Embed Size (px)
Citation preview

Presynaptic SNAP-25 regulates retinal waves andretinogeniculate projection via phosphorylationYu-Tien Hsiaoa, Wen-Chi Shua, Pin-Chun Chena, Hui-Ju Yanga,b, Hsin-Yo Chena, Sheng-Ping Hsua, Yi-Ting Huanga,Cheng-Chang Yanga, Yen-Ju Chena, Ni-Yen Yua, Shih-Yuan Lioua, Ning Chianga,c,d, Chien-Ting Huanga, Tzu-Lin Chenga,Lam-Yan Cheunga, Yu-Chun Line, Juu-Chin Lue,f,1, and Chih-Tien Wanga,b,c,d,1
aInstitute of Molecular and Cellular Biology, National Taiwan University, Taipei 10617, Taiwan; bGenome and Systems Biology Degree Program, NationalTaiwan University and Academia Sinica, Taipei 10617, Taiwan; cDepartment of Life Science, National Taiwan University, Taipei 10617, Taiwan;dNeurobiology and Cognitive Science Center, National Taiwan University, Taipei 10617, Taiwan; eDepartment of Physiology and Pharmacology, College ofMedicine, Chang Gung University, Taoyuan 33302, Taiwan; and fDivision of Endocrinology and Metabolism, Department of Internal Medicine, Chang GungMemorial Hospital, Linkou 33305, Taiwan
Edited by Marla B. Feller, University of California, Berkeley, CA, and accepted by Editorial Board Member Jeremy Nathans January 8, 2019 (received for reviewJuly 17, 2018)
Patterned spontaneous activity periodically displays in developingretinas termed retinal waves, essential for visual circuit refinement. Inneonatal rodents, retinal waves initiate in starburst amacrine cells(SACs), propagating across retinal ganglion cells (RGCs), furtherthrough visual centers. Although these waves are shown temporallysynchronizedwith transiently high PKA activity, the downstream PKAtarget important for regulating the transmission from SACs remainsunidentified. A t-SNARE, synaptosome-associated protein of 25 kDa(SNAP-25/SN25), serves as a PKA substrate, implying a potential roleof SN25 in regulating retinal development. Here, we examinedwhether SN25 in SACs could regulate wave properties and retinoge-niculate projection during development. In developing SACs, over-expression of wild-type SN25b, but not the PKA-phosphodeficientmutant (SN25b-T138A), decreased the frequency and spatial correla-tion of wave-associated calcium transients. Overexpressing SN25b,but not SN25b-T138A, in SACs dampened spontaneous, wave-associated, postsynaptic currents in RGCs and decreased the SACrelease upon augmenting the cAMP-PKA signaling. These resultssuggest that SN25b overexpression may inhibit the strength oftransmission from SACs via PKA-mediated phosphorylation at T138.Moreover, knockdown of endogenous SN25b increased the fre-quency of wave-associated calcium transients, supporting the roleof SN25 in restraining wave periodicity. Finally, the eye-specificsegregation of retinogeniculate projection was impaired by in vivooverexpression of SN25b, but not SN25b-T138A, in SACs. Theseresults suggest that SN25 in developing SACs dampens the spatio-temporal properties of retinal waves and limits visual circuit re-finement by phosphorylation at T138. Therefore, SN25 in SACs plays aprofound role in regulating visual circuit refinement.
SNAP-25 | PKA-mediated phosphorylation | starburst amacrine cells |retinal waves | retinogeniculate projection
During critical developmental periods, correlated, patterned,spontaneous activity influences the refinement of various
developing circuits in many species, all with a similar underlyingmechanism in which a subset of interneurons release neuro-transmitters to postsynaptic neurons (1). In the developingvertebrate retina, this patterned spontaneous activity displaysunique spatiotemporal patterns termed retinal waves. During thefirst postnatal week in rodents, retinal waves are initiated byneurotransmitter release from cholinergic neurons (starburstamacrine cells, SACs), so-called cholinergic waves or stage IIretinal waves (2, 3). These waves on the layer of retinal ganglioncells (RGCs, the retinal output neurons) are essential foractivity-dependent refinement of visual circuits (1), thus playingan instructive role in guiding RGC axonal projections to theirbrain targets such as the dorsal lateral geniculate nucleus ofthalamus (dLGN) (4). Recent in vivo imaging has shown thatstage II retinal waves coordinate patterned spontaneous activ-ity through visual areas (5), confirming the activity-dependent
refinement of visual circuits initiates from developing retinas.However, the molecule in SACs that is sufficient to regulate theoverall visual circuit refinement is yet to be identified.The spatiotemporal patterns of stage II retinal waves depend on
the intrinsic properties of SACs (2). To induce cholinergic waves,cholinergic transmission starts in SACs with spontaneous, periodicdepolarizations that activate voltage-gated Ca2+ channels, thus Ca2+
influx triggering exocytosis. Once Ca2+ ions bind to Ca2+ sensorssuch as synaptotagmin I (Syt I) (6), the vesicle membrane fuses withthe SAC plasma membrane, allowing neurotransmitters to be re-leased and received by neighboring SACs and RGCs. Our previousstudy showed that the frequency of cholinergic waves is dampenedby weakening Ca2+ binding to the C2A/B domain of Syt I in SACs(7), suggesting that a key change in the release machinery of SACs issufficient to alter the temporal patterns of stage II retinal waves. Inaddition to Syt I, the essential fusion machinery during exocytosis isthe soluble N-ethylmaleimide-sensitive factor attachment proteinreceptor (SNARE) complex, consisting of a vesicle membrane
Significance
Patterned spontaneous activity can modify the structure andfunction of neural circuits during development, such as retinalwaves essential for establishing functional visual circuits. Retinalwaves are initiated by presynaptic release, further inducingtemporally synchronized oscillations of PKA activity. However,the PKA substrate regulating visual circuit refinement remainselusive. Here, we describe a biological mechanism in which PKA-mediated SN25 phosphorylation in presynaptic retinal neuronsdown-regulates synaptic strength and retinal waves during de-velopment. These changes in wave spatiotemporal patterns canfurther induce global effects on visual circuit refinement, i.e., theeye-specific segregation of retinogeniculate projection. There-fore, by switching the phosphorylation state by PKA, SN25 inwave-initiating neurons can regulate the plasticity of retinal cir-cuit activity and sculpt developing visual circuits.
