Embed Size (px)
Citation preview

Annals of the Rheumatic Diseases 1993; 52: 285-291
Protective effect of androgens against
inflammation induced cartilage degradation inmale rodents
J A P Da Silva, J-P Larbre, T D Spector, L A Perry, D L Scott, D A Willoughby
Department ofExperimentalPathology,St Bartholomew'sHospital MedicalCollege,London EClM 6BQ,United KingdomJ A P Da SilvaJ-P LarbreD A WilloughbyDepartment ofReproductivePhysiology,St Bartholomew'sHospital MedicalCollege,London EClM 6BQ,United KingdomL A PerryDepartent ofRheumatology,St Bartholomew'sHospital MedicalCollege,London ECIM 6BQ,United KingdomJ A P Da SilvaT D SpectorD L ScottCorrespondence to:Dr J A P Da Silva,Department ofRheumatology,St Bartholomew's HospitalMedical College,Charterhouse Square,London ECIM 6BQ,United Kingdom.Accepted for publication9 December 1992
AbstractObjectives-Rheumatoid arthritis (RA) isa disease which predominantly affectswomen. Interestingly, low serum andro-gen levels and clinical improvement withandrogen replacement have been reportedin male patients. The aetiopathogenic roleof sex hormones in arthritis and theirpotential long term effects on joint de-struction and disability remains unclear,however. This study was designed toinvestigate the potential influence of sexhormones on inflammation induced carti-lage degradation in male rodents.Methods-An in vivo model of cottonwrapped cartilage implants was used toassess the effects of androgen, oestradiol,and progesterone on inflammation in-duced cartilage degradation, and in vitrotechniques were used to investigatethe direct actions on cartilage metab-olism and cytokine production in maleanimals.Results-Orchidectomy resulted in ac-celerated cartilage damage which wasreversed by replacement of physiologicallevels ofandrogens. Granulomatous tissuefrom castrated male rodents producedhigher amounts of interleukin 1. Sex hor-mones reduced spontaneous proteoglycanloss in vitro but did not interfere withthe effects of interleukin 1 on culturedcartilage.Conclusions-Androgens appear to pro-tect cartilage from inflammation inducedbreakdown in male animals. These resultssupport a pathogenic role for hypo-androgenism in rheumatoid arthritis andsuggest that long term androgen replace-ment may help prevent joint damage anddisability.
due to the immunosuppressive properties ofandrogens.4 Androgen receptors have beendescribed in antigen presenting and processingcells in human synovia, suggesting a possiblemodulatory role at the core of local pathogenicmechanisms.5
Cartilage destruction is an early conse-quence of RA and plays a major part in theprogression of disease and resulting disability.The effects of androgens on cartilage damagedue to inflammatory processes have receivedlittle attention despite the demonstration ofsex hormone receptors in articular cartilagefrom different species and experimental evi-dence that these hormones may stimulateproteoglycan and type II collagen synthesis bycultured human articular chondrocytes.6The principal aim of the study was to
investigate the influence of manipulation ofandrogens and other sex hormones on in-flammation induced cartilage degradation inmale animals.
MethodsIN VIVO CARTILAGE DEGRADATION
Castration and sham operationMature male BALB/c mice (Tuck & Sons,United Kingdom) aged 9 to 10 weeks wereused, age matched in each experiment, andkept under controlled light conditions (14hours light, 10 hours dark). All surgicalprocedures were performed under intra-peritoneal anaesthesia with 0 07 ml ofcombined Hypnorm (Janssen) and Hypnovel(Roche) in water (1:1:2, v/v). Orchidectomywas carried out via a single abdominal incisionafter dissection and ligature of the spermaticcord. Sham operated animals had a similaroperation but the gonads were replaced in theabdomen after exposure. Animals were left torecover for two weeks before further use.
(Ann Rheum Dis 1993; 52: 285-29 1)
Rheumatoid arthritis (RA) is clearly affected bygender differences: the disease is not only twoto three times more common in women, butalso tends to be more aggressive and disablingin female patients despite an apparentlybalanced distribution of other risk factors.Furthermore, subclinical hypogonadism hasbeen consistently reported in male patients1 2and androgen replacement resulted in an
improvement of immunological and clinicalparameters of the disease.3 This is likely to be
Preparation of cartilageMale Wistar rats 150-170 g (Tuck & Sons)were killed and both femoral head cartilages(FHCs) were dissected free ofbone and pooledin Hanks's balanced salt solution (HBSS)containing 100 IU/ml penicillin and 100 ,ug/mlstreptomycin. The cartilages were thoroughlywashed in HBSS, trimmed out of any adherenttissue in a laminar flow cabinet and leftovernight in Dulbecco's modified Eagle'smedium (DMEM), without phenol redsupplemented with antibiotics as above, at
285
on Decem
ber 23, 2021 by guest. Protected by copyright.
http://ard.bmj.com
/A
nn Rheum
Dis: first published as 10.1136/ard.52.4.285 on 1 A
pril 1993. Dow
nloaded from
Da Silva, Larbre, Spector, Perry, Scott, Wiloughby
37°C in a humidified incubator (5% C0295%air) before further use for implantation orculture.
Cartilage implantsFemoral head cartilages were asepticallyprepared for implantation by wrapping insterile surgical gauze (Softsorb, Vernon-CarusLtd, United Kingdom) (5±0-15 mg weight)moisturised with HBSS. Each mouse receiveda bilateral subcutaneous implant of eitherwrapped or non-wrapped FHC each side ofthedorsal hump as described previously.7 Anumber of FHCs were kept frozen in DMEMfor control purposes. At the end of theexperiments the granuloma or non-wrappedcartilages were carefully dissected free fromsurrounding tissue. Femoral head cartilageswere processed for glycosaminoglycan (GAG)analysis and the results are expressed as thepercentage loss compared with frozen controls.The granulomas were weighed immediatelyand after drying for 48 hours at 56°C and thefluid volume and dry tissue mass werecalculated.
