Embed Size (px)
Citation preview

4073RESEARCH ARTICLE
INTRODUCTIONThe cranial neural crest (CNC) shapes the vertebrate face. Thesecells possess two exceptional properties: pluripotency and motility.CNC cells differentiate to melanocytes, peripheral neurons, glia,cartilage, bone and the inflow tracts of the heart, based on anorchestrated gene regulatory network and their migratorytrajectories (Sauka-Spengler and Bronner-Fraser, 2008). CNC cellsshare important characteristics with carcinoma cells in undergoingepithelial-mesenchymal transition (EMT) and tissue invasion(Acloque et al., 2009). They express genes involved in tumorformation, such as c-myc, snail1 and snail2 (slug). They alsoupregulate mesenchymal cadherins and metalloproteinases, whichare enriched during metastasis (Kuriyama and Mayor, 2008).Therefore, deciphering the regulatory network of CNCdevelopment would not only help to understand congenital defectsof craniofacial development but also provide insight into genefunctions in tumor formation and progression.
CNC is induced at the neuroectoderm/ectoderm border under theinfluence of defined levels of BMP, canonical Wnt, retinoic acid(RA), FGF and notch signaling (Basch and Bronner-Fraser, 2006).These signals activate the neural plate border specifiers pax3, zic1and dlx, which in turn activate expression of the more restricted,localized CNC specifiers, including c-myc, foxD3, AP-2, snail1 andsox9. The latter are thought to initiate CNC fate specification bycontrolling CNC proliferation and apoptosis, EMT and emigration
from the neural fold. In addition, snail2, sox10 and twist areactivated, which serves to maintain CNC pluripotency and survivalbut also to regulate the expression of genes required for cellmigration and differentiation (Meulemans and Bronner-Fraser,2004; Sauka-Spengler and Bronner-Fraser, 2008; Steventon et al.,2005). This view of hierarchy in neural crest gene activation ismost likely an oversimplification because there is mountingevidence of a cross-regulation between snail1, snail2 and twist inthe induction of neural crest at the gastrula stage (Carl et al., 1999;Zhang et al., 2006; Zhang and Klymkowsky, 2009). Although wehave to reconsider the regulatory network at the level of neuralcrest specification, the upstream requirement of the neural plateborder specifiers appears correct. In addition to their role in theCNC, neural plate border specifiers such as pax3 and zic1 alsopromote hatching gland and preplacodal development and acttogether to stimulate neural crest formation (Hong and Saint-Jeannet, 2007). In addition, gbx2 collaborates with zic1 inactivating neural crest genes and inhibiting the placodal fate (Li etal., 2009).
Not all homologs of neural crest specifier genes that areupregulated in human tumors share common functions with theirorthologs in neural crest cells. snail1 and snail2 behave similarlyby promoting EMT and blocking apoptosis. The proto-oncogene c-myc, however, operates differently. When overexpressed ormutated, c-myc promotes tumor growth by facilitating cell cycleentry (Herold et al., 2009; Morrish et al., 2009) and sensitizes cellsfor apoptosis (Hoffman and Liebermann, 2008). In neural crestdevelopment, c-myc neither influences cell proliferation nor celldeath; rather, it is required for snail1, snail2 and twist expression(Bellmeyer et al., 2003).
The human prohibitin homologs, PHB (PHB1) and PHB2, aretumor suppressors that share 53% amino acid sequence identity;these proteins are localized to the plasma membrane, nucleus andmitochondria (Mishra et al., 2006). Studies in C. elegans haverevealed a crucial mitochondrial function for prohibitins in
Development 137, 4073-4081 (2010) doi:10.1242/dev.053405© 2010. Published by The Company of Biologists Ltd
KIT, Campus South, Zoologicak Institute, Cell and Developmental Biology,Kaiserstrasse 12, D-76131 Karlsruhe, Germany.
*Present address: Department of Biology, University of Erlangen-Nuernberg,D-91058 Erlangen, Germany†Author for correspondence ([email protected])
Accepted 29 September 2010
SUMMARYProhibitin 1 (phb1), which was initially described as an inhibitor of cell proliferation, is a highly conserved protein found inmultiple cellular compartments. In the nucleus it interacts with the transcriptional regulators Rb and E2F1 and controls cellproliferation and apoptosis. Here we unravel an unexpected novel function for phb1 in Xenopus cranial neural crest (CNC)development. Xphb1 is maternally expressed; zygotically expressed neurula stage transcripts accumulate in the CNC and theneural tube. Knockdown of Xphb1 by antisense morpholino injection results in the loss of foxD3, snail2 and twist expression,whereas expression of c-myc, AP-2 and snail1 remains unaffected. Xphb2, its closest relative, cannot substitute for Xphb1,underlining the specificity of Xphb1 function. Epistatic analyses place Xphb1 downstream of c-myc and upstream of foxD3, snail2and twist. To elucidate which subdomain in Xphb1 is required for neural crest gene regulation we generated deletion mutantsand tested their rescue ability in Xphb1 morphants. The E2F1-binding domain was found to be necessary for Xphb1 function inneural crest development. Gain- and loss-of-function experiments reveal that Xphb1 represses E2F1 activity; suppression of E2F1through Xphb1 is required for twist, snail2 and foxD3 expression in the CNC. With the Xphb1 dependency of a subset of CNCspecifiers downstream of c-myc, we have identified a new branching point in the neural crest gene regulatory network.
KEY WORDS: Snail/slug regulation, c-myc, Cranial neural crest, Xenopus
Prohibitin1 acts as a neural crest specifier in Xenopusdevelopment by repressing the transcription factor E2F1Martina Schneider, Alexandra Schambony* and Doris Wedlich†
DEVELO
PMENT

4074
development and lifespan control (Artal-Sanz and Tavernarakis,2009; Artal-Sanz et al., 2003); in mitochondria, the prohibitinslocalize predominantly to the inner membrane where they form amultimeric complex with a chaperone-like function (Osman et al.,2009). Nuclear prohibitins inhibit cell proliferation and suppressapoptosis (Fusaro et al., 2002; Wang et al., 1999a). Here, we reportthat Xenopus prohibitin1 (Xphb1) acts downstream of c-myc inneural crest development to control snail2, foxD3 and twistexpression. Whereas PHB-1/PHB1 and PHB-2/PHB2 appearfunctionally synonymous in C. elegans and human cell lines, thisis not the case in Xenopus. The role of Phb1 in neural crestdevelopment is not associated with cell proliferation or apoptosisbut involves its ability to bind to and repress the activity of thetranscription factor E2F1.
MATERIALS AND METHODSXenopus embryos, micromanipulation and lineage tracingEmbryos were obtained by in vitro fertilization and staged according toNieuwkoop and Faber (Nieuwkoop and Faber, 1967). mRNA for injectionexperiments was synthesized in vitro using the mMessage mMachine Kit(Ambion, Norwalk, CT, USA). Xphb1 (phb1 MO, 5�-ATCCCTGTT -CTTCCACACGGCTAAT-3�) and c-myc [c-myc MO, as previouslydescribed (Bellmeyer et al., 2003)] morpholino antisense oligonucleotideswere purchased from Gene Tools (Philomath, OR, USA). As a control weused the 3�carboxyfluorescein-tagged standard control morpholinooligonucleotide (co MO) designed by Gene Tools.
