Embed Size (px)
Citation preview

Original article
Performance of vegetatively propagatedLarix decidua, L kaempferi and L laricina hybrids
LE Pâques
INRA, Centre d’Orléans, Station d’Amélioration des Arbres Forestiers, 45160 Ardon, France
(Received 25 January 1991; accepted 17 October 1991)
Summary — Interspecific hybridization between the tamarack and the Japanese and Europeanlarches was attempted. Successful crosses between the tamarack and the Japanese larch as wellas the 3-way hybrid (tamarack crossed to European x Japanese larches) were obtained for the firsttime in France but on a limited scale. Hybridization with tamarack seems possible but rather difficultdue to differences in phenology and reproduction potential. Hybrid clones were successfully propa-gated by stem cuttings with high rates of rooting and a good quality of the root system, especially forthe tamarack and its hybrids. Field performances at 8 years reveal that even if their initial growth isvigorous is absolute terms, neither the tamarack nor its hybrids could compete with the hybrid Larixdecidua x L kaempferi for both growth and stem form. One exception is the 3-way hybrid L laricina x(L decidua x L kaempferi) which can be advantageously compared to the latter. An additional advan-tage could be its expected greater tolerance of wet soils. Nevertheless, its creation appeared to bedifficult. So far, the hybrid L decidua x L kaempferi seems to be best for reforestation.
Larix / larch / tamarack / interspecific hybridization / vegetative propagation I clonal test /
stem cutting
Résumé — Comparaison des performances de différents hybrides entre les mélèzes laricinsd’Europe et du Japon multipliés végétatlvement. L’hybridization interspécifique offre à l’améliora-teur des perspectives intéressantes pour la création variétale, notamment pour la combinaison decaractères complémentaires favorables et par valorisation de l’hétérosis. Dans cette perspective,plusieurs croisements contrôlés entre le mélèze laricin (Larix laricina) - adapté aux sols hydro-morphes - et les mélèzes d’Europe (L decidua) et du Japon (L kaempferi) ont été tentés, certainsavec succès tels le croisement entre le mélèze laricin et le mélèze du Japon et celui de l’hybridetriple (L laricina x (L decidua x L kaempferi)). Bien que possible, l’hybridation avec le mélèze laricinest rendue difficile par de très nombreux problèmes de floraison, entre autres : décalage phénologi-que, virescence, absence de pollen. Les différents hybrides obtenus ont ensuite été multipliés parbouturage horticole avec des taux d’enracinement élevés et une bonne qualité du système racinaire.En particulier, le bouturage du mélèze laricin et de ses hybrides se révèle encore plus facile quecelui des hybrides entre le mélèze d’Europe et du Japon. Les premiers résultats en forêt à 8 ansmontrent que malgré une croissance initiale très vigoureuse en termes absolus, ni le mélèze laricinni son hybride avec le mélèze du Japon ne supportent la comparaison avec l’hybride L decidua xL kaempferi. Seul l’hybride triple apparaît prometteur avec une vigueur et une qualité de forme du fûtcomparables. Néanmoins, la difficulté de son obtention limite son intérêt pratique à moins qu’il ne serévèle plus tolérant à l’hydromorphie du sol que son parent hybride.
Larix / mélèze / hybridation interspécifique / bouturage / test clonal

INTRODUCTION
Among the Larix species, two are of com-mon use for reforestation in western Eu-
rope, namely the European larch (Larix de-cidua Mill) and the Japanese larch (Larixkaempferi (Lamb) Carr = Larix leptolepis(Sieb and Zucc) Gord). A third one, ie thetamarack, a species from North America(Larix laricina (Du Roi) K Koch), has beenconsidered too as a potentially valuablespecies because of its tolerance to water-logged soils in its natural range. So far, its
plantation has been largely restricted in
Europe to botanical gardens and experi-ments.
As a result of certain biological limita-tions of each of these species (eg cankersensitivity of Larix decidua, poor stem formand summer drought sensitivity of Larix
kaempferi and poor growth and stem formof Larix laricina; Pâques, in preparation),great emphasis has been put on interspe-cific hybridization in French tree improve-ment programs.
Interspecific hybridization is used bothto develop and combine favourable com-plementary traits from both parent speciesas well as to take advantage of hybrid vi-gour or heterosis.
