Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 288:474-492 (1989)
Primate Spinothalamic Pathways: II. The Cells of Origin of the Dorsolateral and
Ventral Spinothalamic Pathways
A. VANIA APKARIAN AND CHARLES J. HODGE Neurosurgery Department, SUNY Health Science Center, Syracuse, New York 13210
ABSTRACT The cells of origin of the dorsolateral (DSTT) and the ventral (VSTT)
spinothalamic tracts were studied in 11 monkeys. The spinothalamic tract cells were retrogradely labeled by horseradish peroxidase (HRP) injected in the thalamus. All animals also received a midthoracic spinal cord lesion on the side ipsilateral to the thalamic injections.
The distribution of labeled cells found in these animals throughout the cervical segments was similar to animals with no spinal cord lesions. Five ani- mals had ventral quadrant lesions to demonstrate the cells of origin of the DSTT. In macaques with complete ventral quadrant lesions, more than 80%# of the HRP label in the contralateral L4-L7 segments was located in lamina I, while in squirrel monkeys, the label in the contralateral lower lumbar region was distributed between laminae 1-111 and IV-VI. Few labeled cells were found in laminae VII-X.
Six animals received dorsolateral funiculus lesions to demonstrate the cells of origin of the VSTT. In animals with adequate lesions, 84-99% of the contralateral HRP label in L4-L7 was located in laminae IV-X. Macaques had a larger percentage of labeled cells located in lamina I than squirrel monkeys.
The results indicate the existence of two spinothalamic pathways in the primate. The DSTT was calculated to compose about one fourth of the total spinothalamic population.
Key words: dorsolateral funiculus, ventral quadrant, spinal cord, macaque monkey, squirrel monkey
The spinothalamic tract (STT), which is capable of relay- ing nociceptive information directly to the thalamus, is a crossed tract. I t is thought to be composed of axons situated in the ventral quadrant (VQ) of the spinal cord white mat- ter. A large body of evidence supports this notion. Section- ing of the spinal cord VQ results in anterograde degenera- tion in the thalamus in cats, monkeys, and humans (Mott, 1895; Bowsher, '57; Mehler et al., '60; Mehler, '62; Boivie, '71, '79; Kerr and Lippman, '74; Kerr, '75a; Berkley, '80, '83; Mantyh, '83a,b) and results in retrograde chromatolysis of cells in the contralateral spinal cord gray matter below the level of the lesion (Foerster and Gagel, '32; Kuru, '49; Morin et al., '51; Kerr, '75b; Smith, '76). Spinal cord VQ lesions decrease thalamic neuronal responses to peripheral noxious stimuli (Kenshalo et al., '80; Peschanski et al., '85; Chung e t al., '86) and abolish antidromic activation of spinal cord cells from the contralateral thalamus (Giesler et al., '81), while thalamic nociceptive responses can be recorded in ani- mals with only the ventral quadrants spared (Per1 and Whitlock, '61; Kenshalo et al., '80; Peschanski et al., '85;
Chung et al., '86). Antidromic activation of spinothalamic units by thalamic stimulation results in axonal action poten- tials recorded in the ipsilateral VQ of the spinal cord (Ap- plebaum et al., '75). Axons labeled with horseradish peroxi- dase (HRP) are found within the ipsilateral VQ in rats, cats, and monkeys after HRP injections in the thalamus (Willis et al., '79; Giesler et al., '81; Jones et al., '85).
Many studies implicate the spinothalamic pathway in the transmission of nociceptive, potentially painful, signals to the thalamus through the ventral quadrant. A large percent- age of the spinothalamic neurons respond to noxious so- matic and/or visceral stimuli (Willis et al., '74; Giesler et al., '76; Chung et al., '79; Fox et al., '80; Milne et al., '81; Ammons et al., '85; Blair, '85; Craig and Kni f i i , '85; Ammons, '87). Nociceptive responsive neurons are found in thalamic regions where the axons of the spinothalamic path- way terminate in rat (Guilbaud et al., '77, '80; Zemlan et al.,
Accepted May 2,1989.
0 1989 ALAN R. LISS, INC.

PRIMATE SPINOTHALAMIC PATHWAYS: I1
'78; Peschanski et al., '83), cat (Poggio and Mountcastle, '60; Per1 and Whitlock, '61; Honda et al., '83; Knif i i and Mizu- mura, '83; Hodge et al., '87; Knif€ki and Vahle-Hinz, '87), and monkey (Applebaum et al., '75, '79; Kenshalo et al., '80; Chung et al., '86; Yokota et al., '87). Spinal cord lesions of the VQ in monkeys and humans cause elevation of the thresholds of responses to noxious cutaneous stimuli contra- laterally below the level of the lesion (White and Sweet, '69; Vierck and Luck, '79; Greenspan et al., '86, '87). Stimulation of the VQ causes thermal and pain sensations in man (Mayer et al., '75; Tasker et al., '76). And pain can be trans- mitted rostrally in humans with only the VQ intact (Noor- denbos and Wall, '76). Given this large body of evidence indicating that STT axons lie in the VQ, little attention was paid to the possibility that some STT axons could lie out- side the ventral quadrant despite the report of Kuru ('49) describing the existence of a dorsolateral spinothalamic tract in humans and the hypothesis that there exist alter- nate pain pathways to account for the return of pain sensa- tion after anterolateral cordotomies (White and Sweet, '69).
McMahon and Wall ('83) observed that electrical stimu- lation of the cervical spinal cord white matter more fre- quently activated lumbar lamina I cells when electrodes were placed in the dorsolateral funiculus (DLF) compared to when electrodes were placed in the VQ. They concluded that about 80% of the axons of lamina I cells that project cephalad are located in the DLF and, hence, are not part of the spinothalamic pathway. Anatomic experiments in our laboratory (Apkarian et al., '85), performed in the cat, showed that 98% of the axons of lumbar lamina I cells pro- jecting cephalad lie in the DLF in the thoracic spinal cord, implying that any pathway with a lamina I cell component should have axons located in the DLF. Since Carstens and Trevino ('78) demonstrated that lamina I cell axons termi- nate in the thalamus in the cat, we investigated the possibil- ity of the existence of a spinothalamic pathway in the DLF in the cat (Jones et al., '85, '87). When thalamic horseradish peroxidase (HRP) injections were combined with midtho- racic ipsilateral VQ lesions, 87% of the contralateral labeled cells in the lumbar enlargement below the lesion were located in laminae 1-111, most of which were in lamina I. On the other hand, when thalamic HRP injections were com- bined with DLF lesions, only 6% of the total contralaterally projecting cells in the lumbar enlargement were found in laminae 1-111 (Jones et al., '87). Therefore, in the cat, there are two distinct spinothalamic pathways: a dorsolateral tract (DSTT) made up primarily of axons originating from contralateral spinal cord lamina I cells and a ventral spino- thalamic tract (VSTT) made up of axons of cells located in laminae IV-X.
There are important species differences between cats and primates regarding nociception. Sherrington and Laslett ('03) noted that unilateral VQ lesions in cats lead to bilateral deficits in responses to noxious stimuli. On the other hand, humans and nonhuman primates exhibit primarily contra- lateral deficits in nociceptive discrimination following uni- lateral VQ lesions (White and Sweet, '69; Vierck and Luck, '79). Cordotomy studies by Kennard ('54) pointed out that the dorsolateral funiculi were essential for nociceptive re- sponses in cats. Casey and Morrow ('88) have shown that, depending on the extent of the DLF lesion, it is possible to obtain a diminution or increase in nocifensive behavior bilaterally in cats. However, cordotomy studies in monkeys indicate primarily hyperalgesia following DLF lesions
475
(Vierck, personal communication). There are differences between cats and monkeys in the cells of origin of the STT (see Apkarian and Hodge '89a, this issue) as well as in the STT termination sites (in the cat the lateral terminations are to the border zone of the VPL/VPM complex, while the monkey STT terminates within these nuclei; Boivie, '71; Berkley, '80; Mantyh, '83a,b; Craig and Burton, '85; Craig, '87), and possibly even in the responses of STT cells in equivalent populations. (Lamina I STT cells are reported to be nociceptive specific in cats, while in monkeys some lam- ina I STT cells have wide dynamic range type responses; Craig and Knif€ki, '85; Ferrington et al., '87.) Given these differences, the effort to generalize the existence of the DSTT to primates becomes very tenuous until its existence is directly demonstrated.
The purpose of this study was to determine if there is a dorsolateral component of the STT in primates. The label found in the spinal cord of new world and old world mon- keys with thalamic HRP injections and various midthoracic spinal cord white matter lesions was compared to the label seen in animals with no spinal cord lesions as described in Apkarian and Hodge, ('89a). The results indicate, with some differences between new and old world monkeys, the exis- tence of a DSTT and a VSTT in primates.
MATERIALS AND METHODS Ten old world monkeys (Macaca fascicularis) weighing
between 2.0 and 3.5 kg and six new world monkeys (Saimiri sciureus) weighing between 0.6 and 1.4 kg were used. All surgical procedures were performed under sterile conditions in an animal operating room. The animals were anesthetized with intramuscular ketamine (75 mg/kg) and intravenous nembutal (5 mg/kg) and supplemented with nembutal whenever they exhibited withdrawal responses to noxious stimuli. The details of the surgical procedure, thalamic injections, survival times, perfusates used, and the tetrame- thy1 benzidine reaction were identical to those described in the previous paper. All the animals in this study received a midthoracic spinal cord white matter lesion on the side ipsi- lateral to the thalamic HRP injections. The lesions included ventral quadrant (VQ) lesions (i.e., lesions of the cord ante- rior to the dentate ligament), dorsolateral funiculus (DLF) lesions (i.e., lesions of the cord dorsal to the dentate liga- ment and ventral to the dorsal root entry zone), and dorsal column lesions. The thalamic injections were aimed to cover all of the somatosensory thalamus on one side, as described in Apkarian and Hodge ('89a). The injections were targeted to include caudal and oral portions of the ventral posterior lateral thalamus (VPLc, VPLo), the centrolateral nucleus (CL), the medial dorsal nucleus (MD), and the nucleus sub- medius (SM). The coordinates for these targets were calcu- lated by using the atlases of Olszewski ('52) and Szabo and Cowan ('84) for the old world monkeys and the atlas of Emmers and Akert ('63) for the new world monkeys. In some animals the total volume of the 2 % wheat germ agglu- tinin-conjugated horseradish peroxidase (WGA-HRP) in- jected in the thalamus was increased from 1 pl to 2 p1. The thalamus and some of the brainstem tissue as well as most of the spinal cord was cut at 60-80 pm and processed for HRP. Spinal cord tissue containing the lesion site was stored in fixative for at least a week, imbedded in paraffin, cut, and stained with Luxol fast blue and cresyl violet in order to visualize the extent of the spinal cord lesions. Stained and unstained HRP-processed thalamic tissue was used for re-
MF
Vl
SQ
VI
L
MF
V2
S
QV
2
L
MF
V3
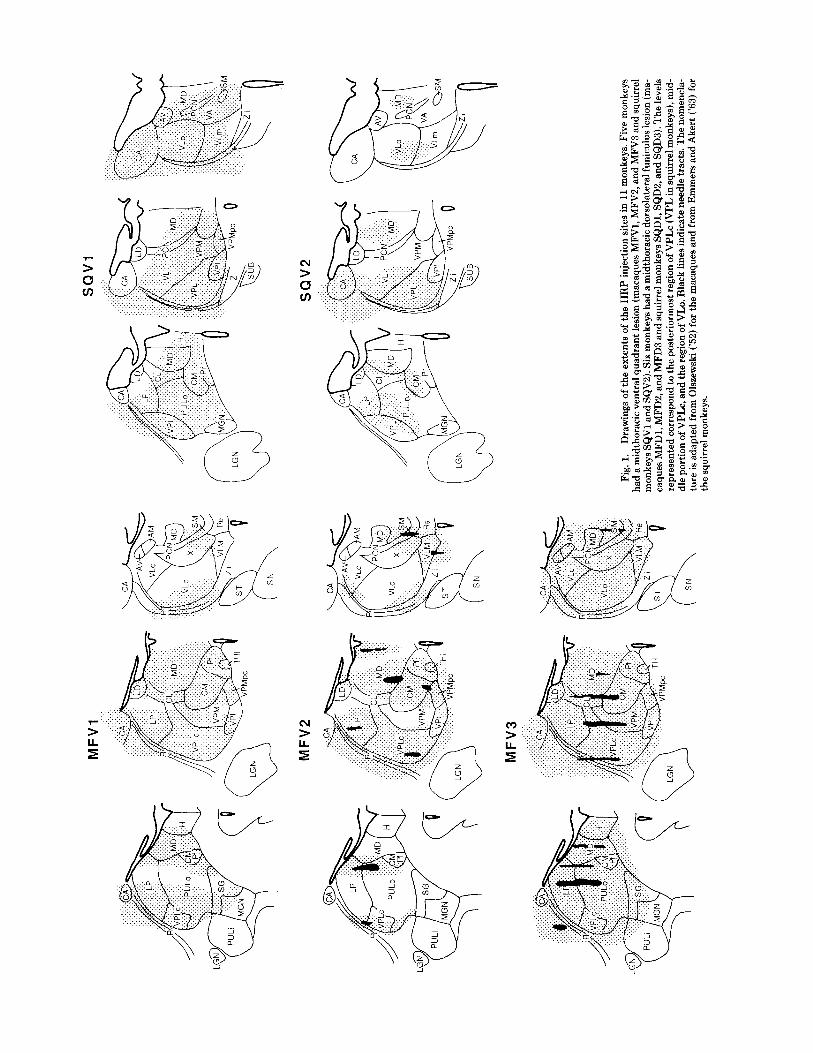
Fig.
1.
Dra
win
gs of
the
ext
ents
of
the
HR
P in
ject
ion
site
s in
11
mon
keys
. Fiv
e m
onke
ys
had
a m
idth
orac
ic v
entr
al q
uadr
ant l
esio
n (m
acaq
ues M
FV1,
MFV
P, a
nd M
FV3
and
squi
rrel
m
onke
ys S
QV
l and
SQ
VZ)
. Six
mon
keys
had
a m
idth
orac
ic d
orso
late
ral f
unic
ulus
lesi
on (m
a-
caqu
es M
FD1,
MFD
Z, a
nd M
FD3
and
squi
rrel
mon
keys
SQ
D1,
SQ
DZ,
and
SQ
D3)
. The
leve
ls
repr
esen
ted
corr
espo
nd to t
he p
oste
rior
mos
t reg
ion
of V
PLc
(VP
L in
squ
irre
l mon
keys
), m
id-
dle
port
ion
of V
PLc,
and
the
regi
on o
f VLo
. Bla
ck li
nes
indi
cate
nee
dle
trac
ts. T
he n
omen
cla-
tu
re is
ada
pted
fro
m O
lsze
wsk
i ('5
2) fo
r the
mac
aque
s an
d fr
om E
mm
ers
and
Ake
rt ('
63)
for
the
squi
rrel
mon
keys
.
L

