Embed Size (px)
Citation preview

(CANCER RESEARCH 48, 913-919. February 15, 1988)
Purification and Characterization of a Bindable Form of Mitochondrial BoundHexokinase from the Highly Glycolytic AS-30D Rat Hepatoma Cell Line1
Richard A. Nakashima, Marco G. Paggi,2 Laura J. Scott, and Peter L. Pedersen3
Laboratory for Molecular and Cellular Bioenergelics. Department of Biological Chemistry, The Johns Hopkins University, School of Medicine, Baltimore, Maryland 21205
ABSTRACT
Recent studies from this laboratory have demonstrated that a form ofhexokinase characteristic of rapidly growing, highly glycolytic tumorcells is bound to an outer mitochondria! membrane receptor complexcontaining a M, 35,000 pore protein (D. M. Parry and P. L. Pedersen,J. Biol. Chem., 258: 10904-10912, 1983; R. A. Nakashima, et a/.,Biochemistry, 25: 1015-1021, 1986). In new studies reported here thespecificity of this receptor complex for binding hexokinase is defined,and a purification scheme is described which leads to a homogeneous andbindable form of the tumor hexokinase.
In the AS-30D hepatoma, hexokinase activity is elevated more than100-fold relative to liver tissue. The relative increase in hexokinaseactivity is 8 times greater than that of any other glycolytic enzyme.Hexokinase is the only glycolytic enzyme of AS-30D cells to exhibit amitochondrial/cytoplasmic specific activity ratio greater than 1, showinga 3.5-fold elevation in the mitochondria!fraction.
Purification of hexokinase is accomplished by preferential solubiliza-tion of the mitochondria! bound enzyme with glucose-6-phosphate, followed by high-performanceliquid chromatographyon gel permeationandaniónexchange columns. The final fraction has a specific activity of 144units per mg of protein, with a A,,,for glucose of 0.13 miviand for ATPof 1.4 mM.The purified tumor enzyme migrates as a single species uponsodium dodecyl sulfate: polyacrylamide gel electrophoresis with an apparent molecular weight of 98,000. Significantly, the purified tumorenzyme retains its activity for mitochondria!binding.
Additional results derived from Chromatographie,polyclonal antibody,and amino acid analysis studies indicate that the predominant rat hepatoma hexokinase species is related most closely to isozymic form(s) ofthe enzyme commonly referredto as type II, and least related to the livertype IV isozyme (glucokinase).
INTRODUCTION
It has been widely observed that transformed cells exhibitincreased rates of glucose utilization compared to normal cellsfrom the same tissue of origin ( 1-8). Elevated glucose catabo-lism is important for the production of both energy (7) andrequired anabolic precursors in rapidly growing tumors. Previous studies from our laboratory (4-6) and others (9-11) haveemphasized the role of the initial enzyme of glycolysis, hexokinase (EC 2.7.1.1), in maintaining high rates of glucose utilization in rapidly growing cancer cells. In comparison withnormal cells from the same tissue of origin there is a largeincrease in hexokinase activity in highly glycolytic tumor celllines (2-7,9,10). This is accompanied by a change in subcellulardistribution of the enzyme from a predominantly cytosolic to amitochondria-bound form (4-6, 9, 11-14). Mitochondria-bound hexokinase has been reported to be less sensitive tofeedback inhibition by glucose-6-phosphate, a potent regulator
Received 6/17/87; revised 10/22/87; accepted 11/18/87.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by NIH Grant CA 32742 to P. L. P. R. A. N. andM. G. P. were supported, respectively, by a fellowship from the Leukemia Societyof America and by CNR Grants 85.02159.44 and AIRC/86 from the AssociazioneItaliana per la Ricerca sul Cancro.
2Permanent address: Istituto Regina Elena per lo Studio e la Cura dei Tumori,V.le Regina Elena 291,00161 Rome, Italy.
3To whom requests for reprints should be addressed.
of hexokinase activity in normal tissues (4, 9, 11). The boundenzyme may also obtain preferential access to mitochondria!generated ATP (13, 15). A direct correlation has been observedbetween mitochondria! bound hexokinase activity, tumor glycolysis, and tumor growth rate in cancer cell lines from a widevariety of tissues and species (3, 5, 6, 10).
A protein which promotes binding of rat brain hexokinasehas been partially purified from the outer membrane of rat livermitochondria (16). Several recent reports have suggested thatthe outer mitochondria! membrane pore protein, also known asVDAC or mitochondrial porin, might be identical to the hexokinase binding protein of rat liver (17, 18). Significantly, wehave recently demonstrated directly that the pore protein formsat least one part of a hexokinase receptor complex on the outermembrane of tumor mitochondria (19).
Despite the apparent importance of mitochondrial boundhexokinase in tumor cell growth and metabolism very little isknown about the molecular characteristics of the tumor enzyme.In the present study, we report the first purification to apparenthomogeneity of mitochondrial bound hexokinase from a transformed cell line, the highly malignant AS-30D rat hepatoma.The purification technique, involving rapid separation of solu-bilized mitochondrial hexokinase by high-performance liquidchromatography, yields an apparently homogeneous enzyme ofhigh specific activity which retains its ability to rebind to theouter mitochondrial membrane. We have also examined thespecificity of the binding interaction between hexokinase andthe outer mitochondrial membrane relative to that of otherglycolytic enzymes.
MATERIALS AND METHODS
Chemicals. Acrylamide, bisacrylamide, sodium dodecyl sulfate, molecular weight standard proteins, Coomassie Brilliant Blue R-2SO,DEAE-cellulose (Cellex D), and 2-mercaptoethanol were obtained fromBio-Rad. Centricon 10 microconcentrators were purchased from Ami-con. Dithiothreitol, NADI' . and glucose-6-phosphate dehydrogenase(Grade I) were from Boehringer Mannheim. Oligomycin, phenylmeth-ylsulfonyl fluoride, bovine serum albumin (Fraction V, fatty acid free),NADH, and P',P'-di(adenosine-5')pentaphosphate were from Sigma
Chemical Co. Phenylisothiocyanate, constant boiling 6 N HC1 andamino acid standards were from Pierce. Eluents for the analysis ofamino acid composition were obtained from the Waters Chromatography Division of the Millipore Corporation (Pico-Tag Eluents A andB). The detergents used were Lubrol WX from Grand Island BiologicalCo. and Triton X-100 from Research Products International Corp. Allother chemicals and substrates used were of analytical grade or betterand were obtained from standard chemical supply companies includingSigma Chemical Co., J. T. Baker Chemical Co., Calbiochem-BehringCorp., K and K Laboratories, Inc., Eastman Kodak Co., and Mallinck-rodt Chemical Works.
Animals and Tumor Cells. The AS-30D rat hepatoma cell line wasobtained from Dr. Baltazar Reynafarje of the Department of BiologicalChemistry, the Johns Hopkins University, School of Medicine, Baltimore, MD. Cells were grown in ascites form by i.p. injection into 100-to 150-g female Sprague-Dawley rats (from Holtzman Laboratories,Madison, WI). AS-30D cells were collected from ascites fluid andpurified according to Parry and Pedersen (14). Animals were fed andwatered ad libitum.
913
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

MITOCHONDRIA!. BOUND HEXOKINASE IN HEPATOMA CELLS
Mitochondria! Isolation. Tumor mitochondria were isolated fromAS-30D cells by the sonication protocol of Parry and Pedersen (14).Rat liver mitochondria were prepared by the high-yield method ofBustamante et al. (20). In both cases after the cell homogenization stepthe mitochondria were resuspended in H medium4 without bovine
serum albumin or EGTA added. Mitochondria were either used freshor else frozen in liquid nitrogen and thawed immediately prior to use.