Hybridization between Larix species isdocumented by numerous natural and arti-ficial interspecific crosses (Bobrov, 1973,cited in Wright, 1976; Avrov, 1982). No in-compatibility barriers, more severe thanthose observed within a species (corres-ponding to low filled seed sets (Kosinski,1987)), seem to exist. Successful crosseshave been made even between geographi-cally distant species as reported by SyrachLarsen (1937) and Delevoy (1949).Among the three Larix species mentio-
ned above, hybridization between the Eu-ropean and the Japanese larches is most
commonly used in tree breeding and its
feasibility and interest have been docu-
mented for nearly a century (Henry andFlood, 1919).
Hybridization between tamarack and ei-ther the European or the Japanese larchesseems to be possible as reported byChowdbury (1931) and Syrack Larsen
(1937) for Larix laricina and Larix decidua,and by MacGillivray (1967) for Larix larici-na and Larix kaempferi. Nevertheless, assuggested by MacGillivray’s (1967) obser-vations, crossing between tamarack andthe Japanese larch was quite unsuccessfuland no seed could be obtained from reci-
procal mating. Moreover, though Avrov
(1982) characterizes crossability betweentamarack and either the European or theJapanese larches as ’partial’, he could getno filled seed from crosses between tama-rack and the Japanese larch and no re-
sults are presented from crosses includingthe European larch.
Several matings including the Europeanand Japanese larches on the one handand the tamarack on the other were at-
tempted in this exploratory study. Objec-tives were to test their interspecific cross-ability and if positive, to study the respec-tive hybrids for use in reforestation. In par-ticular, it was interesting to compare theirperformances with those of pure larch spe-cies.
MATERIALS AND METHODS
Hybridization and material
Hybridization included crosses between Larixdecidua and L kaempferi (coded as ’dec x kae’)and its reciprocal: L kaempferi x L decidua (’kaex dec’). F2 hybrids were also attempted by cros-sing several F1 hybrid clones (’(dec x kae) X(dec x kae)’). Tamarack was involved in three
crossings: L laricina x L decidua (’lar x dec’),L laricina x L kaempferi (’lar x kae’) and the 3-way hybrid: L laricina x (L decidua x L kaempfe-ri) (’lar X (dec x kae’).

Hybridization through controlled crossingtook place at the INRA Forest Tree Improve-ment Research Station in Orléans in spring1979. Hybrids ’lar X (dec x kae)’ and ’(dec x kae)X (dec x kae)’ shared the same father hybridclone. Different tamarack mother clones wereused for obtaining hybrids ’lar x kae’ and ’lar X(dec x kae)’.
Successful hybrids were then field tested andcompared to one provenance of Larix kaempferi(coded as ’kae’) and to two provenances of Larixlaricina (’lar’). No pure European larch was pre-sent in the experiment. In addition, two morespecies were also used as controls of traditionalreforestation material, namely Norway spruce(Picea abies (L) Karst, represented by two pro-venances of southern Poland origin) and Dou-glas fir (Pseudotsuga menziesii (Mirb) Franco,by one provenance from Arlington, Washington).They will be referred to in tables and figures by’P ab’ and’P menz’ respectively. Provenances ofboth species have been recommended for refo-restation in France because of their outstandingperformances. Japanese larch, Norway spruceand Douglas fir are represented by open pollina-ted material collected in natural forest stands.
A brief description of the material is given intable I. It presents only successful hybrids.
Vegetative propagation
Prior to field plantation, part of the materialunder study (ie larch hybrids and tamarack) wasvegetatively propagated by stem cuttings andpart by seedlings (ie Japanese larch, Norwayspruce and Douglas fir). Vegetative propagationhad two objectives: first, to multiply poor repre-sented hybrid material, and secondly, to studythe feasibility of the technique.
Ortets of the different larch hybrids and of ta-marack were selected for total height at 2 yearsin a progeny test. The proportion of ortets
(clones) selected ranged from 1.5 to 35.0% ofeach family. Nevertheless, for hybrids ’lar x kae’,’lar x ’(dec x kae)’ and ’(dec x kae) x (dec xkae)’, all the seedlings were used as stock
plants due to the low number of available seed-lings. In total, 184 ortets were finally chosen.Further details about the origin of the materialare given in table I.
Ortets were potted in autumn 1981 and for-ced under greenhouse conditions before propa-gation.
An average of 30 softwood cuttings per clone(about 10 cm long) were taken in May 1982 anddipped in a 0.5% IBA solution including a fungi-

cide. They were rooted in a medium mixture (2-1-3 in volume) of peat, compost and pouzzolane(volcanic ash) under greenhouse conditions. Anintermittent mist was used and alternating fungi-cide treatments were applied as necessary.
Rooted cuttings were then transferred to theINRA nursery in March 1983 for two years be-fore being field planted while the control species(’kae’, ’P ab’, and ’P menz’) were sown in thesame nursery in May 1983.