Unless indicated otherwise, all injections were performed into the animalhemisphere of one blastomere of 2-cell stage embryos as follows: 700 pgRNA, 16 ng phb1 MO, 11 ng c-myc MO. Embryos were co-injected withlacZ (b-gal) DNA (50 pg) to identify the manipulated side. Embryos werefixed in MEMFA at stage 18 (unless otherwise noted) and successivelyprocessed for b-gal staining (Sive et al., 2000) using X-Gal (Applichem,Darmstadt, Germany) as substrate. In the rescue experiment, in whichHphb1 mRNA was co-injected, dextran-FITC (4 pg; Molecular Probes,Eugene, OR, USA) was used as a lineage tracer.
ConstructsXphb1 and Xphb2 were amplified from neurula stage embryos. Xphb1 wasfused C-terminally with six copies of the c-myc epitope in the pCS2+vector (pCS2+myc). All Xphb1 mutants, Xphb1DN-term (amino acids 116-275), Xphb1DRaf-1 (amino acids 1-243) and Xphb1DC-term (amino acids1-188), were subcloned into pCS2+myc. Primers are listed in Table S1 inthe supplementary material. The mutant Hphb1D185-214, a kind gift of S.Chellappan (Columbia University, New York, USA), was subcloned intothe pCS2+ vector and the Hphb1 (pcDNA3-PHB) plasmid was kindlyprovided by T. Rudel (Julius-Maximilians-Universität, Würzburg,Germany).
Whole-mount analysisIn situ hybridization was performed as described (Gawantka et al., 1995).Antisense DIG-labeled probes were synthesized with the DIG RNALabeling Kit (Roche, Basel, Switzerland) using template cDNA encodingXphb1, twist (Hopwood et al., 1989), snail2 (Mayor et al., 1995), snail1(Essex et al., 1993), foxD3 (Sasai et al., 2001), c-myc (Bellmeyer et al.,2003) AP-2 (Luo et al., 2003), Xbra (Smith et al., 1991), chordin (Sasai etal., 1995), sox2 (Kishi et al., 2000), cytokeratin Xk81A1 (Jonas et al.,1985), pax3 (Bang et al., 1997), zic3 (Nakata et al., 1997) and meis3(Salzberg et al., 1999). Images were captured as described previously(Etard et al., 2005). The probe for cytokeratin Xk81A1 was kindlyprovided by R. Mayor (UCL, London, UK). The results of at least threeindependent experiments were averaged, and statistical significance wascalculated using Student’s t-test. Proliferation was estimated at stage 18 byphospho-histone H3 detection, as previously described (Turner andWeintraub, 1994). Anti-phospho-histone H3 antibody (1:700; Millipore,Temecula, CA, USA) and anti-rabbit antibody conjugated with alkalinephosphatase (1:700; Dianova, Hamburg, Germany) were used. TUNELstaining was carried out at stage 18 as described (Hensey and Gautier,
1998). DIG was detected by anti-DIG Fab fragments conjugated to alkalinephosphatase (1:2000; Roche) and the chromogenic reaction was performedusing NBT/BCIP in single stainings. In double stainings, BM Purple(Roche) and BCIP were used.
RT-PCR and real-time PCRFor RT-PCR assays, RNA was extracted from groups of five embryos usingthe High Pure RNA Isolation Kit (Roche). Total RNA (500 ng) was reversetranscribed (M-MLV, Promega) and cDNA was amplified according tostandard protocols. For real-time PCR, embryos were injected animally inthe dorsal blastomeres at the 4-cell stage and total RNA was extracted fromthree whole embryos at stage 18 and reverse transcribed. Real-time PCRwas performed using iQ SYBR Green Supermix on an iCycler (BioRad,Hercules, CA, USA). Expression levels were calculated relative toornithine decarboxylase (ODC) and were normalized to uninjectedcontrols. Results of at least three independent experiments were averaged,and statistical significance was calculated using Student’s t-test. Primersare listed in Table S1 in the supplementary material.
Protein analysisThe specificity of phb1 MO was analyzed using the TnT In VitroTranslation Kit (Promega, Mannheim, Germany) with radioactive[35S]methionine (GE Healthcare, Little Chalfont, UK). After incubation of1 mg/ml Xphb1, Xphb2 or Hphb1 DNA alone or together with the phb1 MO(1 mM), samples were subject to SDS-PAGE and evaluation performedusing a BAS-1500 phosphorimager (Fujifilm, Düsseldorf, Germany).
Western blots were carried out as described (Unterseher et al., 2004).Lysates corresponding to half an embryo were separated by SDS-PAGE.Rabbit polyclonal anti-prohibitin antibody was obtained from Abcam(1:4000; Cambridge, UK) and alkaline phosphatase-conjugated secondaryantibody from Dianova (1:3000). b-catenin (a kind gift of Ralph Rupp,München, Germany), with Coomassie Brilliant Blue (Serva, Heidelberg,Germany) staining, was used as a loading control.
Reporter gene assayFor luciferase reporter assays, Xenopus embryos were injected animallyinto one blastomere at the 2-cell stage with the pFR-luciferase trans-reporter plasmid Gal4-DBD-E2F1 and different concentrations of Xphb1mRNA. As a negative control, only the pFR trans-reporter plasmid (40 pg)was injected (data not shown). As a positive control, the pFR trans-reporterplasmid (40 pg) was co-injected with Gal4-DBD-E2F1 (50 pg). This valuewas set as 100% in each experiment. The reporter gene assay wasperformed at stage 18 as previously described (Gradl et al., 1999). The pFRtrans-reporter plasmid and the Gal4-DBD-E2F1 plasmid, which is a fusionof the Gal4 DNA-binding domain (DBD) with E2F1, were kind gifts of S.Kang (Korea University, Seoul, South Korea).
RESULTSXenopus prohibitin1 (Xphb1) is expressed in CNCand is required for twist expressionIn a search for genes expressed in migrating cells of the earlyXenopus embryo we identified prohibitin1 (Xphb1) in neural crestcells by whole-mount in situ hybridization (ISH) (Fig. 1). Atblastula stage, Xphb1 transcripts localized in the animalhemisphere, accumulated in the posterior dorsal area at late gastrulaand concentrated in CNC and neural tube at neurula stage (Fig. 1A-C). When gastrula embryos were cut into two halves (semi-sections), Xphb1 was detected in the mesendoderm at stage 10 (Fig.1D,d) and in the posterior neuroectoderm and mesoderm at stage11.5 (Fig. 1E,e). The sense Xphb1 probe yielded no signal (Fig.1d�,e�). Xphb1 expression in CNC was maintained during theirmigration, and additional expression areas were found in the eye,the otic vesicle and the brain (Fig. 1F,G). Double ISH for Xphb1and twist revealed partial overlap of the expression domains atstage 18 (Fig. 1H-H�). Xphb2, which shares 53% amino acididentity with Xphb1, was not detected in the neural crest (data not
RESEARCH ARTICLE Development 137 (23)
DEVELO
PMENT

shown). RT-PCR (Fig. 1I) and immunoblots (see Fig. S1 in thesupplementary material) showed that Xphb1 is expressedthroughout early development.
