Embed Size (px)
Citation preview

THE ~JRNAL. OF ~JIOLOCICAL CHEMISTRY Vol. 254, No. 19. Im~e of October IO. pp. 9695-9702, 1979 Printed m U. S. A.
Membrane Receptors for Aspartate and Serine in Bacterial Chemotaxis”
(Received for publication, March 22, 1979)
Steven Clarke+ and D. E. Koshland, Jr. From the Department of Biochemistry, University of California, Berkeley, California 94720
High affinity binding sites for serine and aspartate have been characterized in membranes from Salmo- nella typhimurium and Escherichia coli. Greater than 80% of these sites have been identified as chemotaxis receptors. Mutants lacking binding sites for these amino acids have been shown to have corresponding defects in taxis. The substrate specificity of each of the receptors in Salmonella is very high; most analogs of serine and aspartate do not bind to these receptor sites and do not affect chemotaxis. The transport of these amino acids is apparently not related to chemotaxis. At least 2600 serine receptors and 1200 aspartate recep- tors with dissociation constants of about 5 pM are pres- ent in the membrane fraction of logarithmically grow- ing cells.
Bacteria can respond to a variety of chemicals in their environment by swimming towards some of them and away from others (l-3). Two of the most effective attractants for enteric species are the amino acids serine and aspartate (4,5). For the carbohydrate attractants such as ribose and galactose in Escherichia coli and Salmonella typhimurium, the behav- ioral response is mediated by periplasmic, soluble binding proteins which function as primary receptors (6, 7). In the case of the amino acids serine and aspartate, however, recep- tors have been identified by behavioral studies (4, 5), but the receptor molecules for these compounds have not been iden- tified and characterized (3). Indeed, present evidence suggests that aspartate is not detected through a periplasmic binding protein (8).
Recently, it has been demonstrated that taxis to amino acids is mediated through a class of carboxyl methylated intrinsic cytoplasmic membrane proteins in E. coli (9, 10). Aspartate taxis is dependent upon a 60,000-dalton methylated polypeptide(s) produced by the tar gene; the bulk of serine taxis is dependent upon a similar polypeptide(s) produced by the tsr gene. Because the nature of the receptors for serine and aspartate chemotaxis is unknown, it is not clear how sensory information can be focused through these methylated proteins in the initial steps of information processing. It is the purpose of this study to develop an assay for the primary receptor molecules for amino acid chemotaxis and to charac- terize their localization and specificity as well as their relation
* This research was supported in part by United States Public Health Grant AM-09765. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ A Fellow of the Miller Institute for Basic Research in Science during the course of the work. Present address, Department of Chem- istry and Molecular Biology Institute, University of California, Los Angeles, Calif. 90024.
to the methylated proteins. We report here the identification of specific high affinity receptor molecules for serine and aspartate in the membrane fraction of S. typhimurium and E. coli.
EXPERIMENTAL PROCEDURES
MuteriaZs-L-[U-‘4C]Aspartic acid (193 mCi/mmol) and L-[U- ‘%]serine (150 mCi/mmol) were obtained from New England Nu- clear. Amino acids and analogs were obtained at the highest available purity from Sigma, Calbiochem, and VegaFox. The level of contami- nation of these compounds with free serine and aspartate was deter- mined by amino acid analysis on a Beckman model 120C system under conditions where a 0.003 to 0.01% impurity could be detected.
The amount of L-serine contaminating n-serine preparations was determined by enzymatic resolution. Incubations were performed in a mixture containing 20 mM D-SWiDe, 125 pg/ml of catalase (Sigma, beef liver, 2~ crystallized), and 250 pg/ml of D-aDhO acid oxidase (Sigma, hog kidney, 19 units/mg of protein) in a buffer of 0.1 M NaPO+ 1 mM EDTA, pH 7, for 28 h at 3O’C. The reaction was stopped by boiling for 2 min. A control was performed where the reaction was stopped at zero time in this same fashion. Amino acid analysis indicated that 1.5% of the total serine was resistant to enzymatic degradation and was probably present as L-isomer impu- rity.
Bacterial Strains-Salmonella typhimurium strains SL 1509 and SL 4044 were obtained from B. A. D. Stocker, Stanford University. All other Salmonella strains used are derivative of wild type strain ST 1 (11) and are described in the text. Escherichia coli strains were obtained from J. S. Parkinson, University of Utah.
Membrane Preparation-Cells were grown with aeration at 37’C in nutrient broth (Difco, 8 g/liter) and NaCl (5 g/liter) and were harvested in late log phase (Aw nnI = 0.8 to 1.0) by Sharples centrif- ugation. Thymine was added when required at 30 pg/ml. Typically, 25 g of frozen cell paste from 25 liters of culture was resuspended in 75 ml of standard buffer (0.1 M NaPO+ 1 mM EDTA, pH 7.0) and sonicated for 6 min at full power with the standard %-inch probe of a Heat Systems model W-220F cell disruptor. The sample was kept below 1O“C in a rosette cooling cell immersed in a stirred ice bath during this procedure. Whole cells were removed by centrifugation at 7500 X g for 15 min at 4°C in a SS-34 rotor of a Sorvall RC-5 centrifuge. Membranes were pelleted from the supernatant by cen- trifugation at 236,000 X g for 30 min at 4°C in a Ti45 rotor of a Beckman L5-75 ultracentrifuge. The membrane fraction was resus- pended in 40 ml of buffer by passage through a 4-inch 16-gauge needle. Whole cells were spun out as before, and membranes were collected by ultracentrifugation.
The final membrane pellet, consisting of cytoplasmic membrane, outer membrane, and cell wall material, was resuspended in 6 ml of buffer and centrifuged once more at low speed to remove any remain- ing whole cells or large debris. The protein concentration of these membranes was determined by a modified Lowry assay (12) using a standard of bovine serum albumin whose concentration was deter- mined by ultraviolet spectroscopy (Ac$,mZim’ = 6.7) (13). All samples for protein determination were first precipitated with 10% trichloro- acetic acid. Membranes were stored at -20°C in small aliquots at protein concentrations of 25 to 35 mg/ml.
