Embed Size (px)
Citation preview
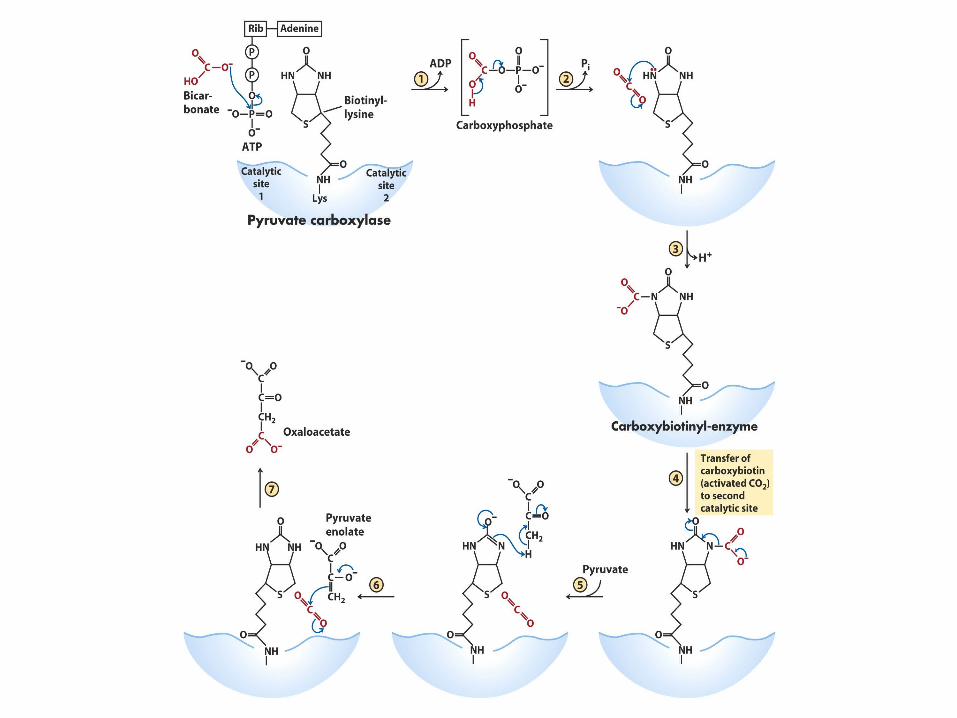
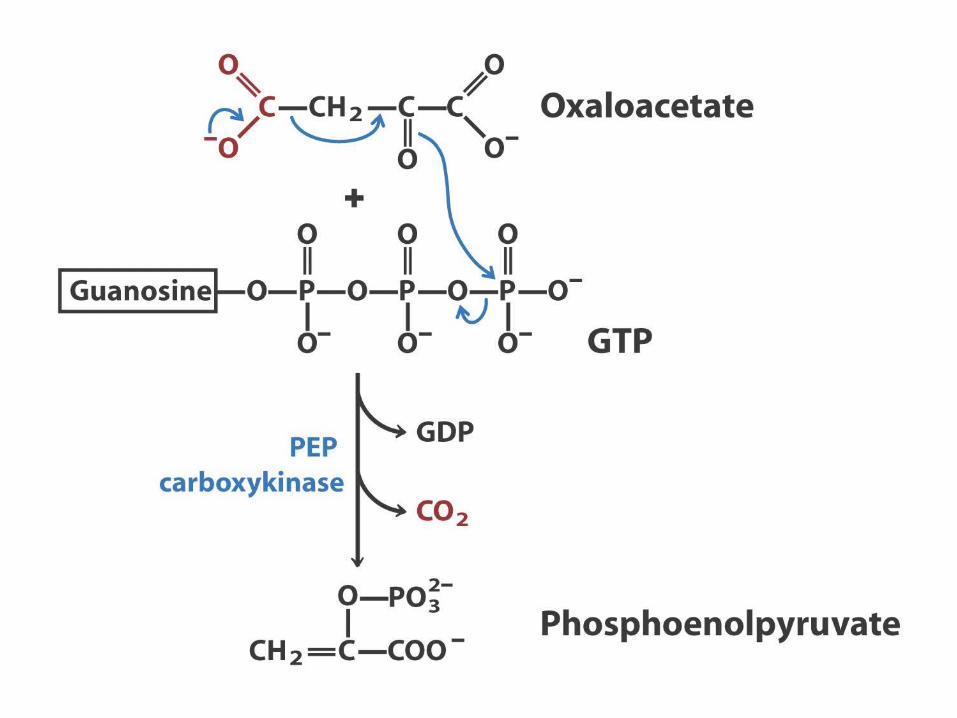