Subcellular Localization of Glycolytic Enzyme Activities. Subcellularfractions of rat liver tissue and AS-30D cells were obtained by differential centrifugation of the homogenized cells according to Parry andPedersen (14). Purified mitochondria! fractions were prepared as described above. Homogenates of rat liver or AS-30D cells were subjectedto low speed (10 min at 20,000 x g) and high speed (45 min at 130,000x g) centrifugations. The high speed supernatants were used as cytosolicfractions.
Enzyme assays were performed at 30 °Cusing a Gilford Model 252
spectrophotometer attached to a circulating water bath. All assay mixtures contained 13 >(K/mlof oligomycin to inhibit F, ATPase activityand 13 ¿IMDAPP to inhibit adenylate kinase. Samples containingmembrane fractions were solubilized with 1 to 5 mg/mg of Lubrol WXprior to assay. Aldolase, enolase, and phosphoglucose isomerase wereassayed according to Bergmeyer et al. (21). Láclate dehydrogenaseactivity was measured by the UV absorbance technique of Vassault(22). Pyruvate kinase was assayed according to Fujii and Miwa (23).Phosphofructose kinase activity was measured as described by Duruibeand Tejwani (24). Hexokinase activity was determined according toParry and Pedersen (14). All enzyme assays were coupled to either theoxidation of NADH to NAD* or the reduction of NADP* to NADPH.
The rate of reaction was determined by following absorbance at 340nm and using a millimolar extinction coefficient of 6.22 for NADHand NADPH. Data are expressed as ¿¿molof substrate utilized per minper mg of protein (units/mg). All assays were performed on freshlyisolated material.
Assay of Hexokinase Activity in the Presence of Glucose-6-phosphate.Since the normal assay for hexokinase activity relies upon a coupledassay with glucose-6-phosphate dehydrogenase, it was necessary to usea new protocol to assay hexokinase activities in the presence of glucose-6-phosphate. The assay conditions developed were essentially as described by Bustamante and Pedersen (4), with the exception that Tris-bicarbonate buffer was replaced with an equimolar concentration ofTris-HCl. The final reaction mix contained 40 mM Tris-HCl, 10 mMMgCl2, 5 mM KCN, 0.2 mM NADH, 0.64 mM phosphoenol pyruvate,2 mM ATP, 55 units pyruvate kinase, 0.5 mM glucose, and 6 unitslactate dehydrogenase, pH 7.9. Production of ADP by hexokinase wascoupled to the oxidation of NADH via pyruvate kinase and lactatedehydrogenase, using phosphoenol pyruvate as substrate. The reactionrate was monitored by following the decrease in absorbance at 340 nm.A mM extinction coefficient of 6.22 was used to calculate ^mol ofproduct formed per min per mg of protein.
Preparation of Solubilized AS-30D Mitochondria! Hexokinase.Freshly prepared AS-30D mitochondria (5 to 10 mg/ml) were suspended in H medium with 1 mM phenylmethylsulfonyl fluoride, 1 mMEGTA, and 1 mM DTT added to inhibit protease activity. After additionof 1 mM glucose-6-phosphate the mitochondria were incubated for 15min at 30°C.This resulted in the selective solubilization of mitochon
dria! hexokinase (12). Following centrifugation of the suspension at20,000 x g for 15 min the resulting supernatant was centrifuged at130,000 x g for 45 min to remove mitochondria and particulatecontaminants. The high speed supernatant containing solubilized tumormitochondrial hexokinase was further purified by HPLC as describedbelow.
Chromatography of Hexokinase on DEAE-Cellulose. Hexokinasefrom either AS-30D mitochondria or normal rat liver was loaded ontoa 0.9 x 6-cm column of DEAE-cellulose (Cellex D, from Bio-Rad)equilibrated with buffer containing 10 mM triethanolamine HC1, pH
*The abbreviations used are: H medium, 210 mM r>-mannitol:70 mM sucrose:5HIM 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid, pH 7.4; EGTA, ethyl-eneglycol-bis-(/J-aminoethylether)-A',A'^''v/V'-tetraacetic acid; HPLC, high-performance liquid chromatography; SDS, sodium dodecyl sulfate; PAGE, polyacryl-amide gel electrophoresis; DIT, dithiothreitol; DAPP, /",/"-di(adenosine-5')pentaphosphate; PTC, phenylthiocarbamyl.
7.6, 1 mM DTT, and 0.5 mM EGTA. The column was washed with 40ml of equilibrating buffer to remove nonadsorbed proteins and thendeveloped with a 40-ml linear gradient from 0 to 0.5 M NaCl in thesame buffer at a flow rate of 18 ml/h. Fractions of 0.5 ml were collectedand assayed for hexokinase activity as described above. The chromatography was performed at a temperature of 4"C.
Purification of AS-30D Mitochondrial Hexokinase by HPLC. AS-30D mitochondrial hexokinase was solubilized as described above.After filtration through a 0.45-Mm HAWP filter (Millipore Corp.) theprotein was injected onto two tandem gel permeation columns (Waters1-60 and 300 SW Protein Pak columns). The columns were prewashedwith Buffer A [10 mM potassium phosphate (pH 7.0): 10 HIMglucose:0.5mM EDTA:2.5 mM DTT], and the enzyme was eluted with the samebuffer. Fractions containing peak hexokinase activity were collectedand loaded directly onto a Waters DEAE-5PW Protein Pak aniónexchange column preequilibrated with Buffer A. After washing withBuffer A for 5 min the enzyme was eluted with a 50-min linear gradientfrom 0 to 0.4 M NaCI in Buffer A. Hexokinase activity typically elutedas a sharp peak near the end of the gradient, of approximately 0.5- to1.0-ml total volume. In experiments where low salt concentration wasnecessary, the enzyme was diluted with distilled water and reconcen-trated by centrifugation for 45 min at 5000 x g in Centricon 10microconcentrators (Amicon). The HPLC system (purchased from theWaters Chromatography Division, Millipore Corporation, Milford,MA) consisted of an M-45 pump, a Model 510 pump, a U6K manualinjector with 2.0-ml sample loop, a Model 440 absorbance detectoroperated at 280 nm, a Model 680 automated gradient controller, anda Model 740 data module. The solvent flow rate was 0.5 ml per min.Buffer reservoirs were kept on ice during the chromatography, andfractions were transferred to an ice bucket immediately after elution inorder to retain maximum hexokinase catalytic activity.
PAGE. Electrophoresis in the presence of SDS was performed asdescribed by Laemmli (25) using a Bio-Rad Protean I slab cell apparatus. Proteins were precipitated with acetone prior to solubilization in2.5% SDS:5% 2-mercaptoethanol:10% glycerol:62.5 mM Tris-HCl, pH6.8. Samples were heated for 2 min at 1()<•"(before loading onto the
gel. Electrophoresis was performed for 2500 V-h overnight in a water-cooled apparatus.
Mitochondrial Binding of Hexokinase. Beef heart hexokinase andyeast hexokinase purchased from the Sigma Chemical Co. were purifiedby HPLC gel permeation chromatography on Waters 1-60 and 300 SWcolumns under the conditions described above. AS-30D hexokinasewas purified by HPLC as described in a preceding section. A constantamount of catalytic activity of each of the hexokinases (0.5 units) wasincubated at room temperature with rat liver mitochondria (1 mgprotein) in 0.5 ml of H medium containing 10 mM MgCl2. After 15min the suspensions were diluted with 0.9 ml of ice-cold H mediumand centrifuged for 2 min at 12,800 x g in an Eppendorf microfuge.The supernatants were decanted, and the mitochondrial pellets containing bound hexokinase were assayed as described above. Mitochondrialbinding of hexokinase was assayed using rat liver mitochondria ratherthan AS-30D mitochondria in order to avoid problems arising fromthe presence of high levels of endogenous bound hexokinase activity inthe tumor mitochondrial fraction.
