Embed Size (px)
Citation preview

Protein Metabolism in Rooted Runner-Bean LeavesAuthor(s): A. C. ChibnallSource: New Phytologist, Vol. 53, No. 1 (Jan., 1954), pp. 31-37Published by: Wiley on behalf of the New Phytologist TrustStable URL: http://www.jstor.org/stable/2430788 .
Accessed: 16/06/2014 16:18
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Wiley and New Phytologist Trust are collaborating with JSTOR to digitize, preserve and extend access to NewPhytologist.
http://www.jstor.org
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions

I 3I 1
PROTEIN METABOLISM IN ROOTED RUNNER-BEAN LEAVES
BY A. C. CHIBNALL Department of Biochemistry, University of Cambridge
(Received i9 March I953)
INTRODUCTION
In my Silliman Lectures, delivered at Yale University (Chibnall, I939), I discussed in some detail the assumption, first given concrete expression by Gregory & Sen (I937), that a protein cycle is operative in leaves. The evidence available at that time was purely cir- cumstantial and was based on two quite different types of investigation. In the one the data were representative of the leaf while still attached to the plant (i.e. conditions were normal); in the other the data illustrate the changes taking place subsequent to detach- ment of the leaf from the plant (i.e. conditions were abnormal).
(i) Gregory, Richards and their collaborators, in a series of experiments with pure- strain barley plants supplied with the nutrient elements at varying levels both singly and in combination, systematically studied the changes in each successive leaf throughout the growth cycle. Respiration measurements, and data for carbohydrate and nitrogen analyses, obtained at successive definite stages during development, were available, showing simultaneously the changes of respiration rate and the drifts of carbohydrate and nitrogen metabolism. Consideration of the data led Gregory & Sen to postulate that a protein cycle was continuously at work in leaves, and that the paths of protein synthesis and of protein breakdown may be separate.
(2) The observation of many workers had shown that detachment of a leaf, with or without its petiole, from the parent plant was always followed by a rapid breakdown of protein in the blade. The rate at which the breakdown occurred varied from species to species, from leaf to leaf with age for the same species, and also with the carbohydrate level. One possible explanation of such phenomena was to assume that a protein cycle is operative in leaves, and that the rate at which this works varies from species to species and from leaf to leaf, and that on detachment from the plant the mechanism of protein synthesis is somewhat impaired, so that protein breakdown predominates.
It was with this second group of experiments that I had been personally concerned, and I concluded my lecture on 'The Regulation of Protein Metabolism in Leaves' with the following statement: 'All the points mentioned in the above discussion could be enlarged upon but at the present time I think without profit, for the overriding fact remains that we do not yet understand the reason why, when certain leaves are detached from the plant, protein decomposition can be detected within a few hours. If this is indeed due to inter- ference with the mechanism of protein synthesis then one is tempted to suggest that some influence of the root system, possibly hormonic, is responsible for the regulation of the protein level in leaves. On this important question, however, I would prefer to reserve judgement until I have had the opportunity to investigate detached leaves which have been induced to form adventitious root systems.'
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions

32 A. C. CHIBNALL
Preliminary studies on these lines were started in I939 (Chakravarti, I939), but the work was interrupted by the war and could not be resumed and completed until I945-6. The results now reported show that the development of adventitious roots on the petiole of a detached leaf will arrest the rapid breakdown of protein in the blade, and that under appropriate conditions this solitary leaf can be kept alive and in a healthy state for many weeks. The possible bearing of these observations on the question of protein regulation in leaves is discussed.
EXPERIMENTAL
Detached leaves and root development Mature leaves of the runner bean (Phaesolus vulgaris) were considered the most suitable for the present investigation, because their behaviour when detached from the plant and kept with their petioles in water was already well known (cf. Chibnall, I939). The later work of Chibnall & Sahai (I93I) and Michael (I935) with other plants, however, had indicated that under these conditions an unsuspectedly large amount of nitrogenous pro- ducts might be translocation from the blade to the petiole, and to determine the magnitude of this it was necessary to pay more attention to controls than hitherto. The earlier work had shown that while detached leaves with their petioles in water remained healthy, there was a slow increase in the fresh weight of the blade due to the uptake of water, together with a concomitant decrease in the dry weight due to respiration. These points are illustrated by the data given later in Table 2. For purposes of comparison between the control and experimental leaf material, therefore, it was necessary to use some basis other than percentage fresh or dry weight. In the following experiment, typical of many, the leaves chosen were about 8 weeks old and had paired opposite pinnae of identical develop- ment as judged by external appearance. Comparative analytical data for collected samples
Table i. Six selected leaves of the runner bean
Opposite paired pinnae
Right side Left side Difference
Fresh wt. in g. 8-22 7-69 -0 53
Dry wt. in g. I-29 I-I0 -O-I9 Total N in mg. 56.56 54-80 -I76 Protein-N in mg. 46.I2 44.I2 -2-0
Non-protein-N in mg. I0-44 io068 +0-24
of opposite pinnae from six leaves are given in Table i. They illustrate the variation to be expected in both fresh and dry weight of similar samples of blade material, suggesting that in the experiments detailed below, the difference in the protein level of the control and treated blades could be most effectively determined by expressing the protein-N of both samples as a percentage of the total N of the control.