Author contributions: Y.-T. Hsiao, J.-C.L., and C.-T.W. designed research; Y.-T. Hsiao,W.-C.S., P.-C.C., H.-J.Y., H.-Y.C., S.-P.H., Y.-T. Huang, C.-C.Y., Y.-J.C., N.-Y.Y., S.-Y.L., N.C.,C.-T.H., T.-L.C., L.-Y.C., Y.-C.L., J.-C.L., and C.-T.W. performed research; H.-Y.C. contributednew reagents/analytic tools; Y.-T. Hsiao, W.-C.S., P.-C.C., H.-J.Y., H.-Y.C., S.-P.H.,Y.-T. Huang, C.-C.Y., Y.-J.C., N.-Y.Y., J.-C.L., and C.-T.W. analyzed data; and Y.-T. Hsiao,J.-C.L., and C.-T.W. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. M.B.F. is a guest editor invited by theEditorial Board.
Published under the PNAS license.1To whom correspondence may be addressed. Email: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1812169116/-/DCSupplemental.
Published online February 6, 2019.
3262–3267 | PNAS | February 19, 2019 | vol. 116 | no. 8 www.pnas.org/cgi/doi/10.1073/pnas.1812169116
Dow
nloa
ded
by g
uest
on
Mar
ch 2
1, 2
020

protein, synaptobrevin (Syb) (i.e., vesicle-associated membraneprotein, VAMP), and two plasma membrane proteins, syntaxin(Stx) and synaptosome-associated protein of 25 kDa (SNAP-25/SN25) (6). Syb and Stx contain a transmembrane domain thatspans the vesicle and the plasma membrane, respectively. In-stead, SN25 anchors to plasma membrane via palmitoyl sidechains at the center of the protein, contributing two α-helical domainsto form SNARE motifs. Although the importance of SNAREs for
vesicle release has been recognized, it is unknown whether any ofthese SNAREs may affect wave patterns or visual circuit refinement.Previous studies showed that SN25 is expressed in developing
SACs within the period of cholinergic waves (8, 9), implying a roleof SN25 in regulating these waves. Moreover, cAMP-PKA signalingis activated by these waves at a periodicity on the order of tens ofseconds, with the occurrence of transiently high PKA activity duringthe interwave intervals (10). Coincidentally, among three SNAREs,only SN25 is shown to function as a PKA substrate by phosphor-ylation at threonine 138 (T138) (11). These led to the hypothesisthat PKA-mediated SN25 phosphorylation at T138 is important forregulating SAC release, likely affecting the spatiotemporal patternsof retinal waves and/or visual circuit refinement. However, to date,there is no direct evidence to support this hypothesis.In this study, by combining the cell type-specific molecular per-
turbation, live imaging, electrophysiology, immunostaining, and ax-onal tracing, we show that SN25 in developing SACs is sufficient tomediate PKA regulation of synaptic strength and wave properties. Inaddition, we demonstrate that PKA-mediated SN25 phosphoryla-tion in SACs can further regulate the eye-specific segregation of theretinogeniculate projection, suggesting a functional role of pre-synaptic SN25 in regulating the visual circuit refinement.