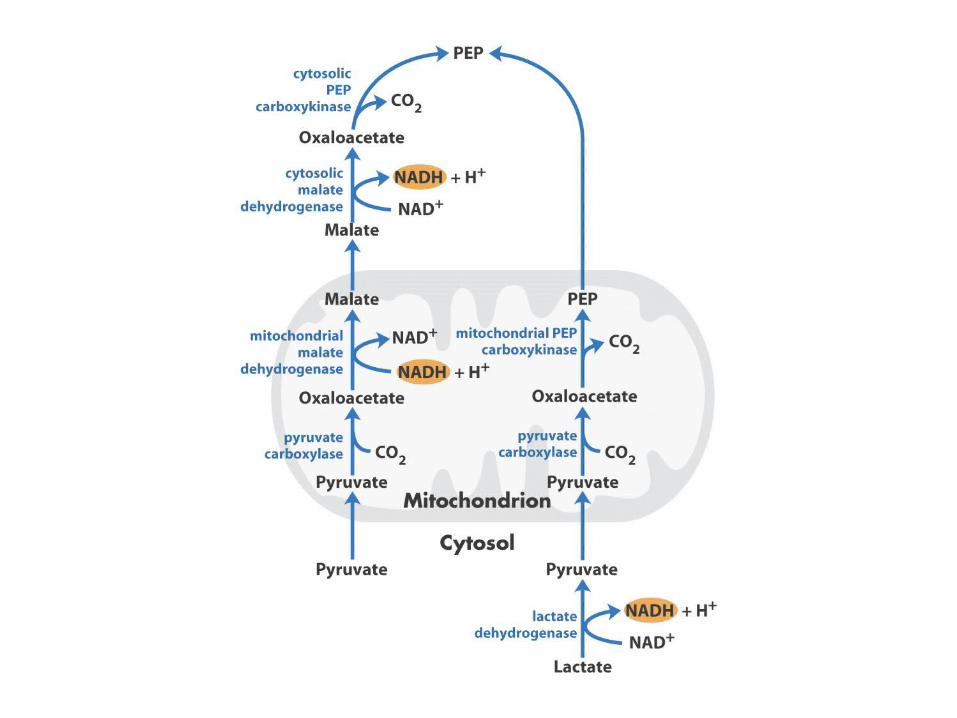
Pyruvate Carboxylase
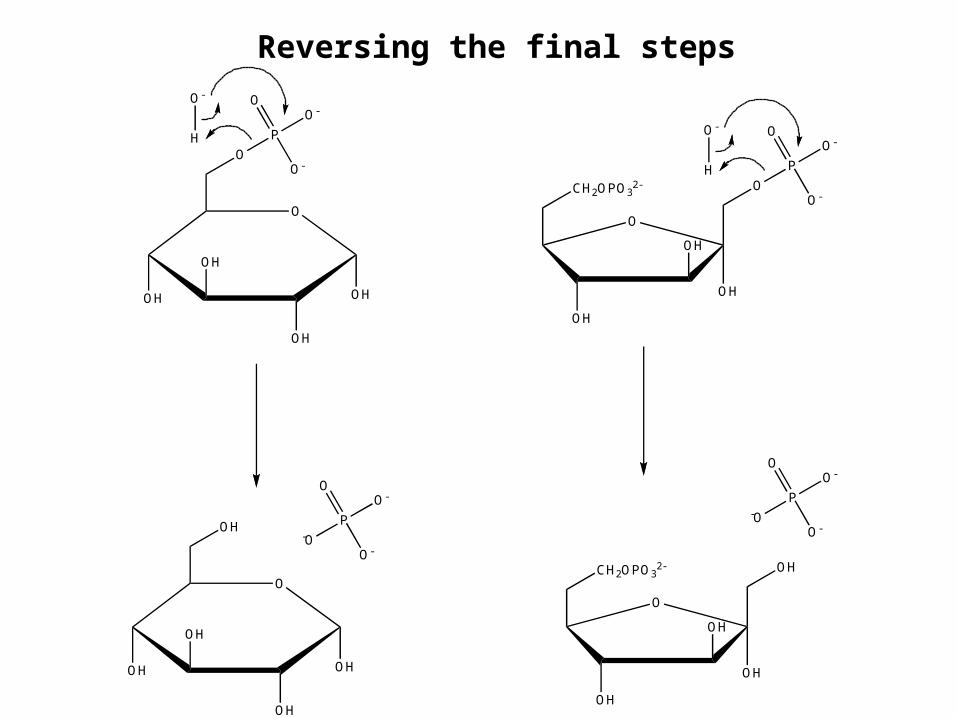
Reversing the final steps
O
OH OH
OH
OH
O