4075RESEARCH ARTICLEXphb1 specifies neural crest by repressing E2F1
Fig. 1. Xphb1 is expressed in the presumptive cranial neural crest(CNC) region. (A-H�) In situ hybridization (ISH) for Xphb1 (A-H) andXphb1 plus twist (H�,H�). (A)At blastula (stage 9) Xphb1 is expressed inthe dorsal animal ectoderm (arrowheads; animal pole is towards the top).(B)At late gastrula stage (stage 11.5) Xphb1 transcripts are found in theposterior dorsal area (arrowhead). (C)At neurula stage (dorsal view)Xphb1 is detected in migrating neural crest (arrowhead) and in the neuraltube. (D,E)Semi-sections of stage 10 (D) and stage 11.5 (E) gastrula stageXenopus embryos (animal pole towards the top). Insets demonstrateXphb1 expression in the mesendoderm (d) and in the posteriorneuroectoderm including mesoderm (e). No Xphb1 mRNA is detectablein comparable regions of sense control embryos (d�,e�). (F,G)Fromneurulation onwards (F, stage 23; G, stage 28; lateral view) Xphb1expression is restricted to the neural crest territory (arrowheads), the eyeand brain. (H-H�) Double ISH at stage 18 shows that Xphb1 (H, purple) ispartially co-expressed with twist (blue) in the neural crest (H�).(H�)Transverse section showing the colocalization of Xphb1 and twist(arrow); Xphb1 (purple arrowhead) and twist (blue arrowhead) are alsoexpressed in non-overlapping regions. Scale bars: 400mm. (I)RT-PCRanalysis of Xphb1 expression at the indicated stages. ornithinedecarboxylase (ODC) was used as an internal control. Additional controlswere performed without reverse transcriptase (–RT) and without cDNA(H2O).
Fig. 2. Xphb1 is required for CNC formation. (A)The phb1 MO,showing its binding site in Xphb1. Nucleotides that are identicalbetween Xphb1 and human PHB1 (Hphb1) or Xphb2 are indicated(asterisks). A standard control morpholino (co MO) tagged with 3�carboxyfluorescein was used as control. (B)In TnT in vitro translation,phb1 MO specifically blocks the translation of Xphb1 (lane 2). The phb1MO had no effect on the translation of Hphb1 (lane 3) or Xphb2 (lane6). (C)Injection (*) of phb1 MO, but not of co MO, efficiently inhibitedtwist expression. This was rescued by co-injection of Hphb1 mRNA orXphb1 mRNA lacking the morpholino binding site, whereas Xphb2mRNA showed no effect. b-gal was used as a lineage tracer (light blue).The co MO-injected side appears fluorescently labeled. Beneath isshown the percentage of Xenopus embryos with reduced twistexpression. n, number of embryos analyzed. **, P<0.005. Error barsindicate standard error. D
EVELO
PMENT

4076
The accumulation of Xphb1 in CNC prompted us to examinewhether depletion of Xphb1 affects neural crest development. Wedesigned an antisense morpholino (phb1 MO) that specificallyblocks Xphb1 translation while Xphb2 synthesis remains unaltered(Fig. 2A,B). Single-sided injections of phb1 MO at the 2-cell stageled to the loss of twist expression in the CNC (Fig. 2C). Co-injection of lacZ DNA served to identify the injected side. NeitherlacZ DNA alone nor control morpholino (co MO) injection affectedtwist expression, pointing to a specific phb1 MO effect. This wasconfirmed by rescue experiments. Both Xphb1 mRNA that lacksthe morpholino binding site and mRNA encoding the humanhomolog PHB1 (Hphb1), which shows 90% amino acid identity toXphb1, rescued the phb1 MO phenotype. Importantly, Xphb2mRNA injection did not rescue the depletion of Xphb1 (Fig. 2C).
Phb1 is known from mammalian cell culture studies to controlcell proliferation and apoptosis (Joshi et al., 2003; McClung et al.,1989; Wang et al., 1999a; Wang et al., 1999b). To address whetherloss of the twist signal is caused by reduced cell proliferation orincreased cell death we performed phospho-histone H3immunostainings and TUNEL assays. As shown in Fig. 3, phb1MO injections influenced neither cell proliferation nor apoptosis.This suggested that Xphb1 might be required in neural crest geneactivation, rather than in controlling CNC proliferation or celldeath.
Neural crest induction takes place during gastrulation anddepends on signals secreted from the dorsolateral mesoderm(Steventon et al., 2009). To exclude the possibility that Xphb1knockdown affects mesoderm formation and consequently neuralcrest induction, we examined the expression patterns of Xbra andchordin by ISH. Both genes were expressed normally upon Xphb1depletion (see Fig. S2 in the supplementary material).
Xphb1 operates downstream of c-myc inregulating snail2, foxD3 and twistNext, we aimed to better allocate Xphb1 into the regulatorynetwork of neural crest genes by expanding ISH studies of markergenes and by performing epistatic analyses. As shown in Fig. 4,depletion of Xphb1 resulted in loss of snail2 and foxD3, in additionto twist, but did not alter c-myc, snail1 or AP-2 expression.Reduction of twist, snail2 and foxD3 expression was observed innearly 80% of the embryos (see Fig. S3A,B in the supplementarymaterial). We further confirmed these findings by real-time PCR.twist mRNA was reduced by 50% and snail2 by 40% in phb1 MO-injected embryos; these effects were rescued by Hphb1 mRNA
injection (see Fig. S4A in the supplementary material). No changesin the amount of c-myc or snail1 transcripts were found in Xphb1morphants; a small decrease in AP-2 mRNA was detected by real-
RESEARCH ARTICLE Development 137 (23)
Fig. 3. Xphb1 regulates CNC formationindependently of proliferation and apoptosis.(A-C,E,F) Phospho-histone H3 staining (A-C) andTUNEL assay (E,F) of single side-injected (*)Xenopus embryos. Dextran-FITC marks the injectedside (C). (D,G)Quantification of phospho-histoneH3 staining (D) and TUNEL assay (G). The averagenumber of cells counted on the uninjected (n.i.)side of the embryo was set at 100%; n, number ofembryos analyzed. Neither proliferation norapoptosis was influenced after phb1 MO injection.Error bars indicate standard error.
