind to certain oligomannoside-containingglycoproteins present on mucosal surfaces, including the Tamm–Horsfall glycoprotein, which is synthesized in the kidney andpresent in urinary slime (Ørskov et al., 1980a); fibronectin, aglycoprotein that is a member of a family of proteins found inthe extracellular matrix (ECM), plasma, and other body fluids;and laminin, a glycoprotein present in basement membranes(Kukkonen et al., 1993). The genes involved in the synthesis andregulation of type 1 fimbriae are located on the chromosome.Expression of type 1 fimbriae is subject to being turned on oroff (phase variation) as a result of the inversion of a 314 base-pair fragment of DNA containing the promoter region of thegene encoding the major fimbrial subunit (fimA). Expression oftype 1 fimbriae is influenced by environmental and growth con-ditions and is controlled by global regulatory factors such asleucine-responsive regulatory protein (Lrp), integration host fac-tor (IHF) and histone-like protein H-NS. See Abraham and Jais-wal (1997) and Low et al. (1996) for a review of the environ-mental factors and genes involved in synthesis and regulation oftype 1 fimbriae.
MR fimbriae are serologically diverse (Ørskov et al., 1980b,1982; Ørskov and Ørskov, 1990) and often function as virulencefactors to mediate adherence that is species- and organ-specific.The genes for these proteins may be located on plasmids or onthe chromosome. When located chromosomally, they often clus-ter together with other virulence genes in regions of the chro-mosome referred to as pathogenicity islands (PAIs). More thanany other virulence factor, the MR pili of E. coli illustrate the
species’ capacity to adapt to the receptor-specific epithelial cellsof certain hosts primarily through horizontal acquisition of genecassettes on plasmids, phages, or other mobile DNA elementsthat will allow for colonization. Evidence for the horizontal trans-fer of fimbrial genes in E. coli comes from the remarkable sim-ilarity between the genetic organization of its fimbrial operonsand those of other members of the family Enterobacteriaceae andfrom the strikingly low C # G content and different codon usagepattern among the fimbrial genes compared to those observedoverall among other genes on the E. coli chromosome.
Fimbriae may also be classified based on their morphology.One group consists of thick, rod-shaped fimbriae with a diameterof 7 nm (range 3.4–8 nm), a length of 0.5–2 lm, and an axialhole diameter of 2.0–2.5 nm. Fimbriae with these dimensionsare represented by the rigid pyelonephritis (P), sialic acid (S),type 1, F6 (987P), colonization factor antigen I (CFA/I), colisurface antigen 1 (CS1) and CS2 pili, and by the bundle-formingCS8 (CFA/III) and CS21 (longus), the latter with homology tothe type 4 fimbrial family. Enteropathogenic E. coli (EPEC) isknown to produce a type 4 fimbria called the bundle-formingpilus (BFP). Another flexible, bundle-forming fimbrial structureof 2–3 nm diameter, designated aggregative adherence fimbriaeI (AAF/I), shows no homology to the type 4 class of fimbriae.Together with another recently described AAF/II fimbria, it ex-hibits sequence and organizational resemblance to the Dr family.Another group consisting of thinner, more flexible fibrillae witha width of 2–5 nm and a length of 0.5–2.0 lm is represented byF4 (K88), F5 (K99), F41, and CS3. Helical fibrillae, where twofibrillae are arranged in a helix, are represented by CS5 and CS7.
Nonfimbrial and related adhesins Bacterial adherence may alsobe mediated by adhesions that are afimbrial (AFA) or nonfim-brial (NFA). Some of these proteins form larger multimers thataggregate around the bacterial cell as an amorphous structurereminiscent of capsular K antigens. Afimbrial adhesins are foundin both uropathogenic E. coli (UPEC) and in diffusely adheringE. coli (DAEC) and represented by the Afa/Dr family consistingof Dr (previously referred to as O75X) and Dr-II (drb), the F1845pilus (daa), and AFAI-IV (afa). Dr fimbriae and related adhesinsrecognize different epitopes of the Dr blood group antigen (Now-icki et al., 1990) and bind to a complement-regulatory protein,the common receptor decay accelerating factor (DAF). E. colistrains expressing Dr fimbriae are able to enter epithelial cellsby interacting with DAF (Goluszko et al., 1997). Other adhesinssuch as the M agglutinin and AIDA-I adhesin, a plasmid-encodedouter membrane protein involved in diffuse adherence of certaintypes of E. coli (Benz and Schmidt, 1989), are commonly present.
Colonial and cultural characteristics; life cycles Dependingon the degree of polymerization of the O antigen polysaccharide,the phenotypes of strains growing on agar media are describedas smooth (S) or rough (R). S forms, which usually grow onnutrient agar as convex, glistening, moist, gray colonies (2–3 mmdiameter) with a defined edge or in fluid medium as turbidgrowth, have developed polysaccharide side chains, while Rforms, which usually grow as flat, dry, dull, wrinkled colonies (1–5 mm diameter) with a blurred edge on agar and agglutinatespontaneously in fluid media, have lost their polysaccharide sidechains by mutation (Luderitz et al., 1966). There are interme-diate forms between these extremes. Mucoid and slime-produc-ing forms occur. E. hermannii is yellow-pigmented, as are half ofthe described E. vulneris strains. See Raetz (1996) and Hull

GENUS I. ESCHERICHIA 611
(1997) for a discussion of the chemical structure, biosynthesis,and biological/virulence properties of LPS.
Nutrition and growth conditions Of the range of tempera-tures, pH values, water activities, and pressures over which bac-terial growth can occur, E. coli strains survive and grow over themid-range (15–45!C) of these environmental conditions. Moststrains can grow over a temperature range of approximately 40!C.The normal temperature range for balanced growth extendsfrom 21! to 37!C; however, strains that can grow at temperaturesas low as 7.5–7.8!C (Shaw et al., 1971) and as high as 49!C (Her-endeen et al., 1979) have been described. A minimum growthtemperature for E. vulneris of 1.6!C (0.8–2.6!C) was reported fora strain isolated from refrigerated meat (Ridell and Korkeala,1997). E. coli is neutrophilic and will grow over the mid-range ofpH, from about pH 5.0 to 9.0 (Ingraham and Marr, 1996).
Metabolism and metabolic pathways Glucose and other car-bohydrates are fermented with the production of pyruvate, whichis further converted into lactic, acetic, and formic acids. Part ofthe formic acid is split by a complex hydrogenlyase system intoequal amounts of CO2 and H2.
Phylogeny The establishment of the Escherichia coli Collec-tion of Reference (ECOR) in 1984 (Ochman and Selander, 1984)and subsequent studies of the strains in ECOR and comparisonswith other E. coli strains have contributed substantially to ourunderstanding of the evolution and population structure of E.coli. ECOR is a collection of 72 strains from humans and 16 othermammalian species from various geographical areas that havebeen grouped into five main groups—A, B (comprising sub-groups B1 and B2), C, D, and E—according to their electro-phoretic types and enzyme allele (allozyme) profiles, based onthe results of multilocus enzyme electrophoresis (MLEE) (Se-lander et al., 1986). The original data for 35 enzymes (Selanderet al., 1987) have been expanded to include allozymes of fouresterase loci (Goullet and Picard, 1989), and, based on allelicvariation at 38 enzyme-encoding loci, multilocus genotypes havebeen used to construct a dendrogram based on the neighborjoining algorithm (Herzer et al., 1990), demonstrating the clonalstructure of the species. The phylogenetic groups are distin-guishable but not identical by random amplified polymorphicDNA (RAPD), restriction fragment length polymorphism (RFLP)of rrn genes (ribotyping) (Desjardins et al., 1995), and to a lesserextent by repetitive-element PCR (rep-PCR) fingerprinting usingERIC2 and BOXA1R primers ( Johnson and O’Bryan, 2000).Generating a phylogenetic tree of the ECOR and 15 O157 strainsby fluorescent amplified-fragment length polymorphism(FAFLP) demonstrated close correlation with the MLEE groupsof ECOR and placed the STEC/VTEC O157 strains on an outlierbranch (Arnold et al., 1999). Thus, there is sufficient evidencethat the ECOR strains broadly represent genotypic variation inclonal groups of E. coli in spite of the fact that many isolates arecommensal forms from healthy carriers and that only 11 out ofthe 72 strains are from human disease: 1 strain from a case ofasymptomatic bacteriuria, 4 from acute cystitis, and 6 from acutepyelonephritis. Other MLEE studies have showed clonal rela-tionships of diarrheagenic E. coli, and a collection of 78 diar-rheagenic E. coli (DEC) representing 15 clonal groups has beenestablished (Whittam et al., 1993). A similar collection of STEC/VTEC strains has been collected by the STEC Center based atthe National Food Safety and Toxicology Center at MichiganState University and is designed to facilitate research on theShiga/Verocytotoxin producing E. coli by providing a standard
reference collection of well-characterized strains and a central,on-line accessible database.*
Numerous studies have indicated that E. coli and the fournamed species of Shigella should be regarded as being one species(Brenner et al., 1972a, b, 1973a; Goullet, 1980; Ochman et al.,1983; Whittam et al., 1983; Hartl and Dykhuizen, 1984; Karaoliset al., 1994; Stevenson et al., 1994; Whittam, 1996; Pupo et al.,1997). Findings from MLEE studies combined with those frommdh (malate dehydrogenase) housekeeping gene sequence stud-ies (Pupo et al., 1997) and ribotyping (Rolland et al., 1998)confirm that the genus Shigella comprises a group of closely re-lated pathogenic E. coli strains and indicate that Shigella, EIEC,and other diarrheagenic E. coli strains do not have a single ev-olutionary origin, but are derived from different ancestral strainsmany times. Furthermore, pathogenic strains belonging to path-ogenic groups of EIEC, EPEC, and ETEC were found to be closelyrelated to ECOR group A strains by MLEE and have mdh se-quences identical to five ECOR strains from group A, which isthought to represent commensal strains. It has been suggestedthat any E. coli strain may acquire virulence factors from nu-merous sources, including plasmids, bacteriophages, and othermobile DNA elements from a large pool of strain-specific geneswhose origin could be outside the species boundaries, and thatthis is how a commensal form is turned into a pathogenic form(Pupo et al., 1997; Hurtado and Rodriguez-Valera, 1999; Don-nenberg and Whittam, 2001). In their analysis, Lawrence andOchman (1998) surmised that about 10% of the E. coli K-12genome consists of genes that were acquired in over 200 eventsof lateral gene transfer, which occurred subsequent to the di-vergence of E. coli and Salmonella some 100 million years ago.While mutations have contributed substantially to the hetero-geneity of E. coli, the importance of these recombinational eventsshould not be underestimated, and transfer of smaller or biggersegments of DNA between different clonal lineages and otherspecies has probably contributed more to the evolution of E. colithan anyone could have imagined. Ongoing and future studieswill be directed toward increasing our understanding of the prin-ciples that govern gene patterns and the relations between in-dividual traits in terms of their significance for virulence andhost adaptation. The combined efforts should contribute to ageneral IDEA (Index of Diversity in Evolution and Adaptation)that will bridge the phylogenetic approach based on the clonalconcept on one side and the horizontal transfer of gene cassetteson the other side.
Mutants, plasmids, phages, and bacteriocins The fact thatE. coli mutants can easily be produced in the laboratory has sub-stantially contributed to our understanding of many geneticmechanisms, ranging from the characterization and function ofan individual gene to the description of complex operons. Plas-mids carrying resistance genes (R plasmids) have been used asvectors and introduced into both laboratory strains and wild-typestrains, and phages and other mobile elements such as trans-posons have been used widely in both research and applied bio-technology. Virulence genes are often found on plasmids (seepathogenicity section below). E. coli strains produce a variety ofsecreted antibiotically active polypeptides, bacteriocins, and mi-crocins, which have the ability to kill or inhibit competing bac-terial strains. Colicins, encoded by plasmids of E. coli, act on other
*Editorial Note: At the time of publication, this information could be obtained athttp://www.shigatox.net.

FAMILY I. ENTEROBACTERIACEAE612
E. coli or closely related bacteria that do not carry that particularCol plasmid. Small lipoproteins are important components ofthe secretory apparatus, which facilitates release of colicin andplasmid- and phage-encoded proteins across the outer mem-brane. For additional information on E. coli mutants, plasmids,and phages see Campbell (1996), Bachmann (1996), Helinski etal. (1996), and Harwood (1993).