Field experimental design
Three-year old rooted cuttings and two-year oldseedlings were planted together in the MassifCentral Mountains during the winter of 1984.Two tests were established: the major one inthe State Forest of Eclache (Puy de Dôme,2°41’E, 45°44’N, 1 000 m), and a minor one inPeyrat-Le-Château (Haute-Vienne, 1 °44’E,45°49’N, 450 m). Because of low representationof certain hybrids in the latter, only the formerwill be considered.
Eclache is a good coniferous forest site witha deep brown forest soil and high precipitationwell distributed throughout the year (averageannual rainfall up to 1 000 mm). Nevertheless,because of the rather short growing season andparticularly windy conditions, the elevation ofthis site is the upper limit where larch can be
planted.An incomplete, balanced randomized block
design was used with 9 blocks, 95 plants perblock and two-tree random non-contiguous plots(see Libby and Cockerham, 1980) per geneticentry and per block. Spacing was 3 by 3 meters.No soil preparation and fertilisation were appliedprior to plantation.
Measurements
The percentage of rooted cuttings per clonewas estimated and the quality of the root sys-tem was assessed according to the followingscale: 1 = numerous roots well distributedaround the stem base; 2 = intermediate; 3 =poor root system with one or two roots and aone-sided distribution; 4 = no roots.
In addition, the number of rooted cuttings perclone judged to be plantable was based on thefrequency of score 1 and 2 for the quality of theroot system and on further survival of ramets inthe nursery.
First measurements in the field took place in1989, 8 years after propagation. They concernsurvival rate (%S), total height (H), the length ofthe 1989 height increment (HI) and heightgrowth of the last 4 years, from 1986 to 1989(HG4)-which corresponds to the growth in theforest after overcoming planting shock. Stemform quality was evaluated by the frequency ofbasal sweep (%BS) and a subjective assess-ment of stem straightness (SS) on a 5-pointscale: class 1 = severe crookedness with morethan two crooks per stem; class 2 = same as 1but no more than one or two crooks per stem;class 3 = light crookedness with more than twocrooks per stem; class 4 = same as 3 but nomore than one or two crooks per stem; and,class 5 = straight stem. The frequency of
straight stems (%SS) for each clone was thenestimated from frequency of ramets score forstem straightness greater than 3. Description ofstem straightness follows Keiding and Olsen’sclassification (1965) but in the reverse order.
When possible, an analysis of variance wasconducted on individual data following themodel:
where
Yijkl = l th observation of clone k of species(’species’ will be used in this paper as a genericterm to design both pure larch species and inter-specific hybrids) j in block i
μ = general mean
Bi = effect of block i (fixed effect),
Sj = effect of species j (fixed effect),
BSij = block x species interaction,
Ck(j) = effect of clone k within species j (randomeffect),
ϵijkl = error term.
Prior to analysis, homogeneity of the pheno-typic variance-covariance matrices of each ge-netic entry was tested according to Kullback’stest (Legendre and Legendre, 1984) in order toverify whether data from the different genetic en-

tries could be included in the same analysis orshould be analysed separately.
Species means were then compared follow-ing Bonferroni’s test after adjustment of data forsignificant block (and interaction) effects if any,and a second analysis of variance on adjusteddata. For qualitative variables (eg %S, %SS),comparisons between species were based onthe original observations according to the 2I orG-test as defined by Sokal and Rohlf (1981). Ittests the complete independence between twodescriptors. Based on contingency tables, 2I is
computed following Arbonnier (1966):
where
xij = number of observations for genetic entry iand score j (0/1),Xi. = marginal sum of observations for species i
X.j = marginal sum of observations for score j,k = number of species in comparison,n = number of scores,
N = total number of observations.
2I is then compared to Chi2 distribution valueswith (k-1) (n-1) degrees of freedom. Used in aniterative way, it allows association of the diffe-rent species in homogeneous groups.
RESULTS
Hybridization success
In addition to crosses between Europeanand Japanese larches (88% of which gavefilled seed with rates of filled seed rangingfrom 16 to 76%), those with tamarack werealso partly successful. These includedcrosses between tamarack and the Japa-nese larch (’lar x kae’) and between tama-rack and the F1 hybrid ’dec x kae’.
The absence of hybrids between tama-rack and the European larch is explainedby the fact that under Orléans conditions,tamarack flowers earlier than the Euro-
pean larch and must therefore be used asa female. Tamarack clones produced noviable pollen and stored pollen of Euro-
pean larch was not available at that time.