Assessment of levels ofsex hormones in serumsamplesLevels of oestradiol were assessed usinga commercially available radioimmunoassay(Diagnostic Products, Abingdon, UnitedKingdom). Serum progesterone was deter-mined by radioimmunoassay as supplied bythe North East Thames radioimmunoassay(NETRIA) unit. Testosterone was assessed byan inhouse radioimmunoassay as describedpreviously.8 Interassay coefficients of variationfor each steroid were less than 10% at threeappropriate concentrations. The same testos-terone assay was used to assess androgen levelsafter implantation of dihydrotestosteronecapsules given that the assay shows 80% crossreactivity between the two steroids.
601 .
50-
40 *
a:a0a
a mOU~~~~~~~~~~~~i
i ~~~~~~~~~~~~~~~~~~~i
_ ° * i i°i °== a-nn
Intact OCx OCx + T OCx + T OCx + DHT OCx + DHTmale male (2 weeks) (5 weeks) (2 weeks) (5 weeks)
animals animals
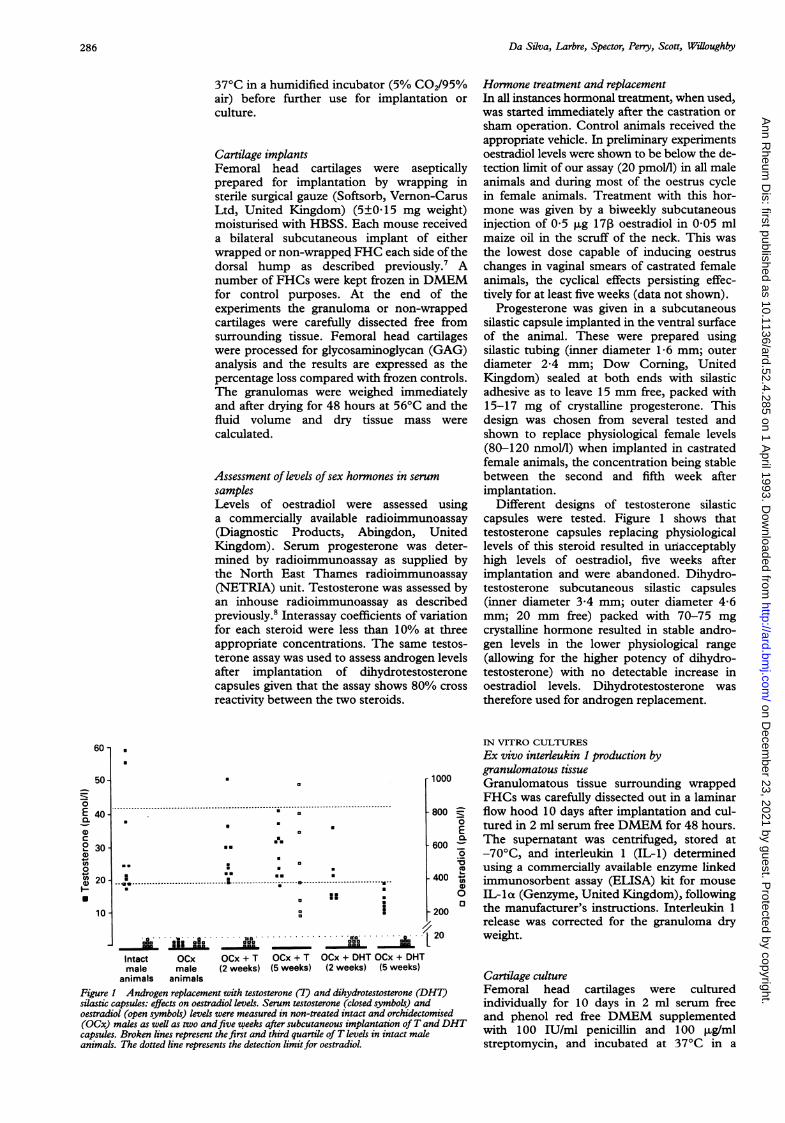
Figure I Androgen replacement with testosterone (T) and dihydrotestosterone (Dsilastic capsules: effects on oestradiol levels. Serum testosterone (closed symbols) antoestradiol (open symbols) levels were measured in non-treated intact and orchidectu(OCx) males as well as two andfive weeks after subcutaneous implantation ofT acapsules. Broken lines represent the first and third quartile of T levels in intact matanimals. The dotted line represents the detection limitfor oestradiol.
Hormone treatment and replacementIn all instances hormonal treatment, when used,was started immediately after the castration orsham operation. Control animals received theappropriate vehicle. In preliminary experimentsoestradiol levels were shown to be below the de-tection limit of our assay (20 pmol/1) in all maleanimals and during most of the oestrus cyclein female animals. Treatment with this hor-mone was given by a biweekly subcutaneousinjection of 0 5 ,ug 17, oestradiol in 0 05 mlmaize oil in the scruff of the neck. This wasthe lowest dose capable of inducing oestruschanges in vaginal smears of castrated femaleanimals, the cyclical effects persisting effec-tively for at least five weeks (data not shown).
Progesterone was given in a subcutaneoussilastic capsule implanted in the ventral surfaceof the animal. These were prepared usingsilastic tubing (inner diameter 1-6 mm; outerdiameter 2A4 mm; Dow Corning, UnitedKingdom) sealed at both ends with silasticadhesive as to leave 15 mm free, packed with15-17 mg of crystalline progesterone. Thisdesign was chosen from several tested andshown to replace physiological female levels(80-120 nmoll) when implanted in castratedfemale animals, the concentration being stablebetween the second and fifth week afterimplantation.
Different designs of testosterone silasticcapsules were tested. Figure 1 shows thattestosterone capsules replacing physiologicallevels of this steroid resulted in unacceptablyhigh levels of oestradiol, five weeks afterimplantation and were abandoned. Dihydro-testosterone subcutaneous silastic capsules(inner diameter 3-4 mm; outer diameter 4-6mm; 20 mm free) packed with 70-75 mgcrystalline hormone resulted in stable andro-gen levels in the lower physiological range(allowing for the higher potency of dihydro-testosterone) with no detectable increase inoestradiol levels. Dihydrotestosterone wastherefore used for androgen replacement.
