Embed Size (px)
Citation preview

Eur. J. Biochem. 63,553-562 (1976)
Proprietks spectrales du plasminoghe porcin gtude de la transition acide
Francis RODIER
Laboratoire de Biologie Physico-Chimique, Universitt de Paris-Sud, Centre d’Orsay
( R e p le 2 mai/30 juillet 1975)
Spectral Properties of Porcine Plusminogeiz : Study of the Acidie Trunsition
The acidic transition of porcine plasminogen, prepared by affinity chromatography, was studied by non-destructive methods. These methods are based on the analysis of the behaviour of the tryptophyls under various conditions. The perturbation of the absorption and emission spectra by pH or temperature and the dynamic quenching of the intrinsic fluorescence are used to obtain information on structural changes which affect the environment of these residues. It is shown that by decreasing pH the fluorescence emission spectra are shifted toward the long wavelengths, with a broadening of the fluorescence band. The same effect can be obtained at constant pH by heating the protein solution. In order to analyze these phenomena, it is assumed that the fluorescence intensities at 355 nm and 328 nm reflect the proportion of the tryptophans which are exposed to the solvent, and buried, respectively. The plot of the ratio of the fluorescence intensities at these wavelengths versus pH or temperature leads to a titration curve showing an unmasking of tryptophans. The proportion of exposed tryptophans is measured by the dynamic fluorescence quenching technique and the data analyzed according to Lehrer. The plot of the fraction of exposed tryptophyls versus pH also shows the unmasking of these chromophores. Thermal perturbation of a solution of plaminogen at neutral pH induces a difference absorption spectrum whose amplitudes at the maxima are proportional to the number of exposed aromatic residues. The comparison with a solution of fully denatured plasminogen in 6 M guanidium chloride, where all the tryptophyls are exposed, shows that the percentage of exposure is equal to 59 %. This number is significantly higher than the percentage found by the fluorescence quenching technique (20 %), indicating that some tryptophyls are located in crevices, exposed to the solvent but not to the iodide. At acidic pH the absorption difference spectra induced by thermal perturbation are not classical, since they show an inversion and a new band between 300 nm and 305 nm. This band is mentioned in the literature as a minor band of tryptophan which appears when this chromophore is located in an asymmetric environment. On plotting the maximum amplitude of these spectra obtained at acidic pH versus temperature, we obtain a curve indicating that two types of antagonistic interactions are involved in the perturbation of the chromophores spectra. The spectrophotometric titration of plasminogen gives classical absorption difference spectra. By plotting the maximum amplitude at 292 nm versus pH, we obtain a titration curve with an apparent pK of 2.9 units. This pK is acidic which respect to the pK value of a normal carboxyl. This low value can be due to a positively charged group in the neigh- bourhood of a carboxyl, which interacts with one or more chromophores. When the carboxyl becomes protonated, this positively charged group is free and available to perturb the environment of some chromophores. When the pH increases, the carboxyl becomes deprotonated and a salt bridge can be formed with the perturbing group. Thus this latter group no longer interacts with one or more chromophores and the absorption difference spectra remain unperturbed. The protonation of the carboxyl breaks the salt bridge and induces a partial unfolding of plasminogen which unmasks some tryptophyls. This unfolding enhances the thermal sensitivity of plasminogen at acidic pH.
Les methodes de spectroscopie d’absorption et ques non destructives pour l’etude des changements d’emission, bakes sur les modifications que subit la de structure des protitines En effet ces macro- lumiere lors de son passage dans la matiere, sont molecules possedent des acides aminks aromatiques utilisees depuis de nombreuses annees comme techni- dont les proprittts spectroscopiques sont intime-

554 Transition acide du plasminoghe porcin
ment likes, non seulement la configuration de ces chromophores, mais aussi a leur environnement. Parmi ces chromophores, le tryptophane semble ttre le residu dont les caracteristiques spectrales sont le plus perturbees par les interactions de l’etat fonda- mental et (ou) de l’ktat excite avec le solvant ou les groupes voisins [1,2]. On sait que l’etude des spectres d’absorption [3,4] et des spectres d’emission des tryptophyles d’une proteine, ainsi que la variation de leur rendement quantique sous l’influence de diffe- rents facteurs peut donner des informations sur leur degre d’exposition au solvant [5] et sur des change- ments globaux ou localises dans la structure proteique par reference A des modeles simples [6].
Au cours de notre etude de la structure du plas- minogene porcin, nous avons observe une transition acide partiellement irreversible qui apparut lors de la modification d’une etape de la preparation de cette proteine. 11 nous a paru interessant d’ktudier ce phenomene plus en detail et, si possible, d’essayer d’en expliquer le mecanisme, en se referant a des protkines connues.
MATERIEL ET METHODES
Prkpurution du plusminog2ne
Le plasminogene a t te prepare a partir de sang de pore defibrink par du chlorure de calcium a 4g / 100 ml par la methode de chromatographie d’affinite de Deutsch et Mertz [7]. Le plasma fut dilue deux fois avec de l’eau distillee et passe sur une colonne de Sepharose-lysine 6B, prkparee selon Cuatrecasas [8]. Dans cette mtthode le Sepharose est couple a de la lysine qui fixe le plasminogene. La colonne a ensuite ete rincee avec un tampon phosphate 0,l M pH 7,4. Lorsque I’absorbance de I’eluat fut inferieure A 0,Ol a 280 nm, le plasminogene fut eluk avec une solution d’acide 8-aminocaproique 0,2 M en tampon phosphate pH 7,4. Des fractions de 2 ml furent collectees et concentrees a l’aide d’un appareil Diaflo-Amicon. L’ultracentrifugation analytique a montrt qu’a ce stade le plasminogene etait accompagne d’un compo- sant lourd de coefficient de sedimentation de 18- 19 S. Ce constituant fut separk du plasminogene par chromatographie sur une colonne de BiogeI A 1,5 m, eluee comme precedemment. Le plasminogene ainsi purifie fut dialyse contre 2 1 de tampon phosphate de sodium 0,01 M pH 7,4, contenant du chlorure de sodium 0,l M. A cette etape de purification le plas- minogene donne un seul pic a l’ultracentrifugation analytique de coefficient de sedimentation egal a 5,8 S.
Afin de verifier que notre methode de preparation donnait une proteine native, I’activation du zymogene en enzyme fut realisee g r k e a l’urokinase ((Leo reagent )) et l’activite plasmique mesuree par le test a
l’azocaseine, selon la methode de Charney et Toma- relli [9] (M. Grimard, a publier).
Toutes les experiences decrites par la suite furent rtalistes sur du plasminogene prepare de cette maniere classique.
