Embed Size (px)
Citation preview

J. Cell Sci. 35, 267-279 (1979) 267Printed in Great Britain © Company of Biologists Limited
PRODUCTION OF SERUM PROTEINS IN
NORMAL DIPLOID FIBROBLAST-HEPATOMA
CELL HYBRIDS AND IN A9-NORMAL
LIVER CELL HYBRIDS
JOSIANE SZPIRER AND CLAUDE SZPIRERDepartment de Biologic Moliculaire, University Libre de Bruxelles,Rue des Chevaux, 67, B 1640 Rhode-St-Genese, Belgium
SUMMARY
Two series of interspecific hybrids have been generated between liver cells (which activelysecrete several serum proteins) and fibroblasts (which do not). In each series, one of the parentalcells was a normal diploid cell: mouse hepatoma cells were fused with normal diploid rat fibro-blasts, and normal rat liver cells were fused with mouse fibroblasts of the permanent line A9.The production of albumin, a-fetoprotein (AFP) transferrin and the third component of com-plement (C3) was analysed in these hybrids. Most hepatoma cell hybrids exhibit extinction ofalbumin, AFP and (to a lesser extent) transferrin; they retain the capacity to secrete C3.Normal liver cell hybrids are also characterized by the absence of albumin and transferrinproduction and by retention of C3 secretion. These results, when compared to previous resultsobtained with hybrids derived exclusively from different differentiated cells of permanent andtransformed lines show that the phenotype of such hybrids is not determined by the abnormalcharacter per se of the aneuploid parental cells. Amongst the rat fibroblast-mouse hepatoma cellhybrids, a few clones retain the capacity to actively secrete mouse albumin, AFP and transferrin,without the concomitant production of the rat serum proteins. These hybrids have lost more rat(fibroblast) chromosomes than the other clones and also have an increased number of mouse(hepatoma) chromosomes. Thus, their phenotype must result from either the complete loss of'extinguisher' chromosomes, or gene dosage effects. The significance of the lack of rat serumprotein production is also discussed, and it is suggested that retention, without concomitantactivation, could be explained in terms of diffusible regulators and heritable differences inchromatin conformation.
INTRODUCTION
Liver functions, like other differentiated traits, generally fail to be expressed inhybrids between hepatoma cells and other cells (for reviews, see Darlington & Ruddle,1975; Davidson,. 1974; Ringertz & Savage, 1976). The hybrids examined werederived with very few exceptions from the fusion of cells of hepatoma lines with cellsof other permanent lines (either fibroblasts, epithelial cells or lymphoma cells...). Theinvestigations have left unanswered the following question: are the phenotypes of suchhybrids determined by the heteroploid and/or neoplastic character of the 2 parentalcells (Ephrussi, 1972) ?The primary aim of the present work is to analyse this question.
We have previously studied hybrids between mouse hepatoma cells (which activelysecrete albumin, a-fetoprotein, transferrin and the third component of complement)and fibroblasts of permanent lines. These hybrids exhibit total extinction of albumin

268 J. Szpirer and C. Szpirer
anda-fetoprotein (AFP) production and partial extinction of transferrin, but do secretethe third component of complement (C3). Moreover, in interspecific hybrids, activa-tion of C3 secretion has been demonstrated (Szpirer & Szpirer, 1975a; Szpirer,Szpirer & Wiener, 1976).
In order to provide a definite answer to the above question, one would ideally studyhybrids derived from fusions between 2 normal diploid differentiated cells (hepatocytexfibroblast for instance). However, such hybrids would be difficult to isolate andwould very probably be incapable of prolonged proliferation. We chose therefore tostudy hybrids derived from either normal diploid fibroblasts fused with hepatoma cellsor normal hepatocytes fused with fibroblasts of permanent lines and asked the ques-tion: will extinction of the serum protein production still be observed ?
Hybrids involving normal diploid cells generally have a less stable karyotype thanhybrids derived from fusions involving 2 permanent cell lines. In particular, hybridsderived from the fusion of mouse hepatoma cells with diploid rat fibroblasts can beexpected to loose preferentially the chromosomes contributed by the diploid rat fibro-blast cell. If some specific chromosomes contributed by the fibroblast are responsiblefor the extinction of hepatic traits, then the loss of these chromosomes might allow thehybrid cell to escape extinction. Identification of these chromosomes would then bepossible in an interspecific cross. Part of this work was aimed at testing this hypothesisby trying to isolate clones secreting albumin and/or AFP.
Re-expression associated with chromosome loss has already been described: re-expression of kidney esterase 2 (Klebe, Chen & Ruddle, 1970), of liver enzymes (Croce,Litwack & Koprowski, 1973; Bertolotti, 1977; Bertolotti & Weiss, 1974; Weiss &Chaplain, 1971) and more recently, re-expression of DMSO-inducible haemoglobinsynthesis, in erythroleukemic x bone marrow cell hybrids, which was found to beassociated with the loss of an X chromosome contributed by the bone marrow cell(Benoff & Skoultchi, 1977).