Lanolin paste containing /-indolylacetic acid was used as the agent to induce rooting in preliminary experiments with detached tomato leaves. While these were in progress a paper by Stuart (I938-9) came to hand in which he showed that cuttings from seedlings of the kidney bean could be induced to form adventitious roots very readily by simple treatment of the hypocotyl for 4 hr. with O.oi % fi-indolylacetic acid. By the use of his procedure it was found that detached mature leaves of the runner bean could also be
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions

Protein metabolism in rooted runner-bean leaves 33 induced to form rootlets at the cut end of the petiole, and having defined the necessary conditions a comprehensive investigation was planned.
Twelve leaves about 8 weeks old, all with opposite paired pinnae of equal development, were detached from the plant so as to leave the main petiole about 3 in. long. The apical leaflets (subsidiary petiole as well as blade) were then removed and the resulting leaves sorted into three groups of four (referred to henceforth as groups A, B and C) for the first three experiments detailed below. In each experiment the first step was to detach one of the paired opposite pinnae from each of the four leaves. The detached blades were collected and used for the group-control analysis. The attenuated leaves, now consisting of no more than the main petiole and the remaining paired pinna, were then submitted to the appropriate treatment.
Group A. The four leaves were placed in beakers containing ordinary tap water so that the lower inch of the main petiole was immersed. The tap water was changed daily, and a very thin slice from the end of each petiole was also removed daily so as to ensure that bacterial contamination did not interfere with the free supply of water through the petiole to the blade. The leaves remained thus for 6 days at room temperature in a room exposed during the day to strong diffused daylight. When removed for analysis at the end of this period the pinnae showed the first signs of incipient yellowing, but were still fully turgid. The conditions of this experiment approximated closely to those of the earlier investiga- tions referred to above.
Group B. The four leaves were planted upright with the end of the petiole buried to a depth of about i in. in moist sand contained in a dish. This dish could be completely covered with a large bell jar so as to maintain the enclosed air in a state of high humidity. The apparatus was placed in a greenhouse at a temperature of about 250 C. for 6 days and the sand was kept moist with tap water. When the pinnae were detached for analysis at the end of this period they were fully turgid and showed no evidence of incipient yellowing. The ends of the petioles showed no signs of swelling.
Group C. The four leaves were first stood for 4 hr. with the lower end of the petiole immersed in a solution of o oi % indolylacetic acid. The petioles were then washed with water and planted in moist sand for treatment as in group B. After 2 days the lower part of each petiole was swollen and a callus had formed over the cut end; by the fifth day these swellings had ruptured in places to permit the emergence of rootlets. Rootlet emergence was also visible at a few other places slightly higher up the petiole. At the end of the sixth day the leaves were removed, washed, and then stood with the petioles in tap water under the same conditions as the leaves in group A. To avoid serious bacterial in- fection the water was changed every second day. During a period of I7 days' water culture the rootlet growth was slow but continuous, and the blades remained to all outward appearances quite healthy. When detached for analysis at the end of the seventeenth day the blades showed the first signs of incipient yellowing and in this respect were com- parable with the leaves of group A at the end of their experimental period.
Group D. At the time when the above-mentioned three experiments were started and when leaves of similar age from the same plants were still available, a fourth experiment was undertaken to find out how long the rooted leaves might survive. For this purpose two leaves were selected in which the paired opposite pinnae of the one were not only comparable in development with each other, but also with those on the second leaf. The paired opposite pinnae on one leaf were then analysed at once as control. The second leaf
New Phytol. 53, I 3
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions

34 A. C. CHIBNALL
was taken, its apical leaflet removed, and the remainder-this time with the main petiole bearing both the opposite paired pinnae-treated in the same way as the leaves in group C. The tap water was changed every other day. Most of the rootlets were nearly an inch long at the end of 2 weeks' water culture, and 2 in. long at the end of 4 weeks. When removed for analysis at the end of the sixth week the blades were still turgid, but were showing the first signs of incipient yellowing; in this respect they were comparable with the blades of groups A and C at the end of their experimental periods.
