Embed Size (px)
Citation preview

AN ABSTRACT OF THE THESIS OF
FLASH GIBSON(Name of Student)
in Zoology
Title:.
(Major)
for the DOCTOR OF PHILOSOPHY(Degree)
presented on May 6, 1971(Date)
BEHAVIORAL PATTERNS AND THEIR TEMPORAL
ORGANIZATION IN BREEDING AMERICAN AVOCETS
Abstract approved:Redacted for privacy
(,John A. Wiens
The organization of behavioral activities in time is important
to a bird's survival and reproduction. To be successful, either
proximately or ultimately, a bird must apportion its behavior in time
so that it may obtain sufficient energy for maintenance activities and
for such activities as migration and reproduction. The organization
of behavioral activities in time can be expressed as a time budget.
I have described the action patterns and displays of breeding American
Avocets (Recurvirostra americana) and have examined the budgeting
of time among these behaviors and its possible adaptiveness. Particu-
lar consideration is given to patterns of seasonal, diurnal and sexual
variations.
The study area was located within an alkaline marsh at Summer
Lake, Oregon. The time budget data were collected by recording at
10 sec. intervals the amount of time spent in 12 categories of behavior.

During the 1969 breeding season (April-July) 422 hrs. of data were
taken on 36 individuals.
The breeding season was divided into four stages ; prenesting,
incubation, parental care and post breeding. Seasonally, all main-
tenance activities combined took 67% of the avocet's time, and repro-
ductive activities took 26%. Of all activities, feeding took the largest
portion of time (36%). Feeding, because of its close relationship to
bioenergetics, is probably one of the most important aspects of the
time budget. Through the breeding season energy requirements
change, and this is reflected in a seasonal flux in time spent feeding.
I suggest that seasonal variation in temperature, activity levels, prey
abundance, and such energetically expensive processes as egg produc-
tion, molting and premigratory fat deposition may determine the
amount of time spent foraging. There was little seasonal fluctuation
in the amount of time spent in resting, preening, or in aggressive
activities. Incubation and caring for the young required similar
amounts of time.
Diurnally, several categories of behavior, such as feeding,
preening and nest building, exhibited morning and afternoon peaks
of activity, separated by a period of inactivity at midmorning.
This pattern held when all "active" action patterns were combined.
I concluded that foraging patterns largely dictate the form of the
diurnal activity pattern. The morning feeding peak may be related

to a need to feed after the night-long fast, while the afternoon peak is
perhaps associated with optimum environmental conditions for feeding,
such as maximum insect availability.
Avocet males and females exhibited few significant differences
in either the seasonal or diurnal time budgets. However, males
appeared to be more aggressive while females spent more time incu-
bating. The general similarities of morphology, behavior, and tem-
poral organization of behavior, indicate that the sexes have similar
roles in the social system, and may be ecologically similar as well.

Behavioral Patterns and Their Temporal Organizationin Breeding American Avocets
by
Flash Gibson
A THESIS
submitted to
Oregon State University
in partial fulfillment ofthe requirements for the
degree of
Doctor of Philosophy
June 19 71

APPROVED:
Redacted for privacy
Associate Profe s r of Zoologyin charge of major
Redacted for privacy
Chairman of the Department of Zoology
Redacted for privacy_VDean diura/ duate School
Date thesis is presented , /77/Typed by Opal Grossnicklaus for Flash Gibson

ACKNOWLEDGEMENTS
Many people have made the completion of this project possible.
I would like to give special thanks to the Oregon State Game Commis-
sion, A. B. Claggett, and W. Olsen for supplying housing for me and
my family and for their valuable council. Dr. R. B. Hamilton gener-
ously supplied me with a copy of his thesis.
I would like to acknowledge Dr. S. G. Martin for helping set up
field equipment and for offering much valuable discussion concerning
the methodology used in this study. Several of my colleagues have
been particularly helpful in their discussions of the theoretical aspects
of this study. In this regard my thanks go to D. Kroodsma, J. States,
M. Scott, and T. Haislip.
My major professor Dr. J. A. Wiens was helpful through all
phases of the study and was invaluable in his supervision of the prepa-
ration of this manuscript. I would like to thank D. Neiss for writing
the computer programs used in the analysis of the data. Special
appreciation goes to Lorna States for her fine drawings of avocet
action patterns. Finally my inexpressable thanks go to my wife,
Linda, wh,, did many of the tedious secretarial duties and offered
continual encouragement. A grant from the Oregon State Computer
Center made analysis of the data possible. Funds for this research
were supplied by a N. D. E. A. Predoctoral Fellowship.

TABLE OF CONTENTS
INTRODUCTION
THE STUDY AREA
1
3
The Summer Lake Management Area 3
Distribution of Water 5
The Vegetation 6
PROCEDURES 8
Constructing the Ethogram 8Breeding Biology 9Marking Individuals 10The Time Budget 11Analysis of the Data 14
BREEDING BIOLOGY 16
Migration and Arrival 16Breeding Chronology 18Territory 21The Nest 25Nest Dispersion 27Egg Laying 29Incubation 3 0Hatching 34Care of the Young 36Preparation for Migration 3 7
THE ETHOGRAM 40
Maintenance Behavior 42Mating Behavior 48Aggressive Behavior 52Diver sionary i3ehavior 59Nesting Behavior 6ZParental Behavior 66Miscellaneous Action Patterns 68

THE TIME BUDGET ANALYSIS69
Behavioral Categories 70The Seasonal Time _budget 72Sexual Differences in the Seasonal Time Budget 79The Diurnal Time Budget 84Sexual Differences in the Diurnal Time Budget 91
DISCUSSION95
Energy and Seasonal Changes in Foraging Activity 96Energy and Seasonal Patterns in Non-feeding Activities 105The Bimodal Nature of the Activity Pattern 106Sexual Aspects of the Time Budget 113
BIBLIOGRAPHY 11 7
APPENDICES
Appendix
1 Seasonal time budget for all action patterns 1 21
2 Diurnal time budget for all action patterns 1 23
3 Seasonal time budget with categories ofbehavior, sexes combined 135
4 Seasonal time budget with categories ofbehavior
5 Diurnal time budget with categories ofbehavior, sexes combined
6 Diurnal time budget with categories ofbe
136
137
139
Calculation of Standard Metabolism 1 43

LIST OF FIGURES
Figure Page
1. The Summer Lake Management Area 4
Z. Habitat features of the Summer Lake ManagementArea 7
3. A portion of the standardized data form 12
4. The diurnal period during which time budget datawere taken. 13
5. The breeding and wintering distribution of theAmerican Avocet 1 7
6. Arrival of migrants at Summer Lake 19
7. Breeding chronology for the avocet population atSummer Lake 20
8. Territories of six pairs of avocets during the incuba-tion period 23
9. Avocet nest dispersion in different habitats atSummer Lake 28
10. Time of nest initiation at Summer Lake in 1968 and1969. 31
11. Avocet action patterns: feeding, flock feeding, preen-ing, resting 43
1 2. Avocet action patterns: bathing, scratching, two-wingstretch, wing-leg stretch 47
13. Avocet action patterns: precopulatory breast preening,mount, crossed bills
50
14. Avocet action patterns: crouch, single crouch-run,paired crouch-run, pursuit attack, retreat 54
15. Avocet action patterns: jump attack, wingspread,bowing, squatting, psuedosleeping 56

FigurePage
16. Avocet action patterns: tightrope, sneak-run, breastdipping, flying 61
17. Avocet action patterns: throwing straws, scraping,groundchecking, nest approach, incubation 65
18. Avocet action patterns: upright, parallel walk,brooding 67
19. The seasonal time budget, sexes combined 73
20. The seasonal time budget for both sexes 81
21. The diurnal time budget, sexes combined 86
22. The diurnal time budget for both sexes 92
23. The seasonal temperature regime for SummerLake during 1969 97
24. Generalized relationships between time spent infeeding by avocets and energy demands 98
25. Seasonal pattern of aquatic insect emergence forthe Whistler's Bend Impoundment, Douglas County,Oregon. 103
26. The apportionment of avocet time to an "active" state 107
27. The diurnal apportionment of time by avocets to 7categories of behavior for the entire breeding season 108
28. The diurnal foraging pattern for avocets; the diurnaltemperature regime; the diurnal pattern of emergenceof Tri.coptera 110

LIST OF TABLES
Table
1. The relationship between nest location and nestlining
Page
26
2. Role of the sexes during the incubation period at 4nests during 1969 33
3. Summary of avocet hatching data for 1969 35
4. Number of chicks in broods of two age classesduring 1969 38
5. Summary of avocet behavior patterns
6. Categories of behavior used in the time budgetanalysis
7. Diurnal incubation pattern for each sex
8. Percent total observation time spent in an "active"state for each stage of the breeding cycle
41
71
85
100

BEHAVIORAL PATTERNS AND THEIR TEMPORALORGANIZATION IN BREEDING
AMERICAN AVOCETS
INTRODUCTION
The time budget of a bird, the way it uses time in the perform-
ance of various activities, is important to survival and reproduction.
To be successful (either proximately,or ultimately), a bird must ap-
portion its time so that it has sufficient energy for metabolic functions
and for activities such as migration and reproduction. The arrange-
ment of the time budget must be closely attuned to environmental
conditions; a bird whose time budget is not properly phased in rela-
tion to environmental conditions will be at a selective disadvantage
(Orians, 1961; Verbeek, 1964; Verner, 1965).
There are two basic types of time budget --annual and diurnal.
Verner (1965) suggests that there is a hypothetical "best" annual and
diurnal time budget for a given set of circumstances and that selec-
tion will favor those individuals whose budgets most closely approach
this optimum. For example, a bird that migrates too early or too
late in the spring and consequently lays eggs too early or too late in
relation to local conditions will probably produce fewer offspring
and thus be at a selective disadvantage. Likewise, diurnal patt,-rns
of behavior are disadvantageous if improperly arranged. Aggressive
neglect is an excellent example of this (Ripley, 1961). An overly

2
aggressive male may spend an excessive portion of his time defend-
ing his territory, thus neglecting courtship and care of the young.
Consequently he produces fewer offspring than his neighbors.
Orians (1961) first emphasized the evolutionary importance of
time budgets and various aspects have been further developed by
other workers (e. g., Verbeek, 1964; Verner, 1965; Morton, 1967;
Smith, 1968; Martin, 1971). Diurnal rhythms of behavior have been
studied for many years with little emphasis on the evolutionary impor-
tance of these rhythms (e.g., Lees, 1948; Palmgren, 1949; Kluyver,
1950; Cullen, 1954; Gibb, 1954, 1956).
The present investigation was undertaken to further develop
the concept that the apportionment of time is an important ecological
and behavioral feature with adaptive significance. The American
Avocet (Recurvirostra americana) was chosen for study partly from
my own interest in the bird and partly because it is large and easily
observed. I have analyzed the avocet's use of time through the bread-
ing cycle with emphasis on seasonal, diurnal and sexual variations.

3
THE STUDY AREA
The study area was located within the Summer Lake Manage-
ment Area, a game refuge administered by the Oregon State Game
Commission (Figure 1). The refuge is located in Lake County,
Oregon (T30S, R16E) at an elevation of 1310 m. The topography
of the area is typical of northern portions of the Great Basin. It is
characterized by north-trending fault-block mountains and basins
with no external drainage (Baldwin, 1964). The Summer Lake Valley
is a graben lying between opposing fault-blocks, one of which is
Winter Rim, rising 940 m above the valley floor. The valley con-
tains a large alkaline lake with a marsh located at its north end. The
majority of the study was carried out in this marsh.
The Summer Lake Management Area
The management area covers approximately 7290 ha. The
Oregon Game Commission has constructed a series of canals and
dikes to regulate water flow in the marsh. Control of water levels
is partially dependent on irrigation practices in other portions of the
valley. Therefore, water levels are not always held at ideal levels.
However, the majority of the water supplying the marsh comes from
the artesian-fed Ana River, thus assuring a fairly constant water
supply even in the driest years.

Figure 1. The Summer Lake Management Area, Lake County,Oregon
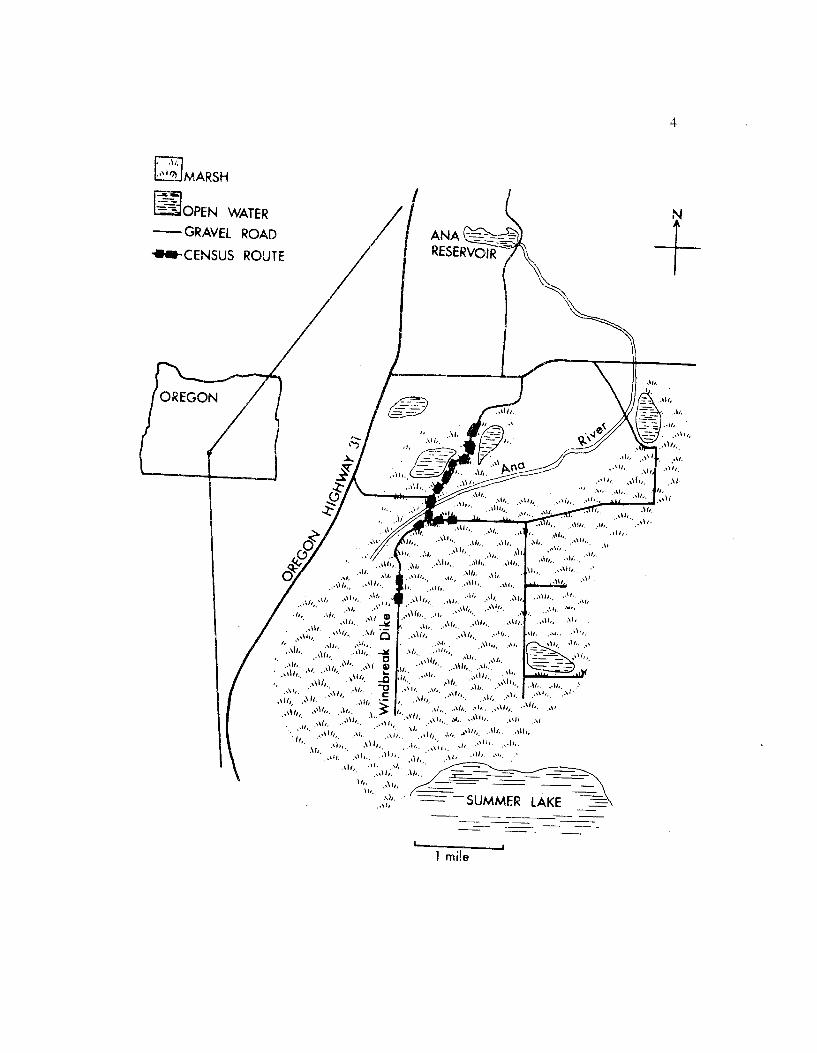
17, MARSH
OPEN WATER
GRAVEL ROAD*-CENSUS ROUTE
ANARESERVOIR
4
.111,
..\.111
".
.\\SUMMER LAKE
I mile

5
Two major dikes traverse the marsh. In the eastern portion
there is Bulgate Dike (4 km long) and in the west, Windbreak Dike
(3. 2 km long) (Figure 1). Both dikes range from 20-50 m in width.
The dikes within the marsh are topped by service roads, making much
of the area accessible by automobile. The majority of this study was
conducted in the Windbreak Dike area.
A large portion of the eastern part of the marsh is characterized
by large alkali flats which are mostly devoid of vegetation. The west-
ern side is extensively vegetated.
Distribution of Water
Water in the marsh is distributed in two ways. In the eastern
and northern portions, water is localized in a patchy distribution of
ponds and canals. In contrast, the region around Windbreak Dike,
extending west to Oregon Highway 31, is extensively covered with
shallow water. The Ana River flows into the area west of Windbreak
Dike; it is also supplied by irrigation run-off from agricultural areas
along the west edge of the valley. These two factors account for the
moister conditions in this area.
Water depths in ponds and waterways throughout most of the
marsh range from 1 to 15 cm. However, some of the larger ponds
in the northern part of the marsh are up to 2 m deep.

6
The Vegetation
The vegetation of the entire marsh is dominated by rushes
(Scirpus sp. ) and salt grass (Distichlis stricta). The eastern half
of the marsh is almost devoid of vegetation except for localized
patches,. The vegetation in the vicinity of Windbreak Dike is char-
acterized by salt grass in dry areas, while three-square tule (Scirpus
americanus) and marsh bullrush (S. robustus) occur in wet soils and
shallow water. Great bullrush (S. validus) and cattail (Typha latifolia)
have a patchy distribution in deeper water. Rabbit brush (Chryso-
thamnus nauseosus) and willows (Salix sp. ) occur sparsely along the
dikes (Figure 2).

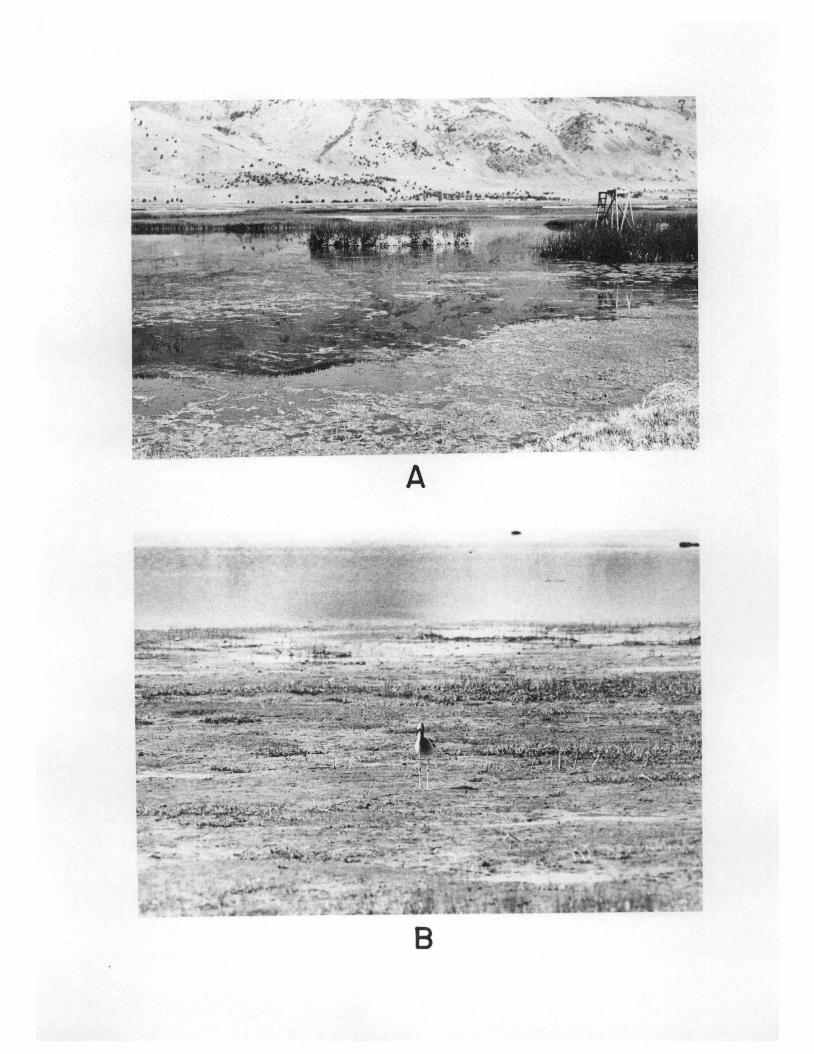
Figure 2. Habitat features of the Summer Lake ManagementArea. A. The western part of the marsh in thevicinity of Windbreak Dike. B. Typical playa onthe eastern edge of the marsh.
S
I

8
PROCEDURES
Field work was conducted over 271 days during the spring and
summer of 1967 (15 May-10 August), 1968 (7 April-13 July), and
1969 (1 April-10 July).
The quantitative data for the time budget analysis were collected
during the 1969 breeding season only. The 1967 and 1968 seasons
were spent constructing an ethogram, collecting breeding biology
information, and devising methods for quantifying behavioral activi-
ties in time.
Constructing the Ethogram
An ethogram consists of a detailed description of a species'
behavior with particular emphasis on stereotyped and semistereo-
typed action patterns. This phase of the study was necessary be-
cause of the lack of adequate descriptive accounts of avocet behavior
in the literature. A few sketchy observations were given in Bent
(1927), and an ethogram of the European Avocet (Recurvirostra
avocetta) was pllblished by Makkink in 1936. The relevance of
Makkink's work to the behavior of the American species could not
be determined until observations on the latter were completed.
It is essential in any study of the apportionment of time to
various activities that all activities be precisely described and

9
categorized. Behavioral observations were made principally from
an automobile or from 3.3 m towers. Observations were either taken
directly as field notes or dictated into a portable tape recorder and
then transcribed into notebooks. Observations were aided by the
use of 7 X50 binoculars, a 20 X spotting scope, and cine and still
photography.
Breeding Biology
Information on breeding biology, like the basic behavioral de-
scriptions, was inadequate prior to this study. Therefore basic life
history data were obtained using the following methods.
I located all nests along Windbreak dike in 1967 and 1969, while
only selected nests were studied in 1968. Eggs were marked with
fingernail polish and their fate followed by twice daily checks during
egg laying and hatching, and alternate day checks during incubation.
Time budget data were used to obtain estimates of the roles of the
sexes in such activities as incubation and parental care.
In order to assess the flux of breeding activities for a larger
segment of the Summer Lake population, a 3.75 km census route
was established (Figure 1), along which all avocets were counted
and their activity recorded. This gave an index to the number of
birds in each stage of the breeding cycle, since particular action
patterns are characteristic of certain breeding stages.

10
To determine the position, boundaries, and patterns of utiliza-
tion of avocet territories, I recorded the position of both members
of a pair within a 15.2 X 15.2 m grid system at 10 sec. intervals.
Observations were collected and mapped to provide information on
flux of territory size and utilization patterns.
Marking Individuals
Individually marked birds were essential to the study so that
individuals could be followed for long periods of time and their breed-
ing schedules monitored. A large number of marking methods were
tried, but most proved unsuccessful. Dyeing adults with dyes such
as Nyazol A (black), picric acid (yellow), rhodamine B (red) and
methyl green (blue) did, however, prove successful. The dye was
applied by placing a piece of cellulose sponge one inch square in the
nest with a nail. The sponge was soaked with dye and when the bird
returned to the nest a large splotch of color was transferred to 5+3
breast. The yellow color of picric acid presented a problem, how-
ever. When an incubating bird found the yellow object in its nest,
it immediately deserted. This may have involved a reaction similar
to that evoked when a bird discovers a nest full of broken eggs with
yolk showing. The problem was averted by mixing a little methyl
green with the picric acid, resulting in a green dye. A day or two

11
after application the green color was gone, leaving yellow. All
dyes except methyl green lasted at least a month.
The Time Budget
A total of 422 hours of time budget data was recorded for 36
different avocets (213 hours on males and 209 hours on females). The
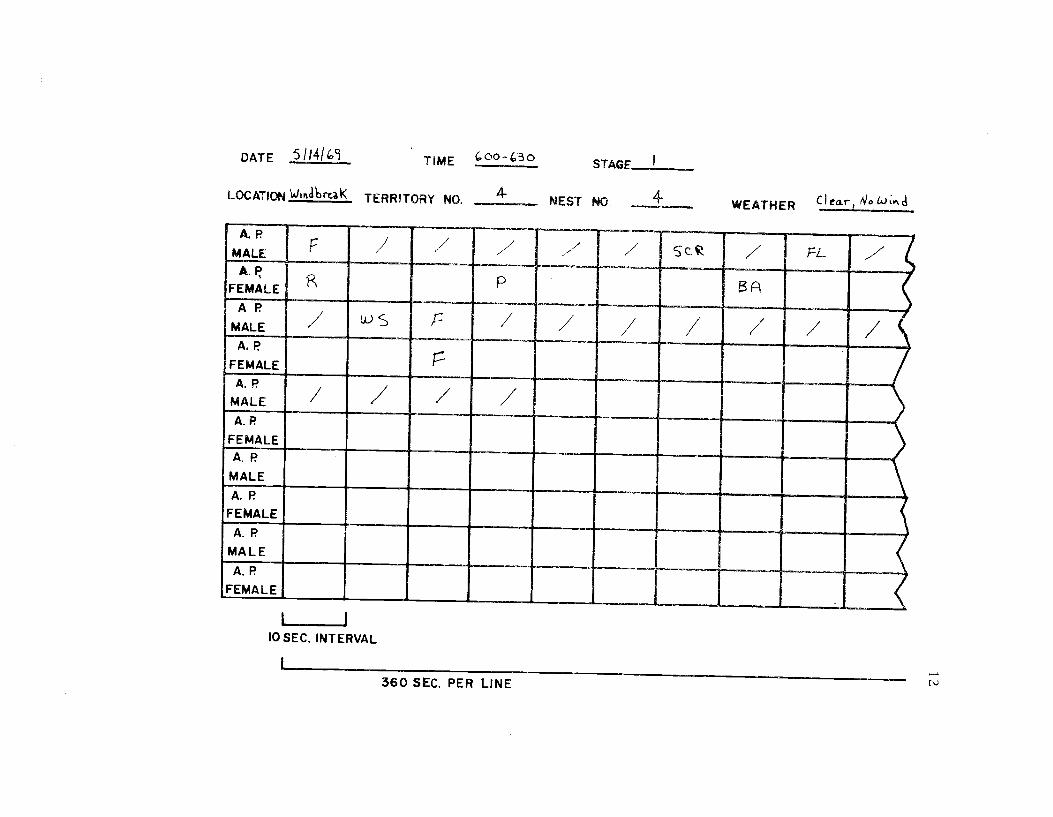
time spent in 40 different categories of behavior (see Ethogram) wereused in the time budget. A system of shorthand symbols was devel-oped so that action patterns could be rapidly recorded on a standard-ized data form (Figure 3). The activities of both members of a pairwere recorded simultaneously at 10 sec. intervals for a 30 min.
observation period. The time base was kept with a periodic metro-
nome timing device (Wiens et al., 1970) which was set to pulse every10 sec.
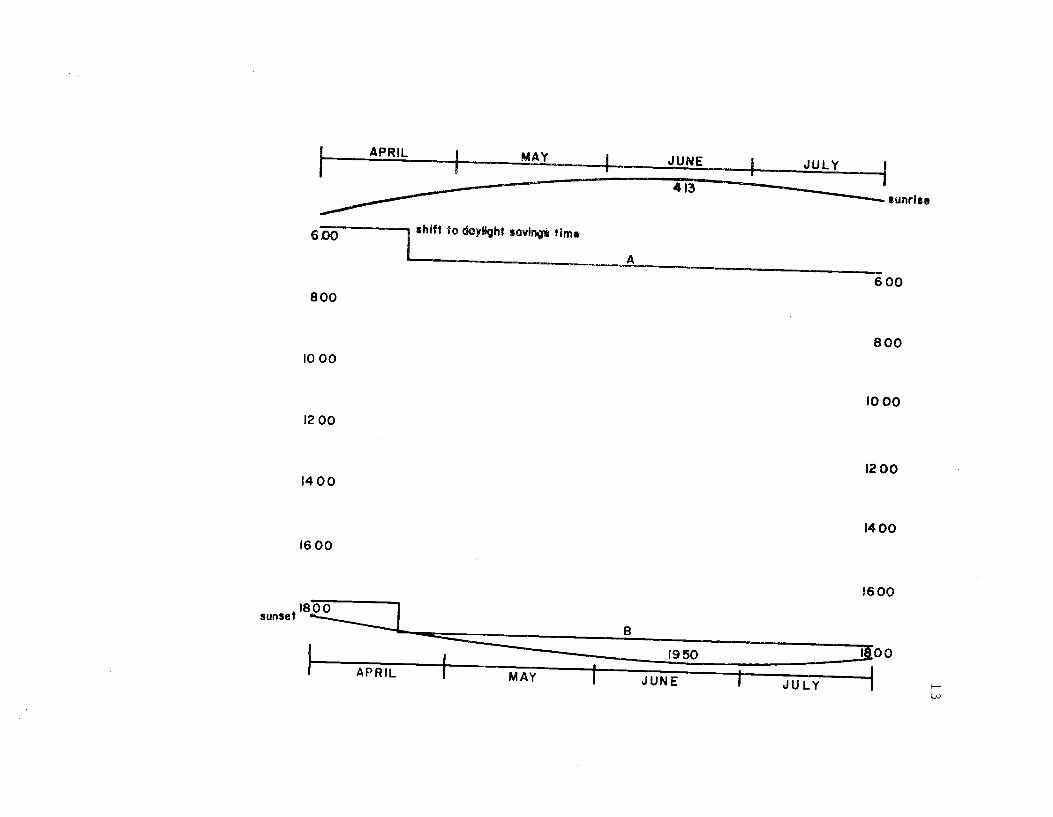
Time budgets were taken systematically from 0600 to 1800 sothat the 12 hr. period was sampled every three days. Sampling the
same 12 hr. period through the breeding season had its drawbacks,
since there were periods of daylight before and after the interval of
observation (Figure 4). Further, the period sampled changed in
relation to sunrise and sunset as the season progressed. This made
the diurnal time budgets somewhat incomplete, but since I examined
mainly general patterns of time utilization, this incompleteness
should not mask overall patterns.

Figure 3. A portion of a standardized data form on whichtime budget information was taken. A completedata sheet was used to record the action patterns(A. P.) of both males and females for 30 min. Ateach 10 sec. interval the action pattern of each sexwas recorded using shorthand symbols. If therewas no change a slash was marked through theinterval. F = feeding; SCR = scratching; Fl =flying; R = resting; P = preening; Ba = bathing;WS = wingshake.
DATE 5114/ " TIME Coo-630STAGE
LOCATION Winlbrca < TERRITORY NO. NEST NO. Cl ear 4o laiWEATHER
10 SEC. INTERVAL
360 SEC. PER LINE N.)

Figure 4. The diurnal period during which behavioralobservations were made. Observations wereinitiated at 0600 (A) and terminated at 1800 (B).The entire daylight period was not sampled.
APRILJUNE 4 JULY
600
800
10 00
12 00
shift to daylight savings time
14 00
16 00
sunset1800
APRIL 1 MAY JUNE 1 JULY
sunrise
6 00
800
to 00
12 00
14 00
1600

14
Eighteen avocet pairs were observed during the season, but
only four pairs were followed through a breeding cycle. The length
of the breeding cycle (60-70 days) for a single pair prevented inten-
sive study of additional pairs. The selection of pairs for observation
was done as systematically as possible to avoid the problems of oppor-
tunistic observations (Wiens et al., 1970). The pairs being used for
collecting time budget data were numbered sequentially and observed
on a rotating basis.
Analysis of the Data
The time budget data were compiled and analyzed on the CDC
3300 operated by the Oregon State Computer Center. The raw data
were coded and punched onto computer cards. A total of 12,000
cards was necessary to handle the 422 hours of data. Computer
analysis was essential because of the difficulty of calculating the
time utilization of 40 action patterns for two sexes for four breeding
cycle stages and for six diurnal time periods within each stage; a
nearly impossible task by hand.
Since I wished to express the time budget in terms of the per-
cent of the total time spent in each activity, the computer output in-
cluded these analyses. But this presented a problem of applying
statistical methods to percentages. Due to lack of funds the data
could not be reanalyzed in such a way so that standard statistical

15
methods could be applied. Therefore, to generate confidence inter-vals an approximation to the binomial was performed on each per-
centage (Huntsberger, 1967). This is valid since percentages are
ratios based on 100 percent. The drawback to this method is thatthe confidence interval generated is strongly dependent on samplesize. Therefore, I had to decide on a unit of time to use for the
analysis. My decision was to use minutes since I felt that this unithad some biological meaning. In other words, I felt minute to minute
changes in activities were more meaningful in the time budget than
second to second or hour to hour changes for many activities. This
selection is obviously a compromise but I feel it is the best of those
available. More specific information about certain analytical pro-
cedures will be covered in each section as they are used.

16
BREEDING BIOLOGY
The American Avocet is a fairly large wader, 39 to 52 cm in
length. In the breeding plumage the avocet has a rust-colored head
and neck with a bold black and white pattern on its back. A major
characteristic is its recurved bill. There is some sexual dimorphism,
in that the female always has a more strongly recurved bill than the
male (Hamilton, 1969 ).
The American Avocet breeds in western North America and
winters along the Pacific coast from San Francisco to Central
America and inland across the southwest United States and northern
Mexico (Figure 5) (Hamilton, 1969). The preferred breeding habitat
is shallow alkaline or saline marshes and lakes. Nesting sites are
usually in open areas with sparse vegetation such as low grass
covered islands, shores or mudflats.
Migration and Arrival
Avocets have both migratory and non-migratory populations.
All populations in Oregon are migratory. Paths of migration and
the respective wintering areas of specific populations are unknown.
Summer Lake supports one of the largest breeding populations in
Oregon.
During my observations, spring migrants began arriving at

Figure 5. The breeding and wintering distribution of theAmerican Avocet (from Hamilton, 1969).
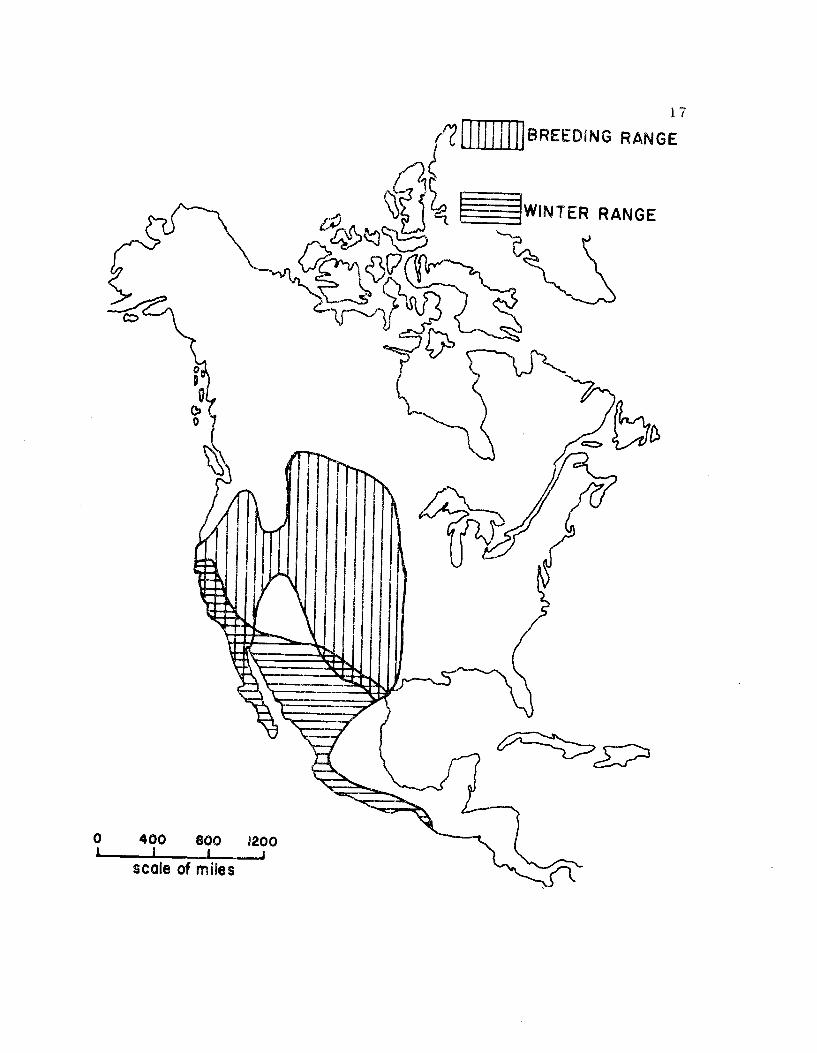
11111111
17
BREEDING RANGE
WINTER RANGE
0
0 400 800 1200
scale of miles

18
Summer Lake around 20-25 March and most had arrived by the end
of April (Figure 6). The greatest influx of birds occurred between
1-15 April. In early April single birds and small flocks of four to
ten were frequently observed. Whether the birds migrate in flocks
or as individuals, at night or during the day, is unclear. Hamilton
(pers. comm.) suggests that they migrate at night since he found
that in certain wintering areas in south San Francisco Bay there
were fewer and fewer birds each day until all birds had gone.
At the peak of migration in mid-April large flocks of avocets
were present in the larger bodies of water within the marsh. The
number of birds varied considerably from day to day, suggesting
that Summer Lake was a stopping-over spot for many avocets con-
tinuing on to other areas.
Breeding Chronology
Avocet breeding activities extended over a four-month perin-2
from late March until late July. Figure 7 summarizes the breeding
chronology for avocets at Summer Lake during the 1969 season.
I have divided the breeding cycle into four stages:
Stage 1: Preresting includes all activities up to the laying of
the first egg. For any single pair, territory establishment, court-
ship, nest site selection, and nest building activities occurred during
the same time period and therefore could not be separated into

Figure 6. An index to avocet population numbers in theSummer Lake Management Area during the 19 69breeding season.
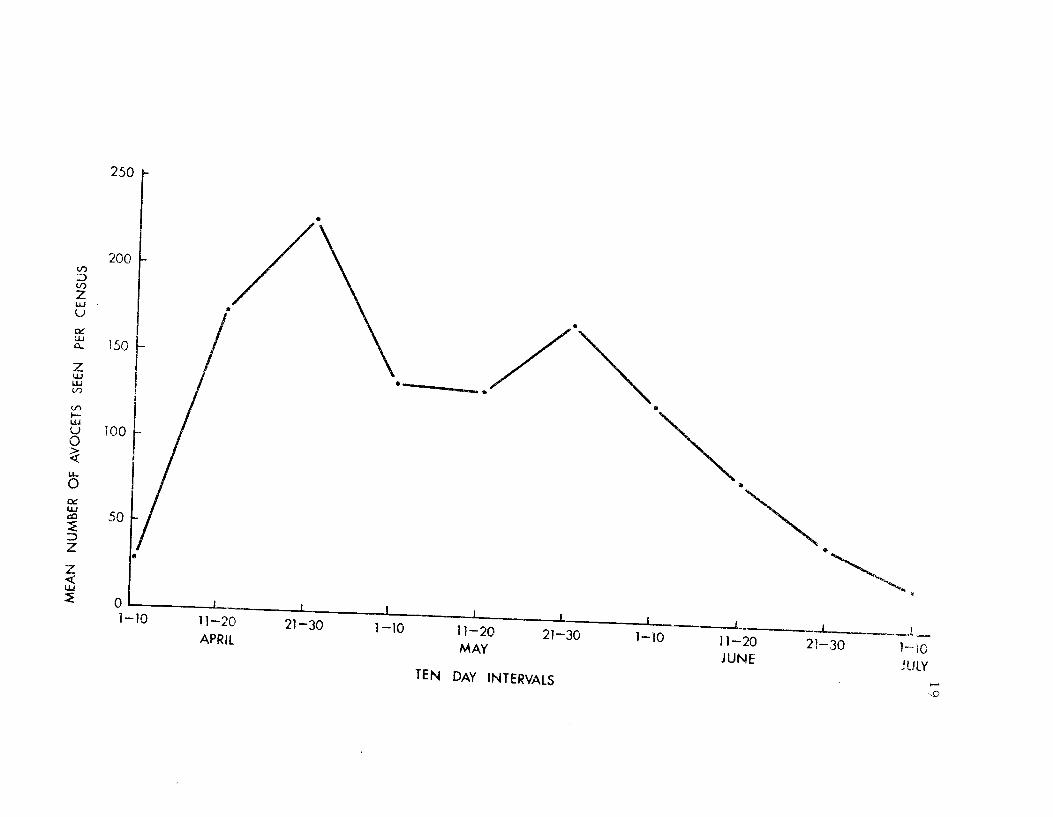
zL1J
250
200
150
100
APRILMAY
TEN DAY INTERVALSJUNE

Figure 7. A. A breeding chronology for the avocet populationat Summer Lake during the 1969 breeding season.B. A breeding chronology for a single pair. Thebeginning and end of each stage are based on dateswhen various breeding activities were first and lastobserved.
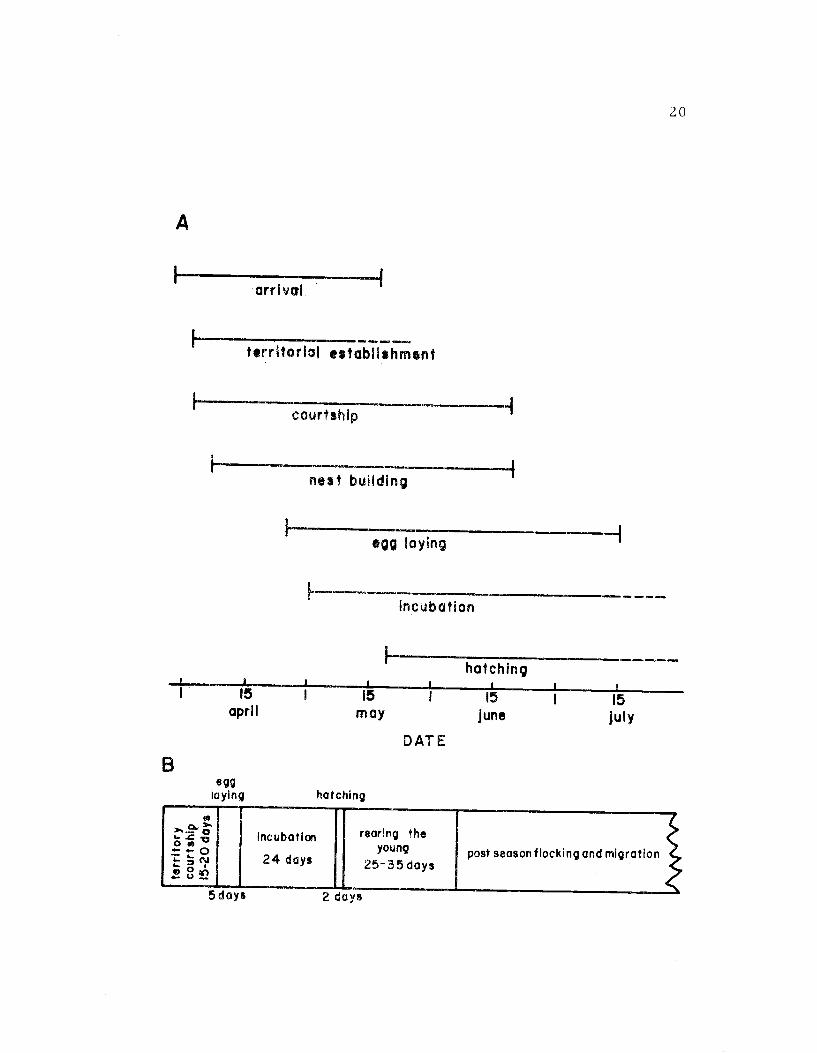
A
I
arrival
H
IMO alb WIZ 7.1M.
territorial establishment
courtship
nest building
1
egg laying
incubation,a,
20
hatching
15 15 15april may June
DATE
egglaying hatching
Incubation
24 days
5 days
rearing theyoung
25-35 days
15
July
post season flocking and migration
2 days

21
stages (Figure 7B). For the majority of the population this stage
lasted from arrival until around May 1st.
Stage 2: Incubation began with the laying of the first egg and
continued until the last egg hatched. The incubation stage was ex-
tended through laying and hatching because avocets sat on the eggs
during both of these periods and it was difficult from behavioral
observations to make a distinction. This stage lasted from about
1 May to approximately 15 June.
Stage 3: Parental Care began when the chicks hatched and
continued until the young were fledged. Precise information about
the duration of this stage is lacking because of the difficulty of observ-
ing family groups. Parental care continued from late May until ap-
proximately 15 July.
Stage 4: Post-breeding Flocking began after breeding ceased
for a particular pair. Usually by late June and Early July a sufficient
portion of the population had completed breeding activities to allow
the formation of flocks. Some avocets remained at Summer Lake un-
til October or November.
Territory
Avocets at Summer Lake established and maintained territories
which were defended by both sexes. Territories assumed three
different forms coinciding with the pre-nesting stage, the incubation

