Embed Size (px)
Citation preview

, . 182: 99–105 (1997)
PROPERTIES OF MULTINUCLEATED GIANT CELLS INA NEW IN VITRO MODEL FOR HUMAN GRANULOMA
FORMATION
1, -**, - ***, 1, 1,̈ 2, - 1 1*
1Department of Immunology and Cell Biology, Forschungszentrum Borstel, Germany2Division of Clinical and Experimental Pathology, Forschungszentrum Borstel, Germany
SUMMARY
Multinucleated giant cells (MGCs) are a key feature of granulomas. They have been studied with respect to the mechanism andregulation of their formation, but the function of these cells still remains elusive. A new method for the in vitro generation of granulomaswas developed and characterized in which L3 larvae of Nippostrongylus brasiliensis, as a target for the cellular response, wereco-incubated with human mononuclear blood cells. The development of epithelioid cells and MGCs was observed and single isolatedMGCs were analysed by the reverse transcriptase polymerase chain reaction method. The presence of tumour necrosis factor alpha(TNFá), interleukin-1 beta (IL-1â), interleukin-6 (IL-6), and inducible nitric oxide synthase (iNOS) transcripts in MGCs wasdemonstrated. It is proposed that MGCs in the granuloma model may in part represent an active cellular constituent involved ingranuloma formation and turnover and in the destruction of the irritant. ? 1997 by John Wiley & Sons, Ltd.
J. Pathol. 182: 99–105, 1997.No. of Figures: 3. No. of Tables: 2. No. of References: 30.
KEY WORDS—multinucleated giant cell; in vitro granuloma; single cell reverse transcriptase polymerase chain reaction (RT-PCR);tumour necrosis factor alpha (TNFá); interleukin-1 beta (IL-1â); interleukin-6 (IL-6); inducible nitric oxide synthase(iNOS)
INTRODUCTION
The granulomatous response is a specialized form ofinflammatory reaction found in many infective, toxic,allergic, autoimmune, and neoplastic diseases.1 Histo-logically, the granulomatous response consists of focallesions containing macrophages, epithelioid cells, multi-nucleated giant cells, and other cell types includingT-cells, granulocytes, and fibroblasts.1 Since the precisemechanism involved in recruitment, activation, andmaintenance of immune cells at the granulomatous fociis not clearly understood, an in vitro model mimickinghuman granuloma formation would be a useful tool toaddress these questions.A large variety of in vivo granuloma models have been
developed in the murine system, including the schisto-some egg model;2 in vitro models include the use ofantigen-coated beads,3 cytokine-coated beads,4 andstimulated monocyte cultures.5–9 However, none ofthese models exactly imitates human granuloma forma-tion with respect to the cellular components involved.
The macrophage and its derivatives, epithelioid andgiant cells, are considered a histological hallmark ofgranulomas. It is believed that these cells are an import-ant source of molecular mediators that dictate theprogression of granuloma formation.4 Studies havedemonstrated the presence of IL-1 and TNFá withingranuloma extracts, or their production by granulomamacrophages,10–12 but little is known about the functionof epithelioid and giant cells. Multinucleated giant cells(MGC) represent an end-point in myeloid cell lineagedevelopment13,14 and are formed by macrophagefusion,13 which is inducible by some cytokines.6–9 Theyhave been credited with an enhanced candidacidalactivity,15 elevated enzyme equipment, and an increasein oxygen free radicals in response to zymosan.13In this study, we introduce a new in vitro model for
human granuloma formation which allows the prep-aration and analysis of MGCs at the mRNA level,using a reverse transcriptase polymerase chain reaction(RT-PCR) method developed for single cells.16
MATERIALS AND METHODS
Preparation of Nippostrongylus brasiliensis larvaeWistar rats were infected by subcutaneous injection
with third-stage larvae of Nippostrongylus brasiliensis(the larvae were a kind gift of Professor K. Ishizaka, LaJolla, CA, U.S.A.). Third-stage larvae were preparedfrom the faeces of infected rats as previously described.2For long-term storage, the larvae were frozen in 100 per
*Correspondence to: Professor Johannes Gerdes, Division ofMolecular Immunology, Forschungszentrum Borstel, Parkallee 22,23845 Borstel, Germany.
**Current address: Department of Rheumatology, University ofBirmingham, U.K.
***Current address: Department of Immunology, University ofBirmingham, U.K.
Contract grant sponsor: Deutsche Forschungsgemeinschaft;Contract grant number: SFB 367/C1.