METHODS OF ANALYSIS
Dry wezght. The fresh leaves were dried rapidly in a steam oven through which a stream of air was drawn. The material was then ground to a fine powder and a sample kept at I02? C. for 24 hr. for determination of dry weight.
Total nitrogen. Thirty mg. of the air-dried material were placed in a micro-Kjeldahl digestion flask, followed by 50 mg. of reduced iron (Kahlbaum) and I ml. of 5000 sul- phuric acid. After standing for io min. the mixture was boiled for 5 mm. to complete the reduction of any nitrate present. On cooling, 0o2 g. of anhydrous potassium sulphate, a small crystal of sodium selenate and 2-5 ml. of sulphuric acid were added. Digestion was allowed to proceed overnight.
Protein-N and non-protein-N. Fifty mg. of the air-dried material were ground in a small mortar with 5 ml. 5 0 N-free aqueous trichloroacetic acid, which was added in small portions over i hr. The mixture was allowed to stand 24 hr. and then filtered. The residue was washed twice with 2 ml. lots of the trichloroacetic acid solution and then digested by the Kjeldahl procedure to give a measure of the protein-N. The filtrate and washings representing the non-protein-N were collected, reduced in volume and submitted to the modified Kjeldahl procedure to allow for nitrate present.
DISCUSSION
The analytical data obtained from the four groups of experimental material are given in Table 2. On the assumption already referred to, that comparison between the nitrogen fractions of the control and treated samples of pinnae is best achieved in terms of the total N of the control, the relevant data are presented in Table 3. Many interesting obser- vations emerge from these studies, of which the following are perhaps the most pertinent to the present inquiry.
When detached leaves are cultured in water until the onset of chloroplast disintegra- tion, the breakdown of protein in the blades is more extensive than earlier work (cf. Chibnall, 1939) had suggested. In the present instance (group A) the loss represents 29 7% of the blade N (Table 3) and 36 2% of the total protein-N. If the data had been interpreted in terms of total N in the respective control and treated samples, which was the procedure adopted in the above-cited earlier work, these losses would have been shown as i9'8 and 23 % respectively. Extensive translocation into the petiole of the pro- ducts of proteolysis has occurred.
When the detached leaves were kept in a humid atmosphere, as in group B, so that transpiration was depressed, the breakdown of protein was much less extensive in spite of the higher temperature prevailing. At the end of the experiment the leaves showed no signs of yellowing and would undoubtedly have remained healthy for a much longer
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions
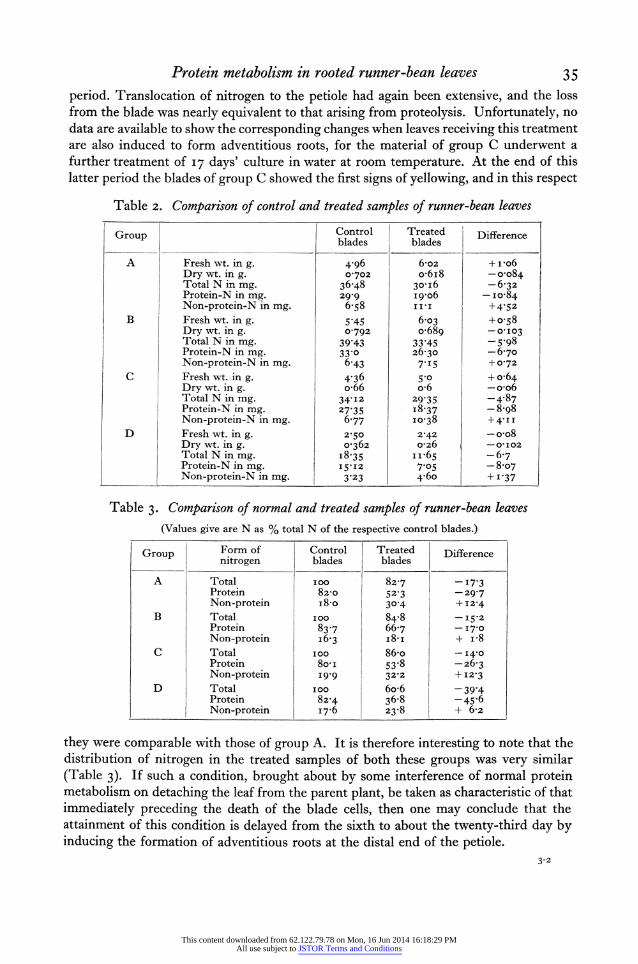
Protein metabolism in rooted runner-bean leaves 35 period. Translocation of nitrogen to the petiole had again been extensive, and the loss from the blade was nearly equivalent to that arising from proteolysis. Unfortunately, no data are available to show the corresponding changes when leaves receiving this treatment are also induced to form adventitious roots, for the material of group C underwent a further treatment of I7 days' culture in water at room temperature. At the end of this latter period the blades of group C showed the first signs of yellowing, and in this respect
Table 2. Comparison of control and treated samples of runner-bean leaves
Group Control Treated Difference Group ~~~~~~~blades blades Dfeec
A I Fresh wt. in g. 4 96 6-o2 + vo6 Dry wt. in g. 0-702 o-6i8 -o-o84 Total N in mg. 36-48 301i6 -6-32 Protein-N in mg. 2999 i9+06 - Io084 Non-protein-N in mg. 6- 8 II*I +4-52