Amino Acid Composition Analysis. The amino acid composition ofpurified AS-30D hexokinase was determined by the technique of Hidlingmeyer et al. (26) using a Pico-Tag work station (Waters Chromatography) to hydrolyze and derivatize the protein and a Waters high-performance liquid chromatography apparatus to analyze the derivati/ed amino acids. The AS-30D mitochondrial enzyme was purified asdescribed above and lyophilized. The dried samples were hydrolyzedwith 6 N constant boiling HC1 (Pierce) in vapor phase for 24 h at110°C.After hydrolysis the free amino acids were dried and derivatized
with phenylisothiocyanate, forming the PTC derivatives. The PTC-amino acids were injected onto a C|8 reverse-phase column (WatersPico-Tag column) and eluted with a convex gradient from 10 to 60%acetonitrile in sodium acetate buffer, pH 6.35. Eluting PTC-aminoacids were detected by absorbance at 254 nm using a fixed-wavelengthdetector (Waters Model 440). The amounts of PTC-amino acids elutedwere calibrated to known injections of 250 pmol each of PTC-deriva-
914
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

MITOCHONDRIA!. BOUND HEXOK1NASE IN HEPATOMA CELLS
tized amino acid standards (from Pierce). The chromatography wasperformed at 38°Cat a flow rate of 1 ml/min. It should be noted that
the conditions used in this analysis tend to result in a low recovery ofcysteine residues.
Preparation of Polyclonal Antibodies to Tumor Hexokinase. Hexoki-nase solubilized from AS-30D mitochondria was purified as describedabove except that the HPLC sizing column was replaced with a 0.9- xSS-cm Sepharose CL-6B column. The resultant preparation showedonly a single band upon SDS:PAGE exhibiting a molecular weight of98,000. Purified AS-30D hexokinase (100 /<«)was mixed with an equalvolume of complete Freund's adjuvant and injected s.c. into the back
of a male New Zealand white rabbit. The rabbit was boosted at 3-wkintervals with 100-^g injections of AS-30D hexokinase mixed with anequal volume of incomplete Freund's adjuvant. Serum containing pol-
yclonal antibodies to hexokinase was prepared by allowing rabbit bloodto clot overnight at 4°C,followed by centrifugaron for 10 min at 700
x g. The resulting clear supernatant was decanted off and diluted 1:250for use in Western blot experiments.
Assay of Immune Cross-Reactivity by Western Blot Analysis. Theability of polyclonal antibodies prepared against AS-30D hexokinaseto cross-react with hexokinases from normal rat tissues was determinedby Western blot analysis. Homogenates of rat brain, rat kidney, and ratliver, along with purified AS-30D hexokinase, were subjected toSDS:PAGE according to Laemmli (25). After electroblotting onto anitrocellulose filter the proteins were incubated with a 1:250 dilutionof rabbit serum containing polyclonal antibodies to AS-30D hexokinase. Antibody-reactive polypeptides were localized on the nitrocellulose filter by autoradiography following incubation with '"I-labeled
Protein A.Other Techniques. Membrane protein was estimated by the biuret
method in the presence of 0.25% sodium cholate using bovine serumalbumin as a standard (27). Soluble protein was measured by the methodof Lowry et al. (28) using bovine serum albumin as standard. Solubleproteins were routinely precipitated with a 9-fold volume of coldacetone and redissolved in 100 ¿ilof 10% SDS prior to performing theLowry assay in order to avoid interference by dithiothreitol.
RESULTS
Comparison of Glycolytic Enzyme Activities in AS-30D Hep-atoma Cells and in Normal Rat Liver Tissue. We have comparedthe specific activities for a number of glycolytic enzymes in theAS-30D hepatoma and in normal rat liver (see Table 1). Themost striking difference between normal and transformed cellsis the great increase in low Km hexokinase in the tumor cell,representing a 136-fold increase in specific activity over normalrat liver. Elevated specific activities in the tumor were notedfor several other key glycolytic enzymes, including phospho-fructose kinase (17-fold increase) and pyruvate kinase (10-fold
increase).
Table 1 Comparison of glycolytic enzyme-specific activities in AS-30D hepatomaand normal rat liver tissue
The specific activities of a number of glycolytic enzymes were determined inhomogenates of normal rat liver tissue and purified AS-30D cells (see "Materialsand Methods").
Specific activity (units/mg)
EnzymeHexokinase
(low KmfPhosphoglucose isomerasePhosphofructose kinaseAldolaseEnolasePyruvate kinaseLáclatedehydrogenaseRat
liver0.005±0.001*
0.35 ±0.040.01 ±0.0010.04 ±0.0010.15 ±0.020.22 ±0.021.83 ±0.32Hepatoma0.68
±0.050.71 ±0.140.17± 0.040.11 ±0.010.66 ±0.082.28 ±0.475.24 ±1.29Ratio136
21734
103
Specificity of Mitochondria! Binding of Hexokinase in AS-30D Hepatoma Cells. It has commonly been observed that mostof the hexokinase in rapidly growing tumor cells is bound tothe outer mitochondrial membrane (4-6, 9, 12-14, 29). Thepossibility that other glycolytic enzymes might also be associated with tumor mitochondria in a multienzyme complex hasnot been examined. Therefore, we investigated the subcellularlocalization of the glycolytic enzymes in the AS-30D hepatomacell. A comparison of specific activities in the cytosolic andmitochondrial fractions is presented in Table 2. It is apparentfrom these data that the only enzyme which shows a markedlyelevated specific activity in the mitochondrial fraction is hexokinase, with a specific activity over 3-fold higher in the mitochondrial fraction than in the cytosol. For all of the otherglycolytic enzymes which we examined the ratio of specificactivities in the mitochondria! fraction versus the cytosol wassubstantially less than one. These results emphasize that thebinding of hexokinase to its outer mitochondrial receptor complex (14, 19) in the AS-30D tumor is a highly specific event.