Because of a very low reproduction po-tential of tamarack under Orléans condi-tions (absence of pollen, numerous prolife-rated strobili and high rates of emptyseed), only one full-sib family per success-ful tamarack hybrid type (’lar x kae’ and ’larX (dec x kae)’) could be produced on twodifferent mother clones. Moreover, thenumber of viable seedlings obtained up tothe nursery stage was low (for ’lar x kae’, 2seedlings out of 623 seeds and 41 clones;for ’lar x (dec x kae)’, 13 seedlings out of56 seeds and 330 clones). This resultconfirms Avrov’s (1982) and MacGillivray’s(1967) observations on the low fertility ofthis type of crossings. For example, Mac-Gillivray’s results on growth performancesof tamarack x Japanese larch are basedon only four viable seedlings.
Rootability and quality of root system
More than 5 200 cuttings were struck inthe rooting medium and nearly 82% roo-ted. All 184 tested clones were successful-
ly rooted, with rooting percentages rangingfrom 9 to 100.
Rate of success was also very variablefrom hybrid to hybrid (table II) with the low-est values for ’kae x dec’ and ’(dec x kae)X (dec x kae)’. 2I-test reveals a heteroge-nous response of rooting ability betweenspecies (21 = 228 01 > χ20.95, 15 df = 25.0)with on one hand ’dec x kae’, ’lar’ and ’lar X(dec x kae)’ associated with the highestrooting rates and on the other hand, ’lar xkae’, ’(dec x kae) X (dec x kae)’ and ’kae xdec’ with the lowest.
As a whole, 62% of the rooted cuttingshad an excellent root system and morethan 88% were judged satisfactory (scores
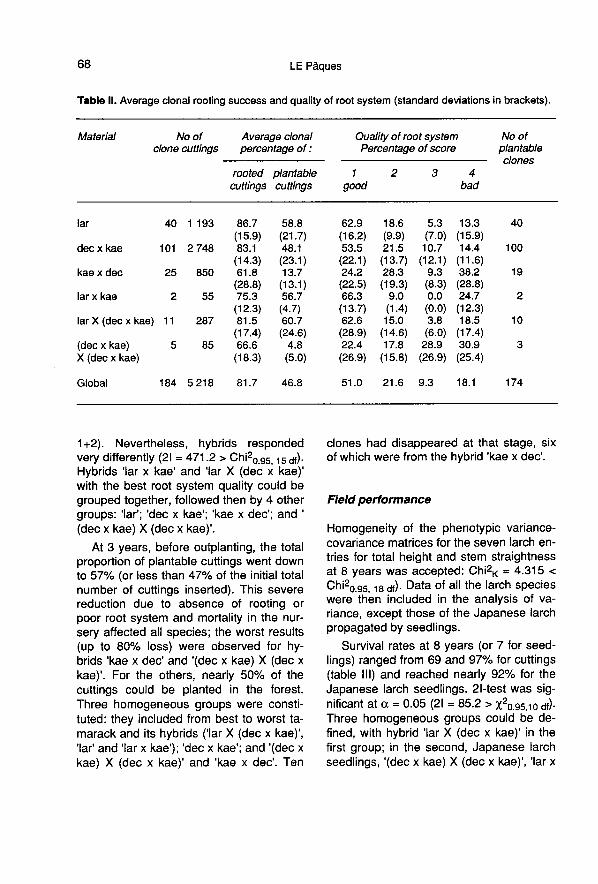
1+2). Nevertheless, hybrids respondedvery differently (21 = 471.2 > Chi20.95, 15 df).Hybrids ’lar x kae’ and ’lar X (dec x kae)’with the best root system quality could begrouped together, followed then by 4 othergroups: ’lar’; ’dec x kae’; ’kae x dec’; and ’(dec x kae) X (dec x kae)’.
At 3 years, before outplanting, the totalproportion of plantable cuttings went downto 57% (or less than 47% of the initial totalnumber of cuttings inserted). This severereduction due to absence of rooting or
poor root system and mortality in the nur-sery affected all species; the worst results(up to 80% loss) were observed for hy-brids ’kae x dec’ and ’(dec x kae) X (dec xkae)’. For the others, nearly 50% of thecuttings could be planted in the forest.
Three homogeneous groups were consti-tuted: they included from best to worst ta-marack and its hybrids (’lar X (dec x kae)’,’lar’ and ’lar x kae’); ’dec x kae’; and ’(dec xkae) X (dec x kae)’ and ’kae x dec’. Ten
clones had disappeared at that stage, sixof which were from the hybrid ’kae x dec’.