B Fresh wt. in g. 5 45 6o03 +?-58 Dry wt. in g. 0?792 o0689 -0-I03 Total N in mg. 39 43 33-45 - 5.98 Protein-N in mg. 33?0 26-30 -670 ! Non-protein-N in mg. 6-43 7 I5 + 0-72
C Fresh wt. in g. 4-36 5-0 + o-64 Dry wt. in g. o66 o6 -oo6 Total N in mg. 34.I2 29 35 -4-87 Protein-N in mg. 27835 -37 -8-98 Non-protein-N in mg. 6&77 10-38 +4'II
D Fresh wt. in g. 2-50 2-42 -o-o8 Dry wt. in g. o-362 o-26 -O'I02 Total N in mg. I8.35 II.65 -6-7 Protein-N in mg. 15.I2 7o05 -8-o7
.______ tNon-protein-N in mg. 3 23 46o + |_'37
Table 3. Comparison of normal and treated samples of runner-bean leaves (Values give are N as % total N of the respective control blades.)
Group Form of Control Treated Difference nitrogen blades blades
A Total I00 82-7 -I7'3 Protein 82o0 52-3 -29,7 Non-protein i8 o 30?4 +I2-4
B Total I 00 84.8 -15* 2 Protein 83-7 66.7 -17-0 Non-protein I6-3 i8-I + I*8
C Total ioo 86-o - 14 0 Protein 8o-I 53 8 -z6-3 Non-protein 19,9 32 2 + I2-3
D Total I00o 6o-6 -39 4 Protein 82.4 36-8 -45 6 Non-protein I7-6 23-8 + 6-2
they were comparable with those of group A. It is therefore interesting to note that the distribution of nitrogen in the treated samples of both these groups was very similar (Table 3). If such a condition, brought about by some interference of normal protein metabolism on detaching the leaf from the parent plant, be taken as characteristic of that immediately preceding the death of the blade cells, then one may conclude that the attainment of this condition is delayed from the sixth to about the twenty-third day by inducing the formation of adventitious roots at the distal end of the petiole.
3-2
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions

36 A. C. CHIBNALL
In the fourth experiment (group D) the leaf material differed from those used in the other three in that both the opposite pinnae remained attached to the petiole. In this case the symptoms of approaching death did not appear until about 40 days after detach- ment from the parent plant. Meanwhile, movement of nitrogen into the petiole for root growth had gone on continuously, and the protein level in the blade was reduced to an astonishingly low value; the loss represents 45-6 00 of its initial total N and 55 3 00 of its initial protein-N. The longer life attained by these particular detached leaves must be ascribed to the presence of the second opposite pinna and the resulting increase in photo- synthetic capacity. This addition to the available carbohydrate must, in its turn, be re- sponsible for the conditions which have permitted the large drain of nitrogen from the blades without lethal effects. Data for carbohydrate analysis were unfortunately not obtained before the work had to be abandoned at the outbreak of war, but the researches of Stuart (I938-9) and of Mitchell & Stuart (I938-9) have shown that in kidney-bean seedling cuttings treated with indolylacetic acid there was a strong drift of carbohydrate as well as of nitrogen from the cotyledons and the young leaflets towards the hypocotyl, which responded by profuse root production. As the experimental material of group D, with two pinnae, differed from that of both groups A and C, it is not surprising that there is a difference in the data given in Table 3. At the time they were submitted to analysis, nevertheless, all three of these treated samples had one characteristic in common, in that they showed signs of incipient chloroplast degeneration. In Table 4 the nitrogen distribu- tion in the treated blades is presented in terms of total N of the respective samples, and not in terms of the controls as in Table 3. All three of the above-mentioned groups exhibit a similar ratio for protein-N to non-protein-N, an indication that degenerative changes may be associated with a mass action regulation.