Nous avons pense que la liberation du plasmino- gene de la colonne d’affinite pouvait ttre rkalisee par un gradient de pH. 11 est raisonnable de penser que la fixation de la lysine sur le plasminogene implique un carboxyle dont la protonation inhibe cette fixation, ainsi qu’il a ett montre pour la trypsine et ses in- hibiteurs [lo]. Le gradient de pH fut simplement obtenu en rinqant la colonne d’affinite par une solu- tion de phosphate 0,2 M pH 3,O. Le plasminogene est apparu comme un pic unique a partir de pH 4. L’elution par l’acide E-aminocaproique etait ainsi evitee. Ces fractions, 2 differents pH acides, furent reunies et dialyskes contre une solution tampon de phosphate de sodium 0,l M pH 7,4, contenant du chlorure de sodium 0,l M. Nous avons alors observe qu’une petite partie precipitait au cours de la dialyse. Le passage de pH acide a pH neutre etait partiellement irreversible et nous avons donc etudi6 cette transition.
Liu et Mertz ont montrk que l’elution de la colonne d’affinite par de l’acide acktique 0,l M pH 3 donnait un plasminogene alter&, dont une partie preci- pitait lors de la dialyse contre un tampon phosphate pH 7,4 [ l l ] . La nature de l’acide utilisi. ne semble donc pas jouer sur l’insolubilisation du plasmino- gene.
Produits utilisks
Le compose modele, N-a-acetyl-L-tryptophane ethylester etait un produit des laboratoires Mann Research. Les tampons utilises pour les titrages ainsi que I’iodure de sodium ont ete fournis par Merck. Les tampons faits avec de l’eau bidistillee furent ajustes avec un pH metre Tacussel. Nous avons vkrifie qu’ils ne contenaient pas d’impuretk fluorescente. Le chlo- rure de guanidine, provenant de chez Merck, a ete purifie deux fois par cristallisation dans l’ethanol. L’absorbance a 280 nm d’une solution 6 M ne dC- passait alors pas 0,2.
Spectrophotomktrie d’uhsorption
Les spectres d’absorption ont ete enregistres sur un spectrophotometre Cary 15. Les spectres de difference d’absorption ont ete obtenus sur I’echelle Ctalee 0-0,l unite d’absorbance, en reglant le voltage des dynodes sur 4, la largeur de fente ne depassant pas 1 mm. Pour ces spectres, la ligne de base fut enregistree en portant la temperature de la cellule de mesure et celle de la cuve de reference a 25 “C. Ensuite l’echantillon etait porte a une autre temperature
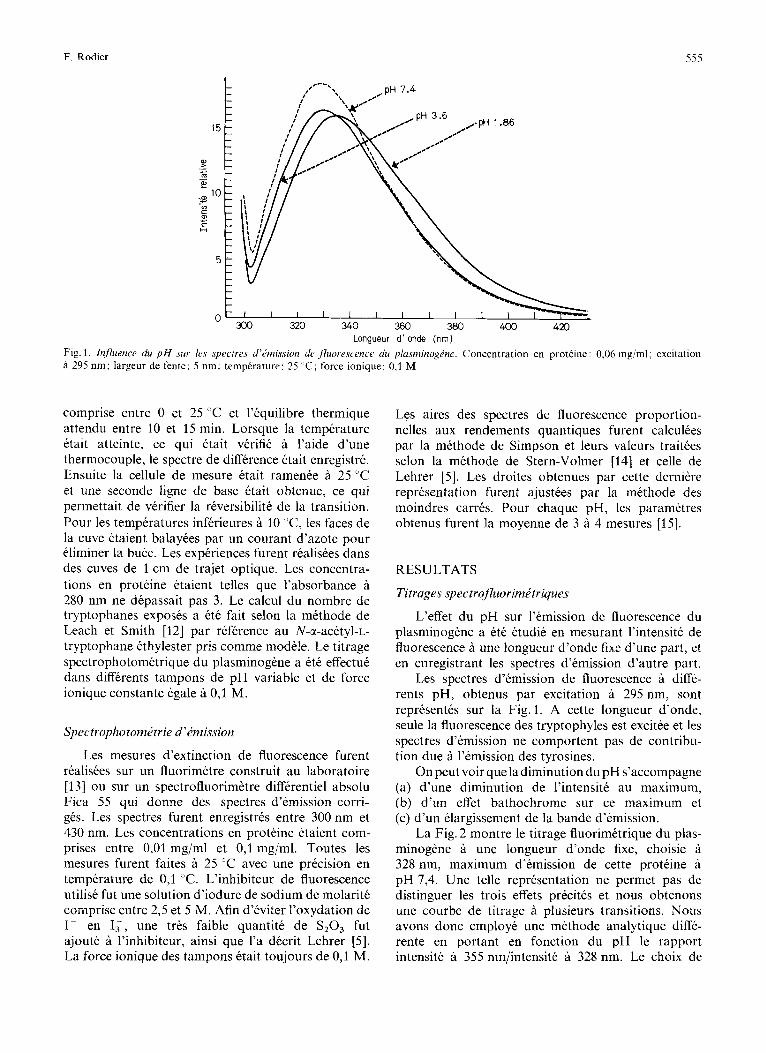
F. Rodier 555
Longueur d ‘ onde (nm) Fig. 1, Influence du p H sur Irs spectres d’imission de fhorescence du plusminogZ.ne. Concentration en proteine: 0,06 mg/ml; excitation a 295 nm; largeur de fente: 5 nm; temperature: 25 ’ C ; force ionique: 0,l M
comprise entre 0 et 25 “C et l’kquilibre thermique attendu entre 10 et 15 min. Lorsque la temperature etait atteinte, ce qui Ctait vkrifie a l’aide d’une thermocouple, le spectre de difference etait enregistre. Ensuite la cellule de mesure Ctait ramenee a 25 “C et une seconde ligne de base etait obtenue, ce qui permettait de verifier la reversibilite de la transition. Pour les temperatures inferieures a 10 “C, les faces de la cuve etaient balayees par un courant d’azote pour eliminer la buee. Les experiences furent realisees dans des cuves de 1 cm de trajet optique. Les concentra- tions en proteine etaient telles que I’absorbance a 280 nm ne depassait pas 3. Le calcul du nombre de tryptophanes exposes a ete fait selon la methode de Leach et Smith [I21 par reference au N-a-acetyl-L- tryptophane Cthylester pris comme modde. Le titrage spectrophotometrique du plasminogene a etk effectui: dans differents tampons de pH variable et de force ionique constante egale d 0,l M.
Spect ropho tomkt rie d’kmission
Les mesures d’extinction de fluorescence furent realisees sur un fluorimetre construit au laboratoire [13] ou sur un spectrofluorimetre differentiel absolu Fica 55 qui donne des spectres d’emission corri- ges. Les spectres furent enregistrks entre 300 nm et 430 nm. Les concentrations en proteine etaient com- prises entre 0,Ol mg/ml et 0,l mg/ml. Toutes les mesures furent faites ri 25 “C avec une precision en temperature de 0,l “C. L’inhibiteur de fluorescence utilise fut une solution d’iodure de sodium de molarite comprise entre 2,5 et 5 M. Afin d’eviter l’oxydation de I - en I;, une tres faible quantite de S,O, fut ajoute a l’inhibiteur, ainsi que l’a decrit Lehrer [ 5 ] . La force ionique des tampons etait toujours de 0,l M.
