Embed Size (px)
Citation preview

Marine Ecology. 2021;00:e12668. wileyonlinelibrary.com/journal/maec | 1 of 18https://doi.org/10.1111/maec.12668
© 2021 Wiley- VCH GmbH
Received:13May2020 | Revised:18April2021 | Accepted:26May2021DOI: 10.1111/maec.12668
O R I G I N A L A R T I C L E
Productivity changes in the Mediterranean Sea drive foraging movements of yelkouan shearwater Puffinus yelkouan from the core of its global breeding range
Francesco Pezzo1 | Marco Zenatello1 | Giulia Cerritelli2 | Augusto Navone3 | Dimitri Giunchi2 | Giovanna Spano3 | Enrica Pollonara2 | Alessandro Massolo2 | Anna Gagliardo2 | Nicola Baccetti1
1Istituto Superiore per la Protezione e RicercaAmbientale(ISPRA),OzzanoEmilia,Italy2EthologyUnit,DepartmentofBiology,UniversityofPisa,Pisa,Italy3AreaMarinaProtetta“Tavolara–PuntaCodaCavallo”,Olbia,Italy
CorrespondenceFrancescoPezzo,IstitutoSuperioreperlaProtezioneeRicercaAmbientale(ISPRA),ViaCa'Fornacetta9,40064,OzzanoEmilia(BO),Italy.Email:[email protected]
AbstractPelagic seabirds are tied to their breeding colonies throughout their long- lasting breedingseason,butatthesametime,theyhavetofeedinahighlydynamicmarineenvironment where prey abundance and availability rapidly change across space and seasons.Here,wedescribetheforagingmovementsofyelkouanshearwaterPuffinus yelkouan, a seabird endemic to theMediterranean Sea that spends its entire lifecyclewithinthisenclosedbasinandwhosefutureconservationisintimatelylinkedtohuman-drivenandclimaticchangesaffectingthesea.Theaimwastounderstandthemainfactorsunderlyingthechoiceofforaginglocationsduringthereproductivephases.Atotalof34foragingtripswereobtainedfrom21breedingadultstaggedandtrackedonTavolaraArchipelago(NSardinia,Italy).Thisisthelargestandmostimportantbreedingareaforthespecies,accountingformorethan50%oftheworldpopulation. The relationships between foraging movements during two differentbreedingstagesandtheseasonalchangesofprimaryproductivityatseaweremod-eled.Movementsappearedtobeaddressedtowardinshore(<20km),highlyproduc-tive,andrelativelyshallow(<200m)foragingareas,ofteninfrontofrivermouthsandatgreatdistancesfromthecolony.Duringincubation,theBonifacioStraitandothercoastalareasclosetoNorthandWestSardiniawerethemostpreferredlocations(upto247kmfromthecolony).Duringthechick-rearingphase,someindividualsreachedareasplacedatgreaterdistancesfromthecolony(upto579km),aimingatfood-richhotspotsplaced as far north as theGulf of Lion (France). Theneed for such longdistanceandlong-lastingforagingtripsishypothesizedtoberelatedtounfavorableconditionsonthelessproductive(andalreadydepleted)Sardinianwaters.
K E Y W O R D S
foragingecology,GPSlogger,marineprimaryproductivity,MediterraneanSea,Puffinus yelkouan,yelkouanshearwater

2 of 18 | PEZZO Et al.
1 | INTRODUC TION
TheMediterraneanSeaisanalmostcompletelyenclosedbasinchar-acterizedbyalowconcentrationofnutrientsespeciallyinitsEasternpart(Boscetal.,2004;Kressetal.,2003)andcanbeclassifiedasanoligotrophicorevenultra-oligotrophicbasin(Pujo-Payetal.,2011).Itsbiologicalproductivityistypicallydominatedbyawinter–springbloom occurring in some restricted areas mostly concentrated in its NW portion (D’Ortenzio & Ribera d’Alcala, 2009; Tanhuaetal.,2013).Asaconsequence,ifcomparedtotheAtlanticOcean,it hosts a simplified community of strictlymarine seabirds (sensuGaston, 2004), both in termsof species diversity andpopulationsabundance,andcharacterizedbyahighproportionofendemictaxa ofmajorconservationconcern(Blondeletal.,2010;Colletal.2010;Zotieretal.,2003).Amongthem,theyelkouanshearwaterPuffinus yelkouan shows a decreasing population trend: It is currently consid-ered as a threatened species and has been categorized as Vulnerable ontheIUCNRedList(BirdLifeInternational,2018).Despitethis,itisstill a poorly monitored species and large undiscovered colonies may existintheEasternMediterraneanorevenintheBlackSea(BirdLifeInternational,2018;Derhé,2012).Knownbreedingsitesaremainlydistributed in the central Mediterranean basin, fromMenorca is-land and the Southern French coasts (Bourgeois & Vidal, 2008;Derhé, 2012) to the Sicilian Channel and the Aegean Sea, with aglobalpopulationsizerecentlyre-assessedat21,000–36,000pairs(Gaudard, 2018). However, as for most other burrowing petrelsbreedingathardlyaccessiblelocationssuchascliffsandcaves,reli-ablelong-termtrendsandpopulationestimatesarescarce(Buxtonetal.,2016).Hence,mostpopulationestimateshavebeenachievedby imprecisemethods such as countingbirdswhile raftingon thewatersurfaceintheproximityofcolonies(Bourgeois&Vidal,2008;Raine et al., 2010). Not surprisingly, available data are subject tosubstantialreassessmentsfollowingsteadyimprovementsinknowl-edge.Ithasbeenreliablyascertainedthattherangeofthisspecies,contrary to other procellariids, is confined to the MediterraneanandBlackSeabothduringthebreedingandnon-breedingseasons(Gaudard,2018;Pérez-Ortega&İsfendiyaroğlu,2017).Asaconse-quence,thewholepopulationappearstobestronglyexposedtotheoverall conditionof this areawhich is currently affectedbymajortransformations(e.g.,Lejeusneetal.,2010;Maciasetal.,2015)andwhich is considereda climate-changehot spot (Giorgi, 2006).Thehumanpressure isconstantly increasingwithanumberof impactsonecosystemsoftheMediterraneanSea(Michelietal.,2013)andon seabirds as a direct consequence. Concerning the yelkouanshearwater,mainthreatshavebeenidentifiedsuchasfisheriesby-catchwithinforagingareas,mortalitybyalienpredatorssuchasratsandcatsatbreedingsites,fishstockdepletionandchronicseapol-lution(Bourgeois&Vidal,2008;Capizzietal.,2010;Gaudard,2018;Ruffinoetal.,2009).Informationonthespatialecologyofthespe-ciesisscarce,andtheknowledgeonfeedingmovementsandfeedingareas is based on observations carried out at diurnal concentra-tionsitesornearbottleneckareas suchas theBosphorousor theBonifacioStrait(Şahinetal.,2012;Zenatelloetal.,2006).Tracking
studiesonyelkouanshearwatersbreedingintheMediterraneanSeaindicatedthatbirdsfromFrenchcoloniesintheHyèresArchipelagomainly move westward along the coast to the adjacent Gulf ofLion(Péronetal.,2013),whereasthosefromtheMaltesecoloniesshow a high individual variability moving both toward the coast ofTunisia/westernLibyaandtotheAegeanSea (Gattetal.,2019;Raineet al., 2013).TheSardiniankey-siteofTavolara-PuntaCodaCavalloMarine Protected Area hosts the largest known breedingpopulationofthespecies,estimatedat9,991–13,424pairs(Zenatelloetal.,2006)which,consideringthemostrecentpopulationestimates(Gaudard,2018),couldrepresentupto55%oftheglobalbreedingpopulation. Conservation actions in this area in the last decades consisted of rat eradication attempts onMolara island (Ragionierietal.,2013;Sposimoetal.,2012a,2012b)andinthesuccessfulde-ratization of Tavolara island (http://www.lifepuffinustavolara.it) in2008 and 2017, respectively.However, no protection on foragingareashasbeenspecificallyenforcedsofar,andinformationonforag-ingstrategiesofthisbreedingpopulationwastotallylacking,despiteitsrelevancefortheconservationofthisandotherpopulations.
Here,weprovidethefirststudydescribingtheforagingmove-mentsof theyelkouan shearwaters from theworld-largest colonyofTavolara,withtheaimsof:(i)identifyingmainforagingandraftingareas bymeans ofGPS loggers; (ii) describing how key ecologicalfactorsaffecttheselectionofforagingandrestingareasand,asaconsequence, the foraging trips lengthandduration in thecourseof incubationandchick rearing; (iii) hypothesizinghowchanges intheMediterraneanhabitatcouldpossiblyaffectspatialecology;(iv)providinginformationforconservationscenariostocome.
2 | MATERIAL S AND METHODS
2.1 | Study site and model species
The studywas carried out from 2011 to 2015 on Tavolara Island(40°54′N,09°42′E;Sardinia), inthe largestknownbreedingsiteoftheyelkouanshearwater(Gaudard,2018;Zenatelloetal.,2012).Allactivitieswereperformedinasinglecavernhostingupto15acces-sibleneststhatwereusuallyinarat-freecondition.Birdswerecap-turedbyhandonthenestandequippedwithGPSloggers(Gypsy-2andGypsy-4byTechnosmartand I-gotUGT120byMobileActionTechnology).GPSloggerswereattachedtothemantlefeathersofbreeding adultsby adhesiveTESA tape (totalweight: 13–20g ac-cordingtobatterysize).TheweightofGPSloggerswas3.1%–4.7%of theaveragemassofadults (424.5g± 28.6 g SD,n =29).Birdswere handled for 15–30 min and then released where trapped(i.e., at their nest). Loggers were retrieved by recapturing breed-ers on their nests or at the entrance of the cavewhen returningfrom their foraging trips. Nests were regularly monitored duringthewholestudyperiod toassess theirbreedingsuccess (Table1).Capture,handling,andtaggingprocedureswereconductedbytheItalianInstituteforEnvironmentalProtectionandResearch(ISPRA),undertheauthorizationofLaw157/1992(Art.4.1andArt7.5),which
| 3 of 18PEZZO Et al.
TAB
LE 1
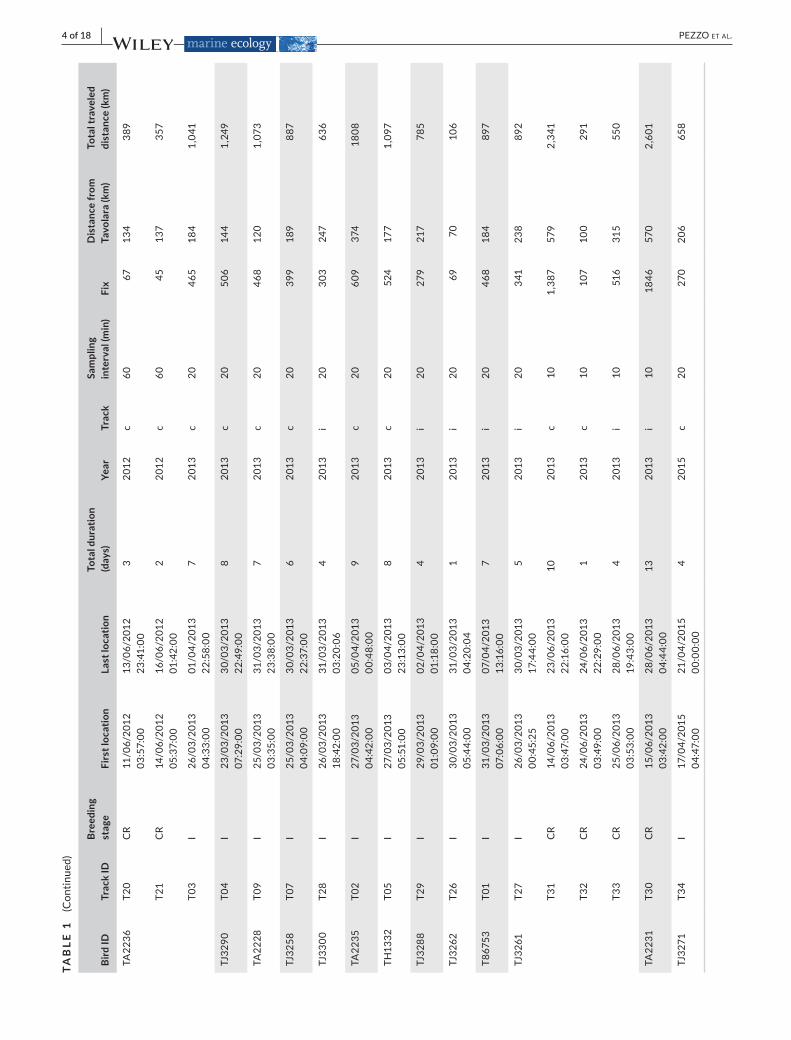
CharacteristicsofthetracksobtainedfromGPSdataloggersmountedonyelkouanshearwatersatTavolaraIsland(Sardinia,IT)from2011to2015.Foreachbird,BirdID,Track
ID(*notincludedintheanalysis),Breedingstage(I,incubation;CR,chickrearing),dateandtimeofFirstandLastlocationrecorded,GPSsamplingrates,andnumberoffixespertrackare
reported.IntheTrack,columnisspecifiedifthetrackingwascomplete(c)orifinterruptedabruptlybeforethebirdreturnedtothecolony(i).Thebeelinebetweenthemaximumdistantpoint
reachedandthecolonyisreportedasDistancefromTavolara,whiletheTotalTraveledDistancewasmeasuredasthesumofallthedistancesbetweensuccessivelocations
Bird
IDTr
ack
IDBr
eedi
ng
stag
eFi
rst l
ocat
ion
Last
loca
tion
Tota
l dur
atio
n (d
ays)
Year
Trac
kSa
mpl
ing
inte
rval
(min
)Fi
xD
ista
nce
from
Ta
vola
ra (k
m)
Tota
l tra
vele
d di
stan
ce (k
m)
TA2226
T24*
I03/04/2011
06:0
6:00
03/04/2011
13:1
6:00
120
11i
1021
6384
TJ3259
T23
I05/04/2011
04:08:00
09/04/2011
10:3
8:00
420
11i
10464
190
538
TA2229
T25
I04/04/2011
05:35:00
08/04/2011
11:55:00
420
11i
1087
165
361
T18
CR
11/0
6/20
1203:37:00
17/06/2012
23:41:00
720
12c
6016
2141
941
T19*
CR
18/0
6/20
1203:43:00
18/0
6/20
1223:42:00
120
12c
6021
69
T06
I27/03/2013
07:23:00
03/04/2013
22:1
9:00
820
13c
20511
225
1,266
TJ3270
T10*
I16/04/2012
04:20:00
17/04/2012
23:35:00
220
12c
6042
18
TJ3267
T13*
I16/04/2012
06:25:00
17/04/2012
00:1
9:00
120
12c
6019
27
T14*
I17/04/2012
07:18:00
18/04/2012
23:3
8:00
220
12c
6042
33
TJ3263
T17
CR
11/0
6/20
1203
:10:
0015/06/2012
00:58:00
420
12c
6094
118
586
TA2223
T22
I17/04/2012
05:30:07
18/04/2012
05:50:08
120
12i
2058
5328
6
T15
CR
11/0
6/20
1203
:03:
0013
/06/
2012
00:56:00
220
12c
6047
107
406
T16
CR
13/0
6/20
1205:54:00
16/0
6/20
1202:56:00
320
12c
6070
128
553
TA2225
T11*
CR
17/04/2012
07:27:00
17/04/2012
22:17:00
120
12c
6016
21
T12*
CR
19/04/2012
07:21:00
19/04/2012
22:56:00
120
12c
6017
26
T08
I26
/03/
2013
06:41:00
30/0
3/20
1321:41:16
520
13c
2030
912
018
9
(Continues)
4 of 18 | PEZZO Et al.
