Embed Size (px)
Citation preview

Preference Hierarchies and the Origin of Geographic Specialization in Host Use in SwallowtailButterfliesAuthor(s): John N. ThompsonSource: Evolution, Vol. 47, No. 5 (Oct., 1993), pp. 1585-1594Published by: Society for the Study of EvolutionStable URL: http://www.jstor.org/stable/2410169 .
Accessed: 23/09/2013 14:31
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Society for the Study of Evolution is collaborating with JSTOR to digitize, preserve and extend access toEvolution.
http://www.jstor.org
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions

Evolution, 47(5), 1993, pp. 1585-1594
PREFERENCE HIERARCHIES AND THE ORIGIN OF GEOGRAPHIC SPECIALIZATION IN HOST USE IN SWALLOWTAIL BUTTERFLIES
JOHN N. THOMPSON
Departments of Botany and Zoology, Washington State University, Pullman, Washington 99164
Abstract. -Four allopatric populations of the widely distributed western anise swallowtail butterfly, Papilio zelicaon, use different plant genera as hosts, but simultaneous choice experiments showed that these populations have diverged only slightly in oviposition preference. Of the four popula- tions-two from southeastern Washington State, one from coastal southwestern Washington State, and one from central California-three use hosts that are not available to any of the others. Although variation for the degree of preference for particular plant species occurred within and among populations, all four populations ranked hosts in the same overall order. Monophagy on a local, low-ranking host outside the range of high-ranking hosts did not necessarily lead to the loss of preference for those high-ranking hosts, thereby indicating that the high-ranking hosts would still be accepted, and in some cases even preferred, if a population encountered them again. Hence, the overall preference hierarchy among P. zelicaon populations appears to be evolutionarily con- servative. Analyses of differences among families within the California population indicated that increased preference for some hosts is inversely correlated, whereas preference for other hosts may be uncorrelated. Positive correlations may also occur but were not observed among the plant species tested. Overall, the results indicate local monophagy on different plant species in P. zelicaon has not involved major reorganizations in the preference hierarchy of ovipositing females, even in populations that may have fed on a low-ranking host for many generations. Instead, small increases in preference for local hosts have occurred within an evolutionarily conservative pref- erence hierarchy.
Key words. -Geographic structure, host selection, host shifts, host specificity, oviposition prefer- ence, Papilio, population divergence, specialization, Umbelliferae.
Received July 17, 1992. Accepted December 14, 1992.
Although many phytophagous insect species feed as larvae on only one plant species or a small group of congeneric species throughout their host ranges, others have long lists of recorded hosts spanning several genera or families. Studies of these broadly oligophagous and polyphagous species have increasingly indicated that many of these apparent generalists are actually collections of populations that use different plant taxa (Fox and Morrow 1981; Scriber et al. 1991; Thomp- son and Pellmyr 1991; Singer et al. 1992). For insects in which only ovipositing females choose the larval hosts, there are two fundamentally dif- ferent ways such geographic specialization could develop. Either populations could evolve genetic differences in how females rank potential hosts, or populations could maintain the same ranking of hosts but differ in the hosts they use only because their most preferred hosts are locally rare or absent. Between these two alternatives, one genetic and the other ecological in origin, is a gradation of genetic and ecological influences de- termining which hosts local insect populations actually use.
Whether most geographic specialization onto
different hosts results from genetic divergence in preference hierarchies or ecological differences in the availability of hosts is unknown. Systematic geographic surveys designed specifically to test for genetic divergence in oviposition preference among populations have been carried out for only a few insects. The available studies, however, show evidence for both genetic and ecological influences on local patterns in host choice. For example, at the one extreme Drosophila tripunc- tata does not differ geographically in its prefer- ence ranking for two hosts (Jaenike 1989a). Sim- ilarly, strains of southern cowpea weevil, Callosobruchus maculatus, associated with dif- ferent leguminous hosts show little differentia- tion in their overall preference ranking, although the strains vary in acceptance of low-ranking hosts in no-choice trials (Wasserman 1986). At the other extreme, the butterfly Euphydryas editha shows significant differences among some pop- ulations in how females rank hosts (Singer 1983; Thomas et al. 1987; Singer et al. 1989, 1992).
When genetic change in preference is posible, it may be constrained in some species by cor- relations in choice among potential hosts, where-
1585
?0 1993 The Society for the Study of Evolution. All rights reserved.
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions

1586 JOHN N. THOMPSON
WASHINGTON
o Leadbetter Wawawai
0 L~~~~~~ittle Q. /Tucannon
OREGON
CAIFONIA
* Sailor Bar
N 0) Kilometers
FIG. 1. The location of the four study populations of Papilio zelicaon in western United States.
as in other species preference and the tendency to accept any one host may evolve independently of the tendency to accept other hosts. Wasserman (1986) and Courtney et al. (1989) have argued that in some species incorporation of a low-rank- ing host may not be possible without incorpo- ration of other, similar hosts. In Drosophila busckii acceptability of two mushroom species is genetically correlated (Courtney and Hard 1990), whereas in E. editha acceptability of at least two hosts, one ancestral and the other novel, is in- dependent (Singer et al. 1992).
An earlier study of the evolutionary genetics of oviposition preference in the Papilio machaon species group of swallowtail butterflies showed that local populations of two species, P. zelicaon and P. oregonius, had completely different pref- erence rankings among potential hosts. In pref- erence trials, each species strongly preferred its
normal host and virtually ignored the other's host(s) (Thompson 1 988a). Interspecific crosses indicated that the differences in preference be- tween these swallowtail species are primarily caused by one or more genes located on the X chromosome, which appear to be modified by one or more genes not on the X chromosome (Thompson 1 988b, 1990). Together these results showed that major genetic reorganization in pref- erence hierarchies had occurred during the evo- lution of these closely related swallowtail species. The initial analysis of preference ranking in these two species, however, was based upon one pop- ulation of each species, and the analysis left open the question of whether these butterflies gener- ally evolve the high degree of preference for their local host that was found in these populations.