MF
D1
S
QD
l
L
SQ
D2
L
MF
D3
S
QD
3
Figu
re 1 c
onti
nued

478 A.V. APKARIAN AND C.J. HODGE
construction of the injection sites. The HRP-processed spi- nal cord tissue was dehydrated and coverslipped without counterstaining. The gracile and cuneate nuclei were exam- ined to ascertain the adequacy of the injections and the suc- cess of the enzymatic reaction.
Spinal cord cells containing retrogradely transported HRP were examined with a microscope equipped with brightfield, darkfield, phase condensers, and polarizing fil- ters. The laminar position and number of labeled HRP neu- rons in each spinal cord section were entered into a PDP11/ 34 computer. In a few animals the diameters of the retro- gradely labeled spinal cord cells were entered into the PDP11/34 computer by using the techniques described ear- lier (Apkarian and Hodge, '89a). This data was used to gen- erate cell diameter distributions and scattergrams of cell shapes.
The spinal segments were divided into six groups and the number of labeled cells was averaged across the segments within each group: 1) upper cervical included cervical seg- ments 1-3; 2) lower cervical included cervical segments 4-8; 3) thoracic cord included a variable number of thoracic seg- ments; 4) upper lumbar included lumbar segments 1-3; 5) lower lumbar included lumbar segments 4-7; and 6) sacro- coccygeal region included all the sacral and the upper coccy- geal segments. The data collected in the thoracic spinal cord was from areas at least two to three segments removed from the lesion sites. The relation between thalamic injections, spinal cord lesions, and the laminar distribution of the STT cells was determined for the contralaterally projecting cells by subdividing the labeled population into three groups: laminae 1-111, laminae IV-VI, and laminae VII-X.
TABLE 1. HRP-Labeled Contralateral and Ipsilateral Cells per 100 pm Sections in Animals with Ventral Quadrant
and Dorsolateral Funiculus Lesions'
Cervical Lumbar Sacral &
Monkey Upper Lower Upper Lower coccygeal
Ventral quadrant lesions a. Contralateral cells/100u section
MFVl 41.90 17.03 5.03 5.00 4.33 MFV2 39.07 12.78 1.53 3.98 3.53 MFVB 42.02 14.92 5.98 7.85 6.65 SQVl 11.99 6.68 NA 1.90 0.59 SQVZ NA 2.00 NA 1.94 NA Mean 33.74 10.68 4.18 4.13 3.78
MFVI 12.20 2.75 0.62 0.55 0.75 MFVZ 14.52 2.40 0.73 3.20 3.68 MFVB 12.47 2.17 1.03 1.52 1.90 SQVl 2.33 0.16 NA 0.06 0.33 SQVZ NA 0.46 NA 0.44 NA Mean 10.38 1.59 0.79 1.15 1.66
b. Ipsilateral cells/lWu section
Dorsolateral funiculus lesions c. Contralateral cellsl100u section
MFDl 4.58' 4.12 NA 4.92 1.60 MFD2 24.703 10.00 NA 11.00 10.35 MFD3 49.62 16.83 NA 20.70 18.72
SQD2 NA NA NA 4.63 NA SQD3 25.80 10.89 2.93 17.08 7.73 Mean 27.31 9.59 2.14 10.57 8.56
MFDl 1.12 0.85 NA 0.57 0.52 MFD2 10.45 2.40 NA 1.80 3.32 MFD3 17.95 2.90 NA 3.40 3.82 SQDl 10.53 0.99 0.26 0.81 2.14 SQDZ NA NA NA 0.19 NA SQD3 6.18 0.83 0.43 1.20 1.36 Mean 9.24 1.59 0.34 1.33 2.23
SQDl 31.85 6.11 1.35 5.10 4.43
d. Ipsilateral cells/lM)u section
'NA = not available. Thoracic values not shown since lesion was placed there. 'C4 only. 3c2 only.
AM AV CA CL CM H LD LGN LP MD MGN PCN Pf PULi PlJLo R Re SG SM SN ST/SUB THI VA VL VLm/VLM VLO VPI VPL VPLC VPM VPMpc X ZI
Abbreviations
anterior medial n. anterior ventral n. caudate n. central lateral n. centre median n. habenula lateral dorsal n. lateral geniculate n. lateral posterior n. medial dorsal medial geniculate n. paracentral n. parafascicular n. inferior division of pulvinar pulvinar oralis reticular n. n. reuniens suprageniculate n. n. submedius substantia nigra subthalamic n. habenulo interpeduncular tract ventral anterior n. ventral lateral n. medial division of ventral lateral n. oral division of ventral lateral n. ventral posterior inferior ventral posterior lateral n. caudal division of ventral posterior lateral n. ventral posterior medial n. parvicellular division of the ventral posterior medial n nucleus x zona incerta
RESULTS Injection sites
Only animals with adequate injections are included in this study. In five additional monkeys inadequate injec- tions, lesions, and/or enzymatic reactions rendered the re- sults unusable. Horseradish peroxidase injections were deemed adequate only if the injections were centered in the thalamus with minimal spread into the mesencephalon. In animals with adequate injections, usually there was HRP spread into the internal capsule laterally. Often HRP also spread along the needle tract into the cortex, but there was no HRP spread into the midbrain or the hypothalamus. The results from six old world monkeys and five new world mon- keys will be presented.
The boundaries of the HRP injections are shown for all 11 monkeys in Figure 1. These injection sites were determined by using thalamic nuclear boundaries described by Ols- zewski ('52), Emmers and Akert ('63), Gergen and Maclean ('62), Szabo and Cowan ('84), Burton and Craig ('83), and Jones ('85). The nomenclature adapted for the various tha- lamic nuclei are those used by Emmers and Akert ('63) for the squirrel monkeys and those used by Olszewski ('52) for the macaque monkeys. Thalamic injections are shown for six macaques (MFV1, MFV2, MFV3, MFD1, MFD2, MFD3) and five squirrel monkeys (SQVl, SQV2, SQD1, SQD2, SQD3). Since all these animals received midthoracic spinal cord white matter lesions interrupting tracts ipsi- lateral to the thalamic injections, only the label found in the cervical segments can directly be compared with the extents of thalamic injections.