22
stage, and the parental stage. Prior to egg laying territories were
centered around feeding areas. Suitable nest sites were generally
available within a few hundred meters of the feeding areas. On my
main study area the feeding and nest sites were in close proximity.
However, on the east side of the marsh where open water was much
more localized, nest sites were not always near the feeding area.
During incubation the form and manner of defense of the terri-
tory changed since it was necessary for one member of the pair to
be present at the nest most of the time. At the Windbreak Dike study
area, where nest sites were surrounded by the feeding areas, a
secondary feeding site was usually established 50 to 130 m from the
nest (Figure 8), usually in a large pond. This secondary foraging
area was also defended against other avocets. The task of defending
two separate areas restricted the effectiveness of territorial defense,
with the result that violations of both territorial spaces by other birds
did occur. Avocets only defended the area on which they were pres-
ent; incubating birds, however, did not leave the nest to defend the
territory. Therefore only one area could be defended at any time,
and intrusions by other birds in the absence of the territory owners
were frequent.
The situation was slightly different at sites where the nest
was located away from the feeding area (Figure 8B). Here only one
foraging site was maintained and there seemed to be only a small

Figure 8. A. Territories of three pairs during the incubationperiod along Windbreak Dike. Each pair defendedtwo territories one which included the nest and asecond in a large pond nearby. B. Territories ofthree pairs located on the eastern edge of theSummer Lake Management Area. The nests wer elocated in an alkali flat some distance from thedefended feeding area resulting in only one foragingsite being defended.
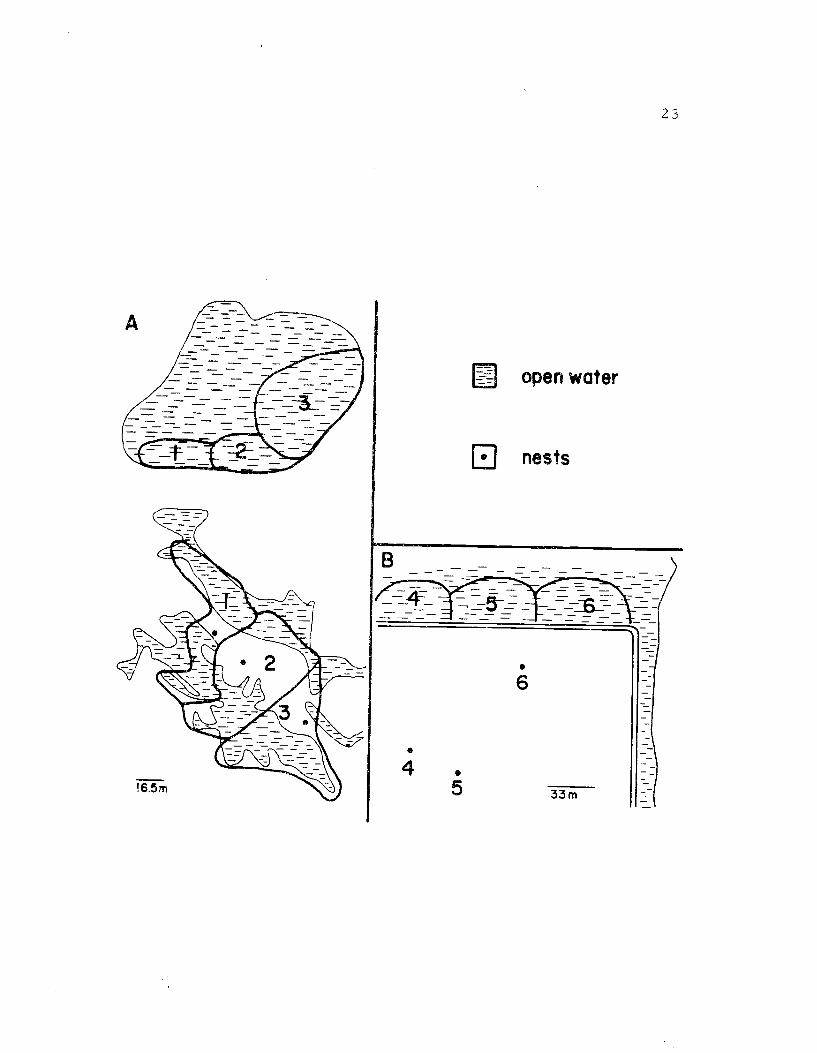
23

24
territory associated with the nest site. The feeding territory was
more exclusive in this case because one member of the pair was
usually in attendance.
Once the eggs hatched the territory became chick-centered
and somewhat mobile. The adults usually moved the chicks to a
suitable foraging site, maintaining a strict territory of roughly 50-
100 m in diameter around them at all times. Defense was inter-
specific as well as intraspecific. Nearly all avian species were
excluded with attacks being most frequent on blackbirds (Icteridae)
and ducks (Anatidae).
Lack (1968) considers members of the family Recurvirostridae
to be monogamous. However, there was not complete fidelity between
members of a pair at Summer Lake. In 1969 I observed three males
and one female (all marked) copulate with birds other than their
mates. These opportunistic copulations always occurred during
the incubation period on the territory of the marked birds. The
breeding condition of the unmarked avocets could not be determined
with accuracy, but in one case a female was followed for several
minutes after a copulation. She intruded other territories and was
driven off, indicating that she might be unpaired. Perhaps the avocet
resembles the oystercatcher (Haematopus ostralegas) in its tendency
to mate with birds outside the pair bond (Makkink, 1942).
Observations on a pair of colorbanded birds in 1967 suggest that

25
pair bonds may last for more than one season. This pair was seen
with young in 1968 but was absent from the study area in 1969. They
were observed together again in May, 1970. Whether they remained
together for the entire year is unknown.
The Nest
Numerous accounts of avocet nest construction have been pub-
lished (Wetmore, 1925; Bent, 1927; Wolfe, 1931; Wheeler, 1955;
Hamilton, 1969). In summary, the nest is a simple scape 15-25 cm
in diameter and 0.5 to 3 cm deep, lined with a variety of materials
from grass to mud chips. The amount of lining is variable; some
nests contain none while others are built up to 38 cm in height
(Wetmore, 1925) in areas subjected to flooding. The largest nest
I found was 7.5 cm high. I sampled 56 nests in order to determine
the materials most commonly used in nest construction and the vege-
tation predominating in the area immediately about the nest (Table I ).
There was a correlation between nest lining and the vegetational sur-
roundings, suggesting that the nest materials were gathered in the
immediate vicinity of the nest. Indeed, behavioral observations
substantiated this view (see Ethogram). The maximum distance
traveled from the nest to obtain materials was only 2-3 m. From
Table 1 it appears that the avocets nesting on Windbreak Dike pre-
ferred salt grass to all other types of vegetation for nesting sites.
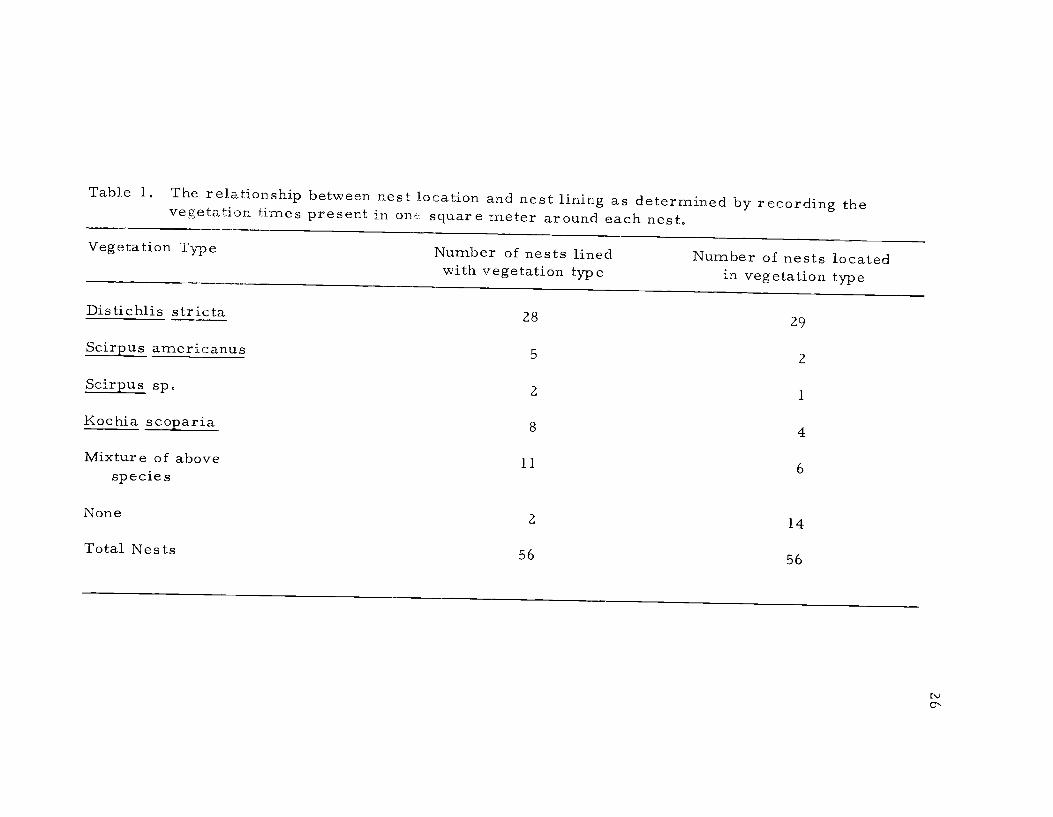
Table 1. The relationship between nest location and nest lining as determined by recording thevegetation times present in onE square meter around each nest.
Vegetation Type Number of nests lined Number of nests locatedwith vegetation type in vegetation type
Distichlis stricta 28 29
Scirpus americanus 5 2
Scirpus sp. 2 1
Kochia scoparia 8 4
Mixture of above 11 6specie s
None 2 14
Total Nests 56 56

27However, in other areas of the refuge avocets nested in other situa-tions as well, including alkali flats devoid of vegetation.
Nest Dispersion
Lack (1968) states that Recurvirostrids nest in loose coloniesand Hamilton (1969) makes a similar statement specifically forR. americana. Loose colonies are generally located on sites whichare more accessible than those of more colonial species, but thenests within the colony are dispersed to aid concealment. Speciesnesting in loose colonies usually do not defend the colony as a group,unlike more colonial birds such as gulls. Therefore, the advantageof a loose colony probably stems from some factor other than safetyfrom predators, presumably feeding behavior (Lack, 1968, p. 140).
Avocets at Summer Lake tended to nest in groups of varyingdensity (Figure 9). Nesting groups were located near foraging areas,suggesting that proximity to suitable feeding sites is an importantfactor in the placement of nests. Hamilton (1969) found that avocetsin his study area also chose nesting sites on dikes which were closestto foraging areas.
The social factors operating in the formation of avocet nestinggroups are unclear. Counter to Lack's (1968) generalization, reac-tion to predators by avocets seems to be an important group activity.When an avian predator, such as a hawk or gull, flew over a nesting

Figure 9. Avocet nest dispersion in different habitats atSummer Lake. A-B. The dispersion of nestsat the same site in 1969 and 19 70 respectively.The area is a salt grass habitat located alongWindbreak Dike. The mean distance betweennests in A was 47. 3 m and in B 29.8 m. C. Nestdistribution in a large salt grass field. The meandistance between nests was 80. 0 m. D. Nestdispersion in an alkali flat where themean distancebetween nests was 50 mo E. Nest distribution onand near Windbreak Dike in 1967 and 1969.
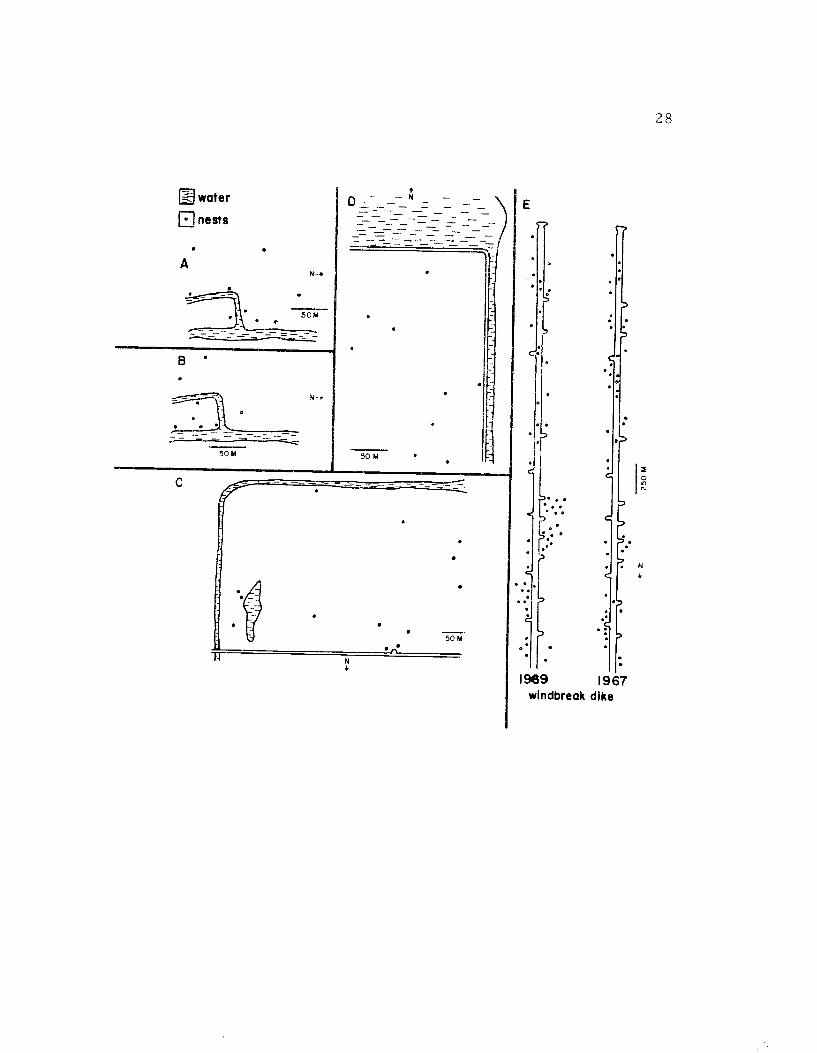
El water
nestsaA
~50M a
C
50
50w
s
1989 1967windbreak dike
28

29
area the avocets responded with loud vocalizations and mobbing.
Many avocets chased and dove at the bird. This form of attack was
quite successful in keeping hawks and gulls away from nesting sites.
Avocets responded to terrestrial predators differently. When a hu-
man approached a nesting group most of the birds began calling and
flew to a spot nearby but away from the nests. The entire group
then began giving distraction displays (Hamilton, 1969). I witnessed
up to 35 birds taking part in this activity at a time. I have seen a
deer and coyote effectively distracted by this activity.
Egg Laying
Bent (1927) and Hamilton (1969) state that avocets normally
have a clutch of four eggs. I found a mean clutch size for 111 nests
o 3. 7 (2-4). Nests containing six or more eggs were not included
in the clutch size calculations, as they probably did not represent
the clutch of a single female. Length and width measurements of
eggs in 50 clutches revealed that the eggs of a single female were
uniform in size. Nests containing six or more eggs had two distinct
size classes, suggesting that two females laid the eggs. Dump nest-
ing in this species requires a violation of territory; unfortunately the
mechanism by which this was accomplished was not observed.
The earliest complete clutches were found in late April or early
May. In 1967 and 1968 the first clutch was found on 7 May and in

30
1969 on 24 April. Egg laying reached its peak between 1 1 and 20 May
in 1968 and 1969 (Figure 10).
The laying period for a clutch of four eggs was usually five
days. The interval between eggs was approximately 24-30 hours.
Nests were checked daily between 0800 and 1600. New eggs were
found at the morning check but not in the afternoon, indicating that
eggs were laid in the 16 hour interval between 1600 and 0800.
Data from three nests deserted due to human disturbance
showed that avocets can renest a short time after losing a clutch.
One pair copulated and initiated nest construction one day after de-
sertion. Two pairs completed second clutches of four eggs in eight
days and a third pair took six days (3 eggs). Each pair had less than
four days remaining in the incubation period when they deserted, so
the effect of the stage of incubation on the rapidity of renesting could
not be determined.
Incubation
Both sexes developed incubation patches and incubated, reliev-
ing each other at the nest rather often. From 480 minutes of obser-
vation at each of the three nests, I found that the female's incubation
intervals averaged 52.2 minutes (3 to 255), while the male's aver-
aged 38.0 minutes (3 to 75). Hamilton (1969) reported that males
incubated 87.6 minutes (N=26 intervals) and females 64.3 (N=18

Figure 10. Time of nest initiation at Summer Lake in 1968and 1969. Nest initiation was defined by thelaying of the first egg.
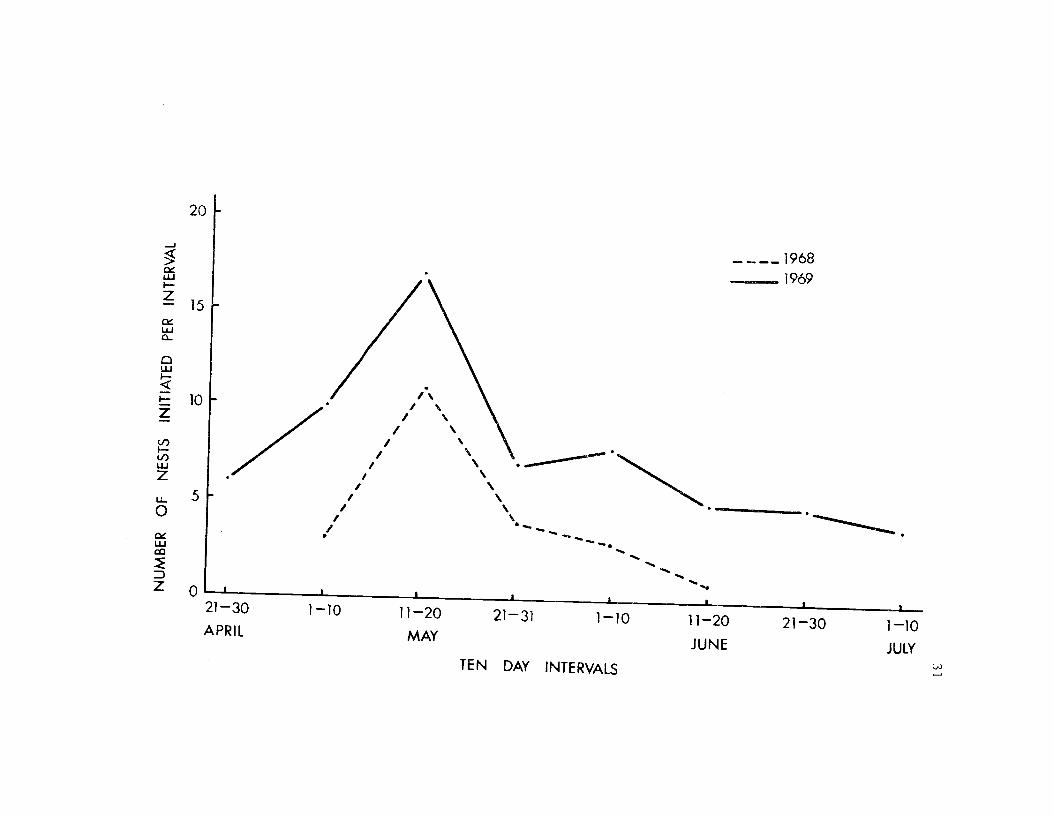
196c3
1969
04
'Z.
21-31
IES DP( It411.1VPA.5

32
intervals). Both estimates are based on small samples and the
large ranges involved indicate that there is considerable variation.
Time budgets of four pairs were analyzed to determine the
amount of time each sex spent on the nest throughout the incubation
period (Table 2). During the first eight days the male spent nearly
twice as much time on the nest as the female. However, these first
eight days were characterized by a high degree of nest inattentive-
ness. The female predominated during the last 16 days of incuba-
tion. Hamilton (1969) found that the male incubated about twice as
much as the female but he gives no indication at which stage of the
incubation period he collected his data.
The incubation period, defined as the interval from laying of
the last egg until that egg hatches, was 24.2 (22-29) days for 28
nests. Effective incubation probably did not begin until the last egg
was laid because all the eggs generally hatched within a day or two.
However, avocets often covered incomplete clutches during especially
cold or hot weather.
As the season progressed the length of the incubation period
tended to decrease. Nests started in late April averaged 27 days of
incubation; by early May the period had dropped to 25; by mid May
to 24; and finally to 23 in early June. The increase in daytime tem-
peratures as the season progressed may have allowed development
to continue even when the eggs were unattended.
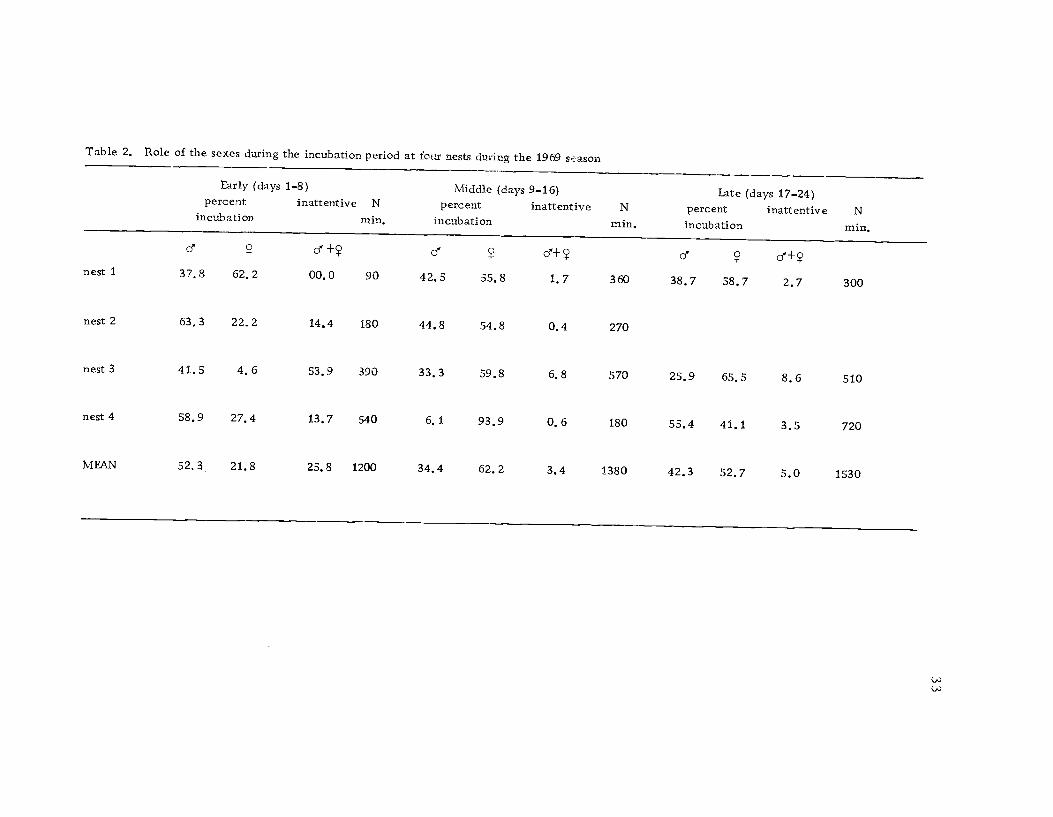
Table 2. Role of the sexes during the incubation period at four nests during the 1969 season
Early (days 1-8) Middle (days 9-16) Late (days 17-24)percent
incubationinattentive N
min.percent
incubationinattentive
min.percentincubation
inattentive N
min.
cf +? d cf+? d cf+Ynest 1 37.8 62.2 00.0 90 42.5 55.8 1.7 360 38.7 58.7 2.7 300
nest 2 63.3 22.2 14.4 180 44,8 54.8 0.4 270
nest 3 41.5 4.6 53.9 390 33.3 59.8 6.8 570 25.9 65.5 8.6 510
nest 4 58.9 27.4 13.7 540 6.1 93.9 0.6 180 55.4 41.1 3.5 720
MEAN 52.3 21.8 25.8 1200 34.4 62.2 3.4 1380 42.3 52.7 5.0 1530

34
Avocets will incubate long past the normal incubation period
if the eggs do not hatch. One pair attended a nest for 39 days before
it was destroyed.
Hatching
Hatching usually occurred over a one or two day period. The
chicks began to pip the eggs four or five days before hatching; they
could be heard calling in the eggs at this time. Shortly before hatch-
ing, a 4-15 mm hole was pipped in the egg. As soon as a chick
freed itself from the egg, a parent removed the shell, dropping it
from 5 to 50 m from the nest. The nidifugous young were usually
dry and able to leave the nest within an hour or two. However the
chicks tended to stay in the nest or near it for at least a day if not
disturbed.
Hatching dates for 78 nests followed during the 1968 and 1969
seasons are summarized in Table 3. Hatching success is similar
to that of other Charadrii (66-96 percent) (Boyd, 1962). Nest losses
due to predation were low; two nests were destroyed by a badger
(Taxidea taxus ); there was no evidence that avian predators de-
stroyed any nests. This may have been a result of the mobbing
behavior of the birds.
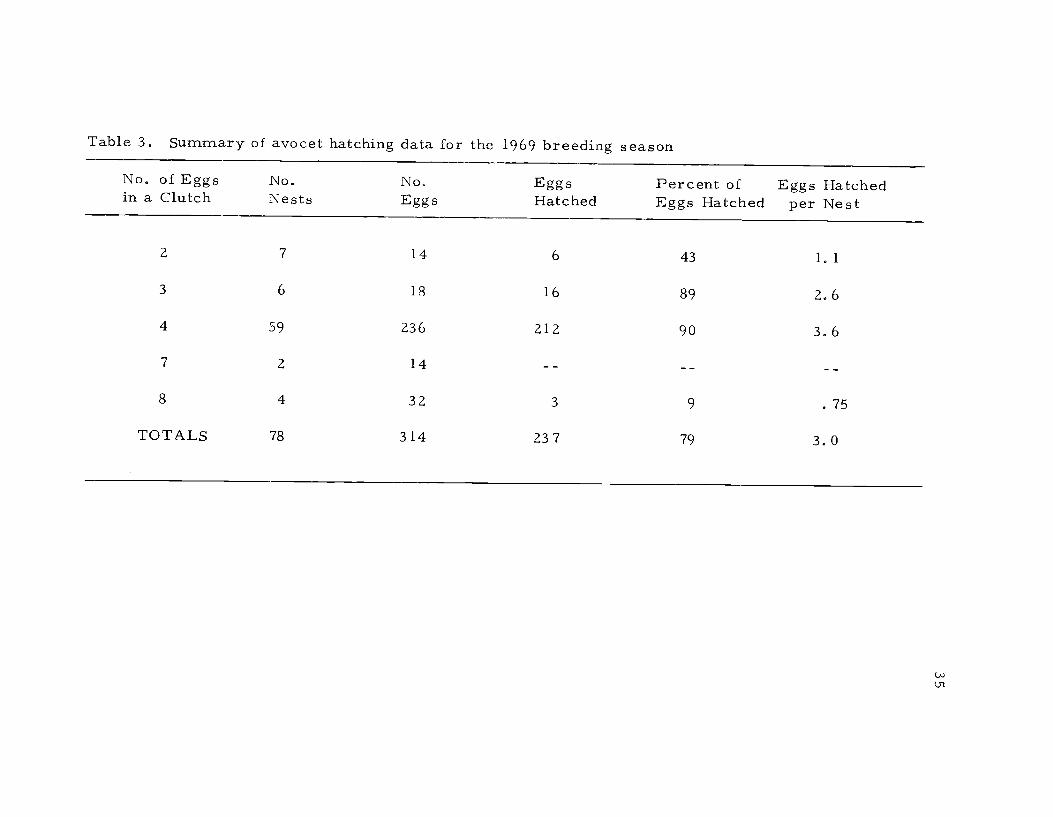
Table 3. Summary of avocet hatching data for the 1969 breeding season
No. of Eggsin a Clutch
No.Nests
NoEggs
EggsHatched
Percent ofEggs Hatched
Eggs Hatchedper Nest
2 7 14 6 43 1.1
3 6 18 16 89 2.6
4 59 236 212 90 3.6
7 2 14
8 4 32 3 9 .75
TOTALS 78 314 23 7 79 3.0

36
Care of the Young
Avocets are attentive parents even though they do not feed their
young. Once the chicks hatched, the parents became aggressive
toward all other species including other avocets.
Hamilton (1969) describes broods which contained two age
classes of chicks. He explains the occurrence of mixed broods by
suggesting that avocets are unable to recognize their own young and
lack territoriality during the parental stage. I observed no mixed
broods at Summer Lake. The mixing of broods would require an
intrusion of territorial boundaries. However in certain situations,
when cover is lacking, a disturbance such as an approaching human
creates chaos, with chicks running in all directions. It seems pos-
sible that under such conditions broods could become mixed.
The young spent most of their time feeding and resting. They
exhibited preening and bathing behavior when less than a week old.
Chicks usually ran for cover and lay flat when approached, making
them very difficult to find (Hamilton, 1969). Hiding behavior lasted
until at least the third week, after which they would just run. Chicks
attempted to dive if chased in the water.
When young avocets became separated from their parents they
were often attacked by other avocets. In one instance, in an open
expanse of shallow water, a lone chick was attacked numerous times

37
by four different adults. The young bird was either struck by the
attacker's feet or picked up in the bill and violently shaken. Hamilton
(1969) also reports this activity, as does Makkink (1936) for R .
avocetta.
Parental care continued until after the chicks could fly (four
to five weeks). The members of one brood were able to fly in 27
days. Parental attentiveness waned as the young matured and the
parents spent less time defending the area around them. The actual
duration of the family group is unknown.
In order to determine how successful avocets were in raising
young once they left the nest, I counted the number of young per
brood for two different age classes, those in downy plumage (one to
two weeks of age) and those in juvenile plumage (three to four weeks
old). Weekly counts were made for three weeks during peak hatching
and rearing periods (Table 4). There was no difference between the
number of chicks in the two age classes of broods, each having ap-
proximately three. This indicated that those pairs which hatch young
and do not lose the entire brood are quite successful in raising young,
at least through the first month following hatching.
Preparation for Migration
Once the young could fly the birds joined a post-breeding flock.
This usually involved moving to some of the larger bodies of water
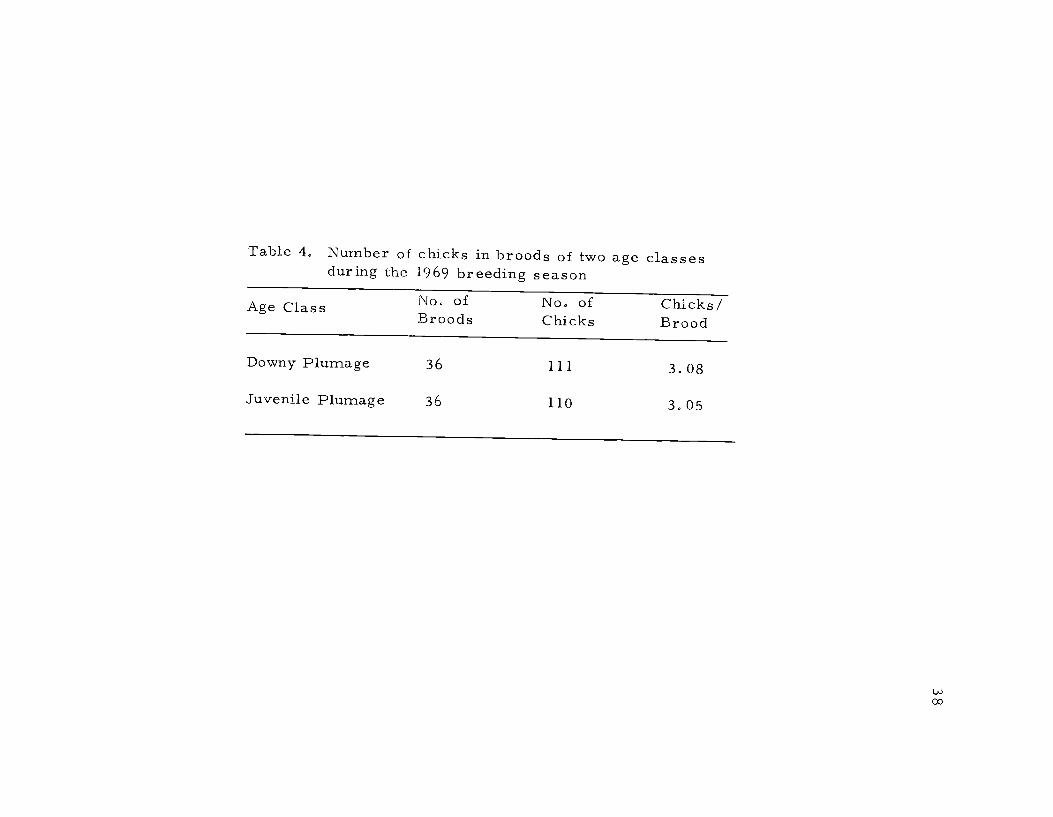
Table 4. Number of chicks in broods of two age classesduring the 1969 breeding season
Age Class No of No. of Chicks/Broods Chicks Brood
Downy Plumage 36 111 3.08
Juvenile Plumage 36 110 3.05

39
within the marsh or to the lake. Flocks began forming in late June
and by mid-July thousands of avocets were congregated along the
west shore of the main lake. The large numbers of birds observed
could not all have come from the marsh. Summer Lake thus seemed
to be a stopping place for migrating avocets, as large numbers could
be seen there in September and even into October (A. B. Claggett,
pers. comm.). The postnuptial molt took place while the birds were
in these flocks.

40
THE ETHOGRAM
Detailed knowledge of a species' behavior is necessary when
studying relationships between its behavior and ecology. An account
of a species' behavioral patterns is classically called an ethogram.
The emphasis in this ethogram will be on descriptions of action pat-
terns and displays characteristic of avocets during the breeding sea-
son. Each phase of behavior will be described, and its overall con-
text and functions given. Many of the action patterns have been previ-
ously described by Hamilton (1969). I have attempted in most cases
to comply with his terminology. The importance of behavioral activi-
ties to this study necessitates that all action patterns used be precisely
described and classified. The separation of behavioral patterns into
discrete units is somewhat subjective. Beginnings and endings of
certain action patterns are indistinct and therefore boundaries must
be established by definition. Precise descriptions make rapid record-
ing of behavioral events in time possible and less subject to error.
Undoubtedly some distortions may occur due to this procedure, but
it is unlikely that it would make a significant difference in the inter-
pretation of the time budgets. For convenience, the avocet's behav-
ioral activities have been divided into broadly functional groupings,
for example maintenance or intraspecific aggressive activities
(Table 5).

Table 5. Summary of avocet behavior patterns
Maintenance BehaviorFeedingFlock feedingFormation FeedingRestingBathingPreeningScratchingTwo-wing StretchWing-leg stretchWings hakeDrinkingFlying
Mating BehaviorPrecopulatory breast preeningNeck ext endedMountCrossed BillsPostcopulatory walk
Aggressive BehaviorIntraspecific aggression
CrouchSingle crouch -runPaired cr ouch -runRetreatPursuit attackBowingJump attackSquatPsuedosleepingParallel walk
Interspecific aggressionPredator chaseDiveWingspread
Diversionary BehaviorTightropeSneak-runBreast dippingAlarm circlingAlarm calling
Nesting BehaviorGround checkingThrowing strawsScrapingNest approachIncubation
Parental BehaviorBroodingLeading
Miscellaneous Action PatternsUprightHead bobbingAbsent
41

42
Maintenance Behavior
Maintenance activities can be considered those behaviors"concerned with locomotion and the general health and efficiencyof the body" (Mar ler, 1956). Maintenance behavior occurs through-out the year.
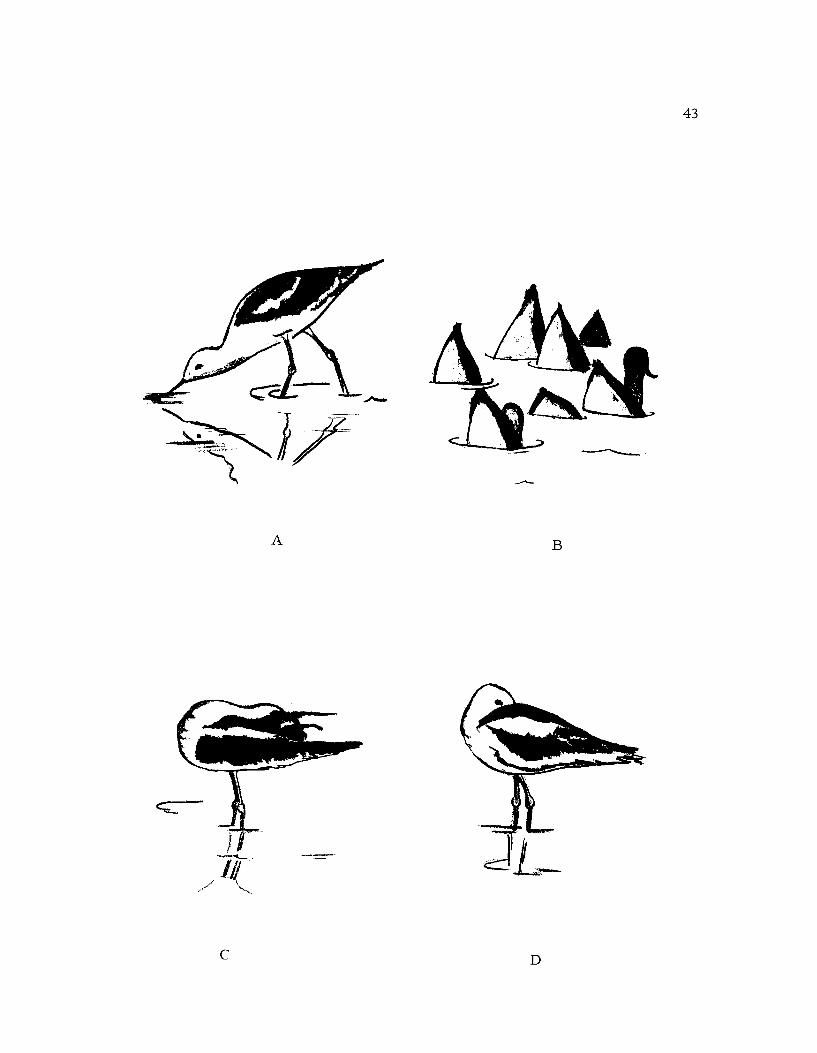
Feeding (Fig. 11A): Avocets fed in shallow alkaline or salinewater (Hamilton, 1969). Two basic patterns of feeding were em-ployed: "sweep-feeding, " which seems to be essentially tactile innature, and visual feeding. In "sweep-feeding" the recurved billwas swept back and forth across the bottom of the water. The body
was kept at about a 30 ° angle but this varied somewhat according tothe depth of the water. When a food item was captured the head andneck were raised from the water and it was swallowed. Avocets feed-ing in this manner did not hesitate to swim if the water became too
deep for wading. In visual feeding, individuals simply walked slowlyand captured prey items by pecking. This type of feeding usuallyoccurred in summer when large numbers of insects were availablein the marsh. Hamilton (1969) describes nine different feeding meth-ods used by avocets, all of which are modifications of these two basicpatterns. For the present study no separation of feeding methods wasmade.
The above mentioned basic feeding patterns were normally

Figure 11. A. Feeding: Here the sweep-feedingmotion is being used.
B. Flock Feeding: The birds are in a compact
feeding flock, each bird feeding by up-ending.
C. Preening: This bird is rubbing its head
over the uropyial gland.
D. Resting.
A
C
B
D
43

44
employed by solitary individuals and pairs, but in certain social
contexts modifications of these patterns occurred. The first, flock
feeding (Figure 11B), was associated with migratory flocks as they
arrived on the breeding grounds. Large aggregations of avocets
occurred in ponds at Summer Lake in April of each year. These
birds often formed feeding flocks of up to several hundred individuals,
although flocks of 10 to 50 were more common. The birds fed close
together (5-10 cm) and the entire flock moved rapidly through the
water as a unit. The water was usually too deep in these ponds for
wading so the whole group swam. Each bird fed by up-ending, much
as ducks do, and rapidly sweeping the bill back and forth in the wa-
ter. The position of the bill and use of the mandibles was quite differ-
ent from that of birds feeding alone. Flock feeding birds held the
bill open and kept it almost vertical. Avocets feeding in this man-
ner were able to catch small fish (Gibson, in prep. ).
Avocets also fed in groups when they were flocked prior to fall
migration. In formation feeding the form of the feeding groups was
considerably different from those of arriving birds. Three to fifty
birds spaced from 30 to 90 cm apart fed in drill-like formation with
all birds parallel to each other and changing directions as a group.
Formation feeding was most often observed along the shores of
Summer Lake in July and August. The formations wer e usually
rather transitory with birds joining and leaving almost continuously.