MATERIALS AND METHODS
Parental cells and culture media
Three permanent lines have been used: A9, an L-cell variant (Littlefield, 1964a) deficient forthe enzyme hypoxanthine guanine phosphoribosyl transferase (HGPRT; EC 2.4.2.8);BWTG3, a clonal line of mouse hepatoma cells, deficient for HGPRT (Szpirer & Szpirer,!975^)> Fa32, a subclone of Faza967 (Deschatrette & Weiss, 1974), a clonal line of HGPRT"rat hepatoma. Fazag67 w a s kindly provided to us by Professor B. Ephrussi and Dr M. Weiss.The mouse and rat hepatoma cells secrete albumin, transferrin and C3; BWTG3 also secretesmouse AFP. BWTG3 was grown in Eagle's Medium, Dulbecco's modification (DMEM),supplemented with 10 % foetal calf serum; A9 and Fa32 were grown in Eagle's minimal essentialmedium supplemented with 10 % foetal calf serum. Freshly isolated adult rat hepatocytes wereprovided to us by Drs Drochmans and Wanson. The cells were isolated according to their enzy-mic perfusion technique (Drochmans, Wanson & Mosselmans, 1975). Rat fibroblasts wereobtained from the skin of embryos and cultured in DMEM for about 2 weeks before fusion.Concentrated culture media from the fibroblasts, or from A9, never gave a positive reactionwhen tested for the presence of albumin, AFP, transferrin or C3.

Serum protein production in cell hybrids 269
Cell fusion and selection of hybrid clones
The rat hepatocytes were kept in suspension before cell fusion. The other cells were harvestedby trypsinization. After rinsing with culture medium without serum, the cells were fused insuspension using u.v.-irradiated Sendai virus (Harris, Watkins, Ford & Schoefl, 1966). Hybridswere selected in DMEM supplemented with io~4 M hypoxanthine, 4 x io~' M aminopterin andI-6XIO 6 M thymidine (HAT medium, Littlefield, 19646): A9 and BWTG3 which lackHGPRT fail to grow in this medium and the normal diploid cells, which have a limited lifespan, do not form clones susceptible of propagation. HAT resistant colonies (one per dish) wereisolated about 2 weeks after fusion with the aid of stainless steel cylinders.
Chromosome preparations
These were made by the standard procedure, described previously (Szpirer & Szpirer, 1975 a).C-banded metaphases were prepared according to the method of Dev, Miller, Allderdice &Miller (1972) modified by Marshall (1975).
Cell incubation and double immunodiffusion
All the antisera were rabbit antisera. Anti-mouse C3 and anti-rat AFP were purchased fromNordic Pharmaceutics end Diagnostics, Tilburg, Holland. Anti-rat transferrin was purchasedfrom Microbiological Associates. Anti-mouse AFP was a gift from Dr R. Hooghe. Anti-mousealbumin, anti-rat albumin and anti-rat C3 were obtained after repeated injections of albumin orof zymosan-rat C3 complex (Mardiney & Muller-Eberhard, 1965).
The methods used were as previously described (Szpirer & Szpirer, 1975 a). At the end of thelog phase, cells were incubated with fresh medium and counted at the end of the incubationperiod (3 days). To estimate the amount of protein secreted by the hybrid cells, the concentratedmedia were serially rediluted. Dilutions of the concentrated media from the hybrid cells werecompared by immunodiffusion with dilutions of concentrated medium from hepatoma cells. Inthis way, and taking into account the number of incubated cells, the relative rates of serumprotein produced by the hybrid clones and the parental hepatoma cells could reproducibly beestimated. The sensitivity of the immunodiffusion tests was determined in the same way; in thecase of the anti-rat antiserum, the sensitivity of the assay was determined using a solution ofpurified rat AFP.
In 3 days, io9 confluent BWTG3 cells secrete about 25 fig of albumin and of AFP. Thesecretion rate of such confluent cells is constant during the first 2 days of incubation (± 10 fig/10* cells/24 h) and then levels off; this secretion rate is slightly higher than during log phase(D. Cassio, personal communication). The approximate rates of transferrin and C3 secretionare 3 and 1 fig/10* cells/24 h respectively (Szpirer & Szpirer, 1975a).
RESULTS
Cross between mouse hepatoma cells and diploid rat embryo skin fibroblasts (BS clones)
Twenty-four clones were isolated and characterized from this type fusion. Theseclones could easily be classified into two groups on the basis of their morphology.Nineteen clones showed a fibroblast-like morphology and grew as monolayers (type Iclones); their saturation density was low (manuscript in preparation). On the otherhand, the cells of three other clones (BS41, 50, 130) were more epithelial-like and veryrefractile; they were inclined to pile up and did not adhere strongly to the surface ofthe culture dish (type II clones). The two remaining clones (BS30 and BS231) appearedto be a mixture of the two cell types described above; they were thus subcloned andsubclones of each type isolated. We have not yet been able to determine whether or notthe original BS30 and BS231 clones were true clones, one cell type being derived fromthe other.
18 CEL 35

270 jf. Szpirer and C. Szpirer
Characterization of type I hybrids. The type I clones possess a complete hepatomacell genome (62 mouse chromosomes or more) combined with at least 29 rat chromo-somes (this represents 70% of the rat genome; 10 clones have retained 90% of the rat
MOUSE CHROMOSOMES RAT CHROMOSOMES
BWTG3
t * ft " « * I ft,* * * * * * «*
* * • • - - -
RSF
A
BS 30.6
litIf A * J
BS41
A A
j
Fig. 1. C-banded metaphases of the parental mouse hepatoma cells (BWTG3), therat fibroblasts (RSF), a Type I subclone (BS30-6), and a Type II clone (BS41).