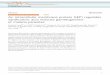
ResultsWave-Initiating Neurons SACs Express SN25, Mainly SN25b, During thePeriod of Stage II Retinal Waves. To investigate the role of pre-synaptic SN25 in regulating cholinergic waves, we confirmed thatSACs expressed SN25 during the cholinergic wave period. Byimmunostaining postnatal (P)2 retinal cross-sections, we foundthat SN25 immunoreactivity was localized in the inner retina,including the inner plexiform layer (IPL) and ganglion cell layer(GCL) (Fig. 1A). SN25 immunoreactivity was found surrounding5 s
A B C D
E GH
I J
K L
F M N
Fig. 1. SN25b in SACs down-regulates patterned spontaneous activity byphosphorylation at T138. (A) Immunostaining of SN25 (green) and cholineacetyltransferase (ChAT, the SAC marker; red) in retinal cross-sections. GCL,ganglion cell layer; IPL, inner plexiform layer; NBL, neuroblast layer. Arrows,the colocalization signal. (B) Immunostaining of SN25 (green) and ChAT (red)in whole-mount retinas (1.5-μm z-section). (C) Immunostaining of SN25(green), ChAT (red), or DAPI (blue) in a dissociated SAC (1.5-μm z-section). BF,bright field. (Scale bars: A, Left, 25 μm; A, Right, B, and C, 10 μm.) (D) Mo-lecular perturbation in P1–P2 rat SACs were performed by transfection withSAC-specific constructs, including Ctrl (pmGluR2-IRES2-egfp), SN25b, andSN25b-T138A. (E) Representative spontaneous Ca2+ transients. (F) SingleCa2+ transients. (G) Ca2+ transient frequency. *P < 0.05; ***P < 0.001, two-tailed Student’s unpaired t test. (H) Cumulative probability of Ca2+ transientfrequency. ***P < 0.001, Friedman test with Dunn post hoc test. (I) Inter-wave interval. *P < 0.05; **P < 0.01, two-tailed Student’s unpaired t test. (J)Cumulative probability of interwave interval. *P < 0.05; ***P < 0.001,Friedman test with Dunn post hoc test. (K) Ca2+ transient duration. (L) Cu-mulative probability of Ca2+ transient duration. (M) Ca2+ transient ampli-tude. (N) Cumulative probability of Ca2+ transient amplitude. For G–N, n,transfection group (no. of cells, no. of retinas, no. of pups): Ctrl (979, 60, 15);SN25b (353, 21, 8); SN25b-T138A (249, 15, 6). Dots beside columns representdata from individual retinas.
A B C
D E F
G H
Fig. 2. SN25b in SACs reduces the intercellular correlation of spontaneousCa2+ transients by phosphorylation at T138. (A–C) Representative rasterplots for spontaneous Ca2+ transients in one imaged region from retinastransfected with SAC-specific constructs. (A) Ctrl (30 regions of interests,ROIs). (B) SN25b (34 ROIs). (C) SN25b-T138A (36 ROIs). (D–F) Representativeactivity plots derived from raster plots. (D) Ctrl. (E) SN25b. (F) SN25b-T138A.(G) Cumulative probability of correlated activity. *P < 0.05, repeated mea-sures ANOVA with Tukey post hoc test. (H) STTC values as a function of in-tercellular distance. ***P < 0.001, repeated measures ANOVA with Tukeypost hoc test. n, transfection group (no. of imaged regions, no. of retinas,no. of pups): Ctrl (41, 25, 9); SN25b (37, 21, 8); SN25b-T138A (24, 14, 6).
Hsiao et al. PNAS | February 19, 2019 | vol. 116 | no. 8 | 3263
NEU
ROSC
IENCE
Dow
nloa
ded
by g
uest
on
Mar
ch 2
1, 2
020

SACs (Fig. 1 B and C), consistent with the hypothesis that at de-veloping cholinergic synapses, SN25 in SACs may regulate stageII waves.SN25 consists of two SN25 splicing variants (SN25a and
SN25b) (6). We found that SN25b was the major SN25 isoformduring this stage (SI Appendix, Fig. S1A). To further determinehow SN25b regulated stage II waves, we manipulated the level ofSN25b expression or phosphorylation in SACs. Postnatal retinaswere transfected with the control vector (designated Ctrl, here-inafter), wild-type SN25b (designated SN25b, hereinafter), orSN25b harboring a PKA-phosphodeficient mutation (replacingthreonine 138 with alanine; designated SN25b-T138A, herein-after) (11), with the SAC-specific promoter (the type II metab-otropic glutamate receptor promoter, pmGluR2) (7, 12).SAC-specific perturbation was effective at 3 d after transfectionfor manipulating the levels of SN25b expression or PKA-mediatedSN25b phosphorylation (SI Appendix, Figs. S1 and S2).
PKA-Mediated SN25b Phosphorylation in SACs Reduces the Frequencyof Spontaneous Ca2+ Transients Associated with Retinal Waves. Todetermine whether the properties of stage II waves could be reg-ulated by the level of SN25b expression or phosphorylation inSACs, we performed live Ca2+ imaging and measured spontaneousCa2+ transients in the RGC layer following the overexpression ofSN25b or SN25b-T138A in SACs (Fig. 1D). Spontaneous, corre-lated Ca2+ transients in individual cells revealed stage II waves in