45
A particular feeding group would only last from one to ten minutes.
The feeding posture of the birds was similar to that of a solitary bird.
Preening (Fig. 11C): Care of the body surface, particularly the
feathers, was accomplished by preening. The feathers were manipu-
lated with the bill. The wings were preened from the lower side by
drawing each feather between the mandibles. The breast and sides
were preened by a chopping action of the mandibles. Oil was ob-
tained from the uropygial gland and spread on the feathers as de-
scribed above; the head was preened by rolling it back on the uro-
pygial gland and then rubbing it over the back. Water was often used
for preening, with the bill being dipped in the water many times dur-
ing a bout.
Resting (Fig. 11D): Resting avocets usually stood on one leg
with the other leg pulled into the plumage of the breast. The head
was turned backward and the bill was placed under the upper edge
of the wing near the center of the back, or at times the head was
merely pulled in close to the body with the bill pointing straight
forward. The birds sometimes rested on both legs, especially dur-
ing strong winds. Resting birds always faced the prevailing wind.
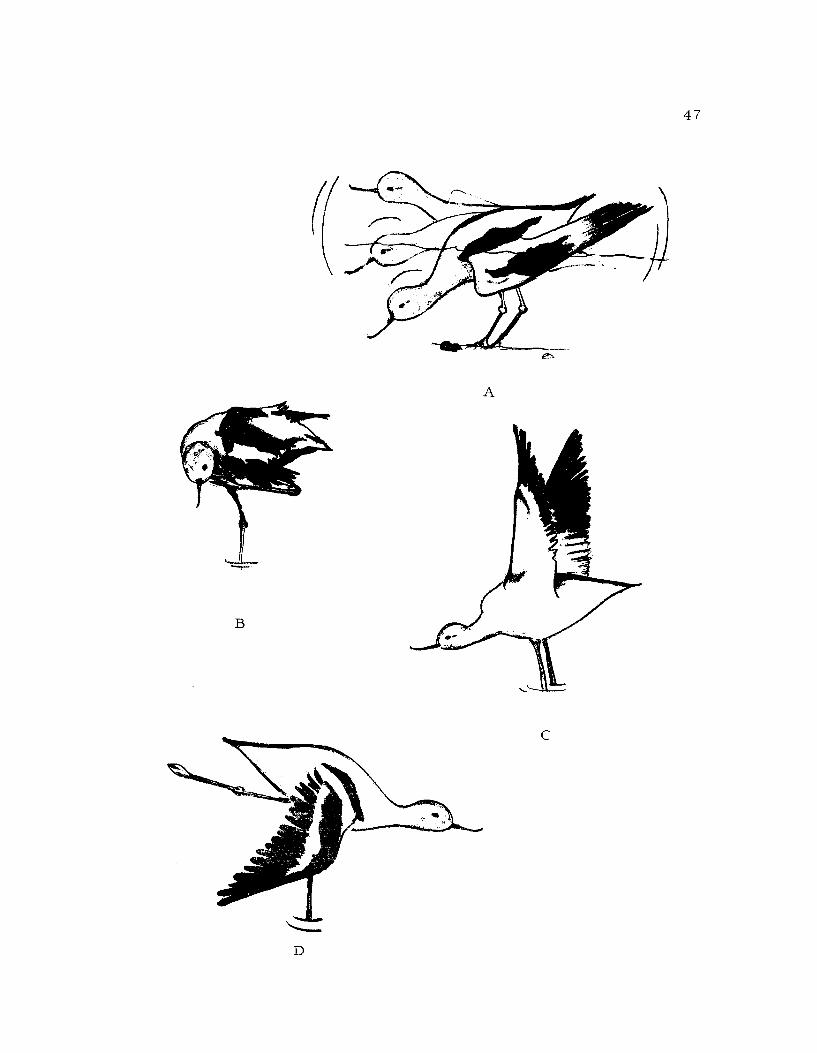
Scratching (Figure 12B): Scratching was used to manipulate
the feathers on the head. Avocets utilized an indirect or over the
wing scratch. One wing was lowered and extended and a leg was
lifted over the wing to scratch at the head and neck. The neck and

46
head were turned toward the scratching foot. Scratching was of
short duration and occurred at any time
Two-wing Stretch (Figure 12C): The two-wing stretch consist-
ed of raising the wings high over the back until they almost touched.
Simultaneously the neck was extended forward so that it had a some-
what downward arch. The function of stretching according to Sauer
and Sauer (1967) is to reactivate the respiratory center after a period
of rest. This type of stretch also occurred just prior to flight and
when the birds were disturbed (Hamilton, 1969).
Wing and Leg Stretch (Figure 12D): Avocets performing the
wing and leg stretch extended one leg backward at about 80 ° angle
and the wing on the same side was extended down beside the leg.
The primaries of the extended wing were fanned out to some extent.
Bathing (Figure 12A), Bathing occurred in shallow water where
the avocet crouched so that the breast was immersed. The wings
were held loosely at the bird's side and it rocked back and forth_
thrusting its head under the water and then raising it, causing water
to roll over its back. This movement was repeated rapidly over and
over. The wings were often rapidly splashed in the water. The wing
beating was usually interspersed with rocking. Sometimes at the
end of a bout of bathing a bird would dive under the water and propel
itself for a meter or two with its wings. Bathing was almost always
culminated with a violent wing shake and a bout of preening.

Figure 12. A. Bathing: This bird is rocking up and down tocascade water over its back.B. Scratching: The leg is placed over the wing.C. Two -win Stretch.D. Wing-leg Stretch
(t
B
A
4D
C
47

48Drinking: When avocets drank they lowered the bill into the
water, keeping it horizontal to the surface. This caused a curve in
the neck as the avocet leaned forward. After the bill was in the
water the avocet raised it, keeping it horizontal to the water's
surface until the bill could be tipped slightly upward and the water
swallowed.
Wingshake: A wingshake consisted of a rapid beating of the
wings while the bird was standing. The wingshaking often caused the
avocets to rise off the ground slightly and sometimes to hop around
to keep their balance. Wingshaking usually occurred immediately
after bathing or following preening. It was most often observed dur-
ing the post-nuptial molt, possibly in an attempt to dislodge loose
feathers.
Flying (Figure 16D): Avocets were strong and able fliers. In
flight the head and neck were extended anteriorly and the feet and
legs were extended posteriorly. Wing beats were rapid and continu-
ous during flight. They never glided except when landing. At the
end of the descending glide they made a short swoop which killed
their air speed and they dropped softly into the water or onto the
ground.
lya;ting Behavior
Avocet courtship was restricted to a sequence of stereotyped
action patterns which immediately preceded copulation. Neither sex

49
seemed to have a dominant role during sexual encounters since
either the male or female initiated the precopulatory sequence.
The male usually began a courtship sequence by preening at
his breast in a stylized manner (precopulatory breast preening;
Figure 13A). He stood fairly erect and preened at his breast with
rapid chopping motions of the mandibles, frequently dipping his bill
in the water. The male sometimes started this display without appar-
ent stimulation from the female, but more often the female initiated
the precopulatory sequence by assuming the neck extended (Figure
13A) position. She began by breast preening similar to the male
but each time she dipped her bill in the water it was extended further
and further forward until she finally assumed a horizontal position
with neck extended and parallel to the surface of the water. The
angle of the body was dependent upon the depth of the water in which
she was standing. As soon as she was in the neck extended position
the male moved to her side and began or continued precopulatory
breast preening. The male moved in a semicircle from one side of
the female to the other many times by walking behind her. As he
did this he dipped his bill in the water repeatedly. The female would
pivot, maintaining a proper orientation to the male. As the bout
progressed the male's preening became more and more intense.
He shook his bill in the water with increasing violence, spraying
water in all directions. At this point his side feathers were raised

Figure 13, A. Precopulatory Breast Preening: The malestands beside the female preening his breast.B. Mount: Front view; the feathers on the male'ssides are raised.Co Mount: Side view; showing the position of themale on the female's back.D. Crossed Bills: The bills are crossed and themale's wing is extended over the female's back.

A
B
50

51
and he mounted the female (Figure 13B and C) by jumping onto her
back. His feet were placed at the level of the anterior position of
her resting wing. His wings were held extended high over his back.
The male began to lower himself toward the female's cloaca immedi-
ately after mounting. Both held their tail feathers to the side just
before copulation took place. When the male mounted, the female
usually raised her neck out of the water and began to swing back and
forth. The male stayed mounted from five to ten seconds. Immedi-
ately following copulation the male dismounted. As he did, he placed
the wing next to the female over her back and they crossed their bills
(crossed bills; Figure 13D) and performed a pirouette. They then
ran a few steps in this position and parted, each walking a few more
steps with their bills held close to the breast (postcopulatory walk).
Copulations took place all through the prenesting period and
continued for a few days after incubation began. Many species have
displays for attracting mates and to aid the pairing process. Avocets
apparently lack displays of this type. Pairing probably occurred on
the wintering grounds or during migration prior to arrival at Summer
Lake. Pairing on the wintering ground appears to be a simple associ-
ation process in which females associate themselves with males and
persist until they are no longer rebuffed (Hamilton, 1969).

52
Aggressive Behavior
Avocets engaged in both intraspecific and interspecific aggres-
sive interactions. Nearly all intraspecific aggressive action patterns
were involved with maintaining spatial relationships between avocets.
In the winter avocets were gregarious and showed almost no aggres-
sive tendencies. In late winter and early spring aggressive activities
began to increase (Hamilton, 1969). At the time of arrival on the
breeding grounds there was still an oscillation between aggressive-
ness and non-aggressiveness. Ultimately avocets spaced themselves
into territories prior to nesting.
Intraspecific Aggression: Avocets had two threat postures
which were elicited when another avocet approached either on foot or
in the air. One was a crouch (Figure 14A) in which the approached
bird slightly flexed its legs and drew its head back close to its body.
The orientation of this display was usually toward the approaching
bird. The second was a simple raising of the interscapular feathers
while feeding. It seemed to function as a warning to reduce the
incidence of overt attacks (Hamilton, 1969). It was seen most often
in early spring in flocks of feeding birds and it also occurred when
two avocets approached one another near territory boundaries while
feeding.
More overt aggressive encounters occurred regularly and had

53
a somewhat ritualized form. If avocets approached too close to a
pair before territories were established, or invaded a territory later
in the season, the trespassers were immediately driven away. This
was accomplished by either one or both members of the pair pursuing
and chasing away the intruder. In either case the basic action pattern
was similar. The pursuing bird or birds ran rapidly after the ad-
versary with the head retracted somewhat and the body inclined
forward at about a 15' angle. When a single avocet pursued I have
termed it a single crouch-run (Figure 14B); and for a pair, paired
crouch-run (Figure 14C). The latter was the most common form of
pursuit before incubation began. It was most often directed at other
avocets, but was also used toward other species, particularly other
shorebirds. A bird being pursued usually ran or flew from the
scene (retreat; Figure 14B). The retreating bird often ran away
using a side step to keep its body perpendicular to the pursuing bird.
If the retreating bird did not leave quickly it would be charged by a
sudden flying assailment in which the attacker struck with its feet
on the other bird's back. This was accomplished by swooping in
close and kicking out with the feet (pursuit attack; Figure 14D).
In cases where spatial dominance was not clearly held by any
pair, complicated aggressive encounters involving several birds
often occurred. If neither pair retreated they met in a bowing
(Figure 15C) position in which all birds faced one another with

Figure 14, A. CrouchBe Single crouch-run: The bird on the left isdriving the bird on the right out of its territory(Retreating).Co Paired crouch-run: The pair on the left isdriving the other bird away.D. Pursuit Attack: The flying bird above isattacking.
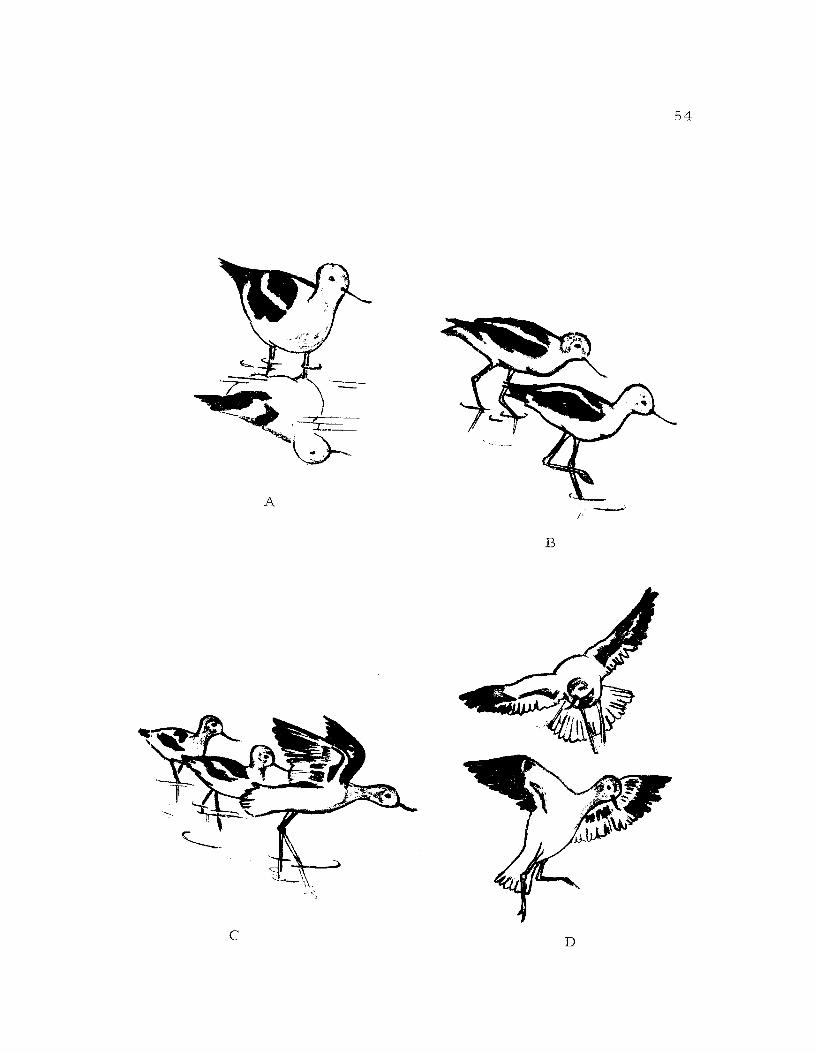
pq
U

55
their bodies tilted forward and their bills near the ground or water.
Mates stayed close together and parallel to one another at all times.
if one pair was dominant, the encounter would go no further, the
dominant pair driving away the others using the paired crouch-run.
Where neither pair retreated, a melee of attacks occurred. One or
both members of a pair would jump at the opponents, either striking
out with a wing or attempting to land on another bird's back (jump
attack; Figure 15A). Immediately after an attack, pair members
quickly moved back to their side-by-side position. A bird trying to
avoid a jump attack either fluttered away or ducked entirely under
the water. If attacks did not occur, several other postures resulted.
Occasionally a bird would squat (Figure 15C) in the water or in some
cases it would squat and place its head in a resting position (pseudo-
sleeping, Figure 15C). One explantaion of these postures is that
they act to reduce aggression or act as appeasement displays. Ap-
peasement displays generally act to reduce fighting or inhibit attacks
and frequently expose some vulnerable area of the body (Manning,
1967). Observations of fighting avocets, however, do not support
the conclusion that these are appeasement displays, since birds in
either posture were often attacked or themselves attacked directly
from this position. A certain amount of pecking in the water, which
may be displacement feeding, also occurred under these stalemate
conditions.

Figure 15. A. Jump Attack: The bird with its wings up isabout to jump at the other bird.B. Wingspread.C. Grouping Display: The two pairs locatedthe right and left are in the bowing posture.Notice the closeness of the members of thepairs. The near avocet to the left is squattingand the near one to the right is pseudosleeping.
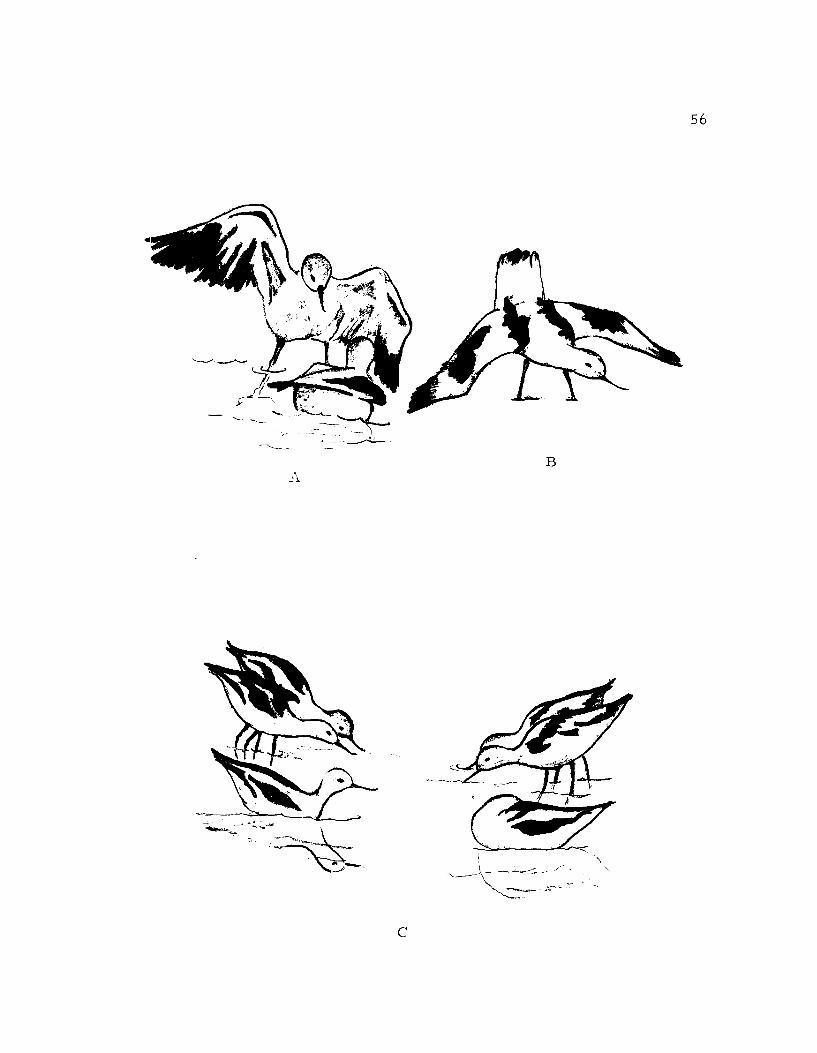
-13
56

57
These group encounters often included more than four birds
and I have observed as many as 12. A large number of birds taking
part suggests that some other function was involved besides spacing.
Birds seldom seek aggressive encounters on their own. There is
usually some advantage in doing so, such as territory establishment
(Mar ler and Hamilton, 1966). When an encounter began, avocets
would fly to the scene to take part, sometimes coming from several
hundred meters away. I feel that these fights have some sexual func-
tion and may act to strengthen the pair bond, but I have no confirma-
tory evidence. Often pairs just involved in aggressive interactions
began courtship activities, indicating that the stimulation generated
in fighting might be carried over as sexual arousal.
Territorial conflicts were usually characterized by single or
paired crouch-runs or a group encounter. However, when two birds
met along a territorial boundary which was not well-defined by land-
marks (such as those in open water), a parallel walk (Figure 18B)
would occur. Both birds walked from 0.5-1.0 m apart along the
territory boundary using a rather stiff deliberate stride with their
necks extended upward. Both frequently pecked at the water. This
usually continued for 20-60 sec and resulted either in both birds
turning and retreating into their own territories or in a series of
jumping attacks followed by retreat.
When avocets reacted to other species they used crouch, single

58
and paired crouch-run or pursuit attack. Most of the season they
were tolerant of non-predatory species such as other shorebirds and
ducks. However, during the early part of the parental care stage
no species were tolerated near the chicks.
Two other aggressive activities were associated with protection
of the nest and egg from predators. One was the predator chase in
which one to many avocets pursued and harassed potential avian
predators such as owls, hawks, and gulls. It included a pursuit
flight and a series of swooping dives at the predator. The harass -
anent persisted until the predator was far from the nest. A loud
alarm call was associated with this activity. If the bird was incu-
bating it returned immediately to the nest. The birds most com-
monly attacked were gulls (Larus californicus and L. delawarensis),
marsh hawks (Cirus and short-eared owls (Asio flamrneus).
The second aggressive action pattern used was the dive,
was directed at terrestrial predators which were in close proximi+7
to the nest. I have actually seen this used only toward humans, but
I suspect that it may also be used against other potential nest preda-
tors. An attacking avocet would swoop at high speed coming very
close to the intruder. I have actually been hit by wing tips as I
banded young in a nest. The flight between dives was characterized
by loud calls.
An inter specific threat display was also sometimes given. An

59
avocet, when suddenly encountering another species, would spread
its wings horizontally, crouch, and extend its neck forward (wing.-
spread; Figure 15B). This display was directed at an opponent and
was often accompanied by short charges. I saw this display directed
toward Killdeer (Chardrius vociferus) and ducks and geese. The
form of the display perhaps functioned to make the avocet appear
larger to an opponent.
Diversionary Behavior
Avocets exhibited several behavior patterns which functioned to
distract potential predators away from the nest or young. These ac-
tivities are those displays most often seen by humans visiting avocet
nesting grounds and have sometimes been misinterpreted in the liter-
ature
The most spectacular of the diversionary action patterns is fhe
tightrope display (Figure 16A). It is probably homologous to the
"broken wing" display of other shorebirds. The avocet held its
wings horizontal to the ground with its neck extended forward. The
tail was fanned and depressed. The bird walked along teetering from
side to side as if it were walking on a tightrope. The aspect of the
wing varied from a horizontal to vertical position. Often the display
was accompanied by a low croaking call.
The sneak-run (Figure 16B) display resembled the aggressive

60crouch-run action pattern, but the context was different. The avocet
ran with its head pulled back next to its body. It usually ran away
from the intruder. The function of this activity is unclear. It may
be similar to the "rodent run" display of some other shorebirds(Williamson, 1 950). To a human it seems to lack much value as a
means of distraction, but, of course, humans do not react in the
same manner as other animals. The crouch-run, tightrope and
breast dipping (Figure 16C) may alternate with one another in a
series of distraction displays. When breast dipping, an avocet ran
or walked in the water, stopping occasionally to bend at the ankle,
thus dipping its breast in the water. Hamilton (1969) feels that the
breast dipping display simulates incubation. This may be the origin
of the display but the fact that I have never seen it performed on land
and that the avocet spends only a second or two in the squatting posi-
tion seems to make fake incubation a doubtful function. Another
possibility is that it simulated an injured bird which was unable to
fly, but repeatedly tried to jump into the air.
When humans and probably other potential predators approached
or disturbed a group of nesting avocets, they were met with alarm
circling. Alarm circling consisted of one to many avocets flying in
circles and giving a loud alarm call. The circles were fairly large
and from 6 to 12 m above the ground. A bird may alarm circle, then
land, perform a tightrope, crouch-run, or breast dipping display,

Figure 16, A. Tightrope,
B. Sneak-run: The bird runs or rapidly walks inthis position when disturbed.
C. Breast dipping.
D. Flying,
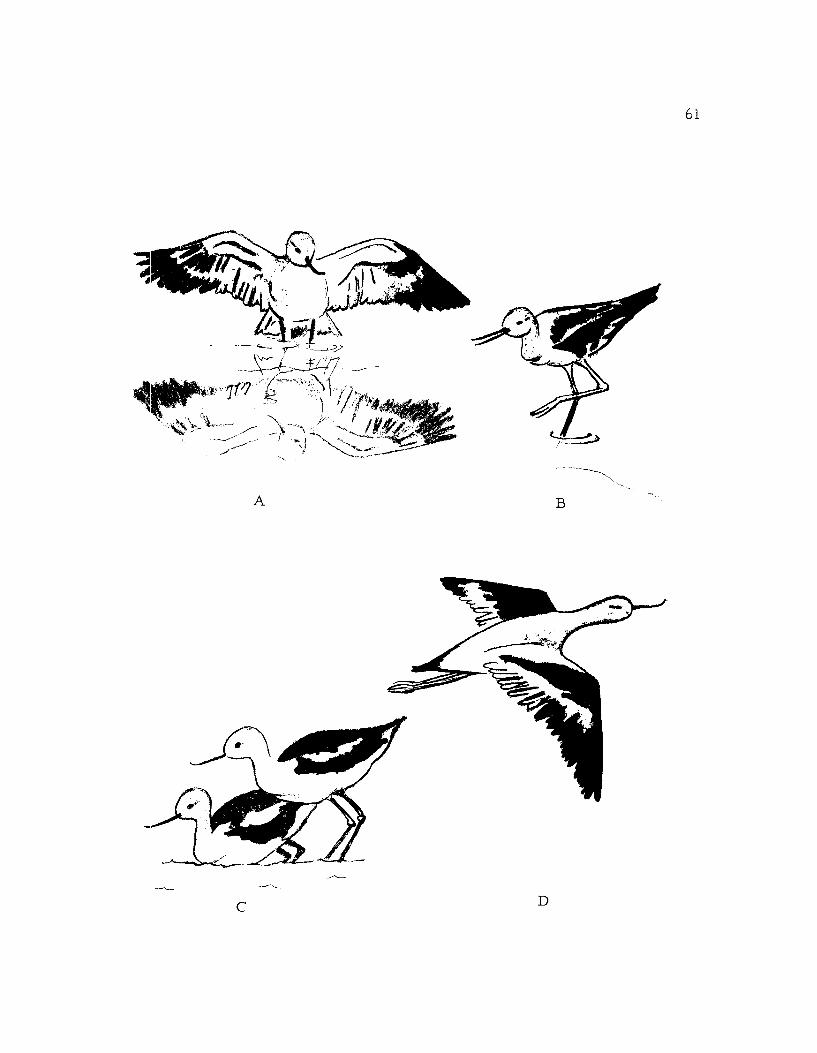
A B
-,,,----
C D
61

62
and circle again. Sometimes a disturbed bird just stood and gave
an alarm call (Alarm calling).
All of the above diversionary activities may be combined in
sequences. Further, up to 50 birds may congregate and perform
distraction displays together. These groups of displaying birds
were usually located away from groups of nests. This activity usu-
ally persisted until the intruder approached close to the nests. Then
the group of displaying birds tended to break up and the pairs whose
nests were most closely approached kept displaying. If the nest was
found, they resorted to diving at the predator.
Nesting Behavior
Under nesting behavior I have included all activities associated
with selecting a nest site, nest building, egg laying, and incubation.
Avocets beg--In searching for a nest site even before they seleet-
ed a territory. Indeed, choice of a nest site may be an important
factor in determining the location of a territory. Although the nest
and the foraging territory (see Breeding Biology) may not be in close
proximity, the placement of the feeding area may be dependent on
having a suitable nesting area nearby. When seeking a nest site,
mates acted together by slowly walking over possible terrain with
their bills placed close to the ground (ground checking; Figure 17B).
They often stopped for varying lengths of time still performing this

63
activity. A characteristic vocalization usually accompanied this ac-
tivity and it seemed that the intensity of the calling was related to
the suitability of the site they were investigating. Ground checking
could change almost imperceptibly into throwing straws (Figure 17A)
in which the birds picked up pieces of grass and other materials in
their bills and tossed them toward the presumed nest site with a
sideways flip of the head. Hamilton (1969) states that the material
is tossed over the bird's back. This can occur on occasion, but it
is more common for items to be tossed sideways.
Throwing straws often led to scraping (Figure 1 7B) by which
the nest scrapes were hollowed out. The avocet rested on its breast
and hollowed out the nest scrape with its feet. Its wings were held
loosely at its side. Hamilton (1969) and Brown (1949) state that the
scrape is made with t the breast. My observations disagree, since
there were obvious scratch marks in newly formed scrapes. Also
scrapes were often found in substrate too hard to have been hollowed
out with the breast alone. Further, the avocets I watched seldom
rocked or rotated on their breast as Hamilton and Brown indicate.
Sometimes many scrapes were made before a final nest site was
chosen. The birds tended to return to favored spots many times
before the eggs were laid; however the final nest site was sometimes
different from any of the "favored" spots. I suspect that the nest
can be built in just a few minutes and the spot and time that the final

64
nest is built may be largely determined by the necessity for the
female to lay. Materials were added to the nest by throwing straws.
A great deal of material could be added to the nest in a short time in
this manner, since a nest could be built up several centimeters over-
night during a flood. Normally nests were built up slightly during
incubation.
Egg laying was observed in only one case. The female was
sitting on the nest. She suddenly stood up and began giving an excited
call similar to that used during ground checking. The male rushed
over and both called for several minutes. When I checked the nest
one egg was present; there had been no egg in the nest 30 minutes
earlier.
Once the eggs were laid and incubation had begun, both sexes
tended the eggs. An incubating (Figure 17D) bird sat on the nes with
its head held upright or in a resting position. During hot weather the
bill was held wide open and gular panting was evident.
An avocet relieving its mate at the nest went through a nest
approach (Figure 17C) ceremony in which it walked in a slow jerky
fashion while occasionally extending a leg posteriorly and shaking it,
As the bird approached the nest it often began to throw straws. At
this point, the incubating bird usually left the nest and began to throw
straws also. The relieving bird then moved to the nest and straddled
it and placed its bill among the eggs. The avocet erected its breast

Figure 17, A. Throwing Straws: The birds are picking uppieces of grass and tossing them with a flip oftheir heads.B. Scraping: The bird sitting on the ground isscraping. The other is ground checking.C. Nest approachD. Incubation
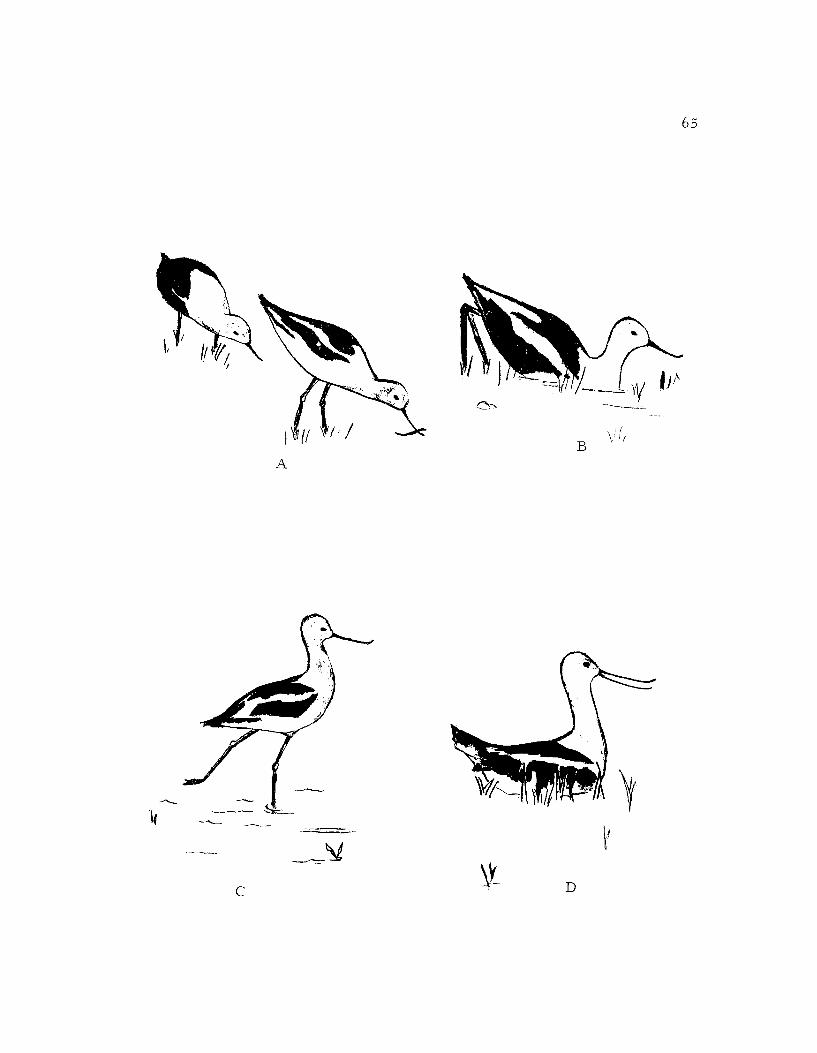
A
65
\B
if

66
feathers and settled on the nest. It then used its legs and feet to
rotate the eggs. Head bobbing sometimes also occurred during the
nest relief ceremony.
Parental Behavior
Avocets were very aggressive while caring for their young.
They excluded all species from the territory around the chicks.
Many aggressive action patterns suchas single crouch-run, pursuit
attack, crouch, parallel walk and wingspread were used.
The young were brooded (Figure 18C) during the first two weeks.
The parent rested on its tarsi and the chicks stood against the breast.
The adult's wings were usually drooped slightly to provide extra pro-
tection for the young. At least one parent was always in attendance
of the chicks. It usually stood neaz by, usually in the taaright (Figure
18A) posture, which is an alert posture. The parents relieved each
other at intervals just as incubating birds did, but there seemed to
be no ceremony involved.
When the nest was located away from a feeding area or the
chicks were in an area prone to disturbances, the parents moved
the chicks. This was done by leading, where both parents walked
ahead of the chicks while continually calling to them. If the adults
were particularly excited they would run or fly ahead, calling, and
then come back to the chicks in an attempt to get them moving.

Figure 18, A. Upright: This bird is standing with its head
extended upward.
B. Parallel walk These two males are walking
side by side in a territorial boundary conflict,
C.
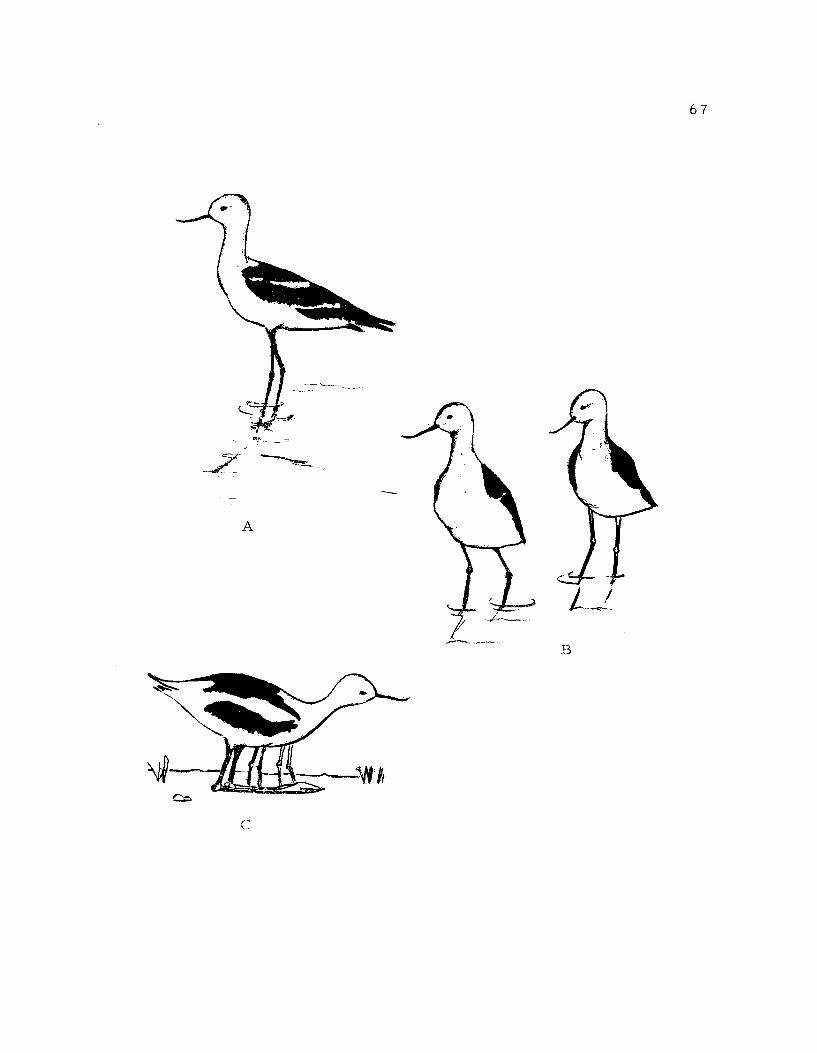
Skt.
A
B
67

68
The parents also had alarm calls which warned the chicks of
danger. The chicks ran for cover and laid flat on the ground upon
hearing the parent's call.
Miscellaneous Action Patterns
In any functional categorization of behavior some action patterns
are difficult to classify or do not fit neatly into any functional group-
ing. The upright posture is one of these. It occurred at all times
of the season and at almost any time A bird in the upright posture
stood or walked with its head and neck extended vertically. This
action pattern seemed to be used when a bird was disturbed or on
the alert.
Another display which does not fit well into any category is
head bobbing, in which the head is pumped up and down rapidly. It
sometimes was used in nest exchange ceremonies, some aggressive
encounters, or when the bird was disturbed in some way, as when a
human approached.

69
THE TIME BUDGET ANALYSIS
Time budgets express the manner in which animals partitiontheir time among various activities. Time budgets can be expressedin two basically different wars. One is a frequency-based time bud-get, the other time-based. The frequency type may give a record ofwhat a bird has done during a one minute or five minute time interval,for example. The result is a sample of the bird's activities throughtime. The frequency with which an animal performs various activitiesduring the time interval may also be recorded. For example, thenumbers of preening movements in an hour or day could be tabulated.In either case, the activity is measured as a frequency pei time inter-val, but the actual amount of time devoted to various activities cannotbe determined.
In the second method, the actual amount of time spent in eachactivity is recorded. It can be expressed as a total time observedfor the activity; for example, if 20 hr, were spent collecting data,the amount of time spent preening might be 30 min.out of the 20 hr.(see Verbeek, 1964). Another approach is to express the time spentin each activity as a percentage of the total observation time. Thelatter method offers the most meaningful representation and it isnot greatly influenced by the length of the activity period of the birds.It also has sufficient abstraction to allow comparisons between stages

70
of the breeding cycle.
I have utilized a combination of the frequency and time ap-
proaches. By selecting a small unit of time (10 sec.) in which to
sample behavior I have essentially measured the total time spent in
each activity. This has allowed me to express the avocet's time
budget as a percentage of the total time available to the bird.
I will consider both seasonal and diurnal time budgets, drawing
inter-sexual comparisons in each case. Proper seasonal and diurnal
apportionment of time requires different strategies and each is acted
upon by slightly different selective pressures, thus demanding that
each be considered separately. Further, many species exhibit very
different male and female roles in their social systems, and thus
examination of sexual differences in the seasonal and diurnal time
budgets is also important.
Time apportionment data for 40 action patterns (Table 5) of
both sexes for both seasonal and diurnal patterns are presented in
Appendices 1 and 2. In order to analyze the time budgets more
meaningfully I have grouped action patterns according to their gen-
eral functions (Table 6).
Behavioral Categories
Several of the categories presented in Table 6 require further
explanation. The maintenance activities have been divided into four

71
Table 6. Categories of behavior used in the time budget analysis
Category Included Action Patterns
Feeding feeding
Resting resting
Preening preening
Other Maintenance bathing wing-leg stretchscratching wingshaketwo-wing stretch drinking
Aggressive Behavior crouch bowingsingle crouch-run squattingpaired crouch-run pseudosleepingpursuit attack parallel walkjump attack wing spreadretreat
Diversionary Behavior
Mating Behavior
predator chase breast dippingtightrope alarm circlingcrouch-run alarm calling
neck extendedprecopulatory preeningmountcrossed billspostcopulatory walk
Nest Building Behavior ground check ingscrapingthrowing straws
Incubation incubation
Upright upright
Parental Care broodingleading
Absent out of view

72
categories. This was done because of the relatively large amount
of time spent in feeding, resting and preening. The aggressive cate-
gory consists of those action patterns most consistently used in intra-
specific encounters. Most interspecific behaviors are used in attack-
ing and fending off predators; therefore they are included in the
diversionary category. The nest building classification consists
of those activities associated with selection of a nest site and build-
ing a nest. Incubation is included as a separate category. The
absent category represents the time when an avocet was lost from
view for more than a few seconds during an observation period.
This category was necessary to maintain continuity during observa-
tions.
The Seasonal Time Budget
The seasonal time budget consists of a representation of the
time spent in each category of behavior for each stage of the breed-
ing cycle. I have combined the data for both sexes for this section
of the analysis. The time budget is graphically represented in
Figure 19 and a tabular presentation of the same data is included
in Appendix 3. The apportionment of time into each category of
behavior will be discussed below with emphasis on the possible eco-
logical and behavioral bases for the patterns found.
Feeding (Figure 19A): From an ecological point of view, the

Figure 19. (A-K). The seasonal time budget (data for bothsexes combined) expressed as the percent oftotal observation time spent in each category ofbehavior. The stages and sample sizes are:Stage 1 Prenesting 4298.0 min.Stage 2 Incubation 18159. 4
Stage 3 Parental Care 2362. 7
Stage 4 Po stbr eeding 276 7. 1
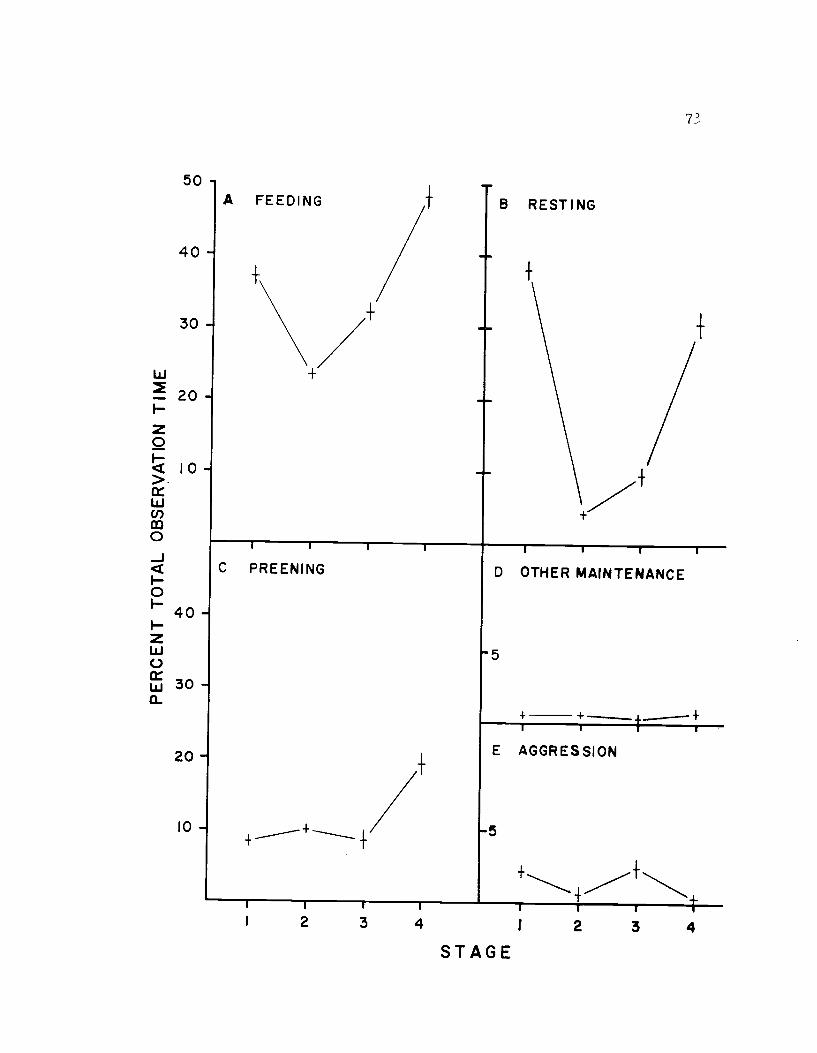
50A FEEDING
40 -
30
0<h-
- 20-I-
I 0 -
w>
030.J
C PREENING D OTHER MAINTENANCEO
40 -
B RESTING
I-
w -5
w 30 -
20 -
10 -
73
E AGGRESSION
-5
I 2 3 4 2 3 4
STAGE
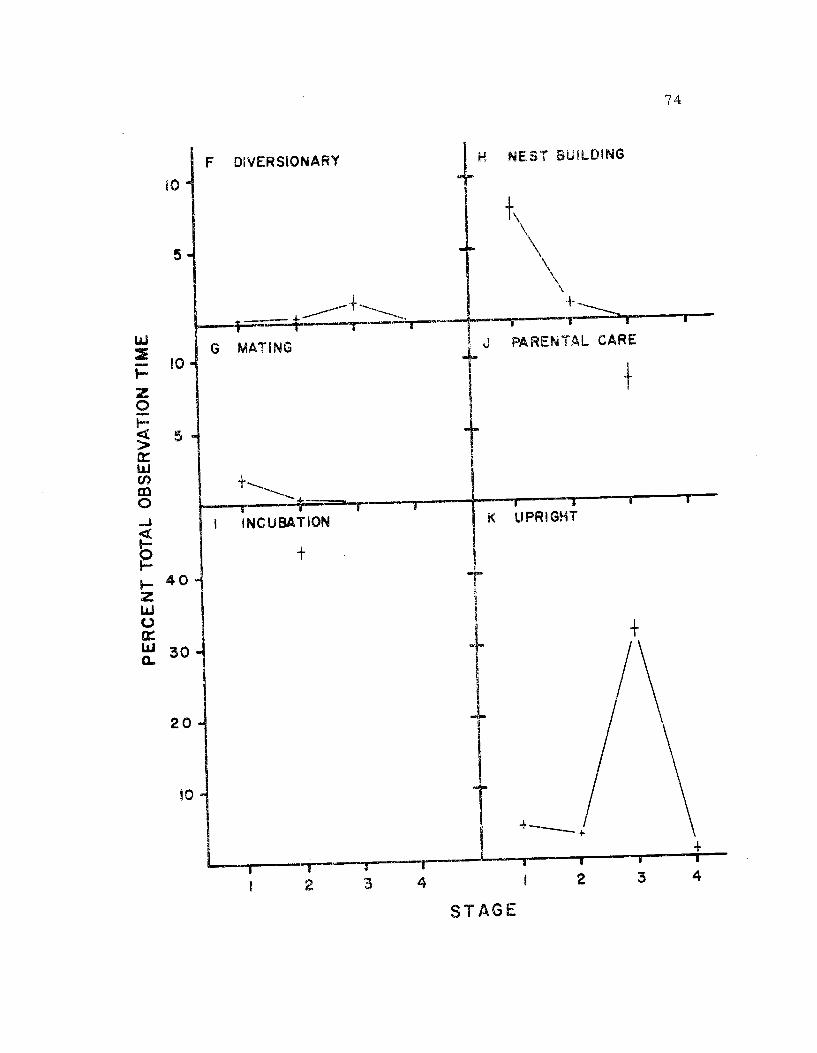
DIVERSIONARY NEST BUILDING
74
G MATING J PARENTAL CARE
UPRIGHT
4
4
STAGE
2 3 4

75
amount of time spent feeding is probably the most interesting and
possibly one of the most important aspects of a bird's time budget.
The expenditure of time in foraging in the four stages of the breeding
cycle is significantly different. There are several possible explana-
tions for these differences, each relating to avocet energy require-
ments. This aspect of the time budget will be considered more fully
in the Discussion.
Resting (Figure 19B): The percentages of time spent resting
through the breeding cycle had a pattern similar to that found for
feeding. The low proportion of resting time in Stage 2 is probably
deceptive. The birds were spending about 43 percent of their time
incubating during this stage. In some large species, such as the
Willow Ptarmigan (Lagopus lagopus) and Ring Dove (Streptopelia
risoria), incubation does not increase metabolism over the resting
level (West, 1968; Brisbin, 1969). Combining rest and incubation
activity for Stage 2 gives a total of 47 percent of the time spent in
an "inactive" state. In Stage 3 only 90 5 percent of the time was
spent in resting. There was, however, a large amount of time
spent in the upright posture, which involves merely standing with
the head and neck extended upward. If upright is considered to be
relatively close to resting in its energy requirement, then the actual
"inactive" time in Stage 3 is close to 41 percent. If this line of
reasoning is correct, avocets at Summer Lake appear to spend a

76
rather constant amount of time through the breeding cycle in an
"inactive" state.
Preening (Figure 19C): The percentage of time spent in preen-
ing remained essentially constant through the first three stages of
the breeding cycle, increasing in Stage 4. Care of the body surface
probably required the same amount of time throughout the breeding
season, but with the onset of post-nuptial molt (Stage 4) there was
an almost two-fold increase in preening time.
Other Maintenance Activities (Figure 19D): Very little time
was spent in the remaining maintenance activities, and there was no
seasonal fluctuation in these activities. Body maintenance should be
constant throughout the breeding season since the body needs continual
care regardless of the time of year.
Aggressive Behavior (Figure 19E): Aggressive activities took
very little of the avocet's time (a maximum of only 2. 4 percent in
Stage 3). Despite this, a casual observer of avocet behavior undoubt-
edly would conclude that they spend a great deal of time in aggressive
activities. This impression stems from the pattern of distribution
of aggressive activities in time. During Stage 3, 18 min/day were
spent in aggressive activities. Each encounter lasted approximately
10 sec; thus an avocet was involved in about 92 aggressive encounters
per day, or about one every 7.5 minutes. Thus while the frequency
of aggressive actions is relatively high, the time expenditure is not.