Serum protein production in cell hybrids 271
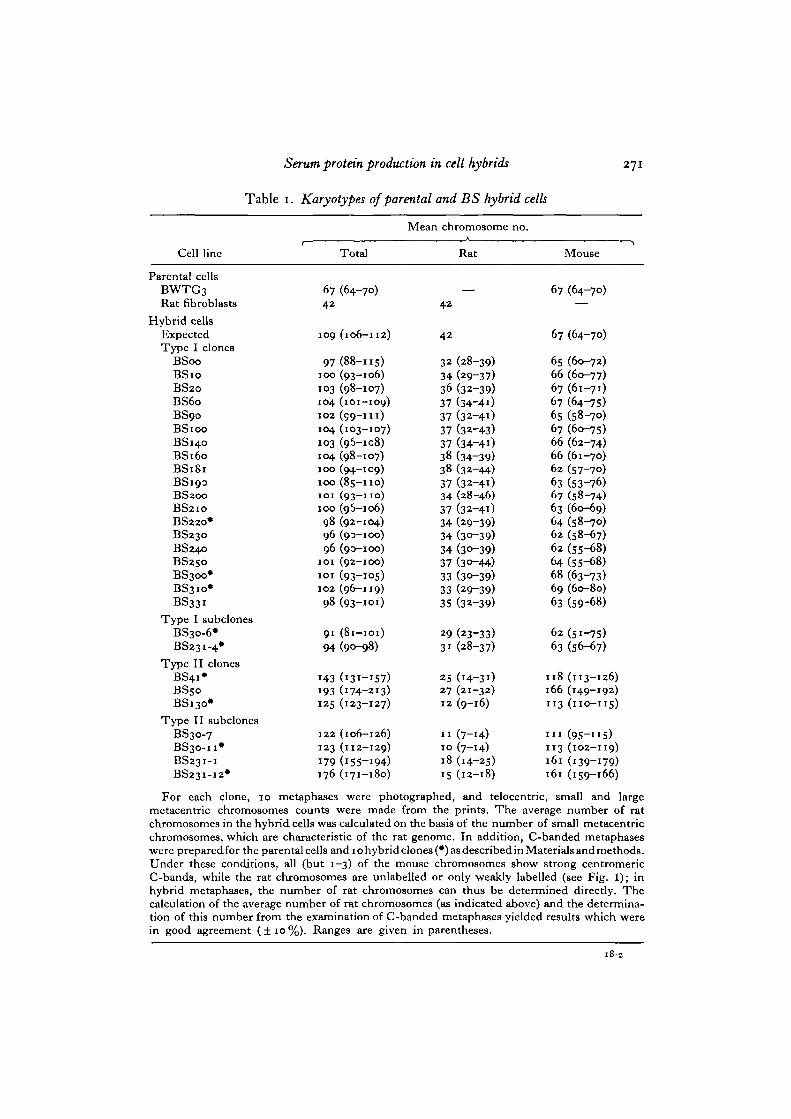
Table 1. Karyotypes of parental and BS hybrid cells
Cell line
Parental cellsBWTG3Rat fibroblasts
Hybrid cellsExpectedType I clones
BSooBS10BS20BS60BS90BS100
BS140BS160BS181BS190BSzooBS210BS22O»BS23OBS240BS25OBS3O0*BS3io»BS331
Type I subclonesBS3o-6*BS23I-4*
Type II clonesBS4i*BS50BSi 3 o #
Type II subclonesBS30-7BS30-H*BS23i-iBS23i-i2#
Total
67 (64-70)4 2
109 (106-112)
97(88-115)100 (93-106)
103 (98-107)104 (101-109)
102 (99-111)104 (103-107)
103 (95-108)104 (98-107)100 (94-1C9)100 (85-110)101 (93—110)
100 (95-i 06)98 (92-104)96 (93-100)
96 (93-100)101 (92-100)
101 (93-105)102 (96-119)
98 (93-101)
91 (81-101)
94(90-98)
143 (131-157)193 (174-213)125 (123-127)
122 (106-126)123 (112-129)
179 (155-194)176 (171-180)
Mean chromosome no.
Rat
—42
42
32 (28-39)34 (29-37)36 (32-39)37 (34-4i)37 (32-41)37 (32-43)37 (34-4i)38 (34-39)38 (32-44)37 (32-41)34 (28-46)37 (32-41)34 (29-39)34 (3O-39)34 (3O-39)37 (3O-44)33 (3O-39)33 (29-39)35 (32-39)
29 (23-33)3i (28-37)
25 (14-31)27 (21-32)
12 (9-16)
11 (7-14)10 (7-14)18 (14-25)15 (12-18)
Mouse
67 (64-70)—
67 (64-70)
65 (60-72)66 (60-77)67 (61-71)67 (64-75)65 (58-70)67 (60-75)66 (62-74)66 (61-70)62 (57-70)63 (53-76)67 (58-74)63 (60-69)64 (58-70)62 (58-67)62 (55-68)64 (55-«8)68 (63-73)69 (60-80)63 (59-68)
62 (51-75)63 (56-67)
118 (113-126)166 (149-192)113 (110-115)
HI (95-ii5)113 (102-119)
161 (139-179)161 (159-166)
For each clone, 10 metaphases were photographed, and telocentric, small and largemetacentric chromosomes counts were made from the prints. The average number of ratchromosomes in the hybrid cells was calculated on the basis of the number of small metacentricchromosomes, which are characteristic of the rat genome. In addition, C-banded metaphaseswere preparedfor the parental cells and iohybridclones(#)asdescribedinMaterialsandmethods.Under these conditions, all (but 1-3) of the mouse chromosomes show strong centromericC-bands, while the rat chromosomes are unlabelled or only weakly labelled (see Fig. 1); inhybrid metaphases, the number of rat chromosomes can thus be determined directly. Thecalculation of the average number of rat chromosomes (as indicated above) and the determina-tion of this number from the examination of C-banded metaphases yielded results which werein good agreement ( ± 1 0 % ) . Ranges are given in parentheses.