Les aires des spectres de fluorescence proportion- nelles aux rendements quantiques furent calculees par la mkthode de Simpson et leurs valeurs traittes selon la methode de Stern-Volmer [14] et celle de Lehrer [ 5 ] . Les droites obtenues par cette derniere representation furent ajustees par la methode des moindres carrks. Pour chaque pH, les parametres obtenus furent la moyenne de 3 A 4 mesures [15].
RESU LTATS
Titrages spectro~iilorimktriques
L’effet du pH sur l’emission de fluorescence du plasminogene a Cte Ctudie en mesurant I’intensitt de fluorescence 8 une longueur d’onde fixe d’une part, et en enregistrant les spectres d’emission d’autre part.
Les spectres d’emission de fluorescence a diffe- rents pH, obtenus par excitation a 295 nm, sont representes sur la Fig. 1. A cette longueur d’onde, seule la fluorescence des tryptophyles est excitee et les spectres d’emission ne comportent pas de contribu- tion due a l’emission des tyrosines.
On peut voir que la diminution du pH s’accompagne (a) d’une diminution de l’intensite au maximum, (b) d’un effet bathochrome sur ce maximum et (c) d’un elargissement de la bande d’emission.
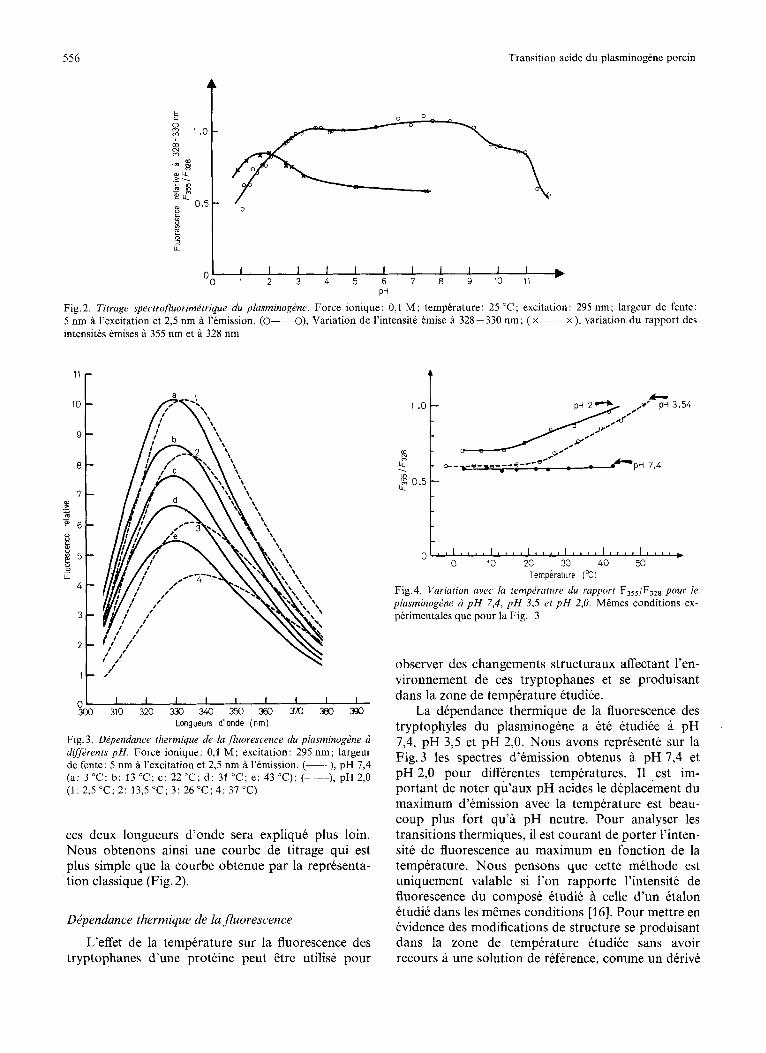
La Fig. 2 montre le titrage fluorimetrique du plas- minogene A une longueur d’onde fixe, choisie d 328 nm, maximum d’emission de cette proteine ii pH 7,4. Une telle representation ne permet pas de distinguer les trois effets precitts et nous obtenons une courbe de titrage 5 plusieurs transitions. Nous avons donc employe une methode analytique diffe- rente en portant en fonction du pH le rapport intensite a 355 nmlintensite a 328 nm. Le choix de
556
10
9 -
a -
J
e %
._ I m -
a,
[ - U
t
-
7 -
6 -
5 -
4 -
3 -
2 -
Transition acide du plasminogene porcin
1 - /
I I I I I I I I I
b PH
Fig. 2. Titrage spectrofluorimttrique du plasminogPne. Force ionique: 0,l M; temperature: 25 ‘C; excitation: 295 nm; largeur de fente: 5 nm a I’excitation et 2,s nm a l’emission. (0-o), Variation de I’intensite tmise a 328 - 330 nm; ( x ~ x ), variation du rapport des intensites emises a 355 nm et a 328 nm
r
Longueurs d‘onde (nm)
Fig. 3. Dependance thermique de la fluorescence du plasminogPne 6 dfftrents pH. Force ionique: 0,l M; excitation: 295 nm; largeur de fente: 5 nm a l’excitation et 2,5 nm a l’emission. (-), pH 7,4 (a: 3 “C; b: 13 “C; c : 22°C; d : 31 “C; e: 43°C); (----), pH 2,O (1 :2 ,5”C;2 : 13 ,5”C;3 :26”C;4 : 37°C)
ces deux longueurs d’onde sera explique plus loin. Nous obtenons ainsi une courbe de titrage qui est plus simple que la courbe obtenue par la representa- tion classique (Fig. 2).
Dipendance thermique de la fluorescence
L’effet de la temperature sur la fluorescence des tryptophanes d’une proteine peut Ctre utilise pour
t
O 0 ,,.... 10 20 Ternpbrature 30 (“c) 40 50
Fig.4. Variation avec la temptraturr du rapport F,,,/F,,, pour le plasrninogPne a p H 7,4, p H 3,5 et p H 2,O. Mdmes conditions ex- perimentales que pour la Fig. 3
observer des changements structuraux affectant l’en- vironnement de ces tryptophanes et se produisant dans la zone de temperature etudike.