77
A time budget gives the more realistic view of how much time was
actually spent in aggression. However, behaviorally, the frequency
at which an action pattern occurs may be functionally more important
than the time actually spent on the activity. For example, it may be
more adaptive for avocets to spread 18 min. of aggressive activity
throughout the day rather than concentrate it in one short time span.
On the other hand, energetically, avocets may not be able to efficiently
spend more than 18 min, on aggression.
The greatest amount of time spent in aggression was during
Stages 1 and 3. This was associated with territorial establishment
in Stage 1 and parental care in Stage 3. Aggressive activity was low
in Stage 2, possibly because the territorial system was stable and
approximately half the birds in the population were incubating and
thus not free to engage in aggressive encounters.
Diversional_2yBk-havior (Figure 19F): An extremely small
amount of time was spent in diversionary activities and that only in
Stages 2 and 3. These stages include incubation and parental care.
Protection of eggs and young may contribute to the appearance of
diversionary activities during these times. The small amount of
time spent in these activities may be due in part to the rigorous
control of predators on the management area.
Mating Behavior (Figure 19G): Mating activities took about 2
percent of the avocet's time in Stage 1. A typical courtship sequence

78
took about 2 min. to complete. Thus, on the average, a pair copulated
or attempted copulation about seven times a day through the month
duration of Stage 1,
Nest Building Behavior (Figure 19H): A fairly large portion
of time was allotted to nest site selection and nest building during
Stage 1. From field observations I suspect that avocets can build
an adequate nest for the laying of the first egg in a few minutes.
They spent most of their nest building time,however, making numer-
ous scrapes and adding lining to some before the first egg was laid.
I doubt that the sole function of this nesting behavior is to build a
nest; strengthening the pair bond may be an important function as
well. The nest building activity which occurred in Stage 2 was asso-
ciated with maintaining the nest structure.
Incubation and Par ntal Care (Figure 19 I -J) I have combined
these two categories here because both are used to care for objects
of the reproductive effort. Incubation took 43.5 percent of the time
in Stage 2 while parental care required 8.4 percent in Stage 3. How-
ever, this does not accurately represent the time spent caring for
the young. The Upright position was used a great deal while watch-
ing over the chicks. This took 31. 7 percent of the time in Stage 3.
If upright is included as part of parental care, this increases paren-
tal care to 40.1 percent of the total time, which is similar to that
spent in incubation. Therefore, care of the eggs and chicks took

79
about the same amount of time.
Upright (Figure 19K): I feel that the upright posture is essen-
tially an alert position assumed when an avocet is watching something
or is wary. It is an indication of the general alertness of the birds
during different stages of the breeding cycle. The birds were most
alert when caring for the young. In Stages 1, 2, and 4 the time spent
in this activity went from 4.5 percent to 1.0 percent. This pattern
is as one might expect. In Stage 1, when the territory was being
established, the birds were comparatively more alert; in Stage 2
the territory was less rigorously defended, while in Stage 4 there
were no territorial activities. Also, a certain undetermined amount
of time spent in the upright posture was due to my being close to the
birds.
Absent (Appendix 3): I only recorded the absent category in
Stages 2 and 3. It represents the time the avocets under observation
were not in my view during an observation period. This accounts for
9 percent of the time in Stage 2. Part of this was due to the bird's
feeding on a territory located some distance from the observation
tower, thus making them difficult to observe.
Sexual Differences in the Seasonal Time Budget
The percentages of time that males and females spent perform-
ing the basic categories of behavior (Table 6) are presented in

80
Figure 20. An asterisk has been placed above each male-female
comparison which shows a significant difference (P <.05). These
data are tabulated in Appendix 4.
A survey of Figure 20 indicates that there are only seven male-
female comparisons which show statistically significant differences.
There are two possible reasons for the small number of differences:
the data may be insufficient to determine if the differences observed
are real, or there may actually be no differences between the sexes.
I suspect both are right, in part. Five of the seven significant differ-
ences observed occurred in Stage 2, where the most data are avail-
able. There is a possibility that with more data for the other stages
there would be more significantly different comparisons. In contrast,
considering the generally similar behavior patterns of the sexes, the
lack of time budget differences is not surprising.
I will not rely heavily on statistical evidence for real differences
because statistical methods do not always reveal biological signifi-
cance. For example, there may be sexual differences in the time
budget which can be shown to be adaptive but are not statistically
significant, or the opposite situation might occur. Therefore, I will
attempt to explain differences between the sexes when there are
plausible explanations.
Feeding (Figure 20A): The male consistently spent more time

Figure 20. (A.K). The seasonal time budget for both sexes,expressed as the percent of the total observationtime spent in each category of behavior by eachsex. Stages are as in Figure 19, Sample sizes(min. ):
Stage 1 23 21 . 0 2076. 7Stage Z 10565, 7 7594. 2Stage 3 1189.7 1173.0Stage 4 13930 5 1373.6
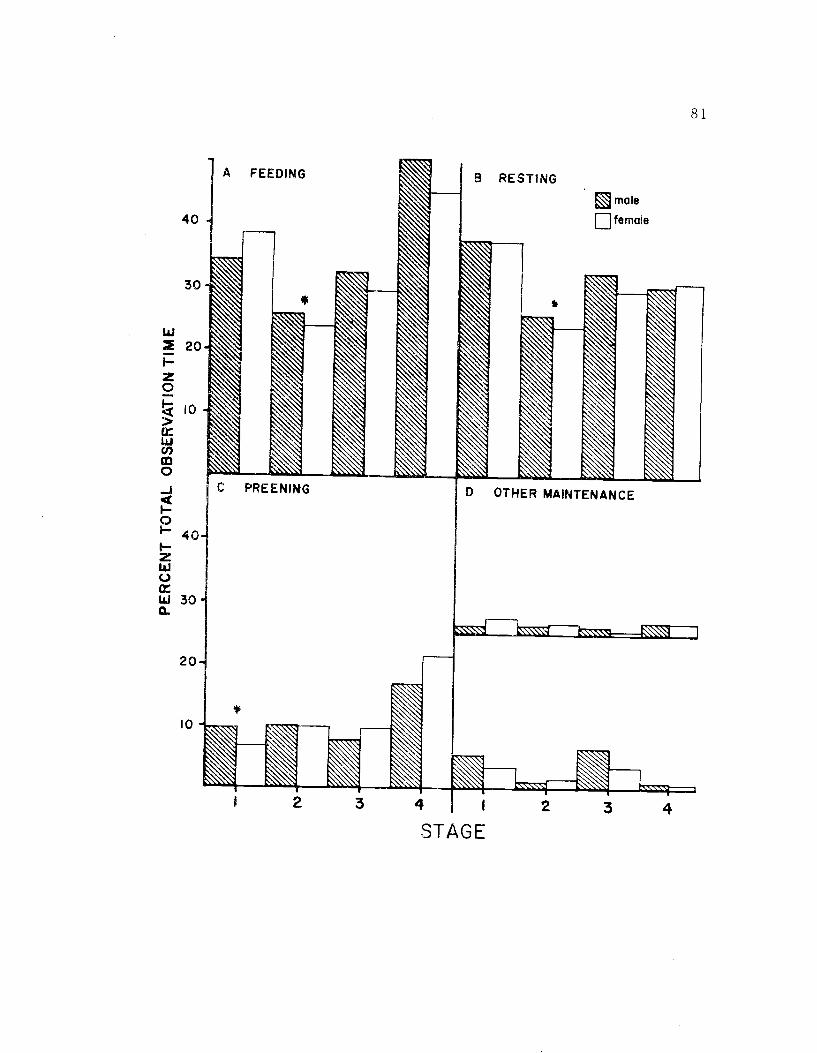
FEEDING
40
30
20\
B RESTING
male
El female
10
40
30
20
10
PREENING
111 011INNIU\
OTHER MAINTENANCE
4
STAGE
81
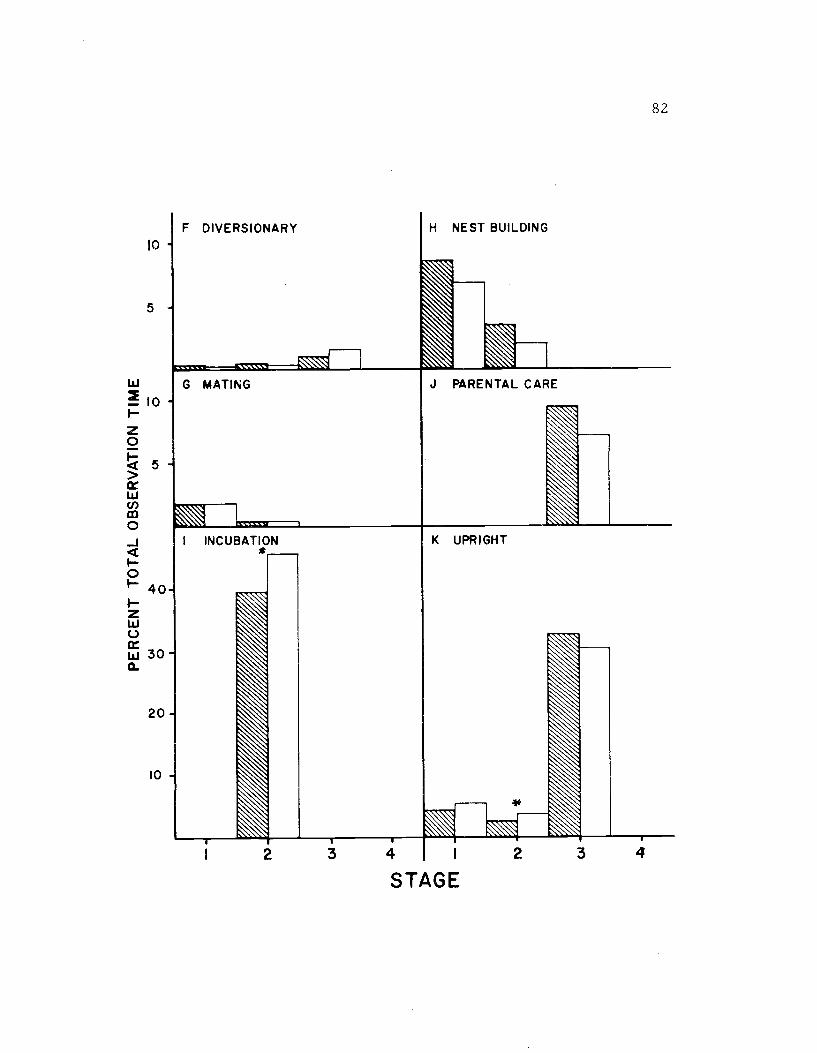
10 -
W
- I -1--
z0F:< 5>CrLUcr)co0I<II 40-Izw0cew 30a_
F DIVERSIONARY
rwrrTini---41. li----MllG MATING
20 -
I0
INCUBATIONI
H NEST BUILDING
82
PARENTAL CARE
1 2
K UPRIGHT
03 4 1
STAGE2 4

83foraging than the female in all stages except Stage 1. The female
may have needed to feed more in Stage 1 because she was producing
eggs, which is an energetically expensive process (El-Wailly, 1966;West, 1968). The reason why the male fed more than the female in
the other stages is unclear.
Resting (Figure 20B): The sexual differences for time spent
in resting are small in every stage of the breeding cycle. The Stage
2 comparison demonstrates a statistical difference. The female may
have rested more in the early phases of Stage 2 because she was
recovering from egg production.
Preening, Other Maintenance Activities, and Aggression (Fig-
ure 20 C-E): There was little difference between the sexes for these
activities.
Nest Building Behavior (Figure 20H): The male spent slightly
more time ground checking, throwing straws, and scraping than thefemale. The sequence of action patterns involved in nest building
may account for this difference. A pair searching for a nest site
usually ground checked together, but when scraping began the male
tended to scrape while the female stood close by in an upright pos-
ture. This seems to be adaptive since the scraping position is
awkward making quick reaction to avian predators difficult.
Incubation (Figure 201): The sexes shared incubation duties,
but not equally. The female spent about 6.5 percent more of her

84
time on the nest than did the male. The difference, however, was
not distributed evenly through the incubation period (Table 7). The
male incubated more during the first third and the female more dur-
ing the latter two-thirds.
The Diurnal Time Budget
The diurnal time budget was partitioned into six 2-hr periods
from 0600-1800. The percent time spent in each category of behavior
for each period is given in Appendices 5 and 6. The seasonal changes
in the diurnal time budget (sexes combined) are presented in Figure
21,
Feeding (Figure 21A): The diurnal distribution of feeding ac-
tivity was bimodal in all stages but Stage 3. Avocet feeding peaks
occurred in the morning and afternoon. The magnitude of the, morn-
ing peak is not really known because I do not know whether the amount.
of time spent feeding was rising to a high point in Period 1 or whether
it was decreasing to that point. The increase in feeding in the after-
noon was dramatic in Stages 1 and 4 and much more muted in Stage 2.
The morning trough in the feeding corresponds closely to the peak
of resting (Figure 21B). The muted peaks seen in Stages 2 and 3 are
probably related to the effects of incubation and parental care. A
constant amount of time was spent incubating throughout the day
(Figure 21 1). This forced the birds to modify their feeding pattern
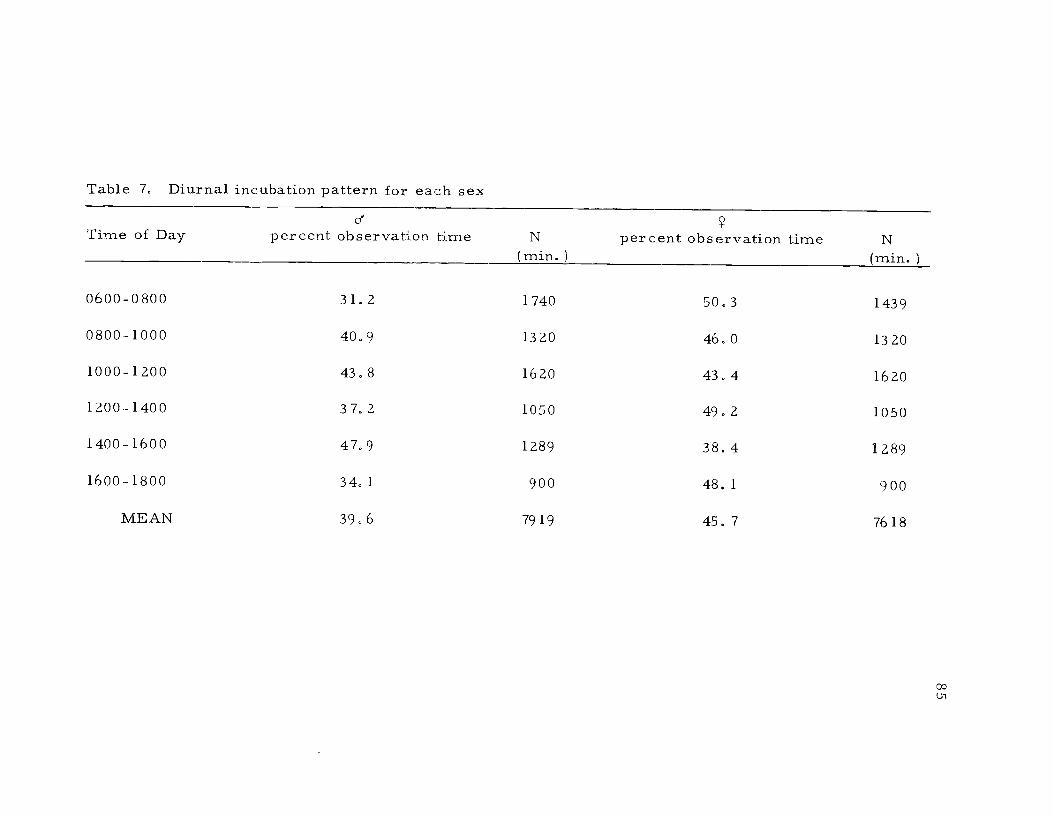
Table 7. Diurnal incubation pattern for each sex
Time of Day percent observation time N(min. )
percent observation time(min. )
0600-0800 3L2 1740 50.3 1439
0800-1000 40.9 1320 46.0 1320
1000-1200 43. 8 1620 43.4 1620
1200-1400 37.2 1050 49.2 1050
1400-1600 47.9 1289 38.4 1289
1600-1800 34.1 900 48. 1 900
MEAN 39.6 7919 45.7 7618
Figure 21, (A-K). The diurnal tim e budget for the four breeding cycle stages expressedas the percentage of the total observation time spent in each category ofbehavior for each of six diurnal time periods (both sexes combined). Stagesare as in Figure 19.
0600-0800
Sample sizes (min):0800- 1000-1000 1200
1200-1400
1400-1600
1600-1800
Stage 1 614. 6 1137.1 776.8 539.5 716. 7 693.2
Stage 2 3060.9 2572.6 3215.8 2088.5 2554.9 2188.0
Stage 3 468.2 60.0 414.3 297.3 118.4 982.6Stage 4 177.6 496.2 472.5 423.2 599.8 478. 3
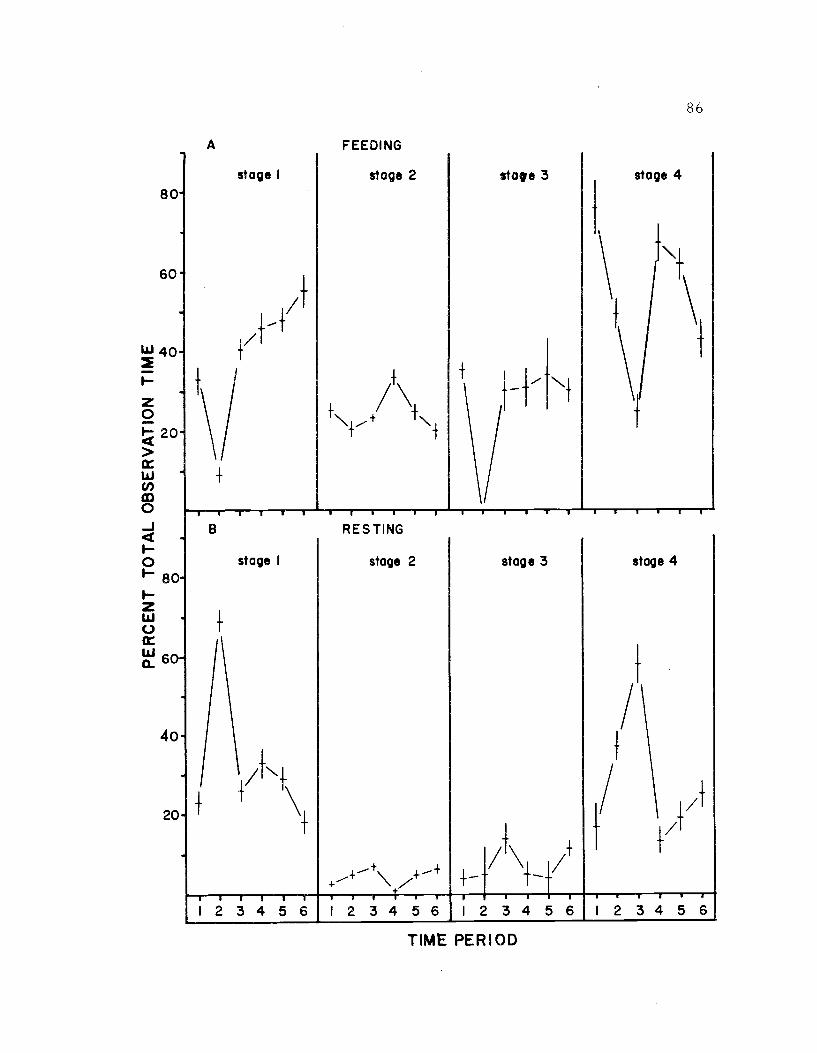
80
60.
40-
I017-: 20
m011OI 801zcrIL
20
A FEEDING
stage I stage 2 stage 3 stage 4
\-[
86
B RESTING
stage I stage 2 stage 3 stage 4
I 2 3 4 5 6 I 2 3 4 5 6 I 2 3 4 5 6 2 3 4 5 6TIME PERIOD
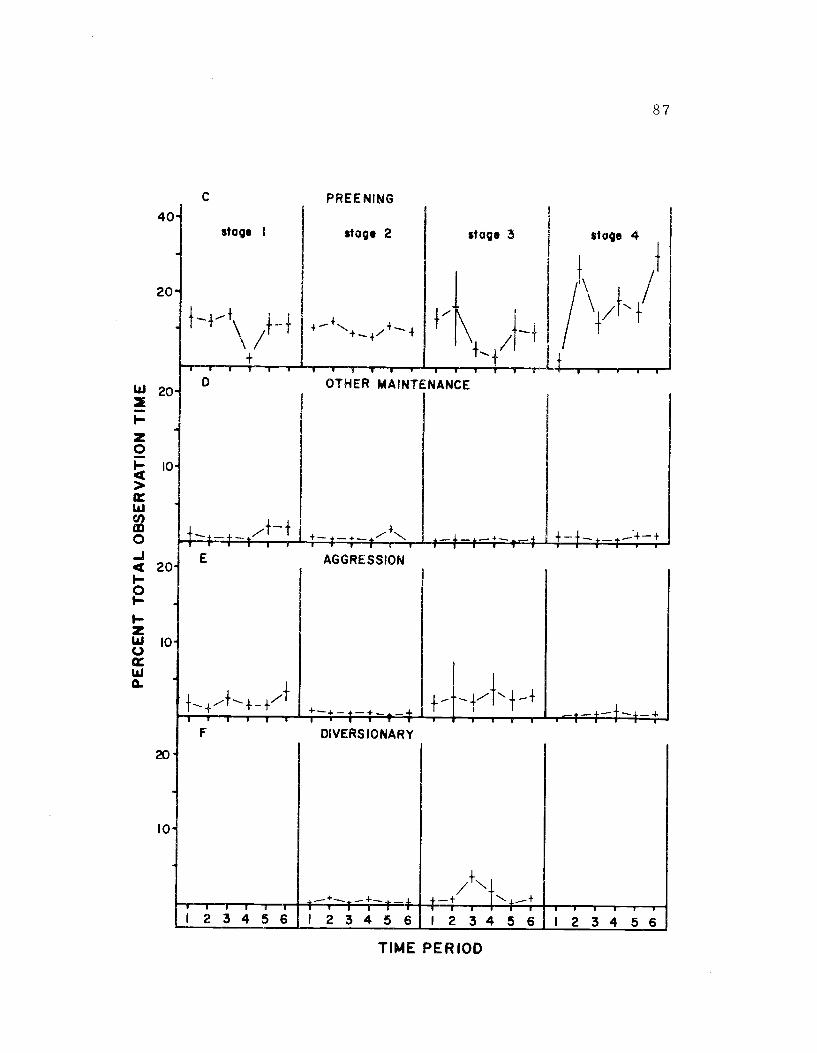
C PREENING
stags I stags 2 stage 3 stage 4
87
OTHER MAINTENANCE
AGGRESSION
DIVERSIONARY
TIME PERIOD
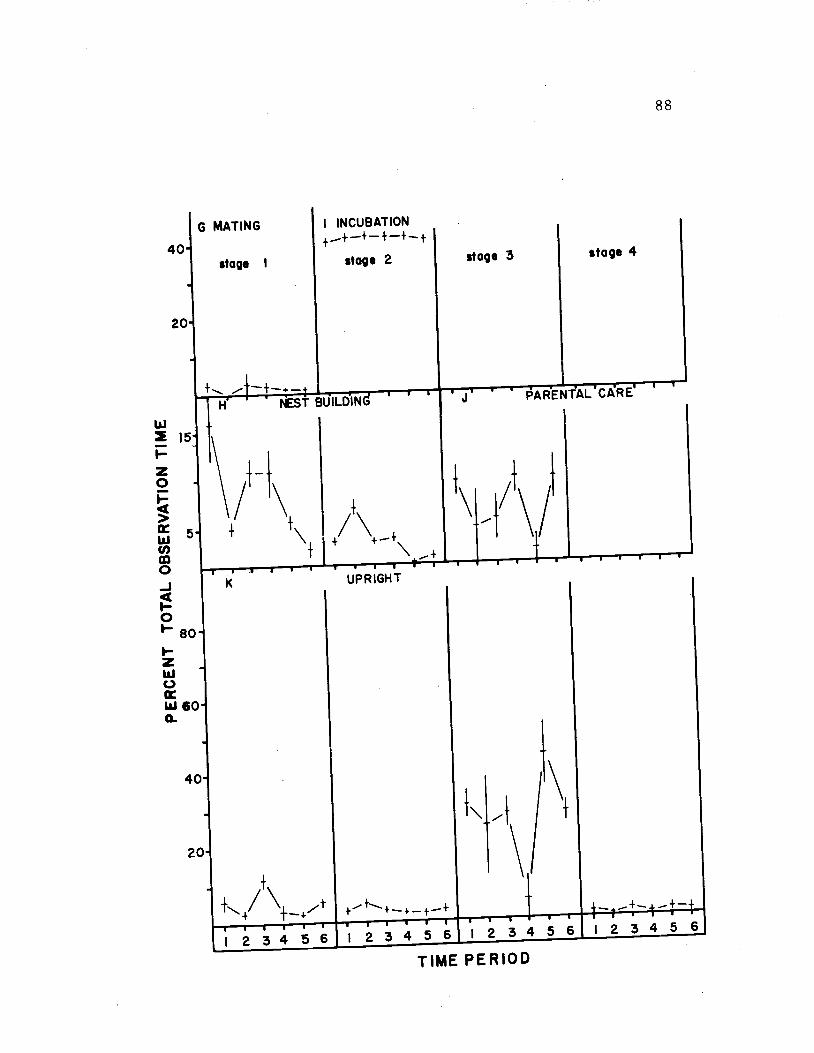
40
20
W2 15p:
z0i=aaxWcocoO-JI-0I 801zWC.)crW 60a.
40
20
G MATING
stage I
+-..... ..."'
I INCUBATION4.--+++-4t
stage 2
. ,NEST BUILDING
K
/+\+---4
stage 3 stage 4
88
// \.\t
UPRIGHT
,4-..... + --- 4. _ + 1.
OAR'ENYAL CA'REI
I 1
1 2 3 4 5 6,
1 2 3 4 5 6 I 2 3 4 5 6. "I
+I 2 3 4 5 6
TIME PERIOD

89
around incubation.
Resting (Figure 21B): Nearly all the resting done by avocets
occurred during a 2-4 hr period in the morning. This peak was not
as evident in Stages 2 and 3. Resting increased somewhat in the
afternoon in Stages 1, 3, and 4. An explanation of the adaptive na-
ture of this resting pattern will be offered in the Discussion section.
Preening (Figure 21C): The percentage of time spent preening
also showed a bimodal distribution around the morning rest period.
The amount of time spent in activities such as preening is probably
dictated by higher priority behavior patterns such as feeding, resting,
and incubation.
Other Maintenance Activities (Figure 21D): The most obvious
feature of this category is the lack of diurnal pattern. The reason
for this is simple. Activities making up this category are such
things as scratching, stretching, drinking, and bathing. These
occur in response to stimuli which may occur at any time.
Aggression (Figure 21E): One would expect aggressive encoun-
ters to be most frequent when avocets are the most active. The data
indicate this to be the case.
Diversionary Behavior (Figure 21F): Avocets only defended
against potential predators during Stages 2 and 3. The diurnal pat-
tern of diversionary activities was probably closely related to the
activity patterns of other species, since avocets reacted to potential

90
predators only when they came near the nest or young.
Mating Behavior and Nest Building (Figure 21G-H): Mating
activities took place predominantly in the morning but were never
observed during the morning rest period. A basically similar pattern
was apparent for nest building. Perhaps during Periods 1 and 3 ac-
tivities such as feeding were not taking a large amount of time during
the morning leaving time for mating and nest building (see Discussion).
The nest building in Stage 2 was concerned with maintaining the nest
and was usually done during nest exchanges. This accounts for the
lack of diurnal variation.
Incubation and parental care (Figure 21 I, 3): As would be ex-
pected, there were no diurnal changes in incubation, since the eggs
were covered almost continually. There was a slight tendency for
the proportion of time spent in parental care to decrease as the day
wore on.
Upright (Figure 21K): There was very little diurnal fluctuation
for the upright posture during Stages 1, 2, and 4. The posture is
used in this stage to watch over the young; the young, like the adults,
tend to feed more in the afternoon. The increased watchfulness may
reflect the need for a closer watch on the chicks as they feed. This
peak may also be a reaction to the increased activity of other species
in the afternoon.

91
Sexual Differences in the Diurnal Time Bud et
A survey of the sexual differences in the diurnal time budget
reveals that the sexes have similar budgets (Figure 22). These data
are tabulated in Appendix 6. Of 180 possible comparisons only nine
were significantly different (P <.05). Considering that the sexes
had very similar seasonal time budgets, this result is not surprising.
The small sample sizes for many time periods and the scat-
tered nature of the statistically significant differences make interpre-
tations difficult. The sexual differences in the diurnal pattern of
incubation are interesting, however. The female incubated more than
the male in the morning and they tended to alternate incubation
duties during the afternoon (Figure 221), There are two possible
explanations for the afternoon alternation. It could represent a
response to heat stress, since incubating avocets were exposed to
high temperatures (> 370 C) in the afternoons. Frequent breaks
from incubation duties could have helped reduce the heat stress.
Another possible explanation is associated with optimal feeding
times. There may be advantages to feeding predominantly in the
afternoon (see Discussion). By alternating incubation duties often
in the afternoon both sexes could exploit this situation. The feeding
pattern in Stage 2 had a similar but opposite pattern of alternation,
suggesting that feeding and incubation patterns are interrelated.
Figure 22. (A-K) The diurnal time budget for the four breeding cycle stages, expressed as the
percentage of the total observation time each sex sp ent in each category of behavior
for each of six diurnal time periods. Stages are as in Figure 19 and time periods as
in Figure 21. Sample sizes (min):
Stage1 2 3
Period4 5 6
300.1 320.2 519.9 569, 4 389.8 389. 7 299.4 269, 2 359.1 363, 1 359.6 365. 6
2 1639.4 1438.6 1469. 1 1319.0 1622.8 1619.6 11049.3 040.1 1288.6 1286.6 1299.2 899. 5
3 239.5 239.7 29.9 29.8 209.4 208. 5 149.9 1 49. 8 58.0 59.0 496, 6 499. 1
4 90, 6 90, 0 299.3 300.5 2 42.3 240.7 213.0 209.5 299.9 299.9 239.7 239.4
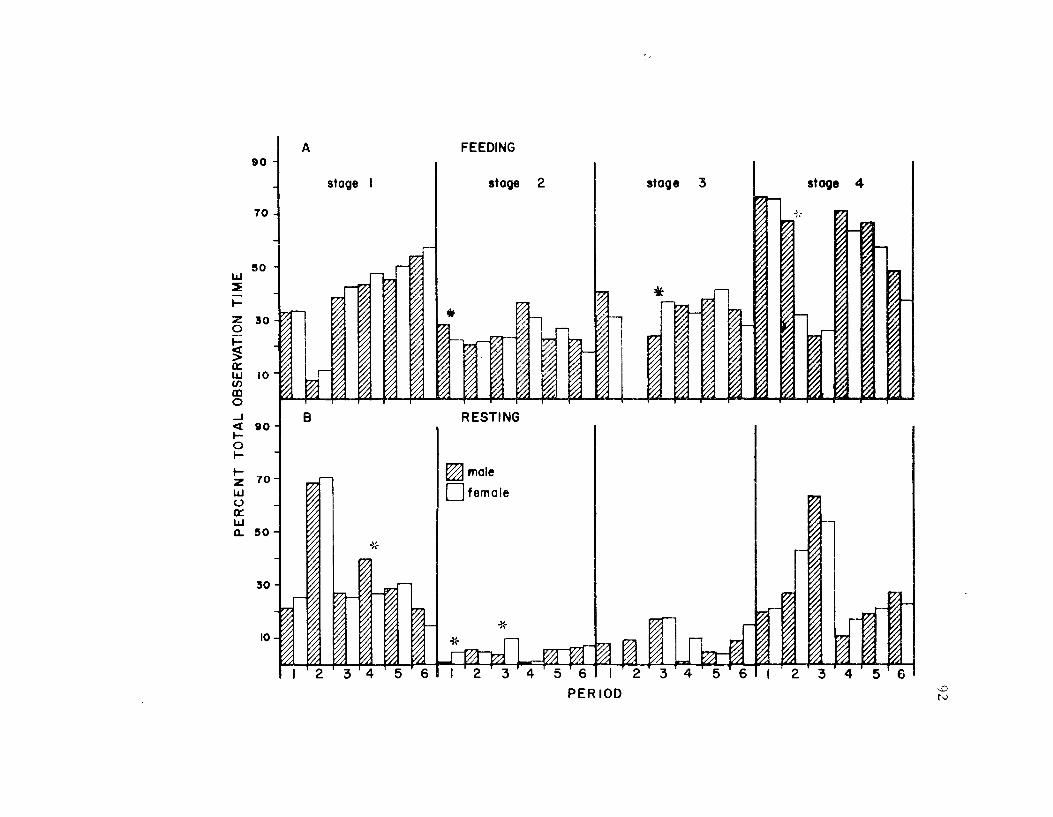
90
70
50
30
10
90
70
50
30
10
A FEEDING
stage I stage 2 stage 3 stage 4
B RESTING
male
ri female
3 4IN'
4
PERIOD4 5 6
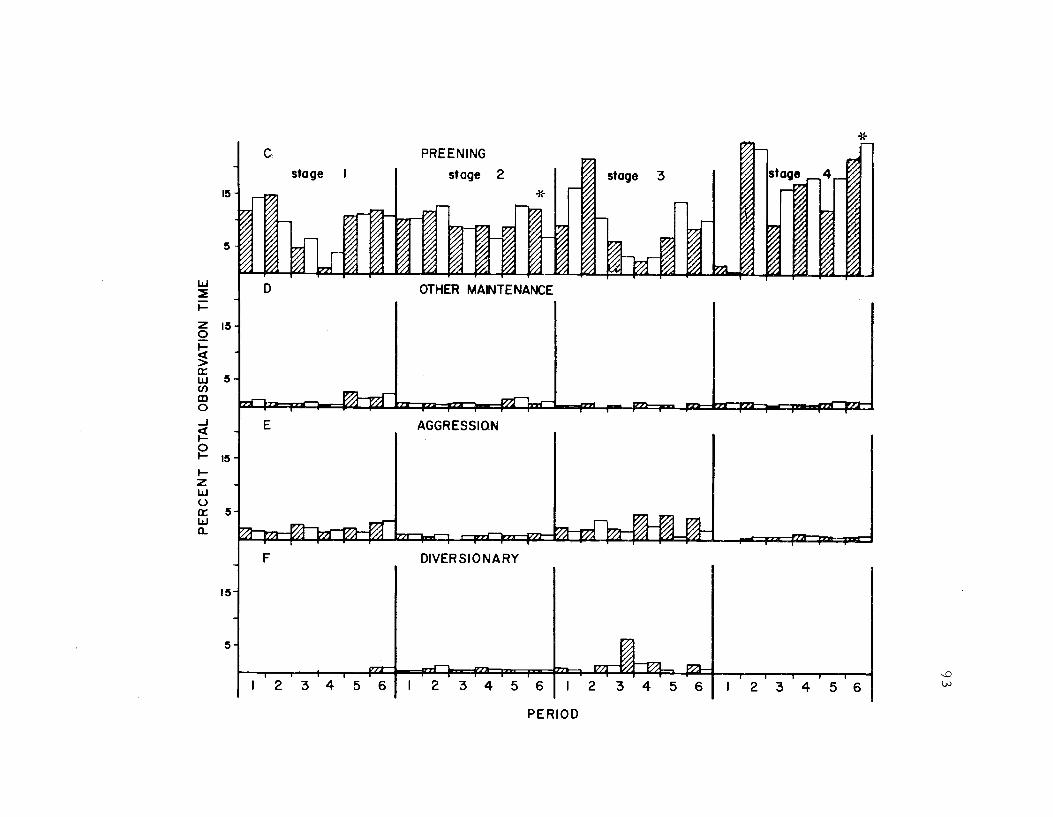
15
15
15
15
C
stage I
PREENING
stage 2 stage 3
.4v A
ftO
OTHER MAINTENANCE
E AGGRESSION
V%2=40; AMAAim
F DIVERSIONARY
3 4 5 6 1 2 3 4 5 6 I 2
PERIOD
" IMP /1' . /a. =I le
4 5 6 3 4 5 6
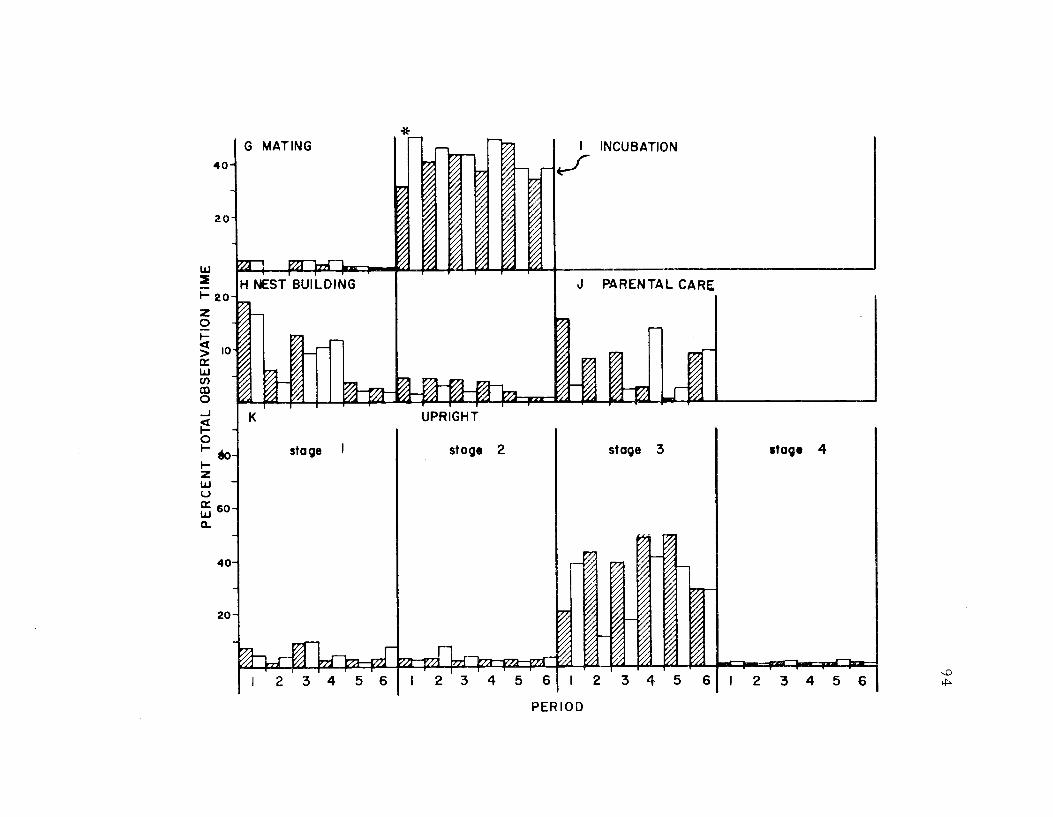
40-
20
w
/7 20z0
10
CCwU)C00
06o
zw
OC 60G.
40
20
VI
G MATING I INCUBATIONV
4 444441.1 /011. , oH NEST BUILDING J PARENTAL CARE
UPRIGHT
stage I stage 2 stage stage 4
MN
3 4 5 6 1 2 3 4 5 6 2 3 4 5 6 1 2 3 4 5 6
PERIOD

95
DISCUSSION
Avocet evolution has resulted in a system of integrated tactics
by which they exploit their environment. These span a broad range,
including the type of mating system, pattern of spatial dispersion,
nesting requirements, breeding season, clutch size, parental care
system, age and size distribution, age at first breeding, longevity,
timing of molts, and migration (Pitelka, pers. comm.). The activi-
ties involved in this tactical design all require energy. Some require
more than others; therefore, the birds must organize their activities
so that they have sufficient time to feed and gain needed energy. The
relationship, then, between energy demands and the allotment of
time to various activities is probably one of the most important ele-
ments in the evolution of a successful exploitation system.
I have measured the time element, or the allocation of time to
activities, and I now will discuss the adaptive nature of the avocet's
seasonal and diurnal foraging pattern in relation to energy require-
ments. I will also briefly delve into the apportionment of time to
non-foraging behavior, and how this relates to the energy budget.
Finally I will attempt to relate the roles of the sexes in the social
system.