the RGC layer of transfected explants (Fig. 1E). Ca2+ transientfrequency was significantly decreased, and the interwave intervalwas significantly increased by the overexpression of SN25b in SACscompared with Ctrl (Fig. 1 G and I). This effect may be attributedto two possibilities, either the increased SN25b expression level orthe elevated SN25b phosphorylation by PKA. To distinguish thesepossibilities, we detected the Ca2+ transient frequency/interval inretinas with SACs overexpressing SN25b-T138A, which confers anincreased expression level but no increase in PKA phosphorylation.As a result, SN25b-T138A showed an increased Ca2+ transientfrequency and a reduced interwave interval compared with SN25b,but similar to Ctrl (Fig. 1 G and I). Moreover, the cumulativeprobability curve for the Ca2+ transient frequency in SN25b wasleft-shifted (Fig. 1H), but the interwave interval was right-shifted(Fig. 1J), compared with Ctrl and SN25b-T138A, indicating that themajority of cells displayed a lower Ca2+ transient frequency butlonger interwave interval in response to SN25b but not SN25b-T138A. These results suggest that SN25b in SACs dampens thefrequency of retinal waves, and T138 phosphorylation by PKA isrequired for the SN25b-dampened wave frequency.The Ca2+ transient size represents the level of Ca2+ influx into
neurons during individual waves. To further determine whether theCa2+ transient size was regulated by SN25b or SN25b-T138A inSACs, we quantified the duration and amplitude of spontaneousCa2+ transients (Fig. 1 F and K–N). The duration and amplitude ofsingle Ca2+ transients were comparable among all groups, suggesting
5% ΔF/F
#
#
#
A B C D E
FG H I
Fig. 3. SN25b inhibits the strength of transmission from developing SACs via PKA-mediated phosphorylation. (A) The RGC, proximal to transfected SACs, wasidentified by Na2+ currents upon depolarizing pulses in whole-cell voltage-clamp recordings. (B, Left) The wave-associated PSCs were recorded in RGCs fromthe retinas transfected with Ctrl (pmGluR2-IRES2-egfp), SN25b, or SN25b-T138A. #, PSCs shown in Right. (C) PSC frequency. *P < 0.05, two-tailed Student’sunpaired t test. (D) PSC duration. *P < 0.05 for Ctrl vs. SN25b, Mann–Whitney test. **P < 0.01, two-tailed Student’s unpaired t test. (E) PSC integral. *P < 0.05for Ctrl vs. SN25b, Mann–Whitney test. *P < 0.05 for SN25b vs. SN25b-T138A, two-tailed Student’s unpaired t test. n, transfection group (no. of RGCs, no. ofretinas, no. of pups): Ctrl (5, 5, 4); SN25b (5, 5, 4); SN25b-T138A (5, 5, 4). Circles represent data from individual RGCs. (F) The adenosine receptor subtype A2ARagonist (CGS 21680) up-regulates A2AR-cAMP-PKA signaling in developing SACs, leading to phosphorylation of SN25b at T138, which may reduce SACexocytosis. (G) The probability of SAC release was determined by coexpressing the exocytosis reporter (VAMPII-pHVenus) and the protein of interest from thesame translational product. The control vector, Ctrl (pmGluR2-vamp2-phvenus-2a), SN25b, or SN25b-T138A was transfected into P1–2 retinas for SAC-specificexpression. (Upper) Heat map in the absence (−CGS) or presence (+CGS) of CGS 21680. (Scale bar: 6.25 μm.) The fluorescence intensity is presented as arbitraryunits (A.U.). (Lower) Changes in the pHVenus fluorescence intensity (ΔF/F) were acquired in SACs expressing Ctrl, SN25b, or SN25b-T138A. Gray area, theperiod of bath-applying CGS 21680. (H) Changes in pHVenus fluorescence intensity (ΔF/F) of SACs before, during, or after CGS application. Black, individualSAC data. Red, average changes in pHVenus fluorescence intensity (ΔF/F). ***P < 0.001, two-tailed Student’s unpaired t test. (I) Average changes in the ratios(ΔR) of pHVenus fluorescence intensity induced by CGS 21680 [(ΔF/F)during CGS − (ΔF/F)before CGS]. Circles indicate the data from individual SACs. **P < 0.01,two-tailed Student’s unpaired t test. n, transfection group (no. of SACs, no. of retinas, no. of pups): Ctrl (13, 8, 8); SN25b (14, 5, 5); SN25b-T138A (9, 5, 5).
3264 | www.pnas.org/cgi/doi/10.1073/pnas.1812169116 Hsiao et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 2
1, 2
020

that PKA-mediated SN25b phosphorylation in SACs did notinfluence the size of spontaneous Ca2+ transients. Thus, SN25bin SACs regulates the temporal properties of stage II waves viaphosphorylation by PKA.
PKA-Mediated SN25b Phosphorylation in SACs Diminishes the SpatialCorrelation of Wave-Associated Ca2+ Transients. Since PKA-mediatedSN25b phosphorylation in SACs regulates the wave temporalproperties, we determined whether SN25b/SN25b-T138A in SACscould alter the wave spatial properties. To address this, we con-structed raster plots (Fig. 2 A–C) and activity plots (Fig. 2 D–F) inneighboring cell pairs across the same imaged regions (5). Wefound that overexpression of SN25b in SACs reduced the occur-rence of the correlated activity compared with Ctrl and SN25b-T138A. The cumulative probability of the averaged correlatedactivity was significantly left-shifted by SN25b compared with Ctrlor SN25b-T138A (Fig. 2G), suggesting that PKA-mediated SN25bphosphorylation in SACs could reduce the intercellular correlationof wave propagation. To further quantify the structure of spatialpropagation, we calculated the spike time tiling coefficient (STTC)of spontaneous Ca2+ transients in neighboring cell pairs (13), anindex of wave spatial correlation. The STTC values were signifi-cantly decreased by SN25b compared with Ctrl or SN25b-T138A(Fig. 2H), suggesting that PKA-mediated SN25b phosphorylation inSACs diminishes the spatial properties of stage II waves.