Membrane Binding Assay-Serine and aspartate binding to mem- branes was determined by a competition centrifugation assay. Mem- branes (200 ~1) were mixed with undiluted ‘%-amino-acid (final concentration 0.2 to 30 pM, 300 cpm/pmol) at 0°C and 25 mg of
9695
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

9696 Amino Acid Chemoreceptors in Salmonella and Escherichia
protein/ml. After 30 s, duplicate SO-p1 aliquots were delivered to lo- ml polycarbonate centrifuge tubes containing either 10 yl of buffer (0.1 M NaP04, 1 mu EDTA, pH 7.0) or 10 gl of a 20 mM solution of the nonisotopically labeled amino acid in the same buffer. The tubes were vortexed and immediately centrifuged at 264,ooO x g for 10 min at 4’C in the Ty65 rotor of the Beckman L5-75 ultracentrifuge. Triplicate aliquots of each supernatant (10 ~1) were mixed with 10 ml of scintillation fluid (Handifluor, Mallinckrodt) and the radioactivity was determined in a Packard 3375 scintillation spectrometer.
To determine “total radioactivity” added measurements were made of the supernatant of membranes diluted with a large excess of cold amino acid. The “free ” “C-amino acid concentration was calculated from the radioactivity in the supernatant after membrane removal. The amount of bound ?amino acid was then determined by sub- tracting the free from the total concentration.
Displacement Assay for Binding of Serine and Aspartate Analogs to Membrane Fractions-The ability of nonisotopically labeled com- pounds to displace bound L-[‘4C]aspartate and L-[14C]serine was measured in experiments similar to those described above. At a total concentration of 1 PM [‘4C]aspartate or [“Clserine, the difference in the amount of radioactivity in the supernatant after centrifugation was measured as a function of the concentration of a nonradioactive analoa added to the membrane/‘4C-amino-acid mixture. This differ- ence was compared to the difference when a large excess of either nonradioactive aspartate or serine was used. At these large concen- trations (generally 2.2 mM), the radioactivity in the supernatant asymptotically reaches a maximal value, representing the total amount of label that can be displaced from the membrane fraction. The fraction of the total bound radioactive species that can be displaced by a given concentration of a nonradioactive compound was calculated in the following way.
pared from Salmonella ST 1038, a strain lacking methyltransferase activity (16), were methylated by incubation for various times at 30°C with a soluble fraction from ST 1 cells and 55 pM S-adenosyl-r$H- methyllmethionine (8 cpm/pmol). The level of alkali-labile-methyl- ation was determined by a modification of the assay protocol of Diliberto and Axelrod (17). Methylated membranes were washed by centrifugation and resuspended to about 15 mg of protein/ml in 0.1 M NaPO+ 1 mM EDTA, pH 7.0.
Transport Assay-The ability of cells to transport L-aspartate and L-serine was determined essentially as described by Stock and Rose- man (18). Cells were grown to log phase in Medium 63, washed twice in this medium by centrifugation, and resuspended to a final AWI “,,, = 0.2. At zero time, cells were mixed at room temperature with 8 pM L-[‘?]aspartate or 8 pM L-[‘?]serine. At 0.2-min intervals, ahquots were withdrawn, diluted into buffer, and passed through Whatman GF/F glass fiber filters. The initial rate of transport was determined from the incorporation of radioactivity into the filters. Controls were performed in which cells were not added to the incubation mixture. In all cases, incorporation was linear for at least 1.2 min and extrap- olated to the control value at zero time.
RESULTS
Identification of Membrane Binding Sites for Serine and Aspartate That Appear to Be Under Flagellar Control-The binding of L-aspartate and L-serine to crude membrane frac- tions of Salmonella typhimurium and Escherichia coli at free concentrations up to 30 pM was quantitated as described under “Experimental Procedures.” To control for nonspecific bind- ing, it was desired to measure binding to cells lacking amino
radioactivity in supernatant at a given
> (
radioactivity in supernatant with no _ concentration of nonradioactive species addition of nonradioactive species >
x=/ d’ ra toactivity in supernatant with a verv\ / radioactivitv in sunernatant with no\ [large excess of Asp or Ser I-\ ” ” addrtion of nonradioactive species I
(1)
In Figs. 3, 4, and 5, this fraction is expressed in percentage terms. For analogs which do not compete for “C-amino-acid binding sites on membranes, there is 0% displacement of bound radioactivity. On the other hand, 100% displacement is obtained for analogs which can compete for all of the radioactive species sites. In these experiments, only aspartate or serine was radioactive. The expression for the dissociation constant for the binding of analogs to “C-amino-acid sites on membranes can be derived from the mass action law as
s(1-x)-2px+(S~-44Px)“~(1-x) L K,I = 0” (2)
where S is the total concentration of the nonradioactive competing species, P is the total concentration of binding sites, and X is the fraction of the radioactive species that is displaced from the mem- brane as described in Equation 1. In the case where P is small compared to S and K,,, this equation simplifies to
K d
= S(1 - X) X (3)
In these experiments, P varies from 1 to 3 pM. The use of Equation 2 was only necessary for experiments in which various concentrations of noniaotopically labeled aspartate and serine were used to compete for L-[‘Qaspartate and r,-[?Y]serine binding sites, respectively.
Chemotaris Assay-A semiquantitative temporal assay of chem- otaxis was performed according to the procedure of Spudich and Koshland (14). An ever-tumbly Salmonella mutant strain (ST 171, cheT-) was grown at 30°C to mid-log phase (A6mnm = 0.2 to 0.5) in Vogel-Bonner citrate medium (15) supplemented with 1% glycerol, 50 pg/ml of L-histidine, and 30 pg/ml of thymine. Cells were washed by centrifugation and resuspended in this medium at an Asso “,,, of 0.5. At zero tke, 5 ~1 of the cell suspension was mixed in a small test tube with 45 al of the comoound to be tested in a buffer of 0.1 M NaP04, 1 mM E’DTA, pH 7.01 A 5.~1 sample was immediately removed for direct microscopic observation as described (14). These cells respond well to both serine and aspartate by a period of smooth swimming followed by a return to the tumbling phenotype. The time required for 80 to 90% of the cells to recover from smooth swimming was taken as the chemotactic response. All observations were made at 30°C.