Bird
IDTr
ack
IDBr
eedi
ng
stag
eFi
rst l
ocat
ion
Last
loca
tion
Tota
l dur
atio
n (d
ays)
Year
Trac
kSa
mpl
ing
inte
rval
(min
)Fi
xD
ista
nce
from
Ta
vola
ra (k
m)
Tota
l tra
vele
d di
stan
ce (k
m)
TA2236
T20
CR
11/0
6/20
1203:57:00
13/0
6/20
1223:41:00
320
12c
6067
134
389
T21
CR
14/06/2012
05:37:00
16/0
6/20
1201:42:00
220
12c
6045
137
357
T03
I26
/03/
2013
04:33:00
01/04/2013
22:58:00
720
13c
20465
184
1,041
TJ3290
T04
I23
/03/
2013
07:29:00
30/0
3/20
1322:49:00
820
13c
20506
144
1,249
TA2228
T09
I25/03/2013
03:35:00
31/0
3/20
1323
:38:
007
2013
c20
468
120
1,073
TJ3258
T07
I25/03/2013
04:09:00
30/0
3/20
1322:37:00
620
13c
2039
918
9887
TJ3300
T28
I26
/03/
2013
18:42:00
31/0
3/20
1303
:20:
064
2013
i20
303
247
636
TA2235
T02
I27/03/2013
04:42:00
05/04/2013
00:48:00
920
13c
2060
9374
1808
TH1332
T05
I27/03/2013
05:51:00
03/04/2013
23:1
3:00
820
13c
20524
177
1,097
TJ3288
T29
I29
/03/
2013
01:0
9:00
02/04/2013
01:1
8:00
420
13i
20279
217
785
TJ3262
T26
I30
/03/
2013
05:44:00
31/0
3/20
1304:20:04
120
13i
2069
7010
6
T86753
T01
I31
/03/
2013
07:06:00
07/04/2013
13:1
6:00
720
13i
20468
184
897
TJ3261
T27
I26
/03/
2013
00:45:25
30/0
3/20
1317:44:00
520
13i
20341
238
892
T31
CR
14/06/2013
03:47:00
23/0
6/20
1322
:16:
0010
2013
c10
1,387
579
2,341
T32
CR
24/06/2013
03:49:00
24/06/2013
22:2
9:00
120
13c
10107
100
291
T33
CR
25/06/2013
03:53:00
28/0
6/20
1319:43:00
420
13i
10516
315
550
TA2231
T30
CR
15/06/2013
03:42:00
28/0
6/20
1304:44:00
1320
13i
101846
570
2,601
TJ3271
T34
I17/04/2015
04:47:00
21/04/2015
00:0
0:00
42015
c20
270
206
658
TAB
LE 1
(Continued)

| 5 of 18PEZZO Et al.
regulates research onwild bird species. Three birdswere trackedinsubsequentyears (Table1).Thedatasetsgeneratedduringand/oranalyzedduringthecurrentstudyareavailableintheMovebankData Repository, https://doi.org/10.5441/001/1.k49p9t60 (Pezzoetal.,2021).
2.2 | Data analysis
Thesampling rateof the loggerschangedduring thestudyperiod(10, 20 or 60 min) depending on GPS receiver, battery size, andbreedingstage (seeTable1 fordetails).Foreach fix, theGPS log-gersrecordeddate,time,speed,andpositionaldata(LongitudeandLatitude).TrackswereplottedonQgis (http://www.qgis.org/),andindividual foragingtripswere identifiedasround-tripsflightsfromthecolonytofeedingareas.Onetofourtripsperindividualwasre-corded. For each fix, the distance from the preceding fix and thedistancefromthecolony(i.e.,theminimaldistanceabirdcanflytoreachthecurrentpoint,assumingshearwatersdidnotflyoverland)werecalculated.Inordertoidentifytheforagingareasandtheac-tivityofthebirds,eachfixwasclassifiedintooneofthefollowingthreecategories:travelling(T),resting(R),foraging(F)correspond-ingtothreedistinctmovementpatterns.While travelling, thebirdmoveswithaconsistentlyhighspeedbetweentwodistantsites.Theresting behavior is characterized by low speed movements within a shortrangeattributabletotheseacurrentsandwaves.Intheforag-ingareas,shearwatersperformshortrangemovementseitheratlowormediumspeed(seebelow).
Indetail,theclassificationwasbasedonthefollowingcriteria:
1. Fixes with a speed greater than 10 km/h have been classifiedas T (Guilford et al., 2008), provided that their distance fromthe preceding fix was greater than 5, 10, and 30 km for thesamplingrateofonefixevery10,20,and60min, respectively.In addition, each fix with a speed greater than 10 km/h notmeeting the previous conditions was anyhow classified as T,providedthattheabsolutedifferencebetweenitsdistancefromthe colony and thepreceding fixdistance from the colonywasgreater than 2.5, 5, and 15 km, for the sampling rate of onefix every 10, 20, and 60min, respectively. The latter criterionwas adopted to discriminate patterns of birds travelling alonga curved path (e.g., as needed to circumnavigate islands) fromthose displayed by birds moving rapidly within a restricted foraging area. All the distance categories for the classificationof the T fixes have been arbitrarily chosen on the basis of thedirect observation of the tracks of birds that were travelingbetween two distant areas.
2. Rfixesincludedallthefixeswithaspeedlowerthan5km/h,pro-videdthatthedistancefromtheprecedingfixwasshorterthan0.5 kmwhen the sampling ratewas every 10 and 20min, and1.5kmwhenthesamplingratewasof1fixevery60min.Thecri-terionbasedonthedistancefromtheprecedingfixwasactually
unreliableincaseofeitherstrongdriftorshortrangemovementswithintheforagingarea(seebelowforthedetectionofforagingareas).
3. AlltheremainingfixeshavebeenassignedtotheFcategory.
EachindividualtrackwassubsequentlyplottedinQgisforvisualinspection.Whenabirdwasrestingforseveralhourssittingontheseasurface,acharacteristicpatternoffixesresulted,duetotheseacurrentand/orwinddriftingthebirdinaconstantdirection(Fayetetal.2015).Inthesecases,themeanvectorlength(Batschelet,1981)forasetofRfixes,computedaveragingthedirectionofmovementbetween two subsequent fixes,was greater than0.90. Therefore,the mean vector length was used as a criterion when a visual inspec-tionrevealedpossibleinconsistenciesinthefixclassassignment.IfthemeanvectorlengthofconsecutiveFfixeswasgreaterthan0.90,thefixeswerere-assignedtotheRcategory.Conversely,ifthemeanvectorlengthofconsecutiveRfixeswassmallerthan0.75,thefixeswerere-assignedtotheFcategory.Foreachtrack,thefirstfixes(1–3)werenotclassifiablewiththeabovecriteria,andtherefore,theyhavebeenexcludedfromtheanalysis.
Thetrackssampledatonefixevery10minwereresampledatonefixevery20minandused,togetherwiththeother20minsamplingratetracks,toperformadensitykernelanalysiswiththoselocations(n =27tracks).ThecoreforagingdistributionswerecalculatedonthebasisofthedistributionofFfixes.ThedensityofthedistributionofFfixeswasmodeledusingthefixedkerneltechnique(Worton,1989)available inR-packageadehabitatHR 4.15 (Calenge,2006).Thead-hocbandwidth for thesmoothingparameter (had- hoc)wasselectedbysequentiallyreducingthereferencebandwidthofthesmoothingfactor(href,i.e.,theoptimalbandwidthundertheassumptionofbi-variatenormality)in0.10incrementsandchoosingthesmallestin-crementofhrefthat:(i)resultedinacontiguousK95%isopleth,and(ii) containedno lacunawithinK95% (Kie,2013).CoreareaswereidentifiedbyapplyingtheAreaIndependentMethoddevelopedbySeamanandPowell(1990).Themethoddividestherangeinareasofhighandlowfixdensityusinganobjectivecriterionwhichisbasedon a graphical representation of the range area in relation to thedensity of the considered fixes. In thisway, it is possible to iden-tifythedividingpointbetweenhigh-andlow-densityareas,asthepointwheretheplotismaximallydistantfromastraightlineofslope+1thatrepresentsadistributionofrandomuse.Weperformedtheanalysis considering steps =5%calculatingsubsequentrangeareasizes using adehabitatHR. On the basis of the point of maximumdistance,wedefinedthecoreareaswithadifferentpercentageofvolumecontourperindividual(median:55%;IQR:50%–55%volumecontours).
Weused theR fixesdistribution toassesswhetherandwhichof the tracked birds rested in areas proximal (within 5 km radius;thedistance range from the colonywithinwhich rafts areusuallyobserved) to yelkouan shearwaters colonies other than those ofTavolaraarchipelagoandadjacentCapeFigari area that areall lo-catedonenexttoanother.A5kmradiusbufferwascreatedwith

6 of 18 | PEZZO Et al.
Qgis aroundeach yelkouan shearwater colony known in Italy andFrance (Baccettietal.,2009;Cadiouetal.,2004).We intersectedthe5kmradiusbufferwiththeRfixesdistribution,inordertocountwhichcolonieswereapproachedbythetrackedbirds. Inaddition,weidentifiedtheareasmostlyusedforrestingbythetrackedbirds.Todoso,a10kmhexagonalgridwascreatedinQgis;foreachindi-vidual,wecomputedthepercentageofRfixesfallingineachhexa-gon.Foreachtrack,thenumberofRfixesineachcellwascomputedand categorized in three categories depending on the proportion ofRfixescontainedinacell: (i)numberoffixesbelowthemedianvalue;(ii)numberoffixesrangingbetweenthemedianvalueandthethirdpercentile;(iii)valueshigherthanthethirdpercentile.Foreachcell,wesummedthescoresobtainedfromallthetracksinordertoidentifyareaswiththehighestscore,thatis,thosemostlikelyfre-quentedbyrestingbirds.
Foreachfixof thetracks, thedistancefromthenearestcoastwascomputed.Inordertoassesswhetherthebirdshadapreferenceforstayingneartoorfarfromthecoast,foreachbirdthepercentageoffixesofeachconsideredcategory(T,R,F)locatedwithin20kmandfurtherthan20kmwascomparedbyWilcoxontestforpaireddata.Whenmorethanonetracksperbirdwereavailable,theindi-vidual mean values were considered.
Eachlocationwasclassifiedas“day”or“night”accordingtotheircorrespondingtimeofnauticaldawnandduskobtainedusingthe"R-package"suncalc 0.5.0(Thieurmel&Elmarhraoui,2019).Differencesin the activity between day and night were tested by applying the Wilcoxontestforpaireddatatothe individualpercentageofeachbehavioralcategory.Asbefore,whenmorethanonetrackperbirdwereavailable,theindividualmeanvalueswereconsidered.
Toanalyzeforaginghabitatselection(sensuManlyetal.,2002)by yelkouan shearwaters, we used remote sensing data to quan-tify Bathymetry (ETOPOGlobal ReliefModel, NOAA) andOceanProductivity(MODIS)ataresolutionof1.6and9km,respectively.Productivitydata(gC·m- 2·day- 1)usedwereaveragedovereightdays(octads)andwereasmuchaspossiblecontemporarytoeachforag-ing trip.Given the resolutionof the remote sensingdata,useandavailability were estimated at the individual and population level,respectively(firstorderresourceselection;Meyer&Thuiller,2006).Onthebasisoftheproductivityrasterdata,agridwith9kmsquaredcellwascreated.ForeachcellwithatleastoneFfix,fivecellswererandomlysampledwithinthepopulation95%range(calculatedwiththe same kernel approach described above) by considering the Ffixesofalltracks.Thisprocesswasrepeatedforallavailableforag-ingtrips.Forallusedandrandomlocations,weextractedthevaluesofbathymetry,distancefromthecolonysitetothecenterofthecell,andproductivity.Duetothelowsamplesize,theyearoftaggingwasnotincludedintheanalysis.Dataexplorationwascarriedoutfollow-ingtheprotocoldescribedinZuurandIeno(2016).
