Embed Size (px)
Citation preview

PertanikaJ. Trop. Agric. Sci. 20(2/3): 121-134(1997) ISSN: 0126-6128© Universiti Putra Malaysia Press
Prey Spectra of Bornean Nepenthes Species (Nepenthaceae) inRelation to their Habitat
JUMAAT H. ADAMEnvironmental Sdence Programme
Faculty of Natural Resource SdenceUniversiti Kebangsaan Malaysia
43600 UKM Bangi, Selangor} Malaysia
Keywords: Prey spectra, Bornean Nepeuthes species, habitat
ABSTRAK
Kajian terhadap kandungan mangsa periuk 18 spesis Nepenthes di Borneo menunjukkan terdapat 17kumpulan fauna utama. Hasil kajian mendapati periuk lantai dan udara memerangkap kedua-dua faunarayap dan fauna terbang, pada umumnya periuk lantai memerangkap lebih banyak fauna rayap sementaraperiuk udara memerangkap lebih banyak fauna terbang. Kandungan jenis mangsa adalah berkorelasi denganhabitat, spesis yang tumbuh pada habitat berbeza dalam satu lokasi yang sama kadangkala menunjukkankandungan jenis mangsa yang berbeza. Formiddae didapati jenis mangsa yang kerap ditemui dan yang palingmelimpah, dan periuk yang dikutip pada altitud bawah daripada 100 m altitud mengandungi bilanganindividu semut yang tinggi tetapi bilangannya didapati menurun dengan peningkatan altitud. Walaubagaimanapun, bilangan semut diperangkap dalam setiap periuk adalah berbeza, contohnya bilangan semutyang ditemui dalam periuk N. rafflesiana adalah daripada beberapa individu sehingga 700 individu perperiuk, Spesis Nepenthes tanah tinggi memerangkap pelbagai jenis mangsa berbanding dengan spesisNepenthes tanah pamah. Spektra mangsa yang pelbagai ini menyediakan lebihan makanan kepada pemangsamenghuni periuk, terutama Arachnida, dan ianya didapati kerap ditemui pada spesis tanah tinggi.
ABSTRACT
Examination of pitcher prey contents of 18 Bornean pitcher plants showed that pitchers attract 17 fauna groups.The upper and lower pitchers trap both flying and creeping fauna but generally the lower pitchers trap morecreeping fauna and upper pitchers more flying prey spedes. Prey composition is generally correlated with habitat;spedes occupying different habitats in the same locality sometimes show striking differences in prey composition.Formiddae are the most abundant and frequently trapped, and pitchers collected below 100 m altitude containedenormous numbers of ants but their number generally decreased with altitude. However, the number of antscaught per pitcher was variable, e.g. from a few ants to 700 per pitcher in N. rafflesiana. Nepenthes spedesgrowing at high altitudes trapped a broader spectrum of prey than spedes at lower altitudes. This broad spectrumof prey provides an ample food source for the pitcher-inhabiting predators, particularly Arachnida, which arecommon in high altitude spedes.
INTRODUCTION
Nepenthes species, commonly known as pitcherplants, are tropical carnivorous plants, which gen-erally grow in areas of infertile soil such as inheath forests, swamp forests, forests on ultrabasicsoils, and in limestone forests. Nepenthes burbidgeae,N kinabaluensis, N, rajah and N. villosa grow onserpentinized ultrabasic rocks and on acid soilson Mt. Kinabalu (Meijer 1965; Mackinnon 1975;Kurata 1976; Kaul 1982). Nepenthes northiana
(Anderson 1965; Adam et al 1992), N clipeata(Danser 1928; Adam et al 1992) and N. mapuluensis(Adam and Wilcock 1990) are endemic to lime-stone in Borneo. The ability of Nepenthes speciesto thrive on poor soils is largely attributed to theircarnivorous habit of trapping prey in the pitch-ers, which are a modification of the leaf tip.
Nepenthes species display a carnivorous syn-drome, i.e. they attract, retain, trap, kill, digest,and absorb useful substances (Juniper 1986).

JUMAAT H. ADAM
The pitchers act as pitfall or passive traps (Lloyd1942), increasing their efficiency by a seductivedevice (Slack 1980), the secretion of nectar bynumerous glands on the under-surface of the lidand the margin of the inner peristome. Theeffectiveness of the trap is enhanced by thepresence of a waxy, slippery surface on theupper half of the inner surface of the pitcherand sharp descending inner peristome teeth.These teeth are very well developed in somespecies, for example, in Nepenthes bicalcarata, N.edxuardsiana, N. kinabaluensis, N. raffUsiana, Nrajah, and N villosa. The pitcher wall secretes adigestive fluid containing enzymes and the prod-ucts of digestion are absorbed by the same glands(Lloyd 1942).
The prey content of pitchers is varied andincludes insects like Hymenoptera (includingFormicidae), Isoptera, Coleoptera, Plecoptera,and Dermaptera, and other faunal groups suchas millipedes, and snails (Jensen 1910; Adamand Wilcock 1994). Pitcher contents of Nepenthesfrom Borneo have not been extensively studied(Spencer 1860; Slack 1980; Phillipps and Lamb1988). Slack (1980) noted that pitchers, notreferring to any particular species, containeddigestive bodies of large insects, such as cock-roaches and centipedes as well as scorpions,small mammals and reptiles. Spencer (1860)and Phillipps and Lamb (1988), in an extremecase, mentioned a large rat being trapped inpitchers of Nepenthes rajah. Jensen (1910) men-tioned the horrible odour arising from pitchersloaded with centipedes, cockroaches, butterflies,and scorpions found in Nepenthes near Tjibodas,Java.
The objectives of this study were as follows:(1) to investigate the prey spectra in pitchers of18 Bornean Nepenthes species; (2) to correlatetheir occurrence and abundance with altitude,habitat and pitcher morphology; (3) to investi-gate the prey-partitioning between upper andlower pitchers of 9 Nepenthes species; (4) to studyprey-partitioning between 18 Nepenthes speciesoccupying the same locality but growing in dif-ferent habitat types.