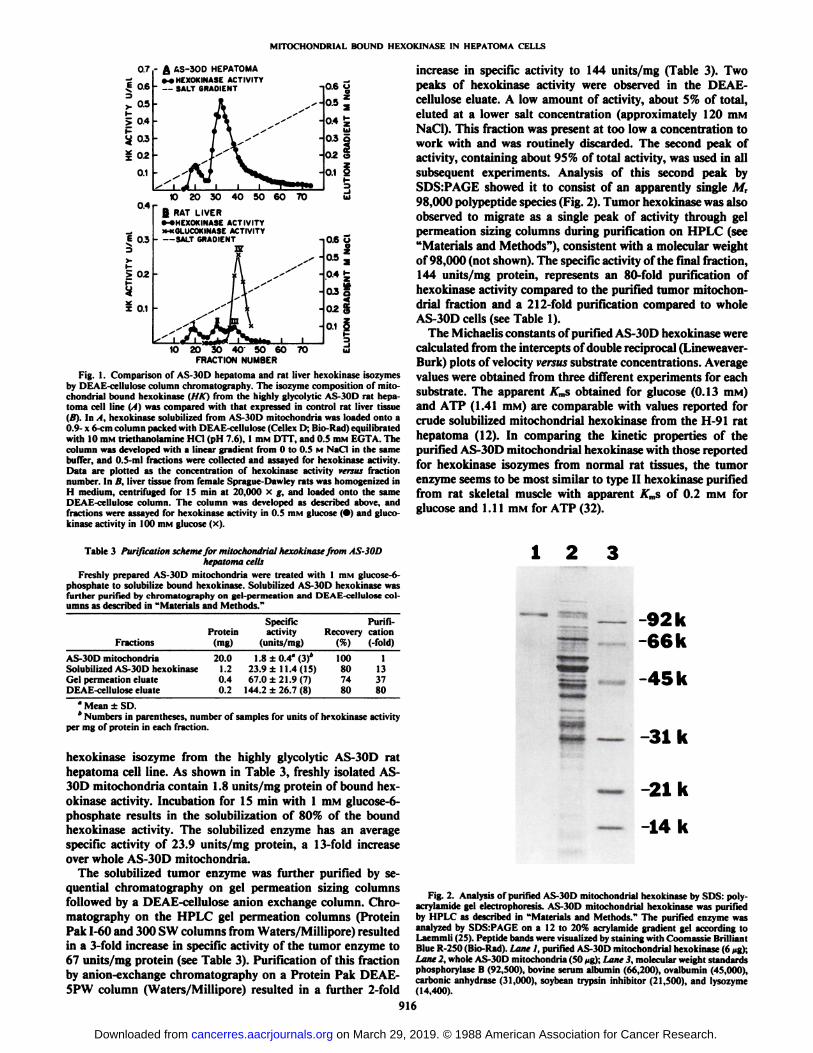
Isozyme Composition of AS-30D Mitochondrial Hexokinase.We have characterized the AS-30D mitochondrial hexokinaseby column chromatography on DEAE-cellulose. The pattern ofelution of hexokinase from DEAE-cellulose columns has beenused as a standard technique to define hexokinase isozymetypes (30, 31). The elution profile of AS-30D mitochondrialhexokinase from DEAE-cellulose is shown in Fig. \A. In theseexperiments the AS-30D hexokinase used was not HPLC purified, but rather was solubilized by glucose-6-phosphate treatment of AS-30D mitochondria. As shown in Fig. Ifi, there arefour isozymes of hexokinase present in normal rat liver tissue.Types I, II, and III represent forms with low Kms for glucoseand can be assayed at a glucose concentration of 0.5 ITIM.TypeIV hexokinase (glucokinase) has a much higher A',,,for glucose
and is assayed in 100 ITIMglucose. The bulk of the hexokinasepresent in normal rat liver is of the type IV isozyme. A comparison of the elution profiles of hepatoma mitochondrial hexokinase (Fig. \A) with normal liver hexokinases (Fig. IB) showsthat there is a shift from predominantly high A',,,isozyme (type
IV) in normal liver to low Km isozymes in the transformed cellline. The bulk of the tumor enzyme coelutes from DEAE-cellulose columns with the type II isozyme of normal rat liver.A minor amount of the tumor mitochondrial hexokinase coelutes with the type I isozyme of normal liver. We have repeatedthis experiment 4 times with essentially identical results.
Purification of Mitochondrial Hexokinase from AS-30D Hepatoma Cells. Treatment of tumor mitochondria with millimolarconcentrations of either ATP or glucose-6-phosphate results inthe selective solubilization of bound hexokinase (9, 12, 14). Wehave used glucose-6-phosphate-solubilized mitochondrial hexokinase as the starting material for the purification of the bound
Table 2 Subcellular localization ofglycolylic enzyme activities in the highlyglycolytic AS-30D hepatoma cell line
Cytosolic and mitochondrial fractions were isolated from AS-30D cells, andenzyme activities were measured as described in "Materials and Methods."
Specific activity (unils/mg)
" At the glucose concentration (0.5 RIM)used for this assay, liver glucokinasewhich has a very high A,,,for glucose (12 m\i) does not contribute to the activitymeasured.
* Mean ±SD (n = 3) for units of activity per mg of cell protein.
EnzymeHexokinasePhosphoglucose
isomerasePhosphofructosekinaseAldolaseEnolasePyruvate
kinaseLactatedehydrogenaseCytosolic0.36
±0.14°0.680.170.180.672.660.300.050.030.100.855.10
1.18Mitochondrial1.25
±0.340.04±0.030.00
1 ±0.000.04±0.020.02±0.020.09±0.040.40±0.14Ratio3.470.060.010.220.030.030.08
"Mean ±SD (n = 3) for units of enzyme activity per mg of cytosolic or
mitochondrial protein.
915
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
MITOCHONDRIAL BOUND HEXOKINASE IN HEPATOMA CELLS
0.7
I 0.6
£0.5
5 0.4
SÃ0.3
M 0.2
0.1
0.4
A AS-30D HEPATOMA
•-•HEXOKINASEACTIVITY— SALT GRADIENT 0.6 <J
0.5*
0.4 £kj
0.3 5
0.2 5
0.1 1
0.3
0.1
K) 20 30 40 50 60 70
fi RAT LIVER•-•HEXOKINASEACTIVITYx-xGLUCOKINASE ACTIVITYSALT GRADIENT
0.6 U
0.5 f
.0*50.3 ¡
0.2 g
0.1 I
10 20 30 40 50 60 70 uFRACTION NUMBER
Fig. 1. Comparison of AS-30D hepatoma and rat liver hexokinase isozymesby DEAE-cellulose column chromatography. The isozyme composition of mitochondria! bound hexokinase (HK) from the highly glycolytic AS-30D rat hepatoma cell line (.11 was compared with that expressed in control rat liver tissueili). In . I. hexokinase solubilized from AS-30D mitochondria was loaded onto a0.9- x 6 mi column packed with DEAE-cellulose (Cellex D; Bio-Rad) equilibratedwith 10 mM triethanolamine HC1 (pH 7.6), 1 mM DTT, and 0.5 mM EGTA. Thecolumn was developed with a linear gradient from 0 to 0.5 M NaCI in the samebuffer, and 0.5-ml fractions were collected and assayed for hexokinase activity.Data are plotted as the concentration of hexokinase activity versus fractionnumber. In I!, liver tissue from female Sprague-Dawley rats was homogenized inH medium, centrifuged for 15 min at 20,000 x g, and loaded onto the sameDEAE-cellulose column. The column was developed as described above, andfractions were assayed for hexokinase activity in 0.5 mM glucose (•)and gluco-kinase activity in 100 mM glucose (x).
Table 3 Purification scheme for mitochondrial hexokinase from AS-30Dhepatoma cells
Freshly prepared AS-30D mitochondria were treated with 1 mM glucose-6-phosphate to solubilize bound hexokinase. Solubilized AS-30D hexokinase wasfurther purified by chromatography on gel-permeation and DEAE-cellulose columns as described in "Materials and Methods."
FractionsProtein
(mg)
Specificactivity
(units/mg)
Purifi-Recovery cation
(%) (-fold)AS-30D mitochondria 20.0 1.8 ±0.4°(3)* 100 1Solubilized AS-30D hexokinase 1.2 23.9 ±11.4 (15) 80 13Gel permeation eluate 0.4 67.0 ±21.9 (7) 74 37DEAE-cellulose eluate 0.2 144.2 ±26.7 (8) 80 80
°Mean ±SD.'' Numbers in parentheses, number of samples for units of hexokinase activity
per mg of protein in each fraction.
hexokinase isozyme from the highly glycolytic AS-30D rathepatoma cell line. As shown in Table 3, freshly isolated AS-30D mitochondria contain 1.8 units/mg protein of bound hexokinase activity. Incubation for IS min with 1 mM glucose-6-phosphate results in the solubilization of 80% of the boundhexokinase activity. The solubilized enzyme has an averagespecific activity of 23.9 units/mg protein, a 13-fold increaseover whole AS-30D mitochondria.