PRIMATE SPINOTHALAMIC PATHWAYS: I1
Relation between cervical STT cells and injection sites
Seven animals, four macaques (MFVl, MFV3, MFD2, MFD3), and three squirrel monkeys (SQVl, SQD1, SQD3) had injections that covered all of the somatosensory thalam- us on one side and resulted in an approximately equal num- ber of HRP labeled cells in the upper cervical and lower cervical regions (Table 1). The number of labeled cells per section in the upper cervical spinal cord was essentially identical between these animals except for SQVl, which had a smaller number of HRP-labeled cells. The number of cells per section in the lower cervical spinothalamic population appears to be larger in macaques than in squirrel monkeys. Animals in which the injections did not include portions of the somatosensory thalamus resulted in a smaller number of labeled cells in the cervical spinal cord (MFD1, SQV2).
The laminar distributions of neurons labeled from contra- lateral thalamic injections in the upper cervical and lower cervical area of the spinal cord are presented in Figure 2 for various animals. Within the upper cervical region, the con- tralateral STT cells were located in laminae deeper than lamina I (88-94% of the contralateral label; Fig. 2A, animals MFV1, MFV2, MFV3, SQV1; Fig. 2B, animals MFD2, MFD3, SQD1, SQD3). The thalamic injections in the eight animals presented were equivalent (see Fig. I), resulting in a similar distribution of label between the three laminar groupings in all the animals (Fig. 2A,B). In the lower cervi- cal area, contralateral STT label was primarily found in laminae 1-111 and laminae IV-VI. (More than 95% of the cells found in laminae 1-111 were in lamina I throughout the spinal cord.) This distribution was especially true in animals in which the thalamic injections covered all somatosensory regions (78-95% of the contralateral label was in laminae I- VI; Fig. 2C, animals MFV1, MFV3, SQV1; Fig. 2D, animals MFD2, MFD3, SQD1, SQD3).
Some of the variability of the cervical label in the remain- ing animals could be correlated with the thalamic injections. In animal SQV1, only 5% of the contralateral label in the lower cervical cord was located in the ventral horn (laminae VII-X) despite a large number of HRP-labeled STT cells found (6.68 cells/100 pm, Table la). The injection in this animal covered most of the thalamus but the ventral most portion of CL was not injected with HRP (Fig. 1). Since a number of studies have indicated that ventral CL receives STT terminations from the lumbar enlargement ventral horn cells (Willis, '87; Apkarian and Hodge, '89b). It is rea- sonable to conclude that the injection in SQVl spared the portion of CL with inputs from the laminae VII-X cells of the lower cervical region. In animal MFDl, the thalamic HRP injection spared large portions of the ventral lateral region of the lateral thalamus resulting in a decrease in the number of lamina I cells labeled in the lower cervical region (Apkarian and Hodge, '89a). Among the eight animals shown in Figure 2C and 2D, animal SQD3 had the largest relative number of lamina I labeled cells in the contralateral lower cervical region; the only difference in the thalamic injections between this animal and the other complete tha- lamic injections (animals MFV1, MFV3, SQV1, MFD2, MFD3, SQD1, SQD3) was the inclusion of zona incerta (Zi) in the injection (Fig. 1). A number of studies have demon- strated spinal projections to Zi in cat and monkey (Berkley, '80; Burton and Craig, '83; see also next paper). Hence it is possible that at least the cervical enlargement lamina I cells preferentially terminate in Zi. In animal MFV2 the injection did not include the SG and PULo region posteriorly, yet the
479
cervical distribution was similar to that of the animals in which this region was injected (animals MFVI, MFV3, MFD2, MFD3, SQVl, SQDl, SQD3).
Ventralquadrant-lesioned animals Five animals, three macaques (MFV1, MFV2, MFV3) and
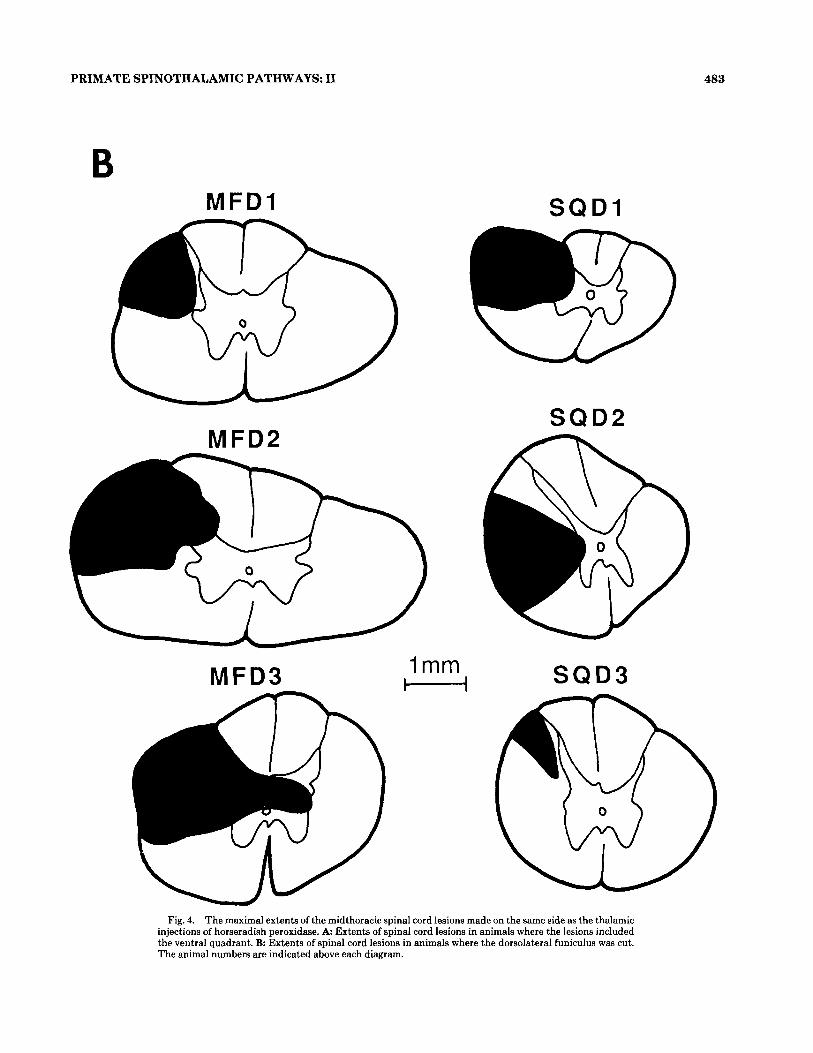
two squirrel monkeys (SQVl, SQVB), had adequate tha- lamic HRP injections (Fig. 1) combined with midthoracic ventral quadrant lesions (Figs. 3,4A) ipsilateral to the tha- lamic injections. The lesions were designed to interrupt the transport of HRP through the ventral quadrant in order to determine the cells of origin of the dorsolateral spinotha- lamic pathway. Figure 3 shows a single 60 pm section of spi- nal cord tissue in the neighborhood of the lesion in animal MFVl. The maximum extents of the lesions for this and the other VQ-lesioned animals are shown in Figure 4A. The number of HRP-labeled cells below the lesion in these ani- mals was lower than in animals with no spinal cord lesions (mean 4.1 cells/100 pm, Table la, vs. mean 10.3 cells/100 pm in unlesioned animals, Table l a Apkarian and Hodge, '89a). In the three macaques (MFV1, MFV2, MFV3), the thalamic injections were similar (Fig. 1) and the number (Table la) and distribution (Fig. 2A,C) of labeled cells in the upper cer- vical and lower cervical regions were also similar. The lesions in animals MFVl and MFV2 were similar and included all of the ventral quadrant (Figs. 3,4A). More than 80% of the label in the contralateral lower lumbar area of these two animals was found in laminae 1-111 (Fig. 5A). In animal MFV3, the midthoracic lesion spared the dorsalmost portion of the VQ (Fig. 4A) resulting in an increase in the proportion of HRP-labeled cells located in laminae IV-VI in the contralateral lower lumbar region (31% in MFV3 as compared to 12% and 10% in MFVl and MFV2, Fig. 5A). In the squirrel monkey SQVl, the thalamic injection and the label above the lesion were similar to those of the macaques (MFV1, MFV2, MFVS), but the label below the lesion was equally distributed between laminae 1-111 and IV-VI (Fig. 5A). Although the injection in animal SQV2 spared large portions of the lateral thalamus (Fig. I), the number of cells per section and pattern of label below the lesion were similar to those of the other squirrel monkey (SQVl). The label found in the contralateral lumbar ventral horn was very low in all five monkeys with VQ lesions (3-11 % of the contralat- erally projecting HRP-labeled cells were in laminae VII-X, Fig. 5A).
The segmental variation in the number of contralaterally projecting HRP-labeled cells in each laminar group is dis- played for one animal in Figure 6A (animal MFVI). Above the lesion, in the upper cervical and lower cervical regions a large number of HRP-labeled cells were evident in all three laminar groupings (Fig. 6A). However, the pattern of label below the lesion is clearly disturbed. Below the lesion, HRP label was concentrated in the marginal zone of the spinal cord throughout segments T10 to the coccygeal cord (Fig. 6A; also compare this pattern of label to Fig. 4 in this issue, Apkarian and Hodge, '89a). Similar results were obtained in three other animals.
Therefore, these results indicate the existence of a dorso- lateral spinothalamic pathway in squirrel and macaque monkeys. In the macaque, this pathway originates primarily from contralateral lamina I cells, while in the squirrel mon- key the DSTT is made up of axons of cells located in the contralateral laminae I-VI. In animal SQV2, the thoracic lesion included the dorsal columns, yet the label below the lesion was not different from animal SQVl. This implies