MATERIALS AND METHODS
The localities, habitat, altitude, number of pitch-ers sampled per species for prey pitcher con-tents and prey partitioning of 18 BorneanNepenthes are listed in Table 1. Prey pitchercontents were sampled 2 weeks after the pitcher
opened. The prey found in the pitchers arelisted in Table 2. A total of 255 pitchers weresampled, ranging from 4-47 pitchers per species.Eighteen species and nine species were investi-gated respectively for prey-partitioning betweenspecies (Table 3) and between upper and lowerpitchers within a species (Tables 4, 5 and 6).
Using a stereo microscope, the intact faunawere carefully separated from the soils and de-cayed insect debris from each sample and subse-quently preserved in 70% alcohol. Prey was iden-tified to class, order, or family level. Keys to theclass of Arthropoda or orders of Insecta (Borroret al 1954), and keys to the class and families ofBritish Insecta (Unwin 1981, 1984) were used.The number of individuals of each taxon in eachsample was counted and numbered accordingly.
Duplicate specimens were sent to the^Sarawak Forest Department Entomology Sectionand National Institute of Health, Kyoto. Speci-mens are deposited in Biology DepartmentMuseum, Universiti Kebangsaan Malaysia SabahCampus and Sarawak Forest Entomology Sec-tion. Analysis of principal component analysis(PCA) used a computer statistical programmeand Sorenson coefficients similarity (CC) wereused to determine fauna group similarity be-tween Nepenthes species (Mueller-Dombois andEllenberg 1974; Brower and Zar 1977).
CC = 2c X 100si + s2
where si and s2 are the number of fauna groupsin Nepenthes species 1 and 2, and c is the numberof faunal groups common in both species.
RESULTS
Diversity of Prey
Of the total 690 taxa collected from 255 pitchersof 18 Nepenthes species sampled, 53.9% and 1.2%were identified to family and generic level re-spectively. A total of 6384 individuals were re-corded of which 79.5% (5077 individuals) wereFormicidae (excluding other Hymenoptera),8.4% Isoptera, 3.8% Diptera, 2.6% in other mis-cellaneous groups, 2.0% Coleoptera, 1.2% Hy-menoptera; and less than 1% in each of theother 11 groups (Table 2).
The mean number of individuals per pitcherwas 25, but varied greatly between species, rang-ing from 1.7 individuals in N. bicalcarata to 168.9individuals in N macrovulgaris. Eight Nepenthesspecies had a mean less than 10 individuals per
122 PERTANIKA J. TROP. AGRIC. SCI. VOL. 20 NO. 2/3 , 1997
PREY SPECTRA OF BORNEAN NEPENTHES SPECIES (NEPENTHACEAE)
TABLE 1Locality, altitude and habitat of pitcher prey sampling sites
No
123456789
101112131415161718
Nepenthes species
N. albomarginataN. ampullarictN. bicalcarata***N. gracilis*N. rafflesiana***N. mirabilis*N. hookeriana**N. macrovulgaris*N. sandakanensis*N. lowii**N. muluensis*N. tentaculata*N. x alisaputmiana***N. curtisii**N. kinabaluensis***N. rajah***N. reinwarditiana*N villosa***
Locality
Weston, SabahWeston, SabahWeston, SabahWeston, SabahWeston, SabahTelupid, SabahTelupid, SabahML Silam, SabahMt. Silam, SabahMt. Mulu, SarawakMt. Mulu, SarawakMt. Mulu, SarawakMt. Kinabalu, SabahMt. Kinabalu, SabahMt. Kinabalu, SabahMt. Kinabalu, SabahMt. Kinabalu, SabahMt. Kinabalu, Sabah
Altitude(m)
5-305-30
55-305-30150150520
700-8001680
1800-23001600-23001900-1930
140028501950960
1600-2300
Habitat
HFDDFDDFSWFHFSVSVRCMFMFMFMFUFSBUFUFRCUF
No. pitcherssampled
1677
201610
47
14121623146
16101047
* Inner surface of lower and upper pitcher partly glandular** Inner surface of upper and lower pitcher wholly glandular, lower pitcher partly glandular*** Inner surface of upper and lower pitcher wholly glandular+ The only known species to have lower pitcher only, wholly glandularHF Heath forestSWF Swamp forestSV Secondary vegetationMF Mossy forestRC Roadside clearingDDF Disturbed dipterocarp forestUF Forest on ultrabasic soil
TABLE 2Prey spectra of eight Bornean Nepenthes species
Nepenthes
species*
(1)(1)(2)(2)(3)(3)(4)(4)(5)(5)(6)(6)(7)(7)(8)
Faunagroup
%No.%
No.%
No.%
No.%
No.%
No. .%
No.%
1
000000140000000
2
000000000000000
3
00
23300140000000
4
000000000000§00
5
0000000000
1.96000
6
000000000000000
7
000000
5.9241.943
00000
8
11.471
69.29
91.71190
36796.2223994.529110060
99.1
9
000000000000000
PERTANIKA J. TROP. AGRIC. SCI. VOL. 20 NO. 2/3, 1997 123
JUMAAT H. ADAM
Table 2 (cont'd)
Nepenthesspecies*
Faunagroup
(8)(9)
(9)(10)
(10)
(11)(11)(12)(12)(13)(13)(14)(14)(15)(15)(16)(16)(17)(17)(18)
(18)
No.
%No.
%No.%No.%No.%No.%No.%No.%No.%No.%No.
0
1.93
16.7400
7.566600
13.8142.5800
13.4
22
0000000000000440000813
0
6.21017417
92016550011
11143
113353
00000000
000
6.260
00000
12
0000000
00001133
000
035
00
0000
0
3.730000
000000
00
03150
8.3
223126
50
000
57585.9193.111
9.215
117142.569
45.811
36.519
41.233
7777
84.581
00
85.527581.7
29300
01
200
00
1080
0000000
270
0
Table 2 (cont'd)
Nepenthesspecies*
Faunagroup
10 11 12 13 14 15 16 17 Total Mean
(1)(1)(2)(2)(3)(3)(4)(4)(5)(5)(6)(6)(7)(7)(8)(8)(9)(9)(10)
(10)
(11)(11)(12)(12)(13)
%No.
%No.%No.%No.%No.%No.%No.%No.%No.%No.%No.