The solubilized tumor enzyme was further purified by sequential chromatography on gel permeation sizing columnsfollowed by a DEAE-cellulose anión exchange column. Chromatography on the HPLC gel permeation columns (ProteinPak 1-60 and 300 SW columns from Waters/Millipore) resultedin a 3-fold increase in specific activity of the tumor enzyme to67 units/mg protein (see Table 3). Purification of this fractionby anion-exchange chromatography on a Protein Pak DEAE-5PW column (Waters/Millipore) resulted in a further 2-fold
increase in specific activity to 144 units/mg (Table 3). Twopeaks of hexokinase activity were observed in the DEAE-cellulose eluate. A low amount of activity, about 5% of total,eluted at a lower salt concentration (approximately 120 mMNaCI). This fraction was present at too low a concentration towork with and was routinely discarded. The second peak ofactivity, containing about 95% of total activity, was used in allsubsequent experiments. Analysis of this second peak bySDS:PAGE showed it to consist of an apparently single M,98,000 polypeptide species (Fig. 2). Tumor hexokinase was alsoobserved to migrate as a single peak of activity through gelpermeation sizing columns during purification on HPLC (see"Materials and Methods"), consistent with a molecular weight
of 98,000 (not shown). The specific activity of the final fraction,144 units/mg protein, represents an 80-fold purification ofhexokinase activity compared to the purified tumor mitochondrial fraction and a 212-fold purification compared to wholeAS-30D cells (see Table 1).
The Michaelis constants of purified AS-30D hexokinase werecalculated from the intercepts of double reciprocal (Lineweaver-Burk) plots of velocity versus substrate concentrations. Averagevalues were obtained from three different experiments for eachsubstrate. The apparent A"msobtained for glucose (0.13 HIM)
and ATP (1.41 HIM) are comparable with values reported forcrude solubilized mitochondrial hexokinase from the II 91 rathepatoma (12). In comparing the kinetic properties of thepurified AS-30D mitochondrial hexokinase with those reportedfor hexokinase isozymes from normal rat tissues, the tumorenzyme seems to be most similar to type II hexokinase purifiedfrom rat skeletal muscle with apparent ATmsof 0.2 mM forglucose and I.I 1 mM for ATP (32).
1 2
-92k-66k
-45k
5 —-31k
— -21k
— -14k
Fig. 2. Analysis of purified AS-30D mitochondrial hexokinase by SDS: poly-acrylamide gel electrophoresis. AS-30D mitochondrial hexokinase was purifiedby HPLC as described in "Materials and Methods." The purified enzyme was
analyzed by SDSrPAGE on a 12 to 20% acrylamide gradient gel according toLaemmli (25). Peptide bands were visualized by staining with Coomassie BrilliantBlue R-250 (Bio-Rad). Lane 1, purified AS-30D mitochondrial hexokinase (6 /ig);Lane 2, whole AS-30D mitochondria (50 jig); Lane 3, molecular weight standardsphosphorylase B (92,500), bovine serum albumin (66,200), ovalbumin (45,000),carbonic anhydrase (31,000), soybean trypsin inhibitor (21,500), and lysozyme(14,400).
916
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
MITOCHONDRIA!. BOUND HEXOKINASE IN HEPATOMA CELLS
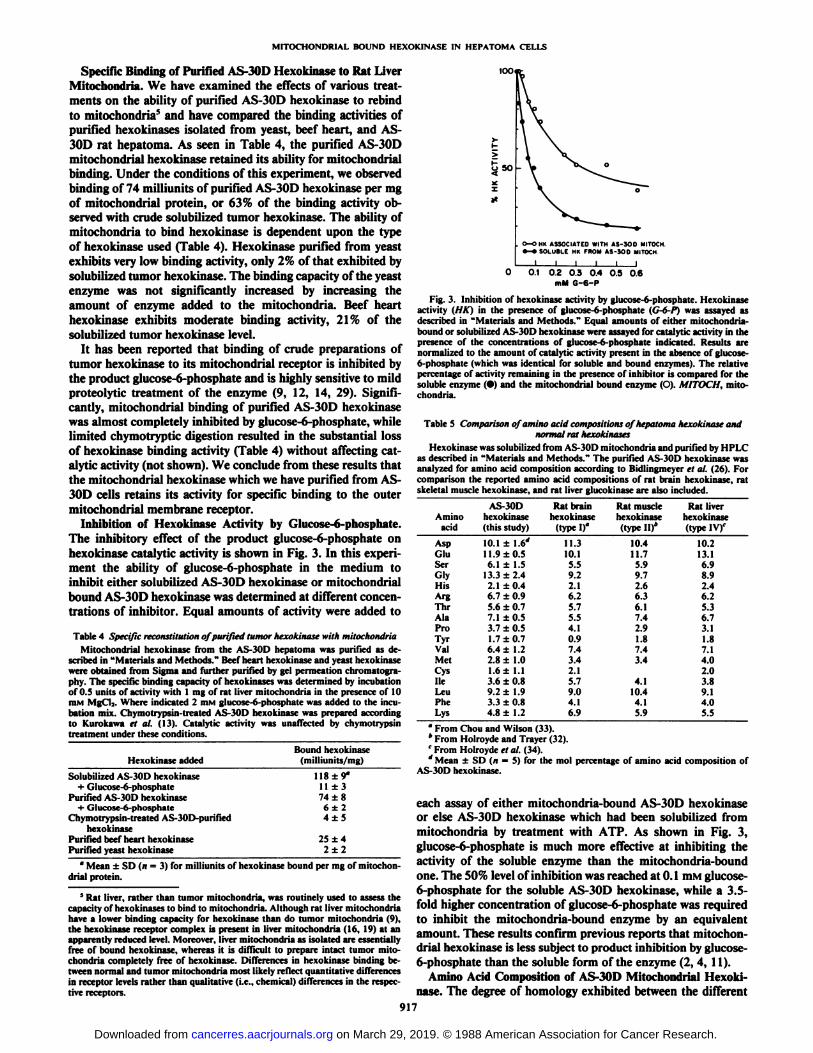
Specific Binding of Purified AS-30D Hexokinase to Rat LiverMitochondria. We have examined the effects of various treatments on the ability of purified AS-30D hexokinase to rebindto mitochondria5 and have compared the binding activities ofpurified hexokinases isolated from yeast, beef heart, and AS-30D rat hepatoma. As seen in Table 4, the purified AS-30Dmitochondria! hexokinase retained its ability for mitochondria!binding. Under the conditions of this experiment, we observedbinding of 74 milliunits of purified AS-30D hexokinase per mgof mitochondria! protein, or 63% of the binding activity observed with crude solubilized tumor hexokinase. The ability ofmitochondria to bind hexokinase is dependent upon the typeof hexokinase used (Table 4). Hexokinase purified from yeastexhibits very low binding activity, only 2% ofthat exhibited bysolubilized tumor hexokinase. The binding capacity of the yeastenzyme was not significantly increased by increasing theamount of enzyme added to the mitochondria. Beef hearthexokinase exhibits moderate binding activity, 21% of thesolubilized tumor hexokinase level.
