Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Putting Toe Clipping into Perspective: A Viable Method for Marking AnuransAuthor(s): T. Ulmar Grafe, Margaret M. Stewart, Kathrin P. Lampert, and Mark-Oliver RödelSource: Journal of Herpetology, 45(1):28-35. 2011.Published By: The Society for the Study of Amphibians and ReptilesDOI: http://dx.doi.org/10.1670/10-016.1URL: http://www.bioone.org/doi/full/10.1670/10-016.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

Putting Toe Clipping into Perspective: A Viable Method for Marking Anurans
T. ULMAR GRAFE,1,2 MARGARET M. STEWART,3 KATHRIN P. LAMPERT,1,4AND MARK-OLIVER RODEL
5
1Department of Animal Ecology and Tropical Biology, University of Wurzburg, Biozentrum, Am Hubland, 97074 Wurzburg, Germany3Department of Biological Sciences, State University of New York at Albany, Albany, New, York USA 12222
5Museum fur Naturkunde, Leibniz Institute for Research on Evolution and Biodiversity at the Humboldt University Berlin, Invalidenstr. 43,
10115 Berlin, Germany; E-mail: [email protected]
ABSTRACT.—Toe clipping is widely used in studies of amphibian ecology and behavior, but its impact on return rates and survival remains
controversial. We evaluated the effect of toe clipping on apparent survival, using four comprehensive mark–recapture data sets of four anuran
species covering four different time scales and varying life histories. The effect of toe clipping was evaluated by comparing frogs with different
numbers of toes removed. Two species, Eleutherodactylus coqui and Hemisus marmoratus, showed minimal effects of toe clipping on apparent
survival with seasonal and annual apparent survival decreasing by 0.1% and 1.5% with toe removal, respectively. In Hyperolius nitidulus, daily
apparent survival increased on average by 4.1% with toe removal, an effect than can be attributed to disproportionate emigration of the lowest
toe-removal group. Finally, in Phrynobatrachus guineensis, individuals with three toes clipped showed an additional 5.0–19.7% decreased
apparent survival between weeks when compared to individuals with only one toe clipped. However, the data set was characterized by an
interaction between the number of toes clipped and week of marking, thus confounding interpretation. To minimize any effects of toe clipping,
we recommend that not more than four toes should be clipped and at most a single toe removed on each leg. In addition, functionally important
toes such as the proximal toes of front feet and fourth toes of hind feet should be spared. Furthermore, toe-removal groups should be equally
distributed over time to facilitate the analysis of potential toe-clipping effects.
Estimation of demographic parameters such as survival andrecapture probabilities are essential for wildlife managementand conservation. Understanding population dynamics re-quires robust methods of marking individuals within popula-tions. With approximately one-third of all amphibian speciescurrently estimated to be in decline (Stuart et al., 2004), theneed to continue and establish new population monitoringprograms is large and will further increase. Toe clipping is awidely used method to mark anurans (Hero, 1989; Donnelly,1994); however, this method has come under scrutiny recently,because of claims that this method reduces the survival ofindividuals across all species and, thus, is unethical (May, 2004;McCarthy and Parris, 2004). This claim is based on a fewstudies that have shown the inability to detect effects of toeremoval because of low statistical power (Parris and McCarthy,2001). Furthermore, a rigorous Bayesian analysis showed lowreturn rates of individuals in which up to eight toes wereremoved (McCarthy and Parris, 2004). The resulting reserve bymany researchers to use toe clipping and the barriers set byethics committees have become a serious impediment foradvancement of our understanding of amphibian populationbiology.
Several studies have voiced opposition to the claim that toeremoval is generally harmful (Funk et al., 2005; Phillott et al.,2007), and this has led to wide spread debate on advantagesand disadvantages of toe clipping (Parris and McCarthy, 2008;Phillott et al., 2008). However, a rigorous analysis of mark–recapture data of toe-clipped anurans spanning different lifehistories, to complement the study by McCarthy and Parris(2004), has been lacking.
Here, we present a comprehensive analysis of four single-sitemark–recapture studies that used toe clipping for individualrecognition to evaluate the effects of toe removal on survival.The studies cover four time scales (daily, weekly, seasonal, andannual) and investigate species with four distinct life historieswith respect to lifespan, habitat, breeding sites, and breeding
system and belonging to four different anuran families Hemisusmarmoratus (Hemisotidae), Eleutherodactylus coqui (Leptodacty-lidae), Hyperolius nitidulus (Hyperoliidae) and Phrynobatrachusguineensis (Phrynobatrachidae) from two different geographicalrealms (Table 1). Thus, our analysis covers a wide range offunctional traits in anuran species, all of which are ofteninvestigated using mark–recapture methodology; thus, resultswill likely be applicable to many other amphibian species.
MATERIALS AND METHODS
Study Sites and Species.—Eleutherodactylus coqui is a terrestrial-breeding direct-developing frog that defends small territoriesin the forest understory and forages in the canopy (Stewart,1995; Stewart and Woolbright, 1996). Data for this analysiswere obtained from a six-year single-site (‘‘River House Plot’’)mark–recapture study conducted in the tabanuco forest zone ofthe Luquillo Experimental Forest in northeastern Puerto Rico(18u229N, 65u529W; Stewart, 1995). Adult males and femaleswere toe clipped using the scheme outlined in Hero (1989) withat most one toe removed on each foot but excluding fourth toeson hind limbs. In total 1, 2, 3, and 4 toes were removed from 16,92, 221, and 191 individuals, respectively. Individuals wererandomly assigned to one of the toe-removal groups. Malesand females were measured for growth rates, longevity, andreproductive behavior between January 1983 and January 1988resulting in a total of 11 sampling periods. Frogs weremonitored biannually over periods of 1–2 months in both thewet and dry seasons in a 10 3 10 m plot that had beensupplemented with small bamboo houses that were used byadults as diurnal retreat sites (Stewart and Pough, 1983).Sampling of the plot was conducted during day and nighthours and involved systematic searches of leaf litter, bamboohouses and other potential retreat sites. Two animals foundwith legs missing were deleted from further analysis. Toestimate the effects of toe clipping on seasonal apparentsurvival, individuals were assigned to one of four groupsdepending on the number of toes removed (1, 2, 3, or 4).
Hemisus marmoratus is a burrowing/fossorial frog withreproductive activity highly synchronized with rains (Grafeet al., 2004). Females construct underground nests in whicheggs are fertilized (Rodel et al., 1995; Kaminsky et al., 1999).Data for analysis were taken from a demographic study
2 Corresponding Author. Present address: Department of Biology,University Brunei Darussalam, Tungku Link, Gadong BE 1410, BruneiDarussalam; E-mail: [email protected]
4 Present address: Department of Animal Ecology, Evolution andBiodiversity, Ruhr-Universitat Bochum, 44780 Bochum, Germany; E-mail: [email protected]
Journal of Herpetology, Vol. 45, No. 1, pp. 28–35, 2011Copyright 2011 Society for the Study of Amphibians and Reptiles

conducted over six years between 1997 and 2002 in thesouthern Guinea savanna region of the Comoe National Park(8u459N, 3u499W; for details, see Grafe et al., 2004). Surveyswere conducted on 48 rain periods each covering 1–3 nights.Male and female Pig-Nosed Frogs were collected duringnightly surveys at a single temporary pond (‘‘Lola’’) along a160-m drift fence that completely encircled the pond. Toes wereremoved from 821 males and females using the scheme of Hero(1989). At most, one toe was removed on each foot resulting ina maximum of four toes removed. In total 1, 2, 3, and 4 toeswere removed from 18, 130, 351, and 323 individuals,respectively. Individuals were randomly assigned to one ofthe toe-removal groups. Proximal toes on forelimbs(‘‘thumbs’’) and fourth toes on hind limbs were not removedbecause they are likely to have specific functions such as aidingamplexus and facilitating molting, respectively. Annual ap-parent survival was estimated and compared between the fourtoe-removal groups.