Field performance
Homogeneity of the phenotypic variance-covariance matrices for the seven larch en-tries for total height and stem straightnessat 8 years was accepted: Chi2K = 4.315 <
Chi20.95, 18 df). Data of all the larch specieswere then included in the analysis of va-riance, except those of the Japanese larchpropagated by seedlings.
Survival rates at 8 years (or 7 for seed-lings) ranged from 69 and 97% for cuttings(table III) and reached nearly 92% for theJapanese larch seedlings. 2I-test was sig-nificant at α = 0.05 (2I = 85.2 > χ20.95,10 df).Three homogeneous groups could be de-fined, with hybrid ’lar X (dec x kae)’ in thefirst group; in the second, Japanese larchseedlings, ’(dec x kae) X (dec x kae)’, ’lar x
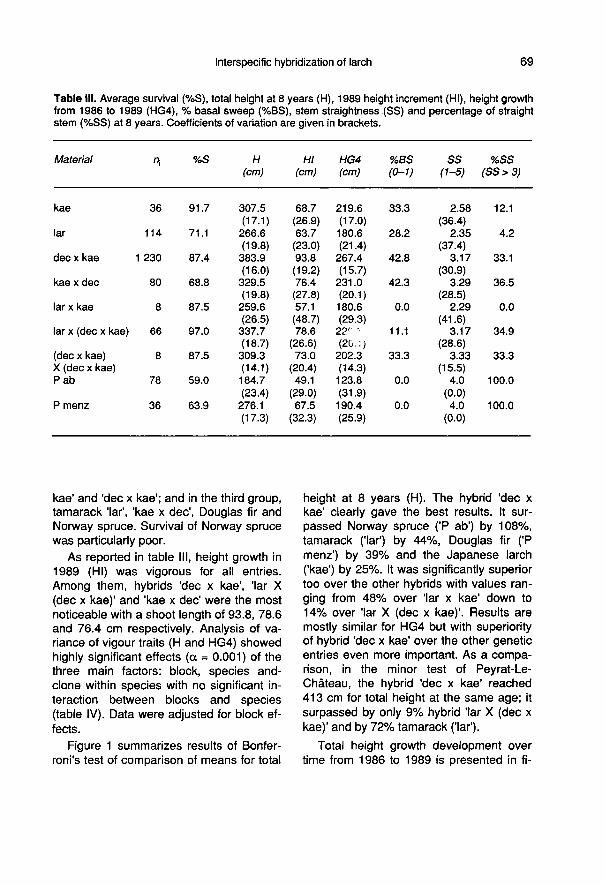
kae’ and ’dec x kae’; and in the third group,tamarack ’lar’, ’kae x dec’, Douglas fir andNorway spruce. Survival of Norway sprucewas particularly poor.
As reported in table III, height growth in1989 (HI) was vigorous for all entries.
Among them, hybrids ’dec x kae’, ’lar X
(dec x kae)’ and ’kae x dec’ were the mostnoticeable with a shoot length of 93.8, 78.6and 76.4 cm respectively. Analysis of va-riance of vigour traits (H and HG4) showedhighly significant effects (α = 0.001) of thethree main factors: block, species and-clone within species with no significant in-teraction between blocks and species(table IV). Data were adjusted for block ef-fects.
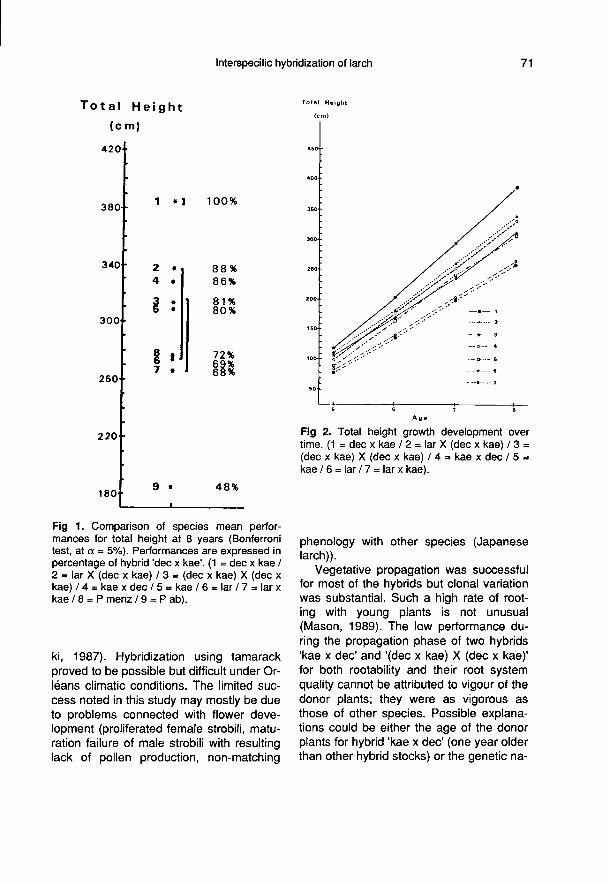
Figure 1 summarizes results of Bonfer-roni’s test of comparison of means for total
height at 8 years (H). The hybrid ’dec xkae’ clearly gave the best results. It sur-
passed Norway spruce (’P ab’) by 108%,tamarack (’lar’) by 44%, Douglas fir (’Pmenz’) by 39% and the Japanese larch
(’kae’) by 25%. It was significantly superiortoo over the other hybrids with values ran-ging from 48% over ’lar x kae’ down to14% over ’lar X (dec x kae)’. Results aremostly similar for HG4 but with superiorityof hybrid ’dec x kae’ over the other geneticentries even more important. As a compa-rison, in the minor test of Peyrat-Le-Château, the hybrid ’dec x kae’ reached413 cm for total height at the same age; it
surpassed by only 9% hybrid ’lar X (dec xkae)’ and by 72% tamarack (’lar’).