CCC 0022–3417/97/050099–07 $17.50 Received 3 May 1996Revised 24 September 1996? 1997 by John Wiley & Sons, Ltd.Accepted 17 October 1996

cent FCS (fetal calf serum) and preserved in liquidnitrogen. The survival rate was 70 per cent over a periodof 14–20 days in culture.
Preparation of human peripheral blood mononuclearcells (MNCs)
MNCs were obtained from heparinized blood fromhealthy donors by centrifugation through Ficoll-Hypaque (Pharmacia) gradients followed by washing.
Cell culture
Iscove’s modified Dulbecco’s medium (IMDM)(Gibco) was supplemented to a final concentration of100 U/ml penicillin G sodium salt, 100 ìg/ml streptomy-cin (Gibco), and 5 per cent heat-inactivated autologoushuman serum.Co-culture of 0·75#106/ml MNCs and 20 worms/well
was performed in a volume of 2 ml in 24-well tissueculture plates. Co-incubations of MNCs and cottonfibres as structural analogues to the nematodes andMNCs alone were cultured in parallel, for a comparativeanalysis of granuloma formation. Experiments wereperformed in triplicate with four determinations each.
Immunoenzymatic and immunochemical staining
Cytospin slides were prepared from worms withadherent cell clusters aspirated from the primary cul-tures under microscopic control. Specimens were im-munostained by the alkaline phosphatase anti-alkalinephosphatase (APAAP) technique,17 using New Fuchsindevelopment.The antibodies used in this study were as follows:
anti-CD14 (HB44, American Type Culture Collection),anti-CD4 (Leu 3a, Becton Dickinson), anti-gp 140 kd(Ber-Mac318), anti-CD8 (Leu 2a, Becton Dickinson),anti-CD19 (MD-37, Dako), anti-IL1 (Fib-319), anti-IL-6(B-E8, Biosource International), and anti-TNFá(TNFE20). Histochemical staining for NADPH-diaphorase was performed according to Vincent andKimura.21
Determination of the fusion index
The number of nuclei within MGCs (>2 nuclei/cell) inat least 1000 nuclei examined was calculated to give thefusion index (%)=(number of nuclei within MGCs)/(total number of nuclei counted)#100.5,9
Electron microscopyIn vitro granulomas embedded in 2 per cent agarose
were fixed in 2 per cent glutaraldehyde (1 h) and 2 percent osmium tetroxide (1 h). Samples were contrasted in2 per cent uranyl acetate (1 h), dehydrated, and embed-ded in Epon. Ultrathin sections were post-contrastedwith lead citrate and examined (EM 300, Philips).
Micropreparation of single MGCsIn vitro granulomas were stained with Hoechst 33342,
propidium iodide (PI), Ber-Mac3 and CY3-conjugated
goat anti-mouse IgG (Dianova), and FITC-conjugatedmouse anti-CD29 (IOT29, Dianova). MGCs wereselected by means of the micromanipulator 5171 andtransjector 5246 (Eppendorf) and transferred singly intothe well of a Terasaki plate for immunofluorescenceanalysis. Selection criteria for further analysis were theverification that only one cell was present, the presenceof more than two nuclei, and negativity for PI andBer-Mac3. Staining with fluorescein-5-isothiocyanatediacetate (Molecular Probes) as a metabolic marker22was performed in a parallel incubation.
Single cell RT-PCR
Single cell RT-PCR was performed as previouslydescribed.16 The primers used in this study wereGAPDH (glycerin-aldehyde-3-phosphate-dehydrogenase)sense: 5*AACAGCGACACCCACTCCTC, antisense:5*GGAGGGGAGATTCAGTGTGGT; TNFá sense:5*AACGGAGGCTGAACAATAGGC, antisense:5*AGCAACCTTTATTTCTCGCCAC; IL-1â sense:5*CTCTCACCTCTCCTACTCACTTAAAGCCCG,antisense: 5*CACACCAGTCCAAATTGAATTGATTCCATAGA; IL-6 sense: 5*GCACCTCAGATTGTTGTTGTTA, antisense: 5*GCCACTTTCCATTATTATTTCA; iNOS sense: 5*AAGTAGCACCCTGGATTGA,antisense: 5*AAGGAATCATACAGGGAAGAC.