SN25b Inhibits Synaptic Transmission from Developing SACs via PKA-Mediated T138 Phosphorylation. Since PKA-mediated SN25bphosphorylation in SACs reduces wave frequency and diminisheswave spatial properties, we next determined whether the effectsmight be attributed to a decrease in the transmission strengthfrom SACs. First, we performed whole-cell voltage-clamp re-cordings on RGCs after SAC-specific expression (Fig. 3 A–E).Whole-cell voltage-clamp recordings from RGCs revealedspontaneous, wave-associated, compound postsynaptic currents(PSCs) (Fig. 3B). We found that the frequency, duration, andintegral of wave-associated PSCs were significantly decreased inretinas transfected with SN25b compared with Ctrl or SN25b-T138A (Fig. 3 C–E). These results suggest that PKA-mediatedSN25b phosphorylation in SACs decreases the wave-associatedperiodic inputs received by postsynaptic RGCs.To further measure the changes in SAC release, we expressed
the exocytosis reporter VAMPII-pHVenus in developing SACs(14). The pH-sensitive Venus (pHVenus) fluorophore was fused tothe C terminus of VAMP-II, facing the intravesicular lumen (pH ∼5). Upon vesicle exocytosis, pHVenus was exposed to the extra-cellular space (pH 7.4). The increase in pH led to an increase influorescence intensity of pHVenus, reflecting the increased prob-ability of vesicle exocytosis. Moreover, to promptly increase thePKA activity for SN25b phosphorylation, we applied the A2ARagonist CGS 21680 (Fig. 3F), shown to enhance A2AR-cAMP-PKAsignaling in retinal neurons (15). We found that overexpression ofSN25b in SACs dampened the CGS-induced changes in the fluo-rescence intensity compared with Ctrl or SN25b-T138A in SACs(Fig. 3 G–I), suggesting that SN25b may dampen the releaseprobability of SACs by PKA phosphorylation at T138A. Together,through PKA-mediated T138 phosphorylation, SN25b can dampenthe transmission from developing SACs.
Knockdown of Endogenous SN25b in Postnatal Retinas Increases theCa2+ Transient Frequency. To further verify that the levels of en-dogenous SN25b affected stage II waves, we depleted endogenousSN25b by expressing two various shRNA plasmids against SN25b(shSN25b-1 and shSN25b-2) in retinal explants. These shRNAplasmids specifically targeted the expression of SN25b rather thanSN25a (SI Appendix, Fig. S3). Subsequent Ca2+ imaging (Fig. 4)revealed that knockdown of SN25b increased the Ca2+ transientfrequency and decreased the interwave interval compared with
shCtrl (Fig. 4 A, C, and E). Consistently, the cumulative probabilityfor the frequency was right-shifted and for the interval was left-shifted by SN25b knockdown (Fig. 4 D and F), indicating that af-ter SN25b knockdown, the majority of cells displayed the increasedCa2+ transient frequency compared with shCtrl. Conversely, SN25bknockdown caused relatively minor effects on the Ca2+ transientsize (duration and amplitude) (Fig. 4 B and G–J) and the in-tercellular correlation of Ca2+ transients (SI Appendix, Fig. S4)compared with shCtrl. These results suggest that depletion of en-dogenous SN25b increases the Ca2+ transient frequency, in oppo-sition to the decrease in Ca2+ transient frequency caused by SN25boverexpression (Fig. 1G). Moreover, SN25 expression reached themaximal level as the developmental program was completed (SIAppendix, Fig. S3). Thus, high levels of SN25 expression may down-regulate periodic activity in the developing rat retina, supporting afunctional role of SN25 in limiting the periodicity of retinal waves.
PKA-Mediated SN25b Phosphorylation in SACs Impairs the Eye-Specific Segregation of the Retinogeniculate Projection. During vi-sual circuit development, RGC axons from the two eyes first
A
B
C D
E F
G H
I J
Fig. 4. Depletion of endogenous SN25b increases the Ca2+ transient frequencyin developing rat retinas. (A) Spontaneous Ca2+ transients following transfectionwith the shRNA control vector (shCtrl) or the shRNA plasmid against SN25b(shSN25b-1 or shSN25b-2). (B) Single Ca2+ transients. (C) Ca2+ transient frequency.*P < 0.05; **P < 0.01, two-tailed Student’s unpaired t test. (D) Cumulativeprobability of Ca2+ transient frequency. *P < 0.05; ***P < 0.001, Friedman testwith Dunn post hoc test. (E) Interwave interval. *P < 0.05 for shCtrl vs. shSN25b-1,two-tailed Student’s unpaired t test. *P < 0.05 for shCtrl and shSN25b-2, Mann–Whitney test. (F) Cumulative probability of interwave interval. *P < 0.05; ***P <0.001, Friedman test with Dunn post hoc test. (G) Ca2+ transient duration. (H)Cumulative probability of Ca2+ transient duration. (I) Ca2+ transient amplitude. (J)Cumulative probability of Ca2+ transient amplitude. Dots represent data from in-dividual retinas. n, transfection group (no. of cells, no. of retinas, no. of pups):shCtrl (240, 12, 4); shSN25b-1 (260, 13, 7); shSN25b-2 (270, 14, 7).