lb
00
0
PER
CEN
T LA
BEL
PE
RC
ENT
LAB
EL
oo
oo
oo
oo
og
gS
D
NP
~W
O
NP
I
c
w n ..
<
w
zb H ?I
j
<
Y
<
H
PERC
ENT
LABE
L PE
RCEN
T LA
BEL
Y
ru
ge
gg
N
P
m
m
En
& PE
RCEN
T LA
BEL
PERC
ENT
LABE
L N
P~
WG
DO
O
v)
I I
w
w
w
w
w c
c
w
<
w
c
Y
Y
c
I
Y
<
w
w
I
w
X
x
x
Y
H
Y
Y
Y
H
Y
<
C
<
c
c
Y
Y H
- x
<
I X
I
PERC
ENT
LABE
L ~
H
I PE
RCEN
T LA
BEL
I N
P~
~O
PE
RCEN
T LA
BEL
I PE
RCEN
T LA
BEL
I N
P~
WO
0
00
00
00
00
00
0
NP
~W
O N
P~
ID
O
00
00
00
00
00
00
<
H
w
I X
Y
<
<
w
<
Y
H
8 x
H
<
c
<
Y
w
I
x
Y
I PE
RCEN
T LA
BEL
t'
""
I N
PO
I
oo
oo
g8
I a
w
w
H
Y
w
w
<
<
H
<
w
w
I X
7 <
H
<
H
H
I
X
PER
CEN
T LA
BEL
cL
PER
CEN
T LA
BEL
I.
."
'
tL
""
oo
g~
$8
,3
gg
%W
N
Y
w
PERC
ENT
LABE
L I 0
00
N
Pg
gZ
z
11
c
W
Y
H
H
H
Y
<
<
<
w
H
X
H
H
Y
H rc
In
0 z
H
<
<
<
H
!-I
H
<
<
<
H
Y
H
H
I
I X
X
z n 8
1 PE
RC
ENT
LAB
EL
I PE
RC
ENT
LAB
EL
NP
UI
WO
~
I"
"'
1
.'
"'
0
00
00
00
~~
~%
8
Y
Y
Y
H
Y
!-I .c
ul D z:
B 8
< - w XI
Y
<
<
Y
<
H
Y
I X

PRIMATE SPINOTHALAMIC PATHWAYS: I1 48 1
Fig. 3. Photomicrograph of a midthoracic spinal cord ventral quadrant lesion in a macaque (MFV1). Tissue is stained with Luxol fast blue and cresyl violet. Calibration bar is 1 mm.
that most axons of the DSTT do not travel in the dorsal col- umns.
Animals with dorsolateral funiculus lesions Six animals, three macaques (MFD1, MFD2, MFD3) and
three squirrel monkeys (SQDl, SQDB, SQD3), had adequate thalamic injections and received midthoracic DLF lesions ipsilateral to the thalamic injections. These lesions were designed to block HRP transport through the DLF in order to demonstrate the cells of origin of the ventral spinotha- lamic pathway. In four of the animals (MFDB, MFD3, SQD1, SQD3) the thalamic injections covered most of the somatosensory thalamus (Fig. l), and the upper and lower cervical spinal cord label was similar in number (Table lc,d)
Fig. 2. The relative distribution of the contralaterally projecting spi- nothalamic cells in the cervical spinal cord, above the midthoracic lesions. The percent label found in three laminar groupings (laminae I- 111, IV-VI, and VII-X) is plotted for individual animals. A Laminar dis- tribution of the contralaterally projecting HRP-labeled cells in C2 in animals with midthoracic ventral quadrant lesions. B: Contralateral HRP label in C2 in animals with midthoracic dorsolateral funiculus lesions. C: Contralateral HRP label in the lower cervical region (C4-C8) in animals with midthoracic ventral quadrant lesions. D: Contralateral HRP label in the lower cervical region in animals with midthoracic dor- solateral funiculus lesions. Animals are indicated at the left corner of each plot.
and distribution (Fig. 2B,D) among them. In three of these four animals (MFDB, MFD3, SQD1) the midthoracic lesion was also similar, including all of the DLF (Fig. 4B). The HRP label in the contralateral lower lumbar region of these animals was found primarily in laminae IV-VI and VII-X (84-9976, Fig. 5B). In the macaques but not in the squirrel monkey there was a substantial population of lamina I cells labeled below the lesion, in the contralateral lower lumbar area. In animal SQDS, the midthoracic lesion covered only the dorsal third of the DLF (Fig. 4B), resulting in no detect- able change in the proportion of the labeled neurons below the lesion (Fig. 5B) as compared to animals with no spinal cord lesions. Therefore, a t least in the squirrel monkey, as already demonstrated in the cat (Stevens et al., 88, '89b), the DSTT axons are located primarily outside the dorsalmost portion of the DLF.
The injection in animal SQD2 did not include all of the thalamus- (Fig. 1). However, the number of labeled cells in the lower lumbar region was similar to that observed in ani- mal SQDl (Table Ic). More importantly, the thoracic lesion in SQDB was complementary to the lesion in animal SQDS since it spared the dorsalmost portion of the DLF (Fig. 4B). Although the dorsalmost portion of the DLF was spared in animal SQD2 there were very few labeled cells found in the contralateral lamina I ( 2 % , Fig. 5B), thus confirming that very few DSTT axons travel in this spared region of the DLF.
The injection in animal MFDl spared a large portion of

482
A M F V I -
A.V. APKARIAN AND C.J. HODGE
S Q V I
MFV2
SQV2
H MFV3
Figure 4
PRIMATE SPINOTHALAMIC PATHWAYS I1 483
B
W
M F D I
SQD2 MFD2
MFD3 1 mm M
S Q D I
SQD3
Fig. 4. The maximal extents of the midthoracic spinal cord lesions made on the same side as the thalamic injections of horseradish peroxidase. A: Extents of spinal cord lesions in animals where the lesions included the ventral quadrant. B Extents of spinal cord lesions in animals where the dorsolateral funiculus was cut. The animal numbers are indicated above each diagram.

484 A.V. APKARIAN AND C.J. HODGE
A
J W ‘111 M F v i _j:l MFvi ~ MFv, - 5 60 m 5 60
+ 40 6 40 6 40
a kf 20 W
n. 20 20
1-111 IV-VI VII-x 1-111 IV-VI VII-x 1-111 IV-VI VII-x 0
100
80
5 60 g 40
J W
t-
U W
a 20
0
B 100
80 A
I- g 40
20
0
U
100
80 A w m 560 +- 9 40 U W
a. 20
0
SOVl
100
80
m 5 60 _1 W
9 40 I 0
1-111 IV-VI VII-x
loo 1 loo 1 WOl
1-111 IV-VI VII-x 1-111 IV-VI VII-x
so01
_[
100
80
5 60 J
Y e 5 40 20
0
a: w
1-111 IV-VI VII-x
sov2
n
L 1-111 IV-VI VII-x
1-111 IV-VI VII-x
100
so02
I
80
3 60 J
L &-
40
B a 20
0 1-111 IV-VI VII-x
SOD3
II 1-111 IV-r VII-x
Fig. 5. The relative distribution of the contralaterally projecting spi- nothalamic cells in the lower lumbar region of the spinal cord, below the midthoracic lesions. The percent label found in three laminar groupings (laminae 1-111, IV-VI, and VII-X) is plotted for individual animals. A
Distributions in animals with midthoracic ventral quadrant lesions. B Distributions in animals with midthoracic dorsolateral funiculus lesions. Animals are indicated at the left corner of each plot.
the thalamus ventrally (Fig. 1) and resulted in a small num- ber of labeled cells throughout the spinal cord (Table lc). Moreover, the percentage of HRP-labeled lamina I cells in the contralateral lower cervical region was the lowest among the six animals (Fig. 2D). Below the lesion very few cells were found in lamina I (276, Fig. 5B), indicating that the lamina I projections to thalamic areas other than the most ventral portion of VPLc in the macaque travel primarily in the DLF.
The segment to segment variation in the number of con- tralaterally projecting HRP-labeled cells in the three lami- nar groups (1-111, IV-VI, VII-X) is displayed for a macaque with a DLF lesion in Figure 6B (animal MFDZ). Above the
lesion, in the lower cervical region, a large number of HRP- labeled cells were evident in all laminar groups (Fig. 6B). Below the lesion the label was concentrated in the deeper laminae although there were a substantial number of labeled cells (11 74 ) found in laminae 1-111.
Therefore, the cells of origin of the ventral spinothalamic pathway are primarily found in the deeper laminae of the contralateral spinal cord, below the marginal zone. In the macaque, some lamina I cells project through the VSTT. The VSTT in the squirrel monkey is almost exclusively made up of axons of cells located outside of lamina I.
In a few animals with VQ lesions and in some animals with DLF lesions cell diameter distributions and cell shape