The Reed Frog Hyperolius nitidulus is a common denizen ofthe West African savanna and well known for its remarkableaestivation behavior and physiology (Linsenmair, 1998). Malesaggregate at temporary ponds during the 4–6-month rainyseason. Mark–recaptures were conducted in the ComoeNational Park, Ivory Coast. A total of 179 males were markedby toe clipping for demographic analysis during the 1998breeding season on 14 nights between 1 August and 16 October(Lampert and Linsenmair, 2002). One to four toes wereremoved using the scheme of Hero (1989) at a single 50 320 m large savanna pond (‘‘Hyperolius Pond’’). As in H.mamoratus, fourth toes on hind limbs were not removed;however, thumbs were clipped. Individuals with only one toeremoved were too few to be included in the analysis. Overall, 2,3, and 4 toes were removed from 22, 73, and 88 individuals,respectively. Thus, apparent survival was estimated for onlythree toe-removal groups corresponding to the loss of 2, 3 or 4toes. More individuals had two toes removed earlier than laterin the study season. Survival was estimated on a daily basis, anappropriate time scale because in this species adults live onlyfor one breeding season (Lampert and Linsenmair, 2002).
Phrynobatrachus guineensis is a rain-forest litter frog, restrictedto the Upper Guinea forests of West Africa that breeds inphytotelmata (Rodel et al., 2004; Rudolf and Rodel, 2005). Datafor analysis of the effects of toe clipping on survival wereobtained from 156 males and females marked over a 29-weekperiod from 9 March to 27 September 2002 (Rudolf and Rodel,2005, 2007). This demographic study was conducted in the TaıNational Park, Ivory Coast, at the ‘‘Station de Recherche enEcologie Tropical’’ (5u50.0039N, 7u20.5369W). Frogs weremonitored at 89 natural and 57 artificial breeding sites (treeholes, fruit capsules, or empty snail shells) distributed on theforest understory up to 2 m high. Males are known tosometimes move between close-by breeding sites (meandistance: 24 m; max. distance: 94 m; Rodel et al., 2004). Siteattendance varied between 1 and 23 nights. In contrast to theother data sets used in this analysis, only toes of hind feet andup to two toes per foot were removed. Only the top 2–3 digits(i.e., the distal and middle phalanges) of a toe were removed.
In total, 1, 2, and 3 toe tips were removed from 9, 45, and 102individuals, respectively. Frogs were monitored on a dailybasis. To avoid over-parameterized models, we pooledrecaptures by week. Weekly apparent survival and recaptureprobabilities were estimated for three toe-removal groupscorresponding to the loss of 1, 2, or 3 toes. The toe-removalgroups were not equally distributed throughout the studyperiod.
Demographic Analyses.—Mark–recapture data of individuallytoe-clipped frogs was analyzed using Cormack-Jolly-Seber(CJS) models (Lebreton et al., 1992). These live-encounter datawere used to derive separate estimates of apparent survival (Q)and recapture probabilities (p). The fully time-dependent CJSmodel makes some fundamental assumptions (Pollack et al.,1990). First, marks are not lost or overlooked. Second, samplesare instantaneous compared to sample intervals. Third, individ-uals of the ith sample have the same p. Fourth, individuals of theith sample have the same probability of surviving to i + 1. Thefour data sets used in this analysis of effects of toe clipping onsurvival were collected by different researchers; thus, someheterogeneity in methodology is natural. Details of study designand implementation are given above under species headings.
Our first step was to evaluate the four assumptions of theCJS model for each study. First, toe clipping was permanent inH. marmoratus and P. guineensis (Grafe et al., 2004; Rodel et al.,2004). Although toes regenerate in Hyperolius (Richards et al.,1975) and E. coqui (Stewart and Woolbright, 1996), regeneratedsecondary toes can be readily distinguished from primarydigits. In addition, in Hyperolius, the study period was too shortfor toe regeneration to take effect. Furthermore, all studiesemployed toe clipping only after mark identification had beenpracticed in pilot studies, and validity was cross-checked byseveral investigators over time. Thus, mark overlook ormisidentification was likely negligible in all four studies.Second, frogs were released immediately after marking in eachstudy fulfilling our second assumption. We used the programU-Care (vers. 2.3.1) to test for any departures of the data fromour third and fourth assumptions by performing goodness-of-fit tests (GOF; Choquet et al., 2003). The test results arereported below for each species. Overall, transience and trap-dependence were limited and were not further incorporated inthe models. The data were also evaluated for overdispersal invariance estimation using U-Care by calculating the varianceinflation factor c. No evidence for marked overdispersion wasdetected in any of the four data sets (c 5 0.77, 0.90, 0.62, and1.12 for E. coqui, H. marmoratus, H. nitidulus, and P. guineensis,respectively). Fates of all individuals were independent; thus, cwas not adjusted.
We used model selection by fitting progressively simplermodels by minimizing the Akaike’s information criterioncorrected for effective sample size (AICc; Anderson andBurnham, 1999) using the program MARK (vers 6.0; Whiteand Burnham, 1999). We modelled the effect of toe clipping onapparent survival and recapture probabilities by groupingindividuals according to the number of toes lost. Because wecould not follow individuals without toe clipping them, ourevaluation of the effect of toe clipping is based on comparing
TABLE 1. Anuran species investigated for the effects of toe removal on survival.
Species/family Habitat/location Time scale Life history
Eleutherodactylus coqui;Leptodactylidae
Neotropical rain forest/Puerto Rico
6 yr Lifespan of several years, direct development,territorial, terestrial, arboreal
Hemisus marmoratus;Hemisotidae
Afro-tropical savanna/Ivory Coast
6 yr Lifespan of several years, eggs develop inunderground nests, aquatic tadpole, fossorial
Hyperolius nitidulus;Hyperoliidae
Afro-tropical savanna/Ivory Coast
11 weeks Maximum 1-yr lifespan, lek mating, eggs andtadpoles develop in ephemeral ponds, arboreal
Phrynobatrachus guineensis;Phrynobatrachidae
Afro-tropical rain forest/Ivory Coast
29 weeks Lifespan of less than 1 yr, phytotelmata breeder,aquatic tadpoles, terrestrial, arboreal
VIABLE METHOD FOR MARKING ANURANS 29

the survival as a function of the number of toes clipped. Thisassumes that mortality increases with the number of digits lostas some studies indicate (Waddle et al., 2008). Our study doesnot address whether toe clipping a single toe decreasessurvival compared to individuals that have not lost toes.
To identify the best fitting model, we employed themethodology outlined by Cooch and White (2006). A list ofcandidate models to explain variation in survival between toe-removal groups was generated for each of the four investigatedspecies. Candidate models included the effects of time, toeremoval, and rainfall on apparent survival and recapture.Because rainfall was previously identified as contributing tosurvival in H. marmoratus (Grafe et al., 2004), we enteredannual rainfall into the models as an external covariate ofsurvival by forcing the time-dependent survival rates to beestimated as linear functions of rainfall.