The resource selection function (Manly et al., 2002) was cal-culated with a Generalized Linear Mixed Model with a binomialerrordistribution,andbird IDandtrack ID (nestedwithinbird ID)asrandominterceptsbymeansoftheR-packageglmmTMP 1.0.2.1 (Brooksetal.,2017).Thevariablesconsideredinthefullmodelwere
asfollows:seaproductivity(PROD,inversetransformed),bathyme-try(BATHY),distancefromthecolony(DCOL),andthereproductivestage(STAGE,two-levelsfactor: incubationandchickrearing).Wehypothesized that the probability of selection usewas higher forcellswith higher PROD, and lowerBATHY andDCOL.We testedwhethertheincubationperiodmodifiedthewaythebirdsusedtheresourcesbyincludingthesecond-orderinteractionsSTAGE:DCOL,STAGE:BATHY,STAGE:PROD.Thefixedpartofthemodelwassim-plifiedbymeansof theAkaike InformationCriterioncorrectedforsmallsamplesize(AICc,Burnham&Anderson,2002),consideringallthemodelsbetweenthefullmodelandthemodelwhichincludedallmaineffects.SignificancewastestedbymeansofthetypeIIWaldχ2 testusingtheR-packagecar 3.0– 10(Fox&Weisberg,2019).Modelfit,overdispersion,collinearity,andspatialautocorrelationofthere-sidualswerecheckedbeforeusingthefinalmodelforinferencebymeansof theR-packagesDHARMa 0.3.3.0 (Hartig, 2020) andper-formance 0.7.0(Lüdeckeetal.,2020).ThemarginalR²(mR²),whichrepresentsthevarianceexplainedbyfixedfactorsonly(Nakagawa& Schielzeth, 2013), was calculated using the R-package MuMIn 1.43.17 (Bartoń, 2020). The performance of the finalmodelswasalsoevaluatedusingtheAreaUndertheCurve(AUC)generatedbytheReceiverOperatingCharacteristic (ROC;Fielding&Bell,1997;Pearce&Ferrier,2000)bymeansoftheR-packageROCR 1.0– 7(Singetal.,2005).AllcalculationswereperformedusingR4.0.4(RCoreTeam,2021).
Tofurtherinvestigatebirdforagingstrategiesinthetwophasesofthenestingperiod,weanalyzedhowseaproductivityvariedincells locatedatdifferentdistancefromthecolonyoverthestudyperiod. Inthisanalysis,weconsideredonlythecellswithat leastoneFfixduringthewholestudyperiod, that is, thecellsusedatleastoncebytrackedbirdsforforaging,andwithnomissingdataonproductivity(n =333).Toreducesamplingbias,weexcludedtheyearswherebirdsweretrackedduringincubationorchickrearingonly(i.e.,2011and2015).Theavailablecellswerethenresampledtoavoidincludingadjacentcellsintheanalysis,inordertoreducespatial autocorrelation, by means of the R-package spThin 0.2.0 (Aiello-Lammens et al., 2015). The number of cells considered intheanalysiswas91.DataweremodeledbymeansofLinearMixedModels consideringproductivity (inverse transformed) as depen-dent variable and cell ID as random intercept. The independentvariablesconsideredinthemodelwereasfollows:octad(OCTAD,5-level factor coded using the first Julian day of the octad: 81,89,105,161,169),distance fromthecolony (DCOL),bathymetry(BATHY) and the second-order interactions OCTAD:DCOL andOCTAD:BATHY. Based on observed productivity variations, wehypothesized that theproductivityof cellsnearest to the colonydecreases over timemuchmore strongly than that of cellsmoredistantfromthecolonysite,thusinducingbirdstoincreasethefre-quency of long foraging trips. The packages and the proceduresused to check model assumptions, to test significance, and toevaluatemodelfitwerethesameasdescribedabove.Throughoutthe text,meansare reportedalongwith their standarddeviation(mean± SD)unlessotherwisespecified.

| 7 of 18PEZZO Et al.
3 | RESULTS
Outofatotalof43captureeventsinsubsequentyearsand60GPSdeployed,netofthebirdsnotrecaptured(n =7),recapturedwith-outlogger(n =14),orrecapturedwithimproperlyfunctioninglog-gers (n = 5),we obtained34 foraging trips from21 birds trackedduring incubation (March-April;n =21trips)andchick-rearingpe-riod (May-June;n = 13 trips).Nobreeding failureswere recordedbetweencaptureandrecapture.Table1reports thedetailsof thetracks recorded, such aswhether theywere complete (the entirejourneyfromthecolonytotheforagingareaandbackwasrecorded),or interrupted(thepowerrunoutbeforethebirdhomedaftertheforagingtrip),andtracksexcludedfromtheanalysisduetoboththelowsamplingrate(1fixevery60min)andlownumberoffixes(<45).
Birds returned to the colony after 6.5 days (median; range1–8 days) during the incubation period (n = 12 complete tracksfrom11birds)andafter2.0days(median;range1–10days)during
the chick-rearing period (n = 11 complete tracks from 6 birds).Considering only the complete tracks included in the analysis (9tracks from9 birds in the incubation period, and 8 tracks from5birdsduringthechick-rearingperiod;seeTable1),itappearedthatduring the incubation and chick-rearing periods, the mean tracklengthwas1,030±140kmand733±225km,respectively,reachingameandistancefromthecolonyof193±24kmand181±53km,respectively.
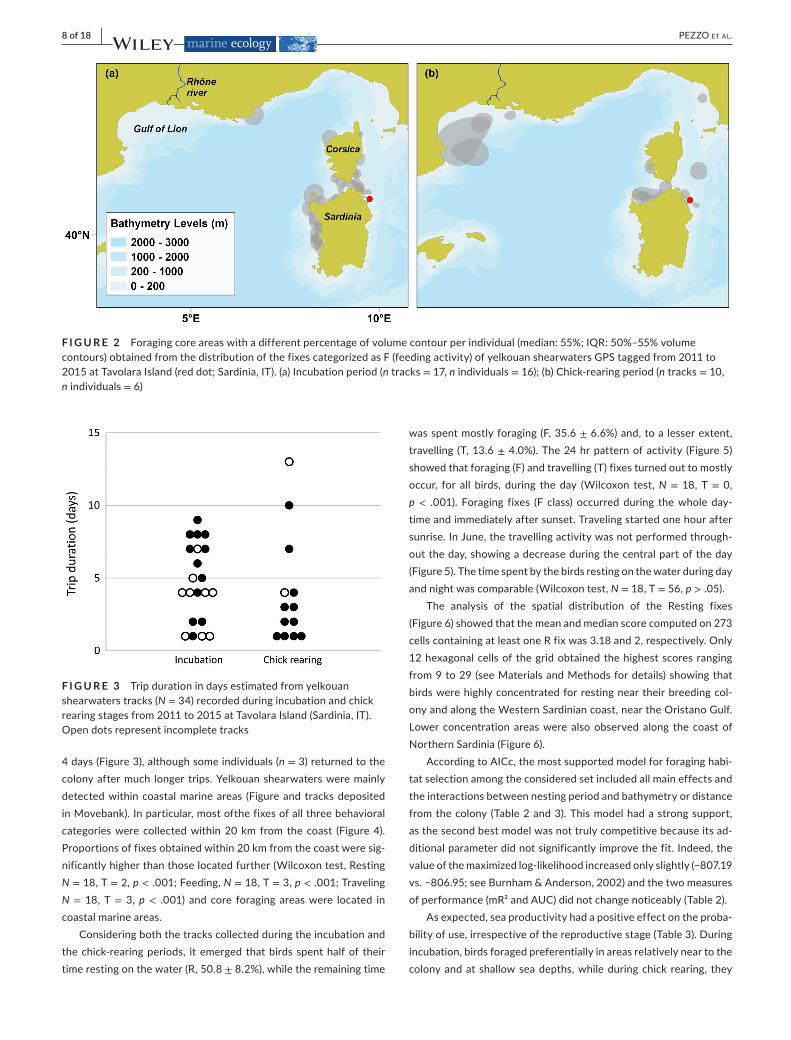
Duringtheincubationperiod,foragingtripsweremostlyconcen-tratedinproximityoftheWesternandNortherncoastsofSardiniaand South Corsica (Figure 1a). During thechick-earing period,WesternSardiniawasalmostentirelydeserted,andnewcorefeed-ingareaswereusedintheFrench/SpanishwatersoftheGulfofLion(maxdistance579Km).AnumberofbirdscontinuedtofeedalongNorthSardiniaandSouthCorsica(Figures1band2).Consideringalltracks,duringtheincubationforagingtripsrangedfrom1to9days,whileduringchick-rearingperiod,mostforagingtripslastedlessthan
F I G U R E 1 TracksofyelkouanshearwatersGPStaggedfrom2011to2015atTavolaraIsland,Sardinia,IT(whitestar).Theblueshadesshowseanetprimaryproductivity(expressedasmgC·m−2·day−1)groupedaccordingtoOctads(8daysperiods)in2011–2015.Eachpanelcontainsthetracksobtainedintheoctadforwhichseaproductivitywascalculatedandincludedintheanalysis(seeTable1).TindicatesthetrackID;theapicalletters(atod)denotetracksfromthesameindividual.Thereproductivestage(a,incubation;b,chickrearing)ofbirdsatthetimeofrecordingiswrittenineachpanel
8 of 18 | PEZZO Et al.
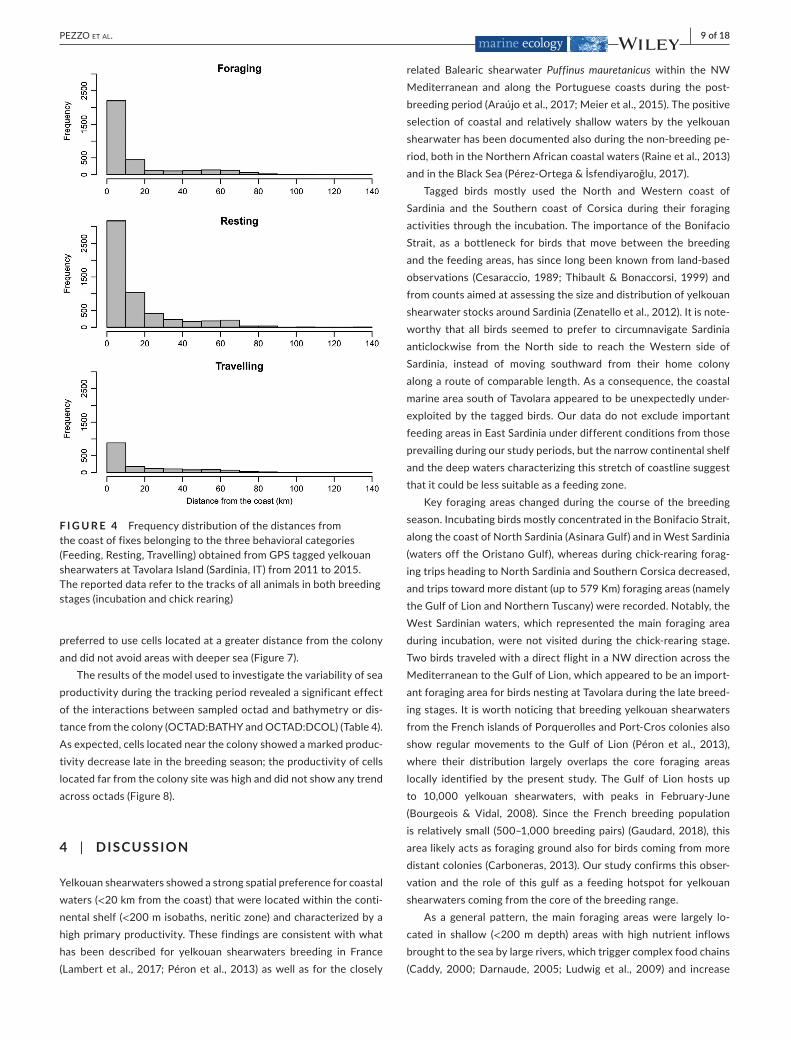
4days(Figure3),althoughsomeindividuals(n =3)returnedtothecolonyaftermuch longertrips.Yelkouanshearwatersweremainlydetectedwithincoastalmarineareas (Figureand tracksdepositedinMovebank).Inparticular,mostofthefixesofallthreebehavioralcategorieswere collectedwithin20km from the coast (Figure4).Proportionsoffixesobtainedwithin20kmfromthecoastweresig-nificantlyhigherthanthoselocatedfurther(Wilcoxontest,RestingN =18,T=2,p <.001;Feeding,N =18,T=3,p <.001;TravelingN = 18, T= 3,p < .001) and core foraging areaswere located incoastal marine areas.