%
No.%
0000000000000000008.320000
2
00000
0000000000016260023
126.250
86.353400000000000000000000004
000000000000000000000000
2
007.11
8.31
000000
000000
4.21
0000
2
000000000000
0000000
000
1.310
o000000
0000
000
000000
001.330
2.31400
001.981.9433.611
00
0.9111.220
00
02.522
100619100
13100
12100
409100
232510030810060100
1182100
16210024100
5210080100
-38.7_
1.9_1.7_
20.4_
145.3„
30.8_
15.0_
168.9
11.6
2.0
3.3
3.5
124 PERTANIKA J. TROP. AGRIC. SCI. VOL. 20 NO. 2 /3 , 1997
PREY SPECTRA OF BORNEAN NEPENTHES SPECIES (NEPENTHACEAE)
Table 2 (cont'd)
Nepenthes
species*
(13)(14)(14)(15)(15)(16)(16)(17)(17)(18)(18)
Faunagroup
No.%
No.%
No.%
No.%
No.%
No.
10
23.1
3000000
3.15
11
00033
1.246
223.1
5
12
4000000
0
000
13
23.1
300000000
14
22,1
255
1.240000
15
00000000000
16
00000000000
17
20033
2.58
3.914
26.443
Total
10010096
100104100322100358100162
Mean
7.1-
16.0-
6.3-
32.2-
35-8-
3.5
KeyFauna group1 Arachnida2 Chilopoda3 Coleoptera4 Dermaptera5 Dictyoptera
6 Diplopoda7 Diptera8 Formicidae9 Homoptera
10 Heteroptera
11 Hymenopters (excluding 8)12 Isoptera13 Mollusca14 Orthoptera15 Plecoptera
16 Pscoptera17 Others
* Nepenthes speciesnumbered as in Table 1
pitcher but two had means of 145.3 (N.rafflesiana) and 168.9 (N macrovulgaris). Fifteenprey groups were identified to family level, oneprey group each to order and unknown faunalgroup respectively (Table 2). Six of these preygroups, Formicidae (which excludes other flyinghymenopteran taxa), Dictyoptera, Isoptera,Chilopoda, Diplopoda and Mollusca are creep-ing fauna. The other prey groups are flyinginsects.
The commonest group was Formicidae,found in abundance in pitchers of 16 Nepenthesspecies. Although the Formicidae were overallthe most common group, they were better rep-resented in lowland habitats where they formed69-100% of pitcher prey in seven of eight speciesfound below 500 m above sea level comparedwith 3-87% for species growing between 700-2300 m, and they were totally absent from pitch-ers in N. villosa (1600-2300 m) and N.kinabaluensis (2850 m).
Diptera were the second most commongroup although in much lower abundance. Inten of the species studied, they comprised 57%of the prey in pitchers of N. kinabaluensis, 31%in N. sandakanensis, and 23% in N. muluensis.While the pitchers of the remaining seven spe-cies contained between 2-9% Diptera. Sixty-fiveDipteron taxa were recognized, belonging tofamilies such as Phoridae, Syrphidae,Cecidomyiidae and Sciaridae.
Coleoptera, the third most common preygroup encountered, were recorded in ten spe-cies, being most abundant in pitchers of N.villosa (33%), N. ampullaria (23%), N tentaculata(20%), N. lowii (17%), N muluensis (17%) andN. kinabaluensis (11%). Eighty-two coleopterantaxa were recognized belonging to families suchas Curculionidae, Scarabaeidae, Elateridae,Chrysomilidae, Pyrochoridae, Odemeridae,Carabidae, Passalidae, Nitidulidae, Buprestidae,Cerambycidae and Lucanidae.