The model with the lowest AICc-value, when DAICc . 2, wasused as the criteria for identifying the best model (Burnham andAnderson, 2002). When DAICc ,2 survival estimates and SEswere weighted by the AICc weights of the candidate models tocounter model uncertainty. This model selection criterion hasrecently gained significant support in ecology and evolution(Johnson and Omland, 2004).
RESULTS
Eleutherodactylus coqui.—We captured and marked 520 frogs ofwhich 182 were recaptured at least once between seasons: a 35%seasonal return rate. GOF tests indicated no trap dependencebetween years (N[0,1] statistic for trap dependence 5 20.587, P 50.557). Likewise, tests of survival homogeneity or transienceshowed that newly marked and previously marked frogs wereequally likely to survive (N[0,1] statistic for transients 5 21.8614,P 5 0.063).
Our general model had separate survival parameters andcapture probabilities for each of the four toe-removal groups foreach capture occasion (Q[g 3 t], p[g 3 t]) and fit the data (GOF:x2
47 5 36.09; P 5 0.876). We used this model as a starting point toinvestigate the effects of toe clipping on survival and recaptureprobabilities.
The most parsimonious model had time-dependent survivaland constant recapture probabilities (model 1; Table 2) andwas 2.3 times better supported than the second best model thathad toe-removal dependent recapture probabilities. None ofthe five best supported models included toe-removal effects onapparent survival. These five best models explained 99% of thevariation. The average apparent seasonal survival estimates ofthe toe-removal groups based on the best model are shown inFigure 1. Over all sample periods, seasonal survival averaged99.5% for toe-removal groups 1–3 and 99.4% for toe-removalgroup 4. Thus, the decrease in apparent survival was 0.1%from individuals that had 1, 2, or 3 toes removed to those that
had 4 toes removed. Apparent seasonal survival estimateswere very high ranging between 99.0% and 99.7%. Apparentsurvival did not differ significantly between dry and wetseasons.
There was also no support for a relationship betweennumber of toes removed and recapture probability based onthe low AIC weights for models including such an effect.Models that included a toe-removal effect on recaptureprobability received only 24.3% of the AICc weight. Themodel-averaged estimates of recapture probabilities variedlittle between toe-removal groups (range 0.65–0.72).
Hemisus marmoratus.—We toe clipped a total of 821 individ-uals over six years. Seventy-four males and females wererecaptured between years resulting in a 9.4% annual returnrate. GOF testing showed no trap dependence between years(N[0,1] statistic for trap dependence 5 21.458, P 5 0.145) ortransience (N[0,1] statistic for transients 5 20.125, P 5 0.900)indicating that our third and fourth assumptions of the CJSmodel were fulfilled.
Our general model had separate survival parameters andcapture probabilities for each of the four toe-removal groupsfor each annual capture occasion (Q[g 3 t], p[g 3 t]), and it fitthe data (GOF: x2
19 5 17.06; P 5 0.586). We used this model asa starting point to investigate the effects of toe clipping onapparent survival and recapture probabilities.
We identified 12 candidate models to investigate the effectsof toe removal on survival (Table 3). Models with time-dependent, constant, or rain-constrained apparent survivalhad the most support (models 1–3, Table 3), whereas toe-specific survival was not highly supported (model 4, Table 3).The best model had time-dependent survival (model 1, AICc 5
628.62) and was 6.4 times better supported than the best toe-specific model (model 4, AICc 5 635.02). Estimated apparent
TABLE 2. The 10 candidate models of seasonal apparent survival (Q) and recapture probabilities (p) in Eleutherodactylus coqui based on a single-site6-yr study in Puerto Rico. AICc 5 Estimated Akaike’s information criterion (lower values indicate more parsimonious models), DAICc 5 differencein AICc between the current model and the top model (model 1), t 5 full time-dependence, g 5 toe-specific variation, . 5 constant.
Model AICc DAICc Weight K Deviance
1 Q (t) p(.) 1,136.34 0.00 0.734 11 185.022 Q (t) p(g) 1,138.60 2.25 0.238 14 181.053 Q (.) p(.) 1,144.77 8.42 0.011 2 211.804 Q (.) p(t) 1,146.23 9.89 0.005 11 194.915 Q (.) p(g) 1,146.54 10.20 0.004 5 207.516 Q (g) p(.) 1,147.73 11.38 0.002 5 208.697 Q (t) p(t) 1,148.75 12.40 0.001 19 180.718 Q (g) p(g) 1,148.89 12.54 0.001 8 203.739 Q (g) p(t) 1,149.03 12.69 0.001 14 191.4810 Q (g 3 t) p(g 3 t) 1,190.81 54.47 0.000 62 125.92
FIG. 1. Mean seasonal apparent survival and the unconditional 95%confidence intervals of adult Eleutherodactylus coqui for each of four toe-removal groups.
30 T. U. GRAFE ET AL.
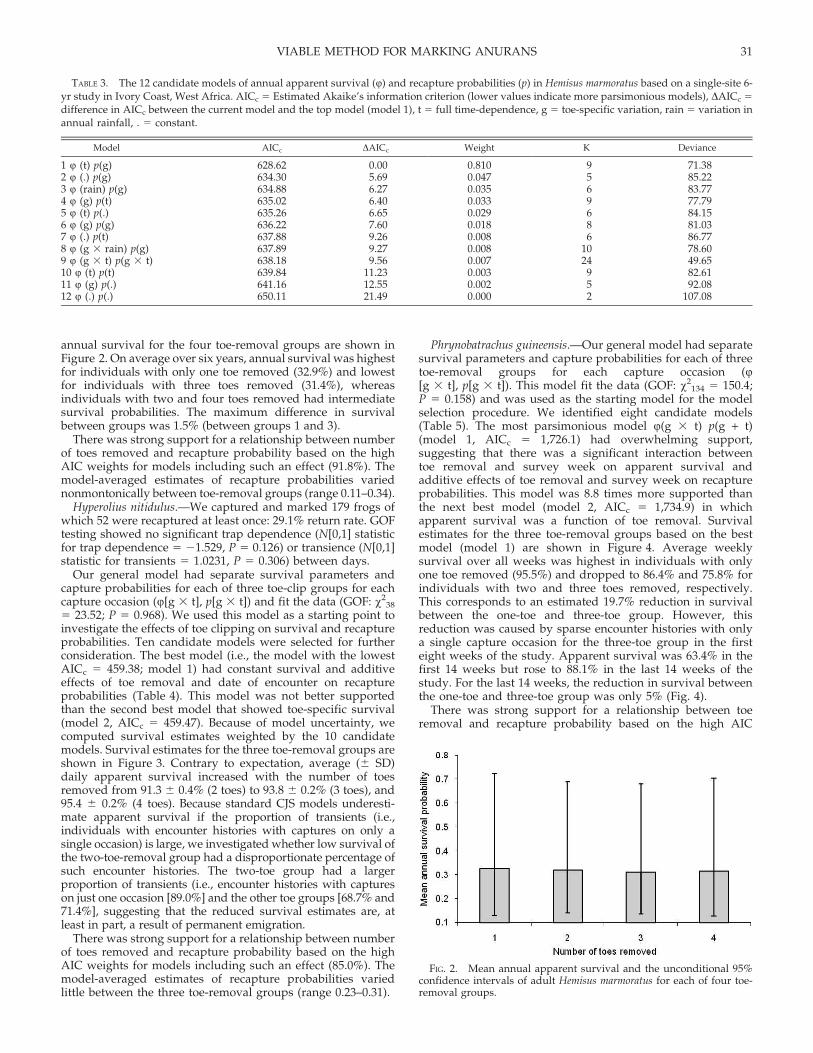
annual survival for the four toe-removal groups are shown inFigure 2. On average over six years, annual survival was highestfor individuals with only one toe removed (32.9%) and lowestfor individuals with three toes removed (31.4%), whereasindividuals with two and four toes removed had intermediatesurvival probabilities. The maximum difference in survivalbetween groups was 1.5% (between groups 1 and 3).