Consideringboththetrackscollectedduringtheincubationandthechick-rearingperiods, itemerged thatbirds spenthalfof theirtimerestingonthewater(R,50.8±8.2%),whiletheremainingtime
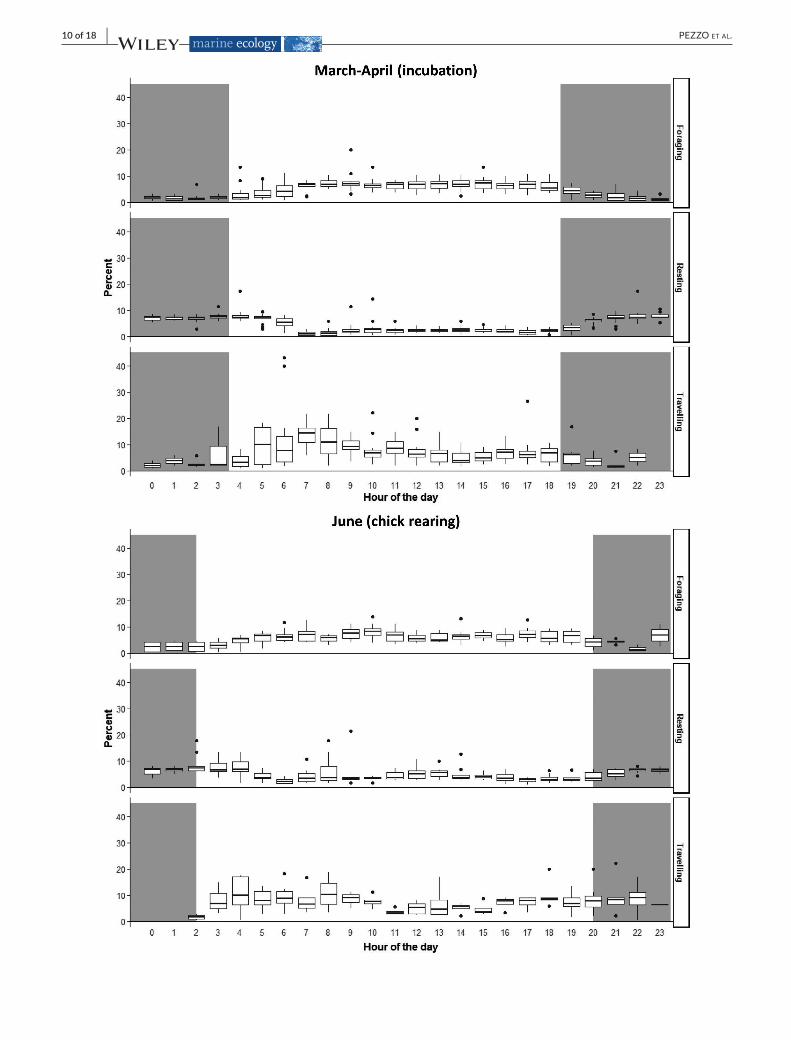
wasspentmostlyforaging(F,35.6±6.6%)and,toa lesserextent,travelling (T,13.6±4.0%).The24hrpatternofactivity (Figure5)showedthatforaging(F)andtravelling(T)fixesturnedouttomostlyoccur, for all birds, during the day (Wilcoxon test,N = 18, T= 0,p < .001). Foraging fixes (F class) occurredduring thewhole day-timeandimmediatelyaftersunset.Travelingstartedonehouraftersunrise.InJune,thetravellingactivitywasnotperformedthrough-outtheday,showingadecreaseduringthecentralpartoftheday(Figure5).Thetimespentbythebirdsrestingonthewaterduringdayandnightwascomparable(Wilcoxontest,N =18,T=56,p >.05).
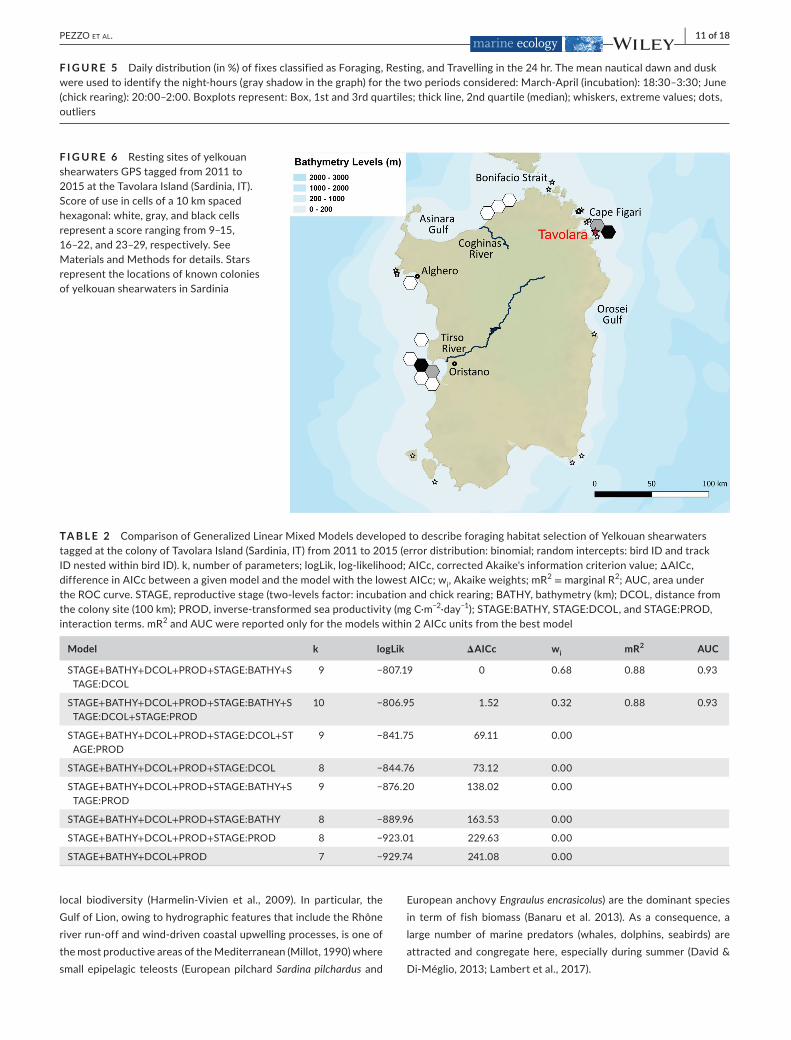
The analysis of the spatial distribution of the Resting fixes(Figure6)showedthatthemeanandmedianscorecomputedon273cellscontainingatleastoneRfixwas3.18and2,respectively.Only12hexagonalcellsof thegridobtainedthehighestscoresrangingfrom9to29(seeMaterialsandMethodsfordetails)showingthatbirdswerehighlyconcentratedforrestingneartheirbreedingcol-onyandalongtheWesternSardiniancoast,neartheOristanoGulf.Lower concentration areaswere alsoobserved along the coast ofNorthernSardinia(Figure6).
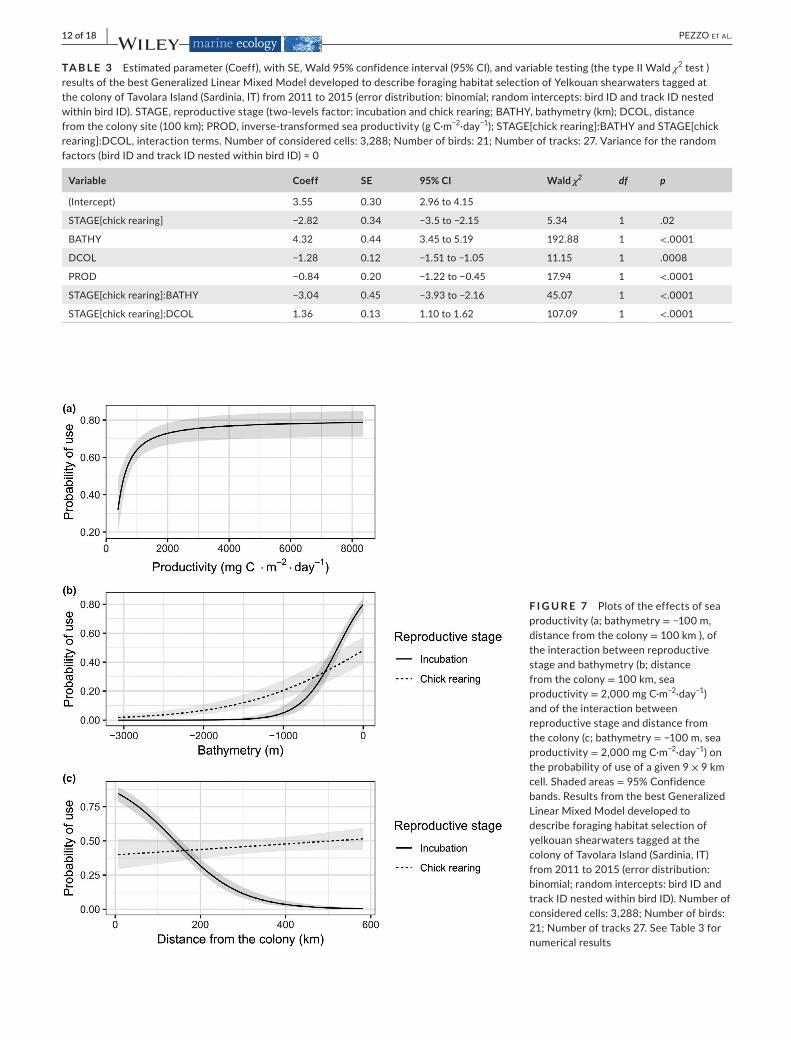
AccordingtoAICc,themostsupportedmodelforforaginghabi-tatselectionamongtheconsideredsetincludedallmaineffectsandthe interactions between nesting period and bathymetry or distance fromthecolony (Table2and3).Thismodelhadastrongsupport,as the second best model was not truly competitive because its ad-ditionalparameterdidnotsignificantlyimprovethefit.Indeed,thevalueofthemaximizedlog-likelihoodincreasedonlyslightly(−807.19vs.−806.95;seeBurnham&Anderson,2002)andthetwomeasuresofperformance(mR²andAUC)didnotchangenoticeably(Table2).
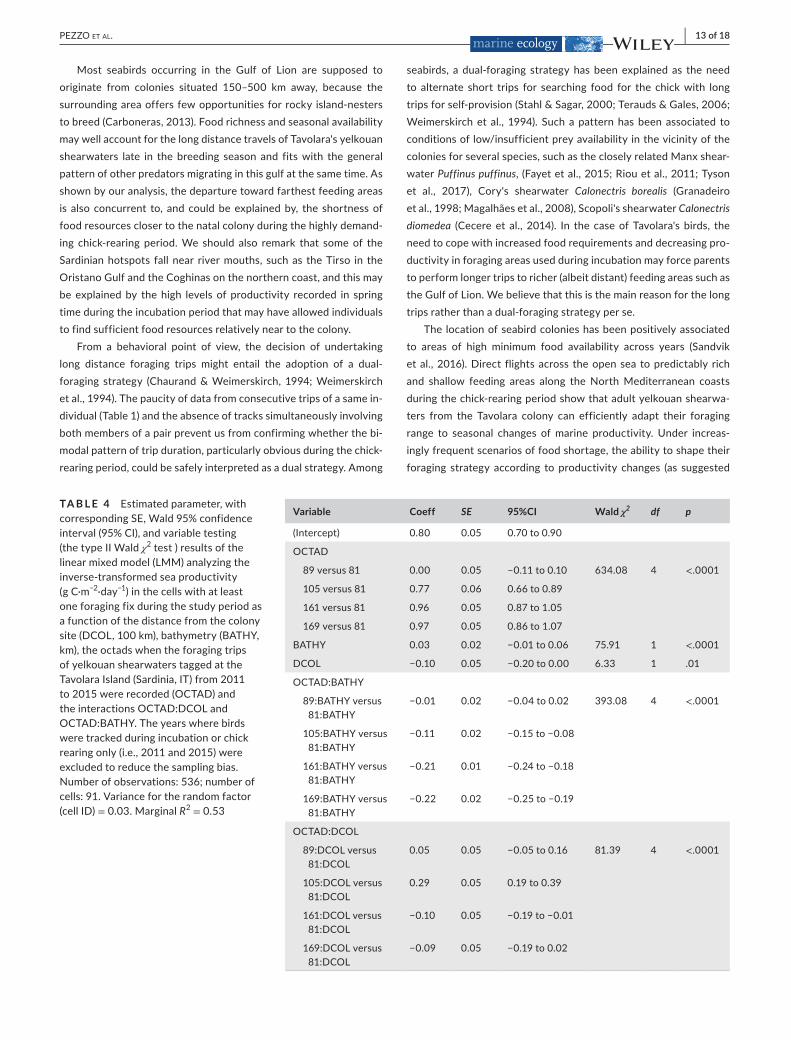
Asexpected,seaproductivityhadapositiveeffectontheproba-bilityofuse,irrespectiveofthereproductivestage(Table3).Duringincubation,birdsforagedpreferentiallyinareasrelativelyneartothecolonyandat shallowseadepths,whileduringchick rearing, they
F I G U R E 2 Foragingcoreareaswithadifferentpercentageofvolumecontourperindividual(median:55%;IQR:50%–55%volumecontours)obtainedfromthedistributionofthefixescategorizedasF(feedingactivity)ofyelkouanshearwatersGPStaggedfrom2011to2015atTavolaraIsland(reddot;Sardinia,IT).(a)Incubationperiod(ntracks=17,n individuals =16);(b)Chick-rearingperiod(ntracks=10,n individuals =6)
F I G U R E 3 Tripdurationindaysestimatedfromyelkouanshearwaterstracks(N =34)recordedduringincubationandchickrearingstagesfrom2011to2015atTavolaraIsland(Sardinia,IT).Opendotsrepresentincompletetracks
| 9 of 18PEZZO Et al.
preferredtousecellslocatedatagreaterdistancefromthecolonyanddidnotavoidareaswithdeepersea(Figure7).