In this paper, I examine geographic variation in oviposition preference in P. zelicaon to eval- uate how commonly and quickly swallowtail populations evolve reorganized preference hi- erarchies as they colonize different plant species in different parts of their geographic range. Pap- ilio zelicaon is particularly well suited for such an analysis because it is one of the most broadly distributed butterflies in western North America and has been recorded from at least 40 plant species in the Umbelliferae and Rutaceae (Em- mel and Shields 1978; W. F. Wehling and J. N. Thompson, unpubl. data). Many populations feed on species of Lomatium or Cymopterus, es- pecially in drier habitats in the Intermountain West and California. In wetter habitats, and in coastal regions where Lomatium and Cymopte- rus are absent, other umbellifers such as Angelica spp. are commonly used. in addition, some pop- ulations have colonized introduced hosts, es- pecially Foeniculum vulgare (fennel) and Citrus, in areas in which the native hosts have been eliminated or reduced to small, remnant popu- lations.
I chose four geographically isolated popula- tions for the analysis (fig. 1). The Little Tucannon population in the Blue Mountains of eastern Washington feeds on Lomatium grayi and Cy- mopterus terebinthinus. This is the population that was used in the earlier comparison of ovi- position preference hierarchies in P. zelicaon and P. oregonius (Thompson 1988a,b, 1990). The Wawawai population occurs in the Snake River canyon of southeastern Washington and feeds on Lomatium grayi. The Leadbetter population oc- curs in coastal Washington more than 100 km from the nearest populations of Lomatium and
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions

EVOLUTION OF HOST PREFERENCE IN PAPILIO ZELICAON 1587
Cymopterus and feeds almost exclusively on An- gelica lucida. Finally, the Sailor Bar population occurs east of Sacramento, California and feeds on fennel (Foeniculum vulgare), which is an in- troduced plant that has escaped from cultivation and now grows as a common weed. Comparison of these populations allows an assessment of whether a high degree of preference for the local host necessarily evolves in swallowtails either soon after a host shift (i.e., use of Foeniculum) or over longer periods of time (i.e., use of either Angelica, Lomatium, or a combination of Loma- tium and Cymopterus).
Specifically, I asked three questions: (1) Does the Sailor Bar population differ from the Little Tucannon population in how it ranks Lomatium, Cymopterus, and Foeniculum? (2) Does the Leadbetter population differ from the Little Tu- cannon and Wawawai populations in how it ranks Lomatium, Cymopterus, and Angelica? (3) Does preference for each of these host plants evolve independently of preference for the others, or are there positive or negative correlations in pref- erences among plant species?
MATERIALS AND METHODS
Study Areas The Little Tucannon study population occurs
in the Little Tucannon drainage just outside the northern edge of the Wenaha-Tucannon Wil- derness Area in the Blue Mountains of south- eastern Washington (Thompson 1 988a,b). The south-facing slopes below the ridges are steppe, and the north-facing slopes are forests dominated by Pinus ponderosa, Pseudotsuga menziesii, and Abies grandis. The host plants, Lomatium grayi and Cymopterus terebinthinus, are long-lived, perennial umbelliferous herbs that grow in patches along the ridge tops (Thompson and Moody 1985; Thompson 1987). The host plants become dormant by early summer, and the Pap- ilio zelicaon population at this site is therefore restricted to one generation each year.
The Wawawai population occurs near the base of Wawawai Canyon along the Snake River about 50 km northeast of the Little Tucannon popu- lation. The habitat is steppe dominated by the grasses Agropyron spicatum and Poa secunda and a variety of dicots including at least six Loma- tium species, of which L. grayi is the most abun- dant. Only a few small patches of C. terebinthinus occur in the part of the canyon used by this pop- ulation. Hence, unlike at Little Tucannon, C.
terebinthinus is essentially unavailable to ovi- positing females.
The Leadbetter population occurs on the northern tip of Long Beach Peninsula within the Leadbetter Point Unit of the Willapa National Wildlife Refuge (Tiritilli and Thompson 1988). The host plant, Angelica lucida, is a perennial, monocarpic umbellifer that grows along the bay side of the peninsula just inland of the grassy dunes. The vegetation in front of the Angelica lucida zone is dominated by several grass and sedge species, including Triglochin maritima, Juncus lesuerii, Poa marcantha, and Ammophila arenaria. The vegetation behind the Angelica zone is dominated by red alders, Alnus rubra, and Pi- nus contorta. The P. zelicaon population at this site has two, and sometimes more, generations each year (Tiritilli and Thompson 1988).
The Sailor Bar population occurs next to Sailor Bar Park along the American River at the eastern edge of Sacramento, California. The vegetation is dominated by alien species, and Foeniculum vulgare (fennel) forms large stands within which P. zelicaon females lay their eggs. This is one of many P. zelicaon populations in California that have evolved to be multivoltine over the past 150 or so years as they have colonized fennel, which, unlike the former native hosts, is avail- able throughout the summer (Shapiro 1974; Simms 1980).
Methods Individuals were collected from each popula-
tion and taken to the laboratory, where adult females were placed in a plastic shoe box under an incandescent light and allowed to oviposit on sprigs of host plants. Larvae were reared initially on fennel sprigs in plastic petri dishes and some families (= offspring of one female) were later transferred to potted fennel plants in cages. [Lar- val food does not affect adult oviposition pref- erence in the P. machaon complex (Wiklund 1974; Thompson, unpubl. data).] Pupae were placed in a cold room set at OL:24D and 5?C, where they were kept until later in the summer or the following spring.
Upon removal from the cold room, pupae were placed in individual cylindrical cages, 150 mm in diameter and 150 mm in height, made of win- dow screen and the top and bottom of a petri dish, and kept in a walk-in growth chamber free of plants. Adults were hand paired on the day after eclosion and then flown in individual cages in a greenhouse free of all plants other than the
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions

1588 JOHN N. THOMPSON
test plants. Trials were run daily from May until July each year. Daytime temperatures in the greenhouse averaged approximately 30?C and nighttime temperatures approximately 20?C. Temperatures in the flight cages were generally about 3?C cooler at midday than outside the cag- es.