RESULTS
Characterization of the in vitro granuloma modelFive blood donors were tested for the in vitro granu-
loma formation capacity of their MNCs. Since therewere no evident differences in morphological or pheno-typical terms (see below), the MNCs of three of thesedonors were used for co-culture in all subsequent exper-iments. Typically, after 1 day of co-incubation, the cellclusters adhering to the nematode consisted predomi-nantly of small cells with dense round nuclei and sparsecytoplasm. The immunophenotype of the cluster-forming cells of one representative experiment issummarized in Table I. At day 1 of in vitro granulomaformation, many of the cells expressed antigens charac-teristic for monocytes/macrophages (CD14, Ber-Mac3).CD4-positive cells and CD8-positive cells, as well asCD19-positive B-lymphocytes, were also present at day1 (Table I). Cell clusters did not form around deadlarvae, which were found to a certain extent in eachculture. After 4 days, the cell clusters were greatlyincreased in size, with no obvious changes in cell mor-phology or immunophenotype. Between the fourth andthe seventh day of co-incubation, remarkable trans-formations could be observed at the cellular level: mostcells had abundant cytoplasm and a small eccentricnucleus (Fig. 1a). This suggested that epithelioid cellformation had taken place in the culture.23,24 Theassumption of epithelioid cell formation was verified bytransmission electron microscopy, which demonstratedthe appropriate subcellular constituents and the pres-ence of characteristic cell–cell interdigitations (Fig. 1b).Multinucleated cells (Fig. 1a) were first observed at days4–7.
100 V. SEITZER ET AL.
? 1997 by John Wiley & Sons, Ltd. J. Pathol. 182: 99–105 (1997)
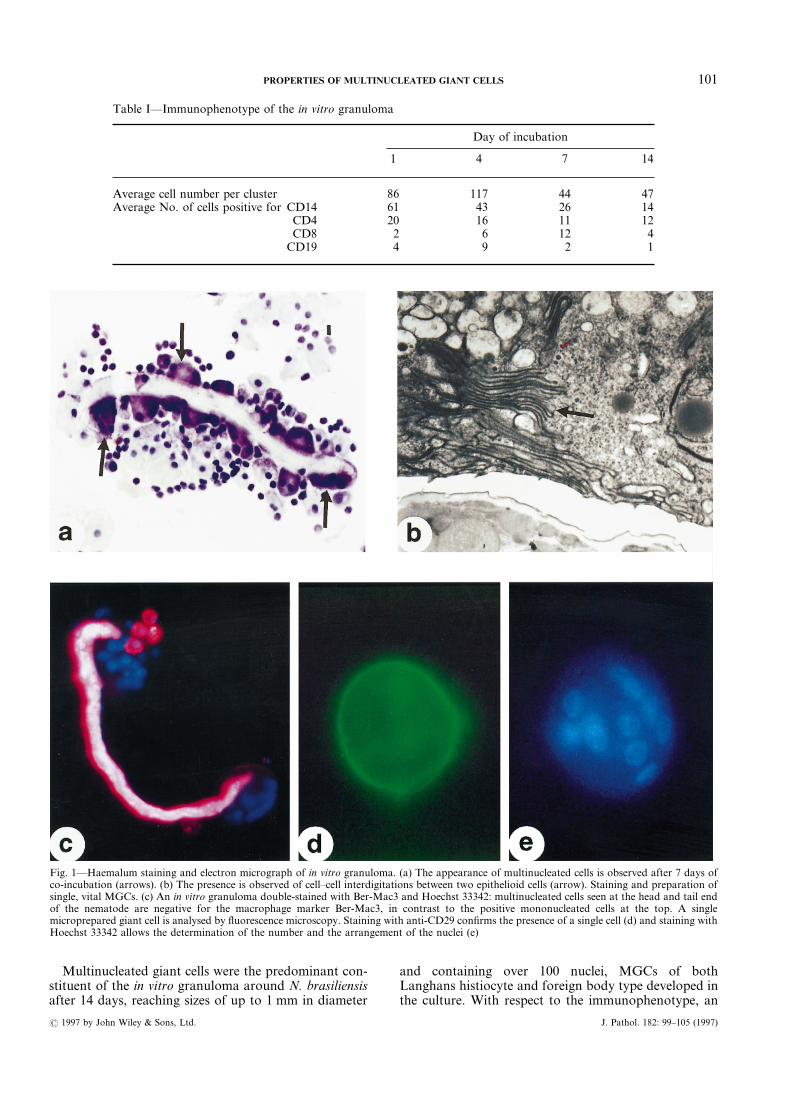
Multinucleated giant cells were the predominant con-stituent of the in vitro granuloma around N. brasiliensisafter 14 days, reaching sizes of up to 1 mm in diameter
and containing over 100 nuclei, MGCs of bothLanghans histiocyte and foreign body type developed inthe culture. With respect to the immunophenotype, an
Table I—Immunophenotype of the in vitro granuloma
Day of incubation
1 4 7 14
Average cell number per cluster 86 117 44 47Average No. of cells positive for CD14 61 43 26 14
CD4 20 16 11 12CD8 2 6 12 4CD19 4 9 2 1
Fig. 1—Haemalum staining and electron micrograph of in vitro granuloma. (a) The appearance of multinucleated cells is observed after 7 days ofco-incubation (arrows). (b) The presence is observed of cell–cell interdigitations between two epithelioid cells (arrow). Staining and preparation ofsingle, vital MGCs. (c) An in vitro granuloma double-stained with Ber-Mac3 and Hoechst 33342: multinucleated cells seen at the head and tail endof the nematode are negative for the macrophage marker Ber-Mac3, in contrast to the positive mononucleated cells at the top. A singlemicroprepared giant cell is analysed by fluorescence microscopy. Staining with anti-CD29 confirms the presence of a single cell (d) and staining withHoechst 33342 allows the determination of the number and the arrangement of the nuclei (e)