18-2
272 J. Szpirer and C. Szpirer
genome). The data on the karyotypes of the BS hybrids are illustrated in Fig. 1 andsummarized in Table 1.
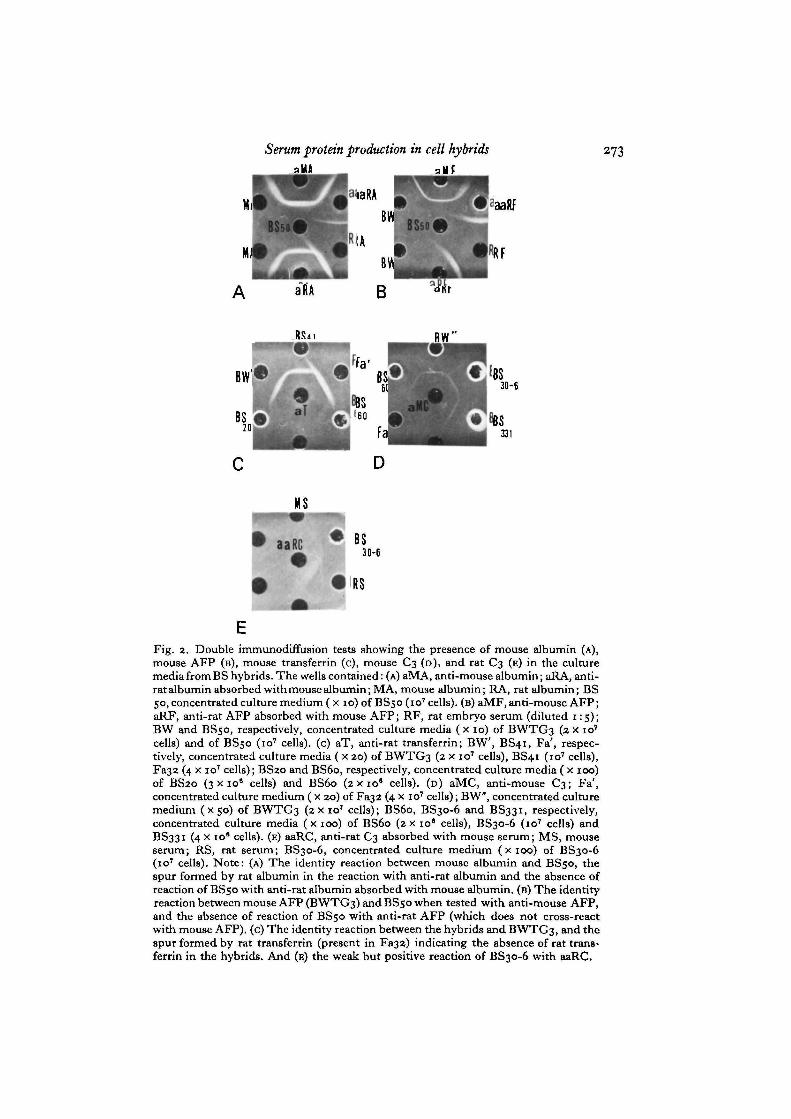
Each clone was analysed for the secretion of albumin, AFP, transferrin and C3.Albumin and AFP could never be detected in any concentrated culture medium fromthe type I hybrids. Given the sensitivity of the method, and taking into account the finalnumber of incubated cells, we can say that these BS cells, depending upon the clone,secrete less than 2-10% of the amount of albumin or AFP produced by the parentalhepatoma cells. It thus appears that albumin and AFP production is, if not totallyextinguished, considerably lowered. Several type I clones were found to secrete mousetransferrin in low amounts, as indicated by the fact that, when present, the precipitinband (see Fig. 2 c) with the anti-transferrin serum was weak or even barely detectable(the strongest reaction was observed with BS60 for which the secretion rate was esti-mated to be about 40 % of that of BWTG3). The type I BS hybrids secrete mouse C3(Fig. 2D), and some lines (5/21) also rat C3. The failure to detect rat C3 in some clonesis not significant, as the sensitivity of the assay for C3 detection is not very high (seeFig. 2E and Table 2).
Table 2. Secretion pattern of the BS hybrids
(A) Type I clones13 hybrids3 hybrids4 hybrids1 hybrid
(B) Type II clonesBS41BS50BS130BS30-7BS30-11BS231-1BS231-12
A 1 \\\ i*Yt tn.rtlULlIIlIIl
(mouse)
————
601 0 0
1 0 0
4 01 0
1 0 0
2 0 0
AFP(mouse)
————
2 0
1 0 0
1 0 0
2
S2 0 0
4 0 0
(mouse)
—20-40
2 0
—
1 0 0
2 0 0
1 0 0
4 0
3 04 0 03 0 0
c3t
Mouse
50-15050-15050-150
1 0 0
1 0 0
2 0 0
1 5 0——
2 0 0
3 0 0
Rat
——
20-50
3°
51525————
The approximate amounts of each serum protein were estimated as described in Materialsand methods. The data are expressed as percentages of the amounts secreted by BWTG3(mouse proteins) or by Fa32, a line of rat hepatoma cells (rat C3). Taking into account thenumber of incubated cells, the limit of sensitivity of the method used is: for albumin and AFP,2-10%; for transferrin, 20%; for mouse C3, 10%; for rat C3, 20% (type I hybrids) or 5 %(type II hybrids).
These results, summarized in Table 2A, show that the type I BS hybrids have apattern of serum protein secretion identical to that of hybrids derived from perma-nent heteroploid fibroblasts: they exhibit extinction of albumin and AFP production,partial extinction of transferrin production and retention of C3 secretion (Szpirer &Szpirer, 1975 a).