The flight cages were the same as those used in earlier experiments (Thompson 1988a,b, 1990) and were a modification of the design developed by Wiklund (1981, 1982). The cages were 1 m3 and made of 1 .3-cm diameter polyvinyl chloride (PVC) pipe surrounded by white nylon chiffon. A maximum of 20 cages could be set up in the greenhouse. Four plant sprigs, each approxi- mately equal in mass and 10 cm in length, were placed in each cage. A sprig was placed 20 cm from each top corner, so that the top of the sprig was approximately 5 cm below the top of the cage. The petioles or stems of each sprig were placed in a florist's tube to maintain leaf turgor, and the tube was held in place in the cage with a length of PVC pipe.
Females were allowed to oviposit freely on the sprigs throughout each day. For comparisons be- tween Little Tucannon and Sailor Bar, the four sprigs in each cage were Lomatium grayi, Cy- mopterus terebinthinus, Foeniculum vulgare, and Artemisia dracunculus. Artemisia dracunculus (Compositae) is the host of Papilio oregonius and is generally ignored by P. zelicaon females when offered in oviposition trials, although there is some variation among P. zelicaon families in the tendency of females to lay a few eggs on this plant in simultaneous choice trials (Thompson 1 988a). For comparisons among Little Tucannon, Wa- wawai, and Leadbetter, Angelica lucida was sub- stituted for Foeniculum. (For ease of reading, the hosts hereafter will be referred to by their genus name.)
The Lomatium and Cymopterus plants used in the trials came from the Little Tucannon study area. The plants used each day came either from plants collected directly from the study area or from plants germinated from seed collected at the study area and grown in a large experimental garden on campus. The Foeniculum, Angelica, and Artemisia sprigs all came from plants grown from seed in the experimental garden. The Foe- niculum used was a commercial variety, Park sweet fennel. The Angelica came from the Lead- better study area, and the Artemisia came from near Palouse Falls State Park about 50 km north of the Little Tucannon study area.
Oviposition preference tests were repeated for each female for 4 consecutive d, and the positions of the plant sprigs in the cages were rotated daily. This protocol allowed an analysis of whether plant position within cages affects preference and whether females become more or less host spe- cific as they age. The plant sprigs were replaced with new ones each morning. Females were fed daily with a honey-water solution. Throughout each day they laid eggs singly, flying away from the plants after each egg was laid. Those females laying at least 20 eggs over the 4-d trial were used in the analyses. Neither position nor number of days in the cage had any effect on preference, and the 4-d total was therefore used for each female.
All tests were performed on laboratory-reared offspring of females or larvae collected earlier in the same year or the year before. Collections from each population were made over at least 2 yr. This was done to maximize the diversity of ge- notypes used in establishing within-population variation in oviposition preference, and thereby minimize the chance that an underestimate of within-population variation would lead to the erroneous conclusion that populations differed. This precaution proved unnecessary, because oviposition preference within populations did not differ among years. Consequently, years were combined for the analyses of differences between populations.
Statistical Analyses The analyses were based upon a total of 348
females, including 41 families with two or more females (mean, 7.8 females per family). Prefer- ence ranking was determined as the percentage of eggs a female laid on each of the four plant species. As in earlier experiments (Thompson 1988a,b), differences between populations and families within populations were determined us- ing multivariate analysis of variance (MANO- VA) on the percentage of eggs laid on each plant species. Percentage data were transformed using an angular transformation. Four test statistics were used in the interpretation of the MANO- VAs (Hotelling-Lawley Trace, Phillai's Trace, Wilk's Criterion, and Roy's Greatest Root). Uni- variate ANOVAs were used to provide an in- dication of which plant species were responsible for the significant differences found in the MAN- OVAs. Null hypotheses were rejected for all tests at P < 0.05. All analyses were performed using SAS. Each MANOVA testing for differences be- tween populations was performed three ways,
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions
EVOLUTION OF HOST PREFERENCE IN PAPILIO ZELICAON 1589
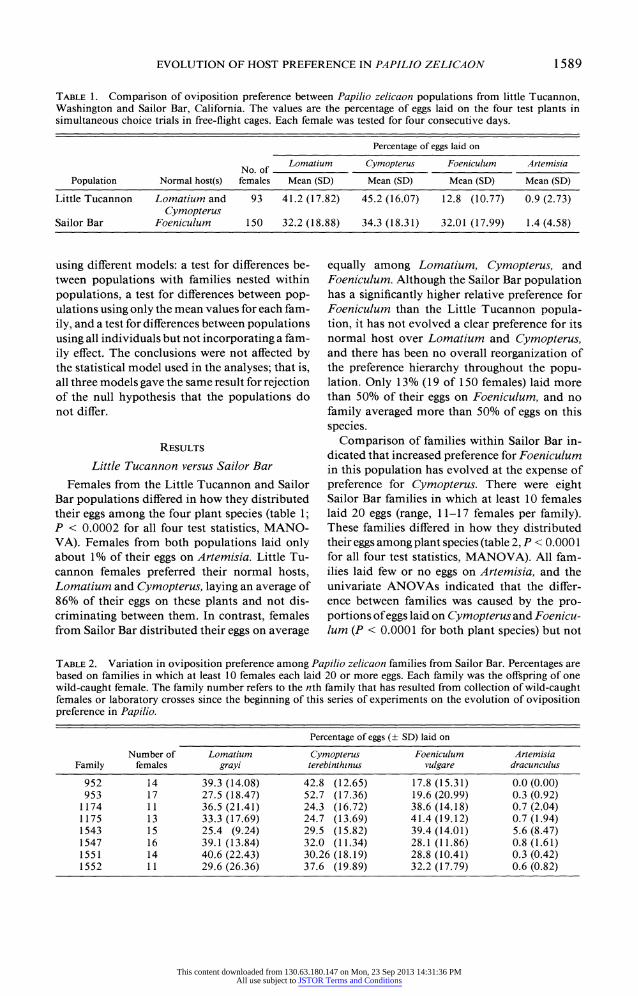
TABLE 1. Comparison of oviposition preference between Papilio zelicaon populations from little Tucannon, Washington and Sailor Bar, California. The values are the percentage of eggs laid on the four test plants in simultaneous choice trials in free-flight cages. Each female was tested for four consecutive days.