96
Energy and Seasonal Changes in Foraging Activity
Since an individual obtains the necessary energy to carry on
life processes by feeding, its energy demands should rather directly
affect the amount of time spent feeding, as modified by food density
and availability. The energy required by any individual is dependent
on several factors; these include weight, ambient temperature, ac-
tivity levels, and such energetically expensive processes as egg
production, molt, and prernigratory fat deposition. Therefore, the
adaptiveness of the allocation of time to foraging during the breeding
season can perhaps be explained by the relationship between the
avocet's energy needs and the availability of food and time (Figure
24).
There is an approximately inverse linear relationship between
temperature and standard metabolic rate for a bird of a certain we,et
(Kendeigh, 1970; King and Farner, 1961). Resting avocets (mean wt,
g) at 30°C require about 33 Kcal/bird/day while at 0°C the
standard metabolic rate is about 92 Kcal/bird/day (see Appendix 7)0
The daily mean ambient temperature at Summer Lake generally
increased throughout the breeding season (Figure 23). One would
therefore expect a corresponding decrease in the standard metabolic
rate from early to late in the breeding season. Other factors aside,
the decrease should be reflected in a decrease in the foraging time

Figure 23. The seasonal temperature regime for SummerLake, Oregon during 19 69. The daily maximumand minimum temperatures were obtained froma U. S. Weather Bureau station located approx-imately 2 mi from the study area. Temperaturesare averaged for 15 day periods.
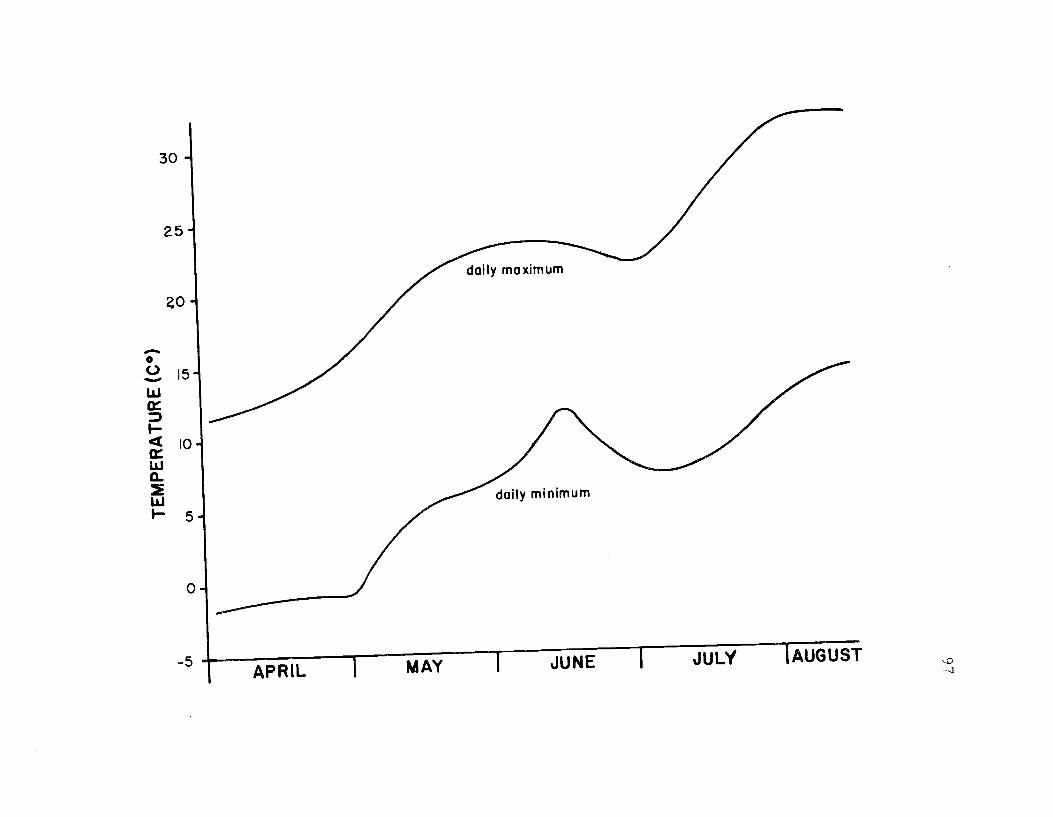
30
25
20
15
I0
daily maximum
daily minimum
APRIL MAY JUNE JULY AUGUST

Figure 24, Generalized relationships between time spent infeeding by avocets and energy demands. The sizeof the boxes under feeding time represents theamount of time spent foraging in each stage of thebreeding cycle. The width of the arrows indicatesthe relative effect each factor has on the amount oftime spent feeding in each stage. The dashed lineindicates a contributing effect.
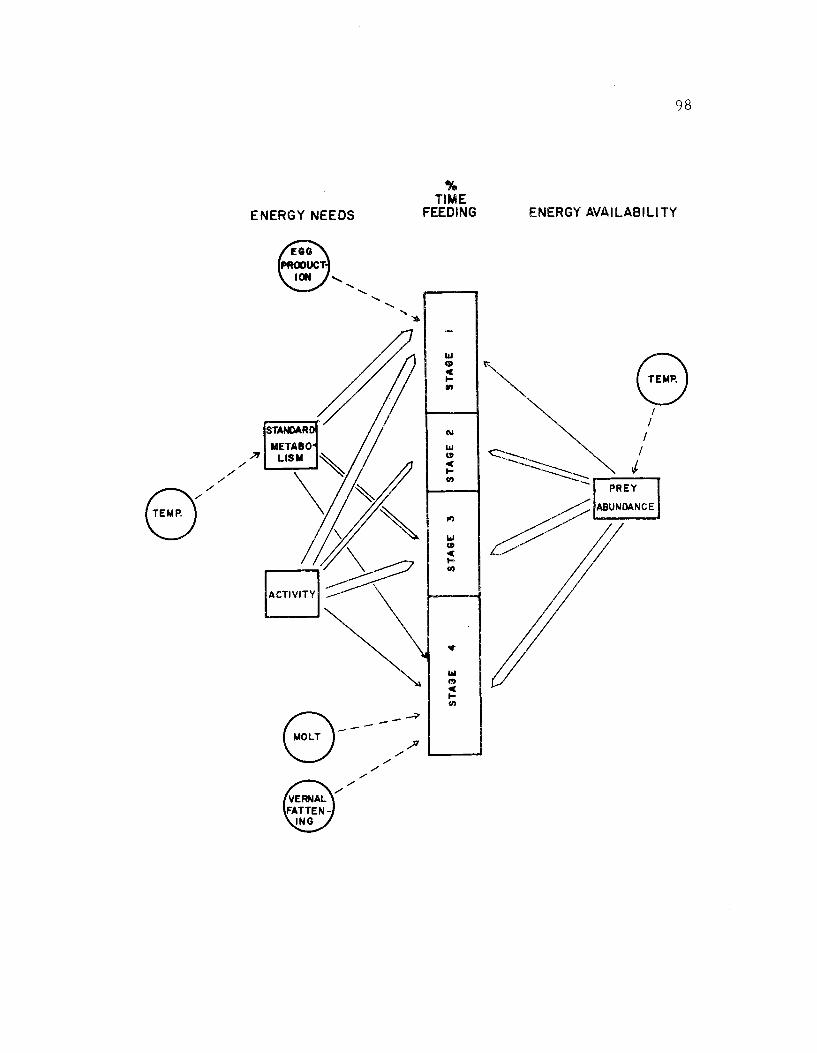
ENERGY NEEDS
98
TIMEFEEDING ENERGY AVAILABILITY
/
PREY
ABUNDANCE
ACTIVITY

99
(Figure 24). Such a relationship has been established for wintering
Brewer's Blackbirds (Euphagus cyanocephalus) under field conditions
(Verbeek, 1964).
Activity or physical exertion is energetically expensive and
adds to energy requirements. Kendeigh (19 70) feels that most of the
increase in energy requirements over standard metabolism is due
to the amount of locomotor activity taking place. For example, fly-
ing increases metabolic rate 5 to 20 times (Tucker, 1969). Avocet
energy requirements should, therefore, be higher in stages of the
breeding cycle with the highest levels of activity (Figure 24, Table 8).
Avocets must go through several energetically expensive proc-
esses during the breeding season. These are egg production, the
postnuptial molt, and premigratory fat deposition. Egg production
has been shown to significantly increase energy needs in the Willow
Ptarmigan (wt. 571 g), which expends about 22 Kcal/female/day in
egg production. This represents a 21 percent increase over standa,rd
energy demands for that time of year (West, 1968). Egg production
has also been shown to be energetically expensive in Zebra Finches
(Taeniopygia castanotis) causing a 13 percent increase in standard
metabolic rate (El-Wailly, 1966).
The post-nuptial molt also increases energy needs. West (1968)
found that Willow Ptarmigan used about the same amount of energy in
molting as in egg production. Domestic fowl increase energy needs
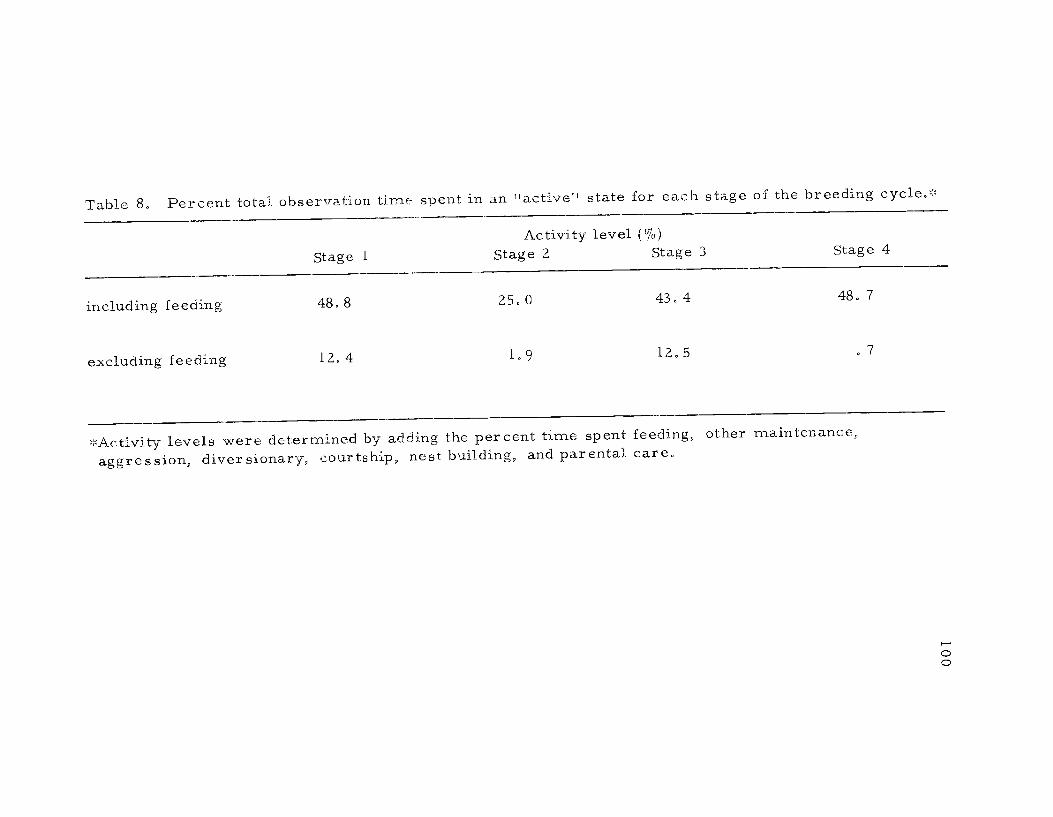
Table 8. Percent total observation time spent in an "active" state for each stage of the breeding cycle.*
Stage 1Activity level (%)
Stage 2 Stage 3 Stage 4
including feeding 48. 8 25.0 43. 4 48. 7
excluding feeding 12.4 1.9 12.5 . 7
*Activity levels were determined by adding the percent time spent feeding, other maintenance,aggression, diversionary, courtship, nest building, and parental care.

1 01
during molting by 45 percent (Perek and Sulman, 1945) and molting
Emberiza hortulana increase metabolism by 62 percent (Wallgren,
1954). Lustick (1970) found that Cowbirds (Molothrus ater) used 13
percent more oxygen while molting. In addition, 13 percent more
oxygen was used for increased thermoregulation due to decreased
feather insulation at 35 C.
During premigratory fat deposition there is a shift in energy
storage from glycogen to fat. This shift is also accompanied by
hyperphagia (King and Farner, 1961). Morton (196 7) found that
White-crowned Sparrows (Zonotrichia leucophrys) increased food
intake by 20 percent during vernal fattening. Egg production, molt,
and premigratory fattening increase energy requirements and they
should influence the amount of time spent feeding in the stages in
which they occur (Figure 24).
Energy needs cannot be directly related to the amount of time
spent foraging, however, because food abundance, availability, ard
quality may vary through the breeding season. Avocets feed pri-
marily on aquatic insects and aquatic vegetation (Wetmore, 1925).
The seasonal changes in the abundance and availability of these food
items should influence the amount of time the avocets must spend
foraging. The patterns of seasonal fluctuation in insect abundance
at Summer Lake are not known, but data are available for the
Whistler's Bend Impoundment, located 17 km northeast of Roseburg

102
in southwestern Oregon (Coche, 1964). The magnitudes of abundance
and the species present in the two areas are probably not the same,
but there should be a general correspondence in the pattern of sea-
sonal abundance. Aquatic insect populations in the impoundment be-
gan to increase in. March and reached high levels by July (Figure 25).
Coche's data indicate insect abundance as numbers of emerging
adults. Therefore the insect larvae available to avocets would be
at their largest size and greatest abundance from 1 to 2 weeks before
the emergence (N. Anderson, pers. com. ). This means that avocets
would probably need to spend more time in feeding early in the breed-
ing season and less later on to obtain the same amount of insect food.
Verner (1965) found that the Long-billed Marsh Wren (Telmatodytes
palustris) consumed 4.3 percent more energy in May than June but
spent 30 percent more time feeding in May. He felt this difference
was due to differences in insect abundance. A similar situation
has been found for Red-winged Blackbirds (Agelaius
(Hintz and Dyer, 1970). It is possible that avocets at Summer Lake
fed primarily on vegetation early in the season due to low abundance
of insect prey. This tactic would probably increase the total amount
of time required for feeding activity, however, since vegetation has
about 20 percent less caloric value per gm dry weight than insects
(Southwood, 1966). It should also be remembered that avocets can
probably not utilize the cellulose portion of plants.

Figure 25. Seasonal pattern of aquatic insect emergence forthe Whistler's Bend Impoundment, Douglas County,Oregon. Estimates of the number of emerginginsects were obtained from trapping. Chironornini,Tanytarsini, Orthocladiinae, Ablaesmyia, Proladius,Ephydridae, Ephemeroptera, and Coenagrionidaewere included. Modified from Coche (1964).
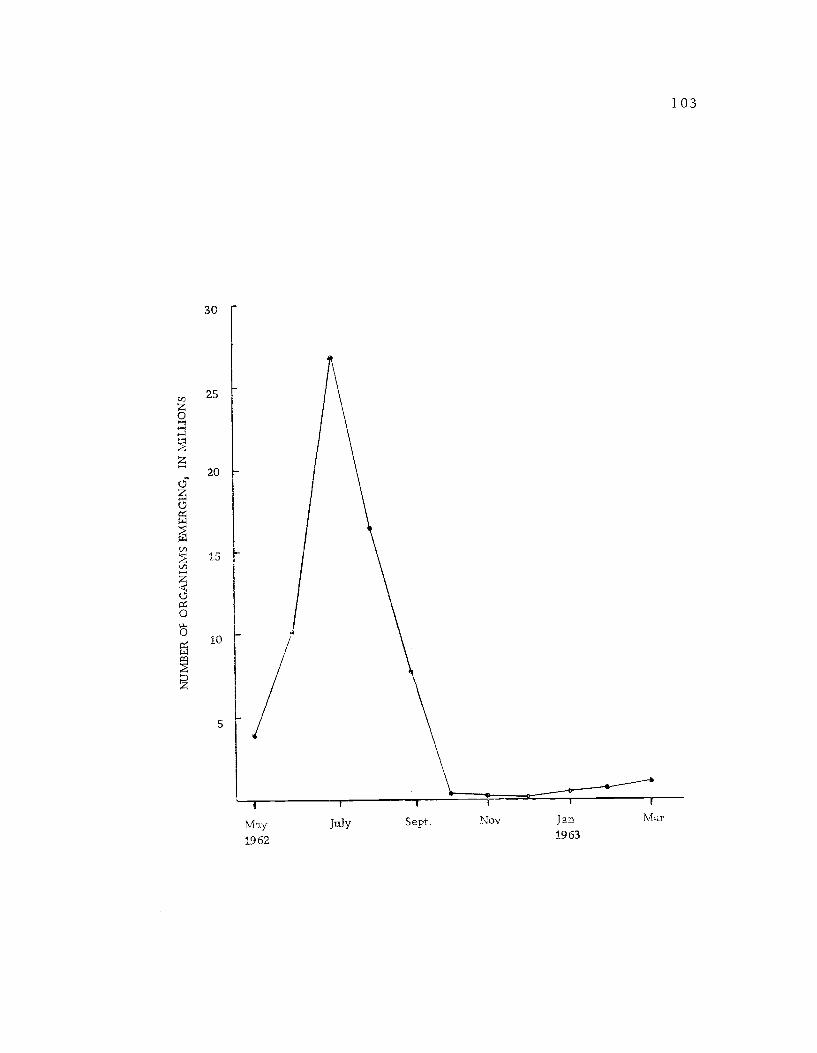
z 0
NU
MB
ER
OF
OR
GA
NIS
MS
EM
ER
GIN
G, I
N M
ILL
ION
S
00

104
Body weight undoubtedly affects energy balance. When energy
requirements are high the birds must either feed enough. to keep up
with energy demands or they will lose weight. This factor may some-
what lower the amount of time spent in foraging during one stage and
feeding might later be increased to rectify the weight loss.
The foraging time budget for the four stages of the breeding
cycle (Figure 19A) appears to be affected by a combination of the
above energy needs. This is summarized in Figure 24. Stage 1 had
the second highest feeding level. Several factors appear to contribute
to this, such as the low environmental temperature and food abun-
dance, high activity levels, plus egg production. In Stage 2 time
spent feeding dropped to its lowest level. This may be accounted
for by the higher ambient temperature, greatly reduced activity
level, and greater prey abundance. The foraging level increased
in Stage 3 in spite of Ligher temperatures and the greatest prey
abundance. The high activity level associated with the care and
protection of the young perhaps accounts for the increased foraging
time. Stage 4 had the highest feeding level, yet temperature and
prey abundance should have been at a high at that time. In addition
activity levels were very low. The post-nuptial molt and premigra-
tory fat deposition were occurring during this stage and I believe
these two processes account for the large allocation of time to
feeding.

1 05
Energy and Seasonal Patterns in Non-feeding Activities
The pattern of the seasonal foraging time budget appears to be
largely controlled by energy requirements during the breeding cycle.
It must be remembered, however, that the amount of time spent
foraging is bacially dependent on the size of the bird, temperature,
and food availability. The allotment of time to all other activities
is determined by the ability of the birds to gain the extra energy
to perform them. A successful species must have evolved a social
system in which the allotment of time and energy to various activities
does not exceed the minimum time needed to obtain sufficient energy.
Therefore, one would expect selection to reduce the time spent in
energy-demanding activities to a minimum. For example, aggres-
sion should not take a large portion of time or energy. Further, one
would expect evolution of a nesting strategy which would offer the
least contact with predators and thus lessen time spent for diversion
and nest defense.
The allocation of time to physically demanding activities such
as aggression and diversionary activities was small for avocets at
Summer Lake (Figure 19). Similar situations have been found for
Marsh Wrens and Dickcissels (Verner, 1965; Schartz and Zimmerman,
19 71).
All animals apparently need rest. The physiological reasons

106
for rest are unclear, but it may have evolved in relation to energy
needs. By reducing metabolic rate to a minimum during a large
portion of the 24-hour cycle, energy can be conserved, especially
during the night when it is not feasible for diurnal animals to feed.
Further, one might expect an animal to rest most when energy de-
mands are the greatest. I found no evidence that avocets changed
the amount of time spent in an "inactive" state during the day in
relation to energy requirements.
The Bimodal Nature of the Activity Pattern
The avocet's diurnal activity pattern was characterized by two
peaks of activity separated by a period of inactivity occurring in mid
to late morning (Figure 26). This pattern was represented in many
categories of avocet's behavior (Figure 21). A similar bimodal
diurnal activity patte r, has also been found for a wide variety of
animal taxa, including insects, arthropods, mammals, and birds
(Aschoff, 1966). The details of these diurnal activity patterns are
different for different species but all are ultimately related to the
time phasing of environmental conditions (Palmgren, 1949; Aschoff,
1964; Verner, 1965). This implies that a diurnal pattern of activity
is adaptive and a product of natural selection.
Figure 27 summarizes the organization of the diurnal time
budget on a seasonal basis for several categories of behavior. The

Figure 26. The apportionment of avocet's time to an "active"state. The percent time active was obtained bysumming the percent total observation time offeeding, mating, nestbuilding, in parental care,aggression, other maintenance, and diversionaryactivities.
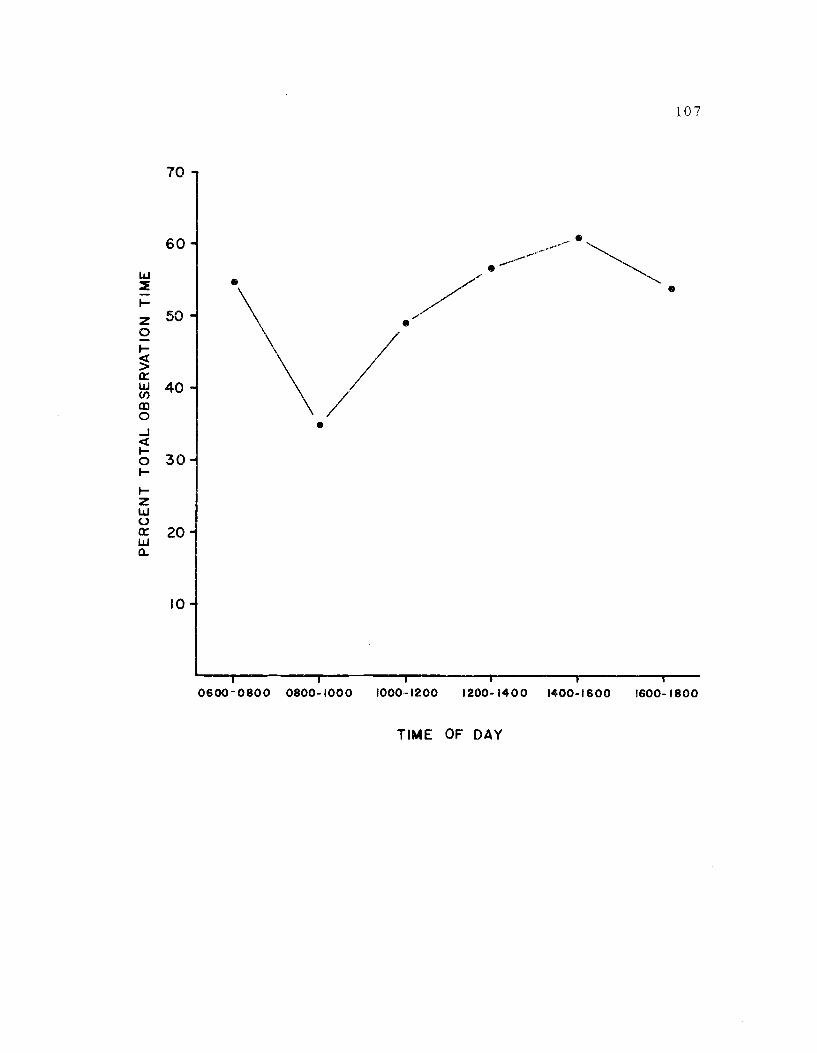
70
60
50
40
30
20
10
107
0600-0800 0800-1000 1000-1200 1200-1400 1400-1600
TIME OF DAY
1600-1800

Figure 27. The apportionment of time by avocets to 7 categoriesof behavior for six diurnal time periods over theentire breeding season. F = feeding; B = resting;P = preening; A = other maintenance, aggressiondiversionary, courtship, and nestbuilding; I = incu-bation; U = upright; PC = parental care; and Babsent plus other unaccounted for time
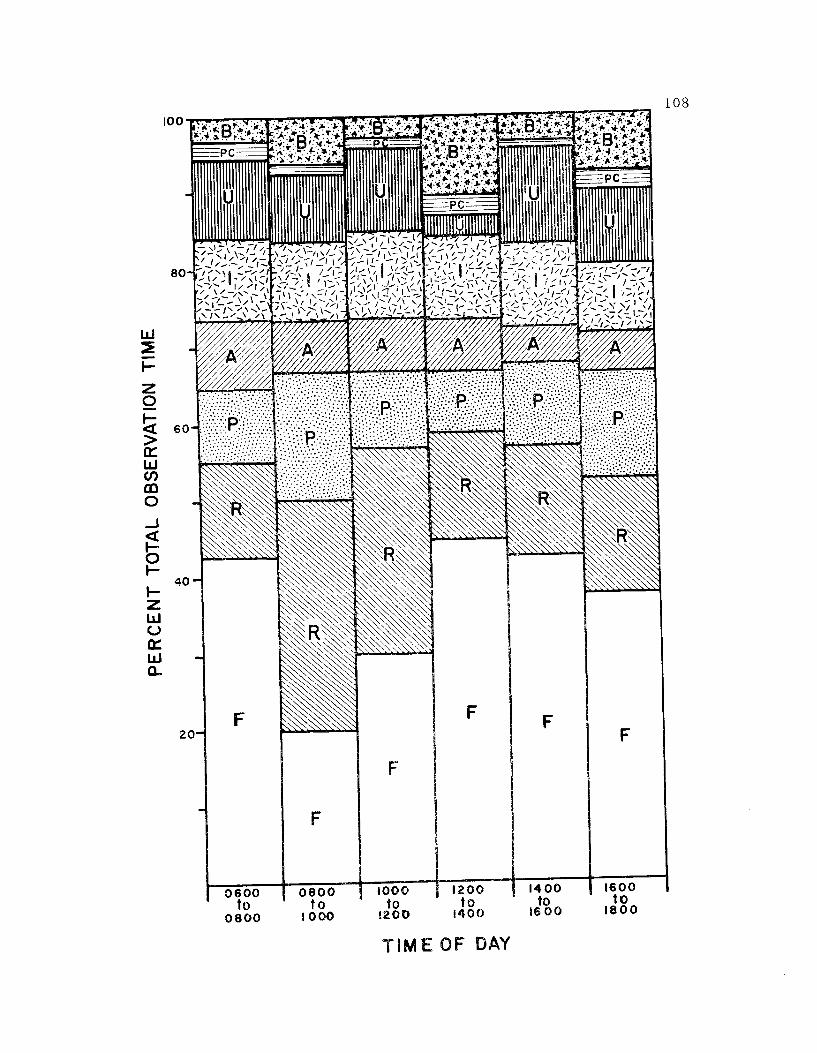
108
0600 0800 1000 1200 1400 1600to to to to to to
0800 1000 1200 1400 1600 1800
TIME OF DAY

1 09
only categories which varied appreciably through the day were feeding
and resting. It is also apparent that the diurnal pattern of feeding
corresponds closely with the overall diurnal activity pattern (Figure
26). Therefore foraging appears to be the major factor in determin-
ing its form. The feeding pattern, in turn, is related to features of
the environment (Figure 24). Therefore a closer look at the adap-
tiveness of the diurnal foraging pattern should offer insight into the
adaptiveness of the bimodal activity pattern.
Since feeding is synonymous with energy procurement, its di-
urnal distribution should reflect patterns of an individual's diurnal
energy requirements given constant food availability. The morning
feeding peak was evident in nearly every breeding cycle stage (Figure
21), I believe that this feeding peak occurred because the birds had
not fed for 6 to 10 hours and they had to feed in the early morning to
make up the energy debt incurred during the night. Red-winged
Blackbirds seem to show a similar response to the night-long fast
(Hintz and Dyer, 19 70). The height of the morning feeding peak
should be related to the energy demands placed on the birds during
the night. The highest morning feeding peaks occurred in Stages 1
and 4. Night temperatures were probably a major factor contributing
to the increased feeding time in Stage 1 (Figure 28) and the demands
of the post-nuptial molt and premigratory fat deposition were probably
important in Stage 4, During Stages 2 and 3 there were lower

Figure 28. The diurnal foraging pattern for avocets atSummer Lake. The diurnal temperatureregime from Cabin Lake, Oregon, located40 mi from Summer Lake. The diurnalpattern of emergence of Tricoptera atMontreal, Quebec (expressed as a percent-age of the occasions on which the highestcatch of the day fell at a certain time) (fromCorbet, 1966). The only relationship betweenthese curves are those made through infer-ence in the text by the author.
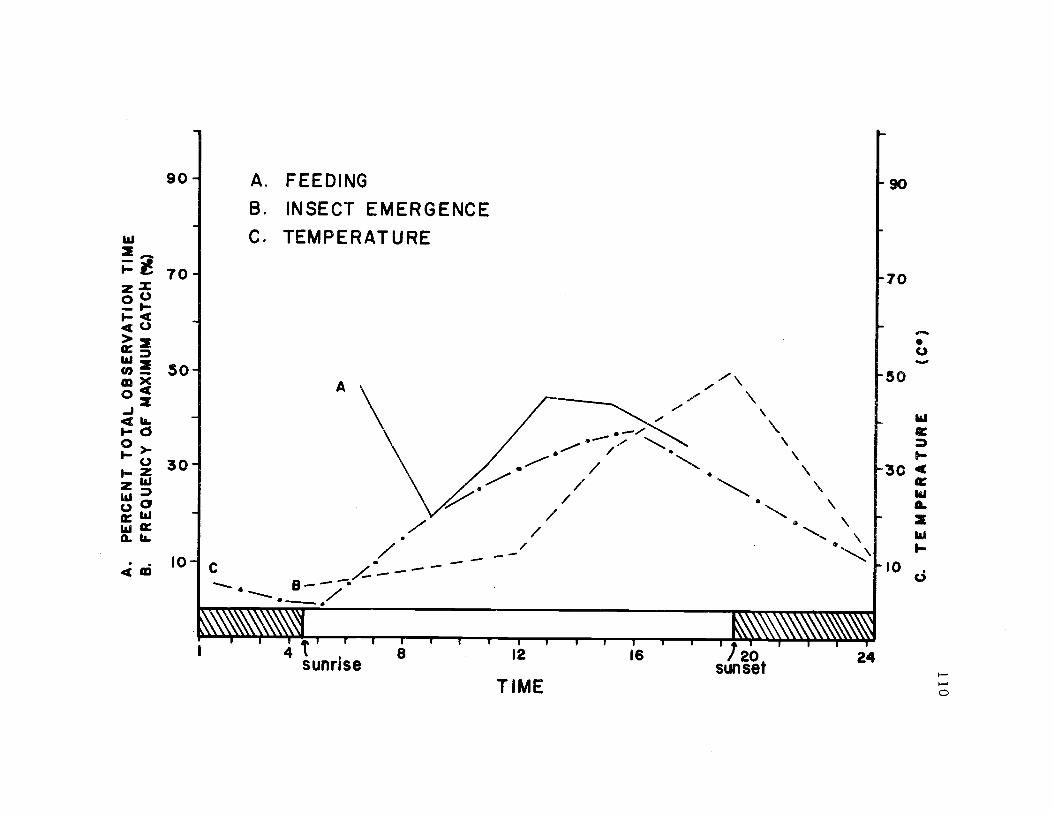
A.
PE
RC
EN
T T
OT
AL
OB
SE
RV
AT
ION
TIM
EB
.F
RE
QU
EN
CY
OF
MA
XIM
UM
CA
TC
H (
%)
0
C-
U)
CD
co
E'7
"m
cn
°
OT
T
0C
s 00
co 0 I
7
T1 m m z
E).
0C
. TE
MP
ER
AT
UR
E
u 0()

111
night-time energy demands due to higher temperatures and a shorter
period of darkness; the morning feeding peaks were correspondingly
lower. These peaks may also be lower because the birds had more
time to feed in the morning prior to the observation periods. The
relative heights of the morning feeding peaks are not due entirely
to the energy demands of early morning. Food abundance is probably
also an important factor determining the proportion of time the birds
spent feeding.
I believe the afternoon feeding peak was correlated with the
optimum time to feed. Several factors contributed to this. Diurnal
temperatures in the central Oregon area reach a maximum in the
afternoon (Figure Z8). Diurnal patterns of aquatic insect emergence
also tend to reach a maximum in the afternoon (e.g. Corbet, 1966)(e. g.
Figure Z8). Further, insects generally are more active at warmer
temperatures (Chapman, 1969), which should increase the ease of
capture by avocets. Similar responses have been observed in other
species which feed on insects; Willow Warblers (Phylloscopus
trochilus), European Robin (Erithacus rubecula), Blue Tits (Parus
caeruleus), Blackbirds (Turdus merula), White-crowned Sparrows,
and Long-billed Marsh Wrens have been shown to have afternoon
feeding maxima (Kuusisto, 1941; Lees, 1948; Morton, 1967; Verner,
1965). The morning foraging peak is not as common, although
Morton (1967) found this peak for White-crowned Sparrows on the

1 1 2
wintering grounds, and Hintz and Dyer (19 70) found the same situa-
tion in breeding Red-winged Blackbirds. It appears that many pas-
serines during the breeding season spend the morning hours in mating
activities (Verner, 1965; Schartz and Zimmerman, 19 71). The ability
to do this requires that the birds be able to store enough energy the
day before to last them through the night and the following morning.
The present evidence suggests that avocets may not be very efficient
in storing enough energy to last through the morning. Also, their
mating activities do not require as much time during the morning
hour s.
The other major feature of the diurnal activity pattern was the
resting phase. A resting or inactive period occurring sometime
during the day occurs in many species of birds. The timing of the
rest period varies, but for most it comes shortly after noon (Palm-
gren, 1949; Aschoff, 1966). However, avocet's inactive period
occurred in mid to late morning. This pattern is in contrast to
the generalization that larger species might be expected to have
afternoon rest periods because they need to feed less (Gibb, 1960).
There has been a great deal of speculation and study to determine why
various species tend to be inactive at a certain time of day. Such
things as avoidance of high mid-day temperatures, low humidities
and so on have been used as explanations.
One assumes, as with the rest of the diurnal time budget, that

1 1 3
the timing of the rest period is subject to natural selection and is
adaptive. In avocets, foraging took the major part of the early
morning as well as most of the afternoon, leaving little time for
resting during these periods. By resting in mid-morning the birds
did not interfere with times when feeding was necessary or environ-
mental conditions were optimal for foraging. Schartz and Zimmerman
(1971) have found a similar resting pattern for the male Dickcissel.
It may be that insectivorous species tend to rest in the morning in
order to take advantage of afternoon insect activity.
Foraging and resting appear to be the dominant activities in con-
trolling the form of the diurnal activity budget. How are other activi-
ties organized in time in response? The major trend is for most
activities to take about the same amount of time in each period of
the day (Figure 27).
Sexual Aspects of the Time Budget
Male and female avocets are similar both morphologically and
behaviorally. This is reflected by the absence of any major differ-
ences between the two sexes in both the seasonal and diurnal time
budgets. Both the behavioral and ecological relationships between
the sexes are probably important in determining the reasons for
the lack of sexual divergence.

114
Several situations may promote the occurrence of sexual
divergence. They include intersexual competition for food and mate
selection (Selander, 1966). The latter is most often seen for poly-
gamous or lek social systems. The opportunity for sexual divergence
due to intersexual competition depends upon the habitat being suffici-
ently complex to be subdivided (Selander, 1966). Avocets may sub-
divide food resources between the sexes by a divergence of bill shape
(Hamilton, 1969). The simplicity of the habitat in which avocets feed
may limit the possibilities for further morphological divergence.
Hamilton has found that both sexes feed in the same depth water, but
I have observed that male and females may feed in somewhat different
areas. A spatial divergence would not be reflected in either morph-
ology or in the time budget.
Considering the extended incubation period of avocets (24 days)
and the necessity for the eggs to be covered almost continually, t!-ere.
is little time for males to court or attend more than one female.
This has possibly reduced selective pressure associated with mate
selection. Also mate selection is not associated with a territory and
neither sex has behavioral patterns which might be used to advertise
a territory. Waders which do use the territory for mate selection
have special behavior patterns for courting females and proclaiming
the territory. Due to the avocet's conspicuousness in the marsh

11 5
habitat they probably do not require special advertising displays.
The simplicity of the habitat, the avocet's particular mating system,
the necessity of covering the eggs almost continuously, and the ex-
tended parental care stage (30+ days) probably account for the general
similarities in the seasonal and diurnal time budgets of the sexes.
This study of the temporal organization of the avocet's behav-
ioral activities in time has illustrated the importance of the time
dimension toward an understanding of both the behavior and ecology
of this species. Further, the relationship between time and energy
appears to be very important in determining the way time utilization
is organized.
The results of this study suggest several lines for further in-
vestigation. I have attempted to indicate the manner in which energy
requirements influence the structure of a time budget. Indeed, as
Collias et al. (19 71) have pointed out, time studies have limited value
unless they can be quantitatively related to behavioral energetics
through metabolic measures. Therefore a logical extension of a time
budget study is an investigation of avocet energy requirements, in-
cluding food abundance and availability.
Another area of considerable interest is the relationship between
the sexes, particularly as it relates to niche subdivision. It is pos-
sible that each sex utilizes the habitat differently, but in a way which
is not reflected in time budget differences. Perhaps a time-space

116
analysis considering how the sexes subdivide the habitat would
elucidate sexual differences, if any are present.

11 7
BIBLIOGRAPHY
Aschoff, J. 1964. Survival of diurnal rhythms. In: Biology ofsurvival, ed. by O. G. Edholm. Symposium of the ZoologicalSociety of London 13:78-98.
Aschoff, J. 1966. Circadian activity pattern with two peaks. Ecol-ogy 47:657-662.
Baldwin, E. M. 1964. Geology of Oregon. 2d ed. Eugene, Univer-sity of Oregon Coop. 165 p.
Bent, A. C. 1927. Life histories of North American shorebirds.Smithsonian Institution United States National Museum Bulletinno. 142. Part 1. 420 p.
Boyd, H. 1962. Mortality and fertility of European Charadrii. Ibis104:368-387.
Brisbin, I. L., Jr. 1969. Bioenergetics of the breeding cycle ofthe ring dove. Auk 86:54-74.
Brown, P. E. 1949. The breeding of avocets in England, 1948.British Birds 42:2-12.
Chapman, R. F. 1969, The insects: Structure and function.London, English Universities Press Ltd. 819 p.
Coche, A. G. 1964. Net production of juvenile steelhead Salmogairdnere Richardson, in a freshwater impoundment. Ph. D.thesis, Corvallis, Oregon State University. 274 numb. leaves.
Collias, N. E., et al. 1971. Some observations on behavioralenergetics in the village weaverbird. II All-day watches inan aviary. Auk 88:133-143.
Corbet, P. S. 1966. Diel periodicities of emergence and ovipositionin riverine Trichoptera. The Canadian Entomologist 9 8: 1025-1034.
Cullen, J. M. 1954. The diurnal rhythm of birds in the arctic sum-mer. Ibis 96:31-46.

118
El-Wailly, A. J. 1966. Energy requirements for egg-laying andincubation in the zebra finch, Taeniopygia castanotis. Condor68 :582 -594.
Gibb, J. 1954. Feeding ecology of tits, with notes on treecreeperand goldcrest. Ibis 96:513 -543.
Gibb, J. 1956. Food, feeding habits and territory of the rock pipetAnthus spinoletta. Ibis 98:506-530.
Gibb, J. 1960. Populations of tits and goldcrests and their foodsupply in pine plantations. Ibis 102:163-208.
Gibson, F . 19 71. The breeding biology of the American avocet incentral Oregon. Condor. [In press]
Hamilton, R. B. 1969. The comparative behavior of the Americanavocet (Recurvirostra americana) and the black-winged stilt(Himantopus himantopus mexicanus). Ph. D. thesis.. Berkeley,University of California. 273 numb. leaves.
Hintz, J. V. and M. I. Dyer. 19 70. Daily rhythm and seasonalchange in the summer diet of adult red-winged blackbirds.Journal of Wildlife Management 34:789-799.
Huntsberger, D. V. 1967. Elements of statistical inference. 2ded. Boston, Allyn and Bacon. 398 p.
Kendeigh, C. S. 19 70. Energy requirements for existence in rela-tion to size of birds. Condor 72:60-65.
King, J. R. and D. S. Farner. 1961. Energy metabolism, tl.ermo-regulation and body temperature. In Biology and comparativephysiology of birds. Vol. 2. ed. by A. J. Marshall. NewYork, Academic press. p. 215-288.
Kluijver, H. N. 1950. Daily routine of the great tit Parus m. majorL. Ardea 38:99-135.
Kuusisto, P. 1941. Studien fiber die Okologie and Tagesrhytrnik vonPhylloscopus trochilus acredula (L. ) mit besonderer Berack-sichtigung der Brutbiologie. Acta Zoologica Fennica 311:1-120.
Lack, D. 1968. Ecological adaptations for breeding in birds.London, Methuen and Company Ltd. 409 p.

119
Lees, J. 1948. Winter feeding hours of robins, blackbirds, andblue tits. British Birds 41:71-76.
Lustick, S. 1970. Energy requirements of molt in cowbirds. Auk
8 7: 742- 746.
Makkink, G. F. 1936. An attempt at an ethogram of the Europeanavocet with ethological and psychological remarks. Ardea 25:1-62.
Makkink, G. F. 1942. Contribution of the knowledge of the behaviorof the oystercatcher (Haematopus ostralegus L. ). Ardea 31:23-74.
Manning, A. 1967. An introduction in animal behavior. Reading,Mass., Addison-Welsey Publishing Co. 208 pa
Marler, P. 1956. Behaviour of the chaffinch (Fringilla coelbs).Behavior sup. no. 5. 184 p.
Marler, P. and W. J. Hamilton, III. 1966. Mechanisms of animalbehavior. New York, John Wiley and Sons, Inc. 771 p.
Martin, S. G. 19 71. Polygmy in the bobolink: habitat quality andthe adaptive complex. Ph. D. thesis. Corvallis, Oregon StateUniversity. 181 numb. leaves.
Morton, M. L. 196 7 Diurnal feeding patterns in white-crownedsparrows Zonotrichia leucophrys gambelii. Condor 69:49 1-512.
Orians, G. H. 1961. The ecology of blackbird (Ageloius ) socialsystems. Ecological Monographs 31 :285 -312.
Palmgren, P. 1949. On the diurnal rhythm of activity and rest inbirds. Ibis 91 :561 --5 76.
Perek, M. and F. Sulman. 1945. The basal metabolic rate in molt-ing and laying hens. Endocrinology 36 :240 -243.
Ripley, S. a 1961. Aggressive neglect as a factor in inter specificcompetition in birds. Auk 78:366-3 71.
Sauer, E. G. F. and E. M. Sauer. 1967. Yawning and other main-tenance activities in the South African ostrich. Auk 84:5 71-
58 7.