It has been reported that binding of crude preparations oftumor hexokinase to its mitochondria! receptor is inhibited bythe product glucose-6-phosphate and is highly sensitive to mildproteolytic treatment of the enzyme (9, 12, 14, 29). Significantly, mitochondria! binding of purified AS-30D hexokinasewas almost completely inhibited by glucose-6-phosphate, whilelimited chymotryptic digestion resulted in the substantial lossof hexokinase binding activity (Table 4) without affecting catalytic activity (not shown). We conclude from these results thatthe mitochondria! hexokinase which we have purified from AS-30D cells retains its activity for specific binding to the outermitochondria! membrane receptor.
Inhibition of Hexokinase Activity by Glucose-6-phosphate.The inhibitory effect of the product glucose-6-phosphate onhexokinase catalytic activity is shown in Fig. 3. In this experiment the ability of glucose-6-phosphate in the medium toinhibit either solubilized AS-30D hexokinase or mitochondria!bound AS-30D hexokinase was determined at different concentrations of inhibitor. Equal amounts of activity were added to
Table 4 Specific reconstitution of purified tumor hexokinase with mitochondriaMitochondria! hexokinase from the AS-30D hepatoma was purified as de
scribed in "Materials and Methods." Beef heart hexokinase and yeast hexokinasewere obtained from Sigma and further purified by gel permeation chromatogra-phy. The specific binding capacity of hexokinases was determined by incubationof 0.5 units of activity with 1 mg of rat liver mitochondria in the presence of 10min MgCl2. Where indicated 2 HIMglucose-6-phosphate was added to the incubation mix. Chymotrypsin-lreated AS-30D hexokinase was prepared accordingto Kurokawa et al. (13). Catalytic activity was unaffected by chymotrypsintreatment under these conditions.
too
Hexokinase addedBound hexokinase
(milliunits/mg)
Solubilized AS-30D hexokinase+ Glucose-6-phosphate
Purified AS-30D hexokinase+ Glucose-6-phosphatc
Chymotrypsin-treated AS-30D-purifiedhexokinase
Purified beef heart hexokinasePurified yeast hexokinase
118 + 9°
11 ±374 ±8
6±24±5
25 ±42±2
" Mean ±SD (n = 3) for milliunits of hexokinase bound per mg of mitochon
dria! protein.
5 Rat liver, rather than tumor mitochondria, was routinely used to assess the
capacity of hexokinases to bind to mitochondria. Although rat liver mitochondriahave a lower binding capacity for hexokinase than do tumor mitochondria (9),the hexokinase receptor complex is present in liver mitochondria (16, 19) at anapparently reduced level. Moreover, liver mitochondria as isolated are essentiallyfree of bound hexokinase, whereas it is difficult to prepare intact tumor mitochondria completely free of hexokinase. Differences in hexokinase binding between normal and tumor mitochondria most likely reflect quantitative differencesin receptor levels rather than qualitative (i.e., chemical) differences in the respective receptors.
o—OHK ASSOCIATED WITH AS-300 MITOCM•-•SOLUBLE MK FROM AS-300 MITOCH
0 0.1 02 0.3 0.4 0.5 0.6mM G-6-P
Fig. 3. Inhibition of hexokinase activity by glucose-6-phosphate. Hexokinaseactivity (HK) in the presence of glucose-6-phosphate (G-6-P) was assayed asdescribed in "Materials and Methods." Equal amounts of either mitochondria-bound or solubilized AS-30D hexokinase were assayed for catalytic activity in thepresence of the concentrations of glucose-6-phosphate indicated. Results arenormalized to the amount of catalytic activity present in the absence of glucose-6-phosphate (which was identical for soluble and bound enzymes). The relativepercentage of activity remaining in the presence of inhibitor is compared for thesoluble enzyme (•)and the mitochondria! bound enzyme (O). MITOCH, mitochondria.
Table 5 Comparison of amino acid compositions ofhepatoma hexokinase andnormal rat hexokinases
Hexokinase was solubilized from AS-30D mitochondria and purified by 11l'I <as described in "Materials and Methods." The purified AS-30D hexokinase was
analyzed for amino acid composition according to Bidlingmeyer et al. (26). Forcomparison the reported amino acid compositions of rat brain hexokinase, ratskeletal muscle hexokinase, and rat liver glucokinase are also included.
AminoacidAspGluSerGlvHisArgThrAlaProTyrValMetCysHeLeuPheLysAS-30D
hexokinase(thisstudy)10.1
±1.6'11.9
±0.56.1±1.513.3
±2.42.1±0.46.7
±0.95.6±0.77.1
±0.53.7±0.51.7
±0.76.4±1.22.8±1.01.6±1.13.6±0.89.2±1.93.3±0.84.8±1.2Rat
brainhexokinase
(typeI)°11.310.15.59.22.16.25.75.54.10.97.43.42.15.79.04.16.9Rat
musclehexokinase(typeII)*10.411.75.99.72.66.36.17.42.91.87.43.44.110.44.15.9Rat
liverhexokinase(typeIV)C10.213.16.98.92.46.25.36.73.11.87.14.02.03.89.14.05.5
* From Chou and Wilson (33).* From Holroyde and Trayer (32).c From Holroyde el al. (34).'' Mean ±SD (n = 5) for the mol percentage of amino acid composition of
AS-30D hexokinase.
each assay of either mitochondria-bound AS-30D hexokinaseor else AS-30D hexokinase which had been solubilized frommitochondria by treatment with ATP. As shown in Fig. 3,glucose-6-phosphate is much more effective at inhibiting theactivity of the soluble enzyme than the mitochondria-boundone. The 50% level of inhibition was reached at 0.1 mM glucose-6-phosphate for the soluble AS-30D hexokinase, while a 3.5-fold higher concentration of glucose-6-phosphate was requiredto inhibit the mitochondria-bound enzyme by an equivalentamount. These results confirm previous reports that mitochondria! hexokinase is less subject to product inhibition by glucose-6-phosphate than the soluble form of the enzyme (2, 4, 11).
Amino Acid Composition of AS-30D Mitochondria! Hexokinase. The degree of homology exhibited between the different
917
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

MITOCHONDRIA!. BOUND HEXOKINASE IN HEPATOMA CELLS
MW Standard! 92 66 45 31UPO) III *
HR*
AS-300 Heiokinose
. Rat Brain Homogenote
Kidney Homogenot«
Rat Liver Homogenate
Fig. 4. Antibody cross-reactivity between tumor and normal rat hexokinases.Polyclonal rabbit antibodies were raised against the AS-30D mitochondrial hex-okinase purified as described in "Materials and Methods." Samples of AS-30D
hexokinase (I; 10 ^g), rat brain homogenate (B; 100 jig), rat kidney homogenate(C; 100 >ig)and rat liver homogenate (l>, 100 >ig)were subjected to SDSrPAGEaccording to Laemmli (25). After electroblotting onto a nitrocellulose filter theproteins were incubated with serum containing anti-hexokinase antibody. Protein-IgG complexes were visualized by incubation with 12!I-Protein A followed byautoradiography using Kodak X-omat X-ray film. The data shown representdensitometric traces of a representative autoradiograph from a sample of threedifferent experiments.
hexokinase isozymes should be reflected in their amino acidcompositions. A direct comparison of sequence homology requires the cloning and sequencing of the various hexokinasegenes, a task which has not yet been performed for any organismabove the level of yeast. In the absence of sequence data therelative homology between the tumor hexokinase isozyme andother isozymes from normal rat tissues can be estimated fromamino acid composition data. The amino acid composition ofpurified AS-30D mitochondrial hexokinase was analyzed byreverse-phase HPLC of the PTC-derivatized amino acids asdescribed in "Materials and Methods." Results for the tumor
enzyme represent averaged values for five different preparationsof purified AS-30D mitochondrial hexokinase. The data arepresented in Table 5, along with the reported amino acidcompositions of hexokinases type I (rat brain) (33), type II (ratskeletal muscle) (32), and type IV (rat liver) (34). It should benoted that these values were obtained from three differentlaboratories by different techniques.