Hymenoptera (excluding Formicidae) werefound in ten species studied, being abundantin Nepenthes muluensis (23%) and N.sandakanensis (16%). Forty-six Hymenopterantaxa recognized belong to families such asChacidoidae, Apidae, Vespoidae,Trichgrammatidae and Icheu monidae. Hymenop-teran species recognized included Apis cerana(in N. muluensis), Trigona sp. (in iV. xalisaputraiana, N gracilis, and N rajah), andDacus sp. (in N gracilis). Bulbitermis sp. (Isoptera)were present in abundance in Nepenthesalbomarginata and comprised 86% of the prey.
The other 12 fauna groups were less com-mon, and each group never comprised morethan 10% of the pitcher prey of the 18 speciesof Nepenthes recorded (Table 2).
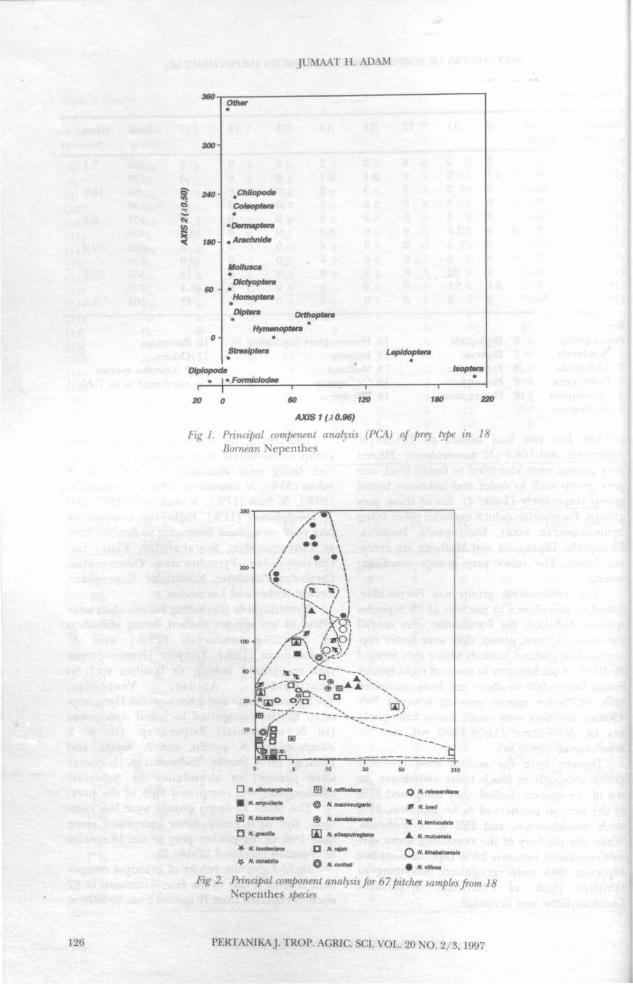
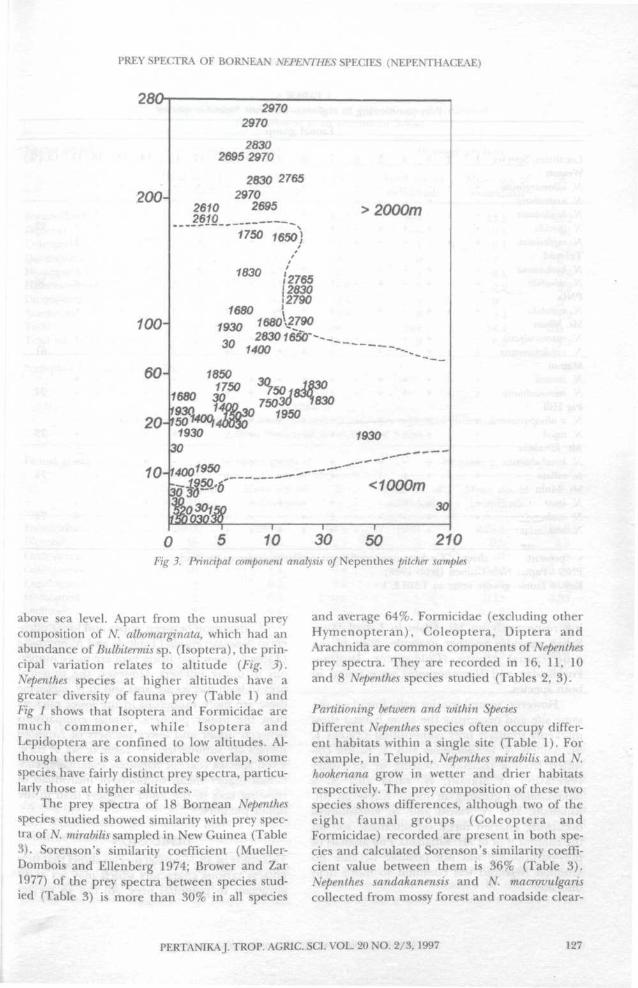
Fig. 1-3 give the results of principal compo-nent analysis (PCA) of the faunal contents of 67pitchers collected from 18 species from 30-2970 m
PERTANIKA J. TROP. AGRIC. SCI. VOL. 20 NO. 2 /3 , 1997 125
JUMAAT H. ADAM
300
240
180
60
Other
^Chilopode
Coieoptera*
•Dermapton
Molluscs
Dictyoptoram
Homoptera
Diptm* orthoptera
Hymenoptera
Stm&iptan
20 0 180 220SO 120
AXIS 1 (A 0.96)
Fig 1. Principal compenent analysis (PCA) of prey type in ISBornean Nepenthes
/f\(t
O1 i
/%0
50
D N. albomsrgtruta gg
• HmmpulW Q N. macrovutgaris
H N.bic*tc*r*u ^ UMmtatommJ*
® M«
0 N.
* N.lowtt
flt N.
• M
O lt
2. Principal component analysis for 67 pitcher samples from 18Nepenthes sp>
126 PERTANIKA J. TROP. AGRIC. SCI. VOL. 20 NO. 2/3 , 1997
PREY SPECTRA OF BORNEAN NEPENTHES SPECIES (NEPENTHACEAE)
28CH
200A
100-
60-
20-
29702970
28302695 2970
2830 27652970
2610 26952610 ^
1750 1650)
1830 I276512830\2790
1680 [
1930
^v *400
18501750 %fm 75^36—1830
>2000m
1930
<1000m30
0 5 10 30 50 210Fig 3. Principal component analysis of Nepenthes pitcher samples
above sea level. Apart from the unusual preycomposition of N. albomarginata, which had anabundance of Bulbitermis sp. (Isoptera), the prin-cipal variation relates to altitude (Fig. 3).Nepenthes species at higher altitudes have agreater diversity of fauna prey (Table 1) andFig 1 shows that Isoptera and Formicidae aremuch commoner, while Isoptera andLepidoptera are confined to low altitudes. Al-though there is a considerable overlap, somespecies have fairly distinct prey spectra, particu-larly those at higher altitudes.
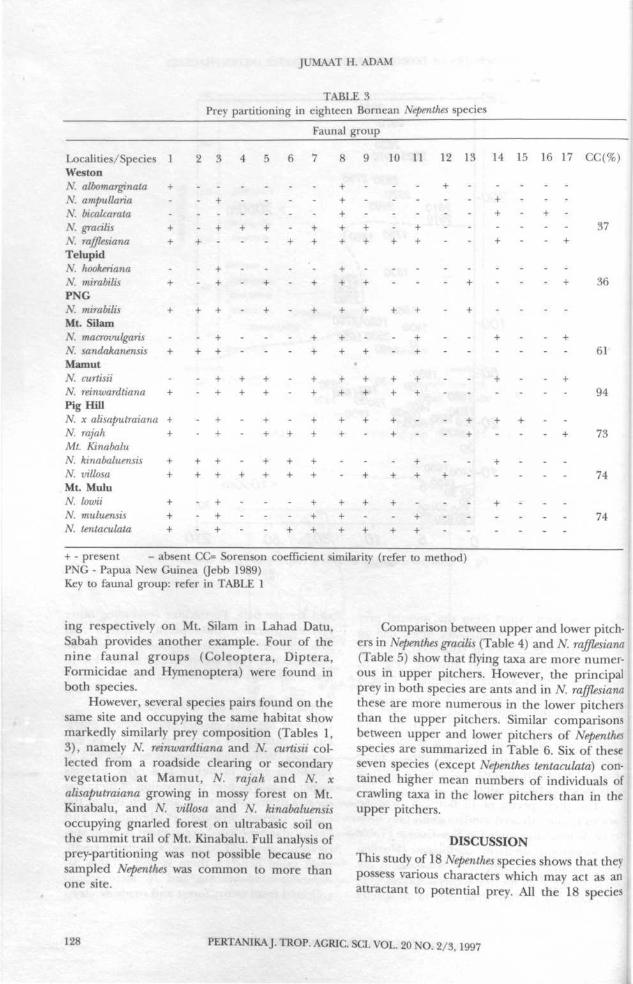
The prey spectra of 18 Bornean Nepenthesspecies studied showed similarity with prey spec-tra of N mirabilis sampled in New Guinea (Table3). Sorenson's similarity coefficient (Mueller-Dombois and Ellenberg 1974; Brower and Zar1977) of the prey spectra between species stud-ied (Table 3) is more than 30% in all species
and average 64%. Formicidae (excluding otherHymenopteran), Coleoptera, Diptera andArachnida are common components of Nepenthesprey spectra. They are recorded in 16, 11, 10and 8 Nepenthes species studied (Tables 2, 3).