In Vitro Methylation of Membrane Fractions-Membranes pre-
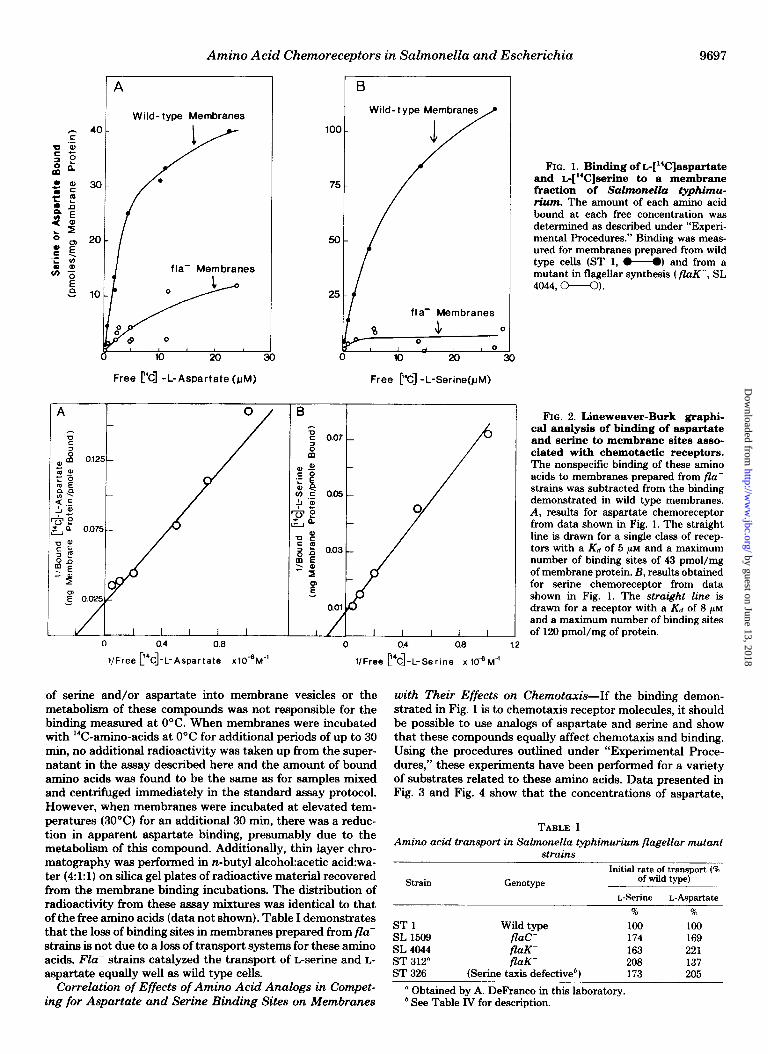
acid receptors. Although such strains had not yet been de- scribed, it has become clear that cells defective in flagellar synthesis (flu-) do not produce or assemble many, if not all, of the proteins required for motility and chemotaxis and it seemed reasonable that these cells might also lack amino acid receptors. That this is the case is shown in Fig. 1, where results obtained when amino acid binding was measured to wild type Salmonella membranes (ST 1) were compared to those obtained with membranes from strain SL 4044 (flaK-). Membranes from two other mutants in flagellar synthesis, SL 1509 (flaC-) and ST 312 (flaK-), gave similar binding curves as SL 4044; membranes from a strain lacking methylation of the 60,000-dalton intrinsic membrane proteins (ST 1038, Ref. 16) gave binding results similar to those seen for wild type membranes. Fig. 2 shows a double reciprocal plot of the binding specific to fla’ strains for L-aspartate and r.-serine with Salmonella membranes.
The large differences in amino acid binding to membranes of flu+ and flu- strains indicate that the bulk of the binding activity (>80%) in wild type cells obtained at low concentra- tions of amino acids is to proteins regulated by a flagellar control system. This control system operates in such a way that mutations in any of a number of flu genes inhibit the synthesis of flagellin (19), the assembly of basal body compo- nents (20), and the synthesis of at least two proteins involved in chemotaxis, including the cheR methyltransferase’ and the cheX methylesterase (21, see also Ref. 22). These mutations in the flu genes do not appear to affect other cellular functions, including transport (see below). The results obtained here suggest that the expression of the se&e and aspartate chemo- receptor genes are also coordinated with those of flagellar and chemotaxis genes.
Controls were performed to demonstrate that the transport ’ S. Clarke, K. T. Sparrow, and D. E. Koshland, Jr., manuscript in
preparation.
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from
Amino Acid Chemoreceptors in Salmonella and Escherichia 9697
1
I Wild- type Membranes Wild- type Membranes
fla- Membranes
Free PC] -L-Aspartate (uM)
0 0.4 0.8
l/Free p’C]-L-Aspartate ~10~‘Me’
Free [‘T] -L-Serine(pM)
30
FIG. 1. Binding of b[“C]aspartate and L@3]serine to a membrane fraction of Salmonella typhimu- rium. The amount of each amino acid bound at each free concentration was determined as described under “Experi- mental Procedures.” Binding was meas- ured for membranes prepared from wild type cells (ST 1, M) and from a mutant in flagellar synthesis ( fluK-, SL 4044, o--o).
FIG. 2. Lineweaver-Burk graphi- cal analysis of binding of aepartate and serine to membrane sites asso- ciated with chemotactic receptors. The nonspecific binding of these amino acids to membranes prepared from flu- strains was subtracted from the binding demonstrated in wild type membranes. A, results for aspartate chemoreceptor from data shown in Fig. 1. The straight line is drawn for a single class of recep- tors with a Kd of 5 CM and a maximum number of binding sites of 43 pmol/mg of membrane protein. B, results obtained for serine chemoreceptor from data shown in Fig. 1. The straight line is drawn for a receptor with a Kd of 8 CM and a maximum number of binding sites of 120 pmol/mg of protein.
0 0.4 0.8 1.2
l/Free PC]-L-Serine x 10m61d
of serine and/or aspartate into membrane vesicles or the metabolism of these compounds was not responsible for the binding measured at 0°C. When membranes were incubated with ‘%-amino-acids at 0°C for additional periods of up to 30 min, no additional radioactivity was taken up from the super- natant in the assay described here and the amount of bound amino acids was found to be the same a8 for samples mixed and centrifuged immediately in the standard assay protocol. However, when membranes were incubated at elevated tem- peratures (30°C) for an additional 30 min, there was a reduc- tion in apparent aspartate binding, presumably due to the metabolism of this compound. Additionally, thin layer chro- matography was performed in n-butyl alcohokacetic acid:wa- ter (4:l:l) on silica gel plates of radioactive material recovered from the membrane binding incubations. The distribution of radioactivity from these assay mixtures was identical to that of the free amino acids (data not shown). Table I demonstrates that the loss of binding sites in membranes prepared from flu- strains is not due to a loss of transport systems for these amino acids. Fla- strains catalyzed the transport of L-serine and L- aspartate equally well as wild type cells.