Immune Cross-Reactivity of Hepatoma Hexokinase and Normal Rat Hexokinases. Polyclonal antibodies were preparedagainst the AS-30D mitochondrial hexokinase as described in"Materials and Methods." The ability of the anti-AS-30D hexokinase antibody to cross-react with hexokinases from normalrat tissues was determined by Western blot analysis using theanti-hexokinase antibody, followed by incubation with 125I-
labeled Protein A. Fig. 4 shows a densitometric trace of arepresentative autoradiograph. As shown in Fig. 4, Lane A, thepurified AS-30D hexokinase reacts strongly with the rabbitantibody which was raised against it. Antibody cross-reactivitywas seen with rat brain hexokinase (Fig. 4, Lane B), and withrat kidney hexokinase (Fig. 4, Lane C). No apparent cross-reactivity was observed with rat liver, the principal hexokinaseisozyme of which is type IV (glucokinase) (Fig. 4, Lane D). Inall three tissues a very strongly antibody-reactive M, ~66,000
peptide is observed (Fig. 4). The identity of this M, 66,000peptide is unknown. However, it may represent a proteolyticproduct of hexokinase. From these results it would appear thattumor hexokinase exhibits some epitopic similarities with bothrat brain and kidney hexokinases.
DISCUSSION
Results of experiments reported here provide new data relevant to the aberrant glucose catabolic properties of rapidlygrowing tumor cells. The data presented demonstrate that lowKmhexokinase activity is greatly elevated in the highly glycolyticAS-30D rat hepatoma cell line, over 100-fold higher than thatdetected in normal rat liver tissue. Previous studies have shownthat the tumor hexokinase binds to a receptor complex locatedin the outer mitochondrial membrane (6, 9, 12, 14, 16-19). Inthe present report we demonstrate that hexokinase is the onlyglycolytic enzyme which apparently binds specifically to thismitochondrial receptor. The specific activity of AS-30D hexokinase in an isolated mitochondrial fraction was 3.5-fold higherthan in a corresponding cytosolic fraction, while for all otherglycolytic enzymes examined this ratio was substantially lessthan one. Mitochondrial binding of hexokinase has been reported to result in an increased affinity of the enzyme forsubstrate, a decrease in inhibition of enzyme activity by theproduct glucose-6-phosphate, and in a preferred access to mitochondrial synthesized ATP (2, 4, 5, 11-13, 15, 35). It hasrecently been demonstrated that the mitochondrial receptor forhexokinase consists at least in part of the outer membranechannel-forming protein (also known as VDAC or porin) (16-19). Since all hydrophilic, low-molecular-weight molecules, including ATP and ADP, must presumably cross the outer mitochondrial membrane through the channel-forming protein, itis easy to envision how hexokinase bound to this same proteincould obtain preferential access to matrix-generated ATP. Thespecificity of this receptor for binding hexokinase would appearto be significant in supporting elevated glucose catabolism intumor cells.
We report a rapid HPLC protocol for purification of AS-30D mitochondrial hexokinase which results in the purificationto apparent homogeneity of a tumor enzyme of high specificactivity (144 units/mg protein). Significantly, the purified tumor mitochondrial hexokinase retains its ability to rebind tothe mitochondrial receptor. Moreover, binding is prevented byeither glucose-6-phosphate or chymotrypsin, characteristic ofhexokinase binding to the mitochondrial receptor (9, 12, 14,29). It would thus appear that the purification technique useddoes not substantially affect the binding ability of AS-30Dhexokinase.
Characterization of the AS-30D mitochondrial hexokinaseby its kinetic and Chromatographie properties indicates that themajor form corresponds to the type II isozyme found in normalrat tissues. However, comparison of the amino acid compositions of normal and hepatoma hexokinases indicates that thetumor enzyme is not identical to any known hexokinase isozyme which has been purified from normal rat tissues. Polakisand Wilson (36) observed that bindable forms of normal ratbrain hexokinase contain an /V-terminal binding domain whichis required for interaction with the mitochondrial receptor.Presumably this binding domain is removed by limited prote-olysis in those tissues in which hexokinase does not bind to theparticulate fraction (9, 13, 36). It is possible that the differencein amino acid composition of tumor hexokinase may be relatedto the presence of a similar binding domain on the A-terminal
918
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

MITOCHONDRIAL BOUND HEXOKINASE IN HEPATOMA CELLS
end of the protein. Since the tumor enzyme is /V-terminallyblocked, it has not yet been possible to sequence this domainof AS-30D hexokinase for comparison with the rat brain enzyme.
Studies with antibodies made against AS-30D hexokinaseshow immune cross-reactivity with normal hexokinases fromrat brain and rat kidney, but not rat liver tissue. It should benoted that, under the conditions used, the results indicate eitherthe presence or absence of epitopic similarities between tumorand normal isozymes, not how similar in structure the isozymesare. Still, we consider it significant that the anti-AS-30D antibody does not cross-react with type IV hexokinase (glucoki-nase), which is the predominant isozyme present in normalliver tissue. The type IV hexokinase of liver differs substantiallyfrom hepatoma hexokinase in other characteristics as well. Themolecular mass of type IV hexokinase is 48,000 daltons (34),in contrast to the 98,000-daIton tumor enzyme. The Km forglucose of type IV hexokinase is two orders of magnitude higherthan the tumor isozyme (12 mivi versus 0.13 HIM). Type IVhexokinase appears to be predominantly localized in the cyto-sol, while 60% of tumor hexokinase occurs bound to the outermitochondria! membrane (14). Finally, the liver enzyme is notinhibited by the product glucose-6-phosphate, whereas the soluble tumor enzyme is significantly inhibited by physiologicalconcentrations of glucose-6-phosphate. (It should be noted thatthe mitochondria! bound tumor enzyme is much less sensitiveto glucose-6-phosphate inhibition). Thus, in the transformationfrom low glucose-utilizing normal liver to high glucose-utilizinghepatoma, the major isozyme of hexokinase shifts to a formwith substantially different kinetic, physical, and regulatoryproperties.
Note Added in Proof.
Since this manuscript was submitted for publication Radojk-ovic and Ureta (37) have reported the purification of hexokinases Type B and C from the Novikoff hepatoma. HexokinaseB is derived from the mitochondria and is considered to behexokinase II.
REFERENCES
1. Warburg, O., Posener, K., and Negelein, E. Ãœberden Stoffwechsel dercarcinomzelle. Biochem. Z., 752: 309-344, 1924.
2. Singh, V. N., Singh, M., August, J. T., and Horecker, B. L. Alterations inglucose metabolism in chick-embryo cells transformed by Rous sarcomavirus: intracellular levels of glycolytic intermediates. Proc. Nati. Acad. Sci.USA, 71: 4129-4132, 1974.
3. Weinhouse, S. Glycolysis, respiration, and anomalous gene expression inexperimental hepatomas: G. H. A. Clowes memorial lecture. Cancer Res.,32: 2007-2016. 1972.
4. Bustamante, 1 .. and Pedersen, P. L. High aerobic glycolysis of rat hepatomacells in culture: role of mitochondria! hexokinase. Proc. Nati. Acad. Sci.USA, 74: 3735-3739, 1977.