Characterization of type II hybrids. In contrast to the type I hybrids, all type IIhybrids retained the capacity to secrete albumin and AFP. Using the appropriate
Serum protein production in cell hybrids 273
BS331
BS30-6
f RS
Fig. 2. Double immunodiffusion tests showing the presence of mouse albumin (A),mouse AFP (B), mouse transferrin (c), mouse C3 (D), and rat C3 (E) in the culturemedia from BS hybrids. The wells contained: (A) aMA, anti-mouse albumin; aRA, anti-rat albumin absorbed with mouse albumin; MA, mouse albumin; RA, rat albumin; BS50, concentrated culture medium ( x 10) of BS50 (io7 cells), (B) aMF, anti-mouse AFP;aRF, anti-rat AFP absorbed with mouse AFP; RF, rat embryo serum (diluted 1:5);BW and BS50, respectively, concentrated culture media (xio) of BWTG3 (2 x 10'cells) and of BS50 (io7 cells), (c) aT, anti-rat transferrin; BW, BS41, Fa', respec-tively, concentrated culture media ( x 20) of BWTG3 (2 x io7 cells), BS41 (10' cells),Fa32 (4 x io7 cells); BS20 and BS60, respectively, concentrated culture media ( x 100)of BS20 (3x10' cells) and BS60 (2 x io8 cells), (D) aMC, anti-mouse C3; Fa',concentrated culture medium ( x 20) of Fa32 (4 x io7 cells); BW", concentrated culturemedium (X50) of BWTG3 (2 x io7 cells); BS60, BS30-6 and BS331, respectively,concentrated culture media (x 100) of BS60 (zx io ' cells), BS30-6 (io7 cells) andBS331 (4 x io ' cells), (E) aaRC, anti-rat C3 absorbed with mouse serum; MS, mouseserum; RS, rat serum; BS30-6, concentrated culture medium (x 100) of BS30-6(io7 cells). Note: (A) The identity reaction between mouse albumin and BS50, thespur formed by rat albumin in the reaction with anti-rat albumin and the absence ofreaction of BS50 with anti-rat albumin absorbed with mouse albumin, (B) The identityreaction between mouse AFP (BWTG3) and BS50 when tested with anti-mouse AFP,and the absence of reaction of BS50 with anti-rat AFP (which does not cross-reactwith mouse AFP), (c) The identity reaction between the hybrids and BWTG3, and thespur formed by rat transferrin (present in Fa32) indicating the absence of rat trans-ferrin in the hybrids. And (E) the weak but positive reaction of BS30-6 with aaRC.

274 J- Szpirer and C. Szpirer
antisera, we determined that only mouse albumin and mouse AFP are produced. Thisis illustrated in Fig. 2 A, B. Given the sensitivity of the assay, we would have detecteda production of rat albumin or AFP equal to ± 2 % of that of mouse albumin or AFPproduced by BWTG3 cells. An estimation of the relative rates of mouse albumin andmouse AFP secretion was made by testing serial dilutions of the concentrated media(see Materials and methods). Table 2 shows that, with the exception of the BS30 sub-clones, type II hybrids actively secrete these 2 proteins: up to 400% of the amountproduced by BWTG3 cells. Similarly, the type II clones actively secrete mouse trans-ferrin as shown in Fig. 2C. Rat transferrin could not be detected in the culture mediumof any of the hybrid clones. As expected, the type II hybrids actively produce mouseC3 (with the exception of the BS30 subclones); 3 of them (BS41, 50 and 130) alsosecrete rat C3, indicating that despite the loss of rat chromosomes, some type II clonesretained at least one chromosome carrying the C3 structural gene(s). These results aresummarized in Table 2B.
The karyotypes of the type II hybrids (see Fig. 1 and Table 1) are strikingly dif-ferent from those of the type I hybrids. Firstly they all have more chromosomes thanexpected: about twice the expected number of mouse chromosomes (or even more, asis the case for BS231 subclones). Secondly, they have undergone a more extensive lossof rat chromosomes than any of the type I clones: they retained at the most 27 ratchromosomes (whereas the type I hybrids retained at least 29 rat chromosomes). Theratio between the mouse and rat chromosome numbers is equal to, or higher than 5.(The high ploidy of BS231 subclones provides an explanation for the fact that theyproduce more albumin and AFP than BWTG3 cells.)
Cross between rat liver cells and mouse heteroploid fibroblasts [LA clones)
Rat liver suspensions, obtained by the enzymic perfusion technique are essentiallymade up of well preserved and functionally active hepatocytes (the proportion of cellsother than parenchymal cells is less than 4%; Drochmans et al. 1975). These cellscontinue to synthesize and secrete serum proteins in vitro for several hours. Albumin,transferrin and C3 were identified in the culture medium and the approximative rate ofalbumin secretion was found to be 2 /ig/h/106 cells; AFP, made only in trace amountsby the adult rat, could not be detected in the culture medium by immunodiffusion tests(May, Wanson, Szpirer and Szpirer, unpublished observations).
Such a cell preparation, freshly isolated from an adult rat liver, was fused with A9mouse fibroblasts, and numerous hybrid clones were isolated. Eleven independentclones were characterized. These hybrids have retained at least 90 and 80 % of themouse and rat chromosome complements respectively.
None of these hybrids were found to secrete albumin or transferrin in detectableamounts (i.e. less than o-oi fig albumin/h/106 cells). On the other hand, rat andmouse C3 could be detected in 4 out of the 6 clones we tested for C3 production.