Antigenic structure
O antigens The main aspect of this analysis is the O antigendetermination based on antigenicity of the LPS; O group des-ignations run from O1 to O173 but O groups O31, O47, O67,O72, O93, O94, and O122 have been removed. Also includedare the provisional O groups OX3 and OX7 listed by Ewing(1986b), which will receive the designations O174 and O175,respectively. An additional six new O groups representing STEC/VTEC strains are currently being investigated and will receive Odesignations O176 through O181 (Scheutz, unpublished data).Subtypes exist within most O groups and are designated ab, ac,etc., e.g., O128ab and O128ac. Many of these O antigens cross-react with other O antigens and to some extent to K antigenswithin E. coli, with other members of the genus and with otherenterobacteria. Recent molecular typing using primers just out-side the O antigen gene cluster (rfb) of 148 representative Ogroups observed unique amplified fragments for each O groupwith sizes ranging from 1.7 to 20 kb (Coimbra et al., 2000).Subsequent MboII digestion of PCR-amplified products resultedin clearly identifiable and reproducible O patterns for the greatmajority of O groups with a variation of band numbers for eachpattern ranging from 5 to 25. Computer analysis identified atotal of 147 O patterns and allowed subdivision of 13 O groups.However, two or more O groups shared a pattern among 13 otherO patterns. The restriction method (rfb-RFLP) is more rapid andmay prove to be more sensitive than conventional serotypingsince 100% of strains are typeable, particularly those that are Orough or nonagglutinating. Additionally, it should facilitate thetyping of strains outside the existing O antigen scheme, whichis restricted to include only O groups of clinical, epidemiological,or scientific relevance. The success and general application ofsuch a typing scheme will require international collaboration todevelop standardized methods for generating, comparing, andmaintaining a database of O patterns.
K antigens The K antigens are the acidic capsular polysac-charide (CPS) antigens. K antigens may be separated into twodistinct groups designated group I and group II. Group I anti-gens, which are composed of high-molecular mass ("100 kDa)CPS, are only found in strains with O groups O8, O9, O20, andO101, and are expressed at both 18! and 37!C. Group I antigensare subdivided according to the absence (IA) or presence (IB)of amino sugars on their CPS. The CPSs of group IA antigensshare structural identity or resemblance to those from Klebsiellaspp., whereas the CPSs of group IB antigens share no structuralresemblance to those from other bacteria. Representative strainsexpressing group IA antigens do not contain the rol (cld) geneencoding the regulator of lipopolysaccharide O-chain length,whereas a similar subset of strains expressing group IB antigenscontains the rol gene (Dodgson et al., 1996). Portions of the CPSsof some group I antigens are attached to the lipid A-core in aform that has been designated KLPS, which will behave similarlyto the traditional O antigens. A good example of a group I KLPS
is K84, which may be operationally defined as an O antigen andwas originally designated as O93.
Group II antigens, which are composed of low molecular mass(#50 kDa) CPS, are found primarily in strains with O groupsthat are associated with extraintestinal disease. The CPSs of manygroup II antigens have structural resemblance or near identityto those from Gram-positive bacteria. These antigens differ widelyin composition and structural features and may be divided intosubgroups based on their acidic components. Twenty to fifty per-cent of the CPS chains are bound to phospholipids. They wereoriginally thought to be temperature dependent, i. e., only ex-pressed at 37!C. However, K2, K3, K10, K11, K19, K54/K96, andK98, which are tentatively classified as group I/II antigens (Finkeet al., 1990), show no temperature regulation of their capsulesand, like group I antigens, do not depend on an elevated CMP-KDO concentration for capsule expression. Based on geneticdata, a subset of the group I/II antigens (K3, K10, and K54/K96)has been designated group III antigens (Pearce and Roberts,1995). A number of K antigens are closely related (indicatedbelow by “$” ) or identical (indicated below by “$” ). The CPSsof some group IB antigens are structurally identical to the sidechains of O antigens and are only considered as K antigens whenco-expressed with another authentic O antigen. The following60 different K antigens are recognized: K1, K2a/ac, K3, K4, K5,K6, K7 ($K56), K8, K9 ($O104), K10, K11, K12 ($K82), K13($ K20 and $ K23), K14, K15, K16, K18a, K18ab ($K22), K19,K24, K26, K27, K28, K29, K30, K31, K34, K37, K39, K40, K41,K42, K43, K44, K45, K46, K47, K49 ($O46), K50, K51, K52, K53,K54 ($ K96), K55, K74, K84, K85ab/ac ($O141), K87 ($O32),K92, K93, K95, K97, K98, K100, K101, K102, K103, KX104, KX105,and KX106. The inclusion of an “X” before the number rep-resents a temporary K antigen designation. A description of theserology, chemistry, and genetics of E. coli O and K antigens isgiven by Ørskov et al. (1977).
H antigens Flagellar or H antigens make up the third maingroup of serotyping antigens. A total of 56 H antigens have beendescribed, but two, H13 and H22, have been removed as beingC. freundii, and H50 has been withdrawn because it is identicalto H10. Cross-reactions are also seen between the H antigens.
Fields et al. (1997) described a tentative molecular methodfor the differentiation of flagellar antigen groups in E. coli basedon restriction fragment length polymorphisms (RFLP) in fliCusing the restriction enzyme RsaI. A wide variety of fliC restrictionfragment patterns was reported among isolates of 53 differentflagellar antigen groups; the majority of the RFLP patterns ob-served corresponded to a unique H antigen group. Limited num-bers of RFLP patterns were observed among members of someH groups, suggesting that the sequence of fliC within certain Hgroups is fairly well conserved. Four patterns were observedamong strains expressing the H7 antigen. Interestingly, the samepattern was detected among all E. coli strains of serotype O157:H7and 16 of 18 of serotype O55:H7, reflecting the common lineageof these strains observed by multilocus enzyme typing (Whittamet al., 1993). Nevertheless, sequencing of a total of 20 H7 fliCgenes, representing 10 different serotypes (Reid et al., 1999;Wang et al., 2000), revealed a notable polymorphism in the fliCgene and identified 10 sequences with differences ranging from0.06% to 3.12%. Recently, a collection of reference strains rep-resenting 48 H types was resolved into 62 patterns (F types) usingHhaI restriction of the fliC gene (Machado et al., 2000). A singleF type was associated with each of 39 H types and more than one

GENUS I. ESCHERICHIA 613
F type was associated with the other nine H types. Antigenicallyrelated H12 and H45 gave a single F type. The determinationof HhaI-fliC F types could allow deduction of all H types andsubdivision of some of these. The two above-mentioned molec-ular typing methods hold promise of a rapid, more refined andspecific typing scheme for the H antigens, which may also behelpful in determining phylogenetic relatedness between differ-ent clones of E. coli. Furthermore, molecular typing has the ad-vantage of allowing typing of nonmotile strains or of strains thatdo not (sufficiently) express the immunoreactive H antigen, andis likely to expand the present number of significantly differentH types. The observed polymorphism in a single determinant,such as the H7 fliC gene, stresses the importance of solid andextensive validation of molecular typing with reference to theexisting serotyping scheme, and calls for caution in the inter-pretation of patterns obtained by DNA fingerprinting methods.
Serotyping Subdivision of E. coli can be carried out in manyways, but serotyping remains one of the most useful ways to sub-divide the species on a global basis. This typing method is basedon the many antigenic differences found in structures on thebacterial surface. A serotype is recorded in the following way:O18ac:K1:H7 or O111:H2 (the latter antigenic formula indicatesthat K antigens are not present in the strain). MR fimbriae, whichare present only in some, often pathogenic, serotypes, can alsobe used for the serological characterization (Ørskov et al., 1977,1980b; Ørskov and Ørskov, 1990) in which case the completeserotype is recorded as O4:K3:H5; F13 or O147:H19; F4ac. Ser-otyping procedures are described in Gross and Rowe (1985) andØrskov and Ørskov (1984a).
Even though complete serotyping involving the many knownO, K, H, and F antigens has been carried out in only a very fewlaboratories, it is well known that the existing number of sero-types is very high.
Antibiotic or drug sensitivity Like other Gram-negative bac-teria, E. coli is intrinsically resistant to hydrophobic antibiotics,such as macrolides, novobiocins, rifamycins, actinomycin D, andfusidic acid (Nikaido, 1996). The structure of the outer mem-brane of E. coli and its role in mediating intrinsic resistance tothese molecules was reviewed by Nikaido (1996). Resistance tothese compounds is attributed, in part, to the low permeabilityof the outer membrane bilayer to lipophilic solutes; however,active efflux mechanisms may have a synergistic effect on resis-tance in certain cases (Nikaido, 1996).
Acquired resistance to aminoglycosides, beta-lactams, chlor-amphenicol, macrolides, sulfonamides, tetracycline, and trimeth-oprim has been described for E. coli strains (reviewed by Quin-tiliani and Courvalin, 1995). Acquired resistance can develop byfour distinct mechanisms: alteration of the target site, enzymaticdetoxification of the antibiotic, decreased drug accumulation,and bypass of an antibiotic-sensitive step. The first three mech-anisms may be mediated by chromosomal mutations or the ac-quisition of plasmids carrying resistance genes. The fourth mech-anism is primarily attributable to the horizontal transfer of anti-biotic resistance genes on a plasmid or transposon. The bio-chemical mechanisms and genetic basis of acquired resistance toantimicrobial agents of clinical importance was discussed byQuintiliani and Courvalin (1995). Genetic methods for the de-tection of antibacterial resistance genes were reviewed by Tenoveret al. (1995).
Pathogenicity E. coli is a natural and essential part of thebacterial flora in the gut of humans and animals. Most E. colistrains are nonpathogenic and reside harmlessly in the colon;however, certain serotypes or clones play an important role inboth intestinal and extraintestinal diseases. The diverse patho-genesis of this bacterium in apparently healthy individuals islargely attributable to its possession of a variety of specific viru-lence factors. In hosts with compromised defenses, E. coli canalso be an excellent opportunistic pathogen.
E. coli in human intestinal diseases E. coli strains isolatedfrom intestinal diseases have been grouped into at least six dif-ferent main categories based on epidemiological evidence, phe-notypic traits, clinical features of the disease they produce, andspecific virulence factors. The currently recognized categories ofdiarrheagenic E. coli include enteropathogenic E. coli (EPEC)(actually a subgroup of attaching and effacing E. coli (A/EEC)defined as eae positive E. coli belonging to both the classical EPECserotypes and nonclassical EPEC serotypes), enterotoxigenic E.coli (ETEC), enteroinvasive E. coli (EIEC), enteroaggregative E.coli (EAggEC), diffusely adherent E. coli (DAEC), and Shiga toxin-producing E. coli (STEC), which are also referred to as Verocytotoxin-producing E. coli (VTEC). These categories are re-viewed below with emphasis on their virulence factors.
Enteropathogenic E. coli (EPEC) Enteropathogenic E.coli was the first category of diarrheagenic E. coli to be recognized.The term enteropathogenic E. coli (EPEC) was originally used torefer to strains belonging to a limited number of O groups thatwere epidemiologically associated with infantile diarrhea (Neteret al., 1955). This rather imprecise definition, which allowed forthe inclusion of a heterogeneous group of pathogens, was usedfor decades and became increasingly problematic as groups ofE. coli that could produce diarrheal disease by the production ofenterotoxins or invasion of intestinal epithelial cells were rec-ognized. The confusion generated by the discovery of new path-ogenic groups of E. coli and the findings that EPEC strains, whichlacked the virulence properties of these newly recognized groups,caused disease in adult volunteers (Levine et al., 1978) promptedresearchers in 1982 to define EPEC as “diarrheagenic E. coli be-longing to serogroups epidemiologically incriminated as patho-gens but whose pathogenic mechanisms have not yet been provento be related to either heat-labile enterotoxins or heat-stableenterotoxins or Shigella-like invasiveness” (Edelman and Levine,1983). As more was learned about the strains associated withinfant diarrhea, the definition was refined to include only certainO:H serotypes associated with illness. Table BXII.c.199 lists someof the O:H serotypes that have been regarded for many years asEPEC. Since 1982, advances in our understanding of the molec-ular aspects of EPEC pathogenesis have allowed researchers tomove beyond the serologic markers that correlate with diseaseto develop a definition based on pathogenic characteristics. Adefinition adopted in 1995 identified the most important char-acteristics of EPEC as its ability to cause attaching and effacing(A/E) histopathology and its inability to produce Shiga toxins(Kaper, 1996). The pathogenesis of EPEC is highlighted belowand has been reviewed in detail by Nataro and Kaper (1998) andWilliams et al. (1997).