IN VITRO CULTURESEx vivo interleukin 1 production bygranulomatous tissue
- 1000 Granulomatous tissue surrounding wrappedFHCs was carefully dissected out in a laminar
- 800 ^ flow hood 10 days after implantation and cul-0 tured in 2 ml serum free DMEM for 48 hours.
a600 The supernatant was centrifuged, stored at. -70'C, and interleukin 1 (IL-1) determinedc using a commercially available enzyme linked
- 400 ,, immunosorbent assay (ELISA) kit for mouseo IL- loa (Genzyme, United Kingdom), following
r 200 0 the manufacturer's instructions. Interleukin 1release was corrected for the granuloma dry
[20 weight.
Cartilage culture)HT) Femoral head cartilages were culturedd individually for 10 days in 2 ml serum freeomised and phenol red free DMEM supplementedendDHT wiith 100 IU/ml penicillin and 100 ,ug/ml
streptomycin, and incubated at 37°C in a
0
E
a.
a)C2 30-0)
0
'A 20-0)
110 -
286
.
.0
on Decem
ber 23, 2021 by guest. Protected by copyright.
http://ard.bmj.com
/A
nn Rheum
Dis: first published as 10.1136/ard.52.4.285 on 1 A
pril 1993. Dow
nloaded from
Effect of androgens on cartilage degradation
humidified incubator (5% C02/95% air), themedium being changed at day 5. Sex hormoneswere added to the medium from stock sol-utions in dimethylsulphoxide (DMSO) to finalconcentrations chosen to cover a range from10-fold below to 10-fold higher physiologicallevels in mature male Wistar rats as determinedin preliminary experiments. The concentrationofDMSO was kept at 0 01% in all groups.When used, mouse recombinant interleukin
lo (Genzyme) prepared in DMEM with 0-5%bovine serum albumin (BSA) was added to themedium in the second five day period to a finalconcentration of 1 ng/ml. A similar volume of0 5% BSA was added to control wells.Cultured FHCs were pulsed with 37 kBq/mlcarrier free Na235SO4 (Amersham Inter-national, United Kingdom) for the last 24hours of culture to evaluate proteoglycansynthesis. Supernatants were kept for thedetermination of GAGs.
Assessment of cartilage and mediumglycosaminoglycanFollowing implant or culture, FHCs were re-peatedly washed and digested overnight at 56°Cin 50 mM phosphate buffer (pH 7T0) con-taining 2 mM N-acetyl cysteine, 1 mM EDTA,and 6 U/ml papain. The GAG content ofcartilage digests and culture supernatants wasdetermined following the method of Farmdaleet al9 adapted to allow the use of a plate reader.Glycosaminoglycan loss from implanted FHCswas subsequently expressed as a percentage ofthe mean of frozen control cartilages. Thepercentage degradation of cultured FHCs wascalculated according to the formula: (GAGmedium/(GAG medium+GAG FHC)) X 100.
Assessment ofproteoglycan synthesis in culturecartilagePulsed FHCs were vigourously washed in cold10 mM MgSO4 and digested in papain asdescribed earlier. Aliquots of the digest wereprecipitated onto filter paper with 1% cetyl-pyridinium chloride and radioactivity in theprecipitate was measured by liquid scintillationcounting. Results are expressed in counts perminute (cpm) per milligram of cartilage weightafter blot drying.
Statistical methodsResults are expressed as mean (SEM) values.Statistical significance was evaluated by theMann-Whitney U test for less than 30 samplesand Student's t test for larger numbers. Valuesof p<0-05 (two tailed) were taken as significant.
ResultsEFFECTS OF CO'TTON PELETT RANTTG MA ON
CARTILAGE DEGRADATION
Groups of intact mice with a double implantofwrapped and non-wrapped FHC were killedat weekly intervals after implantation. Figure 2shows that GAG loss increased progressivelyafter the first week and the presence of cotton
Wrapped
* Non-wrapped
60-
CaCa)a '40 -c 0
coooe 20 -
0
-200 1 2 3 4
Weeks after implantation
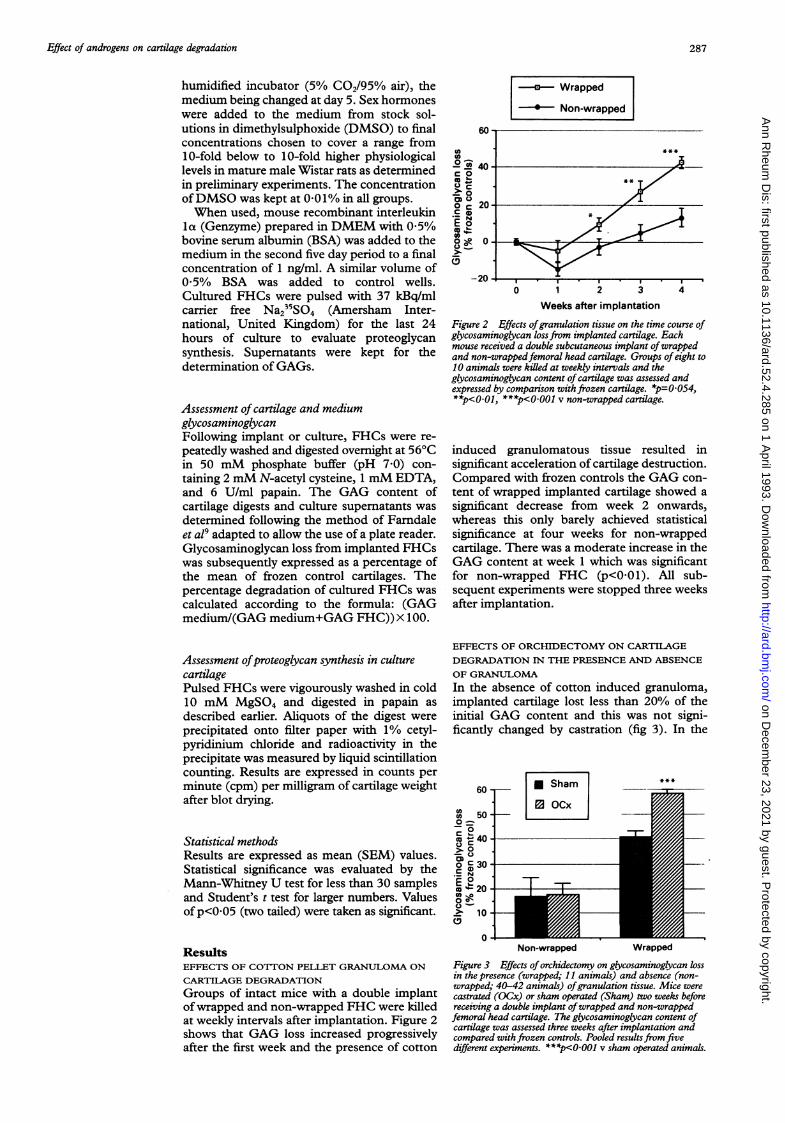
Figure 2 Effects ofgranulation tissue on the time course ofglycosaminoglycan lossfrom implanted cartilage. Eachmouse received a double subcutaneous implant ofwrappedand non-wrappedfemoral head cartilage. Groups of eight to10 animals were killed at weekly intervals and theglycosaminoglycan content ofcartilage was assessed andexpressed by comparison withfrozen cartilage. *p=0 054,**p<001, ***p<OOI0 v non-wrapped cartilage.