There was strong support for a relationship between numberof toes removed and recapture probability based on the highAIC weights for models including such an effect (91.8%). Themodel-averaged estimates of recapture probabilities variednonmontonically between toe-removal groups (range 0.11–0.34).
Hyperolius nitidulus.—We captured and marked 179 frogs ofwhich 52 were recaptured at least once: 29.1% return rate. GOFtesting showed no significant trap dependence (N[0,1] statisticfor trap dependence 5 21.529, P 5 0.126) or transience (N[0,1]statistic for transients 5 1.0231, P 5 0.306) between days.
Our general model had separate survival parameters andcapture probabilities for each of three toe-clip groups for eachcapture occasion (Q[g 3 t], p[g 3 t]) and fit the data (GOF: x2
38
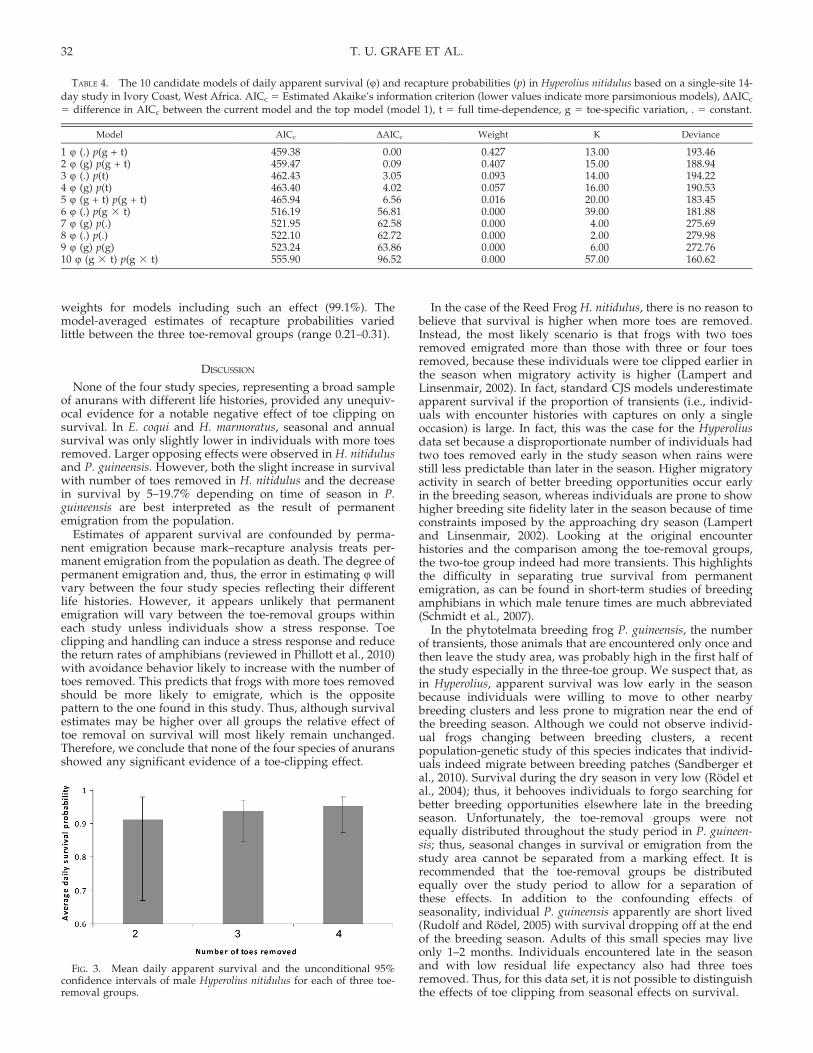
5 23.52; P 5 0.968). We used this model as a starting point toinvestigate the effects of toe clipping on survival and recaptureprobabilities. Ten candidate models were selected for furtherconsideration. The best model (i.e., the model with the lowestAICc 5 459.38; model 1) had constant survival and additiveeffects of toe removal and date of encounter on recaptureprobabilities (Table 4). This model was not better supportedthan the second best model that showed toe-specific survival(model 2, AICc 5 459.47). Because of model uncertainty, wecomputed survival estimates weighted by the 10 candidatemodels. Survival estimates for the three toe-removal groups areshown in Figure 3. Contrary to expectation, average (6 SD)daily apparent survival increased with the number of toesremoved from 91.3 6 0.4% (2 toes) to 93.8 6 0.2% (3 toes), and95.4 6 0.2% (4 toes). Because standard CJS models underesti-mate apparent survival if the proportion of transients (i.e.,individuals with encounter histories with captures on only asingle occasion) is large, we investigated whether low survival ofthe two-toe-removal group had a disproportionate percentage ofsuch encounter histories. The two-toe group had a largerproportion of transients (i.e., encounter histories with captureson just one occasion [89.0%] and the other toe groups [68.7% and71.4%], suggesting that the reduced survival estimates are, atleast in part, a result of permanent emigration.
There was strong support for a relationship between numberof toes removed and recapture probability based on the highAIC weights for models including such an effect (85.0%). Themodel-averaged estimates of recapture probabilities variedlittle between the three toe-removal groups (range 0.23–0.31).