Correlation of Effects of Amino Acid Analogs in Compet- ing for Aspartate and Serine Binding Sites on Membranes
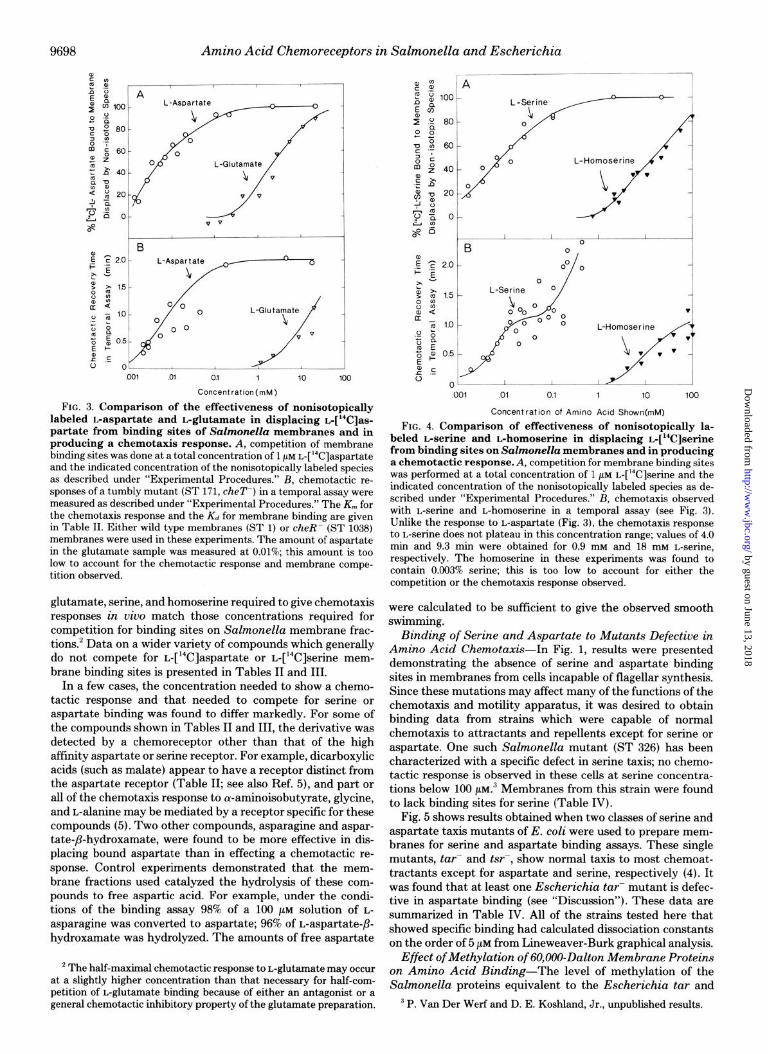
with Their Effects on Chemotaxis-If the binding demon- strated in Fig. 1 is to chemotaxis receptor molecules, it should be possible to use analogs of aspartate and serine and show that these compounds equally affect chemotaxis and binding. Using the procedures outlined under “Experimental Proce- dures,” these experiments have been performed for a variety of substrate8 related to these amino acids. Data presented in Fig. 3 and Fig. 4 show that the concentrations of aspartate,
TABLE I Amino acid transport in Salmonella typhimurium flagellar mutant
strains
Strain
ST 1 SL 1509 SL 4044 ST 312” ST 326
Genotype
Wild type flue- flaK- /i’aK-
(Serine taxis defective’)
Initial rate of transport (R of wild type)
L-Serine L-Aspartate % %
100 100 174 169 163 221 208 137 173 205
a Obtained by A. DeFranco in this laboratory. ‘See Table IV for description.
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from
9698 Amino Acid Chemoreceptors in Salmonella and Escherichia
B
001 ol 01 1 10 loo
Concentratmn(mM)
FIG. 3. Comparison of the effectiveness of nonisotopically labeled kaspartate and Lglutamate in displacing @C]as- partate from binding sites of Salmonella membranes and in producing a chemotaxis response. A, competition of membrane binding sites was done at a total concentration of 1 PM L-[‘4C]aspartate and the indicated concentration of the nonisotopically labeled species as described under “Experimental Procedures.” B, chemotactic re- sponses of a tumbly mutant (ST 171, cheT-) in a temporal assay were measured as described under “Experimental Procedures.” The K,,, for the chemotaxis response and the Kd for membrane binding are given in Table II. Either wild type membranes (ST 1) or cheR- (ST 1038) membranes were used in these experiments. The amount of aspartate in the glutamate sample was measured at 0.01%; this amount is too low to account for the chemotactic response and membrane compe- tition observed.
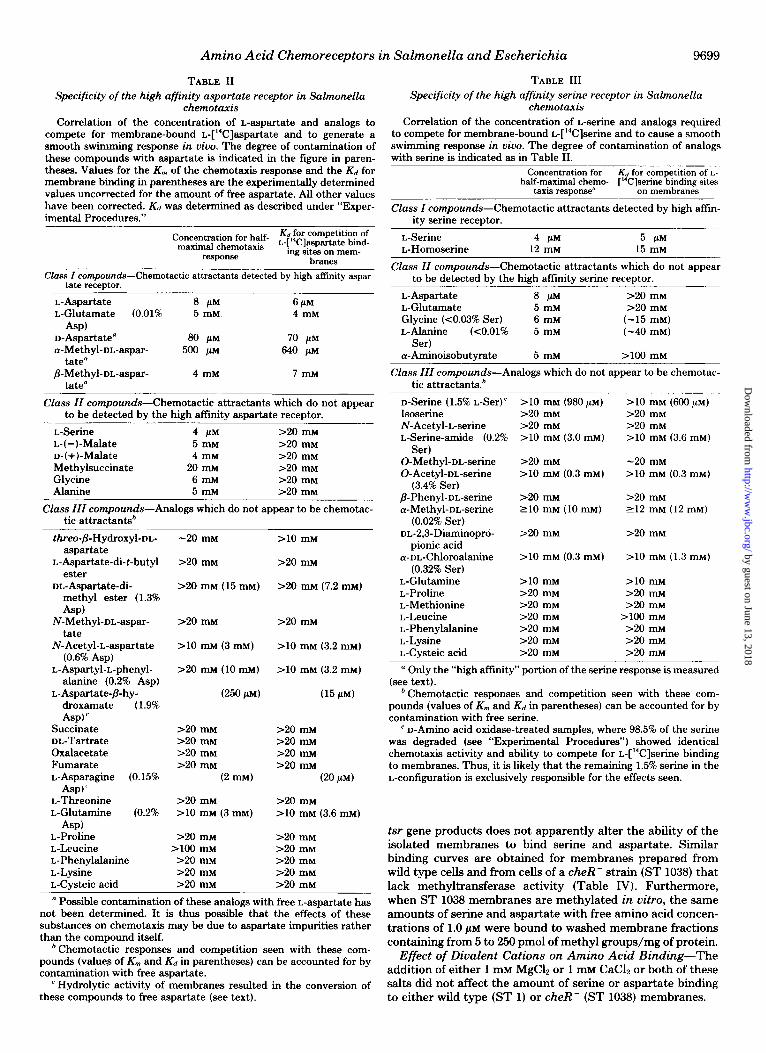
glutamate, serine, and homoserine required to give chemotaxis responses in uiuo match those concentrations required for competition for binding sites on Salmonella membrane frac- tions.2 Data on a wider variety of compounds which generally do not compete for L-[%]aspartate or L-[‘%]serine mem- brane binding sites is presented in Tables II and III.