La dependance thermique de la fluorescence des tryptophyles du plasminogene a ete etudiee a pH 7,4, pH 3,5 et pH 2,O. Nous avons represente sur la Fig. 3 les spectres d’emission obtenus a pH 7,4 et pH 2,O pour differentes temperatures. I1 est im- portant de noter qu’aux pH acides le deplacement du maximum d’emission avec la temperature est beau- coup plus fort qu’a pH neutre. Pour analyser les transitions thermiques, il est courant de porter l’inten- site de fluorescence au maximum en fonction de la temperature. Nous pensons que cette mkthode est uniquement valable si l’on rapporte l’intensite de fluorescence du compose ktudie a celle d’un etalon etudik dans les mCmes conditions [16]. Pour mettre en evidence des modifications de structure se produisant dans la zone de temperature etudike sans avoir recours a une solution de reference, comme un derive
F. Rodier
5 . 9
6 .9
3 . 6
3 . 0
2 . 6
551
I I I I I I I I I I
l / [NaI] (M-’)
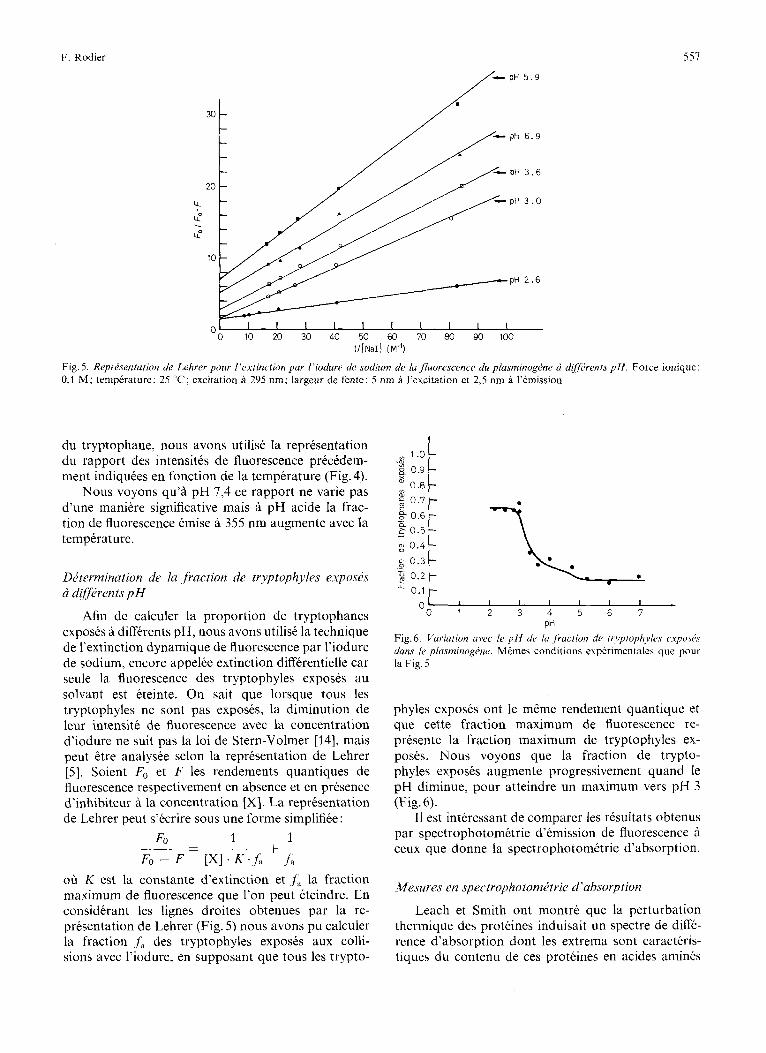
Fig. 5. Reprisentation de Lehrer pour I’extinction par l’iodure de sodium de ia fluorescence du plusrninogtke ri diJffrents p H . Force ionique: 0,l M; temperature: 25 T; excitation a 295 nm; largeur de fente: 5 nm a I’excitation et 2,5 nm a I’emission
0 10 20 30 40 50 60 70 80 90 100 01
du tryptophane, nous avons utilise la representation du rapport des intensites de fluorescence preckdem- ment indiquees en fonction de la temperature (Fig. 4).
Nous voyons qu’a pH 7,4 ce rapport ne varie pas d’une maniere significative mais a pH acide la frac- tion de fluorescence emise a 355 nm augmente avec la temperature.
Ddtermination de la .fraction de trjytophyles exposb ri dffZrentspH
Afin de calculer la proportion de tryptophanes exposes li diffkrents pH, nous avons utilise la technique de I‘extinction dynamique de fluorescence par l’iodure de sodium, encore appelee extinction differentielle car seule la fluorescence des tryptophyles exposes au solvant est eteinte. On sait que lorsque tous les tryptophyles ne sont pas exposes, la diminution de leur intensite de fluorescence avec la concentration d’iodure ne suit pas la loi de Stern-Volmer [14], mais peut Etre analysee selon la representation de Lehrer [5]. Soient Fo et F les rendements quantiques de fluorescence respectivement en absence et en presence d’inhibiteur .h la concentration [XI. La representation de Lehrer peut s’ecrire sous une forme simplifiee :
1 1 + - Fo - F [ X ] . K . b f a
- - ~ _ _ - Fo ~ _ _
oh K est la constante d’extinction et f, la fraction maximum de fluorescence que l’on peut eteindre. En considerant les lignes droites obtenues par la re- presentation de Lehrer (Fig. 5) nous avons pu calculer la fraction f a des tryptophyles exposes aux colli- sions avec l’iodure, en supposant que tous les trypto-
D a 0 . 4 1
0.3 $ 0.2
0.1 L L
0‘ I I I I I I I - 0 1 2 3 4 5 6 7
PH Fig. 6. Variation avec lr p l l de lu .fraction de tvyptophyles c~.xpo.sPs dam lr plasminog6ne. Mdmes conditions experimentales que pour la Fig. 5
phyles exposes ont le mCme rendement quantique et que cette fraction maximum de fluorescence re- presente la fraction maximum de tryptophyles ex- poses. Nous voyons que la fraction de trypto- phyles exposes augmente progressivement quand le pH diminue, pour atteindre un maximum vers pH 3 (Fig. 6) .
11 est interessant de comparer les resultats obtenus par spectrophotometrie d’kmission de fluorescence ii ceux que donne la spectrophotometrie d’absorption.