induced granulomatous tissue resulted insignificant acceleration of cartilage destruction.Compared with frozen controls the GAG con-tent of wrapped implanted cartilage showed asignificant decrease from week 2 onwards,whereas this only barely achieved statisticalsignificance at four weeks for non-wrappedcartilage. There was a moderate increase in theGAG content at week 1 which was significantfor non-wrapped FHC (p<O-Ol). All sub-sequent experiments were stopped three weeksafter implantation.
EFFECTS OF ORCHIDECTOMY ON CARTILAGE
DEGRADATION IN THE PRESENCE AND ABSENCE
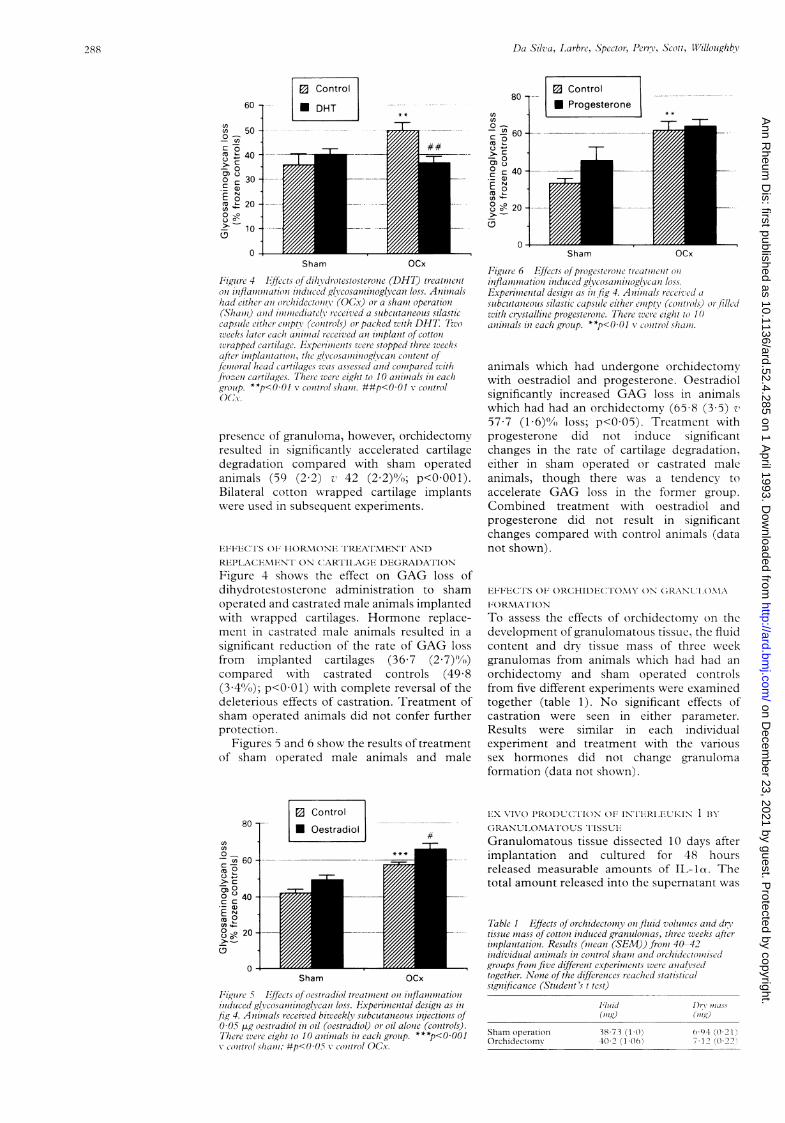
OF GRANULOMAIn the absence of cotton induced granuloma,implanted cartilage lost less than 20% of theinitial GAG content and this was not signi-ficantly changed by castration (fig 3). In the
60 -
CAm 50-o =
o+-X r-4->L.cu
- 0
co r,, 30 -. N
0
E 20
O O-
c0-
| Sham
eaI OCxI
Non-wrapped Wrapped
Figure 3 Effects of orchidectomy on glycosaminoglycan lossin the presence (wrapped; 11 animals) and absence (non-wrapped; 40-42 animals) ofgranulation tissue. Mice werecastrated (OCx) or sham operated (Sham) two weeks beforereceiving a double implant ofwrapped and non-wrappedfemoral head cartilage. The glycosaminoglycan content ofcartilage was assessed three weeks after implantation andcompared with frozen controls. Pooled resultsfrom fivedifferent experiments. ***p<0.001 v sham operated animals.