PRIMATE SPINOTHALAMIC PATHWAYS: I1 485
Normalization of the mean lower lumbar HRP-labeled cells in the unlesioned animals relative to the VQ-lesioned animals indicated that on the average 23 % of the total lower lumbar STT cells remained labeled following VQ lesions (4.13 of 18 cells/100 pm section). This value is an estimate of the relative size of the DSTT. However, the relative size of the DSTT in macaques (26-51% of the total contralateral STT) seems larger than that found for squirrel monkeys (15-16% of the total contralateral STT; compare the num- ber of cells labeled in the lower lumbar region of animals MFV1-MFV3 to animals SQVl and SQV2 in Table la). A similar procedure in animals with DLF lesions indicated that on the average 65% of the total lower lumbar STT cells remained labeled with HRP following DLF lesions (10.57 of 16.24 cells/100 pm section), which is an estimate of the rela- tive size of the VSTT.
Essentially all lamina I STT cells in squirrel monkeys travel in the DSTT since less than 1% were found in the VSTT. In the macaque, however, it was estimated that 78% of the contralateral lower lumbar lamina I STT cells travel in the DSTT while 22% travel in the VSTT. These esti- mates were calculated by comparing the size of the lower lumbar HRP-labeled lamina I cells in unlesioned animals (previous paper) to the size found in animals with VQ or DLF lesions.
Locations of the spinothdamic axons The spinal cord white matter of all the animals used in
this and the previous study were carefully examined, espe- cially the dorsolateral funiculus ipsilateral to the thalamic injections, in order to localize the spinothalamic axons. In a number of animals a large number of HRP-labeled axons were found in the spinal cord white matter throughout the extent of the spinal cord ipsilaterally. Figure 8 shows camera lucida drawings of HRP-labeled axons in various segments of the spinal cord in a macaque (C2, C5, and L1, Fig. 8). This macaque did not receive a spinal cord lesion. The number of axons in the white matter increased at more rostra1 seg- ments. Very few axons were observed at segments below L4. No HRP-labeled axons were observed in the ipsilateral DLF in 15 monkeys examined (Fig. 8). The lesion studies, how- ever, clearly indicate the existence of the DSTT. It is con- cluded, therefore, that HRP label in the small DSTT axons can not be seen with light microscopy.
scattergrams were generated for HRP-labeled cells located above and below the midthoracic lesions. In these animals, the sizes and shapes of the STT cells in both the cervical and lumbar areas were similar to those described for animals with no spinal cord lesion (see preceding paper).
Comparison between unlesioned animals and animals with VQ and DLF lesions
A large portion of this paper as well as the preceding one (this issue), is dedicated to an analysis of the relationship between the location of the thalamic injections and their relationship with the different populations of neurons la- beled in the spinal cord. In order to demonstrate the exis- tence of the DSTT independent of the variations of the thalamic injections, the percentages of the contralaterally projecting HRP-labeled laminae 1-111 neurons in segment C2, lower cervical region, and lower lumbar region were averaged across the three animal groups presented here and in Apkarian and Hodge ('89a). The percent contralateral label in laminae 1-111 in Figure 3A,B, and D in the previous paper were averaged and assumed to represent the control label in laminae 1-111 in the three spinal cord regions. Simi- larly, the percent contralateral label in the animals with VQ or DLF lesions presented here in Figures 2 and 5 were aver- aged. Animal SQD3 was excluded because of the small size of the DLF lesion. The average and 95th percentile interval of the percent of contralaterally projecting cells located in laminae 1-111 in these groups and spinal regions are shown in Figure 7. The data shows a lack of difference in the amount of label found in C2 or the lower cervical region between the three groups of animals, suggesting that the thalamic HRP injections and subsequent transport to the cervical segments of the spinal cord were equivalent be- tween the three groups of animals. Figure 7 also demon- strates a substantial decrease in the percentage of label found in the lower lumbar region laminae 1-111 in animals with DLF lesions. On the other hand, Figure 7 shows a prominent increase in the percentage of label located in laminae 1-111 in the lower lumbar region of animals with VQ lesions compared to the control group. These results strongly suggest that the differences seen in the contralat- era1 lumbar lamina I cell label are due to the placement of the lesions in the thoracic spinal cord white matter.
A similar type of analysis was performed for the labeled lamina I cells in the spinal cord ipsilateral to the thalamic injections. However, the results of this comparison were inconclusive since there was a large amount of variability in the data (mean and standard deviation of the percent lam- ina I cells in the ipsilateral lower lumbar region was: 4 * 4.4 in animals with no spinal lesions, 9.2 * 7.9 in animals with DLF lesions, and 6.5 f 6.2 in animals with VQ lesions). This large variability was mainly due to the paucity of labeled ipsilaterally projecting cells.
Relative sizes of the DSTT and the VSTT The mean numbers of labeled cells per section in the ani-
mals with VQ lesions (Table la,b) and in the animals with DLF lesions (Table lc,d) at regions above the thoracic lesions do not agree with the mean label in comparable areas in animals with no spinal lesions (Table la,b Apkarian and Hodge, '89a). This renders direct comparisons between the animal groups regarding the size of the label found below the lesions unacceptable. However, if the mean label found in the lower lumbar region is normalized relative to the mean label found in the lower cervical region, size compari- sons can be made.
DISCUSSION The results indicate that the STT extends dorsally into
the DLF beyond its classical confines in both new and old world monkeys. We have chosen to label this STT extension the DSTT because of the unique location of its axons. In new world monkeys most lamina I cells project to the tha- lamus through the DLF, and a significant portion of cells in laminae IV-VI also project to the thalamus through the DLF. Therefore, in new world monkeys, the DSTT is made of locally crossing axons of laminae I-VI cells while the VSTT is made of locally crossing axons of cells located in laminae IV-X. On the other hand, in old world monkeys, although the majority of lamina I cell axons project rostrally to the thalamus through the DLF, a significant portion of STT lamina I cells project through the VQ and only a small portion of laminae IV-VI STT cells have axons in the DLF. Hence, in the old world monkeys, the DSTT is made of locally crossing axons of lamina I cells while the VSTT is made of locally crossing axons of laminae I-X cells. As pre- viously shown for the cat (Jones et al., '87), the primate spi-

486 A.V. APKARIAN AND C.J. HODGE
A
Z 1 4 O N H
t-0 ul- W 11114 \ 111111 J H J w w 013
Z * < a:
E
Z Q O N H
I-0 O F W 11114 \ 111111 J H J w w 0(3
Z * < a:
I P S 1
I P S 1
+ + +++- +
a" d.&&LL
T r-
++++++i-l-+ +
C O N T R A
1 1
L A M I N A
1-3
4-6 : .
7
ct-tn
-10
C O N T R A
+ + +-I-+- +
L A M I N A
1-3
4-6
I
7-10
+++++++I+-
Figure 6

PRIMATE SPINOTHALAMIC PATHWAYS: I1 487
100
4 75 al P m +I
2 50 4
!2 u b? 25
0
C2 Segment
A - I w E b L Control DLFles VQ les
75 I Lower Lumbar or
Control DLF les VQ les
Fig. 7. Percent of the contralateral label found in lamina I averaged across animals with no spinal cord lesions (control; data from the previ- ous paper), animals with dorsolateral funiculus lesions (DLF les), and animals with ventral quadrant lesions (VQ les). The data are shown for
nothalamic axons do not seem to travel in the dorsal col- umns.
Technical considerations The extent of contamination of the spinal cord HRP label
with transynaptic or transneuronal transported HRP can- not be determined. However, it is thought that this type of label does not significantly contribute to the overall spinal cord label (see Jones et al., '87; Apkarian and Hodge, '89a). A more important technical issue is the variation of the label found in the spinal cord among animals as a function of dif- ferences in the injection sites. These differences are contam- inated with two factors. One factor is the inherent variation in the organization of the STT among animals. The second is the variation in the precise position of the injection nee- dles, which might lead to differential labeling of terminals of STT cells in the immediate vicinity of the needle tip. Both of these factors contribute, to some unknown extent, to the
Fig. 6. Variation in the density of label of spinothalamic cells in vari- ous segments of the spinal cord in a macaque (MFV1) with ventral quad- rant lesion (A) and in a macaque (MFDS) with a dorsolateral funiculus lesion (B). Each bar represents the mean number of labeled cells per sec- tion in a single segment, for laminae 1-111, IV-VI, and VII-X, and for ipsilaterally projecting (IPSI) and contralaterally projecting (CONTRA) spinothalamic cells. The scale indicated on the left applies for all bar graphs. The standard deviation for each bar is indicated. The mean and standard deviation of the label in C1 laminae 4 4 are shown numerically for scaling reasons. The arrows indicate the spinal segment where the lesion was placed. Pluses indicate segments where data were not collected. UC is upper cervical segments CI-C4. LC is lower cervical segments CSC8. TH is thoracic spinal cord. UL is upper lumbar seg- ments Ll-L3. LL is lower lumbar segments L4-L7. SX is sacral and coc- cygeal segments.
100
* 75 al P (d c;l
2 50 4
V b-? 25
0
Lower Cervical
T I T
Control DLF les VQ les
1
three regions of the spinal cord: C2, lower cervical region, and lower lumbar region. Error bars indicate the 95 percentile confidence inter- vals.
variability of the label found in the spinal cord. Hence, cor- relations between injections and spinal cord label must be considered tentative until experiments with more limited thalamic injections are performed (e.g., see Craig and Bur- ton, '81, for limited HRP injections in SM). Given these pre- cautions the evidence for a preferential lamina I projection to Zi should be regarded as preliminary. On the other hand, the evidence presented for a lamina I projection to the ven- tral border region of the lateral thalamus is corroborated with observations from Apkarian and Hodge ('89a) and ear- lier studies (Willis et al., '79).
Since there was no difference in the average lamina I label in the upper and lower cervical regions between unlesioned, DLF-lesioned, and VQ-lesioned animals, it is unlikely that there was any systematic variation in the degree of HRP uptake, labeling, or tissue processing between these groups. Therefore, the differences in label found in the lower lumbar region are assumed to be primarily a reflection of the mid- thoracic spinal cord white matter lesions.
Species differences in the organization of DS'IT and VS'IT
The DSTT was estimated to be 23% of the total lower lumbar region population of STT cells, while the VSTT was estimated to be 65% of the total lumbar STT in primates. This estimate closely agrees with the relative size of the DSTT in the cat, which was calculated to constitute one fourth of the total spinothalamic pathway (Jones et al., '87). However, unlike the DSTT axons of the cat which have been repeatedly observed labeled with HRP following thalamic HRP injections (Jones et al., '85; Stevens et al., '88), the primate DSTT axons could not be visualized. Therefore,