The first advance in understanding the pathogenesis of EPECinfection was the discovery that EPEC strains adhere to HEp-2cells in cell culture (Cravioto et al., 1979) in a distinctive patterntermed localized adherence (LA) (Scaletsky et al., 1984). Ex-pression of the LA phenotype in EPEC requires a plasmid re-

FAMILY I. ENTEROBACTERIACEAE614
TABLE BXII.c.199. O:H serotypes regarded as classical and newly recognized EPEC O:H serotypesa,b
O group H antigenc Comments
O26 H%; H11 O26:H% and O26:H11 may also be STEC/VTECd (Levine et al., 1987;Scotland et al., 1990; Bitzan et al., 1991)
O55 H%; H6; H7 O55:H7, H10 and H% may also be STEC/VTEC (Dorn et al., 1989)O86 H%; H8; H34 O86:H% may also be EAggEC (Albert et al., 1993b; Tsukamoto and Takeda,
1993; Schmidt et al., 1995b; Smith et al., 1997b)O86:H8 is a new eae- and bfpA-positive type isolated in Denmark (Scheutz,
unpublished data)O88 H%; H25 (Tsukamoto et al., 1992)O103 H2 New EPEC typeO111 H%; H2; H7; H12 O111:H% may also be STEC/VTEC (Dorn et al., 1989; Bitzan et al., 1991;
Caprioli et al., 1994; Cameron et al., 1995; Allerberger et al., 1996) orEAggEC (Scotland et al., 1991, 1994; Tsukamoto and Takeda, 1993; Chanet al., 1994; Schmidt et al., 1995b; Monteiro-Neto et al., 1997; Morabitoet al., 1998)
O114 H%; H2O119 H%; H2; H6O125ac H%; H6 O125 may also be EAggEC (Tsukamoto and Takeda, 1993; do Valle et al.,
1997; Smith et al., 1997b)O126 H%; H2; H21; H27O127 H%; H6; H21; H40O128ab H%; H2; H7; H12 O128:H2 may also be STEC/VTEC (Beutin et al., 1993a)O142 H%; H6; H34O145 H%; H45 New EPEC typeO157 H%; H8; H16; H45 New EPEC typesO158 H%; H23aData from Cravioto et al. (1979), Levine and Edelman (1984), Levine et al. (1985), Scaletsky et al., 1985,Robins-Browne (1987),Gomes et al. (1989b), Knutton et al. (1989, 1991), Scotland et al. (1989, 1992, 1996), Ørskov and Ørskov (1992), Donnenberg(1995).bO18:H%, H7, H14; O26:H34 and O44:H34 have also been listed but only in Knutton et al. (1991). O18 strains are probably notEPEC (Knutton et al., 1989; Ørskov and Ørskov, 1985). O44:H18 is now considered to belong to the group of enteroaggregativeE. coli (Smith et al., 1994).cNonmotile strains of E. coli are regarded as descendants of motile strains that have lost their motility by mutation(s). Theiroriginal H antigen was often deduced from comparison of biochemical reactions (Kauffmann and Dupont, 1950; Staley et al.,1969).dAbbreviations: EPEC, enteropathogenic E. coli; STEC/VTEC, Shiga toxin-producing E. coli /Vero cytotoxin-producing E. coli;EAggEC, enteroaggregative E. coli.
ferred to as the EPEC adherence factor (EAF) plasmid (Baldiniet al., 1983). A 1-kb DNA probe originally thought to encodethe EPEC adherence factor (EAF) necessary for LA was clonedfrom this plasmid (Nataro et al., 1985a) and has been used ex-tensively as a marker to study the prevalence of EPEC infections(Nataro et al., 1985a; Echeverria et al., 1987, 1991; Gomes et al.,1989a, b; Moyenuddin et al., 1989; Senerwa et al., 1989; Craviotoet al., 1991; Kain et al., 1991; Strockbine et al., 1992; Begaud etal., 1993). EAF plasmids have been found in many EPEC sero-types and range in size from 26 to 76 MDa (Scotland et al., 1989)but typically are 50–70 MDa. Evidence supporting a role for theEAF plasmid in pathogenesis was provided by feeding studiesshowing that volunteers ingesting a plasmid-cured EPEC straindeveloped less diarrhea than those ingesting the plasmid-con-taining parental strain (Levine et al., 1985). Genes at two lociare necessary for expression of the LA phenotype: a cluster of14 genes on the EAF plasmid involved in biogenesis of the bundleforming pilus (BFP), a type-IV pilus, which includes genes en-coding bundlin (bfpA), the major structural subunit of the type-IV pilus, a prepilin peptidase, which processes pre-bundlin to itsmature form, and 12 other proteins, and dsbA on the chromo-some (Donnenberg et al., 1997).
A hallmark of the histopathology of EPEC infections is thepresence of attaching and effacing (A/E) lesions in the intestinaltract. On electron micrographs of jejunal biopsies from childreninfected with EPEC, the bacteria are seen intimately attached tothe epithelial cells on cup-like pedestals composed of depoly-merized cytoskeletal proteins (Knutton et al., 1987). Microvilli
are disrupted as a result of the cytoskeletal rearrangements andeffaced by vesiculation (Fig. BXII.c.192). The intimate attach-ment of EPEC is mediated by a protein known as intimin, whichis a 94-kDa outer membrane protein encoded by the eae gene(E. coli attaching and effacing) ( Jerse and Kaper, 1991). A 1-kbfragment of the eae gene referred to as CVD434 has been cloned( Jerse et al., 1990) and used to screen for attaching and effacingE. coli A/EEC (Bokete et al., 1997), and to characterize enter-opathogenic E. coli (Scotland et al., 1996). Intimins belong to agrowing family of proteins. In human EPEC strains, intimins !,b, c, k (d not now used, although it is in the literature becauseit is a variant of the e variant), and e derivatives have been foundto be serotype-specific and exhibit specific different binding af-finities (Agin and Wolf, 1997). Intimins are also found in rabbitEPEC RDEC-1 strains, A/EEC strains from dogs (Beaudry et al.,1996), pigs (Zhu et al., 1995), and in C. freundii (IntCF) andCitrobacter rodentium (IntCR). Five strains initially identified as Haf-nia alvei were reported to contain the eae gene (Albert et al.,1992), and a protein referred to as IntHA has been characterizedand compared to the above intimins. However, the strains areactually unusual biotypes belonging to the genus Escherichia( Janda et al., 1999), most likely E. coli. The eae gene is only oneof many genes located on a pathogenicity island (PAI) known asthe locus of enterocyte effacement (LEE) (McDaniel et al., 1995).
EPEC strains secrete at least four proteins, Esps for EPECsecreted proteins, encoded by LEE. EspA, B, D, and Tir proteinsare secreted via the type III apparatus and are required for at-taching and effacing activity. Protein secretion, the transcription

GENUS I. ESCHERICHIA 615
FIGURE BXII.c.192. EM of cultured human intestinal mucosa infected with EPEC strain E2348 (E) for 8 h. Bacteria are seen intimately attached tothe epithelial cells on cup-like pedestals composed of depolymerized short filament cytoskeletal proteins (arrows). Brush border microvilli (MV) aredisrupted as a result of the cytoskeletal rearrangements and effaced by vesiculation. (Reproduced with permission from S. Knutton et al., Infectionand Immunity 55: 69–77, 1987, 'American Society for Microbiology.)
of the intimin gene (eae), and the synthesis of the bundle-formingpilus are all affected by growth conditions (Haigh et al., 1995;Jarvis et al., 1995; Kenny and Finlay, 1995) and a plasmid-encodedregulatory region. The plasmid locus is either referred to as theper (plasmid-encoded regulator) genes (perABCD) (Gomez-Duarte and Kaper, 1995) or as an integrated part of the bundle-forming pilus (bfp) operon, genes bfpT, V, and W (Tobe et al.,1996).
EPEC interferes with normal pathways of signal transductionin epithelial cells. EPEC will induce tyrosine phosphorylation;phosphorylation of myosin light chain, vinculin, and !-actinin;in vitro release of intracellular calcium; phospholipase C activity,resulting in elevated levels of inositol phosphates and diacylgly-cerol, in turn activating protein kinase C; reactions that inducehost cell proteins to initiate cytoskeletal rearrangement; and bac-terial uptake (Rosenshine et al., 1992; Manjarrez-Hernandez etal., 1996).
EPEC is not invasive in the same way as Shigella, Yersinia, andSalmonella (Geyid et al., 1996), although some EPEC strains mayexhibit in vitro invasion of epithelial cells at levels comparableto those seen in EIEC (Donnenberg et al., 1989; Pelayo et al.,1999). A 4.5-kb fragment from a large so-called EPEC plasmidof an O111:H% strain negative for both EAF and eae will conferepithelial cell invasivity and the attaching and effacing ability ona noninvasive laboratory strain (Fletcher et al., 1990, 1992). Onlya smaller proportion of EPEC strains hybridize with the clonedfragment.
Of the EPEC O:H serotypes shown in Table BXII.c.199, only16 of the possible 175 recognized O groups are represented and
only a few H antigens, e.g., H2, H6, H7, H12, H21, and H34,occur among several of these 16 O groups. The more recentlydescribed serotypes possessing EPEC-associated virulence mark-ers have been added to the list. Some of these are O86:H8,O88:H25, O103:H2, O127:H40, O142:H34, O145:H45, O157:H8,O157:H16, and O157:H45.
Enterotoxigenic E. coli (ETEC) Enterotoxigenic E. colistrains are important causes of diarrhea in both humans anddomestic animals. ETEC strains do not invade epithelial cells butproduce one or more enterotoxins that are either heat-labile(LT), which is closely related to cholera toxin, or heat-stable (ST)(Cohen and Giannella, 1995). LT-I and ST are plasmid-encoded(LT-II is chromosomally encoded) and found alone or together,and are often associated with a limited number of O:K:H sero-types and O groups. Table BXII.c.200 shows some of the mostcommon serotypes from humans. These enterotoxins cause in-testinal secretion either by activation of guanylate (ST) or aden-ylate (LT) cyclase and are subdivided based on their biologicalactivities, receptors, and chemical and antigenic properties. SThand STp are used to indicate strains of human or porcine origin.The gene encoding another heat-stable enterotoxin called en-teroaggregative heat-stable toxin 1 (EAST1), originally thoughtto be produced by EAggEC only, may also be present in additionto the STa gene. The different variants of enterotoxins are sum-marized in Table BXII.c.201.
There is host specificity among ETEC strains causing diarrheain humans or different species of domestic animals. This is mainlydue to the specific recognition between bacterial colonizationfactors and the epithelial receptors during host-parasite inter-
FAMILY I. ENTEROBACTERIACEAE616
TABLE BXII.c.200. O:(K):H serotypes of human ETEC
O group (K):H antigen
O6 H% K15:H16O7 H% H18O8 K47:H% K25:H9; K40:H9; H10; K87:H19O9 H% K9; K84:H2O11 H27O15 H11; H15; H45O17 K23:H45; H18O20 H% H30O21 H21O25 H% K7:H42; H16O27 H7; H20; H27O29 H?O48 H26O55 H7O56 H%
O63 H12; H30O64 H%
O65 H12O71 H36O73 H45O77 H45O78 H% K2; H11; H12O85 H7O86 H2O88 H25O105 H?O114 H% H21O115 H% H21; H40; H51O119 H6O126 H% H9; H12O128ac H7; H12; H19; H21O133 H16O138 K81O139 H28O141 H% H4O147 H?O148 H28O149 H4; H10; H19O153 H10O159 H% H4; H5; H12; H20; H21; H27; H34; H37O166 H27O167 H5O? H2; H10; H28; K39:H32
TABLE BXII.c.201. Variants of enterotoxins found in ETEC strainsa
Toxin type Subtypes Comments
LT: Heat-labile enterotoxinsLT-I LTp (LTp-I) LTh (LTh-I) Associated with disease
in both humansand animals
LT-II LT-IIa LT-IIb No specific associationwith disease. Rarein human isolates
ST: Heat-stable enterotoxinsSTa (STI) STp (STIa) STh (STIb) Produced by ETEC
and several otherGram-negativebacteria. In ETEC,ST is often foundtogether with thegenes for EAST1.
STb (STII) Induces histologicaldamage in theintestinalepithelium. Mostoften found inporcine ETEC.
aSuffixes commonly used are: h, human variant; p, porcine variant.
TABLE BXII.c.202. O:H serotypes of EIEC
O group H antigen
O28ac H%
O29 H%
O112ac H%
O115 H%
O121 H%
O124 H%; H7; H30; H32O135 H%
O136 H%
O143 H%
O144 H%; H25O152 H%
O159 H2O164 H%
O167 H%; H4; H5O173 H%
action. Most of the fimbriae found in ETEC are typically MRfimbriae. In human ETEC strains at least 21 different surfacestructures called CS, for coli surface antigens, or CFA, for col-onization factor antigens, usually plasmid encoded, have beendescribed (Gaastra and Svennerholm, 1996). In animals, the mostcommon fimbriae are F4 (which was originally described as K88),F5 (originally named K99), F6 (originally named 987P), F17, F41,F42, CS31A, F141, and F165. These fimbriae are found in en-terotoxigenic strains from newborn piglets, pigs, lambs, and new-born calves. A close association between the EAST1 toxin andCS31A, which is related to F4, among pathogenic bovine E. colihas been suggested (Bertin et al., 1998).
More recently, the fimbrial F18 antigen (provisionally desig-nated F107, 2134P, or 8813) has been found in both porcineETEC and STEC/VTEC strains. It is characteristic that the fim-briae usually are found in a limited number of serotypes. Theantigenic variants of F18 fimbriae (F18ab and F18ac) are bio-logically distinct. F18ab fimbriae are expressed poorly both invitro and in vivo and are frequently linked with the productionof Stx2e/VT2e and O group O139, while F18ac are more effi-ciently expressed in vitro and in vivo and most often are linkedwith enterotoxin (STa, STb) production, and O groups O141and O157.