287
on Decem
ber 23, 2021 by guest. Protected by copyright.
http://ard.bmj.com
/A
nn Rheum
Dis: first published as 10.1136/ard.52.4.285 on 1 A
pril 1993. Dow
nloaded from
Da Silva, Larbre, Spector-, Perms, Scott, Willouighbv,
60 -
0)0 50 -0 -- 0
8o 30._a)
20 -
0E00
CD 10 -
0 -
| Control
DHT-T-
Sham OCx
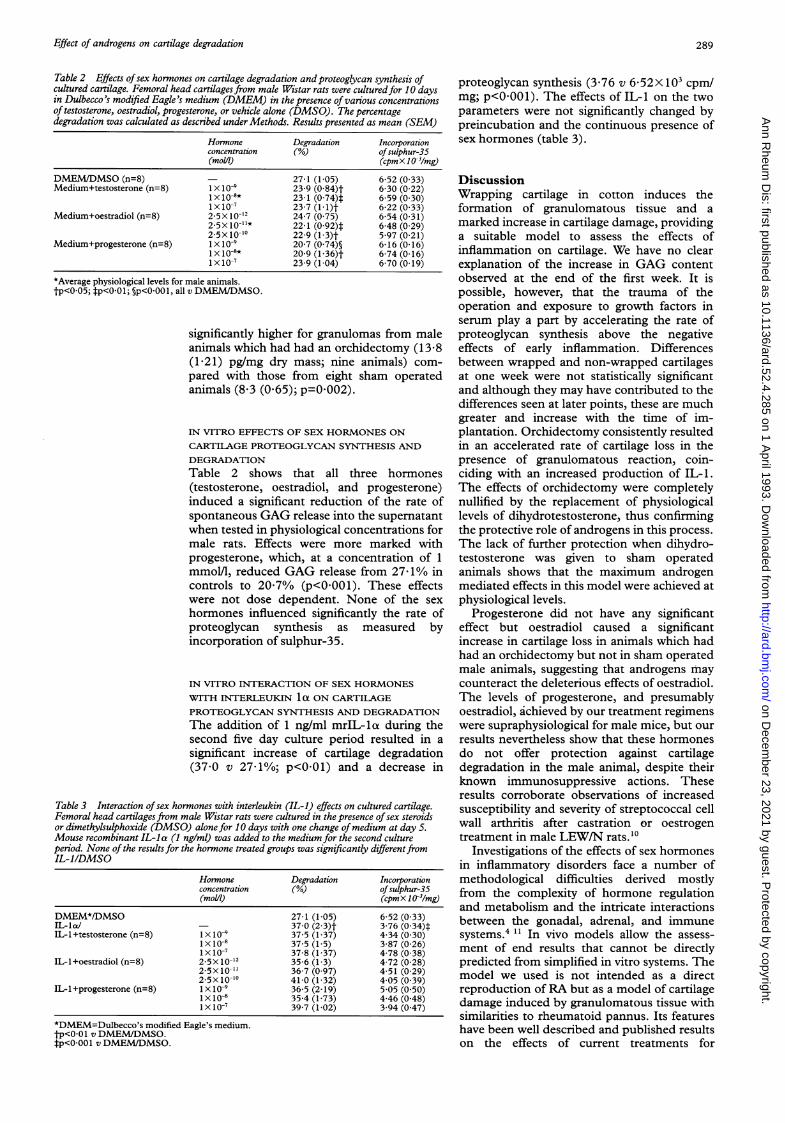
Figue 4 E/fccts of dih/idrotestosterone (DHT) treatmizeniton7 inflaminiation) indoiced glicosaniiniioglvcani loss. Animnalshad eithe/i an or-chiidectoniiv (OCx) or a shanii operatioln(Sham,i) and inuniiediatrel! received a subcutaneous silasticcapsilde eit/lecr emptv (conltrols) or packed with DHT. Twozweeks later cacli aninlial received anl iml'planit of cotton1wrapped cartilage. Fxperimients zere stopped three zveeksafter iniplantation, the glvcosamzinoglvcan content offenioral head cartilages zvas assessed anid coniipared zw1it/ifrozen cartilages. There were eight to 10 aninials in eachgr-oup. **p<OO1 v control shanii. ##p<001 controlOx
presence of granuloma, however, orchidectomyresulted in significantly accelerated cartilagedegradation compared with sham operatedanimals (59 (2 2) v! 42 (2 2)%; p<OOOl).Bilateral cotton wrapped cartilage implantswere used in subsequent experiments.
FEl'1FICTS OF; HORMTOI)NE TPTAEAIMENi ANT)RFlPLACEMINT ON (,ARTIIA(F DEGRADATION
Figure 4 shows the effect on GAG loss ofdihydrotestosterone administration to shamoperated and castrated male animals implantedwith wrapped cartilages. Hormone replace-ment in castrated male animals resulted in asignificant reduction of the rate of GAG lossfrom implanted cartilages (36 7 (2 7)'Vn)compared with castrated controls (49 8(3.4%0); p<OO1) with complete reversal of thedeleterious effects of castration. Treatment ofsham operated animals did not confer furtherprotection.
Figures 5 and 6 show the results of treatmentof sham operated male animals and male
80 --
-X 60 -C:-40_ .
5 O
z c0 0 40 -c c
Cl..
800o 20 -
| Control
* Oestradiol
4+
0 -Sham OCx
FPigutre 5 E'ffects of oestradiol treatment oni injlaniiationiinlduced glcvosaniino&glcan loss. IE,xpevimiental designi as i71fig 4. Animials received bizveekly subcutaneous inijectionzs of0-05 pg oestradiol in oil (oestradiol) or oil alonie (controls).7'There wuere eight to 10 animials in each grroulp. ***p<O0OO1v comtrol sha/ic; #p<0)Ox X conitrol OC,.v.
80 1-cn0)0-
' 0 -
- 00) 0° C 40 -._a)E o
°' - 20-
o -
E Control
* Progesterone
-F
Sham OCx
Figure 6 Effects (f pprogesterone treatmlienit (Oiinflaniniation itiduced gl6,cosanninogglcan loss.Experinienztal designi as in fig 4. Animials received casubcutanzeous silastic capszile either eniptv (controls,) (r fi/ledwzith crystalline progesterone. Thee were l'eg/it to 10aninlials in each group. **p<O0OI vclintrol slham7i.
animals which had undergone orchidectomywith oestradiol and progesterone. Oestradiolsignificantly increased GAG loss in animalswhich had had an orchidectomy (65-8 (3S5) v
57.7 (1 6)% loss; p<0 05). Treatment withprogesterone did not induce significantchanges in the rate of cartilage degradation,either in sham operated or castrated maleanimals, though there was a tendency toaccelerate GAG loss in the former group.Combined treatment with oestradiol andprogesterone did not result in significantchanges compared with control animals (datanot shown).