Mesures en spectrophotonzdtrie d’absorption
Leach et Smith ont montre que la perturbation thermique des proteines induisait un spectre de diffe- rence d’absorption dont les extrema sont caracteris- tiques du contenu de ces proteines en acides aniines
558 Transition acide du plasminogene porcin
I “ I I I 1 I 1 I I
I 1 I 240 250 260 270 280 290 300 310 320 330 340
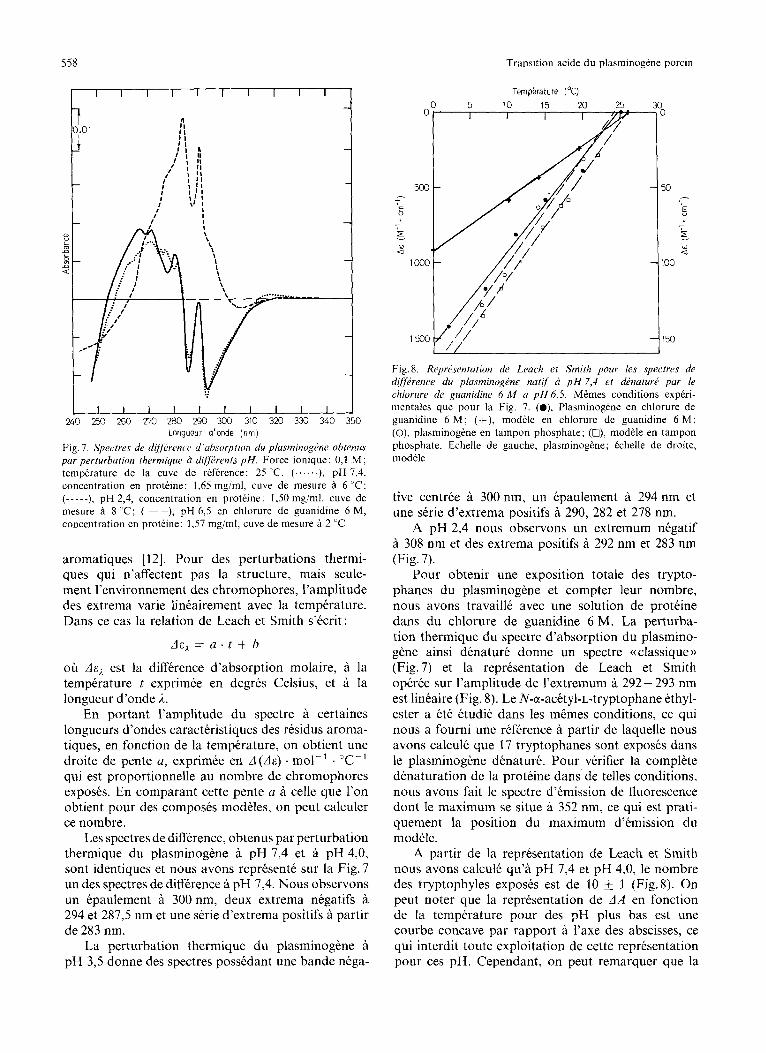
Longueur d‘onde ( n m ) Fig. I. Spectres de dijjiircvw d‘crhsorption du plasminogene ohtenus par perturbation thermique d di@rents p H . Force ionique: 0,1 M ; temperature de la cuve de reference: 25 ”C. (......), pH 7,4, concentration en protkine: 1,65 mg/ml, cuve de mesure a 6 “C; (-----), pH 2,4, concentration en proteine: 1,50 mg/ml, cuve de mesure a 8 “C; (--), pH 6,5 en chloriire de guanidine 6 M, concentration en proteine: 1,57 mg,’ml, cuve de mesure i 2 ”C
aromatiques 1121. Pour des perturbations thermi- ques qui n’affectent pas la structure, mais seule- ment I’environnement des chromophores, l’amplitude des extrema varie linkairement avec la temperature. Dans ce cas la relation de Leach et Smith s’ecrit :
A e , = a . t + b
ou A E , est la difference d’absorption molaire, a la temperature t exprimte en degres Celsius, et a la longueur d’onde A.
En portant l’amplitude du spectre a certaines longueurs d’ondes caracteristiques des residus aroma- tiques, en fonction de la temperature, on obtient une droite de pente a, exprimee en A ( ~ E ) . mol-’ . “C-’ qui est proportionnelle au nombre de chromophores exposes. En comparant cette pente a A celle que l’on obtient pour des composes modeles, on peut calculer ce nombre.
Les spectres de difference, obtenus par perturbation thermique du plasminogene pH 7,4 et 9 pH 4,0, sont identiques et nous avons represente sur la Fig. 7 un des spectres de difference B pH 7,4. Nous observons un epaulement a 300 nm, deux extrema negatifs a 294 et 287,5 nm et une serie d’extrema positifs A partir de 283 nm.
La perturbation thermique du plasminogene a pH 3 3 donne des spectres posskdant une bande nega-
Tempbrature (“C) 0 5 10 15 20 25
- - 5
E a
- W
I 3
50 - - E u - i - w a
100
150
Fig.8. Represenrution de Leach et Smith pour les .ypectres de rliflirence du phsminog6ne natif 2 p H 7 , 4 et denature par le chiorurr de guanidine 6 M d p H 6,5. M&mes conditions experi- mentales que pour la Fig. 7. (O), Plasminogene en chlorure de guanidine 6 M ; (+), rnodele en chlorure de guanidine 6 M ; (O), plasrninogene en tampon phosphate; (O), mod& en tampon phosphate. Echelle de gauche, plasminogene; echelle de droite, modele
tive centrke a 300 nm, un Cpaulement a 294 nm et une serie d’extrema positifs a 290, 282 et 278 nm.
A pH 2,4 nous observons un extremum negatif A 308 nm et des extrema positifs a 292 nm et 283 nm (Fig. 7).
Pour obtenir une exposition totale des trypto- phanes du plasminogene et compter leur nombre, nous avons travail16 avec une solution de proteine dans du chlorure de guanidine 6 M. La perturba- tion thermique du spectre d’absorption du plasmino- gkne ainsi denature donne un spectre ((classique)) (Fig.7) et la representation de Leach et Smith operke sur I’amplitude de I’extremum a 292- 293 nm est linkaire (Fig. 8). Le N-a-acetyl-L-tryptophane ethyl- ester a ete etudie dans les mtmes conditions, ce qui nous a fourni une reference a partir de laquelle nous avons calculk que 17 tryptophanes sont exposes dans le plasminogene denature. Pour verifier la complete denaturation de la proteine dans de telles conditions, nous avons fait le spectre d’kmission de fluorescence dont le maximum se situe A 352 nm, ce qui est prati- quement la position du maximum d’emission du modele.