1 20
Schartz, R. L. and J. L. Zimmerman, 19 71. The time and energybudget of the male dickcissel (Spiza americana). Condor 73:65 -76.
Selander, R. K. 1966. Sexual dimorphism and differential nicheutilization in birds. Condor 68:113-151.
Smith, C. C. 1968. The adaptive nature of social organization in thegenus of three squirrels Tamiasciurus. Ecological Monographs38:31-63.
Southwood, T. R. E. 1966. Ecological Methods. London, Methuenand Co., Ltd. 39 1 p.
Tucker, V. A. 1969. The energetics of bird flight. ScientificAmerican 220: 70- 78.
Verbeek, N. A. M. 1964. A time and energy budget study of thebrewer's blackbird. Condor 66:70--i4.
Verner, J. 1965. Time budget of the male long-billed marsh wrenduring the breeding season. Condor 67:125 -139.
Wallgren, H. 1954. Energy metabolism of two species of the genusEmberiza as correlated with distribution and migration. ActaZoologica Fennica 8 4:1-110.
West, G. C. 1968. Bioenergetics of captive willow ptarmigan undernatural conditions. Ecology 49:1036-1045.
Wetmore, A. 1925. Food of American phalaropes, avocets andstilts. United States Department of Agriculture DepartmentBulletin no. 1359.
Wheeler, R. 1955. Charadriiformes at the Laverton Saltworks,Victoria, 1950-1953. Emu 55:279-295.
Wiens, J. A. et al. 19 70. Metronome timing in behavioral ecolstudies. Ecology 51:350-352.
Williamson, K. 1950. Interpretation of "rodent-run" display. Ibis9 2: 2 8 3 3
Wolfe, L. R. 1931. The breeding Limicolae of Utah. Condor 33:49 -59.