These results suggest that extinction of albumin and transferrin occurred in LA hy-brids, under the influence of thefibroblastgenome. However, we cannot rule out the pos-sibility that the hybrids we tested were derived from cells other than hepatocytes, but thispossibility appearsveryunlikely: hepatocytes represent at least 96 %of the cells present

Serum protein production in cell hybrids 275
in the rat liver preparations isolated by theenzymic perfusion technique. In addition, acontrol fusion was made between such liver cells and mouse hepatoma cells (BWTG3).We isolated 8 hybrid clones (LB clones). Most of them, contrary to the LA hybridsretained only few rat chromosomes (5-20); nevertheless, one LB clone secreted ratalbumin and 3 other LB clones secreted transferrin. This indicates that at least half ofthe LB hybrids were derived from the fusion of hepatocytes with hepatoma cells (a de-tailed report on these hepatocytes x hepatoma cell hybrids will be published separately).
DISCUSSION
Our results show that, in mouse hepatoma cell hybrids, extinction of albumin andAFP production and, to a lesser extent of transferrin production can be achieved bythe genome of a fibroblast whether the latter is diploid and normal, or aneuploid andtransformed. The data on the normal liver hybrids suggest that extinction of albuminand transferrin production takes place in hepatocyte hybrids as well as in hepatoma cellhybrids. These data argue in favour of the idea that the pattern of expression of dif-ferentiated functions in cell hybrids is not determined by the abnormal character per seof the aneuploid parental cells. Our results are in agreement with those of Riddle &Harris (1976) who showed that a liver enzyme, tyrosine aminotransferase, is producedat a reduced level in rat hepatoma cell x normal diploid fibroblast hybrids as well as inhepatoma cell x lymphoma cell hybrids.
The fusion of BWTG3 hepatoma cells with normal diploid fibroblasts allowed us toisolate hybrids producing albumin and AFP (BS hybrids of type II), whereas severalfusions involving heteroploid fibroblasts of permanent lines never gave rise to this type ofhybrid. As expected, the producer BS hybrids have lost a significant number of rat(fibroblast) chromosomes; however, they have also acquired a considerable number ofmouse chromosomes. (These hybrids could have arisen from a triparental fusion, froma fusion involving a 2S BWTG3 cell, from a dikaryon in which the mouse chromosomesreplicated twice before the first cell division whereas the rat chromosomes replicatedonly once, or from a type I BS hybrid in which doubling of the whole chromosomal setwas followed by segregation of rat chromosomes.)
How can the phenotype of the type II BS hybrid be explained ? One possibility isthat the putative 'extinguisher' rat chromosomes (which are supposed to bear gene(s)directing the production of repressor) have all been lost in the type II BS hybrids,whereas at least one copy of these chromosomes have been retained in the type I BShybrids. However, if the loss of rat chromosomes is random, this hypothesis appearsstatistically unlikely.* Another possibility is that the mouse chromosomes, beinglargely supernumerary in the type II BS hybrids, overcome the extinction effect directedby the rat fibroblast chromosomes. In other words, the lack of extinction of albumin
• If only a single pair of rat chromosomes control the extinction of both albumin and AFPproduction, the probability of randomly losing the 2 copies of .this pair, when r chromosomes arelost (out of 42), is equal to Ca.C^/C^ (CJa being the number of combinations of 42 chromo-somes taken r at a time). For BS50, BS41, BS231-12, BS130 and BS30-11 respectively, theseprobabilities are 0-12, 0-16, 0-41, 0-50 and 0-58. The probability that all these 5 type II clonesrandomly lost the 2 putative extinguisher chromosomes is equal to 0-0023.

276 J. Szpirer and C. Szpirer
and AFP production would be due to a gene dosage effect, as is the case for the rathepatoma cell hybrids studied by Weiss and coworkers (Peterson & Weiss, 1972;Malawista & Weiss, 1974). However, such gene dosage effects would be observed inmouse hepatoma cell hybrids only when they have retained hepatoma and fibroblastchromosomes in a ratio of at least 5 whereas a ratio of about 2 appears sufficient in Fu 5(Peterson & Weiss, 1972) and Faza (Malawista & Weiss, 1974) hepatoma cellhybrids.
Gene dosage effects could reflect an overtitration or a dilution of repressors producedby the rat fibroblast genome, or alternatively an increased concentration of activatorsinvolved in the maintenance of the activity of the mouse albumin, AFP and transferringenes.
None of the type II BS hybrids were found to secrete rat albumin, rat AFP or rattransferrin, even though most of them actively produced the mouse serum proteinsand some of them secreted rat C3 (in agreement with our previous results; Szpireret al. 1976). It is statistically very unlikely that all these type II BS hybrids, some ofwhich retained at least one copy of the chromosome(s) bearing the C3 structural gene(s),have lost all the rat chromosomes carrying the structural genes for albumin, AFP andtransferrin (the genetic linkage of these structural genes is unknown). Therefore, thelack of production of rat albumin, rat AFP and rat transferrin in these hybrids shouldnot be considered as resulting from the loss of structural genes but rather as the lack ofactivation of these genes. Retention of differentiated functions without concomitantactivation has previously been described (Riddle & Harris, 1976; Bernstine, Koyama &Ephrussi, 1977). Riddle & Harris concluded that their results could not be easilyexplained by the mere intervention of diffusible repressors or activors, even if theseregulators were species specific. A model in which the superstructure of a gene is animportant factor in the hierarchy of the mechanisms which govern the expression ofthe gene, as discussed by Cook (1973, 1974; Colman & Cook, 1977) would appear to bemore interesting. This proposal is supported by recent studies indicating that in dif-ferentiated cells, active genes are maintained in a conformation that is distinguishablefrom that of the majority of the transcriptionally inactive DNA in the chromatin(Gottesfeld & Partington, 1977; Wallace, Dub.e & Bonner, 1977; Weintraub &Groudine, 1976). Moreover the maintenance of the active conformation does not seemto be a consequence of the presence of RNA polymerase molecules along the genes(Weintraub & Groudine, 1976; Garel, Zolan & Axel, 1977). The conformation of thegenes, whether active or inactive, might be imposed during development and be stablyconserved during mitosis (Cook, 1973, 1974). DNA modifications, such as methylationof bases (Scarano, 1969; Holliday & Pugh, 1975) might play a role in the establishmentand (or) the maintenance of these superstructures.