A partir de la representation de Leach et Smith nous avons calcule qu’a pH 7,4 et pH 4,0, le nombre des tryptophyles exposes est de 10 4 1 (Fig.8). On peut noter que la representation de AA en fonction de la temperature pour des pH plus bas est une courbe concave par rapport A l’axe des abscisses, ce qui interdit toute exploitation de cette representation pour ces pH. Cependant, on peut remarquer que la
F. Rodier 559
-l??rL - 4 -3 0 5 10 15 20 25 30
Temperature ( “ C )
Fig. 9. Reprisenfation de Leach er Smithpour les spectres de dijJirence du plusrnino&e rl p H 3 J et a p H 2,4. Pour pH 2.4, m&mes conditions experimentales que pour la Fig. 7; pour pH 3,5, concen- tration en proteine: 1,28 mg/ml. (a), 292 nm, pH 3.5; (+)> 292 nm, pH 2,4; (A), 305 nm, pH 2,4; (o), 300 nm, pH 3 3
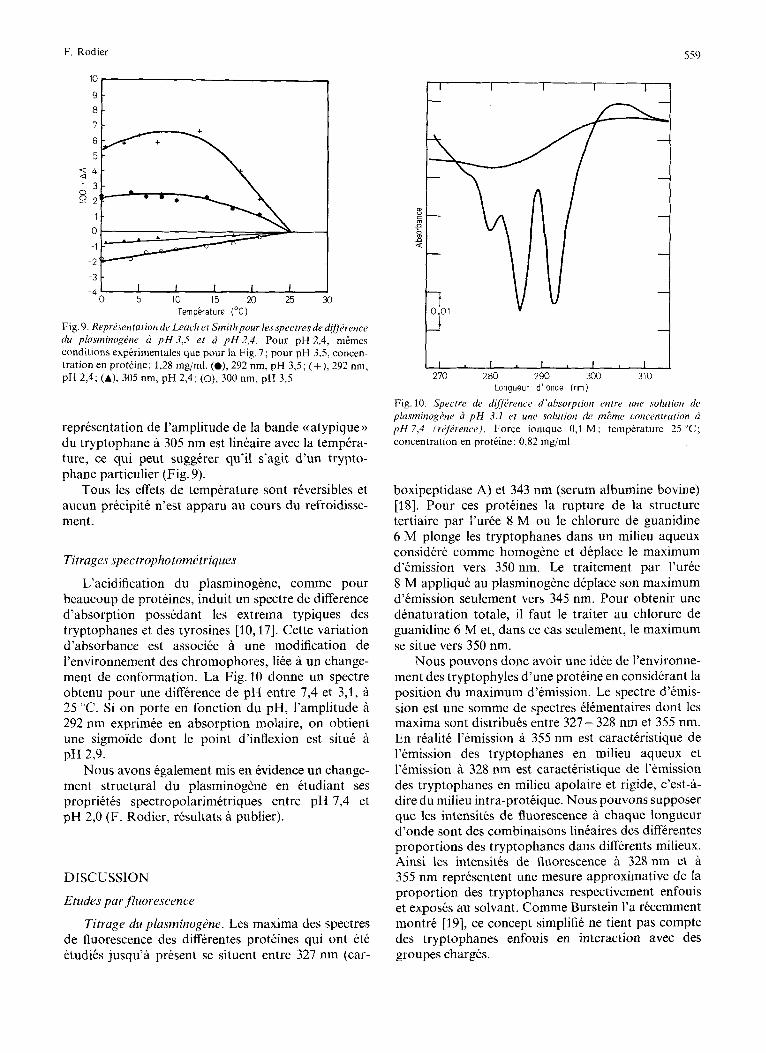
representation de l’amplitude de la bande (( atypique )) du tryptophane a 305 nm est linkaire avec la tempera- ture, ce qui peut suggkrer qu’il s’agit d’un trypto- phane particulier (Fig. 9).
Tous les effets de temperature sont rkversibles et aucun precipite n’est apparu au cours du refroidisse- ment.
Titrages spectrophotomdtriques
L’acidification du plasminogene, comme pour beaucoup de proteines, induit un spectre de difference d’absorption possedant les extrema typiques des tryptophanes et des tyrosines [lo, 171. Cette variation d’absorbance est associee a une modification de l’environnement des chromophores, liee a un change- ment de conformation. La Fig.10 donne un spectre obtenu pour une difference de pH entre 7,4 et 3,1, .& 25 “C. Si on porte en fonction du pH, I’amplitude a 292 nm exprimee en absorption molaire, on obtient une sigmoide dont le point d’inflexion est sit& A pH 2,9.
Nous avons egalement mis en evidence un change- ment structural du plasminogene en etudiant ses proprietks spectropolarimitriques entre pH 7,4 et pH 2,O (F. Rodier, resultats a publier).
DISCUSSION
Etudes par fluorescence
Titrage du plasminogene. Les maxima des spectres de fluorescence des differentes proteines qui ont ete ttudiks jusqu’a present se situent entre 327 nm (car-
7 0.01
270 280 290 300 310 Longueur d ’onde (nrn)
Fig. 10. Spectre de d#?r.ence d’ahsorption entre une solution di, plasrninog6nr 2 p H 3.1 el une solution de mgme concentration p H 7,4 (rijirence). Force ionique 0,1 M: temperature 25 “C; concentration en proteine: 0,82 mg/ml
boxipeptidase A) et 343 nm (serum albumine bovine) [18]. Pour ces proteines la rupture de la structure tertiaire par I’urCe 8 M ou le chlorure de guanidine 6 M plonge les tryptophanes dans un milieu aqueux consider6 comme homogene et deplace le maximum d’emission vers 350 nm. Le traitement par l’uree 8 M applique au plasminogene deplace son maximum d’kmission seulement vers 345 nm. Pour obtenir une denaturation totale, il faut le traiter au chlorure de guanidine 6 M et, dans ce cas seulement, le maximum se situe vers 350 nm.
Nous pouvons donc avoir une idee de I’environne- ment des tryptophyles d’une proteine en considerant la position du maximum d’emission. Le spectre d’emis- sion est une somme de spectres klementaires dont les maxima sont distributs entre 327- 328 nm et 355 nm. En realit6 l’emission a 355 nm est caracteristique de l’bmission des tryptophanes en milieu aqueux et l’emission d 328 nm est caracteristique de l’emission des tryptophanes en milieu apolaire et rigide, c’est-A- dire du milieu intra-proteique. Nous pouvons supposer que les intensites de fluorescence A chaque longueur d’onde sont des combinaisons lineaires des differentes proportions des tryptophanes dam differents milieux. Ainsi les intensites de fluorescence a 328 nm et h 355 nm representent une mesure approximative de la proportion des tryptophanes respectivement enfouis et exposes au solvant. Comme Burstein l’a recemment montrk [19], ce concept simplifie ne tient pas compte des tryptophanes enfouis en interaction avec des groupes charges.

560 Transition acide du plasminogkne porcin
La diminution du pH provoque une extinction de fluorescence du plasminogkne et augmente la contri- bution des tryptophanes (( polaires )) dans le spectre d’emission. L’influence du pH sur la fluorescence du tryptophane et de ses composes modeles a ete 6tudit.e par White [20]. Cet auteur a montre que la protonation du carboxyle Cteint la fluorescence de l’indole. Pour la tryptamine et l’ester methylique du tryptophane, le meme phenomkne se produit a bas pH avec un pK de 1,8. Ce dernier quenching est expli- que par une extinction collisionnelle du noyau indole par les protons de la solution. Ce quenching dynamique se produit pour les pH inferieurs a 2 [21].