Phrynobatrachus guineensis.—Our general model had separatesurvival parameters and capture probabilities for each of threetoe-removal groups for each capture occasion (Q[g 3 t], p[g 3 t]). This model fit the data (GOF: x2
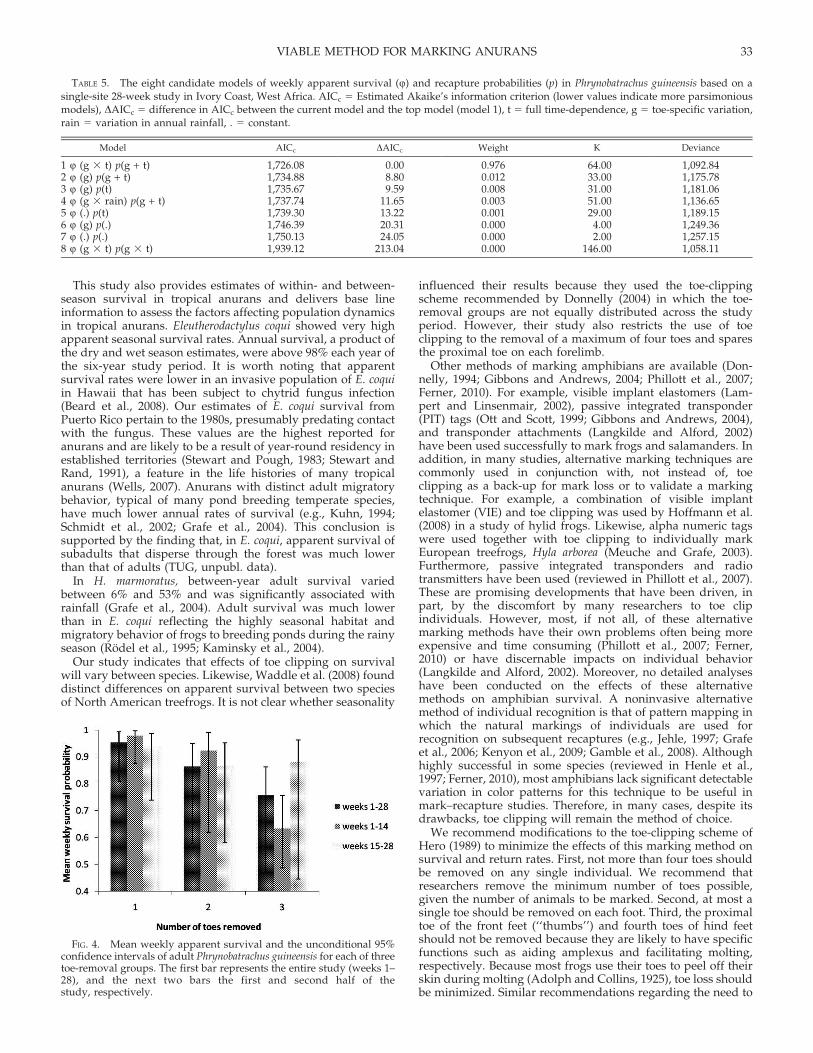
134 5 150.4;P 5 0.158) and was used as the starting model for the modelselection procedure. We identified eight candidate models(Table 5). The most parsimonious model Q(g 3 t) p(g + t)(model 1, AICc 5 1,726.1) had overwhelming support,suggesting that there was a significant interaction betweentoe removal and survey week on apparent survival andadditive effects of toe removal and survey week on recaptureprobabilities. This model was 8.8 times more supported thanthe next best model (model 2, AICc 5 1,734.9) in whichapparent survival was a function of toe removal. Survivalestimates for the three toe-removal groups based on the bestmodel (model 1) are shown in Figure 4. Average weeklysurvival over all weeks was highest in individuals with onlyone toe removed (95.5%) and dropped to 86.4% and 75.8% forindividuals with two and three toes removed, respectively.This corresponds to an estimated 19.7% reduction in survivalbetween the one-toe and three-toe group. However, thisreduction was caused by sparse encounter histories with onlya single capture occasion for the three-toe group in the firsteight weeks of the study. Apparent survival was 63.4% in thefirst 14 weeks but rose to 88.1% in the last 14 weeks of thestudy. For the last 14 weeks, the reduction in survival betweenthe one-toe and three-toe group was only 5% (Fig. 4).
There was strong support for a relationship between toeremoval and recapture probability based on the high AIC
TABLE 3. The 12 candidate models of annual apparent survival (Q) and recapture probabilities (p) in Hemisus marmoratus based on a single-site 6-yr study in Ivory Coast, West Africa. AICc 5 Estimated Akaike’s information criterion (lower values indicate more parsimonious models), DAICc 5
difference in AICc between the current model and the top model (model 1), t 5 full time-dependence, g 5 toe-specific variation, rain 5 variation inannual rainfall, . 5 constant.
Model AICc DAICc Weight K Deviance
1 Q (t) p(g) 628.62 0.00 0.810 9 71.382 Q (.) p(g) 634.30 5.69 0.047 5 85.223 Q (rain) p(g) 634.88 6.27 0.035 6 83.774 Q (g) p(t) 635.02 6.40 0.033 9 77.795 Q (t) p(.) 635.26 6.65 0.029 6 84.156 Q (g) p(g) 636.22 7.60 0.018 8 81.037 Q (.) p(t) 637.88 9.26 0.008 6 86.778 Q (g 3 rain) p(g) 637.89 9.27 0.008 10 78.609 Q (g 3 t) p(g 3 t) 638.18 9.56 0.007 24 49.6510 Q (t) p(t) 639.84 11.23 0.003 9 82.6111 Q (g) p(.) 641.16 12.55 0.002 5 92.0812 Q (.) p(.) 650.11 21.49 0.000 2 107.08
FIG. 2. Mean annual apparent survival and the unconditional 95%confidence intervals of adult Hemisus marmoratus for each of four toe-removal groups.
VIABLE METHOD FOR MARKING ANURANS 31
weights for models including such an effect (99.1%). Themodel-averaged estimates of recapture probabilities variedlittle between the three toe-removal groups (range 0.21–0.31).
DISCUSSION
None of the four study species, representing a broad sampleof anurans with different life histories, provided any unequiv-ocal evidence for a notable negative effect of toe clipping onsurvival. In E. coqui and H. marmoratus, seasonal and annualsurvival was only slightly lower in individuals with more toesremoved. Larger opposing effects were observed in H. nitidulusand P. guineensis. However, both the slight increase in survivalwith number of toes removed in H. nitidulus and the decreasein survival by 5–19.7% depending on time of season in P.guineensis are best interpreted as the result of permanentemigration from the population.
Estimates of apparent survival are confounded by perma-nent emigration because mark–recapture analysis treats per-manent emigration from the population as death. The degree ofpermanent emigration and, thus, the error in estimating Q willvary between the four study species reflecting their differentlife histories. However, it appears unlikely that permanentemigration will vary between the toe-removal groups withineach study unless individuals show a stress response. Toeclipping and handling can induce a stress response and reducethe return rates of amphibians (reviewed in Phillott et al., 2010)with avoidance behavior likely to increase with the number oftoes removed. This predicts that frogs with more toes removedshould be more likely to emigrate, which is the oppositepattern to the one found in this study. Thus, although survivalestimates may be higher over all groups the relative effect oftoe removal on survival will most likely remain unchanged.Therefore, we conclude that none of the four species of anuransshowed any significant evidence of a toe-clipping effect.
In the case of the Reed Frog H. nitidulus, there is no reason tobelieve that survival is higher when more toes are removed.Instead, the most likely scenario is that frogs with two toesremoved emigrated more than those with three or four toesremoved, because these individuals were toe clipped earlier inthe season when migratory activity is higher (Lampert andLinsenmair, 2002). In fact, standard CJS models underestimateapparent survival if the proportion of transients (i.e., individ-uals with encounter histories with captures on only a singleoccasion) is large. In fact, this was the case for the Hyperoliusdata set because a disproportionate number of individuals hadtwo toes removed early in the study season when rains werestill less predictable than later in the season. Higher migratoryactivity in search of better breeding opportunities occur earlyin the breeding season, whereas individuals are prone to showhigher breeding site fidelity later in the season because of timeconstraints imposed by the approaching dry season (Lampertand Linsenmair, 2002). Looking at the original encounterhistories and the comparison among the toe-removal groups,the two-toe group indeed had more transients. This highlightsthe difficulty in separating true survival from permanentemigration, as can be found in short-term studies of breedingamphibians in which male tenure times are much abbreviated(Schmidt et al., 2007).
In the phytotelmata breeding frog P. guineensis, the numberof transients, those animals that are encountered only once andthen leave the study area, was probably high in the first half ofthe study especially in the three-toe group. We suspect that, asin Hyperolius, apparent survival was low early in the seasonbecause individuals were willing to move to other nearbybreeding clusters and less prone to migration near the end ofthe breeding season. Although we could not observe individ-ual frogs changing between breeding clusters, a recentpopulation-genetic study of this species indicates that individ-uals indeed migrate between breeding patches (Sandberger etal., 2010). Survival during the dry season in very low (Rodel etal., 2004); thus, it behooves individuals to forgo searching forbetter breeding opportunities elsewhere late in the breedingseason. Unfortunately, the toe-removal groups were notequally distributed throughout the study period in P. guineen-sis; thus, seasonal changes in survival or emigration from thestudy area cannot be separated from a marking effect. It isrecommended that the toe-removal groups be distributedequally over the study period to allow for a separation ofthese effects. In addition to the confounding effects ofseasonality, individual P. guineensis apparently are short lived(Rudolf and Rodel, 2005) with survival dropping off at the endof the breeding season. Adults of this small species may liveonly 1–2 months. Individuals encountered late in the seasonand with low residual life expectancy also had three toesremoved. Thus, for this data set, it is not possible to distinguishthe effects of toe clipping from seasonal effects on survival.