On the basis of the above evidence, we assume that the genes coding for albumin,AFP and transferrin are in different conformations in the mouse hepatoma cell on theone hand (active conformation), and in the rat fibroblast on the other hand (inactiveconformation). After fusion, the pre-existing conformations would be maintained, butdiffusible repressors actively produced by the fibroblast genome would shut off theexpression of the structural genes coding for albumin and AFP and to a lesser extent,

Serum protein production in cell hybrids 277
the expression of the transferrin gene. This would not necessarily alter the conforma-tion of these genes, so that re-expression might subsequently take place, as is the casefor liver enzymes (Weiss & Chaplain, 1971; Bertolotti & Weiss, 1972, 1974; Croceet al. 1973; Bertolotti, 1977) after the loss of the chromosomes carrying the repressorgene(s).
Hybrids were described in which activation of a gene was shown to accompany lackof extinction (Peterson & Weiss, 1972; Malawista & Weiss, 1974; Brown & Weiss,1975; Darlington, Bernhard & Ruddle, 1974). Such an effect would need a modifica-tion in the conformation of this normally inactive gene. Conformational changes mightor might not occur, depending on the stability of the inactive conformation and on theefficiency of the implicated regulators.
Our working hypothesis reconciles the different results yielded by various hybridiza-tion experiments. For instance, the study of albumin production in hepatoma cellhybrids has revealed the following situations: total extinction (Szpirer & Szpirer, 1975;Deschatrette & Weiss, 1975), partial extinction (Peterson & Weiss, 1972; see alsoConscience, Ruddle, Skoultchi & Darlington, 1977), retention without activation (thispaper) and finally retention with concomitant activation (Malawista & Weiss, 1974;Darlington et al. 1974)-
This hypothesis could be tested by studying the conformation of genes (as detectedby their sensitivity to nucleases) in hybrids displaying these different phenotypes.
We are grateful to Professor J. Brachet, Professor R. Thomas and Dr A. Kinsella for thought-ful reading of the manuscript. We thank Dr J. C. Wanson and Dr C. May for kindly providingus with suspensions of isolated rat hepatocytes, Dr M. Van de Winckel for giving us purifiedrat AFP and Dr C. Wuillemaert for his help in statistical calculations.
This work was carried out under a Euratom-ULB contract, an agreement between the BelgianGovernment and the Universit6 Libre de Bruxelles ('Actions de Recherches concerte'es') andwith support from the FNRS.
REFERENCES
BENOFF, S. & SKOULTCHI, A. (1977). X-linked control of hemoglobin production in somatichybrids of mouse erythroleukemic cells and mouse lymphoma or bone marrow cells. Cell 12,263-274.
BERNSTINE, E. G., KOYAMA, H. & EPHRUSSI, B. (1977). Enhanced expression of alkaline phos-phatase in hybrids between neuroblastoma and embryonal carcinoma. Somatic Cell Genet. 3,217-225.
BERTOLOTTI, R. (1977). Expression of differentiated functions in hepatoma cell hybrids:Selection in glucose-free media of segregated hybrid cells which reexpress gluconeogenicenzymes. Somatic Cell Genet. 3, 579-602.
BERTOLOTTI, R. & WEISS, M. C. (1972). Expression of differentiated functions in hepatoma cellhybrids. Extinction and re-expression of liver alcohol dehydrogenase. Biochimie 54, 195-201.
BERTOLOTTI, R. & WEISS, M. C. (1974). Expression of differentiated functions in hepatoma cellhybrids. Re-expression of aldolase B in vitro and in vivo. Differentiation 2, 5-17.
BROWN, J. E. & WEISS, M. C. (1975). Activation of production of mouse liver enzymes in rathepatoma-mouse lymphoid cell hybrids. Cell 6, 481-494.
COLMAN, A. & COOK, P. R. (1977). Transcription of superhelical DNA from cell nuclei. Eur.J.Biochem. 76, 63-78.
CONSCIENCE, J. F., RUDDLE, F. H., SKOULTCHI, A. & DARLINGTON, G. J. (1977). Somatic cellhybrids between Friend erythroleukemia cells and mouse hepatoma cells. Somatic Cell Genet.3. IS7-I72.

278 J. Szpirer and C. Szpirer
COOK, P. R. (1973). Hypothesis on differentiation and the inheritance of gene superstructure.stature, Land. 245, 23-25.
COOK, P. R. (1974). On the inheritance of differentiated traits. Biol. Rev. 49, 51-84.CROCE, C. M., LITWACK, G. & KOPROWSKI, H. (1973). Human regulatory gene for inducible
tyrosine aminotransferase in rat human hybrids. Proc. natn. Acad. Sci. U.S.A. 70, 1268-1272.