APPENDICES
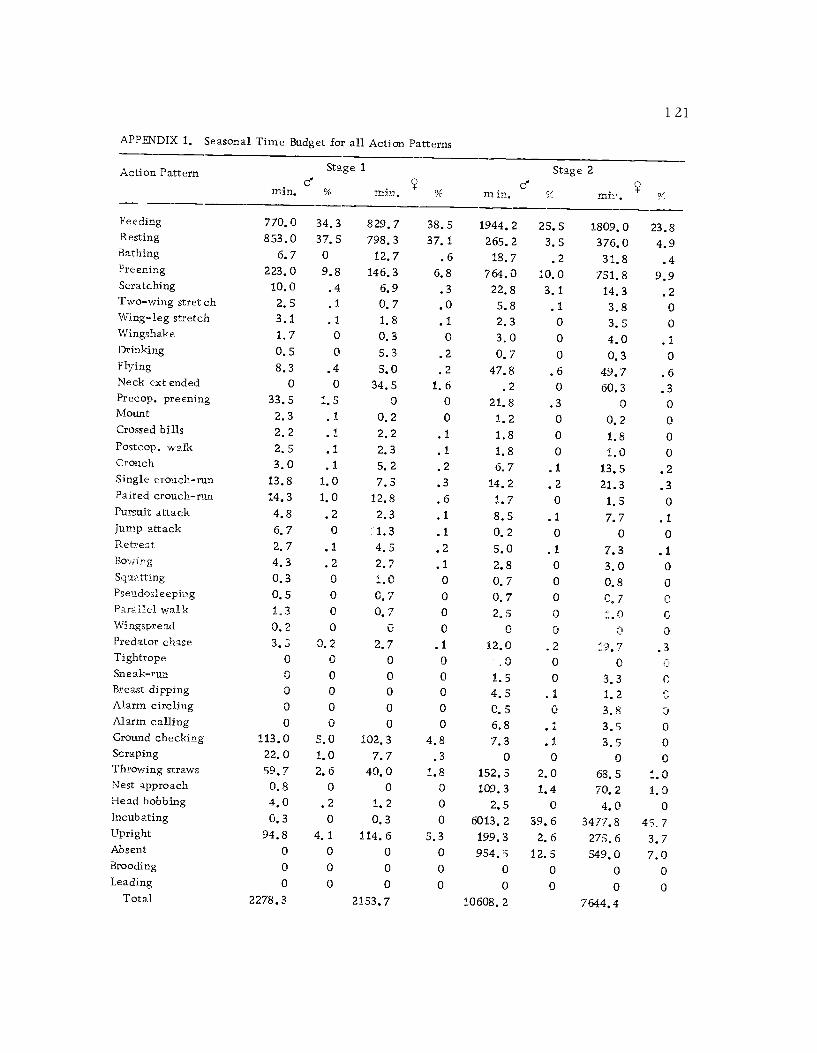
1 21
APPENDIX 1. Seasonal Time Budget for all Action Patterns
Action Patternmin.
Stage 1d'
V rni . na in.
Stage 2d
g mi
Feeding 770. 0 34. 3 829. 7 38.5 1944. 2 25.5 1809. 0 23.8Resting 853, 0 37. 5 798. 3 37. 1 265. 2 3. 5 376.0 4.9Bathing 6.7 0 12. 7 . 6 18.7 . 2 31.8 .4Preening 223, 0 9.8 146.3 6.8 764.0 10.0 751. 8 9.9Scratching 10.0 .4 6.9 .3 22.8 3. 1 14.3 . 2Two-wing stretch 2. 5 . 1 0. 7 . 0 5.8 . 1 3.8 0Wing-leg stretch 3. 1 . 1 1.8 . 1 2.3 0 3. 5 0Wingshake 1.7 0 0, 3 0 3. 0 0 4.0 . 1Drinking 0.5 0 5.3 . 2 0. 7 0 0, 3 0Flying 8.3 .4 5.0 . 2 47.8 . 6 49.7 . 6Neck ext ended 0 0 34. 5 1. 6 . 2 0 60.3 .3Precop. preening 33. 5 1. 5 0 0 21. 8 .3 0 0Mount 2.3 . 1 0, 2 0 1. 2 0 0. 2 0Crossed bills 2. 2 . 1 2. 2 . 1 1.8 0 1.8 0Postcop. walk 2. 5 .1 2.3 .1 1.8 0 1, 0 0Crouch 3. 0 . 1 5. 2 , 2 6. 7 . 1 13.5 . 2Single crouch -run 13.8 1.0 7.5 .3 14.2 . 2 21.3 .3Paired crouch-run 14.3 1, 0 12.8 , 6 1.7 0 1. 5 0Pursuit attack 4. 8 . 2 2. 3 . 1 8. 5 . 1 7. 7 . 1Jump attack 6, 7 0 1. 3 . 1 0. 2 0 0 0Retreat 2. 7 . 1 4. 5 . 2 5.0 . 1 7.3 . 1Bowing 4.3 . 2 2.7 . 1 2.8 0 3. 0 0Squ,Ating 0, 3 0 1.0 0 0. 7 0 0, 8 0Pseudosleeping 0, 5 0 0. 7 0 0. 7 0 0, 7 0Parallel walk 1. 3 0 0. 7 0 2, 5 0 1.0 GWingspread 0.2 0 0 0 0 0 0 0Predator chase 3..; 0. 2 2.7 .1 12.0 . 2 19.7 .3Tightrope 0 0 0 0 ,0 0 0Sneak-run 0 0 0 0 1.5 0 3.3 CBr east dipping 0 0 0 0 4. 5 . 1 1. 2 0Alarm circling 0 0 0 0 0. 5 0 3. 8 0Alarm calling 0 0 0 0 6, 8 . 1 3.5 0Ground checking 113.0 5.0 102. 3 4.8 7.3 . 1 3. 5 0Scraping 22, 0 1.0 7.7 .3 0 0 0 0Throwing straws 59.7 2. 6 40. 0 1.8 152. 5 2. 0 68. 5 1.0Nest approach 0.8 0 0 0 109. 3 1.4 70, 2 1. 0Head bobbing 4, 0 . 2 1. 2 0 2.5 0 4.0 0Incubating 0.3 0 0.3 0 6013. 2 39. 6 3477.8 45. 7Upright 94.8 4.1 114.6 5.3 199.3 2.6 275.6 3.7Absent 0 0 0 0 954.5 12.5 549, 0 7.0Brooding 0 0 0 0 0 0 0 0Leading 0 0 0 0 0 0 0 0
Total 2278.3 2153.7 10608.2 7644.4
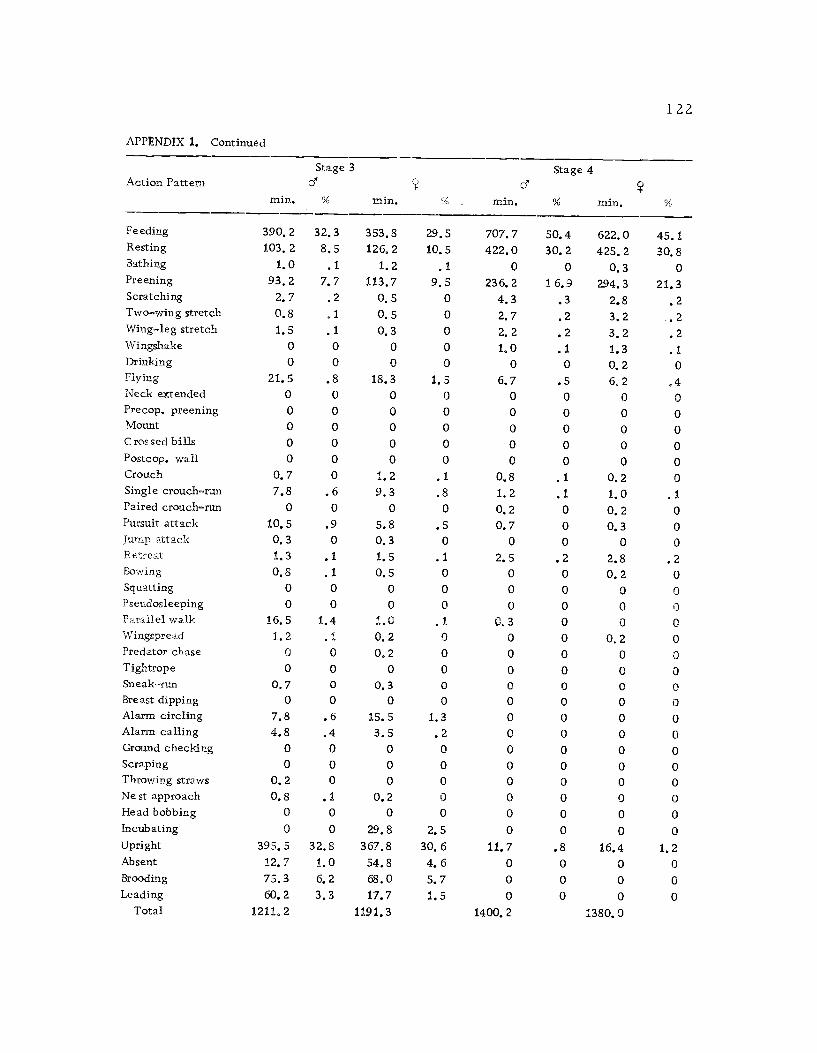
122
APPENDIX 1. Continued
Action PatternStage 3
0 0Stage 4
min. % min. min.
Feeding 390. 2 32.3 353.8 29.5 707. 7 50.4 622.0 45. 1Resting 103. 2 8.5 126. 2 10.5 422.0 30. 2 425. 2 30.8Bathing 1.0 .1 1.2 .1 0 0 0.3 0Preening 93.2 7.7 113.7 9.5 236.2 16.9 294.3 21.3Scratching 2.7 .2 0.5 0 4.3 .3 2.8 .2Two-wing stretch 0.8 . 1 0. 5 0 2. 7 2 3. 2 . 2Wing-leg stretch 1.5 1 0. 3 0 2. 2 . 2 3. 2 . 2Wingshake 0 0 0 0 1.0 1 1.3 .1Drinking 0 0 0 0 0 0 0.2 0Flying 21.5 .8 18.3 1.5 6.7 .5 6.2 .4Neck extended 0 0 0 0 0 0 0 0Precop, preening 0 0 0 0 0 0 0 0Mount 0 0 0 0 0 0 0 0Crossed bills 0 0 0 0 0 0 0 0Postcop. wall 0 0 0 0 0 0 0 0Crouch 0.7 0 1.2 .1 0.8 1 0.2 0Single crouch-run 7.8 .6 9.3 .8 1. 2 . 1 1.0 1
Paired crouch-run 0 0 0 0 0.2 0 0. 2 0Pursuit attack 10. 5 .9 5.8 .5 0. 7 0 0. 3 0jump attack 0. 3 0 0. 3 0 0 0 0 0P etrcat 1.3 .1 1.5 .1 2.5 .2 2.8 .2Bowing 0.8 .1 0. 5 0 0 0 0. 2 0Squatting 0 0 0 0 0 0 0 0Pseudosleeping 0 0 0 0 0 0 0 0Parallel walk 16.5 1.4 1.0 1 0.3 0 0 0Wingspread 1.2 .1 0.2 0 0 0 0.2 0Predator chase 0 0 0. 2 0 0 0 0 0Tightrope 0 0 0 0 0 0 0 0Sneak-run 0.7 0 0. 3 0 0 0 0 0Breast dipping 0 0 0 0 0 0 0 0Alarm circling 7.8 .6 15. 5 1. 3 0 0 0 0Alarm calling 4.8 .4 3. 5 . 2 0 0 0 0Ground checking 0 0 0 0 0 0 0 0Scraping 0 0 0 0 0 0 0 0Throwing straws 0. 2 0 0 0 0 0 0 0Nest approach 0.8 . 1 0.2 0 0 0 0 0Head bobbing 0 0 0 0 0 0 0 0Incubating 0 0 29.8 2. 5 0 0 0 0Upright 395.5 32.8 367.8 30.6 11.7 .8 16.4 1, 2Absent 12.7 I . 0 54.8 4.6 0 0 0 0Brooding 75.3 6. 2 68.0 5. 7 0 0 0 0Leading 60.2 3.3 17.7 1.5 0 0 0 0
Total 1211.2 1191.3 1400.2 1380.0
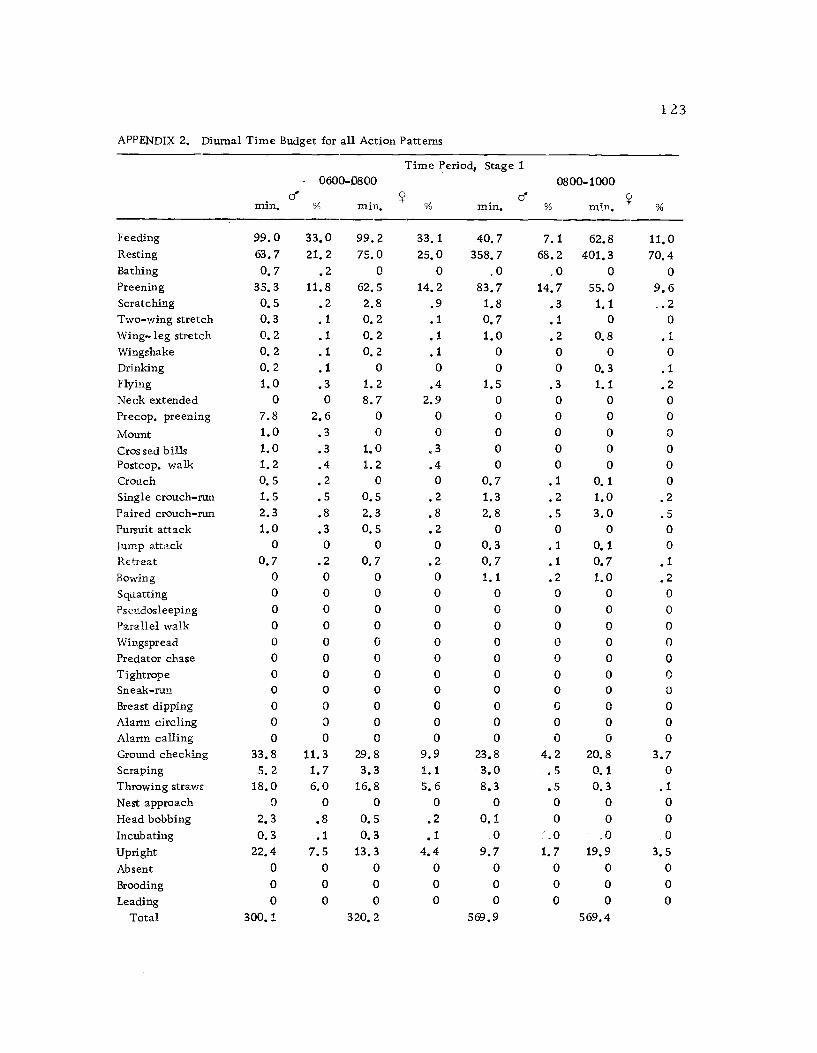
1 23
APPENDIX 2. Diurnal Time Budget for all Action Patterns
min.
0600-0800
9/. min,
Time Period, Stage 1
min.
0800-1000
min.
Feeding 99.0 33.0 99.2 33. 1 40.7 7. 1 62.8 11.0Resting 63.7 21. 2 75.0 25, 0 358.7 68.2 401, 3 70, 4Bathing 0.7 . 2 0 0 .0 , 0 0 0Preening 35, 3 11.8 62.5 14. 2 83.7 14, 7 55.0 9. 6Scratching 0, 5 .2 2.8 .9 1.8 .3 1, 1 . . 2Two-wing stretch 0.3 . 1 0. 2 ,1 0.7 .1 0 0Wing- leg stretch 0. 2 . 1 0. 2 . 1 1. 0 , 2 0.8 . 1
Wingshake 0, 2 . 1 0. 2 . 1 0 0 0 0Drinking 0. 2 ,1 0 0 0 0 0. 3 . 1
Flying 1.0 .3 1. 2 .4 1.5 .3 1. 1 . 2Neck extended 0 0 8.7 2.9 0 0 0 0Precop. preening 7.8 2. 6 0 0 0 0 0 0
Mount 1.0 .3 0 0 0 0 0 0
Crossed bills 1.0 .3 1.0 . 3 0 0 0 0Postcop. walk 1.2 .4 1.2 .4 0 0 0 0Crouch 0, 5 .2 0 0 0. 7 ,1 0, 1 0Single crouch-run 1. 5 .5 0.5 . 2 1.3 . 2 1.0 . 2Paired crouch-run 2.3 .8 2.3 .8 2.8 .5 3.0 , 5
Pursuit attack 1.0 . 3 0.5 .2 0 0 0 0Jump attack 0 0 0 0 0.3 . 1 0. 1 0Reheat 0.7 .2 0.7 .2 0.7 .1 0.7 .1Bowing 0 0 0 0 1. 1 .2 1.0 .2Squatting 0 0 0 0 0 0 0 0
Pseudosleeping 0 0 0 0 0 0 0 0Parallel walk 0 0 0 0 0 0 0 0
Wingspread 0 0 0 0 0 0 0 0
Predator chase 0 0 0 0 0 0 0 0Tightrope 0 0 0 0 0 0 0 0Sneak-run 0 0 0 0 0 0 0 0
Breast dipping 0 0 0 0 0 0 0 0Alarm circling 0 0 0 0 0 0 0 0Alarm calling 0 0 0 0 0 0 0 0Ground checking 33.8 11.3 29.8 9.9 23.8 4. 2 20.8 3.7Scraping 5. 2 1.7 3.3 1. 1 3.0 . 5 0. 1 0Throwing straws 18.0 6.0 16.8 5. 6 8.3 .5 0. 3 . 1
Nest approach 0 0 0 0 0 0 0 0
Head bobbing 2. 3 .8 0. 5 . 2 0. 1 0 0 0
Incubating 0.3 ,1 0. 3 . 1 0 , 0 .0 0
Upright 22.4 7.5 13.3 4.4 9.7 1.7 19.9 3.5Absent 0 0 0 0 0 0 0 0
Brooding 0 0 0 0 0 0 0 0
Leading 0 0 0 0 0 0 0 0
Total 300.1 320.2 569.9 569.4
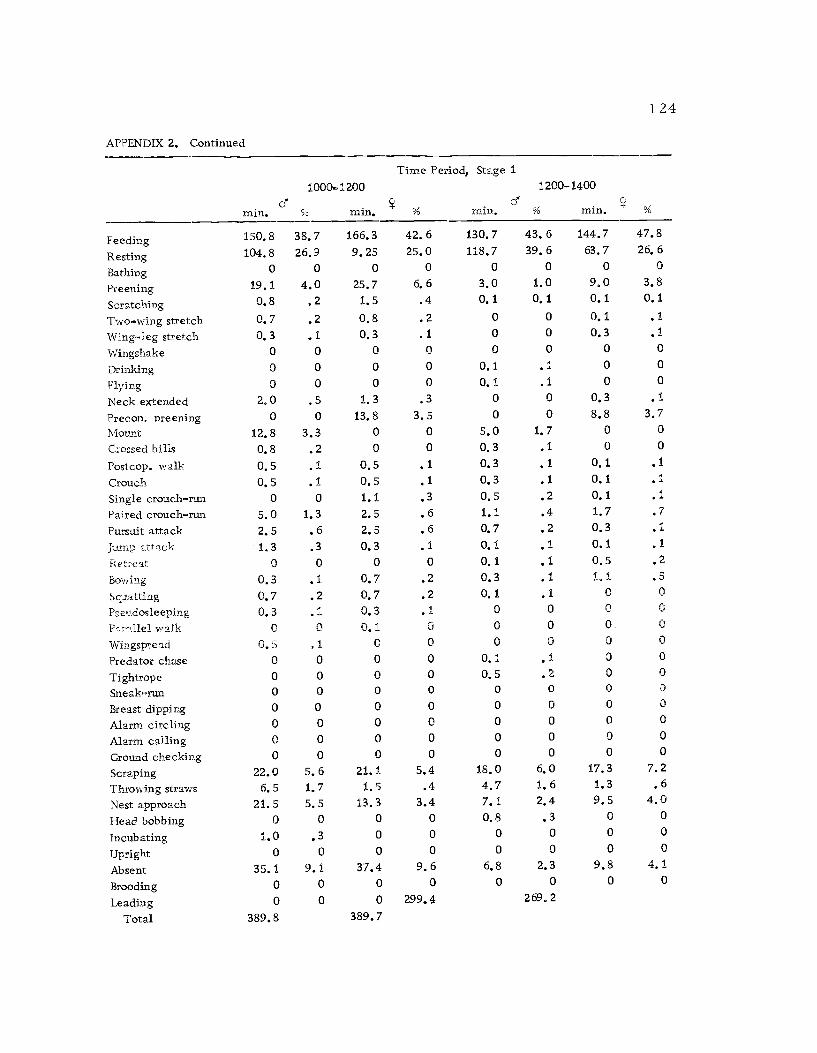
1 24
APPENDIX 2. Continued
ruin.
1000-1200
min.
Time Period, Stage 1
min.
1200-1400
ruin.
Feeding 150.8 38. 7 166.3 42. 6 130. 7 43. 6 144.7 47.8
Resting 104.8 26.9 9. 25 25.0 118.7 39. 6 63. 7 26. 6
Bathing 0 0 0 0 0 0 0 0
Preening 19.1 4.0 25.7 6.6 3.0 1.0 9.0 3.8
Scratching 0.8 .2 1.5 .4 0.1 0.1 0.1 0.1
Two-wing stretch 0.7 .2 0, 8 . 2 0 0 0. 1 . 1
Wing-leg stretch 0. 3 . 1 0.3 . 1 0 0 0.3 . 1
Wingshake 0 0 0 0 0 0 0 0
Drinking 0 0 0 0 0. 1 .1 0 0
Flying 0 0 0 0 0.1 .1 0 0
Neck extended 2.0 .5 1. 3 .3 0 0 0.3 i
Precon. preening 0 0 13.8 3.5 0 0 8.8 3.7Mount 12.8 3.3 0 0 5.0 1.7 0 0
Crossed bills 0. 8 .2 0 0 0. 3 .1 0 0
Postcop. walk 0. 5 .1 0.5 .1 0.3 .1 0. 1 .1Crouch 0. 5 . 1 0.5 . 1 0. 3 . 1 0. 1 . 1
Single crouch-run 0 0 1. 1 .3 0. 5 .2 0. 1 . 1
Paired crouch-run 5.0 1.3 2.5 . 6 1. 1 .4 1. 7 .7Pursuit attack 2. 5 . 6 2.5 . 6 0.7 . 2 0.3 . 1
J7.uulp attack 1. 3 .3 0. 3 . 1 0. 1 . 1 O. 1 . I
Retreat 0 0 0 0 O. 1 . 1 0.5 .2Bowing 0.3 .1 0.7 .2 0.3 .1 1.1 .5Sq:zatting 0.7 . 2 0. 7 . 2 0. 1 ,1 0 0
Ps eudosleeping O. 3 .1 0.3 . 1 0 0 0 0
Ps: ',11e1 walk 0 0 0,1 0 0 0 0 0
Wingspread 0.5 .1 0 0 0 0 0 0
Predator chase 0 0 0 0 0. 1 . 1 0 0
Tightrope 0 0 0 0 0, 5 . 2 0 0
Sneak-run 0 0 0 0 0 0 0 0
Breast dipping 0 0 0 0 0 0 0 0
Alarm circling 0 0 0 0 0 0 0 0
Alarm calling 0 0 0 0 0 0 0 0
Ground checking 0 0 0 0 0 0 0 0
Scraping 22.0 5. 6 21.1 5.4 18.0 6.0 17.3 7. 2
Throwing straws 6.5 1.7 1.5 .4 4.7 1. 6 1.3 . 6
Nest approach 21.5 5. 5 13.3 3.4 7. 1 2.4 9.5 4.0Head bobbing 0 0 0 0 0.8 . 3 0 0
Incubating 1.0 .3 0 0 0 0 0 0
Upright 0 0 0 0 0 0 0 0
Absent 35.1 9. 1 37.4 9. 6 6, 8 2.3 9.8 4. 1
Brooding 0 0 0 0 0 0 0 0
Leading 0 0 0 299. 4 269. 2
Total 389.8 389.7
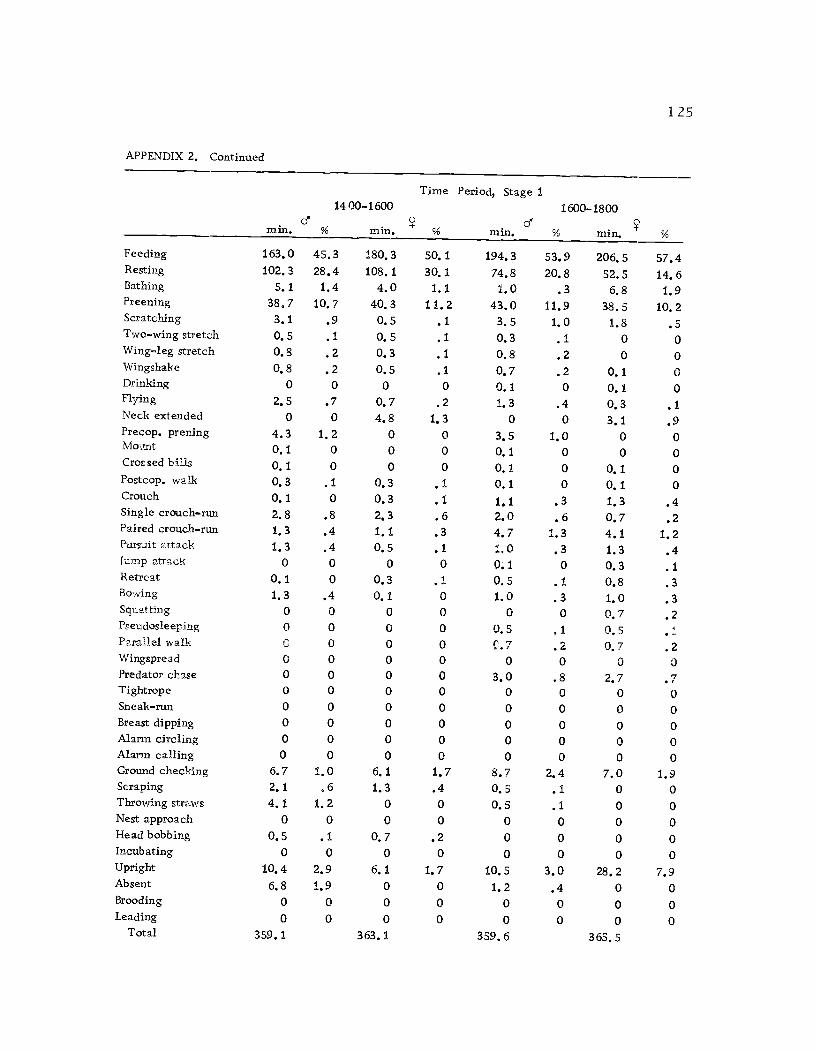
1 25
APPENDIX 2. Continued
min.
14
%
00-1600
min.
Time Period, Stage 11600-1800
d`min. min.
Feeding 163.0 45.3 180.3 50.1 194.3 53.9 206.5 57.4Resting 102. 3 28.4 108. 1 30. 1 74.8 20.8 52.5 14. 6Bathing S.1 1.4 4.0 1.1 1.0 .3 6.8 1.9Preening 38.7 10.7 40.3 1 1.2 43.0 11.9 38.5 10.2Scratching 3.1 .9 0.5 .1 3.5 1.0 1.8 .5Two-wing stretch 0. 5 .1 0. 5 .1 0.3 . 1 0 0Wing-leg stretch 0.8 .2 0. 3 . 1 0.8 .2 0 0Wingshake 0.8 .2 0.5 .1 0.7 .2 0. 1 0Drinking 0 0 0 0 0. 1 0 0. 1 0Flying 2.5 .7 0.7 .2 1.3 .4 0.3 .1Neck extended 0 0 4.8 1.3 0 0 3. 1 .9Precop. prening 4.3 1.2 0 0 3.5 1.0 0 0Mount 0.1 0 0 0 0.1 0 0 0Crossed bills 0. 1 0 0 0 0. 1 0 0. 1 0Postcop. walk 0. 3 .1 0.3 .1 0. 1 0 0. 1 0Crouch 0. 1 0 0.3 . 1 1. 1 .3 1.3 .4Single crouch-run 2. 8 .8 2. 3 .6 2.0 . 6 0.7 .2Paired crouch-run 1.3 .4 1. 1 .3 4.7 1.3 4.1 1.2Pursuit attack 1.3 .4 0.5 . 1 1.0 .3 1.3 .4Jump attack 0 0 0 0 0. 1 0 0.3 .1Retreat 0.1 0 0.3 .1 0.5 .1 0.8 .3Bowing 1.3 .4 0.1 0 1.0 .3 1.0 .3Squatting 0 0 0 0 0 0 0.7 .2Pseudosleeping 0 0 0 0 0.5 . 1 0.5 . 1Parallel walk 0 0 0 0 0.7 .2 0.7 .2Wingspread 0 0 0 0 0 0 0 0Predator chase 0 0 0 0 3.0 .8 2.7 .7Tightrope 0 0 0 0 0 0 0 0Sneak-run 0 0 0 0 0 0 0 0Breast dipping 0 0 0 0 0 0 0 0Alarm circling 0 0 0 0 0 0 0 0Alarm calling 0 0 0 0 0 0 0 0Ground checking 6.7 1.0 6. 1 1.7 8.7 2.4 7.0 1.9Scraping 2.1 .6 1.3 .4 0.5 .1 0 0Throwing straws 4. 1 1.2 0 0 0.5 . 1 0 0Nest approach 0 0 0 0 0 0 0 0Head bobbing 0.5 . 1 0. 7 .2 0 0 0 0Incubating 0 0 0 0 0 0 0 0Upright 10.4 2.9 6.1 1.7 10.5 3.0 28.2 7.9Absent 6.8 1.9 0 0 1.2 .4 0 0Brooding 0 0 0 0 0 0 0 0Leading 0 0 0 0 0 0 0 0
Total 359.1 363.1 359.6 365.5
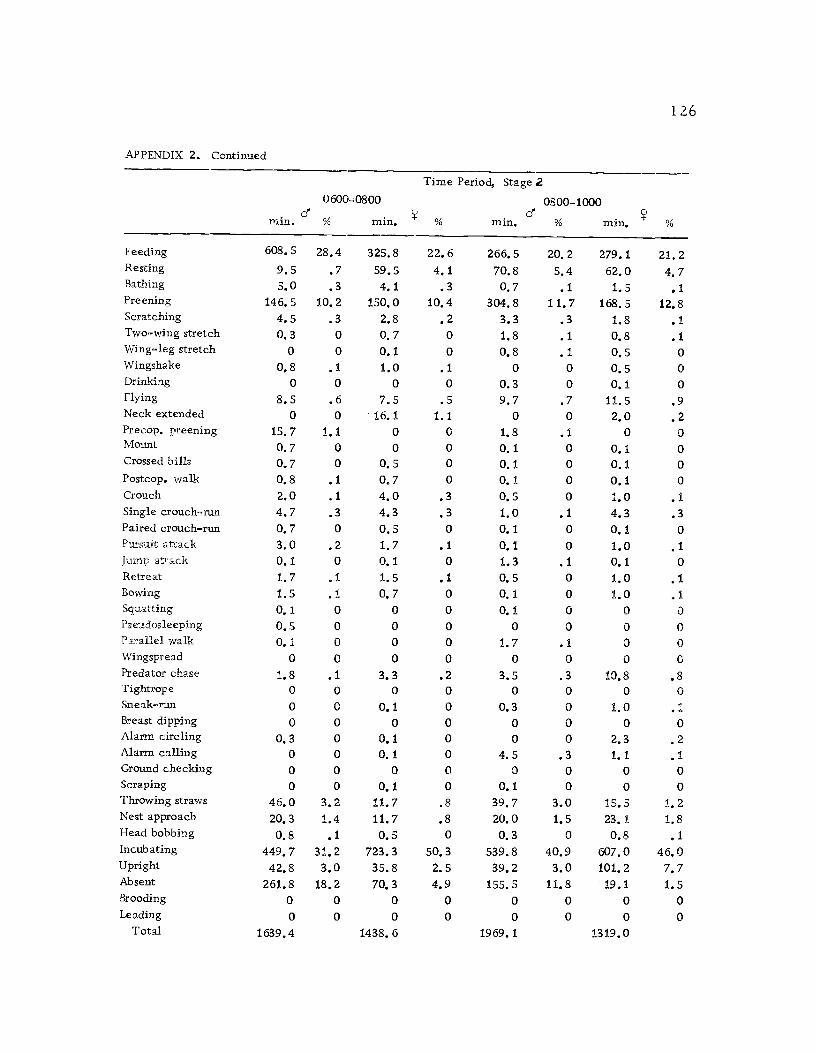
126
APPENDIX 2, Continued
dmin.
0600-; 0800
min.
Time Period, Stage 2
dmin.0800-1000
min.
Feeding 608.5 28.4 325.8 22.6 266.5 20.2 279.1 21,2Resting 9.5 .7 59.5 4.1 70.8 5.4 62.0 4.7Bathing 5.0 . 3 4.1 . 3 0.7 . 1 1.5 . 1Preening 146.5 10.2 150.0 10.4 304.8 1 1.7 168.5 12.8Scratching 4.5 .3 2.8 . 2 3.3 .3 1.8 . 1Two-wing stretch 0.3 0 0.7 0 1.8 ,1 0.8 . 1Wing-leg stretch 0 0 0.1 0 0.8 ,1 0.5 0Wingshake 0.8 . 1 1.0 ,1 0 0 0.5 0Drinking 0 0 0 0 0.3 0 0,1 0Flying 8.5 .6 7.5 .5 9.7 .7 11.5 .9Neck extended 0 0 16.1 1.1 0 0 2.0 . 2Pr econ. preening 15.7 1.1 0 0 1.8 ,1 0 0Mount 0.7 0 0 0 0.1 0 0.1 0Crossed bills 0.7 0 0.5 0 0.1 0 0.1 0Postcop. walk 0.8 ,1 0.7 0 0.1 0 0.1 0Crouch 2.0 . 1 4.0 , 3 0.5 0 1.0 .1Single crouch-run 4.7 .3 4.3 , 3 1.0 . 1 4.3 .3Paired crouch-run 0.7 0 0.5 0 0.1 0 0.1 0Palsuit attack 3.0 . 2 1.7 . 1 0.1 0 1.0 . 1juinp at-rs=_ck 0.1 0 0.1 0 1.3 . 1 0.1 0Retreat 1.7 .1 1.5 . 1 0.5 0 1.0 . 1Bowing 1.5 I 0.7 0 0.1 0 1.0 . 1Squatting 0,1 0 0 0 0.1 0 0 0Pseu.dosleeping 0.5 0 0 0 0 0 0 0Parallel walk 0.1 0 0 0 1.7 . 1 0 0Wingspread 0 0 0 0 0 0 0 0Predator chase 1.8 .1 3.3 .2 3.5 .3 10.8 .8Tightrope 0 0 0 0 0 0 0 0Sneak-run 0 0 0.1 0 0.3 0 1.0 ,1Breast dipping 0 0 0 0 0 0 0 0Alarm circling 0.3 0 0.1 0 0 0 2.3 . 2Alarm calling 0 0 0.1 0 4.5 . 3 1.1 . 1Ground checking 0 0 0 0 0 0 0 0Scraping 0 0 0.1 0 0.1 0 0 0Throwing straws 46.0 3.2 11.7 .8 39,7 3.0 15.5 1.2Nest approach 20.3 1.4 11.7 , 8 20.0 1.5 23.1 1,8Head bobbing 0.8 .1 0.5 0 0.3 0 0.8 . 1Incubating 449.7 31.2 723.3 50.3 539.8 40.9 607.0 46.0Upright 42.8 3.0 35.8 2.5 39.2 3.0 101.2 7.7Absent 261.8 18.2 70.3 4.9 155.5 11.8 19.1 1.5Brooding 0 0 0 0 0 0 0 0Leading 0 0 0 0 0 0 0 0
Total 1639.4 1438.6 1969.1 1319.0
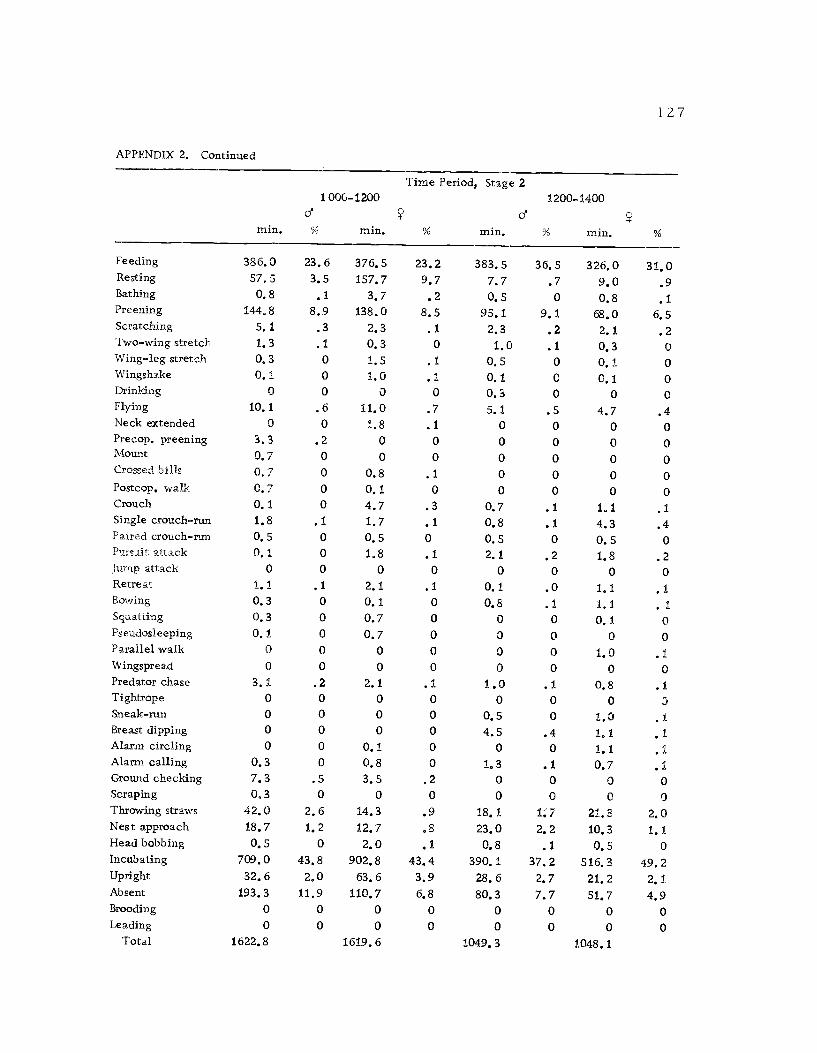
127
APPENDIX 2. Continued
min.
1000 -1200
% min.
Time Period, Stage 2
c/i min.
1200-1400
ruin.
Feeding 386.0 23.6 376.5 23.2 383.5 36.5 326.0 31, 0Resting 57.5 3.5 157.7 9.7 7.7 .7 9.0 .9Bathing 0.8 . 1 3.7 . 2 0.5 0 0.8 . 1Preening 144.8 8.9 138.0 8.5 95. 1 9. 1 68, 0 6.5Scratching 5. 1 .3 2.3 .1 2.3 .2 2. 1 . 2Two-wing stretch 1.3 . 1 0.3 0 1.0 . 1 0.3 0Wing-leg stretch 0.3 0 1.5 . 1 0.5 0 0. 1 0Wingshake 0. 1 0 I . 0 .1 0. 1 0 0. 1 0Drinking 0 0 0 0 0.3 0 0 0Flying 10.1 .6 11.0 .7 5.1 .5 4.7 .4Neck extended 0 0 1.8 . 1 0 0 0 0Precop. preening 3.3 .2 0 0 0 0 0 0Mount 0.7 0 0 0 0 0 0 0Crossed sills 0.7 0 0.8 .1 0 0 0 0Postcop. walk 0.7 0 0. 1 0 0 0 0 0Crouch 0.1 0 4.7 .3 0.7 .1 1.1 .1Single crouch-run 1.8 . 1 1.7 . 1 0.8 . 1 4.3 .4Paired crouch-run 0, 5 0 0. 5 0 0. 5 0 0. 5 0Purs_iit attack 0. 1 0 1.8 . 1 2. I . 2 1.8 .2Jump attack 0 0 0 0 0 0 0 0Retreat 1. 1 . 1 2.1 .1 0. 1 .0 1. 1 . 1Bowing 0.3 0 0. 1 0 0.8 . 1 1. 1Squatting 0.3 0 0.7 0 0 0 0. 1 0Ps euslosleeping 0.1 0 0. 7 0 0 0 0 0Parallel walk 0 0 0 0 0 0 1.0 .1Wingspread 0 0 0 0 0 0 0 0Predator chase 3. 1 .2 2. 1 . 1 1 .0 . 1 0.8 .1Tightrope 0 0 0 0 0 0 0 0Sneak-run 0 0 0 0 0.5 0 1.0 .1Breast dipping 0 0 0 0 4.5 .4 1, 1 , 1Alarm circling 0 0 0. 1 0 0 0 1. 1 .1Alarm calling 0.3 0 0.8 0 1.3 . 1 0.7 . 1Ground checking 7.3 .5 3. 5 . 2 0 0 0 0Scraping 0.3 0 0 0 0 0 0 0Throwing straws 42.0 2. 6 14.3 .9 18. 1 1. 7 21.3 2.0Nest approach 18.7 1. 2 12.7 .8 23.0 2. 2 10.3 1.1Head bobbing 0. 5 0 2, 0 .1 0, 8 .1 0, 5 0Incubating 709.0 43.8 902.8 43.4 390.1 37.2 516.3 49. 2Upright 32.6 2.0 63.6 3.9 28. 6 2.7 21.2 2. 1Absent 193.3 11.9 110.7 6.8 80.3 7.7 51.7 4.9Brooding 0 0 0 0 0 0 0 0Leading 0 0 0 0 0 0 0 0
Total 1622.8 1619.6 1049.3 1048.1
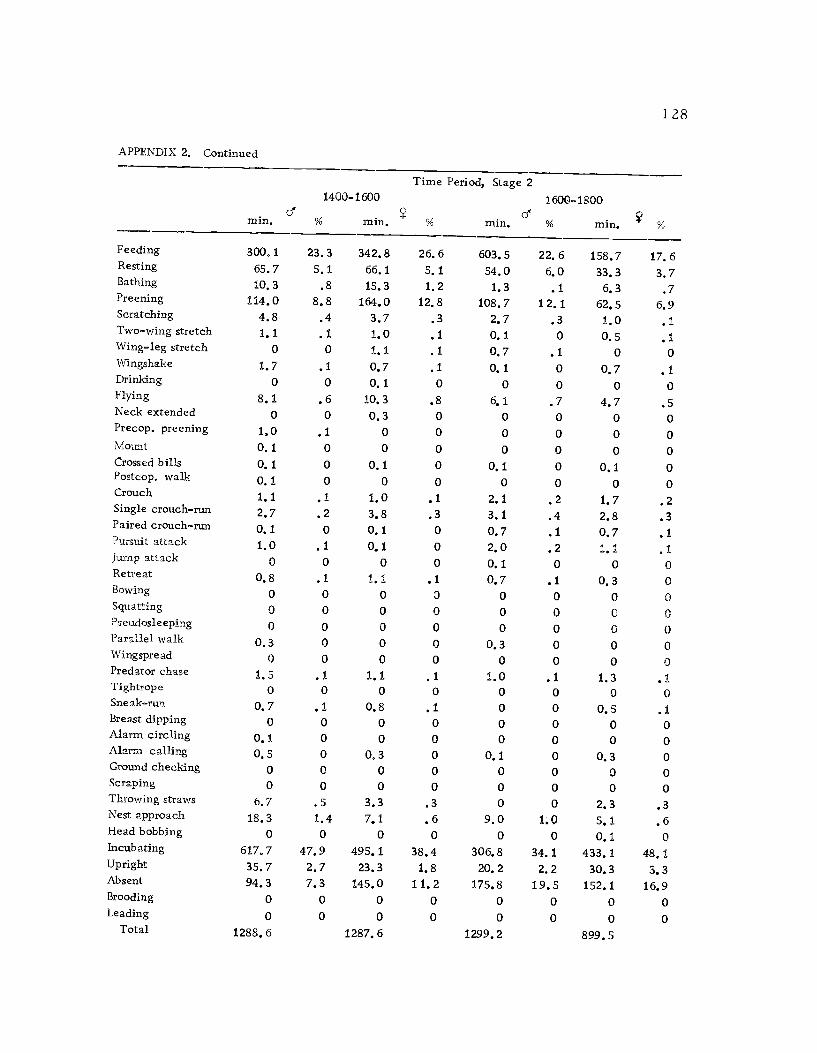
1 28
APPENDIX 2. Continued
min.
1400-1600cf` .mm.
Time Period, Stage 2
cf`min.
1600-1800
min.
Feeding 300. 1 23.3 342.8 26. 6 603.5 22. 6 158.7 17. 6Resting 65.7 5. 1 66.1 5. 1 54.0 6.0 33.3 3.7Bathing 10.3 .8 15.3 1. 2 1.3 . 1 6.3 .7Preening 114.0 8.8 164.0 12.8 108.7 1 2. 1 62.5 6.9Scratching 4.8 .4 3.7 .3 2.7 .3 1.0 ./Two -wing stretch 1. 1 . 1 1.0 . 1 0. 1 0 0. 5 . 1Wing-leg stretch 0 0 1. 1 . 1 0.7 1 0 0Wingshake 1.7 .1 0.7 . 1 0. 1 0 0. 7 ,1Drinking 0 0 0. 1 0 0 0 0 0Flying 8. 1 . 6 10.3 .8 6, 1 . 7 4.7 .5Neck extended 0 0 0.3 0 0 0 0 0Precop. preening 1.0 .1 0 0 0 0 0 0Mount 0.1 0 0 0 0 0 0 0Crossed bills 0. 1 0 0. 1 0 0. 1 0 0,1 0Postcop. walk 0. 1 0 0 0 0 0 0 0Crouch 1. 1 ,1 1.0 .1 2. 1 .2 1.7 .2Single crouch-run 2.7 . 2 3.8 .3 3. 1 .4 2.8 .3Paired crouch-run 0. 1 0 0. 1 0 0.7 .1 0.7 . 1Pursuit attack 1.0 . 1 0. 1 0 2. 0 . 2 1. 1 . 1jump attack 0 0 0 0 0. 1 0 0 0Retreat 0 8 .1 1. 1 .1 0.7 .1 0.3 0Bowing 0 0 0 0 0 0 0 0Squatting 0 0 0 0 0 0 0 0Pseudosleeping 0 0 0 0 0 0 0 0Parallel walk 0.3 0 0 0 0.3 0 0 0Wingspread 0 0 0 0 0 0 0 0Predator chase 1. 5 . 1 1. 1 . 1 1.0 . 1 1.3 . 1Tightrope 0 0 0 0 0 0 0 0Sneak -run 0. 7 . 1 0.8 . 1 0 0 0.5 .1Breast dipping 0 0 0 0 0 0 0 0Alarm circling 0. 1 0 0 0 0 0 0 0Alarm calling 0.5 0 0. 3 0 0. 1 0 0.3 0Ground checking 0 0 0 0 0 0 0 0Scraping 0 0 0 0 0 0 0 0Throwing straws 6.7 . 5 3.3 .3 0 0 2. 3 ,3Nest approach 18.3 1.4 7.1 . 6 9.0 1.0 5. 1 .6Head bobbing 0 0 0 0 0 0 0. 1 0Incubating 617.7 47.9 495. 1 38.4 306.8 34. 1 433. 1 48. 1Upright 35. 7 2.7 23.3 1.8 20. 2 2. 2 30.3 3.3Absent 94.3 7.3 145.0 1 1. 2 175.8 19.5 152.1 16.9Brooding 0 0 0 0 0 0 0 0Leading 0 0 0 0 0 0 0 0
Total 1288.6 1287.6 1299.2 899.5
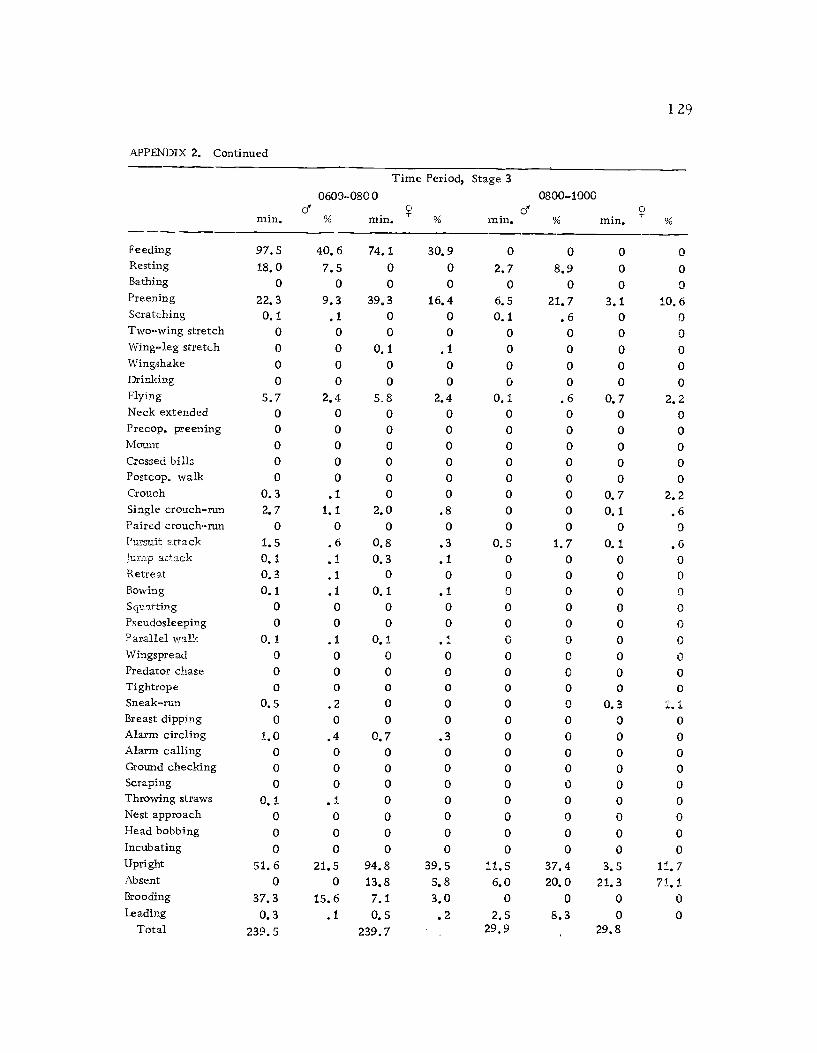
129
APPENDIX 2. Continued
min.
Time Period, Stage 30600-080 0
min. min.
0800-1000
min.
Feeding 97. 5 40. 6 74. 1 30, 9 0 0 0 0Resting 18.0 7, 5 0 0 2.7 8.9 0 0Bathing 0 0 0 0 0 0 0 0Preening 22.3 9.3 39.3 16.4 6.5 21.7 3. 1 10. 6Scratching 0. 1 ,1 0 0 0. 1 . 6 0 0Two-wing stretch 0 0 0 0 0 0 0 0Wing-leg stretch 0 0 0. 1 . 1 0 0 0 0Wingshake 0 0 0 0 0 0 0 0Drinking 0 0 0 0 0 0 0 0Flying 5.7 2.4 5.8 2.4 0. 1 . 6 0.7 2, 2Neck extended 0 0 0 0 0 0 0 0Precop. preening 0 0 0 0 0 0 0 0Mount 0 0 0 0 0 0 0 0Crossed bills 0 0 0 0 0 0 0 0Postcop. walk 0 0 0 0 0 0 0 0Crouch 0. 3 ,1 0 0 0 0 0. 7 2.2Single crouch-run 2. 7 1. 1 2, 0 .8 0 0 0. 1 . 6Paired crouch-run 0 0 0 0 0 0 0 0Pursuit attackck 1. 5 . 6 0.8 .3 0. 5 1. 7 0. 1 .6Jump attack 0, 1 .1 0, 3 ,1 0 0 0 0Retreat 0, 3 .1 0 0 0 0 0 0Bowing 0. 1 .1 0. 1 .1 0 0 0 0Squatting 0 0 0 0 0 0 0 0Pseudosleeping 0 0 0 0 0 0 0 0Parallel 1,t7.1.11z. 0. 1 . 1 0, 1 . 1 0 0 0 0Wingspread 0 0 0 0 0 0 0 0Predator chase 0 0 0 0 0 0 0 0Tightrope 0 0 0 0 0 0 0 0Sneak-run 0.5 .2 0 0 0 0 0.3 1. 1Breast dipping 0 0 0 0 0 0 0 0Alarm circling 1.0 .4 0, 7 .3 0 0 0 0Alarm calling 0 0 0 0 0 0 0 0Ground checking 0 0 0 0 0 0 0 0Scraping 0 0 0 0 0 0 0 0Throwing straws 0, 1 .1 0 0 0 0 0 0Nest approach 0 0 0 0 0 0 0 0Head bobbing 0 0 0 0 0 0 0 0Incubating 0 0 0 0 0 0 0 0Upright 51.6 21.5 94.8 39.5 11, 5 37.4 3.5 11.7Absent 0 0 13.8 5.8 6.0 20.0 21.3 71. 1Brooding 37.3 15.6 7. 1 3, 0 0 0 0 0Leading 0, 3 . 1 0, 5 .2 2.5 8.3 0 0
Total 239.5 239.7 29.9 29.8
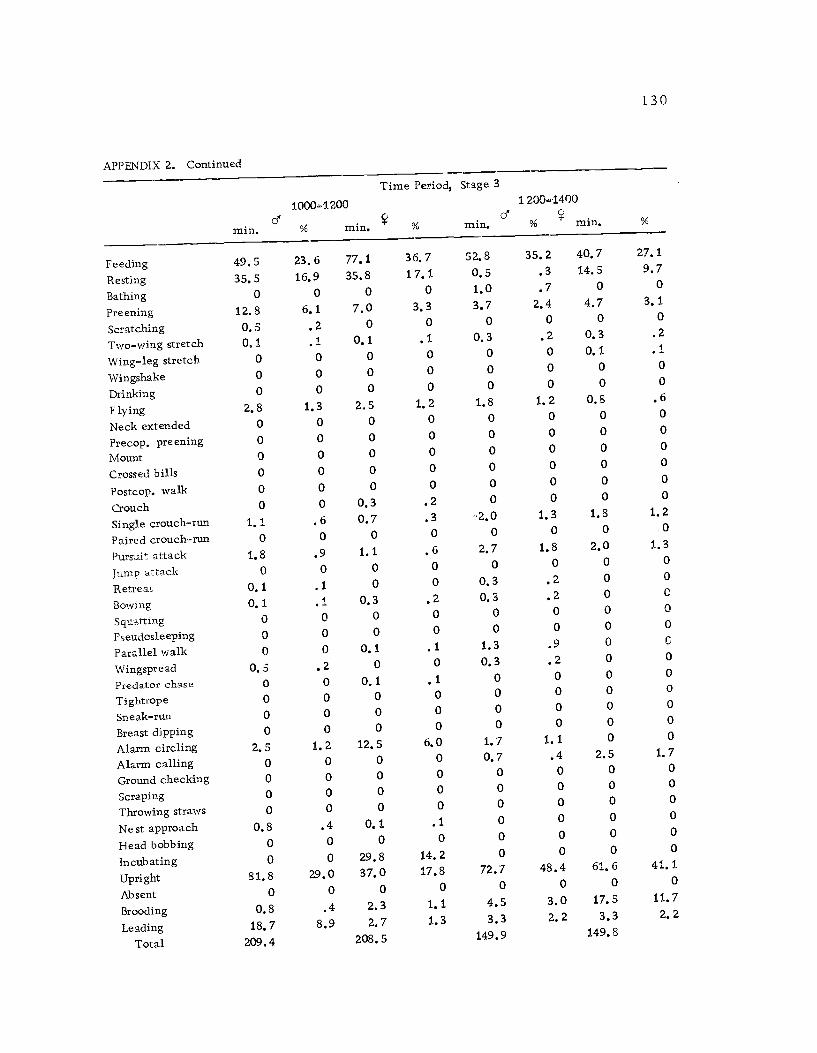
130
APPENDIX 2. Continued
min.
10001200d min.
Time Period, Stage 3
min.
1200.1400d' min.
Feeding 49.5 23. 6 77. 1 36. 7 52.8 35.2 40.7 27. 1
Resting 35,5 16.9 35.8 17. 1 0.5 .3 14.5 9.7
Bathing 0 0 0 0 1.0 .7 0 0
Preening 12.8 6.1 7.0 3.3 3.7 2.4 4.7 3.1
Scratching 0.5 .2 0 0 0 0 0 0
Two-wing stretch 0. 1 ./ 0. 1 . 1 0. 3 .2 0. 3 .2
Wing-leg stretch 0 0 0 0 0 0 0. 1 _ . 1
Wingshake 0 0 0 0 0 0 0 0
Drinking 0 0 0 0 0 0 0 0
F lying 2.8 1.3 2.5 1. 2 1.8 1. 2 0.8 .6
Neck extended 0 0 0 0 0 0 0 0
Precop. preening 0 0 0 0 0 0 0 0
Mount 0 0 0 0 0 0 0 0
Crossed bills 0 0 0 0 0 0 0 0
Postcop. walk 0 0 0 0 0 0 0 0
Crouch 0 0 0.3 .2 0 0 0 0
Single crouch-run 1. 1 . 6 0.7 .3 2.0 1.3 1.8 1. 2
Paired crouch-run 0 0 0 0 0 0 0 0
Pursuit attack 1,8 .9 1. 1 .6 2.7 1.8 2.0 1.3
Jump attack 0 0 0 0 0 0 0 0
Retreat 0.1 .1 0 0 0.3 .2 0 0
Bowing 0. 1 .1 0.3 .2 0. 3 .2 0 0
Sci :luting 0 0 0 0 0 0 0 0
Pseudosleeping 0 0 0 0 0 0 0 0
Parallel walk 0 0 0. 1 .1 1.3 .9 0 0
Wingspread 0. 5 .2 0 0 0, 3 . 2 0 0
Predator chase 0 0 0. 1 .1 0 0 0 0
Tightrope 0 0 0 0 0 0 0 0
Sneak-run 0 0 0 0 0 0 0 0
Breast dipping 0 0 0 0 0 0 0 0
Alarm circling 2. 5 1. 2 12. 5 6.0 1. 7 1. 1 0 0
Alarm calling 0 0 0 0 0.7 .4 2.5 1. 7
Ground checking 0 0 0 0 0 0 0 0
Scraping 0 0 0 0 0 0 0 0
Throwing straws 0 0 0 0 0 0 0 0
Nest approach 0.8 .4 0. 1 .1 0 0 0 0
Head bobbing 0 0 0 0 0 0 0 0
Incubating 0 0 29.8 14.2 0 0 0 0
Upright 81.8 29.0 37.0 17.8 72.7 48.4 61.6 41.1
Absent 0 0 0 0 0 0 0 0
Brooding 0.8 .4 2.3 1.1 4.5 3.0 17.5 11.7
Leading 18.7 8.9 2.7 1.3 3,3 2.2 3.3 2.2
Total 209.4 208.5 149.9 149.8
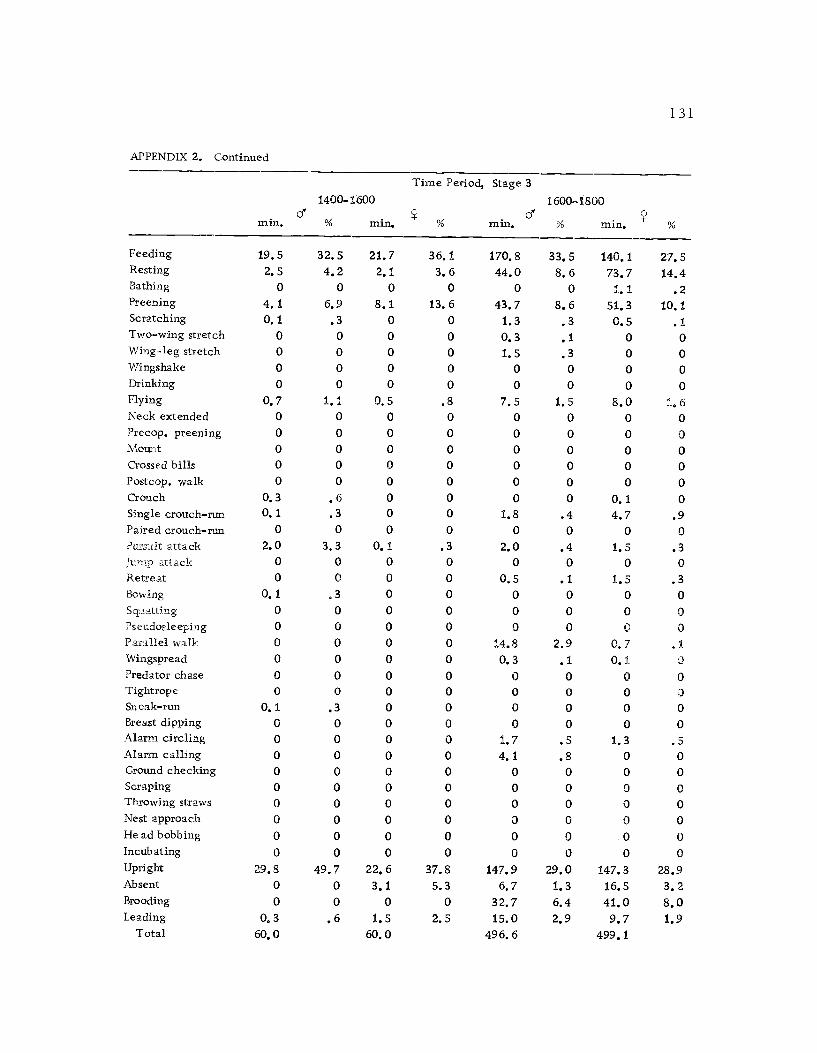
131
APPENDIX 2. Continued
mm.
1400-1600
ox,.
Time Period, Stage 3
min.
1600-1800
min.
Feeding 19.5 32.5 21.7 36. 1 170.8 33.5 140. 1 27.5Resting 2. 5 4.2 2. 1 3. 6 44.0 8. 6 73.7 14.4Bathing 0 0 0 0 0 0 1.1 .2Preening 4. 1 6.9 8. 1 13. 6 43.7 8. 6 51.3 10. 1Scratching 0. 1 .3 0 0 1.3 .3 0.5 . 1Two-wing stretch 0 0 0 0 0.3 . 1 0 0Wing-leg stretch 0 0 0 0 1. 5 .3 0 0Wingshake 0 0 0 0 0 0 0 0Drinking 0 0 0 0 0 0 0 0Flying 0.7 1. 1 0, 5 .8 7.5 1.5 8.0 1.6Neck extended 0 0 0 0 0 0 0 0Precop. preening 0 0 0 0 0 0 0 0Mount 0 0 0 0 0 0 0 0Crossed bills 0 0 0 0 0 0 0 0Postcop. walk 0 0 0 0 0 0 0 0Crouch 0.3 . 6 0 0 0 0 0. 1 0Single crouch-run 0. 1 . 3 0 0 1.8 .4 4.7 .9Paired crouch-rum 0 0 0 0 0 0 0 0P IlfEIJ it attack 2.0 3. 3 0. 1 .3 2.0 .4 1.5 .3rump attack 0 0 0 0 0 0 0 0Retreat 0 0 0 0 0.5 . 1 1.5 .3Bowing 0, 1 3 0 0 0 0 0 0Scatting 0 0 0 0 0 0 0 0Pseadosleeping 0 0 0 0 0 0 0 0Parallel walk 0 0 0 0 14.8 2.9 0. 7 . 1Wingspread 0 0 0 0 0. 3 .1 0. 1 0Predator chase 0 0 0 0 0 0 0 0Tightrope 0 0 0 0 0 0 0 0Sneak-run 0. 1 .3 0 0 0 0 0 0Breast dipping 0 0 0 0 0 0 0 0Alarm circling 0 0 0 0 1.7 .5 1.3 .5Alarm calling 0 0 0 0 4. 1 .8 0 0Ground checking 0 0 0 0 0 0 0 0Scraping 0 0 0 0 0 0 0 0Throwing straws 0 0 0 0 0 0 0 0Nest approach 0 0 0 0 0 0 0 0He ad bobbing 0 0 0 0 0 0 0 0Incubating 0 0 0 0 0 0 0 0Upright 29.8 49.7 22.6 37.8 147.9 29.0 147.3 28.9Absent 0 0 3. 1 5.3 6.7 1.3 16.5 3. 2Brooding 0 0 0 0 32. 7 6.4 41.0 8.0Leading 0.3 .6 1.5 2.5 15.0 2.9 9.7 1.9
Total 60. 0 60. 0 496. 6 499. 1
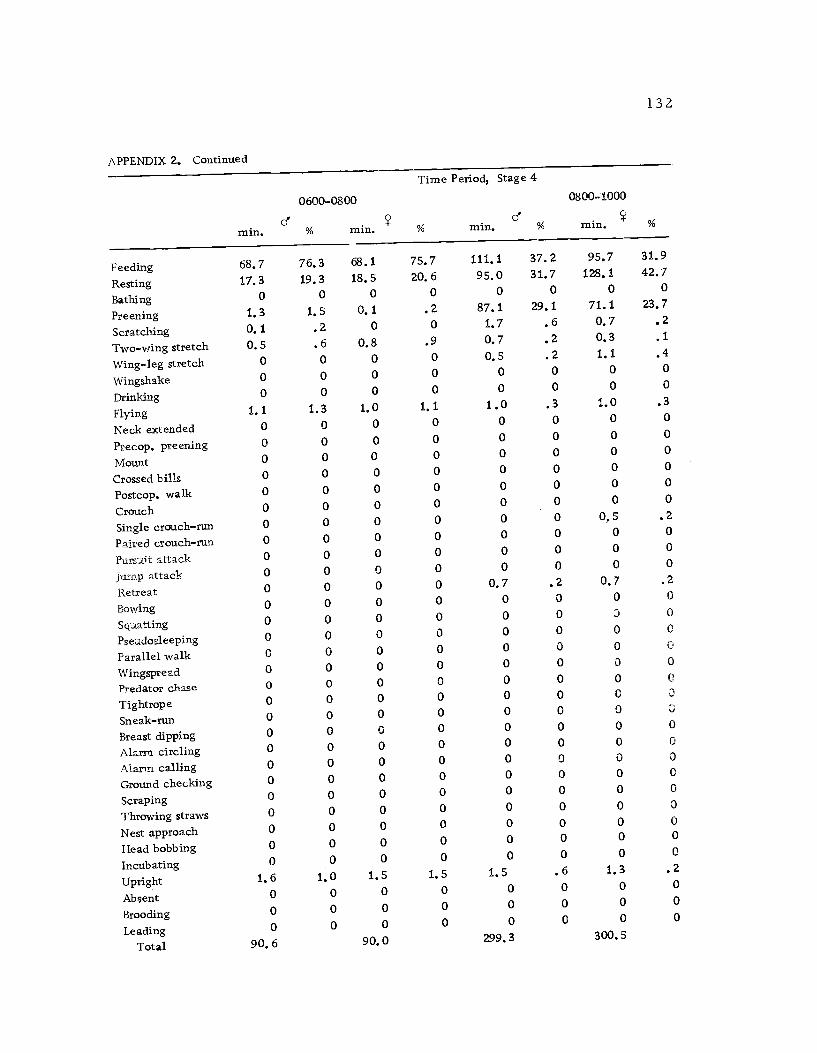
132
APPENDIX 2. Continued
min.
0600-0800
min.d
mm.
Time Period, Stage 4
min.
0800-1000
min.
Feeding 68.7 76.3 68.1 75.7 111.1 37.2 95.7 31.9
Resting 17.3 19.3 18.5 20.6 95.0 31.7 128.1 42.7
Bathing 0 0 0 0 0 0 0 0
Preening 1.3 1.5 0.1 .2 87.1 29.1 71.1 23.7
Scratching 0.1 .2 0 0 1.7 .6 0.7 .2
Two-wing stretch 0.5 .6 0.8 .9 0.7 .2 0.3 .1
Wing-leg stretch 0 0 0 0 0.5 . 2 1.1 .4
Wingshake 0 0 0 0 0 0 0 0
Drinking 0 0 0 0 0 0 0 0
Flying 1.1 1.3 1.0 1.1 1 . 0 .3 1.0 .3
Neck extended 0 0 0 0 0 0 0 0
Precop. preening 0 0 0 0 0 0 0 0
Mount 0 0 0 0 0 0 0 0
Crossed bills 0 0 0 0 0 0 0 0
Postcop. walk 0 0 0 0 0 0 0 0
Crouch 0 0 0 0 0 0 0 0
Single crouch-run 0 0 0 0 0 0 0,5 .2
Paired crouch-run 0 0 0 0 0 0 0 0
Purs'..,.it attack 0 0 0 0 0 0 0 0
Jump p attack 0 0 0 0 0 0 0 0
Retreat 0 0 0 0 0.7 .2 0.7 2
Bowing 0 0 0 0 0 0 0 0
Sq.Aatting 0 0 0 0 0 0 0 0
Pseadosleeping 0 0 0 0 0 0 0 0
Parallel walk 0 0 0 0 0 0 0 0
Wingspread 0 0 0 0 0 0 0 0
Predator chase 0 0 0 0 0 0 0 0
Tightrope 0 0 0 0 0 0 0 0
Sneak-run 0 0 0 0 0 0 0 0
Breast dipping 0 0 0 0 0 0 0 0
Alarm circling 0 0 0 0 0 0 0 0
Alarm calling 0 0 0 0 0 0 0 0
Ground checking 0 0 0 0 0 0 0 0
Scraping 0 0 0 0 0 0 0 0
Throwing straws 0 0 0 0 0 0 0 0
Nest approach 0 0 0 0 0 0 0 0
Head bobbing 0 0 0 0 0 0 0 0
Incubating 0 0 0 0 0 0 0 0
Upright 1.6 1.0 1.5 1.5 1.5 . 6 1.3 .2
Absent 0 0 0 0 0 0 0 0
Brooding 0 0 0 0 0 0 0 0
Leading 0 0 0 0 0 0 0 0
Total 90.6 90.0 299.3 300.5
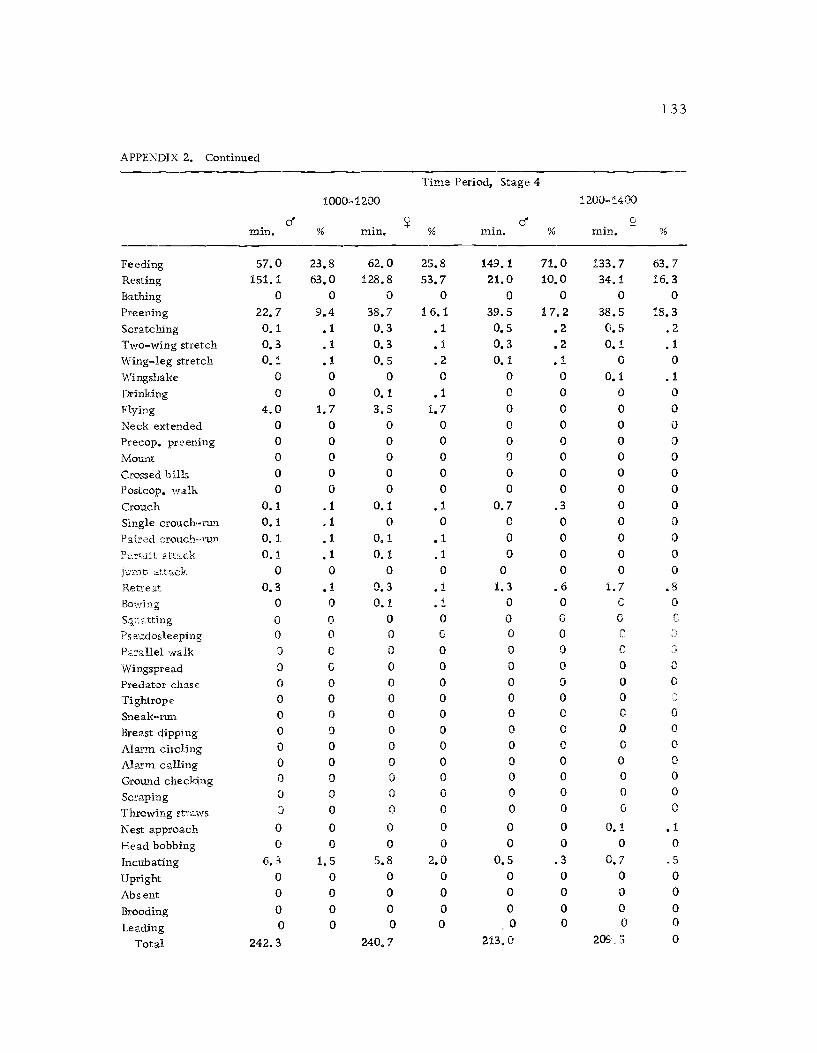
133
APPENDIX 2. Continued
10001200
min.
Time Period, Stage 4
min.
1200-1400
min.
FeedingRestingBathingPreening
57.0151.1
0
22.7
23.863.0
09.4
62.0128.8
038.7
25.853.7
01 6.1
149. 121.0
0
39.5
71.010.0
017. 2
133.734. 1
038.5
63.716.3
018.3
Scratching 0. 1 .1 0, 3 . 1 0. 5 . 2 0. 5 . 2
Two-wing stretch 0, 3 . 1 0, 3 . 1 0, 3 .2 0, 1 . 1
Wing-leg stretch 0. 1 ,1 0, 5 . 2 0. 1 . 1 0 0
Wingshake 0 0 0 0 0 0 0. 1 . 1
Drinking 0 0 0, 1 . 1 0 0 0 0
Flying 4.0 1.7 3.5 1, 7 0 0 0 0
Neck extended 0 0 0 0 0 0 0 0
Precop. preening 0 0 0 0 0 0 0 0
Mount 0 0 0 0 0 0 0 0
Crossed bills 0 0 0 0 0 0 0 0Postcop. walk 0 0 0 0 0 0 0 0
Crouch 0. 1 .1 0. 1 . 1 0.7 .3 0 0
Single crouch -ran 0, 1 . 1 0 0 0 0 0 0
P a ii° c d crouch-run 0. 1 . 1 0. 1 ,1 0 0 0 0
Purai 1. attack 0. 1 . 1 0. 1 . 1 0 0 0 0
J crnp attack. 0 0 0 0 0 0 0 0
Retreat 0, 3 . 1 0.3 . 1 1.3 . 6 1.7 .8Bowing 0 0 0. 1 .1 0 0 0 0
Seic,:_tting 0 0 0 0 0 0 0Pse.:dosleepinsz 0 0 0 0 0 0 0 0
Parallel walk 0 0 0 0 0 0 0
Wingspread 0 0 0 0 0 0 0 0
Predator chase 0 0 0 0 0 0 0 0
Tightrope 0 0 0 0 0 0 0
Sneak-run 0 0 0 0 0 0 0 0
Breast dipping 0 0 0 0 0 0 0 0
Alarm circling 0 0 0 0 0 0 0 0
Alarm calling 0 0 0 0 0 0 0 0
Ground checking 0 0 0 0 0 0 0 0
Scraping 0 0 0 0 0 0 0 0
Throwing straws 0 0 0 0 0 0 0 0
Nest approach 0 0 0 0 0 0 0, 1 . 1
Head bobbing 0 0 0 0 0 0 0 0
Incubating 6.3 1.5 5, 8 2.0 0.5 .3 0.7 .5Upright 0 0 0 0 0 0 0 0
Abs ent 0 0 0 0 0 0 0 0
Brooding 0 0 0 0 0 0 0 0
Leading 0 0 0 0 0 0 0 0
Total 242.3 240.7 213.0 20S', 3 0
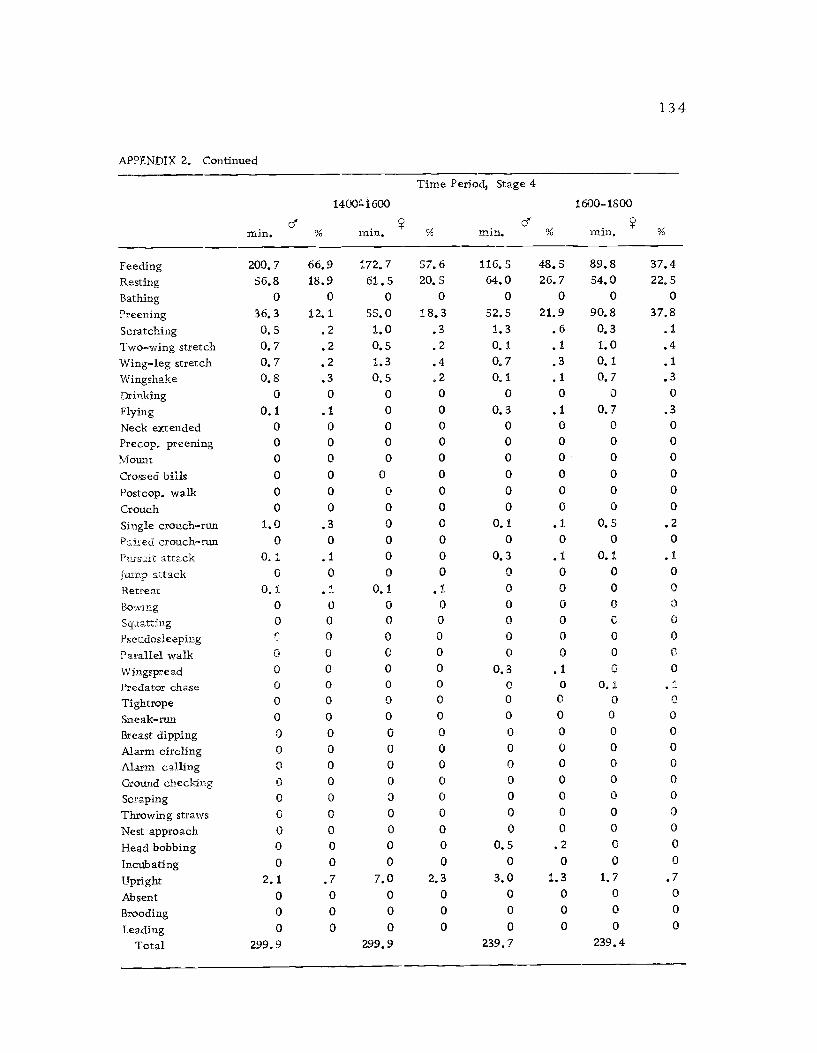
134
APPENDIX 2. Continued
min.
1400::1600
cr
Time Period, Stage 4
cr./D
1600-1800
min.
Feeding 200.7 66.9 172.7 57.6 116.5 48.5 89.8 37.4Resting 56.8 18.9 61.5 20.5 64.0 26.7 54.0 22.5Bathing 0 0 0 0 0 0 0 0
Preening 36.3 12. 1 55.0 18.3 52.5 21.9 90.8 37.8Scratching 0.5 .2 1.0 .3 1.3 .6 0.3 .1l'wo-wing stretch 0. 7 .2 0.5 . 2 0. 1 . 1 1.0 .4Wing-leg stretch 0. 7 .2 1.3 .4 0.7 .3 0. 1 . 1
Wingshake 0.8 .3 0.5 .2 0. 1 .1 0. 7 .3Drinking 0 0 0 0 0 0 0 0
Flying 0.1 .1 0 0 0.3 .1 0.7 .3Neck extended 0 0 0 0 0 0 0 0
Precop, preening 0 0 0 0 0 0 0 0
Mount 0 0 0 0 0 0 0 0
Crossed bills 0 0 0 0 0 0 0 0
Postcop. walk 0 0 0 0 0 0 0 0
Crouch 0 0 0 0 0 0 0 0
Single crouch-run 1.0 .3 0 0 0. 1 . 1 0. 5 .2Paired crouch-run 0 0 0 0 0 0 0 0
F'ris,..it attack 0. 1 .1 0 0 0.3 . 1 0. 1 . 1
Jump attack 0 0 0 0 0 0 0 0
Retreat 0.1 .1 0.1 .1 0 0 0 0
Bowing 0 0 0 0 0 0 0 0
Sciaatting 0 0 0 0 0 0 0 0
Pseudosleeping '') 0 0 0 0 0 0 0
Parallel walk 0 0 0 0 0 0 0 0
Wingspread 0 0 0 0 0. 3 .1 0 0
Predatca. chase 0 0 0 0 0 0 0. 1 .
Tightrope 0 0 0 0 0 0 0 0
Sneak-run 0 0 0 0 0 0 0 0
Breast dipping 0 0 0 0 0 0 0 0
Alarm circling 0 0 0 0 0 0 0 0
Alarm calling 0 0 0 0 0 0 0 0
Ground checking 0 0 0 0 0 0 0 0
Scraping 0 0 0 0 0 0 0 0
Throwing straws 0 0 0 0 0 0 0 0
Nest approach 0 0 0 0 0 0 0 0
Head bobbing 0 0 0 0 0. 5 . 2 0 0
Incubating 0 0 0 0 0 0 0 0
Upright 2.1 .7 7.0 2.3 3.0 1.3 1.7 .7Absent 0 0 0 0 0 0 0 0
Brooding 0 0 0 0 0 0 0 0
Leading 0 0 0 0 0 0 0 0
Total 299.9 299.9 239.7 239.4
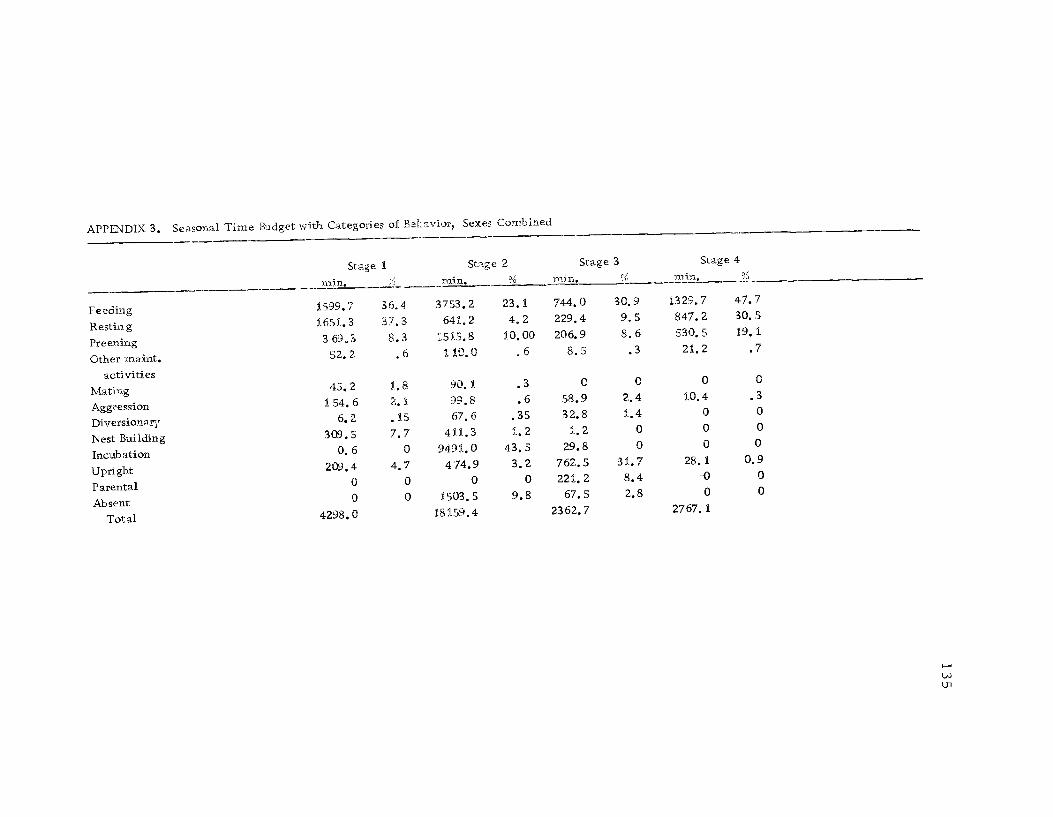
APPENDIX 3. Seasonal Time Radget with Categories of Behavior, Sexes Combined
Stage 1min.
Stage 2nin.
Stage 3 Stage 4rn
Feeding 1599.7 36. 4 3753.2 23.1 744.0 30.9 1329, 7 47.7
Restin g 1651.3 37. 3 641.2 4. 2 229.4 9. 5 847. 2 30. 5
Preening 3 69.3 8.3 .515.8 10.00 206.9 8. 6 530. 5 19. 1
Other maint.activities
52.2 .6 1 10. 0 .6 8.5 .3 21.2 .7
Mating 45.2 1.8 90.1 .3 0 0 0 CI
Aggression 1 54. 6 2.1 99.8 .6 58.9 2.4 10.4 .3
Diversionary 6.2 .15 67.6 .35 32.8 1. 4 0 0
Nest Building 309.5 7. 7 411.3 1, 2 1. 2 0 0 0
Incubation 0. 6 0 9491.0 43.5 29.8 0 0 0
Upright 209.4 4.7 4 74.9 3. 2 762.5 31.7 28.1 0.9
Parental 0 0 0 0 221.2 8.4 -0 0
Absent 0 0 1503.5 9.8 67.5 2.8 0 0
Total 4298.0 18199.4 2362.7 2767. 1
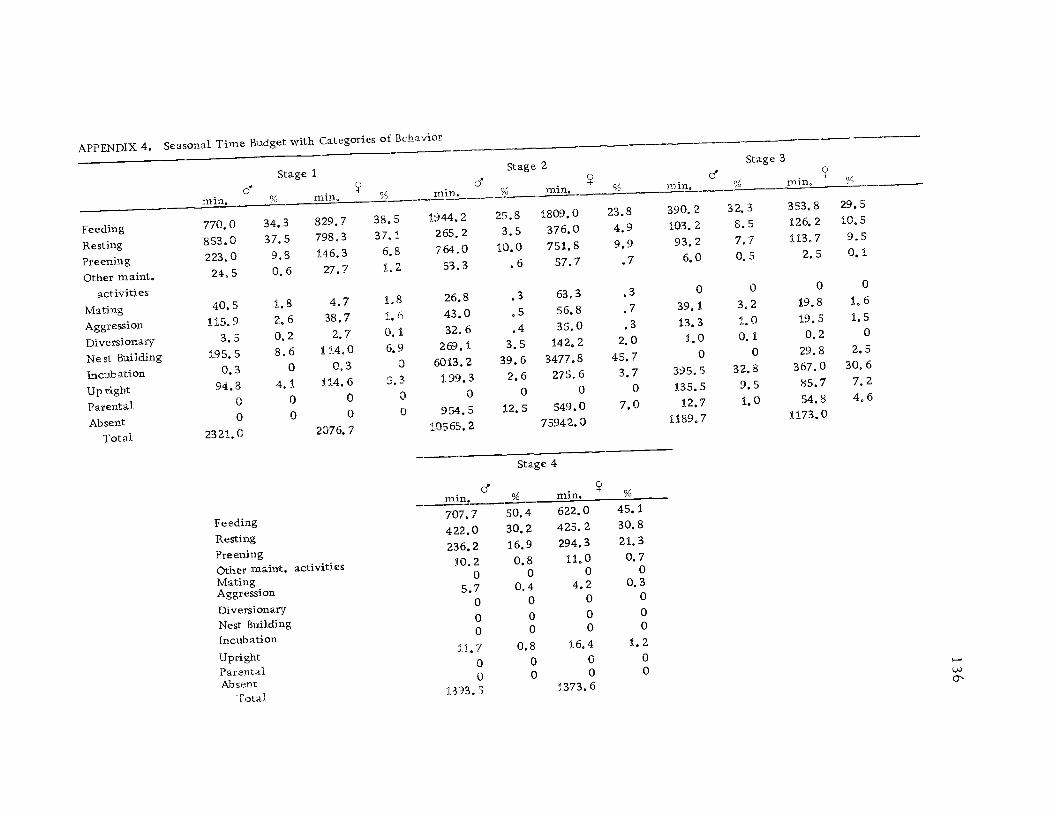
APPENDIX 4. Seasonal Time Budget with Categories of Behavior
Stage 1
min. min, min.
Stage 2
min.
Stage 3
Feeding 770.0 34.3 829.7 38, 5 1944, 2 25.8 1809.0 23,8 390.2 32.3 353.8 29. 5
Resting 853.0 37.5 798.3 37. 1 265, 2 3.5 376.0 4.9 103. 2 8. 5 126. 2 10.5
Preening 223.0 9.8 146.3 6.8 764,0 10.0 751.8 9.9 93.2 7,7 113.7 9.5
Other maint.activities
24.5 0. 6 27.7 1, 2 53.3 . 6 57.7 .7 6.0 0. 5 2.5 0. 1
Mating 40.5 1.8 4.7 1.8 26.8 .3 63.3 .3 0 0 0 0
Aggression 115.9 2. 6 38.7 1. i 43.0 . 5 56.8 .7 39. 1 3. 2 19.8 1. 6
Diversionary 3. 5 0. 2 2.7 0. 1 32. 6 .4 35.0 . 3 13. 3 1. 0 19. 5 1. 5
Nest Building 195.5 8. 6 1 14.0 6.9 269. 1 3. 5 142.2 2. 0 1.0 O. 1 0. 2 0
Incubation 0.3 0 0.3 0 6013.2 39.6 3477.8 45.7 0 0 29.8 2.5
Up right 94.8 4. 1 114.6 5. 3 1 99. 3 2. 6 275. 6 3. 7 395.5 32.8 367.0 30. 6
Parental 0 0 0 0 0 0 0 0 135.5 9, 5 85.7 7. 2
Absent 0 0 0 0 954.5 12, 5 549.0 7.0 12.7 1, 0 54.8 4. 6
Total 2321.0 2076.7 10565.2 75942.0 1189.7 1173.0
Stage 4
dmin. min.
Feeding707. 7 50.4 622.0 45. 1
Resting422.0 30.2 425. 2 30.8
Preening236.2 16.9 294.3 21. 3
Other maint. activities 10. 2 0.8 11.0 0. 7
Mating 0 0 0 0
Aggression 5.7 0.4 4. 2 0. 3
Diversionary0 0 0 0
Nest Building 0 0 0 0
Incubation0 0 0 0
Upright11.7 0.8 16.4 1. 2
Parental0 0 0 0
Absent0 0 0 0
Total1333. 5 1373. 6
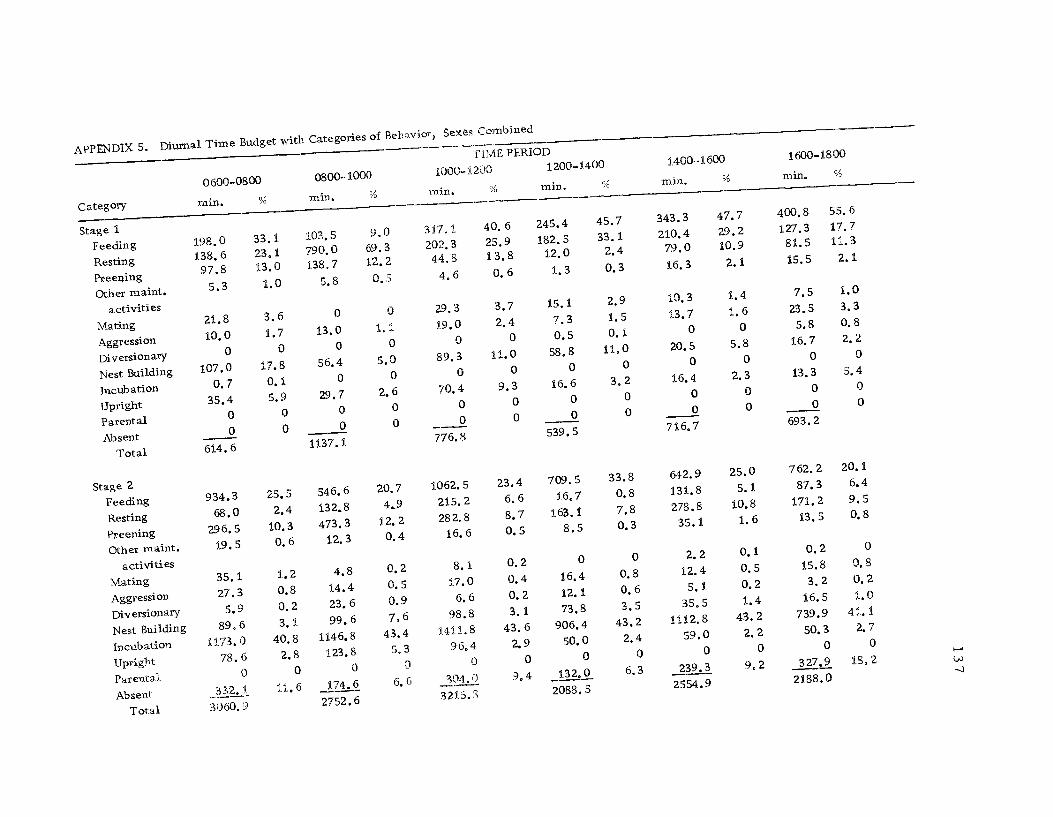
APPENDIX 5. Diurnal Time Budget with Categories of Behavior; Sexes Combined
Category
0600-0800 0800-1000
TIME PERIOD
1000-1200 1200-1400
min. min.
1400-1600
min.
1600-1860
min. CA
Stage 1Feeding 198. 0 33. 1 103.5 9.0 317. 1 40.6 245.4 45.7 343.3 47.7 400.8 55. 6
Resting 138. 6 23. 1 790.0 69.3 202. 3 25.9 182.5 33. 1 210.4 29. 2 127.3 17. 7
Preening 97.8 13.0 138.7 12.2 44.8 1 3. 8 12.0 2.4 79.0 10.9 81.5 11.3
Other maint.activities
5.3 1.0 5.8 0, 5 4. 6 0. 6 1.3 0.3 16. 3 2. 1 15. 5 2. 1
Mating 21.8 3.6 0 0 29.3 3.7 15.1 2.9 10.3 1.4 7.5 1.0
Aggression 10.0 1.7 13.0 1. 1 19.0 2.4 7.3 1.5 13.7 1. 6 23.5 3.3
Diversionary0 0 0 0 0 0 0.5 0. 1 0 0 5.8 0. 8
Nest Building 107.0 17.8 56.4 5.0 89.3 11.0 58.8 11, 0 20.5 5.8 16.7 2. 2
Incubation 0. 7 0. 1 0 0 0 0 0 0 0 0 0 0
Upright 35, 4 5.9 29.7 2. 6 70.4 9.3 16.6 3. 2 16.4 2.3 13.3 5.4
Parental 0 0 0 0 0 0 0 0 0 0 0 0
Absent 0 0 0 0 0 0 0 0 0 0 0 0
Total 614.6 1137.1 776.8 539.5 716.7 693.2
Stage 2Feeding 934.3 25.5 546.6 20.7 1062.5 23.4 709.5 33.8 642.9 25.0 762.2 20. 1
Resting 68.0 2.4 1328 4..9 215. 2 6. 6 16.7 0. 8 131, 8 5. 1 87. 3 6.4
Preening 296.5 10.3 473.3 12.2 282.8 8.7 163.1 7.8 278.8 10.8 171, 2 9.5
Other maint.activities
19.5 0.6 12.3 0.4 16. 6 0.5 8.5 0.3 35. 1 1.6 13. 5 0. 8
Mating 35.1 1.2 4.8 0. 2 8. 1 0.2 0 0 2. 2 0. 1 0.2 0
Aggression 27. 3 0.8 14.4 0, 5 17.0 0.4 16.4 0. 8 12.4 0. 5 15.8 0. S
Diversionary 5.9 0. 2 23.6 0.9 6. 6 0. 2 12. 1 0. 6 5, 1 0. 2 3. 2 0. 2
Nest Building 89.6 3. 1 99.6 7.6 98.8 3. 1 73.8 3. 5 35.5 1.4 16.5 1.0
Incubation 1173.0 40.8 1146.8 43.4 1411.8 43.6 906.4 43. 2 /112.8 43.2 739.9 41. 1
Upright 78, 6 2.8 123.8 5. 3 96.4 2.9 50.0 2.4 59.0 2. 2 50.3 2. 7
Parental 0 0 0 0 0 0 0 0 0 0 0 0
Absent 332,1 11, 6 174.6 6. 6 9, 4 132.0 6.3 239.3 9. 2 327.9 18. 2
Total 3060, 9 2752.6
_304.03213. 3 2088.5 2554.9 2188.0
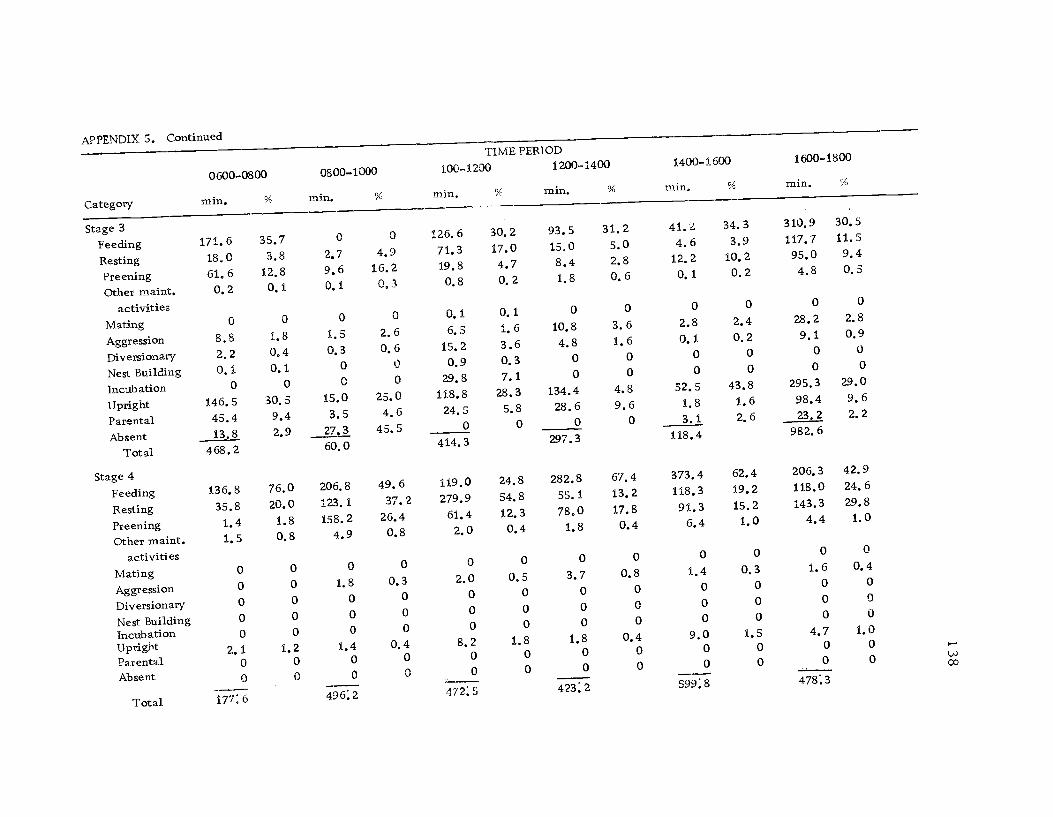
APPENDIX 5. ContinuedTIME PERIOD
0600-0800 0800-1000 100-1200 1200-1400 1400-1600 1600-1800
Category min. min. min. min. min. min. %
Stage 3Feeding 171.6 35.7 0 0 126.6 30.2 93.5 31.2 41.2 34.3 310.9 30, 5
Resting 18.0 3.8 2.7 4.9 71.3 17.0 15.0 5.0 4.6 3.9 117.7 11.5
Preening 61.6 12.8 9.6 16.2 19.8 4.7 8.4 2.8 12.2 10.2 95.0 9.4
Other maint.activities
0.2 0. 1 0. 1 0.3 0.8 0, 2 1.8 0.6 0. 1 0.2 4.8 0. 5
Mating 0 0 0 0 0.1 0. 1 0 0 0 0 0 0
Aggression 8.8 1.8 1.5 2. 6 6.5 1. 6 10.8 3. 6 2.8 2.4 28.2 2.8
Diversionary 2.2 0.4 0.3 0. 6 15.2 3 . 6 4.8 1. 6 O. 1 0. 2 9. 1 0.9
Nest Building O. 1 0. 1 0 0 0.9 0. 3 0 0 0 0 0 0
Incubation 0 0 0 0 29, 8 7. 1 0 0 0 0 0 0
Upright 146.5 30.5 15.0 25.0 118.8 28.3 134.4 4.8 52.5 43.8 295.3 29.0
Parental 45.4 9.4 3. 5 4. 6 24.5 5.8 28.6 9. 6 1.8 1.6 98.4 9. 6
Absent 13.8 2.9 27.3 45.5 0 0 0 0 3. 1 2. 6 23.2 2. 2
Total 468 . 2 60.0 414.3 297.3 118.4 982. 6
Stage 4Feeding 136.8 76.0 206.8 49.6 119.0 24.8 282.8 67.4 373.4 62.4 206.3 42.9
Resting 35.8 20.0 123.1 37.2 279.9 54.8 55.1 13.2 118.3 19.2 118.0 24.6
Preening 1.4 1.8 158.2 26.4 61.4 12.3 78.0 17.8 91.3 15.2 143.3 29.8
Other maint.activities
1.5 0.8 4.9 0.8 2, 0 0.4 1.8 0.4 6.4 1, 0 4.4 1.0
Mating 0 0 0 0 0 0 0 0 0 0 0 0
Aggression 0 0 1.8 0.3 2.0 0.5 3.7 0.8 1.4 0.3 1.6 0.4
Diversionary 0 0 0 0 0 0 0 0 0 0 0 0
Nest Building 0 0 0 0 0 0 0 0 0 0 0 0
Incubation 0 0 0 0 0 0 0 0 0 0 0 0
Upright 2.1 1.2 1.4 0.4 8.2 1.8 1.8 0.4 9.0 1.5 4.7 1.0
Parental 0 0 0 0 0 0 0 0 0 0 0 0
Absent 0 0 0 0 0 0 0 0 0 0 0 0
Total 177; 6 496; 2 472:5 423; 2 599; 8 478; 3
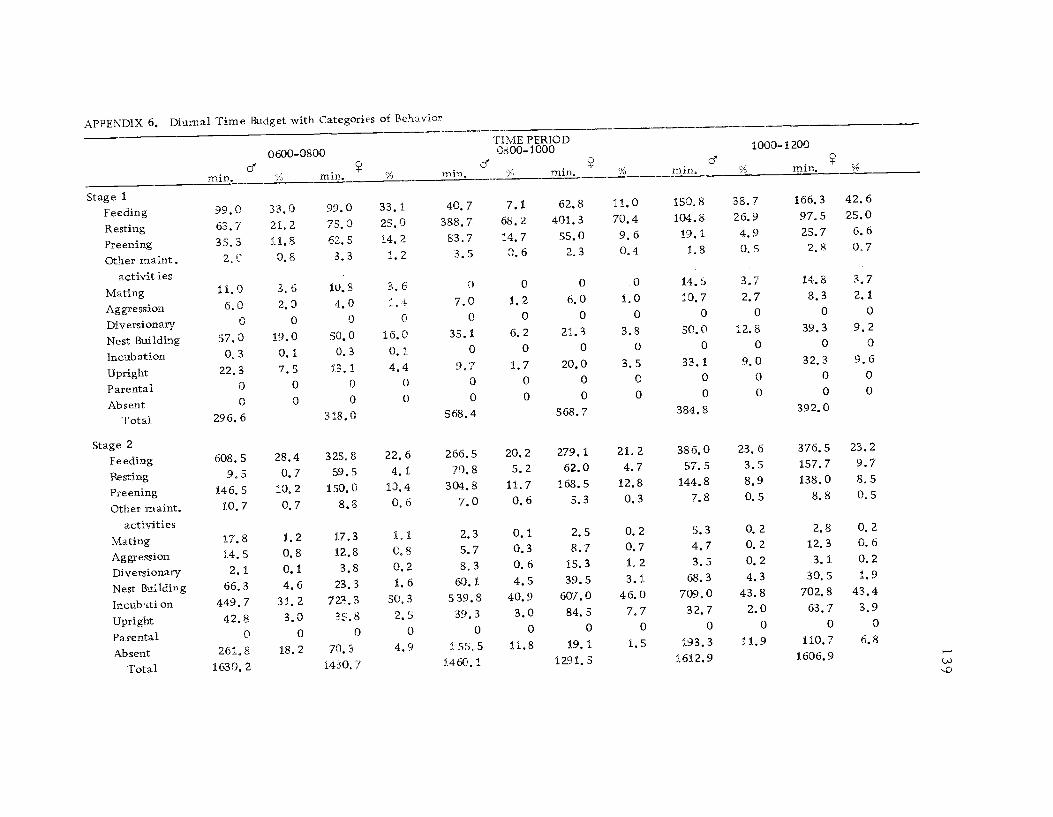
APPENDIX 6. Diurnal Time Budget with Categories of BOlavior
min.
0600-0800cc
°Z, min. '76 min.
TIME PERIOD0800-1000
.mm. min
1000-1200cf min.
Stage 1Feeding 99.0 33.0 99.0 33, 1 40.7 7. 1 62.8 11.0 150.8 38.7 166.3 42. 6
Resting 63, 7 21.2 75.0 25.0 388.7 68.2 401.3 70.4 104.8 26.9 97, 5 25.0
Preening 35.3 11.8 62.5 14.2 83.7 14,7 55.0 9. 6 19.1 4. 9 25.7 6. 6
Other maint .activities
2. 0 0.8 3. 3 1, 2 3, 5 O. 6 2. 3 0, 4 1. 8 O. 5 2. 8 0. 7
Mating 11.0 3. 6 10, 8 3, A 0 0 0 0 14.5 3.7 14. 8 3., 7
Aggression 6.0 2. 0 4, 0 1,4 7. 0 1. 2 6.0 1.0 10.7 2.7 8. 3 2. 1
Diversionary 0 0 0 0 0 0 0 0 0 0 0 0
Nest Building 57.0 19.0 50, 0 16.0 35.1 6. 2 21.3 3.8 50.0 12.8 39.3 9. 2
Incubation 0. 3 0. 1 0. 3 0. 1 0 0 0 0 0 0 0 0
Upright 22.3 7. 5 13.1 4.4 9.7 1.7 20.0 3. 5 33.1 9. 0 32.3 9. 6
Parental 0 0 0 0 0 0 0 0 0 0 0 0
Absent 0 0 0 0 0 0 0 0 0 0 0 0
Total 296.6 318. 0 568, 4 568.7 384.8 392.0
Stage 2Feeding 608.5 28.4 325.8 22.6 266..5 20.2 279.1 21.2 386.0 23.6 376.5 23.2
Resting 9.5 0.7 59.5 4.1 70.8 5.2 62.0 4.7 57.5 3.5 157.7 9.7
Preening 146.5 10.2 150.0 10.4 304.8 11.7 168.5 12.8 144.8 8.9 138.0 8. 5
Other Tri a int .
activities10.7 0. 7 8,8 0, 6 7.0 0. 6 5.3 0.3 7.8 0. 5 8.8 0. 5
Mating 17.8 1. 2 17.3 1. 1 2.3 0. 1 2. 5 0. 2 S. 3 0. 2 2.8 0. 2
Aggression 14.5 0. 8 12.8 0.8 5. 7 0.3 8. 7 0. 7 4, 7 0. 2 12. 3 0. 6
Diversionary 2. / 0, 1 3.8 0. 2 8. 3 0. 6 15.3 1. 2 3. 5 0, 2 3. 1 0. 2
Nest Building 66.3 4. 6 23.3 1. 6 60.1 4. 5 39.5 3. 1 68.3 4, 3 30. 5 1. 9
Incubati on 449.7 31, 2 723.3 50.3 5 39. 8 40.9 607.0 46.0 709, 0 43, 8 702.8 43.4
Upright 42.8 3.0 35.8 2. 5 39.3 3.0 84.5 7. 7 32.7 2.0 63.7 3.9
Parental 0 0 0 0 0 0 0 0 0 0 0 0
Absent 261.8 18.2 70, 3 4.9 1 55.5 11.8 19.1 1.5 193.3 11.9 110.7 6.8
Total 1630. 2 1430.1 1460. 1 1291.5 1612.9 1606.9
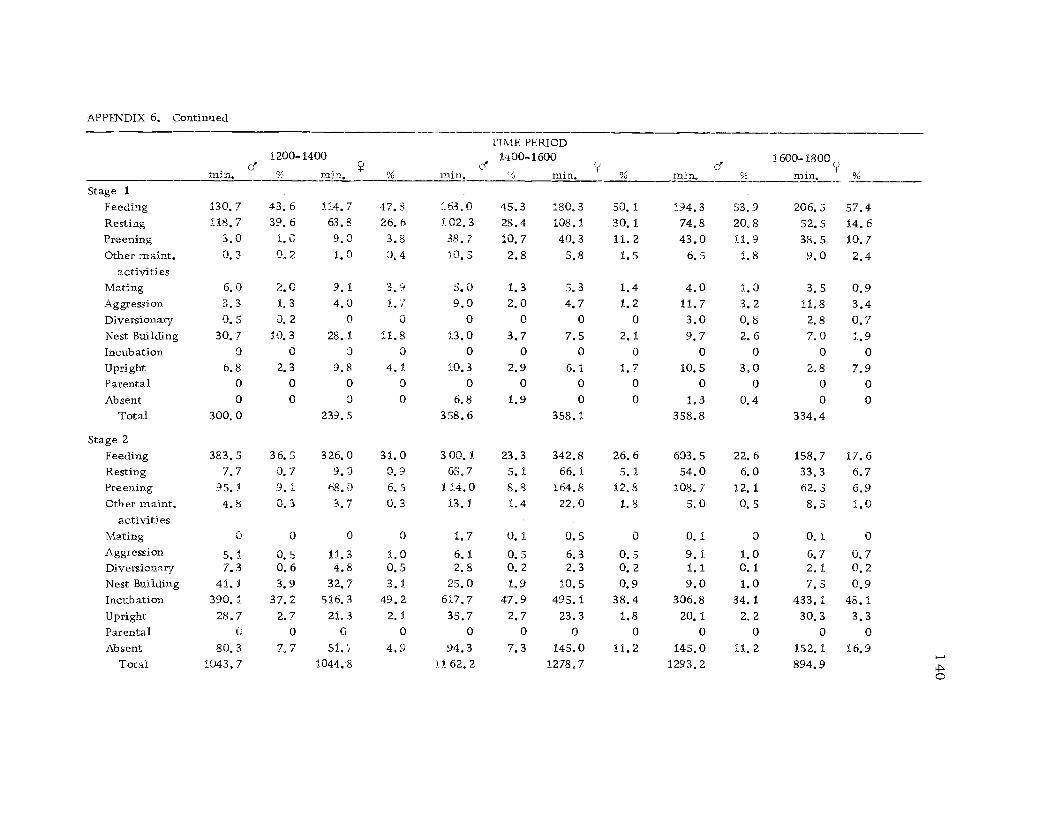
APPENDIX 6. Continued
min,1200-1400
w min, min,
TIME PERIOD1400-1600
min, min.1600-1800
min.Stage 1
Feeding 13O,7 43,h z14.7 4r,8 163, 0 45,S 180. 3 50.1 194,3 53. 9 206.s 57,4Resting 11u,7 39. 6 t3,8 26. 6 1 02. 3 28. 4 108. 1 30. 1 74,8 2O,8 52. 5 t4.6Preening s,0 1.J 9,8 3.B f8,7 1O,7 40,S 11,u 43,O 11.9 38,S 1O,7Other maint.
activities0,3 g,3 1,O 0,4 10. 5 2,8 S,8 1,s 0,s 1^8 9,0 3,4
Mating 6. 0 2. 0 9.1 3^9 5,O 1,3 S,3 1.4 4,0 1,J 3. 5 0,9Aggression *.3 1.3 4,0 1,7 9,O u,0 4,7 1,% 11,7 3.0 11,8 3.4Diversionary 0. 5 0, 2 0 0 0 0 0 0 3,O 0,Q u.8 O,7Nest Building S0,7 1Q,3 28, 1 H,& 13,O 3,7 7,S u,1 9,7 2,h 7.0 1,9Incubation 0 0 0 0 0 0 0 0 0 0 0 0Upright h,8 u,3 9,8 «,1 10.3 2,9 h,1 1,7 10,3 3.0 2.8 7,eParental 0 0 0 0 0 0 0 0 0 0 0 0Absent 0 0 0 0 h.8 1,9 0 0 1.3 0.4 0 0
Total 300,0 239,s 3S8.6 358,1 358. 8 334,4
Stage 2Feeding 383. 5 36. 5 326. 0 31,0 3 00. 1 23. 3 342. 8 26. 6 603, 5 22. 6 1s8,7 17. 6Resting 7,7 0,Y 9.0 0.9 h5.7 s.1 @6.1 s.1 34,0 6.O 33,S 6,7Preening 95. 1 'J. 1 t8,9 6. 5 1 14. 0 8,8 164. 8 12,8 1o8,7 12.1 62. 5 0.9Other maint.
activities4.8 O.3 J,7 8,S 13'1 1.4 z2.0 1.8 5.0 0,S 8,s LO
Mating 0 0 0 0 1,7 0, 1 0,S 0 0, 1 0 0. 1 0Aggression 3,1 0,s 11^3 1.0 h,1 0,s h,3 0,S 9,1 1,0 h.7 0,7Diversiongry 7.3 Q,0 4.8 0,3 u,8 0. 2 2. 3 0,u 1.1 0,1 o,1 0,zNest Building 41,/ 3,9 32, 7 3,1 25, 0 1,9 10,S O,9 9. 0 1,0 7,s 0,pIncubation 390,1 37,2 516. 3 49,u h17,7 47.9 495.1 38.4 30h,8 34,1 43S^1 4B,1Upright 28. 7 m,7 u1.3 u.1 S5.7 u.7 23. 3 1.8 u0,1 2, 2 30,3 3,SParental 0 0 0 0 0 0 0 0 0 0 0 0Absent 80,a 7.7 51. ; 4.9 94,S 7,3 14s,0 11,2 146,0 11, 2 152. 1 1h,9
Total 1043, 7 1044, 8 11 62. 2 1278, 7 1293. 2 894. 9
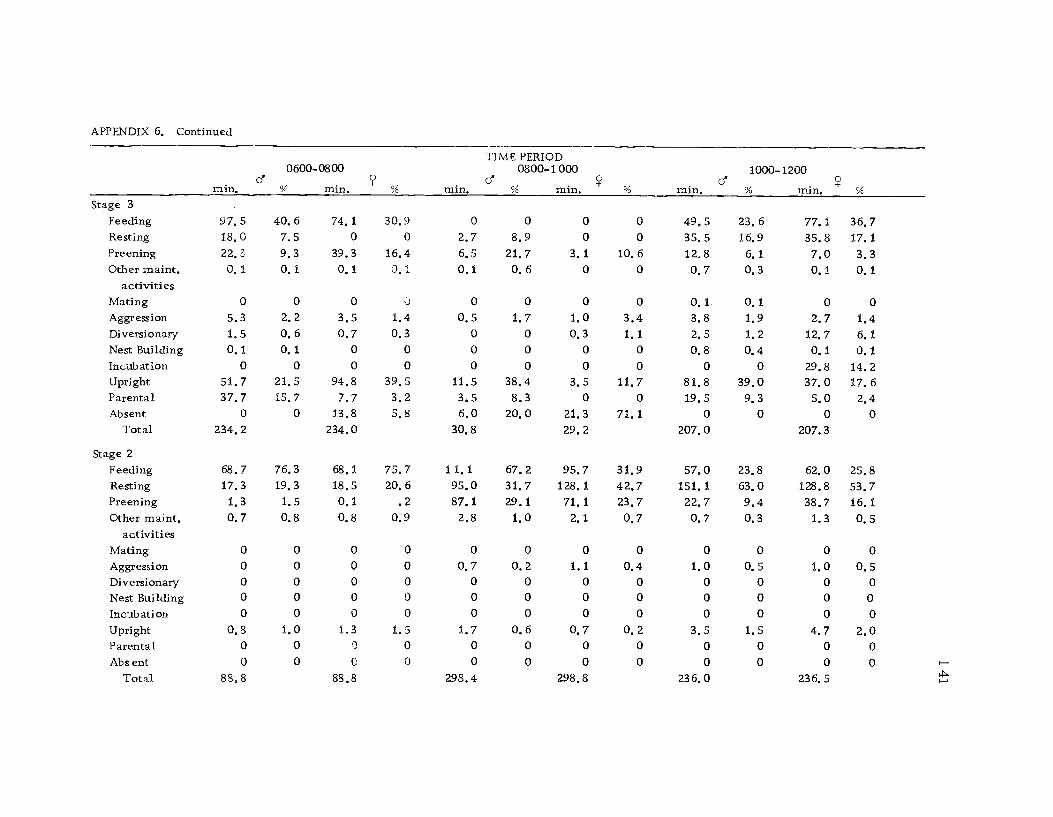
APPENDIX 6. Continued
min,
0600-0800
% min. min.
TIME PERIOD0800-1 000
% min. % min.
1000-1200
% min, %
Stage 3Feeding 97.5 40.6 74.1 30.9 0 0 0 0 49.5 23.6 77.1 36.7Resting 18.0 7.5 0 0 2.7 8.9 0 0 35.5 16.9 35.8 17.1Preening 22.3 9.3 39.3 16.4 6.5 21.7 3.1 10.6 12.8 6.1 7.0 3.3Other maint.
activities0.1 0.1 0.1 0.1 0.1 0.6 0 0 0.7 0.3 0.1 0./
Mating 0 0 0 0 0 0 0 0 0.1 0.1 0 0Aggression 5.3 2.2 3.5 1.4 0.5 1.7 1.0 3.4 3.8 1.9 2.7 1.4Diversionary 1.5 0.6 0.7 0.3 0 0 0.3 1.1 2.5 1.2 12.7 6.1Nest Building 0.1 0.1 0 0 0 0 0 0 0.8 0.4 0.1 0.1Incubation 0 0 0 0 0 0 0 0 0 0 29.8 14.2Upright 51.7 21.5 94.8 39.5 11.5 38.4 3.5 11.7 81.8 39.0 37.0 17.6Parental 37.7 15.7 7.7 3.2 3.5 8.3 0 0 19.5 9.3 5.0 2.4Absent 0 0 13.8 5.8 6.0 20.0 21.3 71.1 0 0 0 0
Total 234.2 234.0 30.8 29.2 207.0 207.3
Stage 2Feeding 68.7 76.3 68.1 75.7 11.1 67.2 95.7 31.9 57.0 23.8 62.0 25.8Resting 17.3 19.3 18.5 20.6 95.0 31.7 128.1 42.7 151.1 63.0 128.8 53.7Preening 1.3 1.5 0.1 .2 87.1 29.1 71.1 23.7 22.7 9.4 38.7 16.1Other maint.
activities0.7 0.8 0.8 0.9 2.8 1.0 2.1 0.7 0.7 0.3 1.3 0.5
Mating 0 0 0 0 0 0 0 0 0 0 0 0Aggression 0 0 0 0 0.7 0.2 1.1 0.4 1.0 0.5 1.0 0.5Diversionary 0 0 0 0 0 0 0 0 0 0 0 0Nest Building 0 0 0 0 0 0 0 0 0 0 0 0
Incubation 0 0 0 0 0 0 0 0 0 0 0 0Upright 0.8 1.0 L3 1.5 1.7 0.6 0.7 0.2 3.5 1.5 4.7 2.0Parental 0 0 9 0 0 0 0 0 0 0 0 0Abs ent 0 0 0 0 0 0 0 0 0 0 0 0
Total 88.8 88.8 298.4 298.8 236.0 236.5
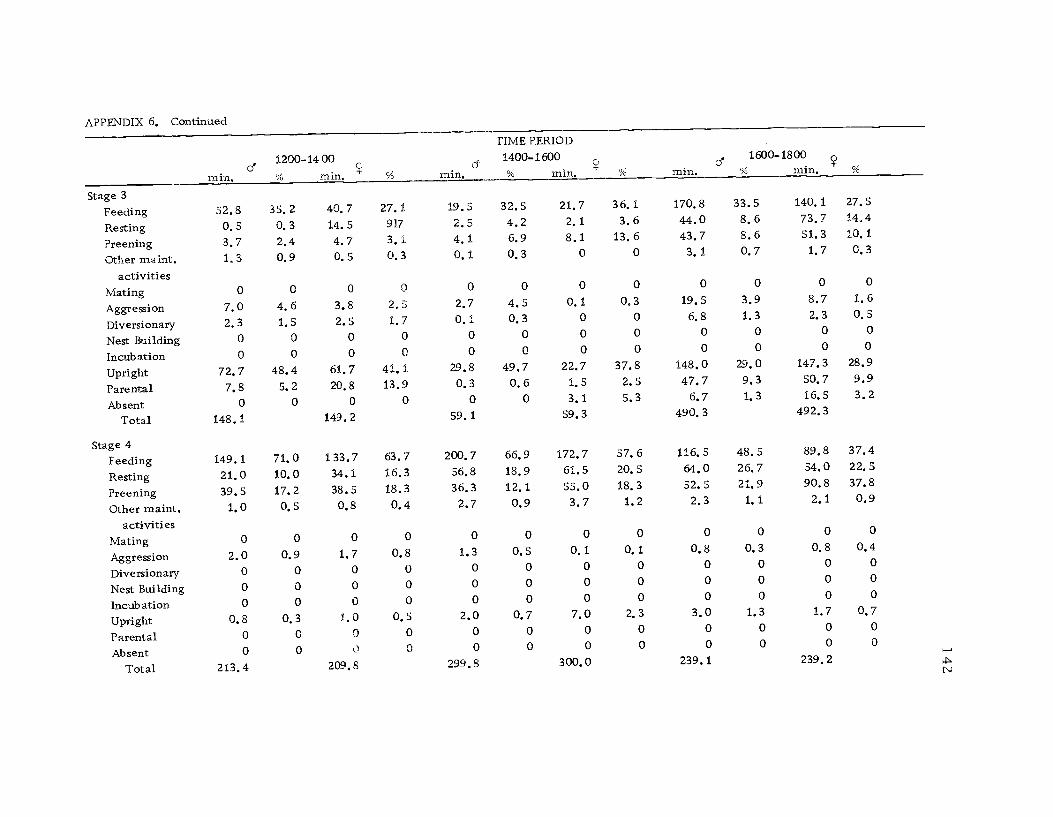
APPENDIX 6. Continued
min.1200-14d%
00-mm. %
cfmin.
TIME PERIOD1400-1600% min. % min.
1600-1800min.
Stage 3Feeding 52.8 35.2 40.7 27.1 19.5 32.5 21.7 36.1 170.8 33.5 140.1 27.5
Resting 0.5 0.3 14.5 917 2.5 4.2 2.1 3.6 44.0 8.6 73.7 14.4
Preening 3.7 2.4 4.7 3.1 4.1 6.9 8.1 13.6 43.7 8.6 51.3 10.1
Other maint.activities
1. 3 0.9 0. 5 0. 3 0. 1 0. 3 0 0 3. 1 0. 7 1. 7 0. 3
Mating 0 0 0 0 0 0 0 0 0 0 0 0
Aggression 7.0 4.6 3.8 2.5 2.7 4.5 0.1 0.3 19.5 3.9 8.7 1.6
Diversionary 2. 3 1.5 2.5 1.7 0. 1 0. 3 0 0 6.8 1. 3 2. 3 0.5
Nest Building 0 0 0 0 0 0 0 0 0 0 0 0
Incubation 0 0 0 0 0 0 0 0 0 0 0 0
Upright 72.7 48.4 61.7 41, 1 29.8 49.7 22.7 37.8 148.0 29.0 147.3 28.9
Parental 7.8 5.2 20.8 13.9 0.3 0.6 1.5 2.5 47.7 9.3 50.7 9.9
Absent 0 0 0 0 0 0 3.1 5.3 6.7 1.3 16.5 3.2
Total 148.1 149.2 59.1 59.3 490.3 492.3
Stage 4Feeding 149.1 71.0 1 33. 7 63.7 200.7 66.9 172.7 57.6 116.5 48.5 89.8 37.4
Resting 21.0 10.0 34.1 16.3 56.8 18.9 61.5 20. 5 64. 0 26. 7 54.0 22. 5
Preening 39.5 17.2 38.5 18.3 36.3 12. 1 55.0 18. 3 52. 5 21.9 90.8 37.8
Other maint.activities
1.0 0. 5 0.8 0.4 2.7 0.9 3. 7 1. 2 2. 3 1. 1 2. 1 0.9
Mating 0 0 0 0 0 0 0 0 0 0 0 0
Aggression 2.0 0.9 1.7 0.8 1.3 0.5 0.1 0.1 0.8 0.3 0.8 0.4
Diversionary 0 0 0 0 0 0 0 0 0 0 0 0
Nest Building 0 0 0 0 0 0 0 0 0 0 0 0
Incubation 0 0 0 0 0 0 0 0 0 0 0 0
Upright 0.8 0.3 1.0 0.5 2.0 0.7 7.0 2.3 3.0 1.3 1.7 0.7
Parental 0 0 0 0 0 0 0 0 0 0 0 0
Absent 0 0 0 0 0 0 0 0 0 0 0 0
Total 213.4 209.8 299.8 300.0 239.1 239. 2