Partitioning between and within Species
Different Nepenthes species often occupy differ-ent habitats within a single site (Table 1). Forexample, in Telupid, Nepenthes mirabilis and N.hookeriana grow in wetter and drier habitatsrespectively. The prey composition of these twospecies shows differences, although two of theeight faunal groups (Coleoptera andFormicidae) recorded are present in both spe-cies and calculated Sorenson's similarity coeffi-cient value between them is 36% (Table 3).Nepenthes sandakanensis and N. macrovulgariscollected from mossy forest and roadside clear-
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 20 NO. 2 /3 , 1997 127
JUMAAT H. ADAM
TABLE 3Prey partitioning in eighteen Bornean Nepenthes species
Faunal group
Localities/SpeciesWestonN. albomarginataN. ampullariaN. bicalcarataN. gracilisN. rafjlesianaTelupidN. hookerianaN. mirabilisPNGN. mirabilisMt. SilamN. macrovulgarisN sandakanensisMamutN. curtisiiN. reinwardtianaPig HillN. x alisaputraianaN. rajahMt. KinabaluN. kinabaluensisN. villosaMt. MuluN. loioiiN. muluensisN. tentaculata
2 3 7 8 9 10 11 12 13 14 15 16 17 CC(%)
37
61
74
74
+ - present - absent G O Sorenson coefficient similarity (refer to method)PNG - Papua New Guinea (Jebb 1989)Key to faunal group: refer in TABLE 1
ing respectively on Mt. Silam in Lahad Datu,Sabah provides another example. Four of thenine faunal groups (Coleoptera, Diptera,Formicidae and Hymenoptera) were found inboth species.
However, several species pairs found on thesame site and occupying the same habitat showmarkedly similarly prey composition (Tables 1,3), namely N. reinwardtiana and N. curtisii col-lected from a roadside clearing or secondaryvegetation at Mamut, N. rajah and N. xalisaputraiana growing in mossy forest on Mt.Kinabalu, and N. villosa and TV. kinabaluensisoccupying gnarled forest on ultrabasic soil onthe summit trail of Mt. Kinabalu. Full analysis ofprey-partitioning was not possible because nosampled Nepenthes was common to more thanone site.
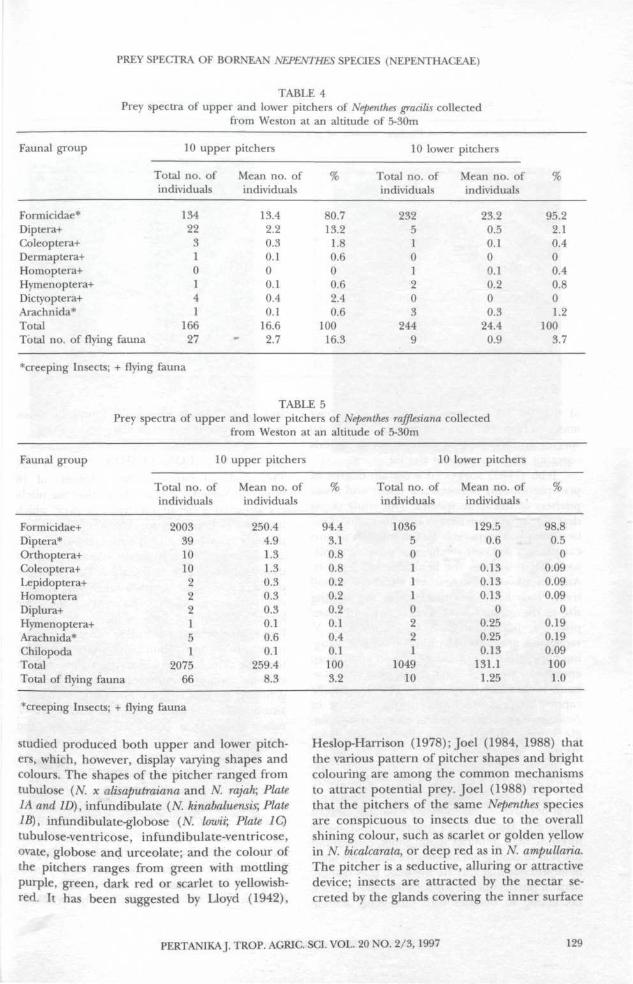
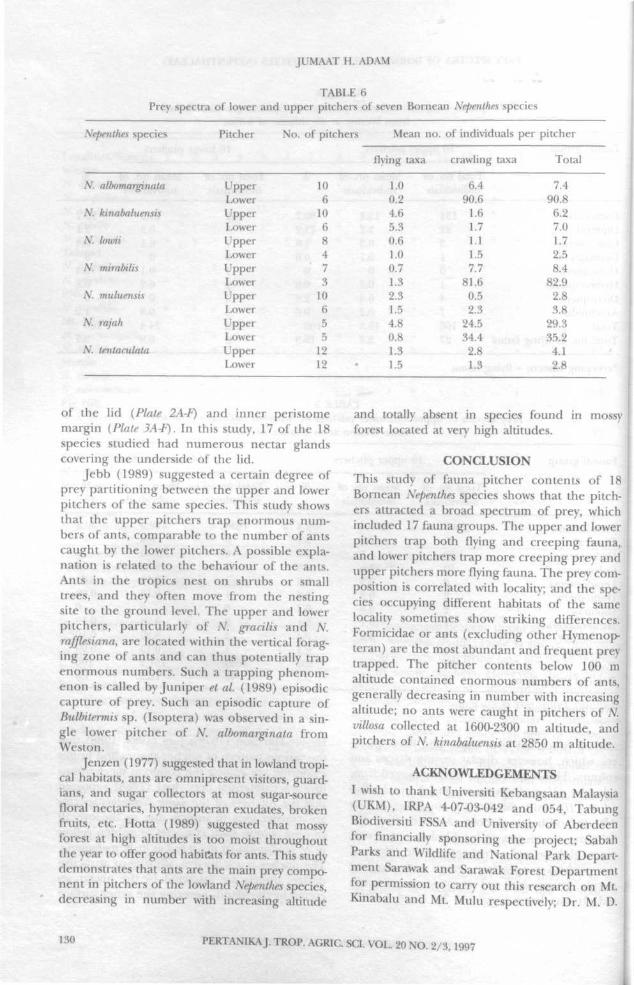
Comparison between upper and lower pitchers in Nepenthes gracilis (Table 4) and N rafjlesiana(Table 5) show that flying taxa are more numer-ous in upper pitchers. However, the principalprey in both species are ants and in N rafjlesianathese are more numerous in the lower pitchersthan the upper pitchers. Similar comparisonsbetween upper and lower pitchers of Nepenthesspecies are summarized in Table 6. Six of theseseven species (except Nepenthes tentaculata) con-tained higher mean numbers of individuals ofcrawling taxa in the lower pitchers than in theupper pitchers.