488 A.V. APKARIAN AND C.J. HODGE
Figure 8

PRIMATE SPINOTHALAMIC PATHWAYS: I1 489
the exact location of DSTT axons is still unknown in pri- mates although it is clear that they are not clustered in the dorsalmost portion of the DLF. Direct visualization of the DSTT axons would permit the assertion that this pathway is not simply a dorsal extension of the VSTT. The STT axons in the cat are distributed into two clusters, one located in the DLF and the other in the VQ (Jones et al., '85; Stevens et al., '88), thus implying that in the cat the DSTT and the VSTT are two separate pathways. Inability to visualize DSTT axons in primates suggests that some cat DSTT axons are significantly larger than primate DSTT axons, and that, in primates, the DSTT axons are smaller than the VSTT axons.
The cells of origin of the DSTT and the VSTT in macaque and squirrel monkeys differ from each other as well as from the organization of the DSTT and the VSTT in cats. In the cat, the lumbar DSTT is made of locally decus- sating axons of cells located primarily within lamina I with a small contribution of deeper STT cells. The cat lumbar VSTT is composed of locally decussating axons of cells located in laminae IV-X with almost no lamina I cell contri- bution (Jones et al., '85, '87). The existence of a DSTT made up of locally crossing axons of lamina I cells has recently been confirmed in another laboratory in the rat (Nahin et al., '87). Hence, two spinothalamic pathways have been demonstrated in every mammalian species examined. I t is reasonable, therefore, to assume that man also has two spi- nothalamic pathways, a DSTT and a VSTT. In 1949, Kuru described two distinct ascending pathways in man, both of which reached the thalamus. Correlating the ascending de- generating fibers and the chromatolytic cells below the spi- nal cordotomies with ensuing deficits, Kuru concluded that the cells of origin of the dorsolateral spinothalamic pathway were found in the contralateral lamina I and transmitted information related to pain and temperature perception. A number of neurosurgeons, based on the extent of hypalgesia following different-sized cordotomies, have concluded that the human STT has two divisions, termed the lateral and the ventral STT (Foerster and Gagel, '32; White, '54; Kerr, '75a,b). In this scheme, the more dorsal and lateral tract was thought to conduct pain and thermal information while the more ventral tract was thought to convey information regarding touch and pressure. Single-unit recordings in monkey VQ have failed to demonstrate a segregation of STT axons carrying low-threshold information from those conveying high-threshold information (Applebaum et al., '75). It is possible that the human lesion studies included the DSTT since the lesions commonly extended dorsal to the dentate ligament. The DLF, however, was not studied in the primate physiologic experiments. White and Sweet ('69) showed in a large number of patients that when cordotomies were extended further ventrally there was better pain relief. This observation together with the failure to physiologically demonstrate a segregation of the STT axons in monkeys has resulted in the belief that there is a single spinothalamic tract the axons of which, located in the VQ, are scattered throughout the white matter.
Fig. 8. Camera lucida drawings of HRP labeled axons in the spinal cord white matter ipsilateral to the thalamic injections in one macaque. Each drawing is from a single 60 pm section taken from the segment indicated (C2, C6,and Ll).
Physiologic differences between the spinothalamic pathways
The differences in the cells of origin of the DSTT and the VSTT in macaque from those of the cat parallel differences in response properties of the STT cells observed in these two species. In an exhaustive study of the response proper- ties of 218 lamina I STT cells in the cat, Craig and KnifFki ('85) determined that lamina I cells with wide dynamic range type responses did not reach the thalamus and that those lamina I cells that did terminate in the thalamus responded either to nociceptive-specific stimuli or to innoc- uous and noxious thermal stimuli (innocuous cooling and noxious heating of the receptive field) or to both types of inputs convergently (mean conduction velocity 3.7 m/sec- ond). Given that the cat DSTT is composed primarily of lamina I cell axons it must be concluded that the DSTT sig- nals primarily nociceptive-specific and thermal information to the thalamus. The majority of cat laminae IV-VI STT cells in the cervical enlargement respond in a graded fash- ion, in relatively small receptive fields, to innocuous and noxious inputs (58% of 56 cells studied, Ferrington et al., '86). Only 13% of this population of STT cells had nocicep- tive-specific responses (mean conduction velocity 38 m/sec- ond). The response properties of cat STT cells in laminae VII and VIII have been investigated in the lumbar enlarge- ment and show striking differences in response properties from STT cells in laminae I-VI (Meyers and Snow, '82). The receptive fields of these ventral horn cells were complex and almost always bilateral, with a large number of the cells hav- ing excitatory and inhibitory fields and with most having convergent responses to both muscle and cutaneous inputs. These cells were antidromically activated from medial and lateral thalamic stimulation sites. Of the 38 ventral horn STT cells (mean conduction velocity 42.2 mhecond; Tre- vino et al., '72, report a mean conduction velocity of 33 m/ second for cat ventral horn STT cells), only three responded exclusively to noxious stimuli, although 46 % of the cells had nociceptive responses and 78% of the cells responded to innocuous stimuli. In contrast to the cat lamina I STT cells, none of the units in this population (laminae VII-X STT cells) responded to innocuous thermal stimuli. It is conceiv- able that the small number of nociceptive-specific STT cells in the deeper laminae project, together with the lamina I cell axons, in the DLF and thus are part of the DSTT. Experi- ments in cats in our laboratory corroborate this hypothesis since a statistically significantly larger number of nocicep- tive-specific thalamic cells were found to receive inputs from the ipsilateral DLF vs. the VQ (Martin et al., '89). The large difference between the conduction velocity of the lam- ina I STT cells is also in agreement with the idea that the axons of these neurons travel in the DLF. It is thought that the conduction velocity of nociceptive-specific STT cells in the deeper laminae are lower than the rest of the STT cells found in these laminae, although data on this subject is lack- ing in the cat.
In contrast to the cat, lamina I STT cells with graded responses to low-threshold and high-threshold inputs have been repeatedly described in the macaque (Giesler et al., '81; Ferrington et al., '86; Yezierski et al., '87; Willis, '87). Most, if not all, of these wide dynamic range type lamina I STT neurons have been antidromically activated from the lateral thalamus. Ferrington et al. ('87) showed that nine of 21 monkey lamina I STT cells were nociceptive specific (mean conduction velocity of 14 m/second) while the rest had wide

490
dynamic responses (mean conduction velocity of 33 m/sec- ond). Some of these units had responses to innocuous cool- ing and/or noxious heating. In the rat more than 90% of 131 lamina 1 cells with axons in the DLF had nociceptive responses only (McMahon and Wall, '88). Recently Willis ('87) summarized results of experiments performed in his laboratory over a large number of years characterizing pri- mate STT cells. Of a total of 318 STT cells projecting to VPL 29 were classified as low threshold, and all but one of these were in laminae IV-VI (10% of laminae IV-VI STT cells). The remainder of the STT cells were grouped into nociceptive-specific or wide dynamic range type. Sixty per- cent of the STT cells in laminae IV-VI were wide dynamic range type. Lamina I cell responses were equally divided between nociceptive-specific and wide dynamic range type. Giesler et al. ('81) identified 21 cells activated from CL; these were located in laminae VI-VIII. Most had large, com- plex, at times bilateral receptive fields; 30% were classified as wide dynamic range type; and the remainder were noci- ceptive-specific units. The mean conduction velocity of the cells projecting to the medial thalamus was 18.6 mhecond, while the conduction velocity of cells projecting to the lat- eral thalamus (cells located in laminae IV-VI and one lam- ina I cell) was 37.1 mhecond.
Accepting the hypothesis that cat DSTT is primarily made up of cells with nociceptive-specific inputs and cat VSTT is made up of cells with graded responses to both nox- ious and innocuous inputs as well as cells with convergent cutaneous and muscle inputs, similar predictions could be made about the response properties of the primate STT pathways. The primate ventral horn cells, like in the cat, have large complex receptive fields (it is not clear if these cells have deep muscle inputs although Foreman et al., '77, '79, have demonstrated muscle afferents on STT cells in pri- mates) and project through the VSTT to the medial tha- lamus. The primate laminae IV-VI cells have small, circum- scribed receptive fields, have wide dynamic range type responses, and primarily project through the VSTT to the lateral thalamus, similar to those of the cat (Willis et al., '74). The small number of nociceptive-specific laminae 1V- VI cells could ascend through the DSTT. The major physio- logic difference between cat and monkey seems to be that a large number of lamina I STT cells in the macaque have wide dynamic range responses to noxious inputs. It is likely that these cells are the lamina I cells that project through the VSTT in the macaque, since the conduction velocity of lamina I cells with axons in the VQ has been reported to be faster (27 m/second) than the mean conduction velocity of lamina I cells with axons in the DLF or the VQ (11 m/ second) in the rat (McMahon and Wall, '83). The segrega- tion of the DSTT and VSTT into physiologically distinct groups remains to be ascertained. Knowledge of the exact termination sites of the DSTT and the VSTT in the tha- lamus would further clarify the various aspects of nocicep- tion that these pathways participate in (see next paper).
Recent experiments in the rat by McMahon and Wall ('88) point to physiologic dissociation in the responses of lamina I cells with axons in the DLF (DSTT is a subset of this population) from cells in laminae IV-VI. A large pro- portion of lamina I cells with axons in the DLF were facili- tated through descending pathways located in the DLF, while cells in the deeper laminae were inhibited. Moreover, stimulation of ascending pathways in the DLF did not affect the excitability of lamina I cells but resulted in inhibiting cells in the deeper laminae. The authors suggest that lamina
A.V. APKARIAN AND C.J. HODGE
I cells projecting through the DLF activate brainstem cir- cuits which in turn inhibit cells in the deeper laminae of the spinal cord. Independent of the details of the circuitry involved, it is clear that the cells subserving the DSTT have distinct connectivity and descending control from those of the VSTT cells.
The terminations of the ascending dorsolateral pathway include the mesencephalon, the dorsolateral pons, and the pontomedullary reticular formation (McMahon and Wall, '85; Hylden et al., '86; Yezierski, '88; personal observations) as well as the thalamus. These areas are associated with descending modulation (Fields and Anderson, '78; Basbaum and Fields, '79; Holstege and Kuypers, '82; Hodge et al., '83a), multisynaptic pathways projecting to the thalamus (Collins et al., '66; Noordenbos, '66), and visceral sensory nuclei with projections to the thalamus (Jordan and Spyer, '86; Cechetto and Saper, '87). The input to the dorsolateral pons seems to be primarily from the dorsolateral lamina I tract (Hylden et al., '86). However, the differences in the extent of input supplied from the dorsolateral pathway in comparison to the ventral pathway to other areas of the brainstem needs to be investigated.
Functional considerations in relation to human cordotomies
The existence of a DSTT in primates raises interesting questions and possibilities in terms of clinical nociception. On the most superficial level, these results provide evidence for yet another ascending alternate pain pathway that may in part be responsible for the return of pain following an ini- tial successful ventrolateral cordotomy. If the DSTT is assumed to be left intact following an anterolateral cordoto- my, then the reason why a cordotomy results in pain relief at all warrants an explanation. One explanation is that the lamina I input to the thalamus (and brainstem) is not suffi- cient for normal pain perception, implying that cells with wide dynamic range type responses are essential for noci- ception, an idea supported by the work of Mayer et al. ('75). Another possible explanation is that a ventral spinal lesion interrupts descending modulatory pathways (e.g., the de- scending noradrenergic system; Stevens et al., '85) as well as ascending pathways, resulting in disruption of spinal pro- cessing. A corollary to the latter explanation would be that long-term changes in the efficacy of the descending modula- tory systems may result in the resumption of the transmis- sion of nociception. Chronic deafferentation, for example, results in a reversal of the effects of descending modulatory circuits from inhibition to excitation (Hodge et al., '83b). These theories need to be tested.
ACKNOWLEDGRlENTS We thank Richard T. Stevens for contributing to the com-
pletion of this manuscript. We also thank Norma J. Horton for excellent technical assistance. This work was supported by NINCDS grant number NS228910, by the Perkins foun- dation, and by a gift from Bristol Laboratories.
LIlXRATURE CITED Ammons, W., M. Girardot, and R. Foreman (1985) T2-T5 spinothalamic neu-
rons projecting to the medial thalamus with viecerosomatic input. J. Neu- rophysiol. 5433-89.
Ammons, W.S. (1987) Characteristics of apinoreticulss and spinothalamic neurons with renal input. J. Neurophysiol. 58:480488.