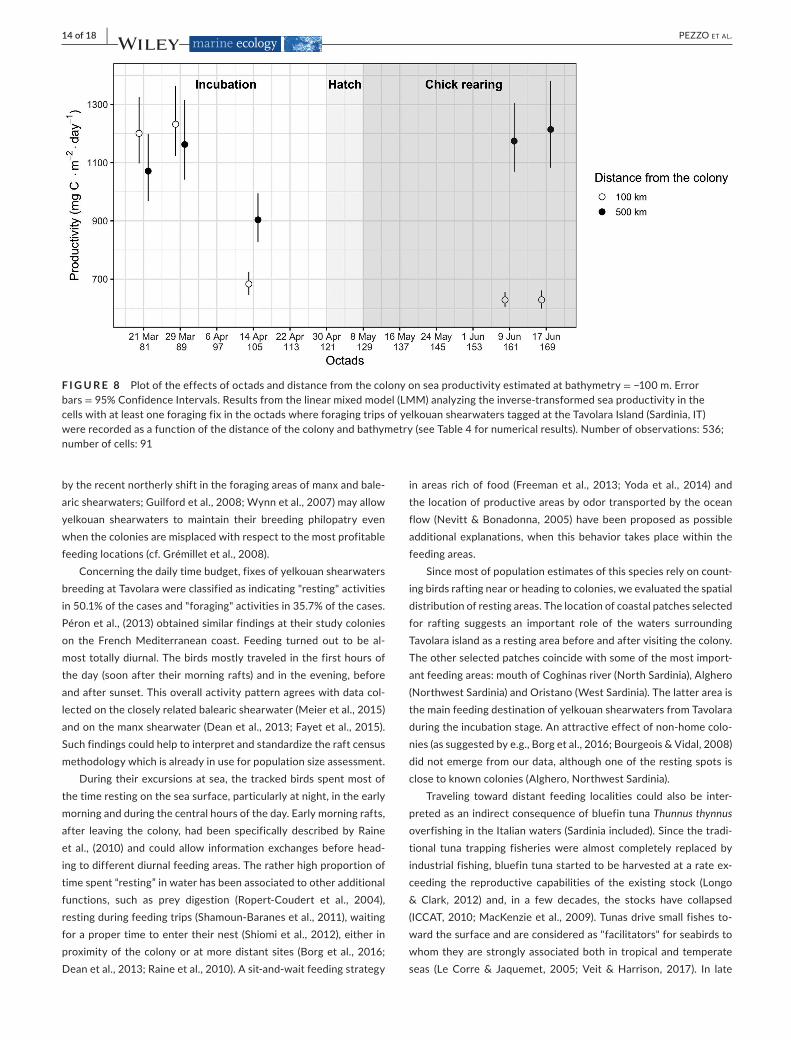
Theresultsofthemodelusedtoinvestigatethevariabilityofseaproductivityduringthetrackingperiodrevealedasignificanteffectoftheinteractionsbetweensampledoctadandbathymetryordis-tancefromthecolony(OCTAD:BATHYandOCTAD:DCOL)(Table4).Asexpected,cellslocatednearthecolonyshowedamarkedproduc-tivitydecreaselateinthebreedingseason;theproductivityofcellslocatedfarfromthecolonysitewashighanddidnotshowanytrendacrossoctads(Figure8).
4 | DISCUSSION
Yelkouanshearwatersshowedastrongspatialpreferenceforcoastalwaters(<20kmfromthecoast)thatwerelocatedwithintheconti-nentalshelf(<200misobaths,neriticzone)andcharacterizedbyahighprimaryproductivity.Thesefindingsareconsistentwithwhathas been described for yelkouan shearwaters breeding in France(Lambertetal.,2017;Péronetal.,2013)aswellas for theclosely
related Balearic shearwater Puffinus mauretanicus within the NWMediterranean and along the Portuguese coasts during the post- breedingperiod(Araújoetal.,2017;Meieretal.,2015).Thepositiveselectionof coastal and relatively shallowwatersby theyelkouanshearwater has been documented also during the non- breeding pe-riod,bothintheNorthernAfricancoastalwaters(Raineetal.,2013)andintheBlackSea(Pérez-Ortega&İsfendiyaroğlu,2017).
Tagged birds mostly used the North and Western coast ofSardinia and the Southern coast of Corsica during their foragingactivitiesthroughtheincubation.TheimportanceoftheBonifacioStrait, as a bottleneck for birds thatmove between the breedingandthefeedingareas,hassincelongbeenknownfromland-basedobservations (Cesaraccio, 1989; Thibault&Bonaccorsi, 1999) andfromcountsaimedatassessingthesizeanddistributionofyelkouanshearwaterstocksaroundSardinia(Zenatelloetal.,2012).Itisnote-worthy that all birds seemed to prefer to circumnavigate Sardiniaanticlockwise from the North side to reach the Western side ofSardinia, instead of moving southward from their home colonyalongarouteofcomparablelength.Asaconsequence,thecoastalmarineareasouthofTavolaraappearedtobeunexpectedlyunder-exploited by the tagged birds. Our data do not exclude important feedingareasinEastSardiniaunderdifferentconditionsfromthoseprevailingduringourstudyperiods,butthenarrowcontinentalshelfandthedeepwaterscharacterizingthisstretchofcoastlinesuggestthatitcouldbelesssuitableasafeedingzone.
Key foragingareas changedduring the courseof thebreedingseason.IncubatingbirdsmostlyconcentratedintheBonifacioStrait,alongthecoastofNorthSardinia(AsinaraGulf)andinWestSardinia(watersofftheOristanoGulf),whereasduringchick-rearingforag-ingtripsheadingtoNorthSardiniaandSouthernCorsicadecreased,andtripstowardmoredistant(upto579Km)foragingareas(namelytheGulfofLionandNorthernTuscany)wererecorded.Notably,theWest Sardinianwaters,which represented themain foraging areaduring incubation,werenotvisitedduring thechick-rearing stage.TwobirdstraveledwithadirectflightinaNWdirectionacrosstheMediterraneantotheGulfofLion,whichappearedtobeanimport-antforagingareaforbirdsnestingatTavolaraduringthelatebreed-ingstages.ItisworthnoticingthatbreedingyelkouanshearwatersfromtheFrenchislandsofPorquerollesandPort-Croscoloniesalsoshow regularmovements to theGulf of Lion (Péron et al., 2013),where their distribution largely overlaps the core foraging areaslocally identified by the present study. TheGulf of Lion hosts upto 10,000 yelkouan shearwaters, with peaks in February-June(Bourgeois & Vidal, 2008). Since the French breeding populationis relativelysmall (500–1,000breedingpairs) (Gaudard,2018), thisarealikelyactsasforaginggroundalsoforbirdscomingfrommoredistantcolonies(Carboneras,2013).Ourstudyconfirmsthisobser-vation and the role of this gulf as a feeding hotspot for yelkouanshearwaterscomingfromthecoreofthebreedingrange.
As a general pattern, themain foraging areaswere largely lo-cated in shallow (<200m depth) areaswith high nutrient inflowsbroughttotheseabylargerivers,whichtriggercomplexfoodchains(Caddy, 2000;Darnaude, 2005; Ludwig et al., 2009) and increase
F I G U R E 4 Frequencydistributionofthedistancesfromthecoastoffixesbelongingtothethreebehavioralcategories(Feeding,Resting,Travelling)obtainedfromGPStaggedyelkouanshearwatersatTavolaraIsland(Sardinia,IT)from2011to2015.Thereporteddatarefertothetracksofallanimalsinbothbreedingstages(incubationandchickrearing)
10 of 18 | PEZZO Et al.
| 11 of 18PEZZO Et al.
local biodiversity (Harmelin-Vivien et al., 2009). In particular, theGulfofLion,owingtohydrographicfeaturesthatincludetheRhôneriverrun-offandwind-drivencoastalupwellingprocesses,isoneofthemostproductiveareasoftheMediterranean(Millot,1990)wheresmallepipelagic teleosts (EuropeanpilchardSardina pilchardus and
EuropeananchovyEngraulus encrasicolus)arethedominantspeciesin termof fish biomass (Banaru et al. 2013).As a consequence, alarge number ofmarine predators (whales, dolphins, seabirds) areattractedandcongregatehere,especiallyduringsummer (David&Di-Méglio,2013;Lambertetal.,2017).
F I G U R E 5 Dailydistribution(in%)offixesclassifiedasForaging,Resting,andTravellinginthe24hr.Themeannauticaldawnandduskwereusedtoidentifythenight-hours(grayshadowinthegraph)forthetwoperiodsconsidered:March-April(incubation):18:30–3:30;June(chickrearing):20:00–2:00.Boxplotsrepresent:Box,1stand3rdquartiles;thickline,2ndquartile(median);whiskers,extremevalues;dots,outliers
F I G U R E 6 RestingsitesofyelkouanshearwatersGPStaggedfrom2011to2015attheTavolaraIsland(Sardinia,IT).Scoreofuseincellsofa10kmspacedhexagonal:white,gray,andblackcellsrepresentascorerangingfrom9–15,16–22,and23–29,respectively.SeeMaterialsandMethodsfordetails.StarsrepresentthelocationsofknowncoloniesofyelkouanshearwatersinSardinia
TA B L E 2 ComparisonofGeneralizedLinearMixedModelsdevelopedtodescribeforaginghabitatselectionofYelkouanshearwaterstaggedatthecolonyofTavolaraIsland(Sardinia,IT)from2011to2015(errordistribution:binomial;randomintercepts:birdIDandtrackIDnestedwithinbirdID).k,numberofparameters;logLik,log-likelihood;AICc,correctedAkaike'sinformationcriterionvalue;ΔAICc,differenceinAICcbetweenagivenmodelandthemodelwiththelowestAICc;wi,Akaikeweights;mR
2 = marginal R2;AUC,areaundertheROCcurve.STAGE,reproductivestage(two-levelsfactor:incubationandchickrearing;BATHY,bathymetry(km);DCOL,distancefromthecolonysite(100km);PROD,inverse-transformedseaproductivity(mgC·m−2·day−1);STAGE:BATHY,STAGE:DCOL,andSTAGE:PROD,interaction terms. mR2andAUCwerereportedonlyforthemodelswithin2AICcunitsfromthebestmodel
Model k logLik ΔAICc wi mR2 AUC
STAGE+BATHY+DCOL+PROD+STAGE:BATHY+STAGE:DCOL
9 −807.19 0 0.68 0.88 0.93
STAGE+BATHY+DCOL+PROD+STAGE:BATHY+STAGE:DCOL+STAGE:PROD
10 −806.95 1.52 0.32 0.88 0.93
STAGE+BATHY+DCOL+PROD+STAGE:DCOL+STAGE:PROD
9 −841.75 69.11 0.00
STAGE+BATHY+DCOL+PROD+STAGE:DCOL 8 −844.76 73.12 0.00
STAGE+BATHY+DCOL+PROD+STAGE:BATHY+STAGE:PROD
9 −876.20 138.02 0.00
STAGE+BATHY+DCOL+PROD+STAGE:BATHY 8 −889.96 163.53 0.00
STAGE+BATHY+DCOL+PROD+STAGE:PROD 8 −923.01 229.63 0.00
STAGE+BATHY+DCOL+PROD 7 −929.74 241.08 0.00
12 of 18 | PEZZO Et al.
TA B L E 3 Estimatedparameter(Coeff),withSE,Wald95%confidenceinterval(95%CI),andvariabletesting(thetypeIIWaldχ2test)resultsofthebestGeneralizedLinearMixedModeldevelopedtodescribeforaginghabitatselectionofYelkouanshearwaterstaggedatthecolonyofTavolaraIsland(Sardinia,IT)from2011to2015(errordistribution:binomial;randomintercepts:birdIDandtrackIDnestedwithinbirdID).STAGE,reproductivestage(two-levelsfactor:incubationandchickrearing;BATHY,bathymetry(km);DCOL,distancefromthecolonysite(100km);PROD,inverse-transformedseaproductivity(gC·m−2·day−1);STAGE[chickrearing]:BATHYandSTAGE[chickrearing]:DCOL,interactionterms.Numberofconsideredcells:3,288;Numberofbirds:21;Numberoftracks:27.Variancefortherandomfactors(birdIDandtrackIDnestedwithinbirdID)≈0
Variable Coeff SE 95% CI Wald χ2 df p
(Intercept) 3.55 0.30 2.96to4.15
STAGE[chickrearing] −2.82 0.34 −3.5to−2.15 5.34 1 .02
BATHY 4.32 0.44 3.45to5.19 192.88 1 <.0001
DCOL −1.28 0.12 −1.51to−1.05 11.15 1 .0008
PROD −0.84 0.20 −1.22to−0.45 17.94 1 <.0001
STAGE[chickrearing]:BATHY −3.04 0.45 −3.93to−2.16 45.07 1 <.0001
STAGE[chickrearing]:DCOL 1.36 0.13 1.10 to 1.62 107.09 1 <.0001
F I G U R E 7 Plotsoftheeffectsofseaproductivity(a;bathymetry=−100m,distancefromthecolony=100km),ofthe interaction between reproductive stageandbathymetry(b;distancefromthecolony=100km,seaproductivity =2,000mgC·m−2·day−1)andoftheinteractionbetweenreproductivestageanddistancefromthecolony(c;bathymetry=−100m,seaproductivity =2,000mgC·m−2·day−1)ontheprobabilityofuseofagiven9×9kmcell. Shaded areas =95%Confidencebands.ResultsfromthebestGeneralizedLinearMixedModeldevelopedtodescribeforaginghabitatselectionofyelkouanshearwaterstaggedatthecolonyofTavolaraIsland(Sardinia,IT)from2011to2015(errordistribution:binomial; random intercepts: bird ID and trackIDnestedwithinbirdID).Numberofconsideredcells:3,288;Numberofbirds:21;Numberoftracks27.SeeTable3fornumerical results
| 13 of 18PEZZO Et al.
Most seabirds occurring in the Gulf of Lion are supposed tooriginate from colonies situated 150–500 km away, because thesurroundingareaoffersfewopportunitiesforrocky island-nesterstobreed(Carboneras,2013).FoodrichnessandseasonalavailabilitymaywellaccountforthelongdistancetravelsofTavolara'syelkouanshearwaters late in the breeding season and fitswith the generalpatternofotherpredatorsmigratinginthisgulfatthesametime.Asshownbyouranalysis,thedeparturetowardfarthestfeedingareasis also concurrent to, and couldbeexplainedby, the shortnessoffoodresourcesclosertothenatalcolonyduringthehighlydemand-ing chick-rearing period.We should also remark that someof theSardinianhotspots fallnear rivermouths, suchas theTirso in theOristanoGulfandtheCoghinasonthenortherncoast,andthismaybeexplainedby thehigh levels of productivity recorded in springtime during the incubation period that may have allowed individuals tofindsufficientfoodresourcesrelativelyneartothecolony.