Dans une proteine, deux classes d’extinction peuvent donc se produire: par protonation d’un car- boxyle au voisinage d’un tryptophane et par collision du noyau indole par les protons de la solution. Nous pensons que la diminution de fluorescence n’est pas necessairement due a un changement de conforma- tion, mais peut etre liee A une extinction de la fluorescence de l’indole des tryptophanes par les protons de la solution et non par les carboxyles de chaines voisines. Ainsi la representation classique de l’intensitk de fluorescence au maximum d’emission ou bien a une longueur d’onde fixe en fonction du pH ne permet pas de distinguer entre ces deux pheno- menes. Nous avons trouve une representation ori- ginale, valable aussi bien pour analyser l’effet du pH que l’effet de la temperature sur la fluorescence des proteines en portant les variations du rapport: emission a 355 nm sur emission a 328 nm. En portant ce rapport en fonction du pH, nous obtenons une courbe de titrage decomposable en deux parties. De pH 7,4 a pH 2, nous observons le demasquage des tryptophyles (augmentation du rapport). Ce demas- quage est dii a un changement de conformation du plasminogene. Pour expliyuer la deuxieme partie, au dessous de pH 2, il est rationnel de penser que l’ex- tinction dynamique par les protons de la solution est plus marquee pour les tryptophyles exposes que pour les tryptophyles enfouis. La forme de la courbe de titrage du plasminogene peut Ctre expliquke en sup- posant qu’elle est la somme de la courbe de dkmas- quage des tryptophyles et de la courbe de titrage des noyaux indoles des tryptophanes externes.
EJfet de la tempirature. Lorsque le plasminogine est chauffe, a pH 7,4, le rapport F35qF328 reste constant jusyu’a 45 “C. Ceci indique qu’il n’y a pas de deformation significative du spectre et que le seul effet de la temperature est un effet d’extinction que l’on interprite classiquement par l’augmentation de conversion interne [22]. D’autre part, il n’y a pas de dkplacement de la position du maximum d’emission du spectre qui reste au voisinage de 328- 330 nm (Fig.3). On peut obtenir le meme phenomene en chauffant une solution aqueuse de tryptophane. Le rendement quantique de fluorescence diminue avec
la temperature, sans qu’il y ait de changement dans la forme du spectre d’emission [16]. Tant qu’il n’y a pas de changement de conformation de la proteine lie a un effet de temperature, le seul effet observable est le quenching de fluorescence des tryptophyles.
A pH 3,5 le quenching de fluorescence existe pour les memes raisons que precedemment, mais le maxi- mum d’emission se dkplace de 330 nm a - 2 “C jusqu’a 338 nm 51 “C. La variation du rapport F355/F328 re- vele une deformation du spectre dans la region des grandes longueurs d’onde.
A pH 2,4, le rapport varie d’une maniere sem- blable et on observe Cgalement un effet bathochrome du spectre d’emission.
La temperature ri laquelle le rapport E j 5 5 / F 3 2 8
commence a augmenter a pH 3 3 est de 22 “C. A pH 2,4 cette temperature tombe a 13 “C. Ceci indique un effet du pH sur la sensibilite thermique du plasminogene. On voit yue le calcul du rapport F 3 5 5 / F 3 2 8 klimine l’effet de temperature sur les chro- mophores (conversion interne) pour ne laisser subsister que l’effet propre a la proteine, a savoir le changement de conformation.
A I’aide de ces premiers resultats, nous pouvons representer schematiquement l’effet du pH sur la conformation du plasminogine par une ouverture partielle de la proteine qui demasque des tryptophyles. A pH constant, un autre demasquage se produit sous l’influence de la temperature et ceci pour les pH acides; plus le pH est bas, plus le demasquage est important.
A temperature constante, la transition induite par la diminution de pH est partiellement reversible car une partie du plasminogene precipite (environ 10 %). I1 ne s’agit donc pas d’un simple equilibre de trans- conformation entre deux formes de plasminogene mais d’un schema plus complexe ou une voie de formation de proteine denaturee doit etre ajoutee.
L’analyse de la courbe obtenue a partir des mesures d’extinction dynamique de l’exposition des trypto- phanes a differents pH revile que le demasquage de ces chromophores est continu et atteint un maximum vers pH 3. Ceci iniplique une correlation entre la variation du rapport F&/F328 et la position du maxi- mum d’emission d’une part et la fraction maximum de la fluorescence que l’on peut eteindre d’autre part.
Etudes par spectvophotomttrie d’absorption
Perturbation de temptrature. L’analyse des spectres de difference Crees par perturbation thermique revele une difference dans la rkponse des chromophores entre pH 7,4 et pH 2,O. I1 est evident que la methode de Leach et Smith ne peut etre utilisee pour calculer le nombre de tryptophanes exposes A pH 2,4 et pH 3,5 car cette methode est basee sur la linearit6 de la varia- tion de AA avec la temperature et I’additivite des

F. Rodier 561
perturbations propres A chaque chromophore. De plus les variations d’absorbance doivent Etre negatives pour les bandes caracteristiques du tryptophane, ainsi qu’il en est pour le compose modele.
Comme l’a montri: Donovan [23], la perturbation thermique produit aussi une pertubation de l’environ- nement des chromophores enfouis, la perturbation se transmettant par la chaine polypeptidique. Ainsi, le nombre de chromophores comptes par cette mtthode peut Etre plus grand que celui compte par la mtthode de l’extinction dynamique de la fluorescence.
C’est effectivement le cas a pH 7,4, puisque nous trouvons par perturbation thermique 10 tryptophanes exposes dans la proteine native et 17 dans la proteine totalement denaturee, soit une fraction d’exposition de 58,8 %, tandis que la methode d’extinction dynami- que donne environ 20%. I1 est evident que cette derniere mkthode est 1imiti.e car l’iodure ne peut pas atteindre des tryptophanes enfouis dans des crevasses, mais sensibles aux variations de tempera- ture. De plus nous avons supposC que tous les trypto- phanes exposes au solvant avaient le meme rende- ment quantique, ce qui n’est qu’une approximation, conduisant A un calcul d’exposition moyenne.
En ce qui concerne les pH 3,5 et 2,4, la reference a des modeles n’est pas suffisante pour expliquer l’inversion apparente des bandes du spectre de diffe- rence. On peut noter dans ces spectres une bande situee a 308 nm a pH 2,4 et une situke a 300 nm a pH 3,5. Cette bande a kti: dkja signalee par Anan- thanarayanan et Bigelow [24,25], comme etant une bande mineure correspondant a une transition du tryptophane interdite normalement pour des raisons de symetrie et devenant permise par suite de la crea- tion d’un environnement assymetrique. Bello [26] a montre que la perturbation thermique de l’a-chymo- trypsinogene et de la P-lactoglobuline donne des spectres possedant aussi un minimum a 303 nm. Cet auteur a prouve qu’un tel minimum pouvait etre Cree par perturbation thermique du tryptophane dans un melange de 80 % de N-mithylacetamide et de 20 % de N , N ’-dimethylacetamide. I1 resulte de ces considera- tions que la reponse spectrale d’un chromophore a une perturbation thermique peut &tre perturbee si une interaction est possible entre le chromophore et un ensemble de groupes polaires et apolaires. La courbure de la representation de AA en fonction de la tem- perature suggere plus schtmatiquement un antago- nisme entre deux types d’interactions, interactions hydrophiles stabilisees a basse temperature et inter- actions hydrophobes stabilisees a haute temperature.