PRIMATE SPINOTHALAMIC PATHWAYS: I1 491
Apkarian, A.V., and C.J. Hodge (1989a) Primate spinothalamic pathways. I. A quantitative study of the cells of origin of the spinothalamic pathway. J. Comp. Neurol. 288t447-473.
Apkarian, A.V., and C.J. Hodge (1989b) Primate spinothalamic pathways: 111. Thalamic terminations of the dorsolateral and ventral spinothalamic pathways. J. Comp. Neurol. 288t493-511.
Apkarian, A.V., R.T. Stevens, and C.J. Hodge (1985) Funicular location of ascending axons of lamina I cells in the cat spinal cord. Brain Res. 334:160-164.
Applebaum, A., R. Leonard, D. Kenshalo, R. Martin, and W. Willis (1979) Nuclei in which functionally identified spinothalamic tract neurons ter- minate. J. Comp. Neurol. 188t575-585.
Applebaum, A.E., J.E. Beall, R.D. Foreman, and W.D. Willis (1975) Organi- zation and receptive fields of primate spinothalamic tract neurons. J. Neurophysiol. 38:572-586.
Basbaum, A.I., and H.L. Fields (1979) The origin of descending pathways in the dorsolateral funiculus of the spinal cord of the cat and rat: Further studies on the anatomy of pain modulation. J. Comp. Neurol. 187:513- 532.
Berkley, K. (1983) Spatial relationships between the terminations of somatic sensory motor pathways in the rostral brainstem of cats and monkeys. 11. Cerebellar projections compared with those of the ascending somatic sen- sory pathways in the lateral diencephalon. J. Comp. Neurol. 22Ot229- 251.
Berkley, K.J. (1980) Spatial relationships between the terminations of somatic sensory and motor pathways in the rostral brainstem in cats and monkeys. I. Ascending somatic sensory inputs to lateral diencephalon. J. Comp. Neurol. 193t283-317.
Blair, R. (1985) Noxious cardiac input onto neurons in medullary reticular formation. Brain Res. 326t335-346.
Boivie, J. (1971) The termination of the spinothalamic tract in the cat. An experimental study with silver impregnation methods. Exp. Brain Res. 12331-353.
Boivie, J. (1979) An anatomical reinvestigation of the termination of the spi- nothalamic tract in the monkey. J. Comp. Neurol. 186:343-370.
Bowsher, D. (1957) Termination of the central pain pathway in man. The conscious appreciation of pain. Brain 80:606-622.
Burton, H., and A.D. Craig (1983) Spinothalamic projections in cat, raccoon and monkey: A study based on anterograde transport of horseradish per- oxidase. In G. Macchi, A. Rustioni, and R. Sprehco (eds): Somatosen- sory Integration in the Thalamus. Amsterdam: Elsevier Science Publish- ers, pp. 1741.
Carstens, E., and D.L. Trevino (1978) Laminar origins of spinothalamic pro- jections in the cat as determined by the retrograde transport of horserad- ish peroxidase. J. Comp. Neurol. 182t151-166.
Casey, K.L., and T.J. Morrow (1988) Supraspinal nocifensive responses of cats: Spinal cord pathways, monoamines, and modulation. J. Comp. Neu- rol. 270:591405.
Cechetto, D.F., and C.B. Saper (1987) Evidence for a viscerotopic sensory representation in the cortex and thalamus in the rat. J. Comp. Neurol. 2622745.
Chung, J.M., D.R. Kenshalo, K.D. Gerhart, and W.D. Willis (1979) Excitation of primate spinothalamic neurons by cutaneous C-fiber volleys. J. Neuro- physiol. 42:135P1369.
Chung, J.M., K.H. Lee, D.J. Surmeier, L.S. Sorkin, and W.D. Willis (1986) Response characteristics of neurons in the ventral posterior lateral nucleus of the monkey thalamus. J. Neurophysiol. 56t370-390.
Collins, W.F., F.E. Nulsen, and C.N. Shealy (1966) Electrophysiological stud- ies of peripheral and central pathways conducting pain. In R.S. Knighton and P.R. Dumke (eds): Pain. Boston: Little Brown, pp. 33-45.
Craig, A., and H. Burton (1985) The distribution and topographical organiza- tion in the thalamus of anterogradely-transported horseradish peroxidase after spinal injections in the cat and the raccoon. Exp. Brain Res. 58227- 254.
Craig, A., and K. Knift'ki (1985) Spinothalamic lumbosacral lamina 1 cells responsive to skin and muscle stimulation in the cat. J. Physiol. (Lond.) 365t197-221.
Craig, A.D., and H. Burton (1981) Spinal and medullary lamina I projection to nucleus submedius in medial thalamus: A possible pain center. J. Neu- rophysiol. 45t443-466.
Craig, A.D. (1987) Medial thalamus and nociception: The nucleus submedius. In J.M. Besson, G. Guilbaud, and M. Peschanski (eds): Thalamus and Pain. Amsterdam: Excerpta Medica, pp. 227-243.
Emmers, R., and K. Akert (1963) A Stereotaxic Atlas of the Brain of the Squirrel Monkey (Saimiri Sciureus). Madison: U Wisconsin Press.
Ferrington, D.G., L.S. Sorkin, and W.D. Willis (1986) Responses of spinothal-
amic tract cells in the cat cervical spinal cord to innocuous and noxious stimuli. Somatosens. Res. 3t339-358.
Ferrington, D.G., L.S. Sorkin, and W.D. Willis (1987) Responses of spinotha- lamic tract cells in the superficial dorsal horn of the primate lumbar spi- nal cord. J. Physiol. (Lond.) 388:681-703.
Fields, H.L., and S.D. Anderson (1978) Evidence that raphe-spinal neurons mediate opiate and midbrain stimulation-produced analgesia. Pain 5t333-349.
Foerster, O., and 0. Gagel (1932) Die vorderseitenstrangdurgschneidung beim menschan. Eine Klinisch-patho-physiologisch-anatomische studie. 2. Ges. Neurol. Psychiatr. 138:l-92.
Foreman, R.D., R.F. Schmidt, and W.D. Willis (1977) Convergence of muscle and cutaneous input onto primate spinothalamic tract neurons. Brain Res. 124:555-560.
Foreman, R.D., R.F. Schmidt, and W.D. Willis (1979) Effects of mechanical and chemical stimulation of fine muscle afferents upon primate spino- thalamic tract cells. J. Physiol. (Lond.) 286215-231.
Fox, R.E., J.A. Holloway, A. Iggo, and S.S. Mokha (1980) Spinothalamic neu- rones in the cat: Some electrophysiological observations. Brain Res. 182t186-190.
Gergen, J.A., and P.D. MacLean (1962) A Stereotaxic Atlas of the Squirrel Monkey's Brain (Saimiri sciureus). Bethesda: US Dept. HEW.
Giesler, G.J., D. MenBtrey, G. Guilbaud, and J.M. Besson (1976) Lumbar cord neurons at the origin of the spinothalamic tract in the rat. Brain Res. 118t320-324.
Giesler, G.J., R.P. Yezierski, K.D. Gerhart, and W.D. Willis (1981) Spinotha- lamic tract neurons that project to medial and/or lateral thalamic nuclei: Evidence for a physiologically novel population of spinal cord neurons. J. Neurophysiol. 46t1285-1308.
Greenspan, J.D., C.J. Vierck, L.A. Ritz, and R.H. Cohen (1987) Long term changes in purposive and reflexive responses to painful stimulation in monkeys following anterolateral chordotomy. Pain 4rS402.
Greenspan, J.D., C.J. Vierck, and L.A. Ritz (1986) Sensitivity to painful and nonpainful electrocutaneous stimuli in monkeys: Effects of anterolateral chordotomy. J. Neurosci. 6~380-390.
Guilbaud, G., D. Caille, J.M. Besson, and G. Benelli (1977) Single units activ- ities in ventral posterior and posterior group thalamic nuclei during noci- ceptive and non nociceptive stimulations in the cat. Arch. Ital. Biol.
Guilbaud, G., M. Peschanski, M. Gautron, and D. Binder (1980) Neurones responding to noxious stimulation in VB complex and caudal adjacent regions in the thalamus of the rat. Pain 8r303-318.
Hodge, C.J., A.V. Apkarian, S. Martini, and R.J. Martin (1987) Lateral tha- lamic nociception: The effects of interruption of transmission through the ventrolateral and the dorsolateral spinothalamic tracts. In L.M. Pubols and B.J. Sessle (eds): Effects of Injury on Trigeminal and Spinal Somato- sensory Systems. New York Alan R. Liss, Inc., pp. 313-320.
Hodge, C.J., A.V. Apkarian, R.T. Stevens, G.D. Vogelsang, 0. Brown, and J.I. Franck (1983a) Dorsolateral pontine inhibition of dorsal horn cell responses to cutaneous stimulation: Lack of dependence on catechol- aminergic systems in cat. J. Neurophysiol. 50:1220-1235.
Hodge, C.J., A.V. Apkarian, M.P. Owen, and B.S. Hanson (1983b) Changes in the effects of stimulation of locus coeruleus and nucleus raphe magnus following dorsal rhizotomy. Brain Res. 288:325-329.
Holstege, G., and H.G.J.M. Kuypers (1982) The anatomy of brain stem path- ways to the spinal cord in cat. A labeled amino acid tracing study. In H.G.J.M. Kuypers and G.F. Martin (eds): Progress in Brain Research. New York Elsevier, pp. 145-175.
Honda, C.N., S. Mense, and E.R. Per1 (1983) Neurons in ventrobasal region of cat thalamus selectively responsive to noxious mechanical stimulation. J. Neurophysiol. 49t662473.
Hylden, J.L.K., H. Hayashi, and G.J. Bennett (1986) Lamina I spinomesen- cephalic neurons in the cat ascend via the dorsolateral funiculi. Somato- sens. Res. 43-41 .
I m a 5 6 .
Jones, E.G. (1985) The Thalamus. New York Plenum Press. Jones, M.W., A.V. Apkarian, R.T. Stevens, and C.J. Hodge (1987) The spino-
thalamic tract: An examination of the cells of origin of the dorsolateral and ventral spinothalamic pathways in cats. J. Comp. Neurol. 260,349- 361.
Jones, M.W., C.J. Hodge, A.V. Apkarian, and R.T. Stevens (1985) A dorso- lateral spinothalamic pathway in cat. Brain Res. 335:18%193.
Jordan, D., and K.M. Spyer (1986) Brainstem integration of cardiovascular and pulmonary afferent activity. Prog. Brain Res. 67:295-314.
Kennard, M.A. (1954) The course of ascending fibers in spinal cord of the cat essential to the recognition of painful stimuli. J. Comp. Neurol. 100:511- 524.