Enteroinvasive E. coli (EIEC) Enteroinvasive E. coli arevery similar to Shigella. Like Shigella, they are capable of invadingand multiplying in the intestinal epithelial cells of the distal largebowel in humans. Approximately two thirds of EIEC strains arelactose negative and virtually all are lysine negative. Multiplechromosomal and plasmid genes are associated with virulence(Acheson and Keusch, 1995). EIEC are restricted to a very limitednumber of serotypes, most of which are nonmotile (TableBXII.c.202).
Enteroaggregative E. coli (EAggEC) EnteroaggregativeE. coli are characterized by a distinct aggregative adherence (AA)pattern to HEp-2 cells in vitro first described by Nataro et al.(1987). This pattern is distinguished by the prominent autoag-glutination of bacterial cells to each other, to the surface of theHEp-2 cells, as well as to the glass cover slip in a characteristiclayering best described as a “stacked brick” configuration. TheAA pattern is plasmid-mediated (Nataro et al., 1985b) and wassuspected to be a putative agent of diarrheal disease as early as1988 (Vial et al., 1988). Volunteer studies with EAggEC haveindicated that certain types will cause diarrhea and other entericsymptoms including borborygmia and cramps (summarized in

GENUS I. ESCHERICHIA 617
TABLE BXII.c.203. O:H serotypes of the most frequently reportedenteroaggregative E. coli (EAggEC)a
O group H antigen References listing these serotypes as EAggEC
O3 H2 Albert et al. (1993b)O15 H18 Vial et al., 1988, Albert et al. (1993b),
Tsukamoto and Takeda (1993), Scotlandet al. (1994)
O44 H%; H18 Scotland et al. (1991, 1994), Tsukamoto andTakeda (1993), Smith et al. (1994),Schmidt et al. (1995b)
O86 H% Albert et al. (1993b), Tsukamoto and Takeda(1993), Schmidt et al. (1995b), Smith etal. (1997b)
O111 H12; H21 Scotland et al. (1991, 1994), Tsukamoto andTakeda (1993), Chan et al. (1994),Schmidt et al. (1995b), Monteiro-Neto etal. (1997), Morabito et al. (1998)
O125 H9; H21 Tsukamoto and Takeda (1993), do Valle etal. (1997), Smith et al. (1997b)
aMany other serotypes of EAggEC have been published by several authors.
Nataro, 1995), and epidemiological studies have implicatedEAggEC as a cause of travelers’ diarrhea (Brook et al., 1994;Scotland et al., 1994). Only about half of the case-control studiesthat have been carried out find significantly higher isolation ratesin cases than in controls (summarized by Law and Chart, 1998).Most recently, EAggEC have been associated with acute andchronic diarrhea and abdominal colic in young children in Ger-many (Huppertz et al., 1997), and with four outbreaks of gas-troenteritis in the UK (Smith et al., 1997b). A DNA probe, re-ferred to as CVD432, has been cloned from the plasmid of strainO42, serotype O44:H18 (formerly an EPEC O:H serotype) andused to identify EAggEC (Baudry et al., 1990). A plasmid en-coded flexible, bundle-forming fimbrial structure, designated ag-gregative adherence fimbria I (AAF/I), is encoded by two regions(Nataro et al., 1992, 1993). Region 2 encodes a transcriptionalactivator of the AraC family of DNA-binding proteins (Nataro etal., 1993). At least one toxin similar to the heat-stable toxin ofETEC, ST designated EAST1, has been identified in an EAggECstrain (Savarino et al., 1991, 1993). The gene encoding EAST1has been shown to be broadly distributed among diarrheagenicE. coli: 100% of 75 O157:H7 STEC/VTEC, 41% of 227 EAggEC,41% of 149 ETEC, 22% of 65 EPEC, and 38% of 47 E. coli fromasymptomatic children hybridized with the EAST1 DNA probe(SS126) (Savarino et al., 1996). Despite the common occurrenceof the EAST1 gene in many diarrheagenic E. coli groups, out-breaks had not been attributed to EAST1-only-producing E. coliuntil recently, when an outbreak in Japan was associated with anO166:H? strain positive for the EAST1 gene (Nishikawa et al.,1999). In Spain, a case-control study suggested that EAST-1 pos-itive E. coli strains are associated with diarrheal diseases in Spanishchildren, whereas EAggEC strains are not (Vila et al., 1998). Thesignificance of these findings remains to be established. Manyserotypes have been observed and some of the most commonlyreported serotypes are listed in Table BXII.c.203.
Diffusely adherent E. coli (DAEC) Diffusely adherent E.coli are defined by the presence of the diffuse adherence (DA)pattern of E. coli strains to HEp-2 cells (Scaletsky et al., 1984;Nataro et al., 1985b). A surface fimbriae designated F1845 con-fers the DA phenotype, and a DNA probe has been cloned (Bilgeet al., 1989). Another adhesin (designated AIDA-I) has also beenassociated with DA of E. coli of serotype O126:H27 (Benz andSchmidt, 1989). The role of DAEC in diarrhea is unclear.
Shiga toxin-producing E. coli (STEC) or Vero cyto-toxin-producing E. coli (VTEC) Shiga toxin-producing E.coli or Vero cytotoxin-producing E. coli strains are characterizedby their ability to produce either one or both of at least twoantigenically distinct, usually bacteriophage-mediated cytotoxinsreferred to as Stx1 or VT1 (first described as Shiga-like toxin I,SLTI) and Stx2 or VT2 (first described as Shiga-like toxin II,SLTII). Whereas STEC/VTEC refers to all E. coli strains that pro-duce Stx/VT in culture supernatants (Konowalchuk et al., 1977,1978), the term enterohemorrhagic E. coli (EHEC) has been usedto refer to strains that have the same clinical and pathogenicfeatures associated with the prototype organism E. coli O157:H7(Levine, 1987). In practice, EHEC is used to describe a subgroupof STEC/VTEC that causes hemorrhagic colitis.
Shiga toxins or Vero cytotoxins belong to the Shiga toxinfamily, comprises the following members: Shiga toxin, which isproduced by Shigella dysenteriae type 1, Stx1/VT1, STx2/VT2, andStx2/VT2 variants (Stxv/VT2v). Stx/VT genes, products, and syn-onyms are summarized in Table BXII.c.204. Stx2e/VT2e, one ofthe variants, is produced by STEC/VTEC strains causing edemadisease (Marques et al., 1987), a usually fatal disease, in weanlingpigs and referred to as Stx2e/VT2e. Unlike all other Stxs/VTs,this is not cytotoxic to HeLa cells and binds to a different re-ceptor, Gb4. The functional receptor for human Stx/VT is theglycolipid globotriosyl ceramide Gb3 (galactose-!-(1-4)-galactose-b-(1-4)-glucose ceramide) (Lingwood et al., 1987; Waddell et al.,1988) found in human renal endothelial cells (Obrig et al.,1993).
Stxs/VTs inhibit protein synthesis by depurination of adeninein 28S rRNA (N-glycosidases), thus inhibiting the elongation fac-tor 1 (EF-1)-dependent aminoacyl-tRNA binding to 60S riboso-mal subunits (Endo et al., 1988). In Vero cells the result is celldeath by apoptosis (Inward et al., 1995).
Nomenclature of Shiga toxins/verocytotoxins Therecognition and investigation of cytotoxin-producing E. coli in-fections by several groups has resulted in the use of differingsystems of nomenclatures for the toxins produced by these bac-teria. In 1994, O’Brien et al. (1994) developed a proposal forrationalizing the nomenclature of the E. coli cytotoxins. In theirproposal, they recommended guidelines for classifying and des-ignating members of the toxin family and stated the toxins shallbe referred to by two alternate but interchangeable names, Shiga-like toxins (SLT) and Vero cytotoxins (VT). Two years later, aproposal to simplify the Shiga-like toxin nomenclature was pub-lished by Calderwood et al. (1996). In this proposal, the word“like” was omitted and the toxins and genes were renamed toreflect their relationship to Shiga toxin, the prototype toxin forthe family. To avoid confusion in the literature, it was suggestedthat cross-reference to existing VT nomenclature could be used.While the omission of the word “like” has received general ac-ceptance in the scientific community, strong arguments for main-taining the existing phenotype nomenclatures for E. coli cyto-toxins were immediately put forward (Karmali et al., 1996), andthe two systems of nomenclature are still being widely used. Tox-ins included in the Shiga toxin/Verocytotoxin family share thefollowing properties: DNA sequence homology and operon struc-ture (A subunit gene immediately upstream of the B subunitgene); polypeptide subunit structure (five B subunits to one Asubunit in the mature holotoxin); enzymatic activity (N-glycosi-dases); binding to specific glycolipid receptors and biologicalproperties, including enterotoxicity in ligated rabbit ileal loops;
FAMILY I. ENTEROBACTERIACEAE618TA
BLE
BX
II.c.
204.
Des
igna
tion
ofstx
/vtx
gene
s,th
eir
prod
ucts
(Stx
/VT
)a ,and
thei
rpr
evio
usde
sign
atio
ns
Toxi
nty
peb
Toxi
nge
neTo
xin
Prot
otyp
eor
gani
sms
(ref
eren
ce)
Sero
type
ofpr
otot
ype
orga
nism
Prev
ious
desi
gnat
ion
orsy
nony
mfo
rto
xin
gene
Prev
ious
desi
gnat
ion
orsy
nony
mfo
rto
xin
1stx
1/vt
x1St
x1/V
T1
EDL9
33(O
’Bri
enet
al.,
1984
)O
157:
H7
slt-I
SLT-
IH
19(K
onow
alch
uket
al.,
1977
)O
26:H
11H
30(K
onow
alch
uket
al.,
1977
)O
26:H
11stx
1-O
103/
vtx1
-O10
3St
x1-O
103/
VT
1-O
103
PMK
1(M
aria
ni-K
urkd
jian
etal
.,19
93)
O10
3:H
2
stx1-
O11
1-P
H/v
tx1-
O11
1-PH
Stx1
-O11
1-PH
/VT
1-O
111-
PHPH
(Pat
onet
al.,
1993
a)O
111:
H%
sltI/
PHSl
t-I/P
Hstx
1-O
X3/
vtx1
-OX
3St
x1-O
X3/
VT
1-O
X3
131/
3(P
aton
etal
.,19
95a)
OX
3:H
8slt
I/O
X3
Slt-I
/OX
3stx
1-O
48/v
tx1-
O48
Stx1
-O48
/VT
1-O
4894
C(P
aton
etal
.,19
95a)
O48
:H21
sltI/
O48
Slt-I
/O48
stx1-
O11
1-C
B16
8/vt
x1-O
111-
CB
168
Stx1
-O11
1-C
B16
8/V
T1-
O11
1-C
B16
8C
B16
8(P
aton
etal
.,19
95a)
O11
1:H
%slt
I/C
BSl
t-I/C
B
2stx
2/vt
x2St
x2/V
T2
EDL9
33(O
’Bri
enet
al.,
1984
)O
157:
H7
slt-II
SLT-
II2c
stx2c
/vtx
2cSt
x2c/
VT
2cE3
2511
(Sch
mitt
etal
.,19
91)
O15
7:H
%St
x2v/
VT
2vslt
-IIc
SLT-
IIc
stx2-
O22
/vtx
2-O
22St
x2-O
22/V
T2-
O22
KY-
O19
(Lin
etal
.,19
93a)
O22
:H%
VT
2v(p
KT
N10
54)
stx2-
O15
7-T
K-5
1/vt
x2-O
157-
TK
-51
Stx2
-O15
7-T
K-5
1/V
T2-
O15
7-T
K-
51T
K-5
1(L
inet
al.,
1993
a)O
157:
H7
VT
2v(p
KT
N10
50)
stx2-
OX
3/a-
031/
vtx2
-OX
3/a-
031
Stx2
-OX
3/a-
031/
VT
2-O
X3/
a-03
103
1(P
aton
etal
.,19
92)
OX
3:H
21stx
2vO
X39
2SL
T-II
/OX
3/V
T2d
-OX
3d;S
LT-
II/O
X3a
stx2-
OX
3/b-
031/
vtx2
-OX
3/b-
031
Stx2
-OX
3/b-
031/
VT
2-O
X3/
b-03
103
1(P
aton
etal
.,19
93b)
OX
3:H
21stx
2vO
X39
3SL
T-II
/OX
3/2/
VT
2d-O
X3/
2c ;SL
T-II
/OX
3bstx
2-O
48/v
tx2-
O48
Stx2
-O48
/VT
2-O
4894
C(P
aton
etal
.,19
95b)
O48
:H21
SLT-
II/O
48stx
2-O
111-
PH
/vtx
2-O
111-
PHSt
x2-O
111-
PH/V
T2-
O11
1-PH
CB
168
(Pat
onet
al.,
1995
b)O
111:
H%
stx2v
O11
1SL
T-II
/O11
1/V
T2d
-O11
1c
stx2-
O11
8/vt
x2-O
118
Stx2
-O11
8/V
T2-
O11
8dEH
250
(Pie
rard
etal
.,19
98)
O11
8:H
12St
x2d/
VT
2d-O
unt
stx2-
O6/
vtx2
-O6
Stx2
-O6/
VT
2-O
6N
V20
6(B
ertin
etal
.,20
01)
O6:
H10
stx2-
NV
206
Stx2
-NV
206
2dd
stx2d
/vtx
2dSt
x2d/
VT
2dB
2F1
(Ito
etal
.,19
90)
O91
:H21
SLT-
IIvh
stx2d
1/vt
x2d1
Stx2
d1/V
T2d
1B
2F1
(Ito
etal
.,19
90)
O91
:H21
stx2v
ha/v
tx2v
haSt
x2vh
a/V
T2v
hastx
2ha
Stx2
vh-a
/VT
2vh-
aV
T2v
-aSL
T-II
vha
stx2d
2/vt
x2d2
Stx2
d2/V
T2d
2B
2F1
(Ito
etal
.,19
90)
O91
:H21
stx2v
hb/v
tx2v
hbstx
2hb
Stx2
vhb/
VT
2vhb
Stx2
vh-b
/VT
2vh-
bV
T2v
-bSL
T-II
vhb
2estx
2e/v
tx2e
Stx2
e/V
T2e
412
(Gyl
eset
al.,
1988
)O
139:
K12
:H1
slt-II
vSL
T-II
vS1
191
(Wei
nste
inet
al.,
1988
)O
139:
H1
slt-II
vaSL
T-II
vae
slt-II
eSL
TII
e/V
TeV
T2v
pV
T2v
p12f
fstx
2f/v
tx2f
Stx2
f/V
T2f
H.I.