1 43
APPENDIX 7
Calculation of Standard Metabolism
There is an approximately inverse linear relationship between standard
metabolism and temperature in birds (King and Farner, 1961). This
relationship is weight dependent. Standard equations have been
calculated to represent these relationships for non-passerine birds
at different temperatures (Kendeigh, 1970):
At 30 ° C, log M = -0. 26 73 + 0. 7545 log W t 0.063
At 0 ° C, log M = O. 63 72 + 0. 5300 log W t 0. 0613
where
M = Standard metabolism as Kcal/bird/day
W = Weight in grams
= Standard error of the estimate of log M
for avocets, W(average) = 316 g (Hamilton, 1969)
Therefor e;
at 30 ° C, M = 33.3 ± 0.063
at 0 ° C, M = 91. 7 ± 0.0613
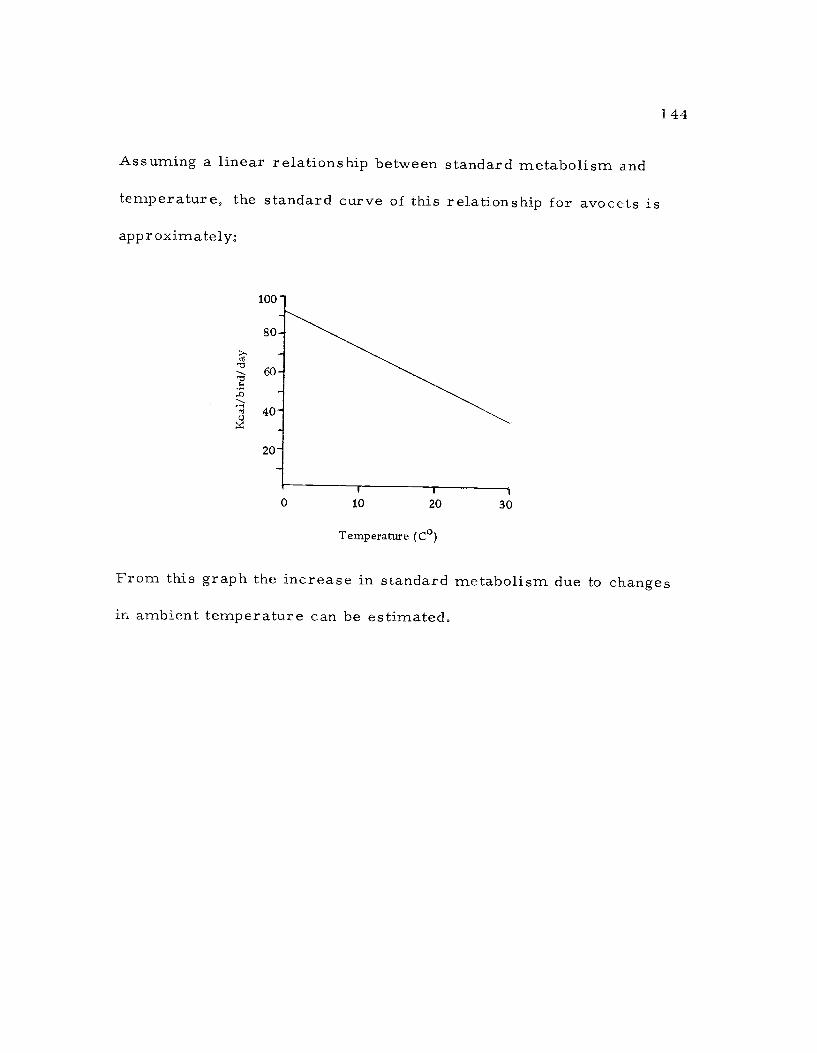
Assuming a linear relationship between standard metabolism and
temperature, the standard curve of this relationship for avocets is
approximately:
0 10 20
Temperature (C°)
30
144
From this graph the increase in standard metabolism due to changes
in ambient temperature can be estimated.





