Fig. 4. Xphb1 knockdown inhibits expression of the CNC markerssnail2 and foxD3 but not of c-myc, snail1 and AP-2. Depletion ofXphb1 mediated by phb1 MO injection (*) specifically inhibited snail2and foxD3 expression. Neural crest markers c-myc, snail1 and AP-2were not affected by Xphb1 knockdown. D
EVELO
PMENT

time PCR, but not by ISH (see Fig. S4B in the supplementarymaterial; compare with Fig. 4).
To examine whether c-myc acts upstream of Xphb1, embryoswere injected with antisense c-myc morpholino (c-myc MO). Therewas a decrease in Xphb1 CNC expression in 90% of the injectedembryos, as monitored by ISH (Fig. 5A). Real-time PCR showeda decrease of ~30% in Xphb1 mRNA (Fig. 5B). As expected,expression of snail2, foxD3 and twist was lost upon c-myc MOinjection. However, when Xphb1 mRNA was co-injected with c-myc MO, the expression of these genes was rescued. Xphb2 wasunable to rescue the effects of c-myc MO injection, confirming a
unique function of Xphb1 (Fig. 5C). Depletion of c-myc alsoresulted in loss of snail1 and AP-2 expression; the expression ofthese genes was not rescued by Xphb1 mRNA injection (Fig. 6).Thus, snail1 and AP-2 are regulated by c-myc independently ofXphb1, whereas c-myc-dependent expression of snail2, foxD3 andtwist requires Xphb1.
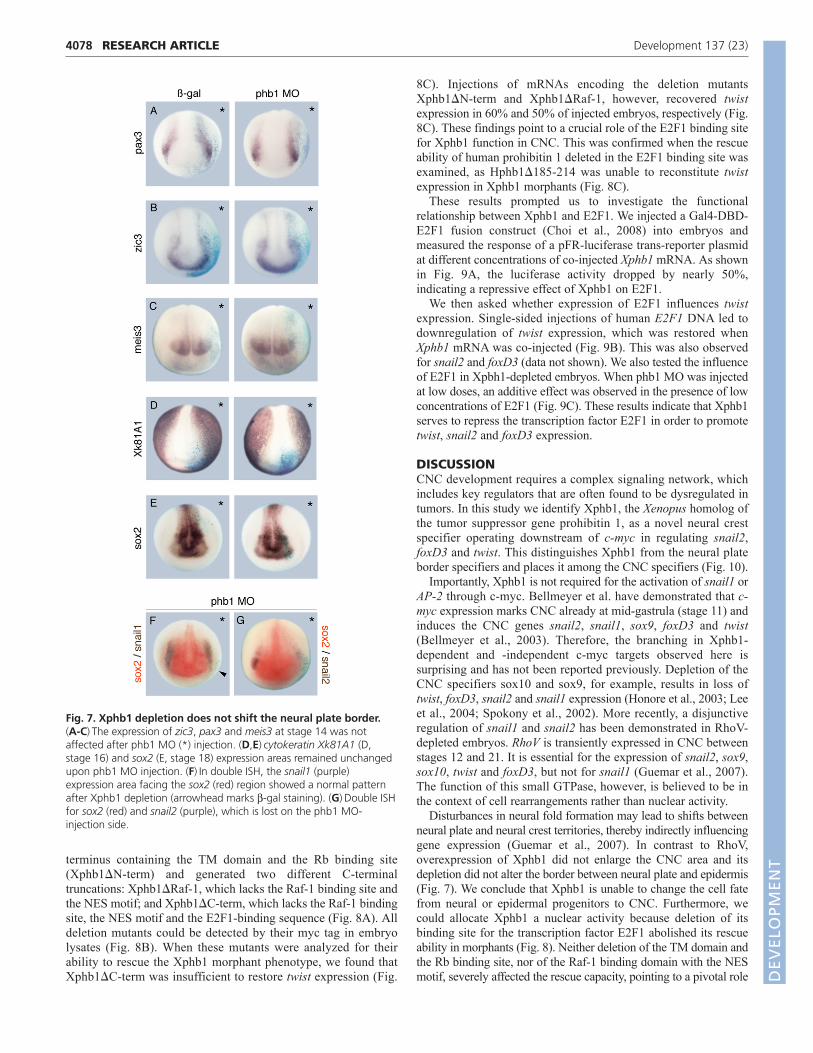
To separate Xphb1 clearly from the group of neural plate borderspecifiers we investigated whether Xphb1 knockdown alters theexpression of pax3, zic3 and meis3. The expression patterns ofthese neural plate border specifiers were unchanged upon Xphb1depletion (Fig. 7A-C). In line with these results, we did not observea broadening or reduction of the epidermal or neural tissue, asexamined by ISH for Xk81A1 (an epidermal marker) and sox2 (aneural marker) (Fig. 7D,E). Double ISH for sox2 and snail1 orsnail2 further confirmed that the ratio between neural plate andCNC areas remains stable in the absence of Xphb1 (Fig. 7F,G).
Xphb1 controls neural crest gene expressionthrough repression of E2F1Next, we aimed to determine whether a specific domain in Xphb1is required to regulate snail2, foxD3 and twist. Prohibitin1 consistsof a transmembrane domain (TM) required for mitochondriallocalization, as well as binding sites for the nuclear transcriptionfactors Rb and E2F1 (Mishra et al., 2006; Wang et al., 1999a;Wang et al., 1999b) (Fig. 8A). Human PHB1 has also beenreported to interact with the ring-finger protein 2 (RNF2) (Choi etal., 2008) and to bind RAF1 (Rajalingam et al., 2005; Wang et al.,1999b). Within the Raf-1 binding site, a nuclear export signal(NES) sequence is located (Mishra et al., 2006). We deleted the N-
4077RESEARCH ARTICLEXphb1 specifies neural crest by repressing E2F1
Fig. 5. Xphb1 acts downstream of c-myc and upstream of twist,snail2 and foxD3. (A)Injection of c-myc MO (*) efficiently inhibitedXphb1 expression (90%, n116). (B)Xenopus embryos injected with c-myc MO and analyzed by quantitative real-time PCR showed areduction in relative Xphb1 expression (normalized to the level of ODCexpression). **, P<0.005. (C)c-myc MO-mediated depletion of twist,snail2 and foxD3 was rescued by co-injection of Xphb1 mRNA (twist,**, P<0.005; snail2, *, P<0.05; foxD3, **, P<0.005), whereas Xphb2had no effect. n, number of embryos analyzed. Error bars indicatestandard error.
Fig. 6. Expression of snail1 and AP-2 is regulated by c-mycindependently of Xphb1. The expression of snail1 and AP-2 wasreduced by injection (*) of c-myc MO. This was not rescued by co-injection of Xphb1 mRNA. n, number of embryos analyzed. **,P<0.005. Error bars indicate standard error.
DEVELO
PMENT
4078
terminus containing the TM domain and the Rb binding site(Xphb1�N-term) and generated two different C-terminaltruncations: Xphb1�Raf-1, which lacks the Raf-1 binding site andthe NES motif; and Xphb1�C-term, which lacks the Raf-1 bindingsite, the NES motif and the E2F1-binding sequence (Fig. 8A). Alldeletion mutants could be detected by their myc tag in embryolysates (Fig. 8B). When these mutants were analyzed for theirability to rescue the Xphb1 morphant phenotype, we found thatXphb1�C-term was insufficient to restore twist expression (Fig.