P
OO-
O-
O-
H
O
OH OH
OH
OH
OH P
OO-
O-
-O
OH
OH
OH
O
CH2OPO32- O
P
OO-
O-
O-
H
OH
OH
OH
O
CH2OPO32- OH
P
OO-
O-
-O
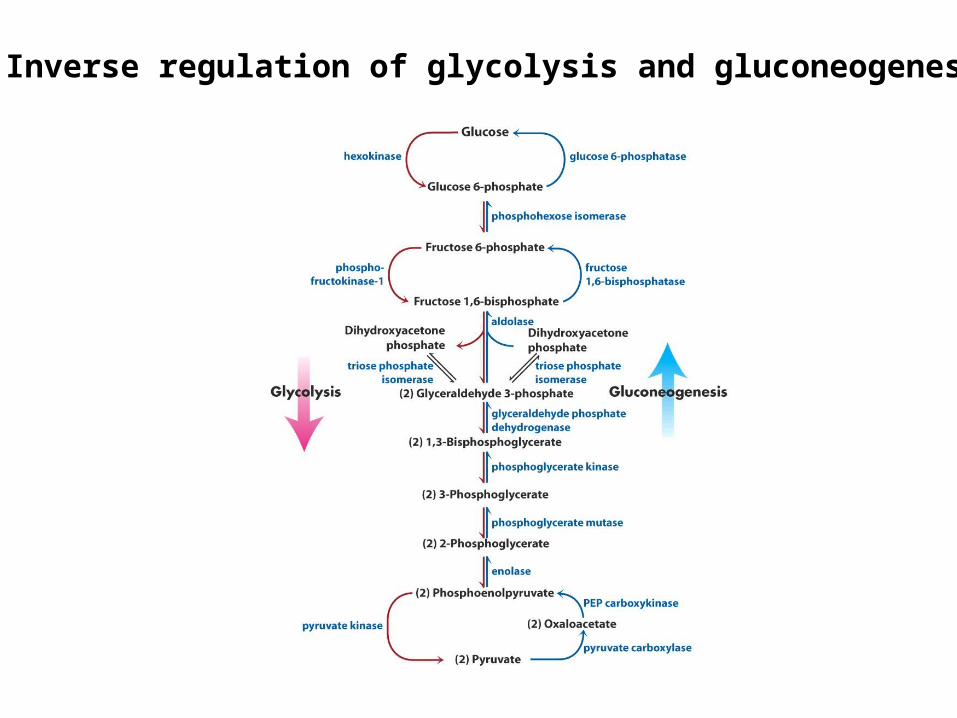
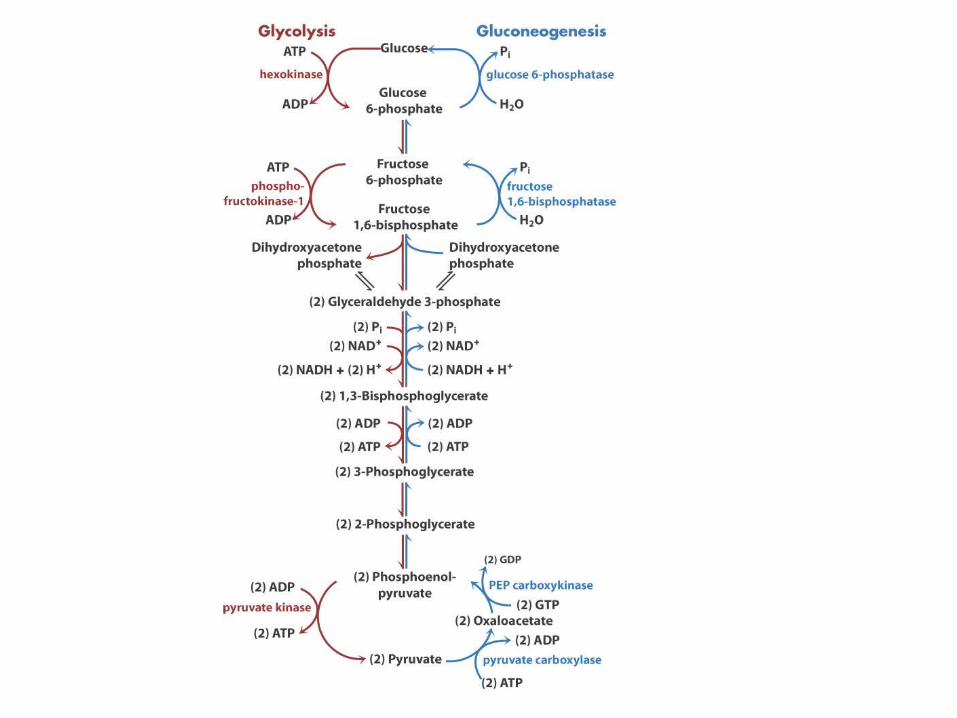
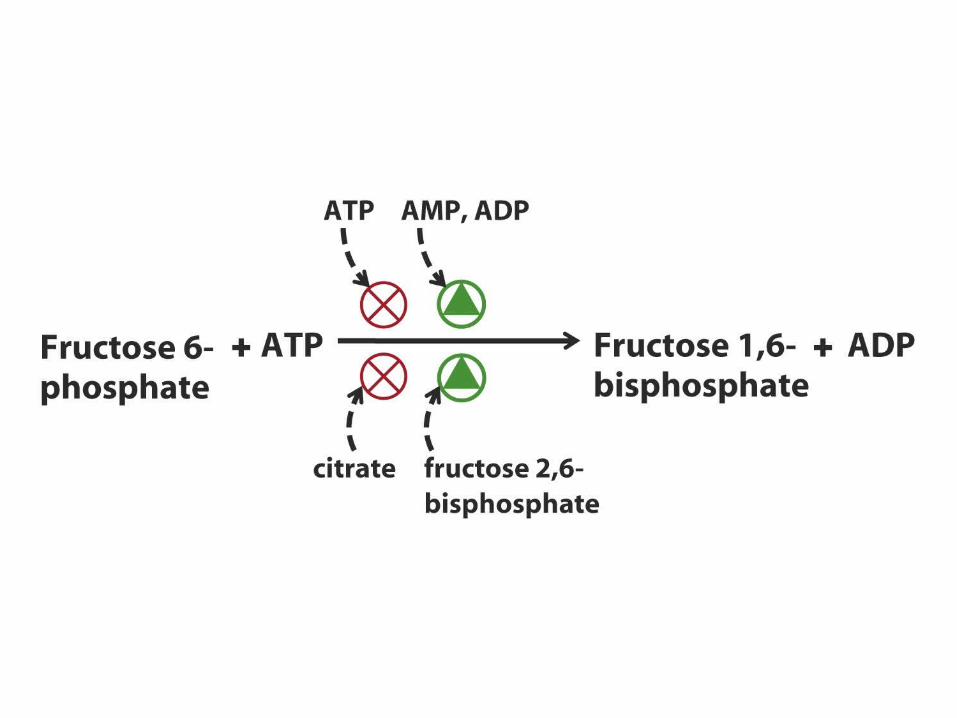