101PROPERTIES OF MULTINUCLEATED GIANT CELLS
? 1997 by John Wiley & Sons, Ltd. J. Pathol. 182: 99–105 (1997)
interesting finding was that macrophages were positivefor Ber-Mac3, whereas epithelioid cells showed weakerexpression and MGCs entirely lacked this marker (Fig.1c). This in vitro finding parallels the results obtainedin situ,18 where Ber-Mac3 reacted strongly with macro-phages in normal and diseased tissue, but not withepithelioid type macrophages or multinucleated giantcells. The negativity of MGCs for this macrophagemarker thus makes Ber-Mac3 a valuable exclusionmarker for these cells. It is noteworthy that even after 2weeks in culture, B-cells, CD4-positive cells, and CD8-positive cells were present to an extent comparable tothe first day of co-incubation (Table I).The cell viability of the in vitro granulomas selected at
the various time points for phenotypical analysis wasassessed by fluorescein-5-isothiocyanate diacetate stain-ing directly in the culture well. The specimens routinelyshowed 95–100 per cent viability of the cells attached inclusters to the nematodes, independent of the incubationperiod.Comparative analysis of MGC formation as a
measure of the stimulus of the culture conditions forgranuloma formation in cultures of mononuclear cells(MNCs) only and of MNCs and cotton fibres, showed aless pronounced attachment of cells to the fibres and lessdifferentiation of the cells in the course of co-incubation.A fusion index of 50&11 per cent (n=3) was observed in
the N. brasiliensis and MNC co-culture after 14 days ofco-incubation. This was significantly higher than the18&6 per cent (n=3) fusion index observed in the MNCplus cotton fibre co-incubation and the 8&4 per cent(n=3) fusion index in the MNC culture alone.
Preparation and transcript analysis of single MGCs
The staining and preparation of single MGCs isdepicted in Figs 1c–1e. Phase contrast microscopy andDNA staining with Hoechst 33342 showed the presenceof single and multinucleated cells attached to the nema-tode. Double staining with CD29 as a membrane markerof the same specimen confirmed that the multinucleatedcells were surrounded by a single membrane and consti-tuted single cells. Staining for DNA and Ber-Mac3showed the presence of Ber-Mac3-positive cells with asingle nucleus and of multinucleated cells negative forBer-Mac3 (Fig. 1c). From specimen suspensions stainedin this manner, only vital (PI negative), multinucleated(>3 nuclei, Hoechst 33342), Ber-Mac3-negative, andsingle (CD29, cell membrane positive) cells selected bymicropreparation were subjected to further analysis byRT-PCR (Figs 1d and 1e).An example of the RT-PCR analysis for a series of