8C). Injections of mRNAs encoding the deletion mutantsXphb1�N-term and Xphb1�Raf-1, however, recovered twistexpression in 60% and 50% of injected embryos, respectively (Fig.8C). These findings point to a crucial role of the E2F1 binding sitefor Xphb1 function in CNC. This was confirmed when the rescueability of human prohibitin 1 deleted in the E2F1 binding site wasexamined, as Hphb1�185-214 was unable to reconstitute twistexpression in Xphb1 morphants (Fig. 8C).
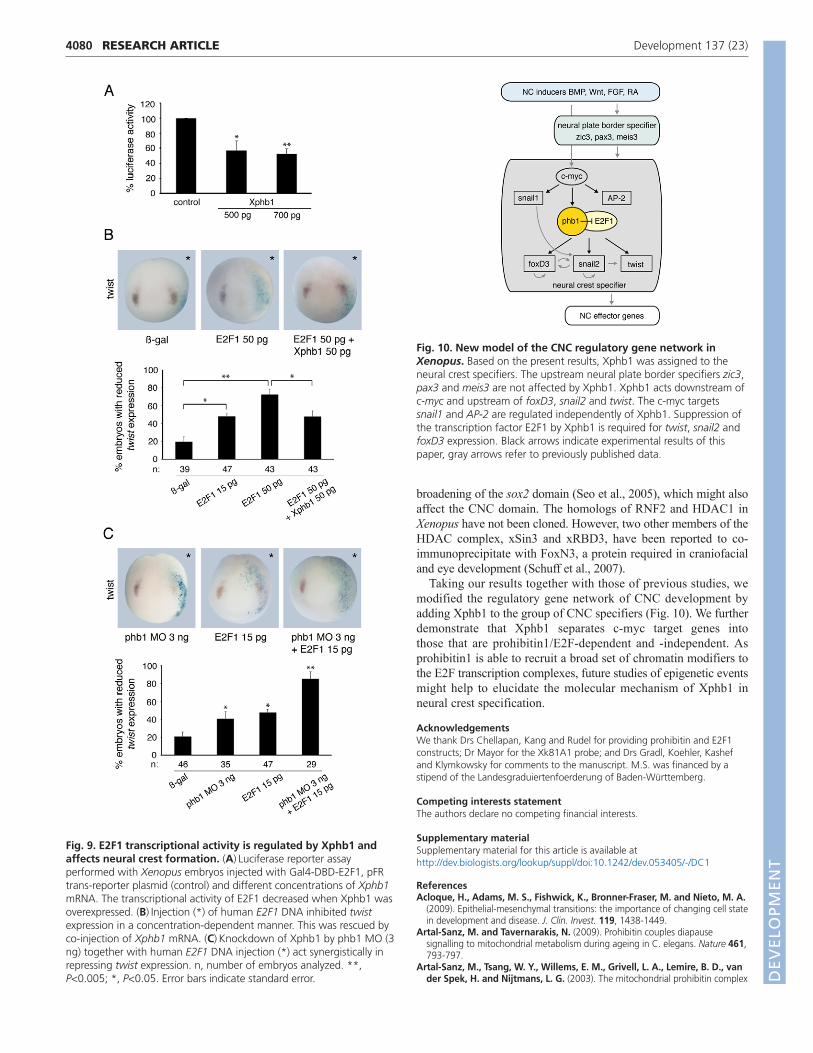
These results prompted us to investigate the functionalrelationship between Xphb1 and E2F1. We injected a Gal4-DBD-E2F1 fusion construct (Choi et al., 2008) into embryos andmeasured the response of a pFR-luciferase trans-reporter plasmidat different concentrations of co-injected Xphb1 mRNA. As shownin Fig. 9A, the luciferase activity dropped by nearly 50%,indicating a repressive effect of Xphb1 on E2F1.
We then asked whether expression of E2F1 influences twistexpression. Single-sided injections of human E2F1 DNA led todownregulation of twist expression, which was restored whenXphb1 mRNA was co-injected (Fig. 9B). This was also observedfor snail2 and foxD3 (data not shown). We also tested the influenceof E2F1 in Xpbh1-depleted embryos. When phb1 MO was injectedat low doses, an additive effect was observed in the presence of lowconcentrations of E2F1 (Fig. 9C). These results indicate that Xphb1serves to repress the transcription factor E2F1 in order to promotetwist, snail2 and foxD3 expression.
DISCUSSIONCNC development requires a complex signaling network, whichincludes key regulators that are often found to be dysregulated intumors. In this study we identify Xphb1, the Xenopus homolog ofthe tumor suppressor gene prohibitin 1, as a novel neural crestspecifier operating downstream of c-myc in regulating snail2,foxD3 and twist. This distinguishes Xphb1 from the neural plateborder specifiers and places it among the CNC specifiers (Fig. 10).
Importantly, Xphb1 is not required for the activation of snail1 orAP-2 through c-myc. Bellmeyer et al. have demonstrated that c-myc expression marks CNC already at mid-gastrula (stage 11) andinduces the CNC genes snail2, snail1, sox9, foxD3 and twist(Bellmeyer et al., 2003). Therefore, the branching in Xphb1-dependent and -independent c-myc targets observed here issurprising and has not been reported previously. Depletion of theCNC specifiers sox10 and sox9, for example, results in loss oftwist, foxD3, snail2 and snail1 expression (Honore et al., 2003; Leeet al., 2004; Spokony et al., 2002). More recently, a disjunctiveregulation of snail1 and snail2 has been demonstrated in RhoV-depleted embryos. RhoV is transiently expressed in CNC betweenstages 12 and 21. It is essential for the expression of snail2, sox9,sox10, twist and foxD3, but not for snail1 (Guemar et al., 2007).The function of this small GTPase, however, is believed to be inthe context of cell rearrangements rather than nuclear activity.
Disturbances in neural fold formation may lead to shifts betweenneural plate and neural crest territories, thereby indirectly influencinggene expression (Guemar et al., 2007). In contrast to RhoV,overexpression of Xphb1 did not enlarge the CNC area and itsdepletion did not alter the border between neural plate and epidermis(Fig. 7). We conclude that Xphb1 is unable to change the cell fatefrom neural or epidermal progenitors to CNC. Furthermore, wecould allocate Xphb1 a nuclear activity because deletion of itsbinding site for the transcription factor E2F1 abolished its rescueability in morphants (Fig. 8). Neither deletion of the TM domain andthe Rb binding site, nor of the Raf-1 binding domain with the NESmotif, severely affected the rescue capacity, pointing to a pivotal role
RESEARCH ARTICLE Development 137 (23)
Fig. 7. Xphb1 depletion does not shift the neural plate border.(A-C)The expression of zic3, pax3 and meis3 at stage 14 was notaffected after phb1 MO (*) injection. (D,E)cytokeratin Xk81A1 (D,stage 16) and sox2 (E, stage 18) expression areas remained unchangedupon phb1 MO injection. (F)In double ISH, the snail1 (purple)expression area facing the sox2 (red) region showed a normal patternafter Xphb1 depletion (arrowhead marks b-gal staining). (G)Double ISHfor sox2 (red) and snail2 (purple), which is lost on the phb1 MO-injection side.