TABLE 4. The 10 candidate models of daily apparent survival (Q) and recapture probabilities (p) in Hyperolius nitidulus based on a single-site 14-day study in Ivory Coast, West Africa. AICc 5 Estimated Akaike’s information criterion (lower values indicate more parsimonious models), DAICc
5 difference in AICc between the current model and the top model (model 1), t 5 full time-dependence, g 5 toe-specific variation, . 5 constant.
Model AICc DAICc Weight K Deviance
1 Q (.) p(g + t) 459.38 0.00 0.427 13.00 193.462 Q (g) p(g + t) 459.47 0.09 0.407 15.00 188.943 Q (.) p(t) 462.43 3.05 0.093 14.00 194.224 Q (g) p(t) 463.40 4.02 0.057 16.00 190.535 Q (g + t) p(g + t) 465.94 6.56 0.016 20.00 183.456 Q (.) p(g 3 t) 516.19 56.81 0.000 39.00 181.887 Q (g) p(.) 521.95 62.58 0.000 4.00 275.698 Q (.) p(.) 522.10 62.72 0.000 2.00 279.989 Q (g) p(g) 523.24 63.86 0.000 6.00 272.7610 Q (g 3 t) p(g 3 t) 555.90 96.52 0.000 57.00 160.62
FIG. 3. Mean daily apparent survival and the unconditional 95%confidence intervals of male Hyperolius nitidulus for each of three toe-removal groups.
32 T. U. GRAFE ET AL.
This study also provides estimates of within- and between-season survival in tropical anurans and delivers base lineinformation to assess the factors affecting population dynamicsin tropical anurans. Eleutherodactylus coqui showed very highapparent seasonal survival rates. Annual survival, a product ofthe dry and wet season estimates, were above 98% each year ofthe six-year study period. It is worth noting that apparentsurvival rates were lower in an invasive population of E. coquiin Hawaii that has been subject to chytrid fungus infection(Beard et al., 2008). Our estimates of E. coqui survival fromPuerto Rico pertain to the 1980s, presumably predating contactwith the fungus. These values are the highest reported foranurans and are likely to be a result of year-round residency inestablished territories (Stewart and Pough, 1983; Stewart andRand, 1991), a feature in the life histories of many tropicalanurans (Wells, 2007). Anurans with distinct adult migratorybehavior, typical of many pond breeding temperate species,have much lower annual rates of survival (e.g., Kuhn, 1994;Schmidt et al., 2002; Grafe et al., 2004). This conclusion issupported by the finding that, in E. coqui, apparent survival ofsubadults that disperse through the forest was much lowerthan that of adults (TUG, unpubl. data).
In H. marmoratus, between-year adult survival variedbetween 6% and 53% and was significantly associated withrainfall (Grafe et al., 2004). Adult survival was much lowerthan in E. coqui reflecting the highly seasonal habitat andmigratory behavior of frogs to breeding ponds during the rainyseason (Rodel et al., 1995; Kaminsky et al., 2004).
Our study indicates that effects of toe clipping on survivalwill vary between species. Likewise, Waddle et al. (2008) founddistinct differences on apparent survival between two speciesof North American treefrogs. It is not clear whether seasonality
influenced their results because they used the toe-clippingscheme recommended by Donnelly (2004) in which the toe-removal groups are not equally distributed across the studyperiod. However, their study also restricts the use of toeclipping to the removal of a maximum of four toes and sparesthe proximal toe on each forelimb.
Other methods of marking amphibians are available (Don-nelly, 1994; Gibbons and Andrews, 2004; Phillott et al., 2007;Ferner, 2010). For example, visible implant elastomers (Lam-pert and Linsenmair, 2002), passive integrated transponder(PIT) tags (Ott and Scott, 1999; Gibbons and Andrews, 2004),and transponder attachments (Langkilde and Alford, 2002)have been used successfully to mark frogs and salamanders. Inaddition, in many studies, alternative marking techniques arecommonly used in conjunction with, not instead of, toeclipping as a back-up for mark loss or to validate a markingtechnique. For example, a combination of visible implantelastomer (VIE) and toe clipping was used by Hoffmann et al.(2008) in a study of hylid frogs. Likewise, alpha numeric tagswere used together with toe clipping to individually markEuropean treefrogs, Hyla arborea (Meuche and Grafe, 2003).Furthermore, passive integrated transponders and radiotransmitters have been used (reviewed in Phillott et al., 2007).These are promising developments that have been driven, inpart, by the discomfort by many researchers to toe clipindividuals. However, most, if not all, of these alternativemarking methods have their own problems often being moreexpensive and time consuming (Phillott et al., 2007; Ferner,2010) or have discernable impacts on individual behavior(Langkilde and Alford, 2002). Moreover, no detailed analyseshave been conducted on the effects of these alternativemethods on amphibian survival. A noninvasive alternativemethod of individual recognition is that of pattern mapping inwhich the natural markings of individuals are used forrecognition on subsequent recaptures (e.g., Jehle, 1997; Grafeet al., 2006; Kenyon et al., 2009; Gamble et al., 2008). Althoughhighly successful in some species (reviewed in Henle et al.,1997; Ferner, 2010), most amphibians lack significant detectablevariation in color patterns for this technique to be useful inmark–recapture studies. Therefore, in many cases, despite itsdrawbacks, toe clipping will remain the method of choice.
We recommend modifications to the toe-clipping scheme ofHero (1989) to minimize the effects of this marking method onsurvival and return rates. First, not more than four toes shouldbe removed on any single individual. We recommend thatresearchers remove the minimum number of toes possible,given the number of animals to be marked. Second, at most asingle toe should be removed on each foot. Third, the proximaltoe of the front feet (‘‘thumbs’’) and fourth toes of hind feetshould not be removed because they are likely to have specificfunctions such as aiding amplexus and facilitating molting,respectively. Because most frogs use their toes to peel off theirskin during molting (Adolph and Collins, 1925), toe loss shouldbe minimized. Similar recommendations regarding the need to
TABLE 5. The eight candidate models of weekly apparent survival (Q) and recapture probabilities (p) in Phrynobatrachus guineensis based on asingle-site 28-week study in Ivory Coast, West Africa. AICc 5 Estimated Akaike’s information criterion (lower values indicate more parsimoniousmodels), DAICc 5 difference in AICc between the current model and the top model (model 1), t 5 full time-dependence, g 5 toe-specific variation,rain 5 variation in annual rainfall, . 5 constant.
Model AICc DAICc Weight K Deviance
1 Q (g 3 t) p(g + t) 1,726.08 0.00 0.976 64.00 1,092.842 Q (g) p(g + t) 1,734.88 8.80 0.012 33.00 1,175.783 Q (g) p(t) 1,735.67 9.59 0.008 31.00 1,181.064 Q (g 3 rain) p(g + t) 1,737.74 11.65 0.003 51.00 1,136.655 Q (.) p(t) 1,739.30 13.22 0.001 29.00 1,189.156 Q (g) p(.) 1,746.39 20.31 0.000 4.00 1,249.367 Q (.) p(.) 1,750.13 24.05 0.000 2.00 1,257.158 Q (g 3 t) p(g 3 t) 1,939.12 213.04 0.000 146.00 1,058.11
FIG. 4. Mean weekly apparent survival and the unconditional 95%confidence intervals of adult Phrynobatrachus guineensis for each of threetoe-removal groups. The first bar represents the entire study (weeks 1–28), and the next two bars the first and second half of thestudy, respectively.