IFFECTS 01F ORCHIDECTFOMY ON GRANLIMA
FORMAFTIONTo assess the effects of orchidectomv on thedevelopment of granulomatous tissue, the fluidcontent and dry tissue mass of three weekgranulomas from animals which had had anorchidectomy and sham operated controlsfrom five different experiments were examinedtogether (table 1). No significant effects ofcastration were seen in either parameter.Results were similar in each individualexperiment and treatment with the varioussex hormones did not change granulomaformation (data not shown).
E'X VIVO PR(ODUCliION OF IN-I FRI,F[UKIN 1 B'Y
GCRANUI ONMATOUOS lISSU E
Granulomatous tissue dissected 10 days afterimplantation and cultured for 48 hoursreleased measurable amounts of IL-1a. Thetotal amount released into the supernatant was
Table I Effects (of orchidectomnv onjluind volum7tes anid dr\tissue niiass of cottoni induced granulomias, three weeks afteriniplanztationz. Results (nieani (SEM)) froni 40-42individual animials in control shami anid orchiidectouiisi.edgroups fronizfive differetnt expe^inients were analysedtogether. None of the differences reached statisticalsignificance (Student's t tcst)
Sham operationOrchidectom\
Fluid(mug)
38 7331(0)40 2 (1 (06)
1)15\ 7t1)0KIng)
0()94 (022i1 02(0 2)
288
4 * *
J.t
on Decem
ber 23, 2021 by guest. Protected by copyright.
http://ard.bmj.com
/A
nn Rheum
Dis: first published as 10.1136/ard.52.4.285 on 1 A
pril 1993. Dow
nloaded from
Effect of androgens on cartilage degradation
Table 2 Effects ofsex hormones on cartilage degradation and proteoglycan synthesis ofcultured cartilage. Femoral head cartilages from male Wistar rats were culturedfor 10 daysin Dulbecco's modified Eagle's medium (DMEM) in the presence of various concentrationsof testosterone, oestradiol, progesterone, or vehicle alone (DMSO). The percentagedegradation was calculated as described under Methods. Results presented as mean (SEM)
Hormone Degradation Incorporationconcentration (%) ofsulphur-35(moN) (cpmX 1(O3/mg)
DMEM/DMSO (n=8) - 27-1 (1-05) 6-52 (0-33)Medium+testosterone (n=8) 1x 10-9 23-9 (0 84)t 6-30 (0 22)
1x1O-8* 23-1 (0-74)t 6-59 (0-30)1X10-7 23-7 (1-1)t 6-22 (0-33)
Medium+oestradiol (n=8) 2 5X10-12 24-7 (0 75) 6-54 (0-31)2-5x10-11* 22-1 (0-92)t 6-48 (0 29)2.5x 10"- 22-9 (1-3)t 5 97 (0-21)
Medium+progesterone (n=8) 1 x 10-9 20-7 (0-74)§ 6-16 (0-16)1 x 10-8* 20-9 (1-36)t 6-74 (0-16)1X 10-7 23-9 (1-04) 6-70 (0 19)
*Average physiological levels for male animals.tp<0-05; tp<0-01; §p<0-001, all v DMEM/DMSO.
significantly higher for granulomas from maleanimals which had had an orchidectomy (13-8(1 21) pg/mg dry mass; nine animals) com-pared with those from eight sham operatedanimals (8&3 (0 65); p=0002).
IN VITRO EFFECTS OF SEX HORMONES ON
CARTILAGE PROTEOGLYCAN SYNTHESIS ANDDEGRADATIONTable 2 shows that all three hormones(testosterone, oestradiol, and progesterone)induced a significant reduction of the rate ofspontaneous GAG release into the supematantwhen tested in physiological concentrations formale rats. Effects were more marked withprogesterone, which, at a concentration of 1mmol/l, reduced GAG release from 27 /1% incontrols to 2077% (p<0 001). These effectswere not dose dependent. None of the sexhormones influenced significantly the rate ofproteoglycan synthesis as measured byincorporation of sulphur-35.
IN VITRO INTERACTION OF SEX HORMONESWITH INTERLEUKIN 1 Ot ON CARTILAGEPROTEOGLYCAN SYNTHESIS AND DEGRADATIONThe addition of 1 ng/ml mrIL-la during thesecond five day culture period resulted in asignificant increase of cartilage degradation(37 0 v 27/1%; p<001) and a decrease in
Table 3 Interaction ofsex hormones with interleukin (IL-1) effects on cultured cartilage.Femoral head cartilagesfrom male Wistar rats were cultured in the presence ofsex steroidsor dimethylsulphoxide (DMSO) alone for 10 days with one change ofmedium at day 5.Mouse recombinant IL-la (1 ng/ml) was added to the medium for the second cultureperiod. None of the results for the hormone treated groups was significantly differentfromIL-1/DMSO
Hormone Degradation Incorporationconcentration (%) ofsulphur-35(molJ) (cpmX 103/mg)
DMEM*/DMSO 271 (1-05) 6-52 (0-33)IL-Ia/ - 37-0 (2-3)t 3-76 (0-34)tIL-1+testosterone (n=8) Ix 10-9 37-5 (1-37) 4-34 (0-30)
1x10-8 37-5 (1-5) 3-87 (0 26)1X10-7 37-8 (1-37) 4-78 (0 38)
IL-1+oestradiol (n=8) 2-5x10-12 35-6 (1-3) 4-72 (0-28)2 5X10-1' 36-7 (0 97) 4-51 (0-29)2-5xI10" 41-0 (1-32) 4 05 (0-39)
ILl+progesterone (n=8) Ix 10-9 36-5 (2-19) 5-05 (0-50)1X10-8 35-4 (1-73) 4-46 (0-48)1X1O-7 39-7 (1-02) 3-94 (0-47)
*DMEM=Dulbecco's modified Eagle's medium.tp<0-01 v DMEM/DMSO.#p<0-00l v DMEM/DMSO.
proteoglycan synthesis (3-76 v 6&52X 103 cpmlmg; p<0-001). The effects of IL-1 on the twoparameters were not significantly changed bypreincubation and the continuous presence ofsex hormones (table 3).