Percentage of eggs laid on
No. of Lomatium Cymopterus Foeniculum Artemisia
Population Normal host(s) females Mean (SD) Mean (SD) Mean (SD) Mean (SD)
Little Tucannon Lomatium and 93 41.2 (17.82) 45.2 (16.07) 12.8 (10.77) 0.9 (2.73) Cymopterus
Sailor Bar Foeniculum 150 32.2 (18.88) 34.3 (18.31) 32.01 (17.99) 1.4 (4.58)
using different models: a test for differences be- tween populations with families nested within populations, a test for differences between pop- ulations using only the mean values for each fam- ily, and a test for differences between populations using all individuals but not incorporating a fam- ily effect. The conclusions were not affected by the statistical model used in the analyses; that is, all three models gave the same result for rejection of the null hypothesis that the populations do not differ.
RESULTS
Little Tucannon versus Sailor Bar Females from the Little Tucannon and Sailor
Bar populations differed in how they distributed their eggs among the four plant species (table 1; P < 0.0002 for all four test statistics, MANO- VA). Females from both populations laid only about 1% of their eggs on Artemisia. Little Tu- cannon females preferred their normal hosts, Lomatium and Cymopterus, laying an average of 86% of their eggs on these plants and not dis- criminating between them. In contrast, females from Sailor Bar distributed their eggs on average
equally among Lomatium, Cymopterus, and Foeniculum. Although the Sailor Bar population has a significantly higher relative preference for Foeniculum than the Little Tucannon popula- tion, it has not evolved a clear preference for its normal host over Lomatium and Cymopterus, and there has been no overall reorganization of the preference hierarchy throughout the popu- lation. Only 13% (19 of 150 females) laid more than 50% of their eggs on Foeniculum, and no family averaged more than 50% of eggs on this species.
Comparison of families within Sailor Bar in- dicated that increased preference for Foeniculum in this population has evolved at the expense of preference for Cymopterus. There were eight Sailor Bar families in which at least 10 females laid 20 eggs (range, 11-17 females per family). These families differed in how they distributed their eggs among plant species (table 2, P < 0.0001 for all four test statistics, MANOVA). All fam- ilies laid few or no eggs on Artemisia, and the univariate ANOVAs indicated that the differ- ence between families was caused by the pro- pottions of eggs laid on Cymopterus and Foenicu- lum (P < 0.0001 for both plant species) but not
TABLE 2. Variation in oviposition preference among Papilio zelicaon families from Sailor Bar. Percentages are based on families in which at least 10 females each laid 20 or more eggs. Each family was the offspring of one wild-caught female. The family number refers to the nth family that has resulted from collection of wild-caught females or laboratory crosses since the beginning of this series of experiments on the evolution of oviposition preference in Papilio.
Percentage of eggs (? SD) laid on
Number of Lomatium Cymopterus Foeniculum Artemisia Family females grayi terebinthinus vulgare dracunculus
952 14 39.3 (14.08) 42.8 (12.65) 17.8 (15.31) 0.0 (0.00) 953 17 27.5 (18.47) 52.7 (17.36) 19.6 (20.99) 0.3 (0.92)
1174 11 36.5 (21.41) 24.3 (16.72) 38.6 (14.18) 0.7 (2.04) 1175 13 33.3 (17.69) 24.7 (13.69) 41.4 (19.12) 0.7 (1.94) 1543 15 25.4 (9.24) 29.5 (15.82) 39.4 (14.01) 5.6 (8.47) 1547 16 39.1 (13.84) 32.0 (11.34) 28.1 (11.86) 0.8 (1.61) 1551 14 40.6 (22.43) 30.26 (18.19) 28.8 (10.41) 0.3 (0.42) 1552 11 29.6 (26.36) 37.6 (19.89) 32.2 (17.79) 0.6 (0.82)
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions
1590 JOHN N. THOMPSON
50.
40 *
20~~~~~~~~~~ . 30 i_ J
E~ 20
10 r2= 0.06, P- 0.55
0- 0 1 0 20 30 40 50
50
o Un (n 40 CY)
0) X 30. oQL
20 BeE 20 C t)
10E r2= 0.74, P = 0.006
0. 0 1 0 20 30 40 50
50
40
U)
E 30
' 20
10 r2= 0.24, P= 0.24
0 -
0 1 0 20 30 40 50
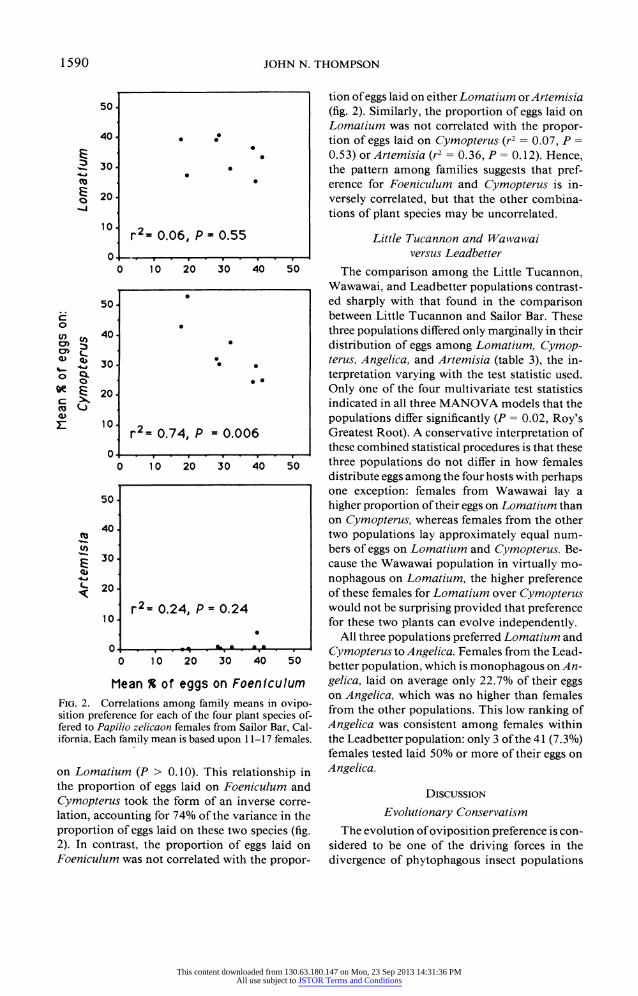
Mean % of eggs on Foeniculum FIG. 2. Correlations among family means in ovipo- sition preference for each of the four plant species of- fered to Papilio zelicaon females from Sailor Bar, Cal- ifornia. Each family mean is based upon 1 1-17 females.