From a behavioral point of view, the decision of undertakinglong distance foraging trips might entail the adoption of a dual-foraging strategy (Chaurand&Weimerskirch, 1994;Weimerskirchetal.,1994).Thepaucityofdatafromconsecutivetripsofasamein-dividual(Table1)andtheabsenceoftrackssimultaneouslyinvolvingbothmembersofapairpreventusfromconfirmingwhetherthebi-modalpatternoftripduration,particularlyobviousduringthechick-rearingperiod,couldbesafelyinterpretedasadualstrategy.Among
seabirds, a dual-foraging strategyhasbeenexplained as theneedto alternate short trips for searching food for the chickwith longtripsforself-provision(Stahl&Sagar,2000;Terauds&Gales,2006;Weimerskirchet al., 1994). Suchapatternhasbeenassociated toconditionsoflow/insufficientpreyavailabilityinthevicinityofthecoloniesforseveralspecies,suchasthecloselyrelatedManxshear-water Puffinus puffinus, (Fayetetal.,2015;Riouetal.,2011;Tysonet al., 2017), Cory's shearwater Calonectris borealis (Granadeiroetal.,1998;Magalhãesetal.,2008),Scopoli'sshearwaterCalonectris diomedea (Cecereet al., 2014). In the caseofTavolara's birds, theneedtocopewithincreasedfoodrequirementsanddecreasingpro-ductivityinforagingareasusedduringincubationmayforceparentstoperformlongertripstoricher(albeitdistant)feedingareassuchastheGulfofLion.Webelievethatthisisthemainreasonforthelongtripsratherthanadual-foragingstrategyperse.
The locationofseabirdcolonieshasbeenpositivelyassociatedto areas of high minimum food availability across years (Sandviketal.,2016).Direct flightsacross theopensea topredictably richand shallow feeding areas along the NorthMediterranean coastsduringthechick-rearingperiodshowthatadultyelkouanshearwa-ters from the Tavolara colony can efficiently adapt their foragingrange to seasonal changes ofmarine productivity. Under increas-inglyfrequentscenariosoffoodshortage,theabilitytoshapetheirforaging strategy according toproductivity changes (as suggested
Variable Coeff SE 95%CI Wald χ2 df p
(Intercept) 0.80 0.05 0.70to0.90
OCTAD
89 versus 81 0.00 0.05 −0.11to0.10 634.08 4 <.0001
105versus81 0.77 0.06 0.66 to 0.89
161 versus 81 0.96 0.05 0.87to1.05
169 versus 81 0.97 0.05 0.86to1.07
BATHY 0.03 0.02 −0.01to0.06 75.91 1 <.0001
DCOL −0.10 0.05 −0.20to0.00 6.33 1 .01
OCTAD:BATHY
89:BATHYversus81:BATHY
−0.01 0.02 −0.04to0.02 393.08 4 <.0001
105:BATHYversus81:BATHY
−0.11 0.02 −0.15to−0.08
161:BATHYversus81:BATHY
−0.21 0.01 −0.24to−0.18
169:BATHYversus81:BATHY
−0.22 0.02 −0.25to−0.19
OCTAD:DCOL
89:DCOLversus81:DCOL
0.05 0.05 −0.05to0.16 81.39 4 <.0001
105:DCOLversus81:DCOL
0.29 0.05 0.19 to 0.39
161:DCOLversus81:DCOL
−0.10 0.05 −0.19to−0.01
169:DCOLversus81:DCOL
−0.09 0.05 −0.19to0.02
TA B L E 4 Estimatedparameter,withcorrespondingSE,Wald95%confidenceinterval(95%CI),andvariabletesting(thetypeIIWaldχ2test)resultsofthelinearmixedmodel(LMM)analyzingtheinverse-transformedseaproductivity(gC·m−2·day−1)inthecellswithatleastoneforagingfixduringthestudyperiodasafunctionofthedistancefromthecolonysite(DCOL,100km),bathymetry(BATHY,km),theoctadswhentheforagingtripsofyelkouanshearwaterstaggedattheTavolaraIsland(Sardinia,IT)from2011to2015wererecorded(OCTAD)andtheinteractionsOCTAD:DCOLandOCTAD:BATHY.Theyearswherebirdsweretrackedduringincubationorchickrearingonly(i.e.,2011and2015)wereexcluded to reduce the sampling bias. Numberofobservations:536;numberofcells:91.Variancefortherandomfactor(cellID)= 0.03. Marginal R2 =0.53
14 of 18 | PEZZO Et al.
bytherecentnortherlyshiftintheforagingareasofmanxandbale-aricshearwaters;Guilfordetal.,2008;Wynnetal.,2007)mayallowyelkouan shearwaters to maintain their breeding philopatry evenwhenthecoloniesaremisplacedwithrespecttothemostprofitablefeedinglocations(cf.Grémilletetal.,2008).
Concerningthedailytimebudget,fixesofyelkouanshearwatersbreedingatTavolarawereclassifiedasindicating"resting"activitiesin50.1%ofthecasesand"foraging"activitiesin35.7%ofthecases.Péronetal.,(2013)obtainedsimilarfindingsattheirstudycolonieson the French Mediterranean coast. Feeding turned out to be al-mosttotallydiurnal.Thebirdsmostlytraveledinthefirsthoursoftheday(soonaftertheirmorningrafts)and intheevening,beforeandaftersunset.Thisoverallactivitypatternagreeswithdatacol-lectedonthecloselyrelatedbalearicshearwater(Meieretal.,2015)andonthemanxshearwater(Deanetal.,2013;Fayetetal.,2015).Suchfindingscouldhelptointerpretandstandardizetheraftcensusmethodologywhichisalreadyinuseforpopulationsizeassessment.
Duringtheirexcursionsatsea,thetrackedbirdsspentmostofthetimerestingontheseasurface,particularlyatnight,intheearlymorningandduringthecentralhoursoftheday.Earlymorningrafts,after leaving the colony, had been specifically described byRaineet al., (2010) andcouldallow informationexchangesbeforehead-ingtodifferentdiurnalfeedingareas.Theratherhighproportionoftimespent“resting”inwaterhasbeenassociatedtootheradditionalfunctions, such as prey digestion (Ropert-Coudert et al., 2004),restingduringfeedingtrips(Shamoun-Baranesetal.,2011),waitingforapropertimetoentertheirnest(Shiomietal.,2012),eitherinproximityof thecolonyoratmoredistantsites (Borgetal.,2016;Deanetal.,2013;Raineetal.,2010).Asit-and-waitfeedingstrategy
in areas richof food (Freemanet al., 2013;Yodaet al., 2014) andthelocationofproductiveareasbyodortransportedbytheoceanflow (Nevitt&Bonadonna,2005)havebeenproposedaspossibleadditionalexplanations,when thisbehavior takesplacewithin thefeedingareas.
Sincemostofpopulationestimatesofthisspeciesrelyoncount-ingbirdsraftingnearorheadingtocolonies,weevaluatedthespatialdistributionofrestingareas.Thelocationofcoastalpatchesselectedfor rafting suggests an important role of the waters surroundingTavolaraislandasarestingareabeforeandaftervisitingthecolony.Theotherselectedpatchescoincidewithsomeofthemostimport-antfeedingareas:mouthofCoghinasriver(NorthSardinia),Alghero(NorthwestSardinia)andOristano(WestSardinia).ThelatterareaisthemainfeedingdestinationofyelkouanshearwatersfromTavolaraduringtheincubationstage.Anattractiveeffectofnon-homecolo-nies(assuggestedbye.g.,Borgetal.,2016;Bourgeois&Vidal,2008)didnotemergefromourdata,althoughoneoftherestingspotsisclosetoknowncolonies(Alghero,NorthwestSardinia).
Traveling toward distant feeding localities could also be inter-pretedasanindirectconsequenceofbluefintunaThunnus thynnus overfishingintheItalianwaters(Sardiniaincluded).Sincethetradi-tional tuna trapping fisherieswere almost completely replacedbyindustrialfishing,bluefintunastartedtobeharvestedatarateex-ceeding the reproductive capabilities of the existing stock (Longo& Clark, 2012) and, in a few decades, the stocks have collapsed(ICCAT,2010;MacKenzieetal.,2009).Tunasdrivesmallfishesto-wardthesurfaceandareconsideredas"facilitators"forseabirdstowhom they are strongly associated both in tropical and temperate seas (Le Corre & Jaquemet, 2005; Veit &Harrison, 2017). In late
F I G U R E 8 Plotoftheeffectsofoctadsanddistancefromthecolonyonseaproductivityestimatedatbathymetry=−100m.Errorbars =95%ConfidenceIntervals.Resultsfromthelinearmixedmodel(LMM)analyzingtheinverse-transformedseaproductivityinthecellswithatleastoneforagingfixintheoctadswhereforagingtripsofyelkouanshearwaterstaggedattheTavolaraIsland(Sardinia,IT)wererecordedasafunctionofthedistanceofthecolonyandbathymetry(seeTable4fornumericalresults).Numberofobservations:536;numberofcells:91

| 15 of 18PEZZO Et al.
spring intheMediterraneanSea,tunas,duringtheirmigration,getclose to thecoastswhenyelkouanshearwatersare raisingchicks.Then, the drop of tuna population may have reduced the feed-ing opportunities for shearwaters, forcing them to move further.Because theyelkouanshearwater isanendemismconfined to theMediterranean and Tavolara island hosts around half of its globalpopulation,ourfindingssuggestthat,besidedirectthreats(mortal-ityduetobycatchandoverfishingofpreyspecies:Gaudard,2018),conservationmeasurestobeenforcedatseashouldaddressthefullsustainabilityofallfisheriesacrossanareaencompassingtheforag-inghotspotsidentified(namelytheOristanoandAlgherowaters,theBonifacioStraitandtheGulfofLion).
In conclusion, despite the limitations associatedwith the rela-tivelylownumberofmarkedindividuals,somerelevantpatternsofthespatialecologyoftheyelkouanshearwatercouldbedescribed.Theirmain value seems that of referring to the globallymost im-portantcolonyknowntodate.Studiesondifferentpopulationsarestronglyneededinordertoassessandimplementaneffectivepan-Mediterranean conservation strategy for this endemic and charis-matic taxon.
ACKNOWLEDG EMENTSFabio Cherchi, Gian Mario Pitzianti, and Massimo Putzu gave aninvaluable fieldsupport forGPSretrieval,challengingbadseaandinclementweatherduringlong-lastingnightsinthefield.Wewarmlythank Francesco Angioni, Roberto Cogoni, Sergio Nissardi, PierPanzalis, ToreVitale for supportingusduringdeployment and re-trieval,joiningmonitoringactivitiesandhelpingorganizingperiodi-calfieldtrips.FPwasfundedbytheItalianMinistryforEnvironment,LandandSeaProtectionofItaly(MATTM)whichalsoprovidedfundsforfieldworkanddataloggers.
CONFLIC T OF INTERE S TTheauthorshavenoconflictinginterests.
AUTHOR CONTRIBUTIONSNB andMZ conceived the project; NB, MZ, FP, AN, and GS co-ordinatedthedatacollection;GC,DG,AM,AG,andEPperformeddatamanagement/analysesanddraftedpartofthemanuscript;FPandMZdraftedthemanuscriptandFPandEPguidedthefinalwrit-ingwithcontributionsfromallauthors.NBsupervisedallphasesoftheproject since itsbeginning.All authors readandapproved thefinalmanuscript.
E THIC AL APPROVALAllproceduresperformedinthisstudywereinaccordancewiththeethicalstandardsoftheinstitutionorpracticeatwhichthestudieswere conducted.
ORCIDFrancesco Pezzo https://orcid.org/0000-0003-4437-3805 Marco Zenatello https://orcid.org/0000-0002-9225-6737 Dimitri Giunchi https://orcid.org/0000-0003-2753-8997
R E FE R E N C E SAiello-Lammens, M. E., Boria, R. A., Radosavljevic, A., Vilela, B., &
Anderson, R. P. (2015). spThin: An R package for spatial thinningof species occurrence records for use in ecological nichemodels.Ecography,38,541–545.https://doi.org/10.1111/ecog.01132
Araújo,H.,Bastos-Santos, J.,Rodrigues,P.C.,Ferreira,M.,Pereira,A.,Henriques,A.C.,Monteiro,S.S.,Eira,C.,&Vingada,J.(2017).Theimportance of Portuguese Continental Shelf Waters to BalearicShearwaters revealed by aerial census. Marine Biology, 164, 55.https://doi.org/10.1007/s00227-017-3089-x
Baccetti, N., Capizzi, D., Corbi, F., Massa, B., Nissardi, S., Spano, G.,& Sposimo, P. (2009). Breeding shearwaters on Italian islands:Population size, island selection and co-existence with their mainalienpredator,theBlackRat.Riv Ital Ornitol,78,83–100.
Banaru,D.,Mellon-Duval,C.,Roos,D.,Bigot,J.L.,Souplet,A.,Jadaud,A.,Beaubrun,P.,&Fromentin,J.M.(2013).TrophicstructureintheGulfofLionsmarineecosystem(north-westernMediterraneanSea)andfishingimpacts.Journal of Marine Systems,111– 112,45–68.https://doi.org/10.1016/j.jmars ys.2012.09.010
Bartoń, K. (2020). MuMIn: Multi- Model Inference. R package ver-sion 1.43.17. Retrieved from https://CRAN.R-project.org/package=MuMIn
Batschelet,E.(1981).Circular statistics in biology.AcademicPress.BirdLife International. (2018).Puffinus yelkouan. The IUCNRed List of
Threatened Species 2018: e.T22698230A132637221. https://doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22698230A132637221.en
Blondel,J.,Aronson,J.,Bodiou,J.Y.,&Boeuf,G.(2010).The Mediterranean region: Biological diversity in space and time,2nded.OxfordUniversityPress.