DiscussionWrapping cartilage in cotton induces theformation of granulomatous tissue and amarked increase in cartilage damage, providinga suitable model to assess the effects ofinflammation on cartilage. We have no clearexplanation of the increase in GAG contentobserved at the end of the first week. It ispossible, however, that the trauma of theoperation and exposure to growth factors inserum play a part by accelerating the rate ofproteoglycan synthesis above the negativeeffects of early inflammation. Differencesbetween wrapped and non-wrapped cartilagesat one week were not statistically significantand although they may have contributed to thedifferences seen at later points, these are muchgreater and increase with the time of im-plantation. Orchidectomy consistently resultedin an accelerated rate of cartilage loss in thepresence of granulomatous reaction, coin-ciding with an increased production of IL-1.The effects of orchidectomy were completelynullified by the replacement of physiologicallevels of dihydrotestosterone, thus confirmingthe protective role of androgens in this process.The lack of further protection when dihydro-testosterone was given to sham operatedanimals shows that the maximum androgenmediated effects in this model were achieved atphysiological levels.
Progesterone did not have any significanteffect but oestradiol caused a significantincrease in cartilage loss in animals which hadhad an orchidectomy but not in sham operatedmale animals, suggesting that androgens maycounteract the deleterious effects of oestradiol.The levels of progesterone, and presumablyoestradiol, achieved by our treatment regimenswere supraphysiological for male mice, but ourresults nevertheless show that these hormonesdo not offer protection against cartilagedegradation in the male animal, despite theirknown immunosuppressive actions. Theseresults corroborate observations of increasedsusceptibility and severity of streptococcal cellwall arthritis after castration or oestrogentreatment in male LEW/N rats.'0
Investigations of the effects of sex hormonesin inflammatory disorders face a number ofmethodological difficulties derived mostlyfrom the complexity of hormone regulationand metabolism and the intricate interactionsbetween the gonadal, adrenal, and immunesystems.4 " In vivo models allow the assess-ment of end results that cannot be directlypredicted from simplified in vitro systems. Themodel we used is not intended as a directreproduction of RA but as a model of cartilagedamage induced by granulomatous tissue withsimilarities to rheumatoid pannus. Its featureshave been well described and published resultson the effects of current treatments for
289
on Decem
ber 23, 2021 by guest. Protected by copyright.
http://ard.bmj.com
/A
nn Rheum
Dis: first published as 10.1136/ard.52.4.285 on 1 A
pril 1993. Dow
nloaded from

Da Silva, Larbre, Spector, Pery, Scott, Willoughby
rheumatic diseases have shown a goodcorrelation with experience in human diseaseand other experimental models.'2 Unlike moreconventional models it allows a reproduciblemeasurement of cartilage destruction offeringappropriate conditions for the study of thisessential component of arthritis.The peripheral transformation of testos-
terone into oestradiol has been shown to bean important confounding factor on studiesaddressing immunological effects of sexhormones.4 Our results show that prolongedtestosterone treatment of experimental maleanimals, in a dose sufficient to attainphysiological levels, will result in extremelyhigh oestradiol levels due to peripheral trans-formation and is, therefore, inadequate tostudy the effects of androgens. The silasticcapsules of 5ux-dihydrotestosterone used hereallowed stable androgen levels to be obtainedin the low physiological range (compensatingfor the higher androgenic activity of dihydro-testosterone) without any detectable change inoestradiol levels.Unsuspected sex hormone action is also a
potential problem in in vitro studies. We haverepeatedly found high levels of oestradiol andprogesterone in different batches of fetal calfserum. Progesterone is also a common com-ponent of most available serum alternatives forculture purposes. Weak oestrogenic propertiesof phenol red, a common pH indicator inculture media, have been shown repeatedlyand we have therefore avoided these com-ponents in our culture conditions.The mechanisms of the protective actions of
androgens are still unclear. The lack ofinfluence of orchidectomy on non-wrappedcartilage suggests that the effects of sexhormones are mainly due to actions on theinflammatory tissue and not directly on thechondrocytes. To clarify this point further weinvestigated the effects of sex steroids and theirinteraction with IL-1 on cartilage metabolismin vitro. The effects seen on spontaneousproteoglycan loss in culture are common to thedifferent sex steroids studied and do not seemto be important in vivo. None of the sexhormones showed any significant interferencewith IL-1 induced effects on proteoglycansynthesis and degradation in vitro, reinforcingthe concept that direct actions on cartilage donot contribute significantly to the overalleffects seen in the in vivo model. The lack ofdose dependency of sex hormone effects oncartilage is probably related to the number andkinetics of the respective receptors in thistissue. To our knowledge only oestradiolreceptors have been described in articularchondrocytes.'3 14 These receptors were foundin small amounts but showed high affinity,which may justify saturation at physiologicalconcentrations. It should also be noted that thelower concentrations used in our in vitrostudies correspond approximately to physio-logical concentrations of free sex steroids,given that these hormones are bound to carrierproteins in excess of 90%.