DEVELO
PMENT

for the interaction with E2F1. Strikingly, E2F1 overexpression led toa decrease in twist, snail2 and foxD3 expression, which wasrecovered by overexpression of Xphb1. Depletion of Xphb1,conversely, promoted a reduction in twist expression through E2F1.These findings, and the decrease in Gal4-DBD-E2F1 activity inpresence of prohibitin in the pFR-luciferase reporter, suggest thatXphb1 is required to repress E2F1 function in CNC development.
In Xenopus, an E2F ortholog is maternally expressed. E2F mRNAspecifically localizes during neurula stage in brain, neural tube andneural crest (Suzuki and Hemmati-Brivanlou, 2000) and E2F seemsto be more widely used in regulating transcription than Xphb1. Thelatter is supported by our findings that E2F1 also reduces c-myc,snail1 and AP-2 expression and that this is not rescued by Xphb1(data not shown). Expression of dominant-negative E2F constructsresults in disturbances of ventral and posterior cell fates (Suzuki andHemmati-Brivanlou, 2000) and in the inhibition of cell cycleprogression from mid-blastula stage onwards (Tanaka et al., 2003).Based on our results from phospho-histone H3 immunostaining andTUNEL assay, we can exclude the possibility that Xphb1, togetherwith E2F1, balances cell proliferation in CNC development (Fig. 3).Furthermore, cell cycle control by prohibitins via binding E2F1 alsoinvolves Rb (Wang et al., 1999a; Wang et al., 1999b). Our Xphb1deletion mutants, however, point to an Rb-independent mechanismbecause the mutant lacking the Rb binding site rescues the Xphb1morphant. This is in line with a recent report which demonstrated
that neither overexpression nor depletion of Rb disturbs Xenopusdevelopment (Cosgrove and Philpott, 2007). They argue that inXenopus, Rb is hyperphosphorylated and therefore inactive untilstage 30 (Cosgrove and Philpott, 2007).
In mammalian cells, multiple mechanisms of prohibitin-mediatedrepression of E2F1 function have been reported, some of which areRb independent. Importantly, all of these mechanisms involvechromatin remodeling. Prohibitin1 is able to recruit histonedeacetylase 1 (HDAC1) and the co-repressor N-CoR into the E2F1transcription complex, thereby abolishing E2F1 function (Wang etal., 2002a). Prohibitin1 has also been shown to recruit Bg-1/Brm,which are part of the SWI/SNF chromatin remodeling complex(Wang et al., 2002b). Choi et al. reported that prohibitin1 bindsRNF2 and recruits it to E2F1-response promoters (Choi et al., 2008).RNF2 belongs to the polycomb family. The presence of RNF2 inpolycomb complexes correlates with histone H2A ubiquitylation(Wang et al., 2004). Therefore, it seems most likely that Xphb1promotes chromatin rearrangements that are essential for theactivation of those CNC specifier genes that contain E2F1-responseelements in their promoters. XBrg1, the catalytic subunit of theSWI/SNF complex, is contributed maternally in Xenopus embryos,but transcripts accumulate in the presumptive CNS, neural crest andin the otic vesicle (Seo et al., 2005). Loss-of-function studiesrevealed a requirement for XBrg1 in neurogenesis. Importantly,single-sided antisense XBrg1 morpholino injections yield a lateral
4079RESEARCH ARTICLEXphb1 specifies neural crest by repressing E2F1
Fig. 8. The E2F1-binding domain is required for CNC formation. (A)Model of prohibitin1 protein domains (according to the human proteinstructure). Beneath are shown the various Xphb1 mutants (Xphb1DN-term, Xphb1DRaf-1, Xphb1DC-term) and a human PHB1 mutant that lacks theE2F1 binding site (Hphb1D185-214). Deletions are indicated by the thin lines. TM, transmembrane domain; NES, nuclear export signal. (B)In westernblots, Xphb1 mutants were detected by anti-myc (9E10) antibody. Coomassie Brilliant Blue (CBB) staining was used to reveal equal loading. (C)Xphb1mutants and Hphb1D185-214 co-injected with phb1 MO (*) and analyzed by twist ISH showed that both of the mutants that lack the E2F1 binding sitewere unable to rescue twist expression. n, number of embryos analyzed. **, P<0.005; *, P<0.05. Error bars indicate standard error.
DEVELO
PMENT
4080
broadening of the sox2 domain (Seo et al., 2005), which might alsoaffect the CNC domain. The homologs of RNF2 and HDAC1 inXenopus have not been cloned. However, two other members of theHDAC complex, xSin3 and xRBD3, have been reported to co-immunoprecipitate with FoxN3, a protein required in craniofacialand eye development (Schuff et al., 2007).
Taking our results together with those of previous studies, wemodified the regulatory gene network of CNC development byadding Xphb1 to the group of CNC specifiers (Fig. 10). We furtherdemonstrate that Xphb1 separates c-myc target genes intothose that are prohibitin1/E2F-dependent and -independent. Asprohibitin1 is able to recruit a broad set of chromatin modifiers tothe E2F transcription complexes, future studies of epigenetic eventsmight help to elucidate the molecular mechanism of Xphb1 inneural crest specification.
AcknowledgementsWe thank Drs Chellapan, Kang and Rudel for providing prohibitin and E2F1constructs; Dr Mayor for the Xk81A1 probe; and Drs Gradl, Koehler, Kashefand Klymkowsky for comments to the manuscript. M.S. was financed by astipend of the Landesgraduiertenfoerderung of Baden-Württemberg.
Competing interests statementThe authors declare no competing financial interests.
Supplementary materialSupplementary material for this article is available athttp://dev.biologists.org/lookup/suppl/doi:10.1242/dev.053405/-/DC1
ReferencesAcloque, H., Adams, M. S., Fishwick, K., Bronner-Fraser, M. and Nieto, M. A.
(2009). Epithelial-mesenchymal transitions: the importance of changing cell statein development and disease. J. Clin. Invest. 119, 1438-1449.
Artal-Sanz, M. and Tavernarakis, N. (2009). Prohibitin couples diapausesignalling to mitochondrial metabolism during ageing in C. elegans. Nature 461,793-797.