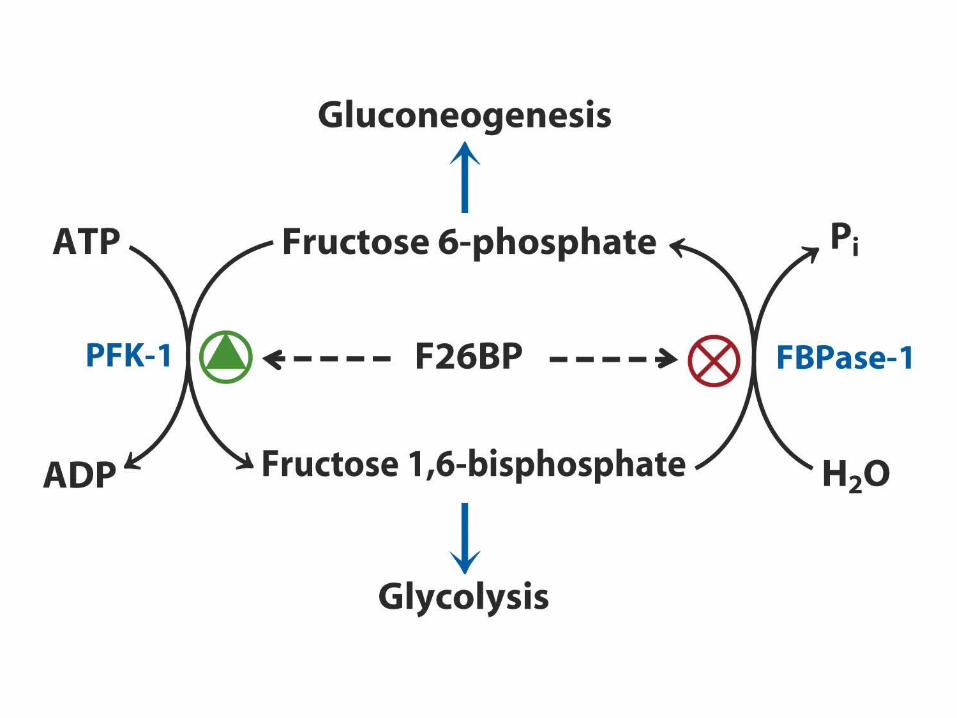
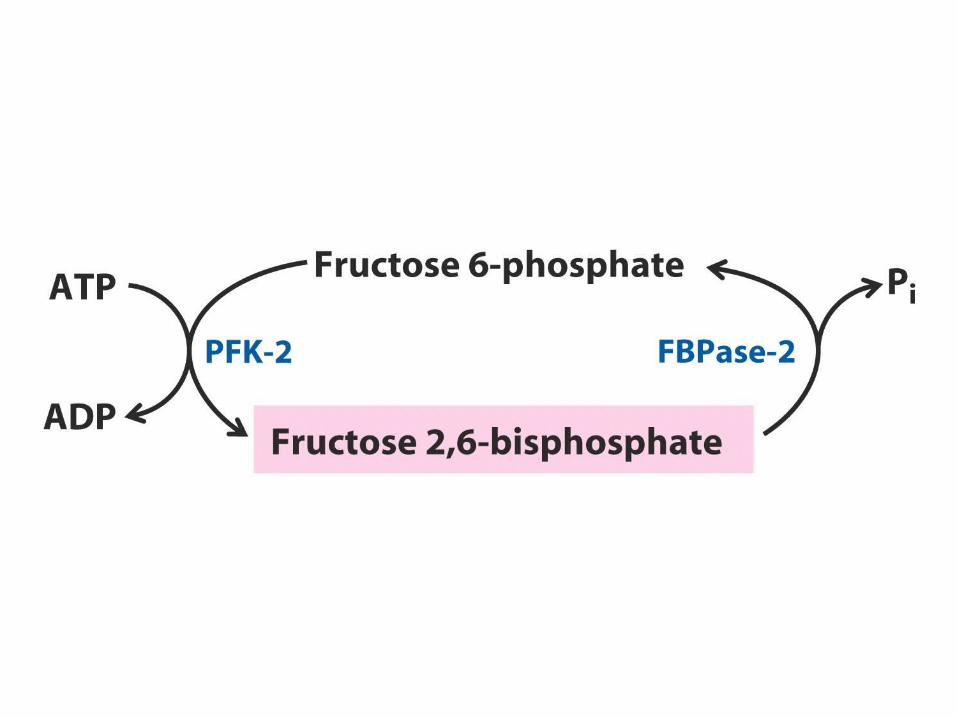
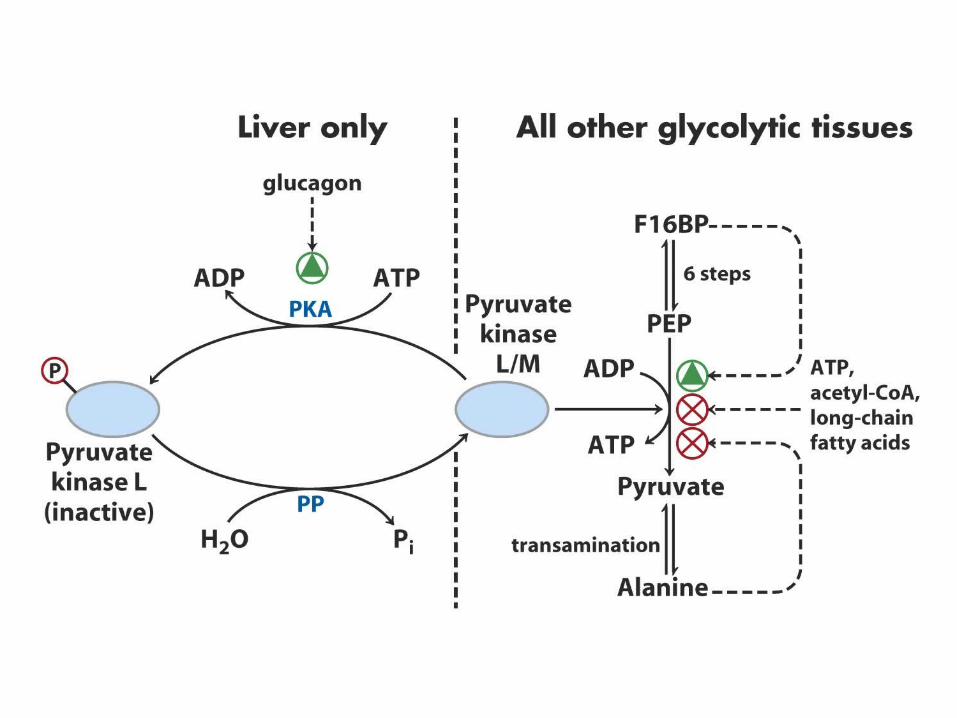
Inverse regulation of