Total height growth development overtime from 1986 to 1989 is presented in fi-

gure 2. Except for the Japanese larch
(’kae’) and the hybrids ’kae x dec’ and ’larx kae’, superiority of the hybrid ’dec x kae’over the other species tended to increase.According to their growth curve pattern,the species might be associated in three
groups corresponding to fast, intermediateand slow growing material. Respectively,these would include ’dec x kae’; ’lar X (decx kae)’, ’kae x dec’, ’(dec x kae) X (dec xkae)’ and ’kae’; and finally ’lar’ and ’lar xkae’.
Highly significant differences (at α =
0.001) are noted too between species forstem straightness (SS) at 8 years as wellas between blocks (table IV). Two entriesincluding the tamarack (’lar’) and its F1 hy-brid with the Japanese larch (’lar x kae’)were characterized by severe crook (tableIII) and were significantly different (at α =
0.01) from the other entries.As well, for the frequency of basal
sweep (%BS) and the rate of straightstems (%SS), the material responded dif-ferently (2 I = 95.8 > Chi20.95, 10 df for %BSand 2I = 195.6 > Chi20.95, 10 df for %SS).The material could be associated in three
homogeneous groups. They included frombest to worst: Norway spruce (P ab) andDouglas fir (P menz); ’lar x kae’ and ’lar X
(dec x kae)’; ’lar’, ’kae’, ’kae x dec’ and ’decx kae’ for basal sweep, with values rangingfrom no defect for the species used ascontrol up to more than 42% for the hybrid’dec x kae’. For frequency of straight stems(%SS), group 1 included Norway spruce (Pab) and Douglas fir (P menz); group 2 ’kaex dec’, ’lar X (dec x kae)’ and ’dec x kae’;and group 3 ’kae’, ’(dec x kae) X (dec xkae)’, ’lar’ and ’lar x kae’. %SS ranged fromno defect for the control species up to
100% for the hybrid ’lar x kae’. brid (’decx kae’) was quite severely affected withnearly two-thirds of crooked stems (tableIII).
DISCUSSION AND CONCLUSION
Hybridization including tamarack is repor-ted here for the first time in France and thetest at Eclache is the first opportunity toobserve tamarack hybrids performance inthe field.
Hybridization between European and
Japanese larches does not raise any parti-cular problems besides those well knownto larch breeders, namely the low set offilled seed per cone which is observed inboth natural and artificial crosses (Kosins-
ki, 1987). Hybridization using tamarack
proved to be possible but difficult under Or-léans climatic conditions. The limited suc-cess noted in this study may mostly be dueto problems connected with flower deve-lopment (proliferated female strobili, matu-ration failure of male strobili with resultinglack of pollen production, non-matching
phenology with other species (Japaneselarch)).
Vegetative propagation was successfulfor most of the hybrids but clonal variationwas substantial. Such a high rate of root-ing with young plants is not unusual
(Mason, 1989). The low performance du-ring the propagation phase of two hybrids’kae x dec’ and ’(dec x kae) X (dec x kae)’for both rootability and their root systemquality cannot be attributed to vigour of thedonor plants; they were as vigorous asthose of other species. Possible explana-tions could be either the age of the donor
plants for hybrid ’kae x dec’ (one year olderthan other hybrid stocks) or the genetic na-

ture of the material or both. Hybrid ’kae xdec’ clones were in fact selected in a
single progeny from open pollinated hybri-dization in a seed orchard while hybrid’(dec x kae) X (dec x kae)’ clones were se-lected in a single F2 full-sib family, the pa-rents of which were full sibs. In bothcases, inbreeding may have taken placeand this could have serious depression ef-fects, as expected on larch (Dieckert,1964). No abnormality of growth (growthdepression, abnormal branchiness) wasnevertheless detected at the time of ortetselection.