on Lomatium (P > 0.10). This relationship in the proportion of eggs laid on Foeniculum and Cymopterus took the form of an inverse corre- lation, accounting for 74% of the variance in the proportion of eggs laid on these two species (fig. 2). In contrast, the proportion of eggs laid on Foeniculum was not correlated with the propor-
tion of eggs laid on either Lomatium orArtemisia (fig. 2). Similarly, the proportion of eggs laid on Lomatium was not correlated with the propor- tion of eggs laid on Cymopterus (r2 = 0.07, P = 0.53) orArtemisia (r2 = 0.36, P = 0.12). Hence, the pattern among families suggests that pref- erence for Foeniculum and Cymopterus is in- versely correlated, but that the other combina- tions of plant species may be uncorrelated.
Little Tucannon and Wawawai versus Leadbetter
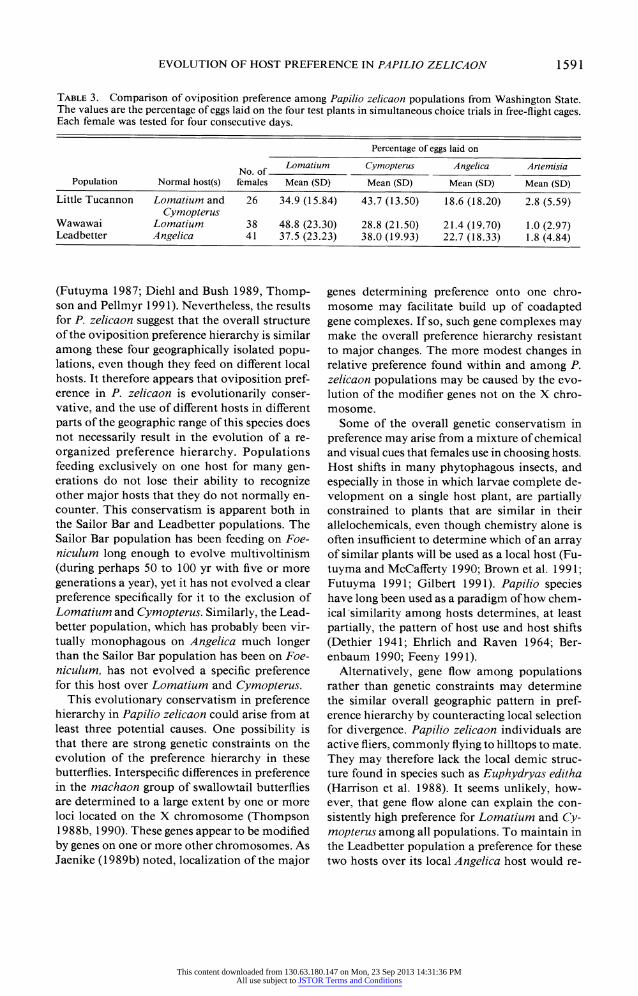
The comparison among the Little Tucannon, Wawawai, and Leadbetter populations contrast- ed sharply with that found in the comparison between Little Tucannon and Sailor Bar. These three populations differed only marginally in their distribution of eggs among Lomatium, Cymop- terus, Angelica, and Artemisia (table 3), the in- terpretation varying with the test statistic used. Only one of the four multivariate test statistics indicated in all three MANOVA models that the populations differ significantly (P = 0.02, Roy's Greatest Root). A conservative interpretation of these combined statistical procedures is that these three populations do not differ in how females distribute eggs among the four hosts with perhaps one exception: females from Wawawai lay a higher proportion of their eggs on Lomatium than on Cymopterus, whereas females from the other two populations lay approximately equal num- bers of eggs on Lomatium and Cymopterus. Be- cause the Wawawai population in virtually mo- nophagous on Lomatium, the higher preference of these females for Lomatium over Cymopterus would not be surprising provided that preference for these two plants can evolve independently.
All three populations preferred Lomatium and Cymopterus to Angelica. Females from the Lead- better population, which is monophagous on An- gelica, laid on average only 22.7% of their eggs on Angelica, which was no higher than females from the other populations. This low ranking of Angelica was consistent among females within the Leadbetter population: only 3 of the 41 (7.3%) females tested laid 50% or more of their eggs on Angelica.
DISCUSSION
Evolutionary Conservatism The evolution of oviposition preference is con-
sidered to be one of the driving forces in the divergence of phytophagous insect populations
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions
EVOLUTION OF HOST PREFERENCE IN PAPILIO ZELICAON 1591
TABLE 3. Comparison of oviposition preference among Papilio zelicaon populations from Washington State. The values are the percentage of eggs laid on the four test plants in simultaneous choice trials in free-flight cages. Each female was tested for four consecutive days.