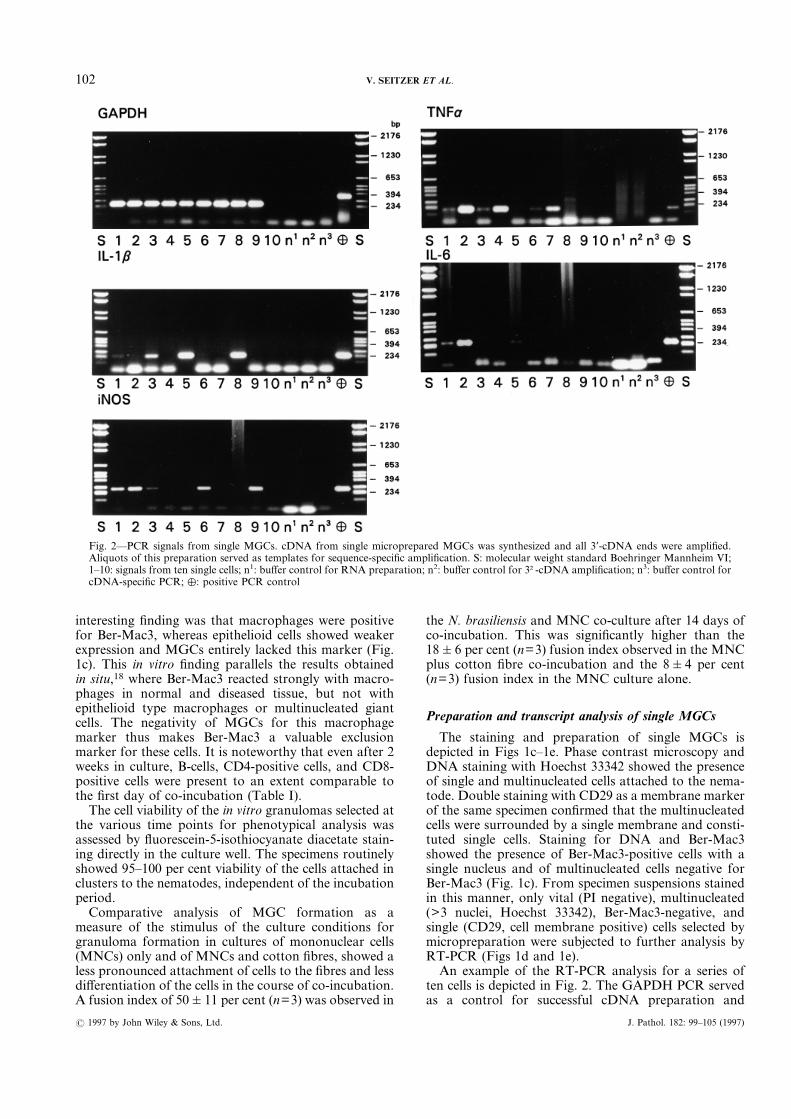
ten cells is depicted in Fig. 2. The GAPDH PCR servedas a control for successful cDNA preparation and
Fig. 2—PCR signals from single MGCs. cDNA from single microprepared MGCs was synthesized and all 3*-cDNA ends were amplified.Aliquots of this preparation served as templates for sequence-specific amplification. S: molecular weight standard Boehringer Mannheim VI;1–10: signals from ten single cells; n1: buffer control for RNA preparation; n2: buffer control for 3†-cDNA amplification; n3: buffer control forcDNA-specific PCR; %: positive PCR control
102 V. SEITZER ET AL.
? 1997 by John Wiley & Sons, Ltd. J. Pathol. 182: 99–105 (1997)

amplification. Only GAPDH-positive cells were in-cluded in the analysis, the summary of which is shownin Table II. Of 55 GAPDH-positive cells prepared fromseven experiments, 19 expressed TNFá transcripts andto a lesser extent IL-1â transcripts (5 cells), IL-6 tran-scripts (8 cells), and iNOS transcripts (10 cells). Whenconsidered separately, FBMGCs displayed a similarratio of the investigated transcripts, whereas LHMGCsdid not express IL-6 transcripts. There was no corre-lation between the number of nuclei and the transcriptspresent.NADPH-diaphorase staining of MGCs from a 14-day
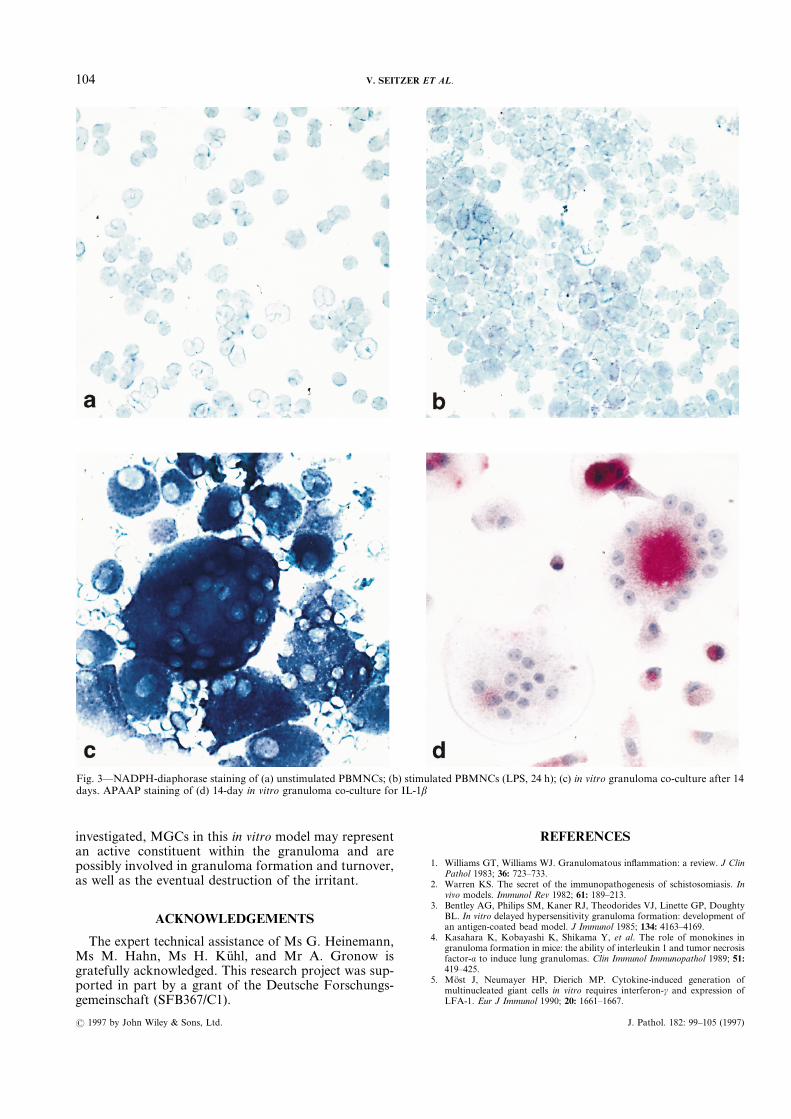
co-culture revealed strong staining of all MGCs (Fig.3c); MGCs attached directly to the nematode showedthe most intense staining. Control unstimulated andLPS-stimulated PBMNCs showed a less pronouncedstaining intensity of stimulated cells only (Figs 3a and3b). Immunocytochemical staining of MGCs for thecytokines of which transcripts were found was per-formed to investigate whether the protein product couldbe detected in the cells. A representative result for IL-1âis shown Fig. 3d. In all stainings performed, MGCspositive and negative for IL-1â, TNFá, and IL-6 couldbe discerned.
DISCUSSION
Characterization of the in vitro granuloma in thismodel showed that its cellular constituents and cellulardevelopment closely resembled human granuloma for-mation in vivo.25,26 Histologically, the in vitro granulomaappeared as an aggregation of macrophages, epithelioidcells, MGCs, and T- and B-cells clustered in a three-dimensional structure around the nematode. These fea-tures, and the use of human MNCs, give this systeman advantage over previously described models, whichpredominantly focused on parameters which influencedthe size of the cell clusters, in which differentiation ofmacrophages to epithelioid cells and MGCs could not beobserved.3,27,28 and the majority of which were notbased on the human system. A variety of studies haveemployed cytokine-induced monocyte fusion, contribut-ing greatly to our understanding of the factors involvedin MGC formation.5–7,9,15 The present model may bereadily used to study factors which affect and modu-late the formation of individual in vitro human granu-lomas, making them readily amenable to molecularanalysis.