Orchidectomy did not affect parameters ofgranulomatous tissue development, in agree-
ment with previous findings with this modelshowing a lack of positive association betweenthe mechanisms responsible for generatingacute exudative inflammation and thosecausing cartilage destruction.7 A similardissociation has been proposed for rheumatoidarthritis.'5 Our results show that sex steroidsaffect the erosive ability of granulomatoustissue, probably by influencing the productionof mediators, as suggested by the increasedrelease of IL-1 associated with orchidectomy.Although we cannot provide firm evidence thatIL-1 is responsible for the acceleration ofcartilage loss induced by castration, there isoverwhelming evidence for a pivotal role of thiscytokine in cartilage degradation in human andexperimental disease and our own results invitro show that mouse IL- lot is capable ofinducing increased loss and decreasedsynthesis of proteoglycan from rat FHCs. It isnot clear how androgens should bring aboutthis effect. In our experience, orchidectomydoes not significantly influence the ex vivorelease of IL-1 by resident mouse peritonealmacrophages (data not shown). Macrophagesin an inflammatory environment, however,may show a different response and the complexinteractions that surround these cells ininflammatory tissue provide a large numberof pathways for indirect modulation-forexample, through T cells and lymphokineproduction. 6 The possibility of directandrogen actions on these cells is supported bythe description of androgen receptors inmacrophage-like cells in normal and rheuma-toid synovia.5 It is unlikely that the effects ofandrogens in this model are dependent on anyinteraction with the hypothalamus-pituitary-adrenal axis, as treatment with dihydro-testosterone results in reduced glucocorticoidresponse to the inflammatory process. ' Thissuggests a direct anti-inflammatory role forandrogens, which is also supported by previousfindings of reduced inflammation and cartilageerosion after intralesional administration oftestosterone in models of air pouch and antigeninduced arthritis in mice. 8
In summary, our results show that andro-gens have a protective role against inflam-mation induced cartilage degradation in maleanimals, probably mediated by interferencewith the immune and inflammatory process.These results offer additional support for apotential pathogenic role of androgens inhuman disease and suggest that in addition toameliorating disease activity, androgens maycontribute to reduce joint destruction anddisability in arthritis.
Dr Da Silva is a visiting physician from Coimbra, Portugal,under a grant from INVOTAM (Lisbon). Dr Larbre issupported by a grant from INSERM, France.
1 Gordon C, Beastall G H, Thomson J A, Sturrock R D.Androgenic status and sexual function in males withrheumatoid arthritis and ankylosing spondylitis. Q 7 Med1986; 60: 67 1-9.
2 Spector T D, Ollier W, Perry L A, et al. Free and serumtestosterone levels in 276 males: a comparative study ofrheumatoid arthritis, ankylosing spondylitis and healthycontrols. Clin Rheumatol 1989; 8: 37-41.
3 Cutolo M, Balleari E, Giusti M, Intra E, Accardo S.Androgen replacement therapy in male patients withrheumatoid arthritis. Arthritis Rheum 1991; 34: 1-5.
290
on Decem
ber 23, 2021 by guest. Protected by copyright.
http://ard.bmj.com
/A
nn Rheum
Dis: first published as 10.1136/ard.52.4.285 on 1 A
pril 1993. Dow
nloaded from

Effect of androgens on cartilage degradation
4 Ansar Ahmed S, Penhale W J, Talal N. Sex hormones,immune responses, and autoimmune diseases. Mech-anisms of sex hormone action. Am Pathol 1985; 121:531-51.
5 Cutolo M, Accardo S. Villaggio B, et al. Evidence for thepresence of androgen receptors in the synovial tissue ofrheumatoid arthritis patients and healthy controls.ArthrinisRheum. 1992; 35: 1007-15.
6 Franchimont P, Bassleer C. Effects of hormones andlocal growth factors on articular chondrocyte metabolism.JRheumatol 1991; 18 (suppl 27): 68-70.
7 De Brito F B, Moore A R, Holmes M J G, Willoughby D A.Cartilage damage by a granulomatous reaction in amurine species. BrJ7ExpPathol 1987; 68: 675-86.
8 Wathen N C, Perry L A, Rubenstein E, Chart T. Arelationship between sex hormone binding globulin anddehydroepiandrosterone sulphate in normally men-struating females. Gynaecol Endocninol 1987; 1: 47-50.
9 Farndale R W, Buttle D J, Barrett A J. Improvedquantitation and discrimination of sulphated glycos-aminoglycans by use of dimethylmethylene blue. BiochimBiophysActa 1986; 883: 173-7.
10 Allen J B, Blatter D, Calandra G B, Wilder R L. Sexhormonal effects on the severity of streptococcal cell wall-induced polyarthritis in the rat. Arthnitis Rheum 1983; 26:560-3.
11 Lesniewska B, Miskowiak B, Nowak M, Malendowicz L K.Sex differences in adrenocortical structure and function.
XXVII. The effect of ether stress on ACTH and cortico-sterone in intact, gonadectomized, and testosterone-or estradiol-replaced rats. Res Exp Med 1990; 190:95-103.
12 Bottomley K M K, Griffiths R J, Rising T J, Steward A. Amodified air pouch model for evaluating the effects ofcompounds on granuloma induced cartilage degradation.BrJPharmacol 1988; 93: 627-35.
13 Rosner I A, Manni A, Malemud C J, Boja B, MoskowitzR W. Estradiol receptors in articular chondrocytes.Biochem Biophys Res Commun 1982; 106: 1378-82.
14 Young P C M, Stack M T. Estrogen and glucocorticoidreceptors in adult canine articular cartilage. ArthritisRheum 1982; 25: 568-73.
15 Krane S M, Simon L S. Rheumatoid arthritis: clinicalfeatures and pathogenetic mechanisms. Med Clin NorthAm 1986; 70: 263-77.
16 Araneo B A, Dowell T, Diegel M, Daynes R A.Dihydrotestosterone exerts a depressive influence on theproduction of interleukin-4 (IL4), IL-5 and y-interferon,but not IL-2 by activated murine T cells. Blood 1991; 78:688-99.
17 Da Silva J A P, Peers S H, Perretti M, Willoughby D A.Sex steroids affect glucocorticoid response to chronicinflammation and to interleukin 1. JT Endocrinol. Inpress.
18 Steward A, Bayley D L. Effects of androgens in models ofrheumatoid arthritis. Agents Actions 1992; 35: 268-72.
291
on Decem
ber 23, 2021 by guest. Protected by copyright.
http://ard.bmj.com
/A
nn Rheum
Dis: first published as 10.1136/ard.52.4.285 on 1 A
pril 1993. Dow
nloaded from












![Cartilage - facultymembers.sbu.ac.irfacultymembers.sbu.ac.ir/rajabi/ppt toPDF/Cartilage [Compatibility Mode].pdfFibrocartilage • Fibrous Cartilage • is a form of connective tissue](https://img.pdfslide.net/doc/110x75/6012989a4318862a0e5813ae/cartilage-topdfcartilage-compatibility-modepdf-fibrocartilage-a-fibrous.jpg)
