In a few cases, the concentration needed to show a chemo- tactic response and that needed to compete for serine or aspartate binding was found to differ markedly. For some of the compounds shown in Tables II and III, the derivative was detected by a chemoreceptor other than that of the high affinity aspartate or serine receptor. For example, dicarboxylic acids (such as malate) appear to have a receptor distinct from the aspartate receptor (Table II; see also Ref. 5), and part or all of the chemotaxis response to cu-aminoisobutyrate, glycine, and L-alanine may be mediated by a receptor specific for these compounds (5). Two other compounds, asparagine and aspar- tate-P-hydroxamate, were found to be more effective in dis- placing bound aspartate than in effecting a chemotactic re- sponse. Control experiments demonstrated that the mem- brane fractions used catalyzed the hydrolysis of these com- pounds to free aspartic acid. For example, under the condi- tions of the binding assay 98% of a 100 pM solution of L-
asparagine was converted to aspartate; 96% of L-aspartate-p- hydroxamate was hydrolyzed. The amounts of free aspartate
’ The half-maximal chemotactic response to L-glutamate may occur at a slightly higher concentration than that necessary for half-com- petition of L-glutamate binding because of either an antagonist or a general chemotactic inhibitory property of the glutamate preparation.
$yJ “~ 0 L-Homoserine /
-J
,001 .Ol 0.1 1 10 100
Concent rat bon of Armno Acid Shown(mM)
FIG. 4. Comparison of effectiveness of nonisotopically la- beled L-serine and Ghomoserine in displacing L-[W]serine from binding sites on Salmonella membranes and in producing a chemotactic response. A, competition for membrane binding sites was performed at a total concentration of 1 pM r$?L!]serine and the indicated concentration of the nonisotopically labeled species as de- scribed under “Experimental Procedures.” B, chemotaxis observed with L-serine and L-homoserine in a temporal assay (see Fig. 3). Unlike the response to L-aspartate (Fig. 3), the chemotaxis response to L-serine does not plateau in this concentration range; values of 4.0 min and 9.3 min were obtained for 0.9 mM and 18 mu L-serine, respectively. The homoserine in these experiments was found to contain 0.003% serine; this is too low to account for either the competition or the chemotaxis response observed.
were calculated to be sufficient to give the observed smooth swimming.
Binding of Serine and Aspartate to Mutants Defective in Amino Acid Chemotaxis-In Fig. 1, results were presented demonstrating the absence of serine and aspartate binding sites in membranes from cells incapable of flagellar synthesis. Since these mutations may affect many of the functions of the chemotaxis and motility apparatus, it was desired to obtain binding data from strains which were capable of normal chemotaxis to attractants and repellents except for serine or aspartate. One such Salmonella mutant (ST 326) has been characterized with a specific defect in serine taxis; no chemo- tactic response is observed in these cells at serine concentra- tions below 100 PM.~ Membranes from this strain were found to lack binding sites for serine (Table IV).
Fig. 5 shows results obtained when two classes of serine and aspartate taxis mutants of E. coli were used to prepare mem- branes for serine and aspartate binding assays. These single mutants, tar- and tsr-, show normal taxis to most chemoat- tractants except for aspartate and serine, respectively (4). It was found that at least one Escherichia tar- mutant is defec- tive in aspartate binding (see “Discussion”). These data are summarized in Table IV. All of the strains tested here that showed specific binding had calculated dissociation constants on the order of 5 pM from Lineweaver-Burk graphical analysis.
Effect of Methylation of 60,000-Dalton Membrane Proteins on Amino Acid Binding-The level of methylation of the Salmonella proteins equivalent to the Escherichia tar and
3 P. Van Der Werf and D. E. Koshland, Jr., unpublished results.
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from
Amino Acid Chemoreceptors in Salmonella and Escherichia 9699
TABLE II TABLE III Specificity of the high affinity aspartate receptor in Salmonella
chemotaxis Correlation of the concentration of L-aspartate and analogs to
compete for membrane-bound @C]aspartate and to generate a smooth swimming response in vivo. The degree of contamination of these compounds with aspartate is indicated in the figure in paren- theses. Values for the K, of the chemotaxis response and the Kd for membrane binding in parentheses are the experimentally determined values uncorrected for the amount of free aspartate. All other values have been corrected. K<, was determined as described under “Exper- imental Procedures.”
Concentration for half- $‘l~I~pm~t$i~n~~ maximal chemotaxis
*ssponae ing sites on mem- branes
Class I compounds-Chemotactic attractants detected by high affinity aspar- tate receptor.
L-Aspartate L-Glutamate (0.01%
Asp)
8 PM 6w 5 mM 4 mM
D-Aspartate” a-Methyl-DL-aspar-
t&e” P-Methyl-DL-aspar-
tate”
80 j.lM 70 /AM 500 PM 640 /AM
4 mM 7 mM
Class II compounds-Chemotactic attractants which do not appear to be detected by the high affinity aspartate receptor.