DARLINGTON, G. J., BERNHARD, H. P. & RUDDLE, F. H. (1974). Human serum albumin pheno-type activation in mouse hepatoma-human leucocyte cell hybrids. Science, N. Y. 185, 859-862.
DARLINGTON, G. J. & RUDDLE, F. H. (1975). Studies of hepatic phenotypes expressed in somaticcell hybrids: a summary. Mod. Trends Human Genet. 2, 111-137.
DAVIDSON, R. (1974). Gene expression in somatic cell hybrids. A. Rev. Genet. 8, 195-217.DESCHATRETTE, J. & WEISS, M. C. (1974). Characterization of differentiated and dedifferentiated
clones from a rat hepatoma. Biodumie 56, 1603-1612.DESCHATRETTE, J. & WEISS, M. C. (1975). Extinction of liver specific functions in hybrids
between differentiated and dedifferentiated rat hepatoma cells. Somatic Cell Genet. 1, 279-292.
DEV, V. G., MILLER, D. A., ALLDERDICE, P. W. & MILLER, O. J. (1972). Method for locating thecentromere of mouse meiotic chromosomes and its application to T163H and T70H trans-locations. Expl Cell Res. 73, 259-261.
DROCHMANS, P., WANSON, j . C. & MOSSELMANS, R. (1975). Isolation and subfractionarion onficoll gradients of adult rat hepatocytes. J. Cell Biol. 66, 1-22.
EPHRUSSI, B. (1972). Hybridisation of Somatic Cells. Princeton, New Jersey: Princeton Univer-sity Press.
GAREL, A., ZOLAN, M. & AXEL, R. (1977). Genes transcribed at diverse rates have similar con-formation in chromatin. Proc. natn. Acad. Sci. U.S.A. 74, 4867-4871.
GOTTESFELD, J. M. & PARTINGTON, G. A. (1977). Distribution of messenger RNA-codingsequences in fractionated chromatin. Cell 12, 953-962.
HARRIS, H., WATKINS, J. F., FORD, C. E. & SCHOEFL, G. I. (1966). Artificial heterokaryons ofanimals cells from different species. .7. Cell Sci. 1, 1-30.
HOLLIDAY, R. & PUGH, J. E. (i975). DNA modification mechanisms and gene activity duringdevelopment. Science, N.Y. 187, 226-232.
KLEBE, R. J., CHEN, T. R. & RUDDLE, F. H. (1970). Mapping of a human genetic regulator ele-ment by somatic cell genetic analysis. Proc. natn. Acad. Sci. U.S.A. 66, 1220-1227.
LlTTLEFlELD, J. W. (1964a). Three degrees of guanylic acid-inosinic acid phosphorylasedeficiency in mouse fibroblasts. Nature, Land. 203, 1142-1144.
LlTTLEFlELD, J. W. (19646). Selection of hybrids from matings of fibroblasts in vitro and theirpresumed recombinants. Science, N.Y. 145, 709—710.
MALAWISTA, S. E. & WEISS, M. C. (1974). Expression of differentiated functions in hepatomacell hybrids: high frequency of induction of mouse albumin production in rat hepatoma-mouse lymphoma hybrids. Proc. natn. Acad. Sci. U.S.A. 71, 927-931.
MARDINEY, M. R. & MCLLER-EBERHARD, H. J. (1965). Mouse /?iC-globulin: production ofantiserum and characterization in the complement reaction. J. Imtmm. 94, 877-882.
MARSHALL, C. (1975). A method for analysis of chromosomes in hybrid cells employing sequen-tial G-banding and mouse specific C-banding. Expl Cell Res. 91, 464-469.
PETERSON, J. A. & WEISS, M. C. (1972). Expression of differentiated functions in hepatoma cellhybrids: Induction of mouse albumin production in rat hepatoma-mouse fibroblast hybrids.Proc. natn. Acad. Sci. U.S.A. 69, 571-575.
RIDDLE, V. & HARRIS, H. (1976). Synthesis of a liver enzyme in hybrid cells. J. Cell Sci. 22,199-215.
RINCERTZ, N. R. & SAVAGE, R. E. (1976). Cell Hybrids. New York: Academic Press.SCARANO, E. (1969). Enzymatic modification of DNA and embryonic differentiation. Ann.
Embryol. Morph., suppl. 1, 51-61.SZPIRER, J. & SZPIRER, C. (1975 a). The control of serum protein synthesis in hepatoma-
fibroblast hybrids. Cell 6, 53-60.SZPIRER, C. & SZPIRER, J. (19756). A mouse hepatoma cell line which secretes several serum
proteins including albumin and a-foetoprotein. Differentiation 4, 85-91.

Serum protein production in cell hybrids 279
SZPIRER, C, SZPIKER, J. & WIENER, F. (1976). The expression of differentiated functions insomatic cell hybrids: retention and activation of C3 production. Cell Differentiation 5, 139—149.
WALLACE, R. B., DUBE, S. K. & BONNER, J. (1977). Localization of the globin gene in thetemplate active fraction of chromatin of Friend leukemia cells. Science, N.Y. 198, 1166—1168.
WEINTRAUB, H. & GROUDINE, M. (1976). Chromosomal subunits in active genes have an alteredconformation. Science, N.Y. 193, 848-856.
WEISS, M. C. & CHAPLAIN, M. (1971). Expression of differentiated functions in hepatoma cellhybrids: reappearance of tyrosine aminotransferase inducibility after the loss of chromo-somes. Proc. natn. Acad. Sci. U.S.A. 68, 3026-3030.
(Received 18 April 1978)






