DISCUSSIONThis study of 18 Nepenthes species shows that theypossess various characters which may act as anattractant to potential prey. All the 18 species
128 PERTANIKAJ. TROP. AGRIC. SCI. VOL. 20 NO. 2/3, 1997
PREY SPECTRA OF BORNEAN NEPENTHES SPECIES (NEPENTHACEAE)
TABLE 4Prey spectra of upper and lower pitchers of Nepenthes gradlis collected
from Weston at an altitude of 5-30m
Faunal group
Formicidae*Diptera+Coleoptera+Dermaptera+Homoptera+Hymenoptera+Dictyoptera+Arachnida*TotalTotal no. of flying fauna
10 upper
Total no, ofindividuals
13422310141
16627
pitchers
Mean no. ofindividuals
13.42.20.30.100.10.40.1
16.62.7
%
80.713.2
1.80.600.62.40.6
10016.3
10 lower
Total no. ofindividuals
2325101
203
2449
pitchers
Mean no. ofindividuals
23.20.50.100.10.200.3
24.40.9
%
95,22,10,400.40.801.2
1003.7
*creeping Insects; + flying fauna
TABLE 5Prey spectra of upper and lower pitchers of Nepenthes rafflesiana collected
from Weston at an altitude of 5-30m
Faunal group
Formicidae+Diptera*Orthoptera+Coleoptera+Lepidoptera+HomopteraDiplura+Hymenoptera+Arachnida*ChilopodaTotalTotal of flying fauna
10
Total no. ofindividuals
2003391010222151
207566
upper pitchers
Mean no. ofindividuals
250.44.91,31.30.30.30.30.10.60.1
259.48.3
%
94.43.10.80.80.20.20.20.10.40.11003.2
10
Total no. ofindividuals
1036501110
221
104910
lower pitchers
Mean no. ofindividuals
129.50.6
00.130.130.13
0
0.250.250.13
131.1L25
%
98.80,5
0
0.090.090.09
0
0.190.190.091001.0
^creeping Insects; + flying fauna
studied produced both upper and lower pitch-ers, which, however, display varying shapes andcolours. The shapes of the pitcher ranged fromtubulose (TV. x alisaputraiana and TV. rajah; Plate1A and ID), infundibulate (TV. kinabaluensisr, PlateIB), infundibulate-globose (TV. lowii\ Plate 1Qtubulose-ventricose, infundibulate-ventricose,ovate, globose and urceolate; and the colour ofthe pitchers ranges from green with mottlingpurple, green, dark red or scarlet to yellowish-red. It has been suggested by Lloyd (1942),
Heslop-Harrison (1978); Joel (1984, 1988) thatthe various pattern of pitcher shapes and brightcolouring are among the common mechanismsto attract potential prey. Joel (1988) reportedthat the pitchers of the same Nepenthes speciesare conspicuous to insects due to the overallshining colour, such as scarlet or golden yellowin TV. bicalcarata, or deep red as in TV. ampullaria.The pitcher is a seductive, alluring or attractivedevice; insects are attracted by the nectar se-creted by the glands covering the inner surface
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 20 NO, 2/3, 1997 129
JUMAAT H. ADAM
TABLE 6Prey spectra of lower and upper pitchers of seven Bornean Nepenthes species
Nepenthes species
N. albomarginata
X. kinabaluensis
N. lowii
N. miraUlis
N. muluensis
N. rajah
N. tentaculata
Pitcher
UpperLowerUpperLowerUpperLowerUpperLowerUpperLowerUpperLowerUpperLower
No. of pitchers
106
1068473
10655
1212
Mean no.