Hsiao et al. PNAS | February 19, 2019 | vol. 116 | no. 8 | 3265
NEU
ROSC
IENCE
Dow
nloa
ded
by g
uest
on
Mar
ch 2
1, 2
020

evenly project to the right and left dLGN and then segregate intodistinct bands called eye-specific layers after activity-dependentrefinement. The critical period for the eye-specific segregation ofretinogeniculate projection is P4–P8 in rodents, i.e., the stage IIwave period. Blocking stage II waves or disrupting the spatiotem-poral patterns of these waves has been shown to interfere withintereye or intraeye competition, thus impairing RGC axonal pro-jections to the eye-specific layers of the dLGN (4, 16–19). Thesestudies have indicated that stage II waves are required for activity-dependent synaptic pruning to accomplish precise retinogeniculateprojection. Since PKA-mediated SN25b phosphorylation in SACsdown-regulates the wave spatiotemporal properties, we further ex-amined whether PKA-mediated SN25b phosphorylation in SACscould affect eye-specific segregation of the retinogeniculate pro-jection. To answer this question, we performed in vivo electro-poration (20) to express Ctrl, SN25b, or SN25b-T138A indeveloping SACs of the right eye of P3 rat pups (Fig. 5A). Thisin vivo electroporation approach allowed molecular perturbationwithin the critical period (P4–P8) of the eye-specific segregation ofretinogeniculate projection, especially in the SACs of nasal retinasof the DNA-injected eye (Fig. 5B and SI Appendix, Fig. S5). Con-sistent with our wave data from ex vivo transfection (Figs. 1 and 2),we found that in vivo transfecting SN25b may dampen the spatio-temporal properties of retinal waves (SI Appendix, Fig. S5 E–I).RGC axonal projections were observed by the intraocular in-
jection of anterograde axonal tracers that can fill axon terminalsin dLGN, i.e., the recombinant cholera toxin B subunit (CTB)conjugated to Alexa Fluor. At P8, each eye was injected withdifferent axonal tracers (with CTB-488 into the left eye andCTB-647 into the right eye), and then the segregation patterns of
the retinogeniculate projection were examined at P9 (Fig. 5A).In the dLGN sections containing labeled retinogeniculate af-ferents, fluorescent images of the contralateral and ipsilateralretinogeniculate termination sites were acquired for each col-ored axonal tracer (Fig. 5C). The degrees of eye-specific segre-gation could be quantified by computing the overlapping pixelsof the contralateral and ipsilateral retinogeniculate projection(Fig. 5C) (21–23), with an increase in the ratio of overlappingpixels demonstrating the disruption of eye-specific segregation(Fig. 5 D and E). Expression of SN25b in SACs impaired eye-specific segregation of the retinogeniculate projection comparedwith the Ctrl or SN25b-T138A, which was especially potent onthe DNA-injected side (Fig. 5 D and E). These results suggestthat PKA-mediated SN25b phosphorylation in SACs affects theactivity-dependent refinement of the retinogeniculate projection.Thus, SN25b in SACs can induce the global effect on visualcircuit refinement by regulating the eye-specific segregation ofthe retinogeniculate projection.
DiscussionIn this study, we show that developing SACs mainly expressSN25b. The level of SN25b expression is important for regula-tion of the stage II wave frequency based on several lines ofevidence. First, knockdown of retinal endogenous SN25b in-creases the frequency of wave-associated spontaneous Ca2+
transients. Second, the expression level of SN25 is gradually up-regulated until the developmental program is finished. Third,overexpression of SN25b in SACs decreases the strength oftransmission, dampens the frequency and spatial correlation ofwave-associated spontaneous Ca2+ transients, and impairs the
B
unin
ject
edin
ject
ed
ChATEGFP MergeA
Retina
Sclera
DNA + Fast Green
P3In vivo electroporation
+_
P8Axonal tracing
CTB488
CTB647
P9Heart perfusion
dLGN
injected sideuninjected side
C
DNA uninj. side DNA inj. side
Ctrl SN25b SN25b- T138A
Ove
rlap
Mer
ge
DNA uninj. side DNA uninj. sideDNA inj. side DNA inj. side
5 10 15 20 25 300
2
4
6
8
% o
verla
p
Contralateral thereshold
Ctrl SN25b SN25b-T138A
5 10 15 20 25 300
2
4
6
8
% o
verla
p
Contralateral threshold
Ctrl SN25b SN25b-T138A
DNA uninj. side DNA inj. sideE
***
D
*********
Fig. 5. SN25b in SACs impairs the eye-specific seg-regation of the retinogeniculate projection by phos-phorylation at T138. (A) Procedures for in vivoelectroporation. The mGluR2 promoter-driven con-structs were injected into the right eyes of P3 ratpups. (Inset) DNA injected into the subretinal spacebetween the retina and sclera. Following DNA in-jection, both eyes were covered by platinum elec-trodes (with the positive electrode on the right eye)for in vivo electroporation. Anterograde axonaltracers were injected into both eyes at P8 (with CTBconjugated-Alexa 488 into the left eye and CTB 647into the right). At P9, the rat brain was fixed by heartperfusion, and dLGN sections were imaged on theDNA-injected (Right) or uninjected side (Left). (B)Immunostaining of EGFP (green) and ChAT (red) inwhole-mount retinas (the 1.5-μm z-section) from theDNA-injected or uninjected eye following trans-fection. The colocalization signal (yellow). (C ) Eye-specific segregation of the retinogeniculate pro-jection in dLGN following SAC-specific expression ofCtrl (pmGluR2-IRES2-egfp), SN25b, or SN25b-T138A.(Upper) Merged axonal projections (yellow) from twoeyes. (Lower) Overlapping pixels shown as dark dotsin the dLGN (included by the yellow dash lines). (Scalebars: B, 30 μm; C, 100 μm.) (D and E) Quantification ofthe overlapping pixels in dLGN. (D) DNA-uninjectedside. ***P < 0.001, Repeated measures ANOVA withTukey post hoc test. (E) DNA-injected side. ***P <0.001, Repeated measures ANOVA with Tukey posthoc test. n = 6 pups for each group.