Percentage of eggs laid on
No. of Lomatium Cymopterus Angelica Artemisia
Population Normal host(s) females Mean (SD) Mean (SD) Mean (SD) Mean (SD)
Little Tucannon Lomatium and 26 34.9 (15.84) 43.7 (13.50) 18.6 (18.20) 2.8 (5.59) Cymopterus
Wawawai Lomatium 38 48.8 (23.30) 28.8 (21.50) 21.4 (19.70) 1.0 (2.97) Leadbetter Angelica 41 37.5 (23.23) 38.0 (19.93) 22.7 (18.33) 1.8 (4.84)
(Futuyma 1987; Diehl and Bush 1989, Thomp- son and Pellmyr 1991). Nevertheless, the results for P. zelicaon suggest that the overall structure of the oviposition preference hierarchy is similar among these four geographically isolated popu- lations, even though they feed on different local hosts. It therefore appears that oviposition pref- erence in P. zelicaon is evolutionarily conser- vative, and the use of different hosts in different parts of the geographic range of this species does not necessarily result in the evolution of a re- organized preference hierarchy. Populations feeding exclusively on one host for many gen- erations do not lose their ability to recognize other major hosts that they do not normally en- counter. This conservatism is apparent both in the Sailor Bar and Leadbetter populations. The Sailor Bar population has been feeding on Foe- niculum long enough to evolve multivoltinism (during perhaps 50 to 100 yr with five or more generations a year), yet it has not evolved a clear preference specifically for it to the exclusion of Lomatium and Cymopterus. Similarly, the Lead- better population, which has probably been vir- tually monophagous on Angelica much longer than the Sailor Bar population has been on Foe- niculum, has not evolved a specific preference for this host over Lomatium and Cymopterus.
This evolutionary conservatism in preference hierarchy in Papilio zelicaon could arise from at least three potential causes. One possibility is that there are strong genetic constraints on the evolution of the preference hierarchy in these butterflies. Interspecific differences in preference in the machaon group of swallowtail butterflies are determined to a large extent by one or more loci located on the X chromosome (Thompson 1 988b, 1990). These genes appear to be modified by genes on one or more other chromosomes. As Jaenike (1 989b) noted, localization of the major
genes determining preference onto one chro- mosome may facilitate build up of coadapted gene complexes. If so, such gene complexes may make the overall preference hierarchy resistant to major changes. The more modest changes in relative preference found within and among P. zelicaon populations may be caused by the evo- lution of the modifier genes not on the X chro- mosome.
Some of the overall genetic conservatism in preference may arise from a mixture of chemical and visual cues that females use in choosing hosts. Host shifts in many phytophagous insects, and especially in those in which larvae complete de- velopment on a single host plant, are partially constrained to plants that are similar in their allelochemicals, even though chemistry alone is often insufficient to determine which of an array of similar plants will be used as a local host (Fu- tuyma and McCafferty 1990; Brown et al. 1991; Futuyma 1991; Gilbert 1991). Papilio species have long been used as a paradigm of how chem- ical similarity among hosts determines, at least partially, the pattern of host use and host shifts (Dethier 1941; Ehrlich and Raven 1964; Ber- enbaum 1990; Feeny 1991).
Alternatively, gene flow among populations rather than genetic constraints may determine the similar overall geographic pattern in pref- erence hierarchy by counteracting local selection for divergence. Papilio zelicaon individuals are active fliers, commonly flying to hilltops to mate. They may therefore lack the local demic struc- ture found in species such as Euphydryas editha (Harrison et al. 1988). It seems unlikely, how- ever, that gene flow alone can explain the con- sistently high preference for Lomatium and Cy- mopterus among all populations. To maintain in the Leadbetter population a preference for these two hosts over its local Angelica host would re-
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions

1592 JOHN N. THOMPSON
quire sustained introgression from Lomatium/ Cymopterus-feeding populations hundreds of ki- lometers away.
Yet a third possibility is that in the absence of other potential hosts, there is little selection to develop strong preferences for Foeniculum in the Sailor Bar population or Angelica in the Lead- better population. This possible explanation, however, explains only half the problem in the conservatism of preference in this species. It ex- plains why a strong preference for the local host has not evolved at Sailor Bar and Leadbetter, but it does not explain why all the populations rank Lomatium and Cymopterus highly. It could be that all populations throughout western North America were initiated by founders from pop- ulations that fed on Lomatium and Cymopterus and that they still retain their preferences. If true, this is another way of arguing that preference hierarchies are evolutionarily conservative. It suggests that specialization on other plants out- side the range of these two highly preferred taxa will not necessarily lead to the loss of preference for these species if they are encountered again sometime in their evolutionary future. That is, oviposition preference among the suite of poten- tial hosts is not like adaptation for, say, sight, in which vision is lost in species like cave fish in environments in which it is not maintained by natural selection. Once acquired, a high prefer- ence for Lomatium and Cymopterus is not easily lost. If instead, a low-ranking species such as Angelica was the ancestral host, it would again imply evolutionary conservatism in the prefer- ence hierarchy, because it would suggest that Leadbetter and similar ancestral populations al- ready had high innate preference for Lomatium and Cymopterus even before descendent popu- lations ever encountered these hosts.
More detailed analyses of the genetics of pref- erence, phylogeography of P. zelicaon popula- tions, and patterns of gene flow among them will be needed to sort out these possible explanations for the evolutionary conservatism in prefrence hierarchy found among these populations. What is clear, however, from the results is that major reorganizations of the preference hierarchy do not necessarily occur among populations of this species as they specialize on different host plants in different parts of their geographic range.
Conservatism versus Flexibility Although the preference hierarchy in P. zeli-
caon is conservative, it is not evolutionarily rig-
id. Within the overall preference hierarchy, mod- erate evolutionary changes are possible, elevating to a limited extent the preference for a local host. All comparisons among and within populations indicated that such moderate shifts can occur: the higher relative preference for Foeniculum in the Sailor Bar population as compared to the Little Tucannon population, the potentially higher relative preference for Lomatium in Wa- wawai as compared with Little Tucannon and Leadbetter, and the variation in relative prefer- ence for Cymopterus and Foeniculum found among families within Sailor Bar all suggest a moderate level of evolutionary flexibility in the preference hierarchy. Moreover, an earlier study of the Little Tucannon population showed that families in this population differ in their tenden- cy to lay a few eggs on the very low ranking Artemisia (Thompson 1 988a). Hence, some vari- ation in relative preference has been found among families or populations for at least four of the five plant species that have been so far tested in these studies of the evolution of oviposition pref- erence in Papilio.