flying taxa
L00.24.65.30.61.00.71.32.31.54.80.81.31.5
of individuals per
crawling taxa
6.490.6
1.61.71.11.57.7
81.60.52.3
24.534.42.81.3
pitcher
Total
7.490.8
6.27.01.72.58.4
82.92.83.8
29.335.24.12.8
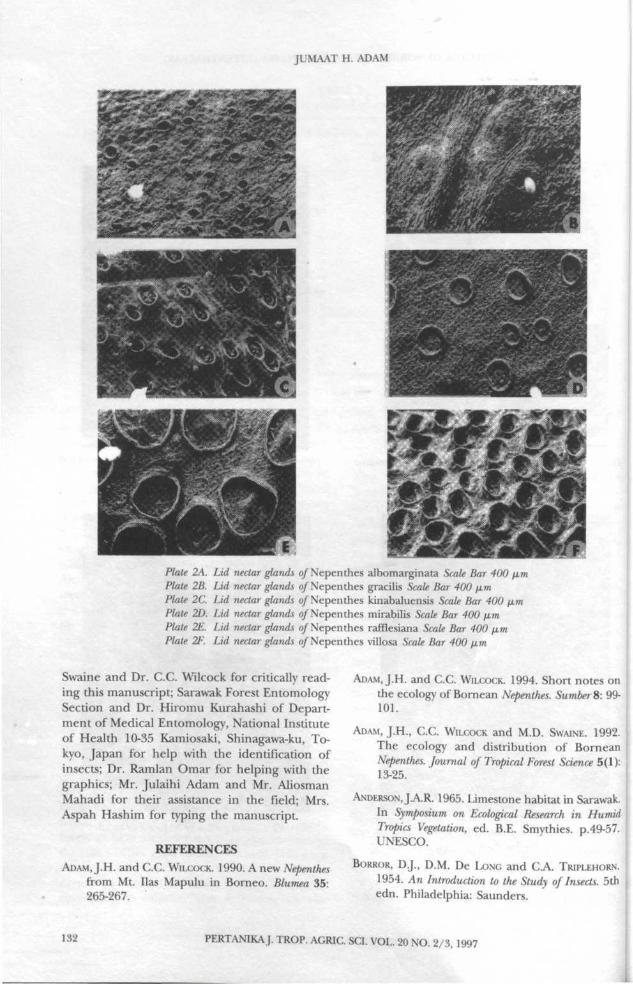
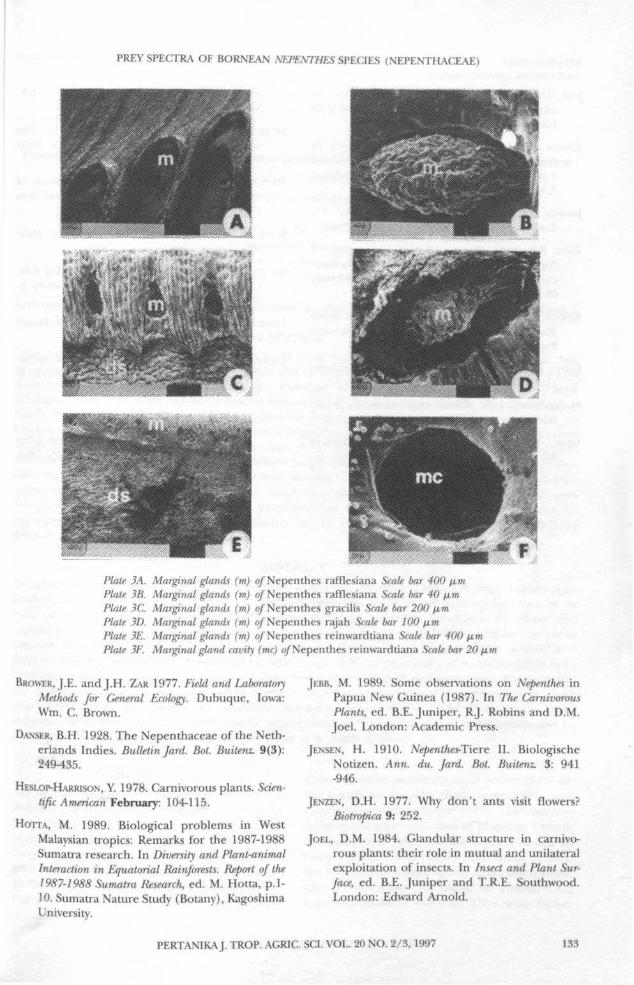
of the lid (Plate 2A-F) and inner peristomemargin (Plate 3A-F). In this study, 17 of the 18species studied had numerous nectar glandscovering the underside of the lid.
Jebb (1989) suggested a certain degree ofprey partitioning between the upper and lowerpitchers of the same species. This study showsthat the upper pitchers trap enormous num-bers of ants, comparable to the number of antscaught by the lower pitchers. A possible expla-nation is related to the behaviour of the ants.Ants in the tropics nest on shrubs or smalltrees, and they often move from the nestingsite to the ground level. The upper and lowerpitchers, particularly of N. gracilis and N.rafflesiana, are located within the vertical forag-ing zone of ants and can thus potentially trapenormous numbers. Such a trapping phenom-enon is called by Juniper et al (1989) episodiccapture of prey. Such an episodic capture ofBulbitermis sp. (Isoptera) was observed in a sin-gle lower pitcher of N. albomarginata fromWeston.
Jenzen (1977) suggested that in lowland tropi-cal habitats, ants are omnipresent visitors, guard-ians, and sugar collectors at most sugar-sourcefloral nectaries, hymenopteran exudates, brokenfruits, etc. Hotta (1989) suggested that mossyforest at high altitudes is too moist throughoutthe year to offer good habitats for ants. This studydemonstrates that ants are the main prey compo-nent in pitchers of the lowland Nepenthes species,decreasing in number with increasing altitude
and totally absent in species found in mossyforest located at very high altitudes.
CONCLUSIONThis study of fauna pitcher contents of 18Bornean Nepenthes species shows that the pitch-ers attracted a broad spectrum of prey, whichincluded 17 fauna groups. The upper and lowerpitchers trap both flying and creeping fauna,and lower pitchers trap more creeping prey andupper pitchers more flying fauna. The prey com-position is correlated with locality; and the spe-cies occupying different habitats of the samelocality sometimes show striking differences.Formicidae or ants (excluding other Hymenop-teran) are the most abundant and frequent preytrapped. The pitcher contents below 100 maltitude contained enormous numbers of ants,generally decreasing in number with increasingaltitude; no ants were caught in pitchers of N.villosa collected at 1600-2300 m altitude, andpitchers of N kinabaluensis at 2850 m altitude.
ACKNOWLEDGEMENTS
I wish to thank Universiti Kebangsaan Malaysia(UKM), IRPA 4-07-03-042 and 054, TabungBiodiversiti FSSA and University of Aberdeenfor financially sponsoring the project; SabahParks and Wildlife and National Park Depart-ment Sarawak and Sarawak Forest Departmentfor permission to carry out this research on Mt.Kinabalu and Mt. Mulu respectively; Dr. M. D.
130 PERTANIKAJ. TROP. AGR1C. SCI. VOL. 20 NO. 2/3, 1997

PREY SPECTRA OF BORNEAN NEPEN7HES SPECIES (NEPENTHACEAE)
Plate 1A. Lower pitcher of Nepenthes x alisaputraianaScale bar 4 cm
Plate IB. I'ppn pitcher of Nepenthes kinabaluensisScale bar 6 cm
Ptate }('.. Upper pitcher of Nepenthes lowiScale bar 6 cm
Plate ID. Ground pitcher of Nepenthes rajahScale bar 8 cm
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 20 NO. 2 /3 , 1997 131
JUMAAT H. ADAM
Plate 2A. Lid nectar glands of Nepenthes albomarginata Scale Bar 400 amPlate 2B. Lid nectar glands of Nepenthes gracilis Scale Bar 400 fxmPlate 2C. Lid nectar glands of Nepenthes kinabaluensis Scale Bar 400 urnPlate 2D. Lid nectar glands of Nepenthes mirabilis Scale Bar 400 fxmPlate 2E. Lid nectar glands of Nepenthes rafflesiana Scale Bar 400 fjumPlate 2F. Lid nectar glands of Nepenthes villosa Scale Bar 400 fxm
Swaine and Dr. C.C. Wilcock for critically read-ing this manuscript; Sarawak Forest EntomologySection and Dr. Hiromu Kurahashi of Depart-ment of Medical Entomology, National Instituteof Health 10-35 Kamiosaki, Shinagawa-ku, To-kyo, Japan for help with the identification ofinsects; Dr. Ramlan Omar for helping with thegraphics; Mr. Julaihi Adam and Mr. AliosmanMahadi for their assistance in the field; Mrs.Aspah Hashim for typing the manuscript.