8(G
anno
net
al.,
1990
)O
128:
H2
stx2e
v/vt
x2ev
Stx2
ev/V
T2e
vT
4/97
(Sch
mid
tet
al.,
2000
)O
128:
H2
Stx2
vp2/
VT
2vp2
VTe
vslt
-IIvh
cSL
TII
vhc
slt-II
ddSL
T-II
d/V
T2d
d
a Dat
aco
mpi
led
from
refe
renc
esB
astia
net
al.(
1998
),B
ertin
etal
.(20
01),
Cal
derw
ood
etal
.(19
96),
Gan
non
etal
.(19
90),
Ito
etal
.(19
90),
W.M
.Joh
nson
etal
.(19
91),
Kar
mal
iet
al.(
1996
),K
okai
-Kun
etal
.(20
00),
Lin
etal
.(19
93a,
b),T
aked
aet
al.(
1993
),an
dT
yler
etal
.(19
91).
b Toxi
nty
pes
are
defin
edac
cord
ing
toan
tigen
icva
riab
ility
,diff
eren
ces
into
xici
tyfo
rtis
sue
cultu
rece
llsan
d/or
anim
als,
thei
rca
paci
tyto
beac
tivat
edby
mou
seel
asta
se,a
nddi
ffer
ence
sin
DN
Aor
amin
oac
idse
quen
ces.
c The
rear
ese
vera
lto
xins
suffi
xed
byd
inth
elit
erat
ure:
The
Stx2
d/V
T2d
toxi
nsof
O91
:H21
(Mel
ton-
Cel
saan
dO
’Bri
en,1
998)
,the
VT
2d($
stx2
-Ogr
oup/
stra
inde
sign
atio
nan
d/or
year
)va
rian
tsby
Pato
net
al.(
1992
,19
93a,
1995
b),P
iera
rdet
al.(
1998
),an
dth
eSL
T-II
d/V
T2d
($St
x2f/
VT
2f)
toxi
npr
oduc
edby
stra
inH
.I.8
(ser
otyp
eO
128:
H2)
aspr
opos
edby
Gyl
es(1
994)
.We
supp
ortt
heus
eof
the
dde
sign
atio
nfo
rac
tivat
able
Stx2
/VT
2to
xins
aspr
opos
edby
Mel
ton-
Cel
saan
dO
’Bri
en,(
1998
).dSt
x2-O
118/
VT
2-O
118
(for
mer
lykn
own
asV
T2d
-Oun
t)is
expe
cted
tobe
nona
ctiv
atab
leba
sed
onan
alys
isof
the
nucl
eotid
ese
quen
ce(D
enis
Pier
ard
and
Ang
ela
Mel
ton-
Cel
sa;u
npub
lishe
d);t
heor
igin
alst
rain
has
been
rety
ped
asO
118:
H12
(Lot
har
Beu
tinan
dFl
emm
ing
Sche
utz,
unpu
blis
hed)
.e T
his
desi
gnat
ion
has
also
been
refe
rred
tofo
rth
eSt
x2f/
VT
2fto
xin
prod
uced
byst
rain
H.I.
8(s
erot
ype
O12
8:H
2).
f The
nucl
eotid
ese
quen
ceof
the
form
erstx
2ev/
vtx2
evof
stra
inH
.I.8
(ser
otyp
eO
128:
H2)
isne
arly
iden
tical
toth
ere
cent
lypu
blis
hed
stx2f
/vtx
2ffo
und
inst
rain
T4/
97(s
erot
ype
O12
8:H
2)fr
omfe
ralp
igeo
ns(S
chm
idte
tal.,
2000
).A
sits
nucl
eotid
ese
quen
ceis
dist
inct
lydi
ffer
ent
from
thos
eof
the
othe
rSt
x2/V
T2
toxi
nsan
dva
rian
tsas
wel
las
Stx1
/VT
1,w
esu
ppor
tth
epr
opos
alof
rena
min
gstx
2ev/
vtx2
evas
stx2f
/vtx
2f.

GENUS I. ESCHERICHIA 619
neurotoxicity in mice; and cytotoxicity to receptor-expressing tis-sue culture cell lines, such as Vero and HeLa cell lines. Classi-fication of the toxins into major toxin types, designated withArabic numbers, is based on differences that result in no cross-neutralization by homologous polyclonal antisera and no DNA–DNA cross-hybridization of their genes under conditions of highstringency. Toxin subtypes, designated with letters added to thetype name, share cross-hybridization of their genes under highstringency but show significant differences in biologic activity,including the capacity to be activated; serologic reactivity; or re-ceptor binding. Table BXII.c.204 summarizes the currently re-ported Shiga toxin/Verocytotoxin genes and toxins, prototypeorganisms, and their previous designations in the literature.
The distinction between toxin types 2c and 2d in TableBXII.c.204 is based on differences in the A subunit determiningwhether the toxin is activatable (Stx2d/VT2d) or nonactivatable(Stx2c/VT2c). The other Stx2/VT2 subtypes described in theliterature, which have not been tested for all properties, havebeen tentatively placed together with toxin type 2c, primarilybased on similarities in nucleotide sequences that place them ina phylogenetically related Stx2/VT2 cluster including all variants.Based on their degree of overall nucleotide sequence similarity,Stx2/VT2 toxins fall into two phylogenetically distinct groups(Bastian et al., 1998; Pierard et al., 1998). Toxins in group 1,which include Stx2/VT2, Stx2c/VT2c (including variants Stx2-O22/VT2-O22, Stx2-O157-TK-51/VT2-O157-TK-51, Stx2-OX3/b-031/VT2-OX3/b-031, Stx2-O48/VT2-O48), and Stx2d/VT2d,share 99.1–99.2% and 95.9–98.5% nucleotide sequence similarityin their A and B subunits, respectively; while toxins in group 2,which include Stx2-OX3/a-031/VT2-OX3/a-031, Stx2-O111-PH/VT2-O111-PH, and Stx2-O118/VT2-O118, share 96.9–99.9 % and99.6–100% nucleotide sequence similarity in their A and B sub-units, respectively. The similarities of individual sequences be-tween group 1 and group 2 are 93.4–96.0% for the A subunitsand 86.2–89.3% for the B subunits. The extent of, and differencesbetween, toxicity for tissue culture cells and/or animals and theircapacity to be activated are not fully established for all the types.Further analysis against all the criteria necessary to allow defin-itive placement with appropriate other toxin types is required.In the absence of this information, suffixes have been addedafter the O group of the source organisms, and, when necessary,the original strain designation. Stx2/VT2 subtype toxins foundin the same original strain are suffixed “/a” , “/b” etc. as in Stx2-OX3/a-031/VT2-OX3/a-031 and Stx2-OX3/b-031/VT2-OX3/b-031.
For a brief presentation and practical use of the subtyping ofStx/VT genes with 4 Stx1/VT1 and 9 Stx2/VT2 oligonucleotideDNA probes, and 3 Stx1/VT1 and 8 Stx2/VT2 primer pairs, seeSmith et al. (1993), Yamasaki et al. (1996), Pierard et al. (1997),and Bastian et al. (1998).
Like EPEC, some STEC/VTEC strains have been shown tocause attaching and effacing lesions in vivo (Hall et al., 1990),in animal models (Francis et al., 1986; Tzipori et al., 1986; Sher-man et al., 1988), and in vitro (Knutton et al., 1989). Two separategroups have cloned, sequenced, and characterized the eae hom-ologue from VTEC O157:H7 (Beebakhee et al., 1992; Yu andKaper, 1992). Homology between EPEC and STEC/VTEC se-quences was 86% and 83% at the nucleotide and amino acidlevels, respectively (Yu and Kaper, 1992), and the STEC/VTECeae sequence was 97% homologous to the EPEC eae gene for thefirst 2200 bp and 59% homologous over the last 800 bp (Bee-bakhee et al., 1992). Both eae sequences show 50% homology to
the central region of the Yersinia pseudotuberculosis inv gene ( Jerseet al., 1990; Beebakhee et al., 1992), and the predicted aminoacid sequence of the STEC/VTEC eae gene share 31% identityand 51% similarity with the invasin molecule of Yersinia pseudo-tuberculosis (Yu and Kaper, 1992).
Serotype specific heterogeneity of the eae gene in STE-C/VTEC strains O55:H7 or H%, O111:H8 and O157:H7 or H%,and in O groups O26, O103, and O157 has been demonstrated(Gannon et al., 1993; Louie et al., 1994).
Almost all STEC/VTEC O157:H7 strains harbor a large 60-65MDa plasmid ( Johnson et al., 1983), designated pO157, thatplays a role in virulence (Karch et al., 1987) and a small plasmidof 6.6 kb found in O157:H7 STEC/VTEC strains appear to syn-thesize colicin D (Bradley et al., 1991). O26:H11 strains alsopossess at least one plasmid in the range of 55–70 MDa. Restric-tion enzyme patterns of plasmids from other O:H serotypes (in-cluding O5, O91, O103, O111, O121, and O127) show a notablesimilarity with the large plasmids in O157 and O26 strains (Levineet al., 1987). A 3.4-kb fragment from a large plasmid of O157:H7(prototype EDL 933) has been cloned and used as a DNA probe(referred to as CVD419) to identify EHEC plasmids (Levine etal., 1987); i.e., large plasmids found in Verocytotoxin producingE. coli strains. DNA probing with gene probes defining the in-compatibility group of plasmids indicates that the STEC/VTECplasmids share an approximately 23-kb fragment with EPEC plas-mids and that the large plasmids of both EPEC and STEC/VTECconstitute a family of transfer-deficient Inc F-IIA plasmids (Haleset al., 1992), while sequencing of pO157 reveals high homologyto the orf1 of the RepFIB replicon (Schmidt et al., 1996a).
The large plasmid of O157 encodes the EHEC-hemolysin(Ehx), which is homologous to the E. coli ! hemolysin (Schmidtet al., 1994, 1995a), and a novel catalase-peroxidase, KatP (Brun-der et al., 1996). In contrast to ! hemolysin, Ehx can be detectedonly on blood agar plates containing washed sheep erythrocytes.The zones of hemolysis on these plates are smaller and moreturbid than those caused by ! hemolysin and require overnightincubation before they become visible (Beutin, 1991; Beutin etal., 1988, 1989). A role for Ehx in the pathogenesis of diarrhealdisease has not been demonstrated. Because ! hemolysin-pro-ducing strains are uncommon in feces, these hemolysins serveas useful phenotypic markers for the detection of the majorityof STEC/VTEC organisms.
The genes encoding the Ehx constitute a typical RTX (repeatsin toxin) determinant, the Ehx-operon, with the gene orderCABD (Schmidt et al., 1996a). The ehxA gene encodes the activeprotein, and ehxB and ehxD share high sequence homology withother RTX transport proteins (Schmidt et al., 1995a, 1996a). Like! hemolysin, the Ehx is a highly active cytolysin of the RTX familywith a similar but not identical pore-forming capacity (Schmidtet al., 1996b). The Ehx plasmid DNA probe (CVD419) coversthe ehxA and part of the ehxB gene (Schmidt et al., 1995a).