To our knowledge, this study is the first showing thepresence of TNFá, IL-1â, IL-6, and iNOS transcriptsand protein in MGCs, implying an active role in thein vitro model. TNFá and IL-1â are both consideredsignificant modulators in granuloma formation,11,12 andiNOS is the producer of nitric oxide, which is regardedas one of the major cytotoxic agents in the eliminationof intracellular parasites.29 Additionally, it has beenclaimed that TNFá has an autoamplifying role in MGCformation8 and IL-6 has an enhancing effect.9 Weare not aware of a significant function for IL-6 in agranulomatous response, but it is noticeable that ourresults do not show the presence of IL-6 transcripts inLHMGCs. Since the morphologically distinct LHMGCsand FBMGCs may originate from different fusionmechanisms,6 it would be reasonable to assume differingfunctions, for which our findings provide the first evi-dence. The results of the transcript analysis were furthersubstantiated by the immunohistochemical detection ofMGCs positive and negative for IL-1â, IL-6, and TNFá.Thus, it appears that these cells synthesize not only theanalysed transcripts, but also the final protein products.The demonstration of intense NADPH-diaphorasestaining of these MGCs may be interpreted as anindication for NO production and thus the presence ofNO synthase in these cells.30We are aware that the detection of the transcripts may
be attributable to the cellular contribution of freshlyfused activated macrophages within an MGC, butseveral points argue against this assumption. Firstly,fluorescein-5-isothiocyanate diacetate staining for meta-bolic activity of MGCs was consistently distributedevenly within the cell, suggesting a uniform cytoplasmicdistribution of cellular activity. Secondly, the meticulouspreparation of single cells, with exclusion criteria ofBer-Mac3 negativity and the staining of the cell mem-brane, eliminated, in our opinion, the possibility of theselection of an MGC freshly fused with active macro-phages. Thirdly, the immunocytochemical results allowthe discrimination of positively and negatively stainedsingle MGCs, underlining the results of the transcriptanalysis.The functional role of MGCs in general has remained
elusive, but there are indications that a change infunctional properties occurs from the macrophage to theMGC7 and that MGCs have an enhanced oxidativecapacity compared with macrophages.13,15 In summary,our results indicate that although the transcriptsanalysed were not expressed uniformly, nor in all MGCs
Table II—Single cell RT-PCR analysis of MGCs
GAPDH TNFá IL-1â IL-6 iNOS
No. cells No. cells % No. cells % No. cells % No. cells %
All MGCs 55 19 35 5 9 8 15 10 18FBMGCs 42 14 33 3 7 8 21 8 21LHMGCs 13 5 38 2 15 0 0 2 15
FBMGCs: MGCs of foreign body type; LHMGCs: MGCs of Langhan’s histiocyte.