L-Serine L-(-)-Malate D-(+)-Mdate Methylsuccinate Glycine Alanine
4 PM >20 rnM 5 rnM >20 rnM 4 rnM >20 rnM
20 mM >20 rnM 6 DIM >20 rnM 5 mM >20 rnM
Class III compounds-Analogs which do not appear to be chemotac- tic attractants”
Lhreo-P-Hydroxyl-DL- aspartate
L-Aspartate-di-t-butyl ester
DL-Aspartate-di- methyl ester (1.3% ASP)
N-Methyl-DL-aspar- tate
N-Acetyl-L-aspartate (0.6% Asp)
L-Aspartyl-L-phenyl- alanine (0.2% Asp)
L-Aspartate-p-hy- droxamate (1.9% ASP)’
Succinate DL-Tartrate Oxalacetate Fumarate L-Asparagine (0.15%
Asp)’ L-Threonine L-Glutamine (0.2%
Asp) L-Proline L-Leucine L-Phenylalanine L-Lysine L-Cysteic acid
-20 rnM
>20 rnM
>20 rnM (15 rnM)
>20 rnM
>lO rnM (3 rnM)
>20 mM (10 mM)
(250 PM)
>20 rnM >20 rnM >20 rnM >20 rnM
(2 mM)
>20 rnM z-10 rnM (3 rnM)
>20 rnM >lOO rnM
>20 rnM >20 rnM >20 rnM
>lO rnM
>20 rnM
>20 mM (7.2 mM)
r20 rnM
>lO mM (3.2 mM)
>lO mM (3.2 mM)
(15 /JIM)
>20 rnM >20 rnM >20 rnM >20 rnM
(20 PM)
>20 rnM >lO mM (3.6 mM)
>20 rnM >20 rnM >20 rnM >20 rnM >20 rnM
” Possible contamination of these analogs with free L-aspartate has not been determined. It is thus possible that the effects of these substances on chemotaxis may be due to aspartate impurities rather than the compound itself.
‘Chemotactic responses and competition seen with these com- pounds (values of K, and Kd in parentheses) can be accounted for by contamination with free aspartate.
’ Hydrolytic activity of membranes resulted in the conversion of these compounds to free aspartate (see text).
Specificity of the high affinity serine receptor in Salmonella chemotaxis
Correlation of the concentration of L-serine and analogs required to compete for membrane-bound L-[%]serine and to cause a smooth swimming response in vivo. The degree of contamination of analogs with serine is indicated as in Table II.
Concentration for K, for competition of L- half-maximal chemo- [“Clserine binding sites
taxis response* on membranes
Class Z compounds-Chemotactic attractants detected by high afiin- ity serine receptor.
L-Serine L-Homoserine
4 PM 5 PM 12 mM 15 mM
Class ZZ compounds-Chemotactic attractants which do not appear to be detected by the high affinity serine receptor.
L-Aspartate 8P >20 rnM L-Glutamate 5mM >20 rnM Glycine (<0.03% Ser) 6 mM (-15 rnM) L-Alanine (<O.Ol% 5 rnM (-40 mM)
Ser) c+Aminoisobutyrate 5 mM 2-100 rnM
Class IZZ compounds-Analogs which do not appear to be chemotac- tic attractants.*
D-Serine (1.5% L-Ser)’ Isoserine N-Acetyl-L-serine L-Serine-amide (0.2%
Ser) 0-Methyl-DL-serine O-Acetyl-DL-serine
(3.4% Ser) P-Phenyl-m-serine cr-Methyl-DL-serine
(0.02% Ser) DL-2,3-Diaminoprd-
pionic acid a-DL-Chlorodanine
(0.32% Ser) L-Glutamine L-Proline L-Methionine L-Leucine L-Phenylalanine L-Lysine L-Cysteic acid
>lO mM (980 PM) >20 rnM >20 rnM >lO mM (3.0 mM)
>20 rnM >lO mM (0.3 mM)
>20 rnM 210 rnM (10 rnM)
>20 rnM
>lO mM (0.3 mM)
>lO rnM >20 rnM >20 rnM >20 rnM 120 rnM >20 rnM >20 mM
>lO mM (600 PM) S-20 mM >20 rnM >lO mM (3.6 mM)
-20 rnM
>lO rnM (0.3 rnM)
>20 rnM 212 mM (12 mM)
>20 rnM
>lO rnM (1.3 rnM)
S-10 rnM 2-20 mM >20 rnM
>lOO rnM >20 rnM >20 rnM >20 mM
” Only the “high aftinity” portion of the serine response is measured (see text).
* Chemotactic responses and competition seen with these com- pounds (values of K, and & in parentheses) can be accounted for by contamination with free serine.
’ D-Amino acid oxidase-treated samples, where 98.5% of the serine was degraded (see “Experimental Procedures”) showed identical chemotaxis activity and ability to compete for L-[‘4C]serine binding to membranes. Thus, it is likely that the remaining 1.5% serine in the L-configuration is exclusively responsible for the effects seen.
tsr gene products does not apparently alter the ability of the isolated membranes to bind serine and aspartate. Similar binding curves are obtained for membranes prepared from wild type cella and from cells of a cheR- strain (ST 1038) that lack methyltransferase activity (Table IV). Furthermore, when ST 1038 membranes are methylated in uitro, the same amounts of serine and aspartate with free amino acid concen- trations of 1.0 pM were bound to washed membrane fractions containing from 5 to 250 pmol of methyl groups/mg of protein.
Effect of Divalent Cations on Amino Acid Binding-The addition of either 1 mM MgClz or 1 mM CaClz or both of these salts did not affect the amount of serine or aspartate binding to either wild type (ST 1) or cheR- (ST 1038) membranes.
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

9700 Amino Acid Chemoreceptors in Salmonella and Escherichia
TABLE IV Membrane binding sites for serine and aspartate in Salmonella
tvDhimurium and Eseherichia coli
Strain
Picomoles amino acid bound per mg Number of spe-
protein at 5 PM free cific chemotaxis Phenotype/gen- concentration receptors/cell”
otype
Swine Aspartate Serine Aspar- tate
Salmonella typhimurium ST 1 wild type 25-49 18-27 2ooo 1100 ST 1036 cheR- (16) 40-49 18-37 2500 1400 SL 4044 4.8 2.4 0 0 SL 1509
FE: a 3.2 3.7 0 0
ST 326 * serine taxis 5.0 44 550 2500 defective
E’scherichia coli RP 437 wild type 50 10 2900 600 RP 4686 g-
1.8 1.0 0 0 RP 4368 74 21 4300 1200 RP 4324 tar- 50 3.0 2900 5100 RP 4372 tsr-tar- 50 0.8 2900 0
“Calculation was made based on 5 x IO-” mg of membrane protein/cell, a Kd of 5 pi for both aspartate and serine receptors, and using flu- binding as the control for nonspecific binding.
‘Obtained from ST 1 by ICR 191 mutagenesis by Van Der Werf and Koshland.3 Normal responses to aspartate and repellents, but approximately lo&fold higher concentrations of serine are required to obtain wild type responses.