Two other enterohemolysins Ehly1 and Ehly2 have been de-scribed (Beutin et al., 1993b; Stroeher et al., 1993). Ehly1 is a33-kDa cell-associated protein encoded by a bacteriophage,UC3888, found in O26:H11 STEC/VTEC. Ehly1 has no knownsequence homology to any other DNA or protein sequence. TheEhly2 enterohemolysin is also encoded by a bacteriophage,UC3208, found in O26:H11. It is in part homologous to DNA ofbacteriophagek; but completely unrelated to Ehly1 (Beutin etal., 1993b).
Most information on the source and transmission of STEC/VTEC has been learned from outbreak investigations. Findings

FAMILY I. ENTEROBACTERIACEAE620
from these investigations showed that most outbreaks are relatedto carriage of the organism in ruminants, especially cattle, whichshow no symptoms of disease. During the period from 1982 to1993, at least 20 outbreaks of O157:H7 have been reported inthe USA (summarized in Anonymous, 1994). These outbreakshave affected 1509 patients, resulting in the hospitalization of346 patients, 86 cases of HUS, and 19 deaths. This increaseddramatically in the following years with 13 outbreaks in 1993 and30 outbreaks in 1994 (Armstrong et al., 1996). The largest mul-tistate outbreak in the USA occurred in early 1993 with morethan 700 illnesses and 4 deaths (Bell et al., 1994; Davis, 1994).It has been estimated that E. coli O157:H7 causes 73,000 illnessesannually in the United States and non-O157 STEC/VTEC, 37,000illnesses; and that 91 deaths occur each year in the USA (Meadet al., 1999). In Canada, 15 outbreaks were reported in 1982–1987 with 242 cases, 24 cases of HUS, and 15 deaths (Karmali,1989). The first recognized community outbreak of O157:H7 inEurope occurred in the UK in the summer of 1985 affecting atleast 24 persons. Eleven patients were hospitalized and one died(Morgan et al., 1988). In England and Wales, O157:H7 was iso-lated from 39% of sporadic cases of hemorrhagic colitis (Smithet al., 1987) and 33% of sporadic HUS cases (Scotland et al.,1988). In an outbreak of HUS in the West Midlands, O157:H7was isolated from 33% of cases (Taylor et al., 1986a; Willshaw etal., 2001). Subsequent outbreaks and sporadic cases in the UKhave been reported (Salmon et al., 1989). Scotland has one ofthe highest rates of infection with O157 increasing from1.37/100,000 of the population in 1989 (Thomas et al., 1996a)to 32.3/100,000 in 1996 (Reilly and Carter, 1997). The worstfood poisoning outbreak with O157 STEC/VTEC in Scotlandoccurred in 1996 with 501 cases; 151 were hospitalized and 20elderly died (Ahmed and Donaghy, 1998).
O157 STEC/VTEC has been isolated from outbreaks andfrom sporadic cases of diarrhea and HUS in many parts of theworld: Canada, UK, Argentina, Germany, Central Europe, Chile,and Italy (summarized by Griffin, 1995; see also Chapters 2–9 inKaper and O’Brien, 1998). Sakai City in Japan experienced thelargest outbreak of O157 STEC/VTEC ever recorded in July1996, which was part of several outbreaks that summer with anestimated number of a little less than 8,000 cases and 6 deaths(Infectious Disease Surveillance Center, Japan, 1997).
Other STEC/VTEC O:H serotypes have caused outbreaks ofdiarrhea and HUS: O111:H%, O145:H%, and O?:H19 in Japan(Kudoh et al., 1994); O26:H11 in the Czech Republic (Bielasz-ewska et al., 1990), USA (Brown et al., 1998), and Ireland (Mc-Master et al., 2001); O103:H2 in France (Mariani-Kurkdjian etal., 1993); O104:H21 in the USA (Centers for Disease Control,1995); O111:H% in Australia (Cameron et al., 1995), Italy (Ca-prioli et al., 1994), USA (Banatvala et al., 1996; Centers for Dis-ease Control, 2000), Spain (Blanco et al., 1996), and France(Boudailliez et al., 1997; Mariani-Kurkdjian et al., 1997);O113:H21 in Australia (Paton et al., 1999); and O119 in France(Deschenes et al., 1996). A clone of sorbitol fermenting O157:H7STEC/VTEC has been isolated from patients with diarrhea andHUS (Gunzer et al., 1992; Karch et al., 1993) and caused anoutbreak in Germany (Ammon et al., 1999). Studies in Europeindicate that non-O157 STEC/VTEC strains are increasing infrequency as a cause of hemolytic-uremic syndrome (HUS) andfound much more commonly in children with diarrhea (Ver-weyen et al., 1999; Scheutz et al., 2001; Tozzi et al., 2001).
Among the over 400 STEC/VTEC serotypes, and apart fromO157:H% and O157:H7, those in O groups O26, O103, O111,
and O145 are most commonly isolated from humans worldwide.These, along with strains that have caused outbreaks, are clearlyrecognized as pathogens. Table BXII.c.205 shows the non-O157STEC/VTEC serotypes that have been isolated from humans.
E. coli in human extraintestinal infections
Extraintestinal pathogenic E. coli (ExPEC) Extrain-testinal pathogenic E. coli (ExPEC) are E. coli strains that possesscurrently recognized extraintestinal virulence factors or havebeen demonstrated to possess enhanced virulence in an appro-priate animal model (Russo and Johnson, 2000). ExPEC strainsprimarily belong to pathogenic clones of a limited number ofO:K:H serotypes (Ørskov and Ørskov, 1975), usually with MRfimbriae (P and S fimbriae), siderophores (e.g., aerobactin), hostdefense-avoidance mechanisms such as capsules (Ørskov and Ør-skov, 1977; Ørskov et al., 1982), O antigens, and serum resistanceand toxins (often !-hemolysin). ExPEC is common in all agegroups and may occur at almost any extraintestinal site. The mostcommon infections include urinary tract infections (UTIs) rang-ing from uncomplicated to febrile to invasive, pyelonephritis,neonatal, and postneurosurgical meningitis and septicemia. Thisgroup is epidemiologically and phylogenetically distinct fromcommensal and intestinal strains of E. coli (Picard et al., 1999).Virulence genes are often located on pathogenicity islands(PAIs), which have the tendency to delete with high frequenciesor may undergo duplications and amplifications. They are oftenassociated with tRNA loci, which may represent target sites forthe chromosomal integration of these elements (Hacker et al.,1997). Many produce toxins that can lyse erythrocytes of differentmammalian species. The best characterized of these is !-hemo-lysin, which is often produced by strains causing UTIs ( J.R. John-son, 1991) and is believed to play an important role. !-Hemolysinis secreted and can be demonstrated in culture fluid filtrates(Beutin, 1991). Hemolytic colonies can also be identified by theclear zone of hemolysis produced on blood agar plates after 3–4 h of incubation.
Uropathogenic E. coli (UPEC) This somewhat misleadingacronym (clones or virulence factors are not syndrome-specific)has been used to refer to the majority of specific clonal groupsof uropathogenic E. coli isolated from UTIs including pyelone-phritis. They are characterized by a number of virulence factorsthat together play a role in their pathogenesis. First, UPEC isdominated by a limited number of O groups with O groups O1,O2, O4, O6, O7, O18ac, O75, O16, and O15 as the most com-monly isolated (Ørskov and Ørskov, 1985). These strains are alsorepresented by a limited number of K antigens: K1, K2, K3, K5,K12, and K13. Common serotypes include O1:K1:H7, O2:K1:H4,O4:K12:H1, O4:K12:H5, O6:K2:H1, O6:K5:H1, O6:K13:H1 (cys-titis), O16:K1:H6, and O18ac:K5:H7. Furthermore, the majorityof UPECs express P fimbriae of F types F7 through F16 and /orS fimbriae.
Neonatal meningitis E. coli (NMEC) Neonatal menin-gitis E. coli is frequently associated with O groups O7, O18ac, O1,and O6 that have the K1 antigen identical to the capsule of N.meningitidis type B (Sarff et al., 1975). O83:K1 strains are alsocommon but apparently only in Europe (Ørskov and Ørskov,1985). One of the most commonly isolated types is an S fimbri-ated clone of serotype O18ac:K1:H7.
E. coli in animal infections As is the case in humans,certain strains of E. coli can cause disease in animals. In farmanimals, E. coli strains are associated with a variety of pathological
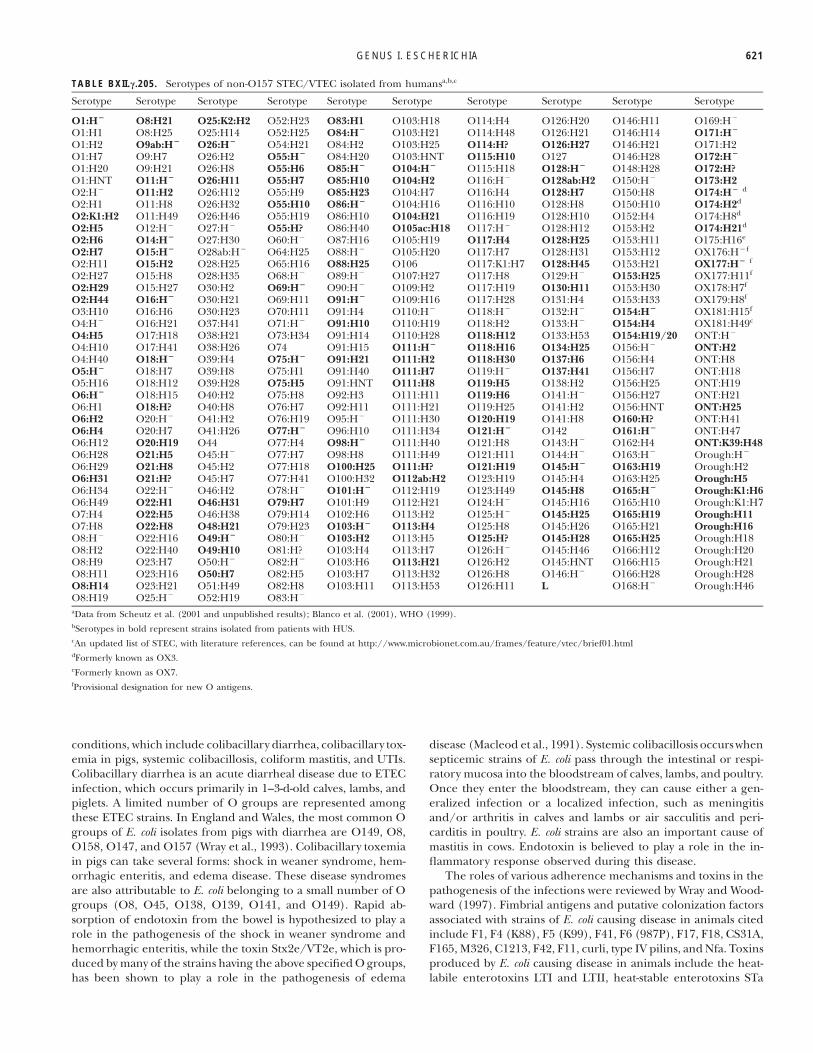
GENUS I. ESCHERICHIA 621
TABLE BXII.c.205. Serotypes of non-O157 STEC/VTEC isolated from humansa,b,c
Serotype Serotype Serotype Serotype Serotype Serotype Serotype Serotype Serotype Serotype
O1:H(
O1:H1O1:H2O1:H7O1:H20O1:HNTO2:H%
O2:H1O2:K1:H2O2:H5O2:H6O2:H7O2:H11O2:H27O2:H29O2:H44O3:H10O4:H%
O4:H5O4:H10O4:H40O5:H(
O5:H16O6:H(
O6:H1O6:H2O6:H4O6:H12O6:H28O6:H29O6:H31O6:H34O6:H49O7:H4O7:H8O8:H%
O8:H2O8:H9O8:H11O8:H14O8:H19
O8:H21O8:H25O9ab:H(
O9:H7O9:H21O11:H(
O11:H2O11:H8O11:H49O12:H%
O14:H(
O15:H(
O15:H2O15:H8O15:H27O16:H(
O16:H6O16:H21O17:H18O17:H41O18:H(
O18:H7O18:H12O18:H15O18:H?O20:H%
O20:H7O20:H19O21:H5O21:H8O21:H?O22:H%
O22:H1O22:H5O22:H8O22:H16O22:H40O23:H7O23:H16O23:H21O25:H%
O25:K2:H2O25:H14O26:H(
O26:H2O26:H8O26:H11O26:H12O26:H32O26:H46O27:H%
O27:H30O28ab:H%
O28:H25O28:H35O30:H2O30:H21O30:H23O37:H41O38:H21O38:H26O39:H4O39:H8O39:H28O40:H2O40:H8O41:H2O41:H26O44O45:H%
O45:H2O45:H7O46:H2O46:H31O46:H38O48:H21O49:H(
O49:H10O50:H%
O50:H7O51:H49O52:H19
O52:H23O52:H25O54:H21O55:H(
O55:H6O55:H7O55:H9O55:H10O55:H19O55:H?O60:H%
O64:H25O65:H16O68:H%
O69:H(
O69:H11O70:H11O71:H%
O73:H34O74O75:H(
O75:H1O75:H5O75:H8O76:H7O76:H19O77:H(
O77:H4O77:H7O77:H18O77:H41O78:H%
O79:H7O79:H14O79:H23O80:H%
O81:H?O82:H%
O82:H5O82:H8O83:H%
O83:H1O84:H(
O84:H2O84:H20O85:H(
O85:H10O85:H23O86:H(
O86:H10O86:H40O87:H16O88:H%
O88:H25O89:H%
O90:H%
O91:H(
O91:H4O91:H10O91:H14O91:H15O91:H21O91:H40O91:HNTO92:H3O92:H11O95:H%
O96:H10O98:H(
O98:H8O100:H25O100:H32O101:H(
O101:H9O102:H6O103:H(
O103:H2O103:H4O103:H6O103:H7O103:H11
O103:H18O103:H21O103:H25O103:HNTO104:H(
O104:H2O104:H7O104:H16O104:H21O105ac:H18O105:H19O105:H20O106O107:H27O109:H2O109:H16O110:H%
O110:H19O110:H28O111:H(
O111:H2O111:H7O111:H8O111:H11O111:H21O111:H30O111:H34O111:H40O111:H49O111:H?O112ab:H2O112:H19O112:H21O113:H2O113:H4O113:H5O113:H7O113:H21O113:H32O113:H53
O114:H4O114:H48O114:H?O115:H10O115:H18O116:H%
O116:H4O116:H10O116:H19O117:H%
O117:H4O117:H7O117:K1:H7O117:H8O117:H19O117:H28O118:H%
O118:H2O118:H12O118:H16O118:H30O119:H%
O119:H5O119:H6O119:H25O120:H19O121:H(
O121:H8O121:H11O121:H19O123:H19O123:H49O124:H%
O125:H%
O125:H8O125:H?O126:H%
O126:H2O126:H8O126:H11
O126:H20O126:H21O126:H27O127O128:H(
O128ab:H2O128:H7O128:H8O128:H10O128:H12O128:H25O128:H31O128:H45O129:H%
O130:H11O131:H4O132:H%
O133:H%
O133:H53O134:H25O137:H6O137:H41O138:H2O141:H%
O141:H2O141:H8O142O143:H%
O144:H%
O145:H(
O145:H4O145:H8O145:H16O145:H25O145:H26O145:H28O145:H46O145:HNTO146:H%
L