Considering the traits related to vegeta-tive propagation, tamarack as well as its
hybrids perform better than the other hy-brids, ’dec x kae’ included. This is particu-larly true for the quality of the root system(table II).
In the field test, no serious adaptationproblems have been noted as survival upto 8 years is judged satisfactory, except fortamarack with nearly 30% mortality. Simi-lar results were observed for seedlings ofthe same two tamarack provenances usedin this study (and others from the same la-titude) in an arboretum located in the Mas-sif Central Mountains (Margeride, Lozère,elevation: 1470 m) (Imbert, 1988). Al-
though the latitudes of origin of the testedprovenances are similar to those of thetest sites, the local weather conditions aresevere (windy) and a better choice of pro-venances from higher latitudes and lessoceanic influenced zones might be recom-mended. In any case, larches showed amuch higher survival rate than Norwayspruce and Douglas fir. This is not surpri-sing for this Douglas fir provenance (Ar-lington) which may be at its ecological limithere. For Norway spruce which is traditio-nally used in this region, no obvious expla-nation can be given.
With regard to growth and vigour (tableIII), neither tamarack nor its hybrids ex-
ceeded performances of better known hy-brids (’dec x kae’ or its reciprocal) with theexception of hybrid ’lar X (dec x kae)’. Ta-marack was very poor but in any case, its
height growth was faster than that of Nor-way spruce. The three-way hybrid ’lar X
(dec x kae)’ was vigorous with perfor-mances intermediate (and significantly dif-ferent) to those of its parental species (’lar’and ’dec x kae’): -13,7% compared to ’decx kae’, + 26.7% with respect to ’lar’ for totalheight at 8 years. In contrast, the single hy-brid ’lar x kae’ performances were evenlower (but not significantly different at α =
0.01) than those of its poorer parent spe-cies (’lar’). MacGillivray (1967) observed amuch more favourable performance of thathybrid in south-central New Brunswickwhere it was 48.0% and 57.4% taller res-
pectively than L laricina and L kaempferi at7 years. But his observations were basedon only four seedlings.
The hybrid between L decidua andL kaempferi is known for its remarkable vi-gour (Pâques, 1989) which is confirmed bythe present study in rather severe ecologi-cal conditions. As a comparison, at two
other sites in somehow milder conditionsof the western range of the Massif Central
Mountains, total height at the same ageand for a similar genetic material exceededby 76 and 27% respectively total height re-corded on the site of Eclache.
The F2 hybrid ’(dec x kae) X (dec x kae)’had an intermediate growth. Its successseems highly dependent on the coancestrylevel of its F1 parent clones (Dietze, 1974).
If tamarack and its hybrids presentedfew basal sweep defects as compared toL decidua x L kaempferi, they were on av-erage affected by serious stem form prob-lems with a much higher frequency ofcrooked stems and more severe defects.
Hybrid ’lar X (dec x kae)’ was once againan exception as its stem form quality wasvery close to that of ’dec x kae’ (table III).

Stem form is nevertheless a generalproblem in larch breeding (Keiding and Ol-sen, 1965) and as shown in table III, no hy-brid or pure species is really free from thisdefect, the frequency of which goes up to50% even for the best tested material. Thiscondition is not restricted to vegetativelypropagated material (Dietze, 1974). More-over, straightness and vigour have been ob-served in many of our experiments to benegatively correlated and for certain hy-brids, correlations are significantly differentfrom zero (for α = 0.05) (table V). Fortu-nately, a rather broad variability exists forthis trait at the various levels (ramet (tableIII), clone, family and population). There-fore, further selection will certainly improvethis trait.
Tamarack is a poorly known species inFrance. These first results from experi-ments indicate that choice of provenancesand proper sites for reforestation underFrench conditions is important (Imbert,1988; Pâques, in preparation). Althoughvegetative propagation and adaptation donot seem to be serious problems in this ex-periment, field performance for the mostimportant traits (vigour and stem form) ofboth tamarack and its hybrids was neversuperior to better known hybrids betweenthe European and Japanese larches (evento the second generation hybrid ’(dec x
kae) X (dec x kae)’). One major exceptionis for the 3-way hybrid (L laricina X (L de-
cidua x L kaempferi)) which combines ex-cellent adaptability, fast growth (second toL decidua x L kaempferi) and good stemform (% BS, SS, %SS). Similar observa-tions both on the remaining trees in the mi-nor test site of Peyrat-Le-Château and inOrléans clone collection confirm its quality.