Borg,J.J.,Sultana,J.,Metzger,B.,&Barbara,N.(2016).Populationes-timates inMaltesepelagicbreedingbirds: numbers, trends andanappealtoassessthesecautiously.InP.Yesou,J.Sultana,J.Walmsley,&H.Azafzaf (Eds.),Conservation of Marine and Coastal Birds in the Mediterranean (pp.38–42).Proceedingsof theUnep-Map-Rac/SpaSymposium,Hammamet,Tunisia
Bosc,E.,Bricaud,A.,&Antoine,D.(2004).Seasonalandinterannualvariabil-ityinalgalbiomassandprimaryproductionintheMediterraneanSea,asderivedfrom4yearsofSeaWiFSobservations.Global Biogeochem Cy,18,GB1005.https://doi.org/10.1029/2003GB002034v
Bourgeois,K.,&Vidal,E.(2008).TheendemicMediterraneanyelkouanshearwater Puffinus yelkouan: Distribution, threats and a plea formore data. Oryx,42,187–194.https://doi.org/10.1017/S0030605308006467
Brooks,M.E.,Kristensen,K.,Benthem,K. J.,Magnusson,A.,Berg,C.W., Nielsen, A., Skaug, H. J.,Mächler,M., & Bolker, B.M. (2017).glmmTMBbalancesspeedandflexibilityamongpackagesforzero-inflated generalized linear mixedmodeling. The R Journal, 9, 378–400.https://doi.org/10.32614/RJ-2017-066
Burnham,K.P.,&Anderson,D.R.(2002).Model selection and multimodel inference: A practical information- theoretic approach. Springer- Verlag.
Buxton,R.T.,Gormley,A.M.,Jones,C.J.,&Lyver,P.O.(2016).Monitoringburrowingpetrelpopulations:A sampling scheme for themanage-ment of an island keystone species: Petrel SurveyDesign. Journal of Wildlife Management, 80, 149–161. https://doi.org/10.1002/jwmg.994
Caddy,J.(2000).Marinecatchmentbasineffectsversusimpactsoffish-eries on semi- enclosed seas. ICES Journal of Marine Science,57,628–640.https://doi.org/10.1006/jmsc.2000.0739
Cadiou,B.,Pons,J.M.,&Yésou,P.(Eds.)(2004).Oiseaux marins nicheurs de France métropolitaine (1960– 2000).ÉditionsBiotope.
Calenge,C.(2006).ThepackageadehabitatfortheRsoftware:Atoolfortheanalysisofspaceandhabitatusebyanimals.Ecological Modelling,197,516–519.https://doi.org/10.1016/j.ecolmodel.2006.03.017
Capizzi,D., Baccetti,N.,& Sposimo, P. (2010). Prioritizing rat eradica-tiononislandsbycostandeffectivenesstoprotectnestingseabirds.

16 of 18 | PEZZO Et al.
Biological Conservation, 143, 1716–1727. https://doi.org/10.1016/j.biocon.2010.04.020
Carboneras,C. (Ed.) (2013).Seabirds in the Gulf of Lions shelf and slope area.UNEP-MAP-RAC/SPA.
Cecere,J.G.,Gaibani,G.,&Imperio,S.(2014).Effectsofenvironmentalvariabilityandoffspringgrowthonthemovementecologyofbreed-ing Scopoli’s shearwaterCalonectris diomedea. Current Zoology,60,622–630.https://doi.org/10.1093/czoolo/60.5.622
Cesaraccio,G.(1989).Avifauna dell'Arcipelago della Maddalena.QuadernidiItaliaNostraVol1.ItaliaNostra,LaMaddalena.
Chaurand,T.,&Weimerskirch,H.(1994).TheregularalternationofshortandlongforagingtripsinthebluepetrelHalobaena caerulea:Aprevi-ouslyundescribedstrategyoffoodprovisioninginapelagicseabird.Journal of Animal Ecology,63,275–282.https://doi.org/10.2307/5546
Coll, M., Piroddi, C., Steenbeek, J., Kaschner, K., Ben Rais Lasram,F., Aguzzi, J., Ballesteros, E., Bianchi, C. N., Corbera, J., Dailianis,T., Danovaro, R., Estrada, M., Froglia, C., Galil, B. S., Gasol, J.M., Gertwagen, R., Gil, J., Guilhaumon, F., Kesner-Reyes, K., …Voultsiadou,E. (2010).Thebiodiversityof theMediterraneanSea:Estimates, patterns, and threats.PLoS One,5, e11842. https://doi.org/10.1371/journal.pone.0011842
D’Ortenzio,F.,&Riberad’Alcala,M. (2009).OnthetrophicregimesoftheMediterranean Sea: A satellite analysis.Biogeoscience,6, 139–148.https://doi.org/10.5194/bg-6-139-2009
Darnaude,A.M.(2005).Fishecologyandterrestrialcarbonuseincoastalareas:Implicationsformarinefishproduction.Journal of Animal Ecology,74,864–876.https://doi.org/10.1111/j.1365-2656.2005.00978.x
David,L.,&Di-Méglio,N.(Eds.)(2013).Important areas for the conserva-tion of cetaceans in the Gulf of Lions shelf and slope area: Synthesis of existing data on cetaceans and threats.UNEP-MAP-RAC/SPA.
Dean, B., Freeman, R., Kirk,H., Leonard, K., Phillips, R.A., Perrins, C.M.,&Guilford,T.(2013).Behaviouralmappingofapelagicseabird:CombiningmultiplesensorsandahiddenMarkovmodelrevealsthedistributionofat-seabehaviour.Journal of the Royal Society, Interface,10,20120570.https://doi.org/10.1098/rsif.2012.0570
Derhé,M.A.(2012).DevelopingapopulationassessmentforYelkouanShearwaters Puffinus yelkouan.InP.Yésou,N.Baccetti,&J.Sultana(Eds.),Ecology and Conservation of Mediterranean Seabirds and other bird species under Barcelona Convention (pp. 65–73). Proceedingsof the 13th Medmaravis Pan-Mediterranean Symposium, Alghero(Sardinia)14-17Oct.2011,Medmaravis,Alghero
Fayet,A.L.,Freeman,R.,Shoji,A.,Padget,O.,Perrins,C.M.,&Guilford,T. (2015). Lower foraging efficiency in immatures drives spatialsegregation with breeding adults in a long- lived pelagic seabird. Animal Behavior, 110, 79–89. https://doi.org/10.1016/j.anbehav.2015.09.008
Fielding,A.H.,&Bell,J.F.(1997).Areviewofmethodsfortheassess-ment of prediction errors in conservation presence/absencemod-els. Environmental Conservation,24,38–49.https://doi.org/10.1017/S0376892997000088
Fox,J.,&Weisberg,S.(2019).An R companion to applied regression,3rded. Sage.
Freeman,R.,Dean,B.,Kirk,H.,Leonard,K.,Phillips,R.A.,Perrins,C.M.,&Guilford,T.(2013).Predictiveethoinformaticsrevealsthecomplexmigratorybehaviourofapelagicseabird.Journal of the Royal Society, Interface,10,20130279.
Gaston,A.J.(2004).Seabirds: A natural history.London,YaleUniversityPress,NewHaven.
Gatt,M.C., Lago, P., Austad,M., Bonnet-Lebrun,A. S.,&Metzger, B.J. (2019).Pre-layingmovementsofYelkouanShearwaters (Puffinus yelkouan) intheCentralMediterranean.Journal of Ornithology,160,625–632.https://doi.org/10.1007/s10336-019-01646-x
Gaudard,C.(compiler)(2018).Single International Species Action Plan for the Yelkouan Shearwater Puffinus yelkouan. Project LIFE 14 PRE/
UK/000002.CoordinatedEffortsforInternationalSpeciesRecoveryEuroSAP.LPO/BirdLifeFrance,.
Giorgi,F.(2006).Climatechangehot-spots.Geophysical Research Letters,33,L08707.https://doi.org/10.1029/2006GL025734
Granadeiro,J.P.,Nunes,M.,Silva,M.C.,&Furness,R.W.(1998).FlexibleforagingstrategyofCory’sshearwater,Calonectris diomedea,duringthechick-rearingperiod.Animal Behavior,56,1169–1176.https://doi.org/10.1006/anbe.1998.0827
Grémillet,D., Lewis,S.,Drapeau,L., vanderLingen,C.D.,Huggett, J.A.,Coetzee,J.C.,Verheye,H.M.,Daunt,F.,Wanless,S.,&Ryan,P.G. (2008).Spatialmatch-mismatchintheBenguelaupwellingzone:Shouldwe expect chlorophyll and SST to predictmarine predatordistributions? Journal of Applied Ecology, 45, 610–621. https://doi.org/10.1111/j.1365-2664.2007.01447.x
Guilford,T.C.,Meade, J., Freeman,R.,Biro,D., Evans,T.,Bonadonna,F., Boyle,D., Roberts, S.,&Perrins,C.M. (2008).GPS tracking ofthe foraging movements of Manx Shearwaters Puffinus puffinus breeding on Skomer Island,Wales. Ibis,150, 462–473. https://doi.org/10.1111/j.1474-919X.2008.00805.x
Harmelin-Vivien,M.L.,Banaru,D.,Dierking,J.,Hermand,R.,Letourneur,Y., & Salen-Picard, C. (2009). Linking benthic biodiversity to thefunctioning of coastal ecosystems subjected to river runoff (NWMediterranean).Animal Biodiversity and Conservation,32,135–145.
Hartig, F. (2020).DHARMa: Residual Diagnostics for Hierarchical (Multi- Level/Mixed) Regression Models.Rpackageversion0.3.3.0.Retrievedfromhttps://CRAN.R-project.org/package=DHARMa
ICCAT.(2010).Report of the standing committee on research and statistics (SCRS). International Commission for the Conservation of AtlanticTuna.
Kie,J.G.(2013).Arule-basedadhocmethodforselectingabandwidthinKernelhome-rangeanalyses.Anim Biotelem,1,1–12.https://doi.org/10.1186/2050-3385-1-13
Kress,N.,Manca,B.B.,Klein,B.,&Deponte,D.(2003).ContinuinginfluenceofthechangedthermohalinecirculationintheeasternMediterraneanon the distribution of dissolved oxygen and nutrients: Physical andchemicalcharacterizationofthewatermasses.Journal of Geophysical Research,108,8109.https://doi.org/10.1029/2002JC001397
Lambert,C.,Laran,S.,David,L.,Dorémus,G.,Pettex,E.,VanCanneyt,O., & Ridoux, V. (2017). How does ocean seasonality drive habi-tat preferences of highlymobile top predators? Part I: The north-western Mediterranean Sea. Deep Sea Res Part II Top Stud Oceanogr,141,115–132.https://doi.org/10.1016/j.dsr2.2016.06.012
LeCorre,M.,&Jaquemet,S.(2005).Assessmentoftheseabirdcommu-nityoftheMozambiqueChannelanditspotentialuseasanindicatoroftunaabundance.Estuarine, Coastal and Shelf Science,63(3),421–428.https://doi.org/10.1016/j.ecss.2004.11.013
Lejeusne, C., Chevaldonné, P., Pergent-Martini, C., Boudouresque, C.F.,&Pérez,T.(2010).Climatechangeeffectsonaminiatureocean:The highly diverse, highly impacted Mediterranean Sea. Trends in Ecology & Evolution, 25, 250–260. https://doi.org/10.1016/j.tree.2009.10.009
Longo,S.B.,&Clark,B.(2012).ThecommodificationofBluefinTuna:ThehistoricaltransformationoftheMediterraneanfishery.J Agrar Change,12,204–226.https://doi.org/10.1111/j.1471-0366.2011.00348.x
Lüdecke,D.,Makowski,D.,Waggoner,P.,&Patil,I.(2020).Assessment of regression models performance.CRAN.Retrievedfromhttps://easystats.github.io/performance/
Ludwig, W., Dumont, E., Meybeck, M., & Heussner, S. (2009). Riverdischargesofwaterandnutrients to theMediterraneanandBlackSea: Major drivers for ecosystem changes during past and futuredecades? Progr Oceanogr, 80, 199–217. https://doi.org/10.1016/j.pocean.2009.02.001
Macias,D.M.,Garcia-Gorriz,E.,&Stips,A.(2015).Productivitychangesin theMediterraneanSea for the twenty-first century in response

| 17 of 18PEZZO Et al.
to changes in the regional atmospheric forcing.Frontiers in Marine Science,2.https://doi.org/10.3389/fmars.2015.00079
MacKenzie, B. R., Mosegaard, H., & Rosenberg, A. A. (2009).Impending collapse of Bluefin Tuna in the Northeast Atlanticand Mediterranean. Conservation Letters, 2, 25–34. https://doi.org/10.1111/j.1755-263X.2008.00039.x
Magalhães,M.C., Santos, R. S., &Hamer, K. C. (2008).Dual-foragingofCory’sshearwatersintheAzores:Feedinglocations,behaviouratseaandimplicationsforfoodprovisioningofchicks.Marine Ecology Progress Series,359,283–293.https://doi.org/10.3354/meps07340
Manly,B.F.J.,McDonald,L.L.,Thomas,D.L.,McDonald,T.L.,&Erickson,W.P.(2002).Resource selection by animals: Statistical analysis and de-sign for field studies,2nded.Kluwer.