O146:H11O146:H14O146:H21O146:H28O148:H28O150:H%
O150:H8O150:H10O152:H4O153:H2O153:H11O153:H12O153:H21O153:H25O153:H30O153:H33O154:H(
O154:H4O154:H19/20O156:H%
O156:H4O156:H7O156:H25O156:H27O156:HNTO160:H?O161:H(
O162:H4O163:H%
O163:H19O163:H25O165:H(
O165:H10O165:H19O165:H21O165:H25O166:H12O166:H15O166:H28O168:H%
O169:H%
O171:H(
O171:H2O172:H(
O172:H?O173:H2O174:H( d
O174:H2d
O174:H8d
O174:H21d
O175:H16e
OX176:H%f
OX177:H( f
OX177:H11f
OX178:H7f
OX179:H8f
OX181:H15f
OX181:H49c
ONT:H%
ONT:H2ONT:H8ONT:H18ONT:H19ONT:H21ONT:H25ONT:H41ONT:H47ONT:K39:H48Orough:H%
Orough:H2Orough:H5Orough:K1:H6Orough:K1:H7Orough:H11Orough:H16Orough:H18Orough:H20Orough:H21Orough:H28Orough:H46
aData from Scheutz et al. (2001 and unpublished results); Blanco et al. (2001), WHO (1999).bSerotypes in bold represent strains isolated from patients with HUS.cAn updated list of STEC, with literature references, can be found at http://www.microbionet.com.au/frames/feature/vtec/brief01.htmldFormerly known as OX3.eFormerly known as OX7.fProvisional designation for new O antigens.
conditions, which include colibacillary diarrhea, colibacillary tox-emia in pigs, systemic colibacillosis, coliform mastitis, and UTIs.Colibacillary diarrhea is an acute diarrheal disease due to ETECinfection, which occurs primarily in 1–3-d-old calves, lambs, andpiglets. A limited number of O groups are represented amongthese ETEC strains. In England and Wales, the most common Ogroups of E. coli isolates from pigs with diarrhea are O149, O8,O158, O147, and O157 (Wray et al., 1993). Colibacillary toxemiain pigs can take several forms: shock in weaner syndrome, hem-orrhagic enteritis, and edema disease. These disease syndromesare also attributable to E. coli belonging to a small number of Ogroups (O8, O45, O138, O139, O141, and O149). Rapid ab-sorption of endotoxin from the bowel is hypothesized to play arole in the pathogenesis of the shock in weaner syndrome andhemorrhagic enteritis, while the toxin Stx2e/VT2e, which is pro-duced by many of the strains having the above specified O groups,has been shown to play a role in the pathogenesis of edema
disease (Macleod et al., 1991). Systemic colibacillosis occurs whensepticemic strains of E. coli pass through the intestinal or respi-ratory mucosa into the bloodstream of calves, lambs, and poultry.Once they enter the bloodstream, they can cause either a gen-eralized infection or a localized infection, such as meningitisand/or arthritis in calves and lambs or air sacculitis and peri-carditis in poultry. E. coli strains are also an important cause ofmastitis in cows. Endotoxin is believed to play a role in the in-flammatory response observed during this disease.
The roles of various adherence mechanisms and toxins in thepathogenesis of the infections were reviewed by Wray and Wood-ward (1997). Fimbrial antigens and putative colonization factorsassociated with strains of E. coli causing disease in animals citedinclude F1, F4 (K88), F5 (K99), F41, F6 (987P), F17, F18, CS31A,F165, M326, C1213, F42, F11, curli, type IV pilins, and Nfa. Toxinsproduced by E. coli causing disease in animals include the heat-labile enterotoxins LTI and LTII, heat-stable enterotoxins STa

FAMILY I. ENTEROBACTERIACEAE622
and STb, cytotoxic necrotizing factors 1 and 2 (CNF1 and CNF2),and Shiga/Verocytotoxins.
Other typing methods For a description of other methodsfor subdivision of E. coli, i.e., phage typing, colicin typing, bio-typing, typing by outer membrane protein (OMP) pattern, typingby antibiotic resistance patterns, and typing by direct hemagglu-tination, see Ørskov and Ørskov (1984a) and Sussman (1985).Phage typing is very useful for certain antigens because antiseraare difficult to produce. This is particularly true for K1 (Grosset al., 1977), K3, K5, K7, K12, K13 (Nimmich et al., 1992), andK95 (Nimmich, 1994). A phage-typing scheme for STEC/VTECO157:H7 established in 1987 (Ahmed et al., 1987) and extendedin 1990 (Khakhria et al., 1990) has proven very useful in theepidemiological surveillance of STEC/VTEC O157 (Frost et al.,1993; Saari et al., 2001) infections and is applicable even in low-technology laboratories. In general, phage typing of O157 shouldbe supplemented with one of the molecular typing methods men-tioned below. Because of their discriminatory power, speed, useof commercially available reagents and equipment, and amen-ability to automation and electronic networking, molecular sub-typing methods have become very popular for subtyping of E.coli, particularly strains involved in causing outbreaks of food-borne disease. Molecular subtyping methods for E. coli O157:H7and other foodborne bacterial pathogens were reviewed by Bar-rett (1997). Some recently described methods include macro-restriction endonuclease analysis with PFGE (Preston et al., 2000;Zhang et al., 2000; Swaminathan et al., 2001); detection of in-sertion sequences and characterization of virulence genes byDNA probes or PCR (Thompson et al., 1998; Zhang et al., 2000);detection of amplified fragment length polymorphisms (Iyodaet al., 1999); computer identification by rRNA gene restrictionpatterns (Machado et al., 1998); and the analysis of randomlyamplified polymorphic DNA (Hopkins and Hilton, 2001). In1996, a molecular subtyping network in the United States, des-ignated PulseNet, for the electronic comparison of DNA finger-prints generated by macrorestriction endonuclease analysis withPFGE was developed to subtype E. coli O157:H7. PulseNet hasproved an exceptionally valuable tool for detecting outbreaks ofE. coli O157:H7 infection. The development of standardized lab-oratory and data analysis protocols and their successful use inproviding surveillance for E. coli O157:H7 and other foodbornebacterial pathogens was reviewed by Swaminathan et al. (2001).
ENRICHMENT AND ISOLATION PROCEDURES
Many simple agar media can be used for isolation. Media usedfor selective isolation from feces usually contain substances thatpartly or completely inhibit growth of bacteria other than Ente-robacteriaceae (tetrathionate, deoxycholate, bile salts, etc.). Theaddition of Maranil (dodecylbenzolsulfonate) at a concentrationof 0.005% will inhibit swarming of Proteus organisms. For details,see Edwards and Ewing (1972) or Kauffmann (1966) or any cat-alogue from one of the medium-producing companies. At StatensSerum Institut, Copenhagen, a medium developed in the mediadepartment of the institute, bromothymol blue (BTB) agar, isused.1
MAINTENANCE PROCEDURES
E. coli strains can be kept viable for many years in beef extractagar stabs (tightly closed, e.g., by corks soaked in melted paraffinwax) or on Dorset egg medium. Cultures are initially incubatedat 37!C followed by storage in the dark at room temperature (20–22!C). After a few weeks or months, such cultures often containmany mutational forms such as R forms and acapsular forms;consequently, we prefer to store important cultures in beef brothcontaining 10% glycerol at %80!C. Screw-capped vials are usedfor easy access.
PROCEDURES FOR TESTING SPECIAL CHARACTERS
Kilian and Bulow (1976) found that a very high percentage(97%) of Escherichia coli and the majority (57%) of Shigella strains,exclusively among the Enterobacteriaceae, produce b-glucuronidase(GUD). Prolonged incubation of 28 h increases positivity to99.5% (Rice et al., 1990), which is in accordance with the pres-ence of the uidA (GUD) gene in all E. coli strains (McDaniels etal., 1996). This test (referred to as the GUD-, PGUA-, MUG-,GUR-, or GLUase-HR test) therefore is very suitable as a screen-ing test for E. coli, with the unfortunate exception of most STEC/VTEC O157 strains, which are phenotypically negative. None ofthe other four species of the genus Escherichia are positive forthis enzyme (Rice et al., 1991). Both genotypic and phenotypicassays for glutamate decarboxylase (GAD) used by environmentalscientists have been described as highly specific for E. coli. Un-fortunately, these tests have only recently been shown to exhibitthe same specificity on a smaller collection of pathogenic isolatesof E. coli (Grant et al., 2001).
DIFFERENTIATION OF THE GENUS ESCHERICHIA FROM OTHER
GENERA
See Table BXII.c.193 of the family Enterobacteriaceae for charac-teristics that can be used to differentiate this genus from othergenera of the family.
TAXONOMIC COMMENTS
The identification of Escherichia strains seldom causes problems;however, many studies have shown that “Escherichia is a genus (orspecies) made up of phenotypically variable strains” (Farmer andBrenner, 1977). DNA–DNA hybridization studies have been aninvaluable tool for solving problems in this field. The genus Shi-gella is closely related to Escherichia, and only historical reasonsmake it acceptable that these two genera are not united. Severaltypical EIEC types have been found that have pathogenic traitsthat are similar to those of Shigella. The Sereny test (Sereny,1957), which demonstrates the capacity to cause keratoconjunc-tivitis in the guinea pig, typical of Shigella strains, is also foundin these special Escherichia strains. Day et al. (1981) described atissue culture technique that can be used as a substitute for theSereny test. Typically, such dysentery-associated E. coli strains haveO antigens that are closely related or identical to Shigella O an-tigens. Brenner et al. (1972a), by DNA reassociation studies,found species-level relatedness between Shigella strains and thesespecial Escherichia strains, as well as nonpathogenic E. coli strains.
1. Bromothymol blue agar (selective for Enterobacteriaceae). Combine the followingingredients: peptone (Orthana Ltd., Copenhagen), 10.0 g; NaCl, 5.0 g; yeast extract(Oxoid), 5.0 g; and distilled water, 1000 ml. The pH is adjusted to 8.0, agar powderis added, and the preparation is autoclaved at 120!C for 20 min. The followingcomponents are then added aseptically from sterile stock solutions: Maranil solution[Paste A75 (dodecylben-zolsulfonate), Henkel, Germany], 1.0 ml; sodium thiosul-
fate (50% solution), 2.0 ml; bromothymol blue (Riedel de Haen, Germany; 1.0%solution), 10.0 ml; lactose (33% solution), 27 ml; and glucose (33% solution), 1.2ml. The pH is adjusted to 7.7–7.8. To obtain optimum results, the amount of glucosemust be adjusted for every new batch of yeast extract, peptone, and agar. Thismedium is very useful for differentiation of lactose-fermenting colonies based ontheir color.