Artal-Sanz, M., Tsang, W. Y., Willems, E. M., Grivell, L. A., Lemire, B. D., vander Spek, H. and Nijtmans, L. G. (2003). The mitochondrial prohibitin complex
RESEARCH ARTICLE Development 137 (23)
Fig. 9. E2F1 transcriptional activity is regulated by Xphb1 andaffects neural crest formation. (A)Luciferase reporter assayperformed with Xenopus embryos injected with Gal4-DBD-E2F1, pFRtrans-reporter plasmid (control) and different concentrations of Xphb1mRNA. The transcriptional activity of E2F1 decreased when Xphb1 wasoverexpressed. (B)Injection (*) of human E2F1 DNA inhibited twistexpression in a concentration-dependent manner. This was rescued byco-injection of Xphb1 mRNA. (C)Knockdown of Xphb1 by phb1 MO (3ng) together with human E2F1 DNA injection (*) act synergistically inrepressing twist expression. n, number of embryos analyzed. **,P<0.005; *, P<0.05. Error bars indicate standard error.
Fig. 10. New model of the CNC regulatory gene network inXenopus. Based on the present results, Xphb1 was assigned to theneural crest specifiers. The upstream neural plate border specifiers zic3,pax3 and meis3 are not affected by Xphb1. Xphb1 acts downstream ofc-myc and upstream of foxD3, snail2 and twist. The c-myc targetssnail1 and AP-2 are regulated independently of Xphb1. Suppression ofthe transcription factor E2F1 by Xphb1 is required for twist, snail2 andfoxD3 expression. Black arrows indicate experimental results of thispaper, gray arrows refer to previously published data.
DEVELO
PMENT

is essential for embryonic viability and germline function in Caenorhabditiselegans. J. Biol. Chem. 278, 32091-32099.
Bang, A. G., Papalopulu, N., Kintner, C. and Goulding, M. D. (1997). Expressionof Pax-3 is initiated in the early neural plate by posteriorizing signals produced bythe organizer and by posterior non-axial mesoderm. Development 124, 2075-2085.
Basch, M. L. and Bronner-Fraser, M. (2006). Neural crest inducing signals. Adv.Exp. Med. Biol. 589, 24-31.
Bellmeyer, A., Krase, J., Lindgren, J. and LaBonne, C. (2003). The protooncogenec-myc is an essential regulator of neural crest formation in xenopus. Dev. Cell 4,827-839.
Carl, T. F., Dufton, C., Hanken, J. and Klymkowsky, M. W. (1999). Inhibition ofneural crest migration in Xenopus using antisense slug RNA. Dev. Biol. 213, 101-115.
Choi, D., Lee, S. J., Hong, S., Kim, I. H. and Kang, S. (2008). Prohibitin interactswith RNF2 and regulates E2F1 function via dual pathways. Oncogene 27, 1716-1725.
Cosgrove, R. A. and Philpott, A. (2007). Cell cycling and differentiation do notrequire the retinoblastoma protein during early Xenopus development. Dev. Biol.303, 311-324.
Essex, L. J., Mayor, R. and Sargent, M. G. (1993). Expression of Xenopus snail inmesoderm and prospective neural fold ectoderm. Dev. Dyn. 198, 108-122.
Etard, C., Gradl, D., Kunz, M., Eilers, M. and Wedlich, D. (2005). Pontin andReptin regulate cell proliferation in early Xenopus embryos in collaboration with c-Myc and Miz-1. Mech. Dev. 122, 545-556.
Fusaro, G., Wang, S. and Chellappan, S. (2002). Differential regulation of Rbfamily proteins and prohibitin during camptothecin-induced apoptosis. Oncogene21, 4539-4548.
Gawantka, V., Delius, H., Hirschfeld, K., Blumenstock, C. and Niehrs, C. (1995).Antagonizing the Spemann organizer: role of the homeobox gene Xvent-1. EMBOJ. 14, 6268-6279.
Gradl, D., Kuhl, M. and Wedlich, D. (1999). The Wnt/Wg signal transducer beta-catenin controls fibronectin expression. Mol. Cell. Biol. 19, 5576-5587.
Guemar, L., de Santa Barbara, P., Vignal, E., Maurel, B., Fort, P. and Faure, S.(2007). The small GTPase RhoV is an essential regulator of neural crest induction inXenopus. Dev. Biol. 310, 113-128.
Hensey, C. and Gautier, J. (1998). Programmed cell death during Xenopusdevelopment: a spatio-temporal analysis. Dev. Biol. 203, 36-48.
Herold, S., Herkert, B. and Eilers, M. (2009). Facilitating replication under stress:an oncogenic function of MYC? Nat. Rev. Cancer 9, 441-444.
Hoffman, B. and Liebermann, D. A. (2008). Apoptotic signaling by c-MYC.Oncogene 27, 6462-6472.
Hong, C. S. and Saint-Jeannet, J. P. (2007). The activity of Pax3 and Zic1 regulatesthree distinct cell fates at the neural plate border. Mol. Biol. Cell 18, 2192-2202.
Honore, S. M., Aybar, M. J. and Mayor, R. (2003). Sox10 is required for the earlydevelopment of the prospective neural crest in Xenopus embryos. Dev. Biol. 260,79-96.
Hopwood, N. D., Pluck, A. and Gurdon, J. B. (1989). A Xenopus mRNA related toDrosophila twist is expressed in response to induction in the mesoderm and theneural crest. Cell 59, 893-903.
Jonas, E., Sargent, T. D. and Dawid, I. B. (1985). Epidermal keratin gene expressedin embryos of Xenopus laevis. Proc. Natl. Acad. Sci. USA 82, 5413-5417.
Joshi, B., Ko, D., Ordonez-Ercan, D. and Chellappan, S. P. (2003). A putativecoiled-coil domain of prohibitin is sufficient to repress E2F1-mediated transcriptionand induce apoptosis. Biochem. Biophys. Res. Commun. 312, 459-466.
Kishi, M., Mizuseki, K., Sasai, N., Yamazaki, H., Shiota, K., Nakanishi, S. andSasai, Y. (2000). Requirement of Sox2-mediated signaling for differentiation ofearly Xenopus neuroectoderm. Development 127, 791-800.
Kuriyama, S. and Mayor, R. (2008). Molecular analysis of neural crest migration.Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 1349-1362.
Lee, Y. H., Aoki, Y., Hong, C. S., Saint-Germain, N., Credidio, C. and Saint-Jeannet, J. P. (2004). Early requirement of the transcriptional activator Sox9 forneural crest specification in Xenopus. Dev. Biol. 275, 93-103.
Li, B., Kuriyama, S., Moreno, M. and Mayor, R. (2009). The posteriorizing geneGbx2 is a direct target of Wnt signalling and the earliest factor in neural crestinduction. Development 136, 3267-3278.
Luo, T., Lee, Y. H., Saint-Jeannet, J. P. and Sargent, T. D. (2003). Induction ofneural crest in Xenopus by transcription factor AP2alpha. Proc. Natl. Acad. Sci.USA 100, 532-537.
Mayor, R., Morgan, R. and Sargent, M. G. (1995). Induction of the prospectiveneural crest of Xenopus. Development 121, 767-777.