5. Pedersen, P. L. Tumor mitochondria and the bioenergetics of cancer cells.Prog. Exp. Tumor Res., 22: 190-274, 1978.
6. Bustamante, E., Morris, H. P., and Pedersen, P. L. Energy metabolism oftumor cells. Requirement for a form of hexokinase with a propensity formitochondrial binding. J. Biol. Chem., 25«:8699-8704, 1981.
7. Nakashima, R. A., Paggi. M. G., and Pedersen, P. L. Contributions ofglycolysis and oxidative phosphorylation to adenosine 5'-triphosphate production in AS-30D hepatoma cells. Cancer Res., 44: 5702-5706. 1984.
8. Burk, D., Woods. M., and Hunter, J. On the significance of glucolysis for
cancer growth, with special reference to Morris rat hepatomas. J. Nati.Cancer Inst., 38: 839-863, 1967.
9. Rose, I. A., and Warms, J. V. B. Mitochondrial hexokinase: release, rebind-ing, and location. J. Biol. Chem., 242: 1635-1645,1967.
10. Knox, W. E., Jamdar, S. C. and Davis, P. A. Hexokinase, differentiation,and growth rates of transplanted rat tumors. Cancer Res.. 30: 2240-2244,1970.
11. Gumaa, K. A., and McLean, P. A possible interrelationship between bindingof hexokinase and the site of ATP formation in Krebs ascites cells. Biochem.Biophys. Res. Commun., 36: 771-779, 1969.
12. Bustamante, E., and Pedersen, P. L. Mitochondrial hexokinase of rat hepatoma cells in culture: solubilization and kinetic properties. Biochemistry. 19:4972-4977, 1980.
13. Kurokawa, M., Tokuoka, S., Oda, S., Tsubotani, E.. and Ishibashi, S.Difference in efficiency of function between mitochondria-bound hexokinaseand non-bound one. Biochem. Int.. 2:645-650, 1981.
14. Parry, D. M., and Pedersen, P. L. Intracellular localization and properties ofpaniculate hexokinase in the Novikoff ascites tumor. J. Biol. Chem., 258:10904-10912, 1983.
15. lnui. M., and Ishibashi. S. Functioning of mitochondria-bound hexokinasein rat brain in accordance with generation of ATP inside the organelle. J.Biochem., 85: 1151-1156, 1979.
16. Felgner, P. L., Messer, J. L., and Wilson, J. E. Purification of a hexokinase-binding protein from the outer mitochondrial membrane. J. Biol. Chem.,254:4946-4949, 1979.
17. Fiek, C., Benz, R., Roos, N., and Brdiczka. D. Evidence for identity betweenthe hexokinase-binding protein and the mitochondrial porin in the outermembrane of rat liver mitochondria. Biochim. Biophys. Acta, 6SS: 429-440,1982.
18. Linden, M., Gellerfors, P.. and Nelson, B. D. Pore protein and the hexokinase-binding protein from the outer membrane of rat liver mitochondria areidentical. FEBS Lett., 141: 189-192, 1982.
19. Nakashima, R. A., Mangan, P. S., Colombini, M., and Pedersen. P. L.Hexokinase receptor complex in hepatoma mitochondria: evidence from N,/V'-dicyclohexylcarbodiimide-labeling studies for the involvement of the pore-forming protein VDAC. Biochemistry. 25: 1015-1021, 1986.
20. Bustamante, E., Soper, J. W., and Pedersen, P. L. A high-yield preparativemethod for isolation of rat liver mitochondria. Anal. Biochem., 50:401-408,1977.
21. Bergmeyer, H. U., Gawehn, K., and Grassi, M. Enzymes as biochemicalreagents. In: H. U. Bergmeyer (ed.). Methods of Enzymatic Analysis, Ed. 2,Vol. 1, pp. 425-522. New York: Academic Press, 1974.
22. Vassault, A. Láclatedehydrogenase: UV-method with pyruvate and NADH.In: H. U. Bergmeyer (ed.). Methods of Enzymatic Analysis, Ed. 3, Vol. 3,pp. 118-125. Deerfield Beach, FL: Verlag Chemie, 1983.
23. Fujii, H.. and Miwa, S. Pyruvate kinase: assay in serum and erythrocytes. In:H. U. Bergmeyer (ed.). Methods of Enzymatic Analysis, Ed. 3, Vol. 3, pp.496-501. Deerfield Beach, FI: Verlag Chemie, 1983.
24. Duruibe, V., and Tejwani, G. A. The effect of ethanol on the activities of thekey gluconeogenic and glycolytic enzymes of rat liver. Mol. Pharmacol., 20:621-630, 1981.
25. Laemmli, U. K. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature (Lond.), 227: 680-685, 1970.
26. Bidlingmeyer. B. A., Cohen, S. A., and Tarvin, T. L. Rapid analysis of aminoacids using pre-column derivatization. J. Chromatogr., 336:93-104, 1984.
27. Jacobs, E. E., Jacob, M., Sanadi. D. R.. and Bradley, L. B. Uncoupling ofoxidative phosphorylation by cadmium ion. J. Biol. Chem., 223: 147-156,1956.
28. Lowry, O. H., Rosebrough. N. J., Fair, A. L., and Randall. R. J. Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem., 193: 265-275,1951.
29. Saito, M., and Sato, S. Studies on particle-bound hexokinase in rat asciteshepatoma cells. Biochim. Biophys. Acta. 227: 344-353, 1971.
30. Katzen, H. M., and Schimkc. R. T. Multiple forms of hexokinase in the rat:tissue distribution, age dependency, and properties. Proc. Nati. Acad. Sci.USA, 54: 1218-1225, 1965.
31. Ureta, T. The comparative isozymology of vertebrate hexokinases. Comp.Biochem. Physiol., TIB: 549-555, 1982.
32. Holroyde, M. J., and Trayer, I. P. Purification and properties of rat skeletalmuscle hexokinase. FEBS Lett., 62: 215-219, 1976.
33. Chou, A. C., and Wilson, J. E. Purification and properties of rat brainhexokinase. Arch. Biochem. Biophys., 151:48-55, 1972.
34. Holroyde, M. J., Allen, M. B., Storer, A. C., Warsy, A. S., Chesher, J. M.E., Trayer, I. P., Cornish-Bowden, A., and Walker, D. G. The purificationin high yield and characterization of rat hepatic glucokinase. Biochem. J.,153: 363-373, 1976.
35. Gots, R. E., and Bessman, S. P. The functional compartmentation of mitochondrial hexokinase. Arch. Biochem. Biophys.. 163: 7-14, 1974.
36. Polakis, P. G., and Wilson, J. E. An intact hydrophobic Ai-terminal sequenceis critical for binding of rat brain hexokinase to mitochondria. Arch. Biochem.Biophys., 236: 328-337, 1985.
37. Radojkovic'. J., and Ureta, T. Hexokinase isozymes from the Novikoff
hepatoma. Purification, Kinetic and Structural Characterization with Emphasis on Hexokinase C. Biochem. J. 242:895-903, 1987.
919
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

1988;48:913-919. Cancer Res Richard A. Nakashima, Marco G. Paggi, Laura J. Scott, et al. AS-30D Rat Hepatoma Cell LineMitochondrial Bound Hexokinase from the Highly Glycolytic Purification and Characterization of a Bindable Form of
Updated version
http://cancerres.aacrjournals.org/content/48/4/913
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/48/4/913To request permission to re-use all or part of this article, use this link
on March 29, 2019. © 1988 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from