REFERENCESADAM, J.H. and C.C. WILCOCK. 1990. A new Nepenthes
from Mt. Has Mapulu in Borneo. Blumea 35:265-267.
ADAM, J.H. and C.C. WILCOCK. 1994. Short notes onthe ecology of Bornean Nepenthes. SumberS: 99-101.
ADAM, J.H., C.C. WILCOCK and M.D. SWAINE. 1992.The ecology and distribution of BorneanNepenthes. Journal of Tropical Forest Science 5(1):15-25.
ANDERSON, J.A.R. 1965. Limestone habitat in Sarawak.In Symposium on Ecological Research in HumidTropics Vegetation, ed. B.E. Smythies. p.49-57.UNESCO.
BORROR, D.J., D.M. De LONG and C.A. TRIPLEHORN.
1954. An Introduction to the Study of Insects. 5thedn. Philadelphia: Saunders.
132 PERTANIKAJ. TROP. AGRIC. SCI. VOL. 20 NO. 2/3, 1997
PREY SPECTRA OF BORNEAN NEPENTHES SPECIES (NEPENTHACEAE)
Plate 3A. Marginal glands (m) of Nepenthes rafHesiana Scale bar 400 fimPlate 3B. Marginal glands (m) q/" Nepenthes rafflesiana Scale bar 40 fimPlate 3C. Marginal glands (m) of Nepenthes gracilis Scale bar 200 fimPlate 3D. Marginal glands (m) of Nepenthes rajah Scale bar 100 jxmPlate 3E. Marginal glands (m) of Nepenthes reinwardtiana Scale bar 400 fimPlate 3F. Marginal gland cavity (me) o/Nepenthes reinwardtiana Scale bar 20 fim
BROWER, J.E. and J.H. ZAR 1977. Field and Laboratory
Methods for General Ecology. Dubuque, Iowa:Wm. C. Brown.
DANSER, B.H. 1928. The Nepenthaceae of the Neth-
erlands Indies. Bulletin Jard. Bot. Buitenz. 9(3):249-435.
HESLOP-HARRISON, Y. 1978. Carnivorous plants. Scien-tific American February: 104-115.
HOTTA, M. 1989. Biological problems in WestMalaysian tropics: Remarks for the 1987-1988Sumatra research. In Diversity and Plant-animalInteraction in Equatorial Rainforests. Report of the1987-1988 Sumatra Research, ed. M. Hotta, p.l-10. Sumatra Nature Study (Botany), KagoshimaUniversity.
JEBB, M. 1989. Some observations on Nepenthes inPapua New Guinea (1987). In The CarnivorousPlants, ed. B.E. Juniper, RJ. Robins and D.M.Joel. London: Academic Press.
JENSEN, H. 1910. Nepenthes-Tiere II. Biologische
Notizen. Ann. du. Jard. Bot. Buitenz. 3: 941-946.
JENZEN, D.H. 1977. Why don't ants visit flowers?Biotropica 9: 252.
JOEL, D.M. 1984. Glandular structure in carnivo-rous plants: their role in mutual and unilateralexploitation of insects. In Insect and Plant Sur-face, ed. B.E. Juniper and T.R.E. Southwood.London: Edward Arnold.
PERTANIKAJ. TROP. AGRIC SCI. VOL. 20 NO. 2/3, 1997 133

JUMAAT H. ADAM
JOEL, D.M. 1988. Mimicry and mutualism in car-nivorous pitcher plants. Biological Journal of theLinnean Society 35(2): 185-197.
JUNIPER, B.E. 1986. The path to plant carnivory. InInsect and Plant Surface, ed. B.E. Juniper andTR.E. Southwood, p.195-218. London: EdwardArnold.
JUNIPER, B.E., RJ. ROBINS and D.M.JOEL 1989. The
Carnivorous Plants. London: Academic Press.
KAUL, R.B. 1982. Floral and fruit morphology ofNepenthes lowii and Nepenthes villosa, montanecarnivores of Borneo. American Journal of Hot a ti y69: 793-803.
KURATA, S. 1976. Nepenthes of Mt. Kinabalu. SabahNational Parks Publication No.2. Sabah NationalPark Trustees.
, F.E. 1942. The Carnivorous Plants. Waltham,Mass: Chronica Botanica.
MACKINNON, J. 1975. Borneo. Amsterdam: Time LifeBooks.
MEIJFT, W. 1965. A botanical guide to the flora ofMt. Kinabalu. In Symposium on Ecological Re-
search in Humid Tropics Vegetation, ed. B.E.Smythies, p. 325-364. UNESCO.
Mi i I.LER-DOMBOIS, D. and H. ELLENBERG. 1974. Aims
and Methods of Vegetation Ecology. New York:Wiley.
PHILLIPPS, A. and A. LAMB. 1988. Pitcher-plants of
East Malaysia and Brunei. Nature Ma/aysiana13(4): 8-27.
SLACK, A. 1980. Carnivorous Plants. Cambridge, Mass:MIT Press.
SPENCER, J. 1860. Life in the forests of the Far East.Gardener's Chronicle and Agriculture Gazette p.599-600.
UNWIN, D.M. 1981. Key to the families of British jDiptera. Field Studies 5: 513-533.
UNWIN, D.M. 1984. Key to the families of BritishColeoptera and Strepsiptera. Field Studies 6:149-197.
(Received 6 March 1996)(Accepted 10 July 1997)
,
134 PERTANIKAJ. TROP. AGRIC. SCI. VOL. 20 NO. 2/3, 1997





