VIABLE METHOD FOR MARKING ANURANS 33

evaluate toe use have been made by ethics committees (e.g.,HACC, 2004). Finally, we recommend the use of a markingscheme in which toe-removal groups are equally distributedacross time. This will facilitate later analysis of effect ofremoval on survival. The P. guineensis data set presented hereemphasizes this point. It was not possible to distinguishseasonal variation in survival from toe-removal effects in thisdata set because the toe-removal groups were not equallydistributed across the duration of each study. Thus, variationbetween toe-removal groups in apparent survival is likely tohave been confounded by variation in residual life expectancy.
Excessive amount of toe clipping in some studies (McCarthyand Parris, 2004) have led to the conclusion that toe clipping isgenerally problematic. However, many researchers have usedtoe clipping in a responsible way over the past decades byminimizing the number of toes cut, reusing numbers insituations where it is very unlikely that individuals previouslymarked with that code were still alive and recognizing that sometoes are likely to be more useful than others for survival andreproduction.
The use of mark–recapture methodology to estimate survivaland recapture probabilities is a powerful and robust tool tounderstand the effects of marking methods. The importance ofinvestigating the consequences of variation in survival forpopulation dynamics and life-history characteristics are undis-puted and the marking of individual animals will remain anecessary prerequisite. In the absence of alternative markingmethods, careful use of toe removal will prove instrumental inadvancing amphibian conservation and management.
Acknowledgments.—The P. guineensis data set was collected aspart of the BIOLOG program of the German Ministry ofEducation and Science (BMBF; BIOTA-West amphibian pro-jects, 01 LC0410). Lodging facilities in Taı National Park (TNP)were provided by the ‘‘Centre de Recherche en Ecologie.’’Research permission was given by the ‘‘Ministere de l’En-seignement Superieur et de la Recherche Scientifique’’ of theRepublic of Ivory Coast. The access permit to TNP was issuedby the ‘‘Ministere de la Construction et de l’Environnement.’’V. H. W. Rudolf, G. G. Gbamlin, and C. Y. Ouoro were ofinvaluable help with fieldwork. The H. nitidulus part of thestudy was supported by the Deutsche Forschungsgemeinschaft(Research Grant Li 150/19-1). We thank the government of theIvory Coast for granting the necessary research permits tostudy H. marmoratus. For help with fieldwork, we thank M.Boutros, A. Dill, M. Kaltenpoth, K. Kouadio, C. Meyer, M.Mosl, C. Muller, and A. Schmidt. This study was supported bygrants from the Deutsche Forschungsgemeinschaft (SFB 251/B3) and a seed grant from the Declining Amphibian PopulationsTask Force (DAPTF). Finally, the E. coqui demographic data wascollected thanks to a National Science Foundation grant (DEB 77-211349). Logistical support was provided by the U.S. Departmentof Agriculture and the University of Puerto Rico. Numerousstudents assisted with fieldwork, in particular L. Woolbright andD. Townsend. TUG, KPL, and M-OR dedicate this paper to thememory of Margaret Stewart.
LITERATURE CITED
ADOLPF, E. F., AND H. H. COLLINS. 1925. Molting in an amphibian,Diemyctylus. Journal of Morphology 40:575.
ANDERSON, D. R., AND K. P. BURNHAM. 1999. General strategies for theanalysis of ringing data. Bird Study 46 (Suppl):14–21.
BEARD, K. H., R. AL-CHOKHACHY, N. C. TUTTLE, AND E. M. O’NEILL. 2008.Population density estimates and growth rates of Eleutherodactyluscoqui in Hawaii. Journal of Herpetology 42:626–636.
CHOQUET, R., A. M. REBOULET, R. PRADEL, O. GIMENZ, AND J. D. LEBRETON.2003. U-Care [Internet]. Vers. 2.0. CEFE/CNRS, Montpellier(Quebec) [cited 2008 Jul 8]. Available from: ftp://ftp.cefe.cnrs-mop/fr/biom/Soft-CR/.
COOCH, E., AND G. WHITE. 2006. Program MARK, a gentle introduction[Internet]. Computer Associates International, Ithaca, NY [cited2008 Oct 6]. Available from: http://www.phidot.org/software/mark/docs/book/.
DONNELLY, M. A. 1994. Estimating population size. In W. R. Heyer, M.A. Donnelly, R. W. McDiarmid, L.-A. C. Hayek, and M. S. Foster(eds.), Measuring and Monitoring Biological Diversity, pp. 183–206.Smithsonian Institution Press, Washington, DC.
FERNER, J. W. 2010. Measuring and marking post-metamorphicamphibians. In C. K. Dodd (ed.), Amphibian Ecology andConservation: A Handbook of Techniques, pp. 123–141. OxfordUniversity Press, Oxford.
FUNK, W. C., M. A. DONNELLY, AND K. R. LIPS. 2005. Alternative views ofamphibian toe-clipping. Nature 433:193.
GAMBLE, L., S. RAVELA, AND K. MCGARIGAL. 2008. Multi-scale features foridentifying individuals in large biological databases: an applicationof pattern recognition technology to the Marbled SalamanderAmbystoma opacum. Journal of Applied Ecology 48:170–180.
GIBBONS, J. W., AND K. M. ANDREWS. 2004. PIT tagging: simple technologyat its best. BioScience 5:447–454.
GRAFE, T. U., S. K. KAMINSKY, J. H. BITZ, H. LUSSOW, AND K. E. LINSENMAIR.2004. Demographic dynamics of the afro-tropical Pig-Nosed Frog,Hemisus marmoratus: effects of climate and predation on survivaland recruitment. Oecologia 141:40–46.
GRAFE, T. U., T. C. WANGER, J. M. DEHLING, S. PROKSCH, AND T. BRAASCH.2006. Use of individual markings in the study of the Foot-FlaggingFrog, Staurois guttatus. Scientia Bruneiana 7:3–8.
HACC (HERPETOLOGICAL ANIMAL CARE AND USE COMMITTEE). 2004.Guidelines for use of live amphibians and reptiles in field andlaboratory research [Internet]. American Society of Ichthyologistsand Herpetologists, Lawrence, KS [cited 2010 Aug 7]. Availablefrom: www.asih.org/files/hacc-final.pdf.
HENLE, K., J. KUHN, R. PODLOUCKY, K. SCHMIDT-LOSKE, AND C. BENDER.1997. Individualerkennung und markierung mitteleuropaischeramphibian und reptilien: ubersicht und bewertung der methoden;empfehlungen aus natur- und tierschutzsicht. Mertensiella7:133–184.
HERO, J.-M. 1989. A simple code for toe clipping anurans. Herpetolog-ical Review 20:66–67.
HOFFMANN, K., M. E. MCGARRITY, AND S. A. JOHNSON. 2008. Technologymeets tradition: a combined VIE-C technique for individuallymarking anurans. Applied Herpetology 5:265–280.
JEHLE, R. 1997. Markierung und individualerkennung metamorphisier-ter amphibien. Stapfia 51:103–118.
JOHNSON, J. B., AND K. S. OMLAND. 2004. Model selection in ecology andevolution. Trends in Ecology and Evolution 19:101–108.
KAMINSKY, S. K., K. E. LINSENMAIR, AND T. U. GRAFE. 1999. Reproductivetiming, nest construction and tadpole guidance in the African Pig-Nosed Frog Hemisus marmoratus. Journal of Herpetology 33:119–123.