103PROPERTIES OF MULTINUCLEATED GIANT CELLS
? 1997 by John Wiley & Sons, Ltd. J. Pathol. 182: 99–105 (1997)
investigated, MGCs in this in vitro model may representan active constituent within the granuloma and arepossibly involved in granuloma formation and turnover,as well as the eventual destruction of the irritant.
ACKNOWLEDGEMENTS
The expert technical assistance of Ms G. Heinemann,Ms M. Hahn, Ms H. Kühl, and Mr A. Gronow isgratefully acknowledged. This research project was sup-ported in part by a grant of the Deutsche Forschungs-gemeinschaft (SFB367/C1).
REFERENCES
1. Williams GT, Williams WJ. Granulomatous inflammation: a review. J ClinPathol 1983; 36: 723–733.
2. Warren KS. The secret of the immunopathogenesis of schistosomiasis. Invivo models. Immunol Rev 1982; 61: 189–213.
3. Bentley AG, Philips SM, Kaner RJ, Theodorides VJ, Linette GP, DoughtyBL. In vitro delayed hypersensitivity granuloma formation: development ofan antigen-coated bead model. J Immunol 1985; 134: 4163–4169.
4. Kasahara K, Kobayashi K, Shikama Y, et al. The role of monokines ingranuloma formation in mice: the ability of interleukin 1 and tumor necrosisfactor-á to induce lung granulomas. Clin Immunol Immunopathol 1989; 51:419–425.
5. Möst J, Neumayer HP, Dierich MP. Cytokine-induced generation ofmultinucleated giant cells in vitro requires interferon-ã and expression ofLFA-1. Eur J Immunol 1990; 20: 1661–1667.
Fig. 3—NADPH-diaphorase staining of (a) unstimulated PBMNCs; (b) stimulated PBMNCs (LPS, 24 h); (c) in vitro granuloma co-culture after 14days. APAAP staining of (d) 14-day in vitro granuloma co-culture for IL-1â
104 V. SEITZER ET AL.
? 1997 by John Wiley & Sons, Ltd. J. Pathol. 182: 99–105 (1997)

6. McNally AK, Anderson JM. Interleukin 4 induces foreign body giant cellsfrom human monocytes/macrophages. Differential lymphokine regulationof monophage fusion leads to morphological variants of multinucleatedgiant cell. Am J Pathol 1995; 147: 1487–1499.
7. Fais S, Burgio VL, Silvestri M, Capobianchi MR, Pacchiarotti A, Pallone F.Multinucleated giant cell generation induced by interferon ã. Changes in theexpression and distribution of the intercellular adhesion molecule-1 duringmacrophage fusion and multinucleated giant cell formation. Lab Invest1994; 71: 737–744.
8. Takashima T, Ohnishi K, Tsuyuguchi I, Kishimoto S. Differential regu-lation of formation of multinucleated giant cells from concanavalinA-stimulated human blood monocytes by IFN-ã and IL-4. J Immunol 1993;150: 3002–3010.
9. Enelow RI, Sullivan GW, Carper HT, Mandell GL. Induction of multi-nucleated giant cell formation from in vitro culture of human monocyteswith interleukin-3 and interferon-ã: comparison with other stimulatingfactors. Am J Respir Cell Mol Biol 1992; 6: 57–62.
10. Kobayashi K, Allred C, Cohen S, Yoshida T. Role of interleukin 1 inexperimental pulmonary granuloma in mice. J Immunol 1985; 134: 358–364.