492 A.V. APKARIAN AND C.J. HODGE
Kenshalo, D.R., G.J. Giesler, R.B. Leonard, and W.D. Willis (1980) Responses of neurons in primate ventral posterior lateral nucleus to nox- ious stimuli. J . Neurophysiol. 43:15941614.
Kerr, F.W.L. (1975a) The ventral spinothalamic tract and other ascending systems of the ventral funiculus of the spinal cord. J. Comp. Neurol. 159:335-356.
Kerr, F.W.L. (1975b) Neuroanatomical substrates of nociception in the spi- nal cord. Pain 1:325-356.
Kerr, F.W.L., and H.H. Lippman (1974) The primate spinothalamic tract as demonstrated by anterolateral cordotomy and commissural myelotomy. Adv. Neurol. 4:147-156.
Kniffki, K.-D., and C. Vahle-Hinz (1987) The periphery of the cat’s ventro- posteromedial nucleus (VPMp): Nociceptive neurones. In J.-M. Besson, G. Guilbaud, and M. Peschanski (eds): Thalamus and Pain. Amsterdam: Excerpta Medica, pp. 245-257.
Kniffki, K.D., and K. Mizumura (1983) Responses of neurons in VPL and VPL-VL region of the cat to algesic stimulation of muscle and tendon. J. Neurophysiol. 49:649-661.
Kuru, M. (1949) Sensory Paths in the Spinal Cord and Brainstem in Man. Japan: Sogensya.
Mantyh, P.W. (1983a) The spinothalamic tract in the primate: A re-examina- tion using wheatgerm agglutinin conjugated to horseradish peroxidase. Neuroscience 9;847-862.
Mantyh, P.W. (1983b) The terminations of the spinothalamic tract in the cat. Neurosci. Lett. 38:119-124.
Martin, B., A.V. Apkarian, and C.J. Hodge (1989) Nociceptive neurons in the cat thalamus with inputs through the ipsilateral dorsolateral funiculus. In preparation.
Mayer, D.J., D.D. Price, and D.P. Becker (1975) Neurophysiological charac- terization of the anterolateral spinal cord neurons contributing to pain perception in man. Pain 1:51-58.
McMahon, S.B., and P.D. Wall (1983) A system of rat spinal cord lamina I cells projecting through the contralateral dorsolateral funiculus. J. Comp. Neurol. 214217-223.
McMahon, S.B., and P.D. Wall (1985) Electrophysiological mapping of brain- stem projections of spinal cord lamina I cells in the rat. Brain Res. 333~19- 26.
McMahon, S.B., and P.D. Wall (1988) Descending excitation and inhibition of spinal cord lamina I projection neurons. J. Neurophysiol. 59:1204.
Mehler, W.R. (1962) The anatomy of the so-called “pain tract” in man: An analysis of the course and distribution of the ascending fibers of the fasci- culus anterolateralis. In J.D. French and R.W. Porter (eds): Basic Research in Paraplegia. Springfield, Ill.: Charles C. Thomas, pp. 26-55.
Mehler, W.R., M.E. Feferman, and W.F.H. Nauta (1960) Ascending axon degeneration following anterolateral cordotomy. An experimental study in the monkey. Brain 83:71&750.
Meyers, D.E.R., and P.J. Snow (1982) The response to somatic stimuli of deep spinothalamic tract cells in the lumbar spinal cord of the cat. J. Phy- siol. (Lond.) 329:355-371.
Milne, R.J., R.D. Foreman, G.J. Giesler, and W.D. Willis (1981) Convergence of cutaneous and pelvic visceral nociceptive inputs onto primate spino- thalamic neurons. Pain 11:163-183.
Morin, F., H.G. Schwartz, and J.L. O’Leary (1951) Experimental study of the spinothalamic and related tracts. Acta Physiol. Neurol. 26:371-396.
Mott, F.W. (1895) Experimental enquiry upon the afferent tracts of the cen- tral nervous system of the monkey. Brain 18:l-20.
Nahin, R.L., F. Anton, and J.L.K. Hylden (1987) Lamina I spinothalamic tract (STT)/spinomesencephalic tract (SMT) projection neurons in the rat ascend via the dorsolateral funiculi (DLF). SOC. Neurosci. Abstr. 13:114.
Noordenbos, W. (1966) Some aspects of anatomy and physiology of pain. In R.S. Knighton and P.R. Dumke (eds): Pain. Boston: Little Brown, pp. 249-254.
Noordenbos, W., and P.D. Wall (1976) Diverse sensory functions with an almost totally divided spinal cord. A case of spinal cord transection with preservation of part of one anterolateral quadrant. Pain 2:185-195.
Olszewski, J. (1952) The Thalamus of the Macaca Mulatta. Basel: Karger.
Perl, E.R., and D.G. Whitlock (1961) Somatic stimuli exciting spinothalamic projections to thalamic neurons in cat and monkey. Exp. Neurol. 3256- 296.
Peschanski, M., A. Briand, M. Gautron, and G. Guilbaud (1985) Electro- physiological evidence for a role of the anterolateral quadrant of the spi- nal cord in the transmission of noxious messages to the thalamic ventro- basal complex in the rat. Brain Res. 34277-84.
Peschanski, M., P.W. Mantyh, and J.M. Besson (1983) Spinal aEerents to the ventrohasal thalamic complex in the r a t An anatomical study using wheat-germ agglutinin conjugated to horseradish peroxidase. Brain Res. 278240-244.
Poggio, G.F., and V.B. Mountcastle (1960) A study of the functional contribu- tions of the lemniscal and spinothalamic systems to somatic sensibility. Bull. Johns Hopkins Hosp. 106266-316.
Sherrington, C.S., and E.E. Laslett (1903) On the anatomical course of reflex connections in the spinal cord. J. Physiol. (Lond.) 29;5&&96.
Smith, M.C. (1976) Retrograde cell changes in human spinal cord after ante- rolateral cordotomies. Location and identification after different periods of survival. Adv. Pain Res. Ther. 1:91-98.
Stevens, R.T., A.V. Apkarian, and C.J. Hodge (1985) Funicular course of cat- echolamine fibers innervating the lumbar spinal cord of the cat. Brain Res. 336243-251.
Stevens, R.T., A.V. Apkarian, and C.J. Hodge (1989) Medial, intralaminar and lateral terminations of lumbar spinothalamic tract neurons: A fluo- rescent double label study. Somatosens. Motor Res. 6:285-308.
Stevens, R.T., A.V. Apkarian, and C.J. Hodge (1988) The location of spino- thalamic axons within the dorsolateral funiculus of cat. Soc. Neurosci. Abstr. 14:120.
Szabo, J., and W.M. Cowan (1984) A stereotaxic atlas of the brain of the Cynomolgus monkey (Macaca fascicularis). J. Comp. Neurol. 300:222- 265.
Tasker, R.R., L.W. Organ, LH. Rowe, and P. Hawrylyshyn (1976) Human spi- nothalamic tract-stimulation mapping in the spinal cord and brainstem. In J.J. Bonica and D. Albe-Fessard (eds): Advances in Pain Research and Therapy. New York Raven, pp. 251-257.
Trevino, D.L., R.A. Maunz, R.N. Bryan, and W.D. Willis (1972) Location of cells of origin of the spinothalamic tract in the lumbar enlargement of cat. Exp. Neurol. 34:64-77.
Vierck, C.J., and M.M. Luck (1979) Loss and recovery of reactivity to noxious stimuli in monkeys with primary spinothalamic cordotomies, followed by secondary and tertiary lesions of other cord sectors. Brain 102.233-248.
White, J.C. (1954) Conduction of pain in man. Arch. Neurol. Psychiatry 71:l- 23.
White, J.C., and W.H. Sweet (1969) Pain and the Neurosurgeon. Springfield, Illinois: C. Thomas.
Willis, W.D., D.R. Kenshalo, and R.B. Leonard (1979) The cells of origin of the primate spinothalamic tract. J. Comp. Neurol. 188:543-574.
Willis, W.D., D.L. Trevino, J.D. Coulter, and R.A. Maunz (1974) Responses of primate spinothalamic tract neurons to natural stimulation of hindlimb. J. Neurophysiol. 37;35&372.
Willis, W.D. (1987) The spinothalamic tract in primates. In J.-M. Besson, G. Guilbaud, and M. Peschanski (eds): Thalamus and Pain. Amsterdam: Excerpta Medica, pp. 35-47.
Yezierski, R.P. (1988) Spinomesencephalic tract: Projections from the lum- hosacral spinal cord of the rat, cat, and monkey. J. Comp. Neurol. 267:131-146.
Yezierski, R.P., L.S. Sorkin, and W.D. Willis (1987) Response properties of spinal neurons projecting to midbrain or midbrain-thalamus in the mon- key. Brain Res. 437:165-170.
Yokota, T., N. Koyama, and Y. Nishikawa (1987) Nociceptive neurons in the shell region of the ventrohasal complex of the thalamus. In L.M. Pubols and B.J. Sessle (eds): Effects of Injury on Trigeminal and Spinal Somato- sensory Systems. New York Alan R. Liss, Inc., pp. 305-312.
Zemlan, F.P., C.M. Leonard, L.-M. Kow, and D.W. Pfaf€ (1978) Ascending tracts of the lateral columns of the rat spinal cord A study using the silver impregnation and horseradish peroxidase techniques. Exp. Neurol. 62:29a334.