Table 4. Comparison of variously treated samples of runner-bean leaves
(Values given are N as % total N of the respective treated blades.)
Group Form of N Treated blades
A Protein 63-2 Non-protein 36.8
C Protein 62-8 Non-protein 37-2
D Protein 6o-5 ____________ Non-protein 39 5
To summarize the evidence that can be drawn from these four experiments it would appear that on detachment of the leaf from the parent plant there is initiated almost at once a strong directional shift of nitrogen (and almost certainly of carbohydrate) from the blades towards the distal end of the petiole (cf. Michael, I935). This is presumably the normal response of a plant organ to wounding, just as it is known to be in animal organs. If conditions do not favour the healing of the wound and the initiation of root growth (group A) the drain of nitrogen from the blades, accompanied by breakdown of protein, carries on at a high rate and toxic symptoms become apparent within a few days, irre- spective of the carbohydrate supply (cf. Chibnall, I939). On the other hand, should conditions, brought about for instance by treatment with hormones, favour rooting, the rate of outflow of nitrogen to the petiole and also the rate of protein breakdown in the
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions

Protein metabolism in rooted runner-bean leaves 37 blade is much retarded, and the onset of toxic symptoms may be delayed many days if not weeks. The development of adventitious roots on the petiole of a detached leaf therefore does indeed influence the rate of protein breakdown in the blade.
This observation provides us with an answer to one of the queries underlying the present investigation. But in itself it provides no direct evidence bearing on the larger question of whether a protein cycle is operative in these detached, and hence by inference in the normal, leaf blades. A cycle implies continuous synthesis as well as breakdown, and it was to obtain evidence that protein synthesis might be going on concomitantly with the observed rapid protein breakdown in the detached leaves that the studies recorded in the following paper were carried out.
SUMMARY
i. When a mature leaf of the runner bean is detached from the plant and kept with the cut end of the petiole in water or mounted in moist sand there is a rapid breakdown of protein in the blade, accompanied by an outflow of non-protein nitrogen into the petiole. Chloroplast degeneration occurs within a few days.
2. Treatment with indolylacetic acid brings about the development of adventitious roots on the petiole; under these conditions the rate of protein breakdown in the blade is much reduced and the rooted leaf will survive for many weeks.
3. The bearing that these observations have on the concept of a protein cycle in leaves is discussed.
REFERENCES
CHAKRAVARTI, M. G. (1939). Investigations on the protein metabolism in plants. Ph.D. Thesis, London University.
CHIBNALL, A. C. (1939). Protein Metabolism in the Plant. New Haven: Yale University Press. CHIBNALL, A. C. & SAHAI, P. N. (I93 I). Observations on the fat metabolism of leaves. i. Detached and
starved mature leaves of brussels sprout. Ann. Bot., Lond., 45, 489. GREGORY, F. G. & SEN, P. K. (1937). Physiological studies in plant nutrition. VI. The relation of respira-
tion rate to the carbohydrate and nitrogen metabolism of the barley plant as determined by nitrogen and potassium deficiency. Ann. Bot., Lond., I, 521.
MICHAEL, G. (I935). tber die Beziehungen Zwischen Chlorophyll- und Eiweissabbau im vergilbenden Laubblatt von Tropaeolum. Z. Bot. 29, 385.
MITCHELL, J. W. & STUART, N. W. (1938-9). Growth and metabolism of bean cuttings subsequent to rooting with indoleacetic acid. Bot. Gaz. zOO, 627.
STUART, N. W. (1938-9). Nitrogen and carbohydrate metabolism of kidney-bean cuttings as affected by treatment with indoleacetic acid. Bot. Gaz. 100, 298.
This content downloaded from 62.122.79.78 on Mon, 16 Jun 2014 16:18:29 PMAll use subject to JSTOR Terms and Conditions