11. Kindler V, Sappino A-P, Grau GE, Piguet P-F, Vassalli P. The inducingrole of tumor necrosis factor in the development of bactericidal granulomasduring BCG infection. Cell 1989; 56: 731–740.
12. Chensue SW, Otterness IG, Higashi GI, Shmyr Forsch C, Kunkel SL.Monokine production by hypersensitivity (Schistosoma mansoni egg) andforeign body (Sephadex bead)-type granuloma macrophages. Evidence forsequential production of IL-1 and tumour necrosis factor. J Immunol 1989;142: 1281–1286.
13. Kreipe H, Radzun HJ, Rudolph P, et al. Multinucleated giant cellsgenerated in vitro. Terminally differentiated macrophages with down-regulated c-fms expression. Am J Pathol 1988; 130: 232–243.
14. Mechtersheimer G, Möller P. Phenotyping of human epithelioid cells,alveolar macrophages, and foamy cells by means of mAb of the myeloidsection. In: Knapp W, Dörken B, Gilks WR, et al. eds. Leucocyte TypingIV. White Cell Differentiation Antigens. Oxford: Oxford University Press,1989; 918–921.
15. Enelow RI, Sullivan GW, Carper HT, Mandell GL. Cytokine-inducedhuman multinucleated giant cells have enhanced candidacidal activity andoxidative capacity compared with macrophages. J Infect Dis 1992; 166:664–668.
16. Toellner K-M, Scheel-Toellner D, Seitzer U, et al. The use of reversetranscription polymerase chain reaction to analyse large numbers ofmRNA-species from a single cell. J Immunol Methods 1996; 191: 71–75.
17. Cordell JL, Fallini B, Erber WN, et al. Immunoenzymatic labelling ofmonoclonal antibodies using immune complexes of alkaline phosphataseand monoclonal anti-alkaline phosphatase antibodies (APAAP complexes).J Histochem Cytochem 1984; 32: 219–229.
18. Backé E, Schwarting R, Gerdes J, Ernst M, Stein H. Ber-Mac3: newmonoclonal antibody that defines human monocyte/macrophage differen-tiation antigen. J Clin Pathol 1991; 44: 936–945.
19. Herzbeck H, Blum B, Rönspeck W, Brandt E, Ulmer AJ, Flad H-D.Functional and molecular characterization of a monoclonal antibodyagainst the 165–186 peptide of human IL-1â. Scand J Immunol 1989; 30:549–562.
20. Bringman TS, Aggarwal BB. Monoclonal antibodies to human tumornecrosis factors á and beta: application for affinity purification, immuno-assays, and as structural probes. Hybridoma 1987; 6: 489–507.
21. Vincent SR, Kimura H. Histochemical mapping of nitric oxide synthase inthe rat brain. Neuroscience 1992; 46: 755–784.
22. Bronner-Fraser M. Alterations in neural crest migration by a monoclonalantibody that affects cell adhesion. J Cell Biol 1985; 101: 610–617.
23. Adams DO. The structure of mononuclear phagocytes differentiatingin vivo. Am J Pathol 1975; 80: 101–116.
24. Papadimitriou JM, Spector WG. The origin, properties and fate of epithe-lioid cells. J Pathol 1971; 105: 187–203.
25. Ramaswamy K, De Sanctis GT, Green F, Befus D. Pathology of pulmonaryparasitic migration: morphological and bronchoalveolar cellular responsesfollowing Nippostrongylus brasiliensis infection in rats. J Parasitol 1991; 77:302–312.
26. Warren KS. The cell biology of granulomas (aggregates of inflammatorycells) with a note on giant cells. In: The Cell Biology of Inflammation,Amsterdam: Elsevier/North Holland Biomedical Press, 1980; 543–557.
27. Franklin GF, Goghill G, McIntosh L, Cree IA. Monocyte aggregationaround agarose beads in collagen gels: a 3-dimensional model of earlygranuloma formation? J Immunol Methods 1995; 186: 285–291.
28. Shikama Y, Kobayashi K, Kasahara K, et al. Granuloma formation byartificial microparticles in vitro. Am J Pathol 1989; 134: 1189–1199.
29. Hibbs J Jr, Taintor RR, Vavrin Z. Macrophage cytotoxicity: role forL-arginine deiminase and imino nitrogen oxidation to nitrite. Science 1987;235: 473–476.
30. Beesley JE. Histochemical methods for detecting nitric oxide synthase.Histochem J 1995; 27: 757–769.
105PROPERTIES OF MULTINUCLEATED GIANT CELLS
? 1997 by John Wiley & Sons, Ltd. J. Pathol. 182: 99–105 (1997)





