Meier,R.,Wynn,R.,Votier,S.,McMinnGrive,M.,Rodriguez,A.,Maurice,L.,VanLoon,E.,Jones,A.,Suberg,L.,Arco,J.,Morgan,G.,Josey,S.,&Guilford,T.(2015).ConsistentforagingandcommutingcorridorsofthecriticallyendangeredBalearicshearwaterPuffinus mauretanicus in the northwest Mediterranean. Biological Conservation,190,87–97.https://doi.org/10.1016/j.biocon.2015.05.012
Meyer,C.B.,&Thuiller,W.(2006).Accuracyofresourceselectionfunc-tions across spatial scales. Diversity and Distributions,12, 288–297.https://doi.org/10.1111/j.1366-9516.2006.00241.x
Micheli,F.,Halpern,B.S.,Walbridge,S.,Ciriaco,S.,Ferretti,F.,Fraschetti,S., Lewison,R.,Nykjaer, L.,&Rosenberg,A.A. (2013).Cumulativehuman impacts on mediterranean and black sea marine ecosys-tems:Assessingcurrentpressuresandopportunities.PLoS One,81,e79889.https://doi.org/10.1371/journal.pone.0079889
Millot,C.(1990).TheGulfofLions’hydrodynamics.Continental Shelf Research,10,885–894.https://doi.org/10.1016/0278-4343(90)90065-T
Nakagawa,S.,&Schielzeth,H.(2013).AgeneralandsimplemethodforobtainingR2fromgeneralizedlinearmixedeffectsmodels.Methods in Ecology and Evolution,4,133–142.
Nevitt,G.A.,&Bonadonna,F. (2005).Sensitivity todimethylsulphidesuggestsamechanismforolfactorynavigationbyseabirds.Biology Letters,1,303–305.https://doi.org/10.1098/rsbl.2005.0350
Pearce, J., & Ferrier, S. (2000). Evaluating the predictive perfor-mance of habitat models developed using logistic regression.Ecological Modelling,133, 225–245. https://doi.org/10.1016/S0304-3800(00)00322-7
Pérez-Ortega, M., & İsfendiyaroğlu, S. (2017). Predicting foraginghotspotsforYelkouanShearwaterintheBlackSea.Deep Sea Research Part II: Topical Studies in Oceanography, 141, 237–247. https://doi.org/10.1016/j.dsr2.2016.07.007
Péron, C., Grémillet, D., Prudor, A., Pettex, E., Saraux, C., Soriano-Redondo, A., Authier,M., & Fort, J. (2013). Importance of coastalMarineProtectedAreasfortheconservationofpelagicseabirds:ThecaseofVulnerableyelkouanshearwatersintheMediterraneanSea.Biological Conservation, 168, 210–221. https://doi.org/10.1016/j.biocon.2013.09.006
Pezzo,F.,Zenatello,M.,Cerritelli,G.,Navone,A.,Giunchi,D.,Spano,G.,Pollonara,E.,Massolo,A.,Gagliardo,A.,&Baccetti,N.(2021).Datafrom:ProductivitychangesintheMediterraneanSeadriveforagingmovementsofyelkouanshearwaterPuffinusyelkouanfromthecoreof itsglobalbreedingrange.Movebank Data Repository. https://doi.org/10.5441/001/1.k49p9t60
Pujo-Pay, M., Conan, P., Oriol, L., Cornet-Barthaux, V., Falco, C.,Ghiglione,J.F.,Goyet,C.,Moutin,T.,&Prieur,L.(2011).Integratedsurvey of elemental stoichiometry (C, N, P) from the western toeastern Mediterranean Sea. Biogeosciences,8,883–899.https://doi.org/10.5194/bg-8-883-2011
RCoreTeam.(2021).R: A language and environment for statistical comput-ing.RFoundationforStatisticalComputing.Retrievedfromhttps://www.R- proje ct.org/
Ragionieri, L., Cutuli, G., Sposimo, P., Spano, G., Navone, A., Capizzi,D., Baccetti, N., Vannini, M., & Fratini, S. (2013). Establishing the
eradicationunitofMolaraIsland:AcaseofstudyfromSardinia,Italy.Biological Invasions, 15, 2731–2742. https://doi.org/10.1007/s10530-013-0487-y
Raine, A. F., Borg, J. J., Raine, H., & Phillips, R. A. (2013). Migrationstrategies of the Yelkouan Shearwater Puffinus yelkouan. Journal of Ornithology, 154, 411–422. https://doi.org/10.1007/s10336-012-0905-4
Raine,A.,Raine,H.,Meirinho,A.,&Borg,J.J.(2010).Raftingbehaviourof Yelkouan Shearwater Puffinus yelkouan breeding at Rdum tal- Madonna,Malta.Il- Merrill,32,26–30.
Riou,S.,Gray,C.M.,Brooke,M.L.,Quillfeldt,P.,Masello,J.F.,Perrins,C.,&Hamer,K.C.(2011).Recentimpactsofanthropogenicclimatechange on a higher marine predator in western Britain. Marine Ecology Progress Series, 422, 105–112. https://doi.org/10.3354/meps0 8968
Ropert-Coudert,Y.,Grémillet,D.,Kato,A.,Ryan,P.G.,Naito,Y.,&LeMaho, Y. (2004). A fine-scale time budget of Cape gannets pro-vides insights into the foraging strategies of coastal seabirds.Animal Behavior, 67, 985–992. https://doi.org/10.1016/j.anbehav.2003.09.010
Ruffino,L.,Bourgeois,K.,Vidal,E.,Duhem,C.,Paracuellos,M.,Escribano,F., Sposimo,P., Baccetti,N., Pascal,M.,&Oro,D. (2009). Invasiveratsandseabirdsafter2,000yearsofanunwantedcoexistenceonMediterranean. Biological Invasions,11,631–1651.
Şahin,D.,Bacak,E.,Bilgin,S.,Atay,C.,Boyla,K.A.,&Tavares,J.(2012).PresenceandbehaviourofYelkouanShearwatersPuffinusyelkouanattheBosphorus.InP.Yésou,N.Baccetti,&J.Sultana(Eds.),Ecology and Conservation of Mediterranean Seabirds and other bird species under Barcelona Convention (pp. 54–57). Proceedings of the 13thMedmaravisPan-MediterraneanSymposiumAlghero(Sardinia)14–17October2011,Medmaravis,Alghero.
Sandvik, H., Barrett, R. T., Erikstad, K. E.,Myksvoll,M. S., Vikebø, F.,Yoccoz, N. G., Anker-Nilssen, T., Lorentsen, S. H., Reiertsen, T.K., Skarðhamar, J., Skern-Mauritzen, M., & Systad, G. H. (2016).Modelleddriftpatternsoffishlarvaelinkcoastalmorphologytosea-bird colony distribution. Nature Communications,7, 11599. https://doi.org/10.1038/ncomms11599
Seaman,D.E.,&Powell,R.A.(1990).Identifyingpatternsandintensityof home range use. International Association for Bear Research and Management,8,243–249.
Shamoun-Baranes, J., Bouten, W., Camphuysen, C. J., & Baaij, E.(2011). Riding the tide: Intriguing observations of gulls rest-ing at sea during breeding. Ibis, 153, 411–415. https://doi.org/10.1111/j.1474-919X.2010.01096.x
Shiomi, K., Yoda, K., Katsumata,N., & Katsufumi, S. (2012). Temporaltuningofhomewardflightsinseabirds.Animal Behavior,83,355–359.https://doi.org/10.1016/j.anbeh av.2011.11.010
Sing, T., Sander, O., Beerenwinkel, N., & Lengauer, T. (2005). ROCR:Visualizing classifier performance in R. Bioinformatics, 21, 3940–3941.https://doi.org/10.1093/bioinformatics/bti623
Sposimo,P.,Spano,G.,Navone,A.,Fratini,S.,Ragionieri,L.,Putzu,M.,Capizzi, D., & Baccetti, N. (2012a). Rodent eradication onMolaraIslandandsurroundingislets(NESardinia):Fromsuccesstotheriddleofreinvasion.Aliens,32,33–38.
Sposimo, P., Spano, G., Navone, A., Fratini, S., Ragionieri, L., Putzu,M., Capizzi, D., Baccetti, N., & Lastrucci, B. (2012b). Rat eradica-tion at Yelkouan Shearwater Puffinus yelkouan colonies on NESardinianislets:Successfollowedbyunexplainedre-appearance.InP.Yésou,G.Sultana,&N.Baccetti (Eds.),Ecology and Conservation of Mediterranean Seabirds and other bird species under the Barcelona Convention (pp.58–64).Proceedingsof the13thMedmaravisPan-MediterraneanSymposium.Medmaravis,Alghero.
Stahl,J.C.,&Sagar,P.M.(2000).ForagingstrategiesofsouthernBuller’salbatrosses Diomedea b. bulleribreedingontheSnares,NewZealand.Journal of the Royal Society of New Zealand,30,299–318.

18 of 18 | PEZZO Et al.
Tanhua, T., Hainbucher, D., Schroeder, K., Cardin, V., Álvarez, M., &Civitarese,G.(2013).TheMediterraneanSeasystem:Areviewandan introduction to the special issue. Ocean Science, 9, 789–803.https://doi.org/10.5194/os-9-789-2013
Terauds,A.,&Gales,R.(2006).Provisioningstrategiesandgrowthpat-terns of light-manted sooty albatrosses Phoebetria palpebrata on MacquarieIsland.Polar Biology,29,917–926.
Thibault,J.C.,&Bonaccorsi,G.(1999).The Birds of Corsica: An annotated checklist.BOUChecklistNumber17.BritishOrnithologists'Union,.
Thieurmel,B.,&Elmarhraoui,A. (2019).Suncalc: Compute Sun Position, Sunlight Phases, Moon Position and Lunar Phase. R package version0.5.0.Retrievedfromhttps://CRAN.R-project.org/package=suncalc
Tyson,C.,Kirk,H.,Fayet,A.,VanLoon,E.E.,Shoji,A.,Dean,B.,Perrins,C.M.,Freeman,R.,&Guilford,T.(2017).Coordinatedprovisioninginadual-foragingpelagicseabird.Animal Behavior,132,73–79.https://doi.org/10.1016/j.anbehav.2017.07.022
Veit,R.R.,&Harrison,N.M.(2017).Positiveinteractionsamongforag-ingseabirds,marinemammalsandfishesand implicationsfortheirconservation. Frontiers in Ecology and Evolution,5, 121. https://doi.org/10.3389/fevo.2017.00121
Weimerskirch, H., Chastel, O., Ackermann, L., Chaurand, T.,Cuenotchaillet,F.,Hindermeyer,X.,&Judas,J.(1994).Alternatelongandshortforagingtripsinpelagicseabirdparents.Animal Behavior,47,472–476.https://doi.org/10.1006/anbe.1994.1065
Worton,B.J. (1989).Kernelmethodsforestimatingtheutilizationdis-tribution in home range studies. Ecology,70, 164–168. https://doi.org/10.2307/1938423
Wynn,R.B.,Josey,S.A.,Martin,A.P.,Johns,D.G.,&Yésou,P.(2007).Climate-drivenrangeexpansionofacriticallyendangeredtoppreda-torinnortheastAtlanticwaters.Biology Letters,3,529–532.https://doi.org/10.1098/rsbl.2007.0162
Yoda,K.,Shiomi,K.,&Sato,K.(2014).Foragingspotsofstreakedshear-watersinrelationtooceansurfacecurrentsasidentifiedusingtheir
driftmovements.Progress in Oceanography,122,54–64.https://doi.org/10.1016/j.pocean.2013.12.002
Zenatello, M., Spano, G., Zucca, C., Navone, A., Putzu,M., Azara, C.,Trainito,E.,Ugo,M.,&Baccetti,N.(2012).Movementsand“moving”population estimates of Yelkouan Shearwater Puffinus yelkouan at Tavolara,Sardinia.InP.Yésou,N.Baccetti,&J.Sultana(Eds.),Ecology and Conservation of Mediterranean Seabirds and other bird species under the Barcelona Convention(pp39–47).Proceedingsofthe13thMedmaravispan-MediterraneanSymposium,Alghero,Sardinia,Italy
Zenatello,M.,Zucca,C.,Nissardi,S.,&Baccetti,N.(2006).Distribuzione di Berta maggiore e Berta minore in Sardegna (Giugno 2006). Relazione tecnica INFS. Retrieved from http://www.infs-acquatici.it/PDF/berte/bertesardegna2006_31ago2006.pdf
Zotier,R.,Bretagnolle,V.,&Thibault,J.C.(2003).Biogeographyofthema-rinebirdsofaconfinedsea,theMediterranean.Journal of Biogeography,26,297–313.https://doi.org/10.1046/j.1365-2699.1999.00260.x
Zuur,A.F.,&Ieno,E.N.(2016).Aprotocolforconductingandpresentingresultsofregression-typeanalyses.Methods in Ecology and Evolution,7(6),636–645.https://doi.org/10.1111/2041-210X.12577
How to cite this article:Pezzo,F.,ZenatelloM.,CerritelliG.,NavoneA.,GiunchiD.,SpanoG.,PollonaraE.,MassoloA.,GagliardoA.,&BaccettiN.(2021).ProductivitychangesintheMediterraneanSeadriveforagingmovementsofyelkouanshearwaterPuffinus yelkouanfromthecoreofitsglobal breeding range. Marine Ecology,00e1–18.https://doi.org/10.1111/maec.12668























![Welcome! [] - Opportunitie… · • Champion of Tourism Productivity Nexus (TPN), an initiative under Malaysia Productivity Blueprint Mr. Uzaidi Udanis has been involved in tourism](https://img.pdfslide.net/doc/110x75/5f0b0fab7e708231d42ea89e/welcome-opportunitie-a-champion-of-tourism-productivity-nexus-tpn.jpg)