3266 | www.pnas.org/cgi/doi/10.1073/pnas.1812169116 Hsiao et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 2
1, 2
020

eye-specific segregation of retinogeniculate projection. Theseeffects require PKA-mediated phosphorylation of SN25b atT138. Therefore, during the critical developmental period, SN25bcan serve as a PKA substrate that mediates PKA regulationof retinal circuit activity and visual circuit refinement.Neural activity can modify the structure and function of neural
circuits during development (24). Vigorous neural activity elicitshigh PKA activity. PKA-mediated phosphorylation has beenshown to modulate the gating of ion channels, thereby regulatingthe circuit activity (24). The present study demonstrates that inaddition to ion channels, SN25b can serve as the PKA substratein presynaptic neurons, further regulating the circuit activityduring development. Particularly, developing neurons displayoscillated PKA activity (25), such that PKA activity is turned onand off following depolarizations on the timescale of ∼40 s in thedeveloping retinal neurons (10). In this scenario, PKA activity ishigh during the interwave interval, implying that a certain PKAsubstrate may be involved in decreasing the SAC release to causewave quiescence. Here, we found that PKA phosphorylation ofSN25b at T138 can inhibit the transmission from developingSACs, thus serving as a PKA substrate to down-regulate the wavespatial-temporal properties. Remarkably, the cAMP-PKA sig-naling displays dynamic spatial-temporal distributions in thedeveloping neurons (25). In addition to cAMP-dependent peri-odic depolarizations in regulating the wave refractory period,here we show that PKA-mediated SN25b phosphorylation candown-regulate wave periodicity from SACs. These results suggestthat multiple cAMP-dependent mechanisms may be involved indiverse regulation of the spatiotemporal properties of retinalwaves. Through dynamic changes in its phosphorylation state,SN25b may have the capacity to rapidly regulate the releaseprobability as well as the large-scale circuit activity, leading toalterations in activity-dependent synaptic refinement for theentire visual circuits.Misconnected circuits are common features of neurodevelop-
mental disorders such as schizophrenia and autism, so it is im-portant to elucidate the mechanisms by which synapse elimination
and synaptic refinement are regulated during development. Retinalwaves propagate through developing retinas and the thalamus to thevisual cortex, and they are essential for refining the visual sensorymap (1, 5). The stage II retinal waves from P0 to P9/10 in rodents(10, 26) have been completed by the time that SN25b expression isup-regulated (8, 9), suggesting that the expression level of SN25b inSACs during this critical period plays an important role in regulatingretinal waves and the refinement of eye-specific retinogeniculateprojections. Coincidentally, the level of SN25 expression has beenassociated with schizophrenia (27, 28). A recent study has revealedthat the SN25b/SN25a ratio is altered in schizophrenia patients (29).Therefore, the effects of PKA-mediated SN25b phosphorylation onvisual circuit refinement would provide insights into the neural cir-cuit development as well as the etiology of neurodevelopmentaldiseases (30).
Materials and MethodsPostnatal Sprague–Dawley rats were used in this study (P1–P2 for ex vivoretinal transfection; P3 for in vivo retinal transfection). All procedures wereperformed in accordance with protocols approved by the institutionalanimal care and use committees of National Taiwan University. Immuno-staining, retinal explant culture and transfection, live Ca2+ imaging of retinalwaves, and whole-cell voltage-clamp recordings were modified from pre-vious studies (7, 15). Detailed procedures regarding plasmid information,imaging of SAC exocytosis, in vivo electroporation, anterograde axonaltracing and tissue processing, analysis of the retinogeniculate projection, RT-qPCR, Western blotting, coimmunoprecipitation, and statistics are described inSI Appendix, SI Materials and Methods.
ACKNOWLEDGMENTS. We thank Drs. Meyer Jackson, Marla Feller, ShigetadaNakanishi, Hiroyuki Kamiguchi, and Li-Chung Hsu for the gift of the plasmid;the RNAi Core, Academia Sinica, Taiwan for shRNA plasmids; the staff ofTechnology Commons, College of Life Science, National Taiwan University(NTU) for help with confocal microscopy; and Dr. Eliza Wang and members ofthe C.-T.W. laboratory for help and discussion. This work was supportedby NTU; Ministry of Science and Technology Grants MOST-103-2311-B-002-026-MY3 and MOST-106-2311-B-002-007 (to C.-T.W.); the Chang GungMedical Research Project Grants CMRPD1G0061, CMRPD1G0501, CMRPD1G0502,and BMRPC03; and the Ministry of Science and Technology Grant MOST-107-2320-B-182-041 (to J.-C.L.).
1. Blankenship AG, Feller MB (2010) Mechanisms underlying spontaneous patternedactivity in developing neural circuits. Nat Rev Neurosci 11:18–29.