DISCUSSION
Identification of Serine and Aspartate Receptors in the Membrane-One might expect that there would be many molecular species in the bacterial membrane capable of bind- ing serine and aspartate. The results presented here demon- strate that the bulk (>80%) of the high affinity binding sites for these amino acids are associated with chemotaxis recep- tors. These binding sites are absent in membranes prepared from cells defective in flagellar synthesis (see’below), and the specificity of the binding essentially parallels that of the chemotactic response. Furthermore, one serine-defective mu- tant of SuZmonelZa which displays normal chemotaxis to attractants and repellents other than serine shows greatly reduced serine binding to the membrane fraction.
Number of Different Receptor Proteins for Serine and Aspartate-The binding measurements performed in this study were made at amino acid concentrations below 30 pM
and would not detect receptor systems with KO values of more than 50 pM. It is thus possible that there may be multiple receptor systems of which we have described only the high affinity forms. Nevertheless, the aspartate binding and chem- otaxis data do suggest that there may be only a single class of asparfate receptors with dissociation constants of about 5 PM. If lower affinity receptors are present, they do not appear to contribute markedly to the chemotactic response (see Fig. 3). On the other hand, the serine chemotaxis response is not saturated at these low concentrations-increasing responses are seen at concentrations above 10 mM (Fig. 4).4 Thus, other serine receptors must exist for chemotaxis which have lower affinities. In fact, the response to serine over many log units requires either negative cooperativity or multiple receptors.
Table V summarizes our present knowledge of amino acid chemoreceptors in Salmonella. These results should be con- trasted to those obtained in Bacillus subtilis, where all 20 amino acids appear to be detected by a complex system of receptors (23, 24).
Substrate Specificity of Binding and Chemotanis-The
’ B. Rubik and D. E. Koshland, Jr., unpublished results.
results obtained here suggest that the aspartate and serine receptors are highly specific for these amino acids. The high affinity serine receptor can respond to approximately lOOO- fold higher concentrations of L-homoserine, but not to the variety of the other amino acids and serine analogs shown in Table III. Apparently, modification of any of the functional groups of L-serine results in the loss of its ability to bind to the high affinity serine receptor or to generate a chemotaxis response. The responses reported in the past to many of the derivatives could be accounted for in our tests by the contam- ination of these samples with free serine. It has also been shown that L-isomer impurity can account for the chemotaxis effects apparently identified with D-serine.
A similar specificity was found for the aspartate chemore- ceptor. Most derivatives of aspartate were found neither to compete for membrane binding sites nor to give chemotaxis responses when corrected for the amount of contaminating free aspartate (Table II). Analogs which did appear to com- pete for the aspartate receptor include glutamate, a-methyl- DL-aSpahk?, P-methyl-DL-aspartate, and D-aspartate, al- though the latter three compounds have not been tested for the amount of free L-aspartate present as a contaminant. All other modifications of aspartate tested were ineffective as chemoattractants.
The specificities of aspartate chemotaxis (and membrane binding) measured here are clearly different from that previ- ously reported for Salmonella typhimurium (8). The results obtained here for both serine and aspartate chemotaxis sug- gest that the specificities of these receptors, at least in Sal- monella, are much greater than that suggested by the results of Melton et al. (5), or Mesibov and Adler (4). Two factors which may influence the determination of the apparent spec- ificity of amino acid receptors include problems in contami- nation of analogs with free aspartate and/or serine and me- tabolism of analogs by the cells or membrane fractions. For example, the purity of many of the derivatives used in the previous studies were not determined to rule out effects from free aspartate or serine. Additionally, data from capillary assays may reflect chemotaxis to chemically modified sub- stances since incubation at 30°C for 30 to 60 min may allow bacterial metabolism of the analogs. Although the binding assay described here can be done in a relatively short time at O”C, analogs are potentially exposed to the enzymes on inter- nal membrane surfaces. This appears to be the case for L-
asparagine and L-aspartate /?-hydroxamate, where hydrolytic activity of cells and membranes apparently readily converts these analogs into free aspartate.
Relation of Amino Acid Receptors to Methylated Mem- brane Proteins-It has recently been shown that sensory input from amino acid and other chemoreceptors is focused through one of a class of carboxyl methylated proteins in the cytoplasmic membrane of Escherichia coli. These signaling components are thought to collect sensory input from a num- ber of receptors as the first step of sensory processing. There is some specificity to this system. Information on aspartate, maltose, Ni’+, and Co’+ levels are processed through a meth- ylated tar gene product, while responses to most repellents and the bulk of the serine are processed through a tsr gene product (9, 10). The results presented here indicate that the nonstimulated level of methylation of these proteins does not itself affect the binding of serine and aspartate to the mem- branes. Examination of amino acid taxis defective strains was then undertaken to further elucidate the relation of the pri- mary receptor molecules and these methylated membrane proteins.
The aspartate-blind E. coli tar- mutants tested here do not appear to have specific binding sites for aspartate (Fig. 5,
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Amino Acid Chemoreceptors in Salmonella and Escherichia 9701
I / tsr‘
5 10 15 0 10 20
Free f‘C] -L-Aspartate(&t) Free p’C]-L-Serinehht)
FIG. 5. Binding of b[“C]aspartate and L$‘C]serine to membrane frac- tions of amino acid taxis-defective mutants of the 60,000-dalton meth- ylated proteins of Escherichia coli. A, aspartate binding to membranes. Data are shown for a wild type strain, RP 437 (A-A); a fluA-mutant, RP 4686 (M); mutants in classes of membrane-methylated proteins which do not respond to aerine or repellents (tsr-) RP 4368 (M) or do not re- spond to aspartate or repellents (tar-) RP 4324 (V-V); or a double mutant constructed from these strains (tar- tsr-) RP 4372.M). B, serine binding to the strains described above.
TABLE V Receptor systems for amino acid chemotaxis in Salmonella
typhimurium Receptor class Chemoeffectors detected KSII
L-Serine specitic L-Serine 5 PM L-Homoserine 15 mM
L-Aspartate specific” L-Aspartate 7 PM L-Glutamate 2.5 mM
Ala-Gly L-Alanine 5 mM Glycine 6 mru o-Aminoisobutyric acid 5 rnM
n a-Methyl-m-aspartate (K, = 0.6 mM) and P-methyl-Dr.-aspartate (If,,, = 5 mM) may also be detected by this receptor class (see text).
Table IV). These results open the possibility that this meth- ylated protein is the aspartate receptor itself; alternatively, it is possible that the aspartate receptor cannot function unless the tar gene product is also present. On the other hand, membranes from a mutant in the membrane-methylated pro- tein primarily associated with serine taxis in E. coli (tsr) demonstrate normal or above-normal serine binding (Table III). Here, either the methylated species functions only as a signaling component, passing on sensory information collected at the chemoreceptor, or is a receptor itself whose binding activity is not affected by the mutation. Thus, the question as to whether the tar and tsr gene products are themselves receptors for aspartate and serine is not yet established.