The inverse correlation in preference for Foe- niculum and Cymopterus found among families at Sailor Bar is particularly intriguing, because it indicates that increases in preference for any one species may sometimes come at the expense of preference for one other particular species rather than as a proportionate decrease in relative pref- erence for all other plant species. Hence, the pref- erence hierarchy in P. zelicaon may be con- structed from a combination of two kinds of relationship among plant species; (1) those in which plants are inversely correlated with an- other, and (2) those in which plants are uncor- related with one another. At least among the plants tested in this study, there were no positive correlations among host plant species. But if yet more species were tested, some positive corre- lations among some species could possibly occur, especially among plants similar in chemistry, morphology, and life history.
The special inverse relationship between Foeniculum and Cymopterus would have been masked if the analysis of the increase in Foenicu- lum use at Sailor Bar had not been structured by family. A simpler experimental design placing all 150 Sailor Bar females individually into the correlations would have suggested that Loma- tium, Cymopterus, and Foeniculum are all in- versely correlated, although only weakly with each accounting for 220/o-27% of the variance. Ana-
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions

EVOLUTION OF HOST PREFERENCE IN PAPILIO ZELICAON 1593
lyzed in this way, however, the inverse correla- tions become an artifact of the use of proportions in the correlations and the fact that some indi- viduals laid up to 90% of their eggs on one plant species, thereby forcing an inverse correlation among plant species. Only by partitioning the population into families was it possible to par- tition the variance sufficiently to see how the increase in preference for Foeniculum has come about. There are so far no other comparable stud- ies of other insect species-involving both ovi- position among more than two plant species and analysis partitioned by family-to know if a combination of negatively correlated and uncor- related preferences is a part of the preference hierarchy of other insects.
Preference Hierarchies in Other Insects and the Experimental Evaluation of Preference The oviposition preference in P. zelicaon, then,
appears to be neither fully constrained nor readi- ly open to wholesale reorganization. This view of moderate evolutionary lability in preference within P. zelicaon appears to be intermediate between that found in Drosophila tripunctata or Callosobruchus maculatus, which show little variation in preference ranking among popula- tions or strains, and that found in Euphydryas editha, which shows considerable variation (Jae- nike and Grimaldi 1983; Wasserman 1986; Jae- nike 1989a; Singer et al. 1991). There are, how- ever, too many differences in the experimental designs used to study these different insect spe- cies to allow direct comparisons among the re- sults. Jaenike's analyses of preference in D. tri- punctata have been based upon simultaneous choice trials between pairs of hosts, and are therefore the most similar in experimental design to this study; Wasserman's analysis of C. ma- culatus was based upon no-choice trials; and the studies of E. editha by Singer and his colleagues have used sequential choice trials in which plants are presented one at a time rather than simul- taneously to individual females. These differ- ences in experimental protocols caution against direct comparisons until the different designs have all been compared on one insect species.
The results for these four P. zelicaon popula- tions provide a working hypothesis for the rel- ative degrees of constraint and evolutionary flex- ibility in preference within P. zelicaon and related Papilio species. Evaluating the hypothesis more fully will now require a broader survey of pref- erence among P. zelicaon populations to deter-
mine if any populations have evolved a complete reorganization in the preference hierarchy, and a more extensive survey of host species to eval- uate the extent to which natural selection can act independently on preference for particular plant species.
ACKNOWLEDGMENTS I thank R. Dowell for help in collecting but-
terflies at Sailor Bar; W. Wehling for help in collecting at Little Tucannon and Leadbetter; D. Althoff, H. Hansel, A. Joshi, R. Lynch, L. Patten, L. Valburg, and W. Wehling for help in the pref- erence experiments and larval rearings, and H. Hansel for help with data anslyses. I am grateful to H. Hansel, R. Holt, M. Singer, and C. Thomas for very helpful comments on an earlier draft of the manuscript. This work was supported by Na- tional Science Foundation grants BSR 8705394 and BSR 9019884 and United States Depart- ment of Agriculture grant 88-37153-4468.
LITERATURE CITED
Berenbaum, M. R. 1990. Evolution of specialization in insect-umbellifer associations. Annual Review of Entomology 35:319-343.
Brown, K. S., Jr., J. R. Trigo, R. B. Francini, A.B.B. De de Morais, and P. C. Motta. 1991. Aposematic insects on toxic host plants: coevolution, coloni- zation, and chemical emancipation. Pp. 375-402 in P. W. Price, T. M. Lewinsohn, G. W. Femandes, and W. W. Benson, eds. Plant-animal interactions: Evolutionary ecology in tropical and temperate regions. Wiley, New York.
Courtney, S. P., G. K. Chen, and A. Gardner. 1989. A general model for individual host selection. Oi- kos 55:55-65.
Courtney, S. P., and J. J. Hard. 1990. Host acceptance and life-history traits in Drosophila busckii: Tests of the hierarchy-threshold model. Heredity 64:371- 375.
Dethier, V. G. 1941. Chemical factors determining the choice of food plants by Papilio larvae. Amer- ican Naturalist 75:61-73.
Diehl, S. R., and G. L. Bush. 1989. The role of habitat preference in adaptation and speciation. Pp. 345- 365 in D. Otte and J. Endler, eds. Speciation and its consequences. Sinauer Associates, Sunderland, Mass.
Ehrlich, P. R., and P. H. Raven. 1964. Butterflies and plants: A study in coevolution. Evolution 18:586- 608.
Emmel, J. F., and 0. Shields. 1978. Larval foodplant records for Papilio zelicaon in western United States, and further evidence for the conspecificity of P. zelicaon and P. gothica. Journal of Research on the Lepidoptera 17:56-67.
Feeny, P. 1991. Chemical constraints on the evolu- tion of swallowtail butterflies. Pp. 315-340 in P. W. Price et al., eds. Plant-animal interactions: Evo-
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions

1594 JOHN N. THOMPSON
lutionary ecology in tropical and temperate regions. Wiley, New York.