2. Zheng J, Lee S, Zhou ZJ (2006) A transient network of intrinsically bursting starburstcells underlies the generation of retinal waves. Nat Neurosci 9:363–371.
3. Ford KJ, Félix AL, Feller MB (2012) Cellular mechanisms underlying spatiotemporalfeatures of cholinergic retinal waves. J Neurosci 32:850–863.
4. Kirkby LA, Sack GS, Firl A, Feller MB (2013) A role for correlated spontaneous activityin the assembly of neural circuits. Neuron 80:1129–1144.
5. Ackman JB, Burbridge TJ, Crair MC (2012) Retinal waves coordinate patterned activitythroughout the developing visual system. Nature 490:219–225.
6. Südhof TC, Rizo J (2011) Synaptic vesicle exocytosis. Cold Spring Harb Perspect Biol 3:a005637.
7. Chiang CW, et al. (2012) Synaptotagmin I regulates patterned spontaneous activity inthe developing rat retina via calcium binding to the C2AB domains. PLoS One 7:e47465.
8. West Greenlee MH, Finley SK, Wilson MC, Jacobson CD, Sakaguchi DS (1998) Tran-sient, high levels of SNAP-25 expression in cholinergic amacrine cells during postnataldevelopment of the mammalian retina. J Comp Neurol 394:374–385.
9. Greenlee MH, Roosevelt CB, Sakaguchi DS (2001) Differential localization of SNAREcomplex proteins SNAP-25, syntaxin, and VAMP during development of the mam-malian retina. J Comp Neurol 430:306–320.
10. Dunn TA, et al. (2006) Imaging of cAMP levels and protein kinase A activity revealsthat retinal waves drive oscillations in second-messenger cascades. J Neurosci 26:12807–12815.
11. Risinger C, Bennett MK (1999) Differential phosphorylation of syntaxin andsynaptosome-associated protein of 25 kDa (SNAP-25) isoforms. J Neurochem 72:614–624.
12. Watanabe D, et al. (1998) Ablation of cerebellar Golgi cells disrupts synaptic in-tegration involving GABA inhibition and NMDA receptor activation in motor co-ordination. Cell 95:17–27.
13. Cutts CS, Eglen SJ (2014) Detecting pairwise correlations in spike trains: An objectivecomparison of methods and application to the study of retinal waves. J Neurosci 34:14288–14303.
14. Tojima T, et al. (2007) Attractive axon guidance involves asymmetric membranetransport and exocytosis in the growth cone. Nat Neurosci 10:58–66.
15. Huang PC, et al. (2014) Adenosine A(2A) receptor up-regulates retinal wave fre-quency via starburst amacrine cells in the developing rat retina. PLoS One 9:e95090.
16. Shatz CJ, Stryker MP (1988) Prenatal tetrodotoxin infusion blocks segregation ofretinogeniculate afferents. Science 242:87–89.
17. Sretavan DW, Shatz CJ, Stryker MP (1988) Modification of retinal ganglion cell axonmorphology by prenatal infusion of tetrodotoxin. Nature 336:468–471.
18. Penn AA, Riquelme PA, Feller MB, Shatz CJ (1998) Competition in retinogeniculatepatterning driven by spontaneous activity. Science 279:2108–2112.
19. Xu HP, et al. (2011) An instructive role for patterned spontaneous retinal activity inmouse visual map development. Neuron 70:1115–1127.
20. Matsuda T, Cepko CL (2007) Controlled expression of transgenes introduced by in vivoelectroporation. Proc Natl Acad Sci USA 104:1027–1032.
21. Torborg CL, Feller MB (2004) Unbiased analysis of bulk axonal segregation patterns.J Neurosci Methods 135:17–26.
22. Torborg CL, Hansen KA, Feller MB (2005) High frequency, synchronized burstingdrives eye-specific segregation of retinogeniculate projections. Nat Neurosci 8:72–78.
23. Stevens B, et al. (2007) The classical complement cascade mediates CNS synapseelimination. Cell 131:1164–1178.
24. Ganguly K, Poo MM (2013) Activity-dependent neural plasticity from bench to bed-side. Neuron 80:729–741.
25. Dunn TA, Feller MB (2008) Imaging second messenger dynamics in developing neuralcircuits. Dev Neurobiol 68:835–844.
26. Wang CT, et al. (2007) GABA(A) receptor-mediated signaling alters the structure ofspontaneous activity in the developing retina. J Neurosci 27:9130–9140.
27. Corradini I, Verderio C, Sala M, Wilson MC, Matteoli M (2009) SNAP-25 in neuropsy-chiatric disorders. Ann N Y Acad Sci 1152:93–99.
28. Thompson PM, Sower AC, Perrone-Bizzozero NI (1998) Altered levels of the synap-tosomal associated protein SNAP-25 in schizophrenia. Biol Psychiatry 43:239–243.
29. Houenou J, et al. (2017) A multilevel functional study of a SNAP25 at-risk variant forbipolar disorder and schizophrenia. J Neurosci 37:10389–10397.
30. Lewis DA, Levitt P (2002) Schizophrenia as a disorder of neurodevelopment. Annu RevNeurosci 25:409–432.
Hsiao et al. PNAS | February 19, 2019 | vol. 116 | no. 8 | 3267
NEU
ROSC
IENCE
Dow
nloa
ded
by g
uest
on
Mar
ch 2
1, 2
020