Quantitation of the number of aspartate and high affinity serine receptors (Table IV) indicate that there are apparently fewer binding sites for these amino acids than methyl groups on 60,000-dalton proteins. In vitro labeling of ST 1038 mem- branes gives 200 to 400 pmol of methyl groups/mg of protein.’ If each methylated protein has a single methyl group, then there are from 6,000 to 12,000 molecules/cell. The estimate for the number of serine and aspartate receptors is, on the other hand, 2,500 and 1,4OO/cell, respectively. If, however, the methylated membrane proteins have multiple sites of meth- ylation, or the binding sites are lost by inactivation, solubili- ration, or internalization in vesicles, the ratio could be altered considerably.
Relation of Amino Acid Chemotaxis to Transport-There do not appear to be any common components in the bacterial systems responsible for amino acid chemotaxis and amino acid transport. Mesibov and Adler have demonstrated that both serine- and aspartate-blind chemotaxis mutants of Esch-
erichia display normal transport of these compounds (4). Conversely, no chemotaxis defects are present in a variety of Escherichia aspartate transport mutants (25). Recently, Ordal et al. have presented evidence that proline chemotaxis in B. subtilis is also not related to the transport of this amino acid (26). The results presented here further confirm this general picture for S. typhimurium. In several fla- mutants and in one serine taxis deficient strain, normal rates of serine and aspar- tate transport were measured (Table I).
Amino Acid Receptors Appear to Be Under Flagellar Control-Flagellar and chemotaxis genes appear to be coor- dinately expressed in enteric bacteria (see above). The /Ia- mutations have been shown to lack the carboxyl methyltrans- ferase activity (16) coded by the cheR gene of Salmonella and the cheX gene of E. coli and the methylesterase activity (21) coded by the cheX gene of Salmonella ap,d the cheB gene of E. coli. The tar and tsr genes are also under similar coordinate control.’ It is now apparent that the receptors for serine and aspartate are also a part of this regulatory system.
Acknowledgments-Special thanks are due to Kristen Sparrow for expert technical assistance, to Dr. Sharon Panasenko for helpful discussions, to Dr. J. S. Parkinson and Dr. B. A. D. Stocker for supplying bacterial strains, and to Benedict Clark for the large scale fermentation of cells.
REFERENCES
1. Adler, J. (1975) Annu. Rev. B&hem. 44.341-356 2. Koshland, D. E., Jr. (1977) Adu. Neurochem. 2.277-341 3. Hazelbauer, G. L., and Parkinson, J. S. (1977) in Microbial
Interactions (Reissig, J. L., ed) Series B, Vol. 3, pp. 61-98, Chapman.and Hall, London
4. Mesibov, R., and Adler, J. (1972) J. Bacterial. 112.315-326 5. Melton, T., Hartman, P. E., Stratis, J. P., Lee, T. L., and Davis,
A. T. (1978) J. Bacterial. 133, 708-716 6. Hazelbauer, G. L., and Adler, J. (1971) Nature New Biol. 230,
101-104 7. Aksamit, R. R., and Koshland, D. E., Jr. (1974) Biochemistry 13,
4473-4478 8. Aksamit, R. R., Howlett, B. J., and Koshland, D. E., Jr. (1975) J.
Bacterial. 123, 1090-1005 9. Silverman, M., and Simon, M. (1977) Proc. Natl. Acad. Sci. U. S.
A. 74, 3317-3321 10. Springer, M. S., Coy, M. F., and Adler, J. (1977) Proc. Natl. Acad.
Sci. U. S. A. 74,3312-3316 11. Aswad, D., and Koshland, D. E., Jr. (1974) J. Bacterial. 118,640-
645 12. Bailey, J. L. (1967) Techniques in Protein Chemistry, p. 340,
American Elsevier, New York 13. Foster, J. F., and Yang, J. T. (1954) J. Am. Chem. Sot. 76, 1015-
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

9702 Amino Acid Chemoreceptors in Salmonella and Escherichia
1019 14. Spudich, J. L., and Koshland, D. E., Jr. (1975) Proc. N&Z. Acad.
Sci. U. S. A. 72, 710-713 15. Vogel, H., and Bonner, D. (1956) J. Biol. Chem. 218,97-106 16. Snrineer. W. R.. and Koshland. D. E.. Jr. (1977) Proc. Natl. Acad.
*S&.-U. S. A. +4,533-537 17. Diliberto, E. J., Jr., and Axelrod, J. (1976) J. Neurochem. 26,
1159-1165 18. Stock, J., and Roseman, S. (1971) Biochem. Biophys. Res. Com-
mun. 44, 132-137 19. Suzuki, H., and Iino, T. (1975) J. Mol. Biol. 95,549-556 20. Suzuki, T., Iino, T., Horiguchi, T., and Yamaguchi, S. (1978) J.
Bacterial. 133, 904-915 21. Stock, J., and Koshhmd, D. E., Jr. (1978) Proc. Natl. Acad. Sci.
U. S. A. X,3659-3663 22. Silverman, M., Matsumura, P., Hilmen, M., and Simon, M. (1977)
J. Bacterial. 130, 877-887 23. OrdaI, G. W., and Gibson, K. J. (1977) J. Bacterial. 129,151-155 24. OrdaI, G. W., VilIani, D. P., and Gibson, K. J. (1977) J. Bacterial.
129, 156-165 25. Schellenberg, G. D. (1977) Ph.D. thesis, University of California,
Riverside 26. Ordal, G. W., Villani, D. P., Nicholas, R. A., and Hamel, F. G.
(1978) J. Biol. Chem. 253,4916-4919
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

S Clarke and D E Koshland, JrMembrane receptors for aspartate and serine in bacterial chemotaxis.
1979, 254:9695-9702.J. Biol. Chem.
http://www.jbc.org/content/254/19/9695Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/19/9695.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

















![IP7 [호환 모드] - CHERIC · 2012. 2. 28. · with Ni(II) for the binding of histidine-tagged receptors), biotin, benzenesulfonamide, sugar, or RGD (arginine-glycine-aspartate)](https://img.pdfslide.net/doc/110x75/60fada1fefa4ac4bca4583da/ip7-eeoe-cheric-2012-2-28-with-niii-for-the-binding-of-histidine-tagged.jpg)