Fox, L. R., and P. A. Morrow. 1981. Specialization: species property or local phenomenon? Science 211: 887-893.
Futuyma, D. J. 1987. The role of behavior in host- associated divergence in herbivorous insects. Pp. 295-302 in M. D. Huettel, ed. Evolutionary ge- netics of invertebrate behavior. Plenum, New York.
1991. Evolution of host specificity in her- bivorous insects: Genetic, ecological, and phylo- genetic aspects. Pp. 431-454 in P. W. Price et al., eds. Plant-animal interactions: Evolutionary ecol- ogy in tropical and temperate regions. Wiley, New York.
Futuyma, D. J., and S. S. McCafferty. 1990. Phylog- eny and the evolution of host plant associations in the leaf beetle genus Ophraella (Coleoptera, Chrys- omelidae). Evolution 44:1885-1913.
Gilbert, L. E. 1991. Biodiversity of a Central Amer- ican Heliconius community: Pattern, process, and problems. Pp. 403-427 in P. W. Price, et al., eds. Plant-animal interactions: Evolutionary ecology in tropical and temperate regions. Wiley, New York.
Harrison, S., D. D. Murphy, and P. R. Ehrlich. 1988. Distribution of the bay checkerspot butterfly, Eu- phydryas editha bayensis: Evidence for a metapopu- lation model. American Naturalist 132:360-382.
Jaenike, J. 1987. Genetics of oviposition-site pref- erence in Drosophila tripunctata. Heredity 59:363- 369.
1989a. Genetic population structure of Dro- sophila tripunctata: Patterns of variation and co- variation of traits affecting resource use. Evolution 43:1467-1482.
. 1989b. Genetics of butterfly-host associa- tions. Trends in Ecology and Evolution 4:34-35.
Jaenike,J.,andD.Grimaldi. 1983. Geneticvariation for host preference within and among populations of Drosophila tripunctata. Evolution 37:1023-1033.
Scriber, J. M., B. L. Biebink, and D. Snider. 1991. Reciprocal latitudinal clines in oviposition behav- ior of Papilio glaucus and P. canadensis across the Great Lakes hybrid zone: Possible sex-linkage of oviposition preferences. Oecologia 87:360-368.
Scriber, J. M., R. C. Lederhouse, and R. H. Hagen. 1991. Foodplants and evolution within Papilio glaucus and Papilio troilus species groups (Lepi- doptera: Papilionidae). Pp. 341-373 in P. W. Price et al., eds. Plant-animal interactions: Evolutionary ecology in tropical and temperate regions. Wiley, New York.
Shapiro, A. M. 1974. The butterfly fauna of the Sac- ramento Valley, California. Journal of Research on the Lepidoptera 13:73-82, 115-122, 137-148.
Simms, S. R. 1980. Diapause dynamics and host plant suitability of Papilio zelicaon (Lepidoptera: Papilionidae). American Midland Naturalist 103: 375-384.
Singer, M. C. 1983. Determinants of multiple host use by a phytophagous insect population. Evolution 37:389-403.
Singer, M. C., D. Ng, and R. A. Moore. 1991. Genetic variation in oviposition preference between but- terfly populations. Journal of Insect Behavior 4:531- 535.
Singer, M. C., D. Ng, D. Vasco, and C. D. Thomas. 1992. Rapidly evolving associations among ovi- position preferences fail to constrain evolution of insect diet. American Naturalist 139:9-20.
Singer, M. C., C. D. Thomas, H. L. Billington, and C. Parmesan. 1989. Variation among conspecific in- sect populations in the mechanistic basis of insect breadth. Animal Behavior 37:751-759.
Thomas, C. D., D. Ng, M. C. Singer, J.L.B. Mallet, C. Parmesan, and H. L. Billington. 1987. Incorpo- ration of a European weed into the diet of a North American herbivore. Evolution 41:892-901.
Thompson, J. N. 1987. The ontogeny of flowering and sex expression in divergent populations of Lomatium grayi. Oecologia 72:605-611.
-. 1988a. Variation in preference and specificity in monophagous and oligophagous swallowtail but- terflies. Evolution 42:118-128.
1988b. Evolutionary genetics of oviposition preference in swallowtail butterflies. Evolution 42: 1223-1234.
1990. Coevolution and the evolutionary ge- netics of interactions among plants and insects and pathogens. Pp. 249-271 in J. J. Burdon and S. R. Leather, eds. 1990. Pests, pathogens, and plant communities. Blackwell, Oxford, UK.
Thompson, J. N., and M. E. Moody. 1985. Assessing probability of interaction in size-structured popu- lations: Depressaria attack on Lomatium. Ecology 66:1597-1607.
Thompson, J. N., and 0. Pellmyr. 1991. Evolution of oviposition behavior and host preference in Lep- idoptera. Annual Review of Entomology 36:65-89.
Tiritilli, M. E., and J. N. Thompson. 1988. Variation in swallowtail/plant interactions: Host selection and the shapes of survivorship curves. Oikos 53:153- 160.
Wasserman, S. S. 1986. Genetic variation in adap- tation to foodplants among populations of the southern cowpea weevil, Callosobruchus macula- tus: Evolution of oviposition preference. Entomo- logia Experimentalis et Applicata 42:201-212.
Wiklund, C. 1974. Oviposition preferences in Papilio machaon in relation to the host plants of the larvae. Experimentalis et Applicata 17:189-198.
. 1981. Generalist vs. specialist oviposition behaviour in Papilio machaon (Lepidoptera) and functional aspects of the hierarchy of oviposition preferences. Oikos 36:163-170.
1982. Generalist versus specialist utilization of host plants among butterflies. Pp. 181-191 in J. H. Visser and A. K. Minks, eds. Proceedings of the Fifth International Symposium of Insect-Plant Re- lationships, PUDOC, Wageningen.
Corresponding Editor: R. Holt
This content downloaded from 130.63.180.147 on Mon, 23 Sep 2013 14:31:36 PMAll use subject to JSTOR Terms and Conditions





