Perturbation acide. Le titrage spectrophotometri- que du plasminogene est classique en ce sens qu’il donne des spectres identiques a ceux obtenus par Delaage et al. [27] dans le cas du chymotrypsinogene A et de la trypsine. On note Cgalement un extremum posi- tif a 305 nm, ainsi qu’il a tte observi: par d’Albis [lo]
et Foucault [17] dans le cas de la trypsine. Par conse- quent, la presence de cet extremum apparaissant entre 300 et 310 nm, anormal par rapport au modele, n’est pas due au type de perturbation utilisee, mais semble caracteristique d’un certain type de chromophores qui se trouve plus ou moins signal6 dans la littkrature. De plus le titrage spectrophotomktrique du plasmino- gene revele un pK de transconformation de 2,9 unites. Ce pK est apparent car I’acidification du plasminogene n’est que partiellement reversible. Cette faible valeur, comparee au pK de 4,6 d’un carboxyle normal, peut s’expliquer par la presence d’une charge positive au voisinage d’un carboxyle, ce qui a pour resultat d’augmenter la probabilite d’existence de la forme de- protonire -COO-, donc d’abaisser le pK d’ionisation de ce groupe.
Proposition d’un mkcanisme de transconformation
Nous expliquons les precedents resultats en sup- posant qu’il y a un groupe charge positivement au voisinage d’un carboxyle, en interaction avec un ou plusieurs chromophores. Lorsque le pH augmente, le carboxyle se deprotone et une liaison peut se former entre ce groupe anionique et un groupe charge posi- tivement. On forme ainsi un pont salin, phenomene deja connu dans le cas de la chymotrypsine Aa dont I’aspartate 184 est enfoui dans la forme native et interagit avec le groupe chargk positivement de l’isoleucine. I1 est d’ailleurs aussi probable qu’un tel pont salin existe pour la trypsine [27].
Par suite de la deprotonation du carboxyle, le groupe charge positivement ne peut plus interagir avec un ou plusieurs chromophores et les spectres de perturbation thermique ne sont pas anormaux. La protonation du carboxylate, rompant le pont salin, s’accompagne d’une ouverture partielle du plasmino- gene, entrainant un demasquage des tryptophanes. La rupture de ce pont salin a pour deuxikme consequence d’augmenter aux pH acides la sensibilite thermique de la proteine.
L’auteur tient a remercier le Dr S. Guinand, le Professeur G. Nemethy et le Dr G. Johannin pour leurs critiques sur ce manuscrit et les multiples discussions durant ce travail. Les ex- periences d’extinction differentielle ont &ti. faites en collaboration avec Mr G. Rossis.
Ce travail a beneficit. de credits du Centre National de la Recherche Scientifique (E.R.A. no 480).
BI BLI OG RAPHIE
1. Elkana, Y. (1968) J . Phys. Chem. 72, 3654-3661. 2. Lehrer, S. S. & Fasman, G. D. (1966) Biochem. Biophys. Res.
3. Herskovits, T. T. & Laskowski, M. (1962) J . Bid. Chem. 237,
4. Herskovits, T. T. & Sorensen, M., Sr (1968) Biochemistry, 7,
Commun. 23, 133-139.
2481 - 2482.
2533- 2542.

562 F. Rodier: Transition acide du plasminogene porcin
5 . Lehrer, S. S. (1971) Biochemistry, 10, 3254-3263. 6. Gerard, D., Laustriat, G. & Lami, H. (1972) Biochim. Biophys.
7. Deutsch, D. G. & Mertz, E. T. (1970) Science (Wash. D.C.)
8. Cuatrecasas, P., Wilchek, M. & Anfinsen, G. B. (1968) Proc.
9. Charney. J. & Tomarelli, R. M. (1974) J . Biot. Chem. 171,
10. D’Albis, A. (1968) These d’Etat, Universitt. Paris-Sud, Centre
11. Liu, T. H. & Mertz, E. T. (1971) Can. J. Biochem. 49, 1055-
12. Leach, S . J. & Smith, J. A. (1972)Znt. J . Protein Res. 4, 11-19. 13. Rodier, F. (1969) These de Specialite, Universitt Paris-Sud,
Centre d’orsdy. 14. Forster, Th. (1 951) Fluoreszenz organischer Verbindungen,
pp. 206-214, Vandenhoeck & Ruprecht, Gottingen. 15. Rossis, G. (1974) Rapport de stage de Diplame d’Etudes
Approfondies, Universitt de Paris-Sud, Centre d’Orsay. 16. Gally, J. A. & Edelman, G. M. (1962) Biochim. Biophys. Acfa,
Acta, 263,482-495.
170, 1095- 1096.
Natl Acad. Sci. U S . 61, 636-643.
501 - 505.
dOrsay.
1061.
60,499- 509.
17. Foucault, G. (1972) These de Specialite, Universite Paris-Sud, Centre d’Orsay.
18. Kronman, M. J. & Holmes, L. G. (1971) Photochenz. Photobiol. 14,113-134.
19. Burstein, E. A,, Vedenkina, N. S . & Ivkova, M. N. (1973) Photochem. Photobiol. 18, 263 - 269.
20. White, A. (1959) Biochem. J. 244, 359-365. 21. Weinrib, 1. & Steiner, R . F. (1971) in Excited States of’Proteins
and Nucleic Acid.s (R. F. Steiner & Weinrib, I., eds) pp. 277- 312, the MacMillan Press Ltd, London and Basingstoke.
22. Wehry, E. L. (1967) in Fluorescence, Theory, Practice and Instrumentation (Guilbault, C. C., ed.) p. 121, Marcel Dekker, Inc. New York.
23. Donovan, J . W. (1973) Methods in Enzymology, 27, 508. 24. Anathanarayanan, V. S . & Bigelow, C. C. (1969) Biochemistry, 8,
25. Anathanarayanan, V. S . & Bigelow, C. C. (1969) Biochemistry,
26. Bello, J. (1970) Biochemistry, 9, 3562-3568. 27. Delaage, M., Abita, J. P. & Lazdunski, M. (1968) Eur. J.
3717- 3723.
8, 3723 - 3728.
Biochenz. 5, 285-293.
F. Rodier, Laboratoire de Biologie Physico-Chimique, Bbtiment 433, Universitt de Paris-Sud, F-91405 Orsay, France





