McClung, J. K., Danner, D. B., Stewart, D. A., Smith, J. R., Schneider, E. L.,Lumpkin, C. K., Dell’Orco, R. T. and Nuell, M. J. (1989). Isolation of a cDNAthat hybrid selects antiproliferative mRNA from rat liver. Biochem. Biophys. Res.Commun. 164, 1316-1322.
Meulemans, D. and Bronner-Fraser, M. (2004). Gene-regulatory interactions inneural crest evolution and development. Dev. Cell 7, 291-299.
Mishra, S., Murphy, L. C. and Murphy, L. J. (2006). The prohibitins: emerging rolesin diverse functions. J. Cell. Mol. Med. 10, 353-363.
Morrish, F., Isern, N., Sadilek, M., Jeffrey, M. and Hockenbery, D. M. (2009). c-Myc activates multiple metabolic networks to generate substrates for cell-cycleentry. Oncogene 28, 2485-2491.
Nakata, K., Nagai, T., Aruga, J. and Mikoshiba, K. (1997). Xenopus Zic3, aprimary regulator both in neural and neural crest development. Proc. Natl. Acad.Sci. USA 94, 11980-11985.
Nieuwkoop, P. D. and Faber, J. (1967). Normal Table of Xenopus laevis (Daudin),2nd edn. Amsterdam: North Holland.
Osman, C., Merkwirth, C. and Langer, T. (2009). Prohibitins and the functionalcompartmentalization of mitochondrial membranes. J. Cell Sci. 122, 3823-3830.
Rajalingam, K., Wunder, C., Brinkmann, V., Churin, Y., Hekman, M., Sievers,C., Rapp, U. R. and Rudel, T. (2005). Prohibitin is required for Ras-induced Raf-MEK-ERK activation and epithelial cell migration. Nat. Cell Biol. 7, 837-843.
Salzberg, A., Elias, S., Nachaliel, N., Bonstein, L., Henig, C. and Frank, D.(1999). A Meis family protein caudalizes neural cell fates in Xenopus. Mech. Dev.80, 3-13.
Sasai, N., Mizuseki, K. and Sasai, Y. (2001). Requirement of FoxD3-classsignaling for neural crest determination in Xenopus. Development 128, 2525-2536.
Sasai, Y., Lu, B., Steinbeisser, H. and De Robertis, E. M. (1995). Regulation ofneural induction by the Chd and Bmp-4 antagonistic patterning signals inXenopus. Nature 377, 757.
Sauka-Spengler, T. and Bronner-Fraser, M. (2008). A gene regulatory networkorchestrates neural crest formation. Nat. Rev. Mol. Cell Biol. 9, 557-568.
Schuff, M., Rossner, A., Wacker, S. A., Donow, C., Gessert, S. and Knochel, W.(2007). FoxN3 is required for craniofacial and eye development of Xenopus laevis.Dev. Dyn. 236, 226-239.
Seo, S., Richardson, G. A. and Kroll, K. L. (2005). The SWI/SNF chromatinremodeling protein Brg1 is required for vertebrate neurogenesis and mediatestransactivation of Ngn and NeuroD. Development 132, 105-115.
Sive, H. L., Grainger, R. M. and Harland, R. M. (2000). Fate mapping and lineagelabeling. In Early Development of Xenopus laevis: a Laboratory Manual (ed. S.Curtis and M. Cozza), pp.143-170. Cold Spring Harbor, NY: Cold Spring HarborLaboratory Press.
Smith, J. C., Price, B. M., Green, J. B., Weigel, D. and Herrmann, B. G. (1991).Expression of a Xenopus homolog of Brachyury (T) is an immediate-early responseto mesoderm induction. Cell 67, 79-87.
Spokony, R. F., Aoki, Y., Saint-Germain, N., Magner-Fink, E. and Saint-Jeannet,J. P. (2002). The transcription factor Sox9 is required for cranial neural crestdevelopment in Xenopus. Development 129, 421-432.
Steventon, B., Carmona-Fontaine, C. and Mayor, R. (2005). Genetic networkduring neural crest induction: from cell specification to cell survival. Semin. CellDev. Biol. 16, 647-654.
Steventon, B., Araya, C., Linker, C., Kuriyama, S. and Mayor, R. (2009).Differential requirements of BMP and Wnt signalling during gastrulation andneurulation define two steps in neural crest induction. Development 136, 771-779.
Suzuki, A. and Hemmati-Brivanlou, A. (2000). Xenopus embryonic E2F is requiredfor the formation of ventral and posterior cell fates during early embryogenesis.Mol. Cell 5, 217-229.
Tanaka, T., Ono, T., Kitamura, N. and Kato, J. Y. (2003). Dominant negative E2Finhibits progression of the cell cycle after the midblastula transition in Xenopus.Cell Struct. Funct. 28, 515-522.
Turner, D. L. and Weintraub, H. (1994). Expression of achaete-scute homolog 3 inXenopus embryos converts ectodermal cells to a neural fate. Genes Dev. 8, 1434-1447.
Unterseher, F., Hefele, J. A., Giehl, K., De Robertis, E. M., Wedlich, D. andSchambony, A. (2004). Paraxial protocadherin coordinates cell polarity duringconvergent extension via Rho A and JNK. EMBO J. 23, 3259-3269.
Wang, H., Wang, L., Erdjument-Bromage, H., Vidal, M., Tempst, P., Jones, R. S.and Zhang, Y. (2004). Role of histone H2A ubiquitination in Polycomb silencing.Nature 431, 873-878.
Wang, S., Nath, N., Adlam, M. and Chellappan, S. (1999a). Prohibitin, a potentialtumor suppressor, interacts with RB and regulates E2F function. Oncogene 18,3501-3510.
Wang, S., Nath, N., Fusaro, G. and Chellappan, S. (1999b). Rb and prohibitintarget distinct regions of E2F1 for repression and respond to different upstreamsignals. Mol. Cell. Biol. 19, 7447-7460.
Wang, S., Fusaro, G., Padmanabhan, J. and Chellappan, S. P. (2002a). Prohibitinco-localizes with Rb in the nucleus and recruits N-CoR and HDAC1 fortranscriptional repression. Oncogene 21, 8388-8396.
Wang, S., Zhang, B. and Faller, D. V. (2002b). Prohibitin requires Brg-1 and Brm forthe repression of E2F and cell growth. EMBO J. 21, 3019-3028.
Zhang, C. and Klymkowsky, M. W. (2009). Unexpected functional redundancybetween Twist and Slug (Snail2) and their feedback regulation of NF-kappaB viaNodal and Cerberus. Dev. Biol. 331, 340-349.
Zhang, C., Carl, T. F., Trudeau, E. D., Simmet, T. and Klymkowsky, M. W.(2006). An NF-kappaB and slug regulatory loop active in early vertebratemesoderm. PLoS ONE 1, e106.
4081RESEARCH ARTICLEXphb1 specifies neural crest by repressing E2F1
DEVELO
PMENT





