KAMINSKY, S. K., T. U. GRAFE, M. SPIELER, AND K. E. LINSENMAIR. 2004. Anew method for immobilizing fossorial frogs after radio-transmitterimplantation and notes on movement patterns in the Pig-NosedFrog, Hemisus marmoratus. Herpetological Review 35:146–148.
KENYON, N., A. D. PHILLOTT, AND R. A. ALFORD. 2009. Evaluation of thephotographic identification method (PIM) as a tool to identify adultLitoria genimaculata (Anura: Hylidae). Herpetological ConservationBiology 4:403–410.
KUHN, J. 1994. Lebensgeschichte und demographie von Erdkroten-weibchen Bufo bufo bufo (L.). Zeitschrift fur Feldherpetologie 1:3–87.
LAMPERT, K. P., AND K. E. LINSENMAIR. 2002. Alternative life cyclestrategies in the West African Reed Frog Hyperolius nitidulus: theanswer to an unpredictable environment? Oecologia 130:364–372.
LANGKILDE, T., AND R. A. ALFORD. 2002. The tail wags the frog: harmonicradar transponders affect movement behavior in Litoria lesueuri.Journal of Herpetology 36:711–715.
LEBRETON, J. D., K. P. BURNHAM, J. COLBERT, AND D. R. ANDERSON. 1992.Modeling survival and testing biological hypothesis using markedanimals: a unified approach with case studies. Ecological Mono-graphs 62:67–118.
LINSENMAIR, K. E. 1998. Risk spreading and risk reducing tactics of WestAfrican anurans in an unpredictable and stressful environment. InD. M. Newbery, N. Brown, and H. H. T. Prins (eds.), Dynamics ofTropical Communities, pp. 221–241. Blackwell Science, Oxford.
MAY, R. M. 2004. Ethics and amphibians. Nature 431:403.
34 T. U. GRAFE ET AL.

MCCARTHY, M. A., AND K. M. PARRIS. 2004. Clarifying the effect of toeclipping on frogs with Bayesian statistics. Journal of AppliedEcology 41:780–786.
MEUCHE, I., AND T. U. GRAFE. 2003. Nummerierte Hautimplantate—einealternative Markierungsmethode fur den Laubfrosch (Hyla arborea)?Zeitschrift fur Feldherpetologie S5:153–158.
OTT, J. A., AND D. E. SCOTT. 1999. Effects of toe-clipping and PIT-taggingon growth and survival in metamorphic Ambystoma opacum. Journalof Herpetology 33:344–348.
PARRIS, K. M., AND M. A. MCCARTHY. 2001. Identifying effects of toe-clipping on anuran return rates: the importance of statistical power.Amphibia–Reptilia 22:275–289.
———. 2008. Toe clipping of anurans for mark–recapture studies:acceptable if justified. Herpetological Review 39:148–149.
PHILLOTT, A. D., L. F. SKERRATT, K. R. MCDONALD, F. L. LEMCKERT, H. B.HINES, J. M. CLARKE, R. A. ALFORD, AND R. SPEARE. 2007. Toe-clippingas an acceptable method of identifying individual anurans in mark–recapture studies. Herpetological Review 38:305–308.
———. 2008. Toe clipping of anurans for mark–recapture studies:acceptable if justified. That’s what we said! Herpetological Review39:149–150.
PHILLOTT, A. D., K. R. MCDONALD, AND L. F. SKERRATT. 2010. Return ratesof male hylid frogs Litoria genimaculata, L. nannotis, L. rheocola andNyctimystes dayi after toe-tipping. Endangered Species Research11:183–188.
POLLACK, K. H., J. D. NICHOLS, C. BROWNIE, AND J. E. HINES. 1990.Statistical inference for capture–recapture experiments. WildlifeMonographs 107:1–197.
RICHARDS, C. M., B. M. CARLSON, AND S. L. ROGERS. 1975. Regeneration ofdigits and forelimbs in the Kenyan Reed Frog Hyperolius viridiflavusferniquei. Journal of Morphology 146:431–445.
RODEL, M.-O., M. SPIELER, K. GRABOW, AND C. BOCKHELER. 1995. Hemisusmarmoratus (Peters, 1854) (Anura: Hemisotidae), Fortpflanzungs-strategien eines Savannenfrosches. Bonner zoologische Beitrage45:191–207.
RODEL, M.-O., V. H. W. RUDOLF, S. FROSCHAMMER, AND K. E. LINSENMAIR.2004. Life history of a West-African tree-hole breeding frog,Phrynobatrachus guineensis Guibe and Lamotte, 1961 (Amphibia:Anura: Petropedetidae). Miscellaneous Publications of the Museumof Zoology, University of Michigan 193:31–44.
RUDOLF, V. H. W., AND M.-O. RODEL. 2005. Oviposition site selection in acomplex and variable environment: the role of habitat quality andconspecific cues. Oecologia 142:316–325.
———. 2007. Phenotypic plasticity and optimal timing of metamor-phosis under uncertain time constraints. Evolutionary Ecology21:121–142.
SANDBERGER, L., H. FELDHAAR, K. P. LAMPERT, D. LAMATSCH, AND M.-O. RODEL.2010. Small, specialised and highly mobile? The tree-hole breedingfrog, Phrynobatrachus guineensis, lacks fine-scale population structure.African Journal of Herpetology 59:79–94.
SCHMIDT, B. R., M. SCHAUB, AND B. R. ANHOLT. 2002. Why you should usecapture–recapture methods when estimating survival and breedingprobabilities: on bias, temporary emigration, overdispersion, andcommon toads. Amphibia–Reptilia 23:375–388.
SCHMIDT, B. R., M. SCHAUB, AND S. STEINFARTZ. 2007. Apparent survival ofthe salamander Salamandra salamandra is low because of highmigratory activity. Frontiers in Zoology 4:19.
STEWART, M. M. 1995. Climate driven population fluctuations in rainforest frogs. Journal of Herpetology 29:437–446.
STEWART, M. M., AND F. H. POUGH. 1983. Population density of tropicalforest frogs: relation to retreat sites. Science 221:570–572.
STEWART, M. M., AND A. S. RAND. 1991. Vocalizations and the defense ofretreat sites by male and female frogs, Eleutherodactylus coqui.Copeia 1991:1013–1024.
STEWART, M. M., AND L. L. WOOLBRIGHT. 1996. Amphibians. In D. P.Reagan and R. B. Waide (eds.), The Food Web of a Tropical RainForest, pp. 273–320. University of Chicago Press, Chicago.
STUART, S. N., J. S. CHANSON, N. A. COX, B. E. YOUNG, A. S. L. RODRIGUES,D. L. FISCHMAN, AND R. W. WALLER. 2004. Status and trends ofamphibian declines and extinctions worldwide. Science306:1783–1786.
WADDLE, J. H., K. G. RICE, F. J. MAZZOTTI, AND H. F. PERCIVAL. 2008.Modeling the effect of toe clipping on treefrog survival: beyond thereturn rate. Journal of Herpetology 42:467–473.
WELLS, K. D. 2007. The Ecology and Behavior of Amphibians. Universityof Chicago Press, Chicago.
WHITE, G. C., AND K. P. BURNHAM. 1999. Program MARK: survivalestimation from populations of marked animals. Bird Study46:120–139.
Accepted: 12 September 2010.
VIABLE METHOD FOR MARKING ANURANS 35





