Due to the low number of entries (ram-ets, clones and families) for some hybrids,precise comparison of material was not al-ways possible. In particular, it is difficult todetect whether the observed performancedifferences are the result either of clone
sampling or of the species themselves.Testing on more sites and of other clonesfrom a wider range of hybrid familiesshould be undertaken to confirm these ear-
ly results and to give indications about theinterest of tamarack hybrids over L decid-ua x L kaempferi.
But in any case, the difficulty - con-firmed by later attempts - to obtain hybridsusing tamarack either as female or as
male under Orléans conditions is a majorlimiting factor whatever the interest ofsome of its hybrids.
Transfer of favourable traits from one
species to another through interspecific hy-bridization is of major interest in larch
breeding as the species commonly used,seem complementary for a number ofcharacteristics (eg canker resistance, stemform). In particular, it is important to checkif the tamarack tolerance to waterlogged

soils might be transferred to other larchspecies through hybridization. Further re-search work with tamarack will be orientedin this direction.
ACKNOWLEDGMENTS
The technical assistance of P Legroux andM Faucher for data collection and the help ofCH Schneider for statistical analysis were great-ly appreciated.
REFERENCES
Arbonnier P (1966) L’analyse de l’information.Aperçu théorique et application à la loi multi-nomiale. Ann Sci For 23, 950-1017
Avrov FD (1982) Crossability of various larchspecies at a collection-parent plantation insouthern Siberia. Lesovedenie 5, 23-29 (inRussian)
Bobrov EG (1973) Introgressive Hybridisation.Sippenbildung und Vegetationsänderung.Fed Rep 84, 273-294
Chowdbury KA (1931) Anatomical studies of thewood of Larix eurolepis. J For 29, 797
Delevoy G (1949) Larix eurolepis (Henry A,Flood M, eds). Bull Soc Centr For Belg 1949,178-194
Dieckert H (1964) Einige Untersuchungen zurSelbsterilität Und Inzucht bei Fichte undLärche. Silvae Genet 13, 77-86
Dietze W (1974) Beurteilung von züchterischenMöglichkeiten zur Verbesserung quantitati-ver und qualitativer Eigenschaften bei euro-paischer Lärche (Larix decidua Mill). Disser-tation, Forstwissensschftl. Fakultät, UnivMunich
Henry A, Flood MG (1919) The history of theDunkeld larch. Proc R Ir Acad Sec B 35
Keiding H, Olsen HC (1965) Assessment ofstem form in clones and progenies of larch.Silvae Genet 14, 115-122
Kosinski G (1987) Empty seed production in Eu-ropean larch (Larix decidua). For Ecol Manag19, 241-246
Imbert P (1988) Choix des Espèces de Reboise-ment en Haute-Margeride. Premier bilan del’Arboretum Curie (Lozère-Col des 3 Sœurs,Alt 1470 m). INRA-Orléans, document n° 397
Legendre L, Legendre P (1984) Écologie Numé-rique. 2. La Structure des Données Écologi-ques. Masson, Québec, 182-183, 2nd edn
Libby WJ, Cockerham CC (1980) Random non-contiguous plots in interlocking field layouts.Silvae Genet 29, 183-190
MacGillivray HG (1967) Hybrid between tama-rack and Japanese larch appears promisingin south central New Brunswick. Res Notes
Dep For Can 23, 2-3Mason WL (1989) Vegetative propagation of hy-
brid larch (Larix x eurolepis Henry) using win-ter cuttings. Forestry 62 (suppl), 189-198
Pâques LE (1989) A critical review of larch hy-bridization and its incidence on breeding stra-tegies. Ann Sci For 46, 141-153
Pâques LE, Cornu D (1991) Effect of vegetativepropagation on field performance up to age 8of hybrid larch (Larix x eurolepis) clones. AnnSci For 48, 469-482
Sokal RR, Rohlf FJ (1981) Biometry - The Prin-ciples and Practice of Statistics in BiologicalResearch. Freeman, San Francisco, 859,2nd edn
Syrach Larsen C (1937) The Employment ofSpecies, Types and Individuals in Forestry.Copenhagen,1937
Wright JW (1976) Introduction to Forest Gene-tics. Academic Press, New York, 322-323