GENUS I. ESCHERICHIA 623
Not unexpectedly, many strains are phenotypically intermediatebetween Escherichia and Shigella, but for obvious reasons a specialtaxonomic status for such strains is not warranted. In the olderliterature the name Alkalescens-Dispar is used, but, as stated byBrenner (1978), this group is virtually indistinguishable from E.coli strains and is, in fact, a biogroup of E. coli that is anaerogenic,lactose-negative (or delayed), and nonmotile.
While most or all characters that classically have been usedfor definition of the genus Escherichia are chromosomally deter-mined, several traits that are not characteristic of Escherichia havebeen found in otherwise typical Escherichia strains. Lautrop et al.(1971) and Layne et al. (1971) described H2S-positive strains ofEscherichia, and this character was plasmid-determined. It is notknown which selective forces account for the simultaneous iso-lation of H2S-positive Escherichia strains in different parts of theworld.
Other “forbidden” phenotypic traits have similarly been de-scribed in Escherichia, many of which are plasmid-determined.Ørskov et al. (1961) found many urease-producing strains amongtypical serotypes from piglet diarrhea. Wachsmuth et al. (1979)demonstrated the plasmid-determined nature of a similar urease-positive phenotype in human E. coli strains. Citrate-utilizing E.coli strains were described by Washington and Timm (1976) andwere found to be plasmid determined in similar strains by Satoet al. (1978). Carbon dioxide-dependent cultures can be found(Eykyn and Phillips, 1978). A citrate-positive, malonate-positivebiogroup and a biogroup negative in these reactions were de-scribed (Burgess et al., 1973).
FURTHER READING
Abraham, S.N. and S. Jaiswal. 1997. Type-1 fimbriae of Escherichia coli. InSussman (Editor), Escherichia coli: Mechanisms of Virulence, Cam-bridge University Press, Cambridge. pp. 169–192.
Cowan, S.T., K.J. Steel, G.I. Barrow and R.K.A. Feltham (Editors). 1995.
Cowan and Steel’ s Manual for the Identification of Medical Bacteria,University of Cambridge, Cambridge.
de Graaf, F.K. and W. Gaastra. 1997. Fimbriae of enterotoxigenic Esch-erichia coli. In Sussman (Editor), Escherichia coli: Mechanisms of Vir-ulence, Cambridge University Press, Cambridge. pp. 193–211.
Dodson, K.W., F. Jacob-Dubuisson, R.T. Striker and S.J. Hultgren. 1997.Assembly of adhesive virulence-associated pili in Gram-negative bac-teria. In Sussman (Editor), Escherichia coli: Mechanisms of Virulence,Cambridge University Press, Cambridge. pp. 213–236.
Duffy, G., P. Garvey and D.A. McDowell (Editors). 2001. Verocytotoxi-genic E. coli, Food & Nutrition Press, Inc., Conneticut. pp. 1–457.
Ewing, W.H. 1986. The Genus Escherichia. In Edwards and Ewing (Edi-tors), Edwards and Ewing’ s Identification of Enterobacteriaceae, 4th Ed.,Elsevier Science Publishing Co., New York. pp. 93–122.
Gyles, C.L. (Editor). 1994. Escherichia coli in Domestic Animals and Hu-mans, CAB International, Wallingford.
Jann, K. and B. Jann (Editors). 1990. Bacterial Adhesins. Current Topicsin Microbiology and Immunology, Springer-Verlag, Berlin. 151 pp.
Kaper, J.B. and A.D. O’Brien (Editors). 1998. Escherichia coli O157:H7and other Shiga Toxin-Producing Strains, ASM Press, Washington,D.C. 465 pp.
Klemm, P. (Editor). 1994. Fimbriae: Adhesins, Genetics, Biogenesis, andVaccines, CRC Press, Boca Raton.
Low, D., B. Braaten and M. van der Woude. 1996. Fimbriae. In Neidhardt,Curtiss, Ingraham, Lin, Low, Magasanik, Reznikoff, Riley, Schaechterand Umbarger (Editors), Escherichia coli and Salmonella: Cellular andMolecular Biology, 2nd Ed., ASM Press, Washington, D.C. pp. 146–157.
Nataro, J.P. and J.B. Kaper. 1998. Diarrheagenic Escherichia coli. Clin.Microbiol. Rev. 11 : 142–201.
Neidhardt, F.C., R. Curtiss, III, J.L. Ingraham, E.C.C. Lin, K.B. Low, B.Magasanik, W.S. Reznikoff, M. Riley, M. Schaechter and H.E. Um-barger (Editors), 1996. Escherichia coli and Salmonella: Cellular andMolecular Biology, 2nd Ed., ASM Press, Washington, D.C.
Sussman, M. (Editor). 1997. Escherichia coli: Mechanisms of Virulence,Press syndicate of the University of Cambridge, Cambridge. 639 pp.
DIFFERENTIATION OF THE SPECIES OF THE GENUS ESCHERICHIA
Characteristics useful in distinguishing the five species of Esch-erichia are given in Table BXII.c.193 of the family Enterobacteriaceaeand in Table BXII.c.197 of the genus Escherichia.
List of species of the genus Escherichia
1. Escherichia coli (Migula 1895) Castellani and Chalmers1919, 941AL (Bacillus coli Migula 1895, 27.)co!li. Gr. n. colon large intestine, colon; M.L. gen. n. coli ofthe colon.
The characteristics are as described for the genus andas listed in Table BXII.c.193 of the family Enterobacteriaceae.Occurs naturally in the lower part of the intestine of warm-blooded animals, and as intestinal (some foodborne) andextraintestinal pathogens of humans and animals.
The mol% G # C of the DNA is: 48.5–52.1 (Tm).Type strain: ATCC 11775, CCM 5172, CIP 54.8, DSM
30083, IAM 12119, NCDO 1989, NCTC 9001. SerotypeO1:K1(L1):H7.
GenBank accession number (16S rRNA): X80725.Additional Remarks: Other sequences are listed in Table
BXII.c.206.
2. Escherichia blattae Burgess, McDermott and Whiting 1973,4AL
blat!tae. L. fem. n. blatta cockroach; L. gen. n. blattae of thecockroach.
The characteristics are as described for the genus andas listed in Table BXII.c.193 of the family Enterobacteriaceae.E. blattae was isolated from the hindgut of healthy cock-roaches, Blatta orientalis, in England (Burgess et al., 1973)and on Easter Island (Nogrady and Aubert, personal com-

FAMILY I. ENTEROBACTERIACEAE624
TABLE BXII.c.206. rrn operon sequences of Escherichia strains
Source and straina, b EMBLc Methodd
Escherichia coli:J01859 rRNAJ01695 rrnBV00348 rrnB
(PK3) X80731 PCR(MC4100) X80732 PCRCIP (ATCC 11775T) X80725 PCRATCC 25922 X80724 PCR(K-12) M87049 rrnA(K-12) U00006 rrnB(K-12) L10328 rrnC(K-12) U18997 rrnD(K-12) U00006 rrnE
M29364 rrnG(K–12) D15061 rrnHBioM X80733 PCR(PK3) X80721 rrnA(PK3) X80722 rrnB(PK3) X80723 rrnC(PK3) X80727 rrnD(PK3) X80728 rrnE(PK3) X80729 rrnG(PK3) X80730 rrnH
Escherichia fergusonii:ATCC 35469 AF530475 NAe
Escherichia hermannii:BioM X80675 rRNA
Escherichia vulneris:CIP (ATCC 33821T) X80734 PCR
aSome strain numbers have been lost; sequences were most probably obtained usingE. coli K-12.bBacterial collection from which each strain is deposited: ATCC (American TypeCulture Collection); BioM (BioMerieux, Marcy l’Etoile, France), CIP (Collectionde l’ Institut Pasteur).cAccession numbers under which sequence is available.dMethod by which each sequence has been obtained: rRNA (total rRNA sequencedusing reverse transcriptase), PCR (total PCR products sequenced using T7-DNApolymerase); rrnX (sequence of a single operon, X).eNot available.
munication). It appears as two biotypes, one of which iscitrate and malonate positive, the other negative, and it isthe only species within Escherichia that is gluconate positive(Burgess et al., 1973). E. blattae has not been associated withdisease either in humans or in cockroaches.
The mol% G # C of the DNA is: not determined.Type strain: ATCC 29907, CDC 9005-74, DSM 4481,
NCTC 12127.GenBank accession number (16S rRNA): X87025.
3. Escherichia fergusonii Farmer, Fanning, Davis, O’Hara,Riddle, Hickman-Brenner, Asbury, Lowery and Brenner1985c, 223VP (Effective publication: Farmer, Fanning, Davis,O’Hara, Riddle, Hickman-Brenner, Asbury, Lowery andBrenner 1985b, 77.)*fer.gu.so!ni.i. M.L. masc. (substantive) fergusonii coined tohonor the American microbiologist William W. Ferguson,who made many contributions to enteric bacteriology andwas one of the first to show the role of certain strains of E.coli in infantile diarrhea (Farmer et al., 1985b).
The characteristics are as described for the genus andas listed in Table BXII.c.193 of the family Enterobacteriaceae.
*Editorial Note: This species was formerly known as Enteric Group 10.
Has been isolated from human clinical specimens (stool,urine, blood, and an abdominal wound), the feces of captiveraptors belonging to the order Falconiformes or Strigiformes(Bangert et al., 1988), and from unspecified sites for otherwarm-blooded animals.
The mol% G # C of the DNA is: not determined.Type strain: ATCC 35469, CDC 0568-73.Additional Remarks: Other sequences are listed in Table
BXII.c.206.
4. Escherichia hermannii Brenner, Davis, Steigerwalt, Riddle,McWhorter, Allen, Farmer, Saitoh and Fanning 1983a,438VP (Effective publication: Brenner, Davis, Steigerwalt,Riddle, McWhorter, Allen, Farmer, Saitoh and Fanning1982a, 705.)*her.man!ni.i. M.L. hermannii of Hermann, named in honorof George J. Hermann, former chief of the Enteric Sectionat the CDC, for his many contributions to enteric bacteri-ology, and Lloyd G. Herman, formerly of the EnvironmentalServices Branch, National Institutes of Health, Bethesda,MD, for his contributions to the study of yellow-pigmentedbacteria (Brenner et al., 1982a).
The characteristics are as described for the genus andas listed in Table BXII.c.193 of the family Enterobacteriaceae.Those that together distinguish it from most other membersof this family include growth in the presence of KCN, fer-mentation of cellobiose, and production of yellow pigment.Has been isolated from human clinical specimens (wounds,sputum, lung, stool, blood, and spinal fluid) and recentlyfrom the sludge of an industrial wastewater treatment plant(Kiernicka et al., 1999). The sludge isolate shows promisefor bioremediation; it grows in and degrades high concen-trations of chlorobenzene.
The mol% G # C of the DNA is: 53–58 (Tm).Type strain: ATCC 33650, CDC 980-72, DSM 4560.Additional Remarks: Other sequences are listed in Table
BXII.c.206.
5. Escherichia vulneris Brenner, McWhorter, Leete Knutsonand Steigerwalt 1983d, 438VP (Effective publication: Bren-ner, McWhorter, Leete Knutson and Steigerwalt 1982b,1137.)*vul.ner!is. L. n. vulnus a wound; L. gen. n. vulneris of awound; Escherichia vulneris the Escherichia of a wound.
The characteristics are as described for the genus andas listed in Table BXII.c.193 of the family Enterobacteriaceae(Brenner et al., 1982b). Has been isolated from humanclinical specimens, primarily wounds, the majority of whichoccurred on the arms or legs, but also blood, throat, spu-tum, vagina, urine, and stool, and other warm-blooded an-imals. The type species was isolated from the intestine of acowbird in Michigan, USA.
The mol% G # C of the DNA is: 58.5–58.7 (Tm).Type strain: ATCC 33821, CDC 875-72, DSM 4564, NIH
580.GenBank accession number (16S rRNA): X80734.Additional Remarks: Other sequences are listed in Table
BXII.c.206.
*Editorial Note: This species was formerly known as Enteric Group 11.
*Editorial Note: This species was formerly known as Enteric Group 1.





























![Bergey's Manual of Systematic Bacteriology [Vol 2 Part A] 2nd ed (Springer, 2005) WW](https://img.pdfslide.net/doc/110x75/613cab999cc893456e1e9a48/bergeys-manual-of-systematic-bacteriology-vol-2-part-a-2nd-ed-springer.jpg)