glycolysis and gluconeogenesis
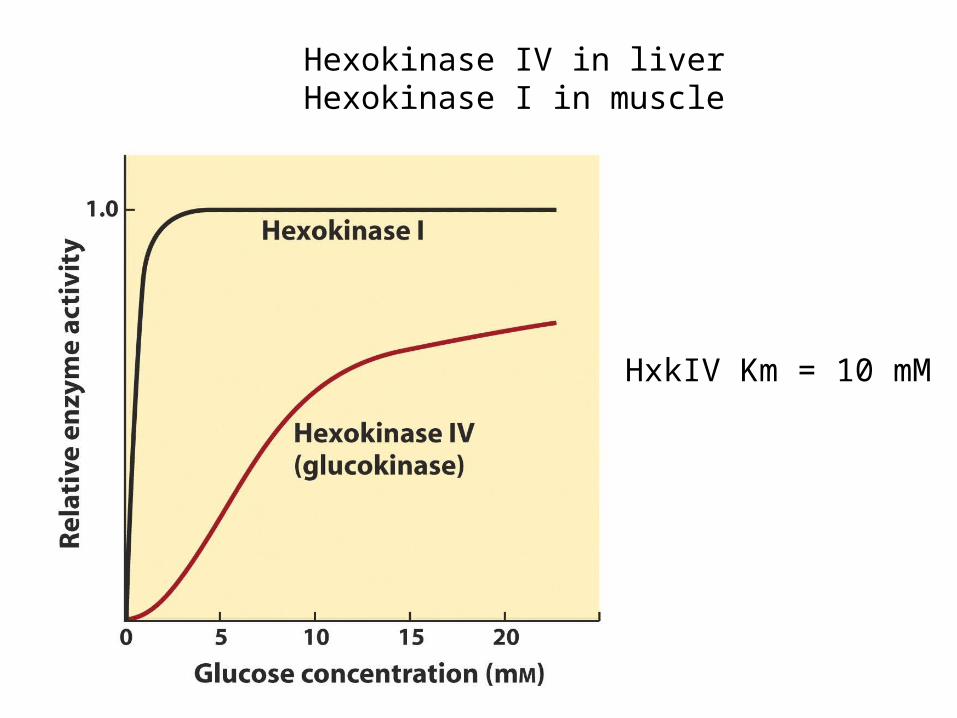
Hexokinase IV in liverHexokinase I in muscle
HxkIV Km = 10 mM
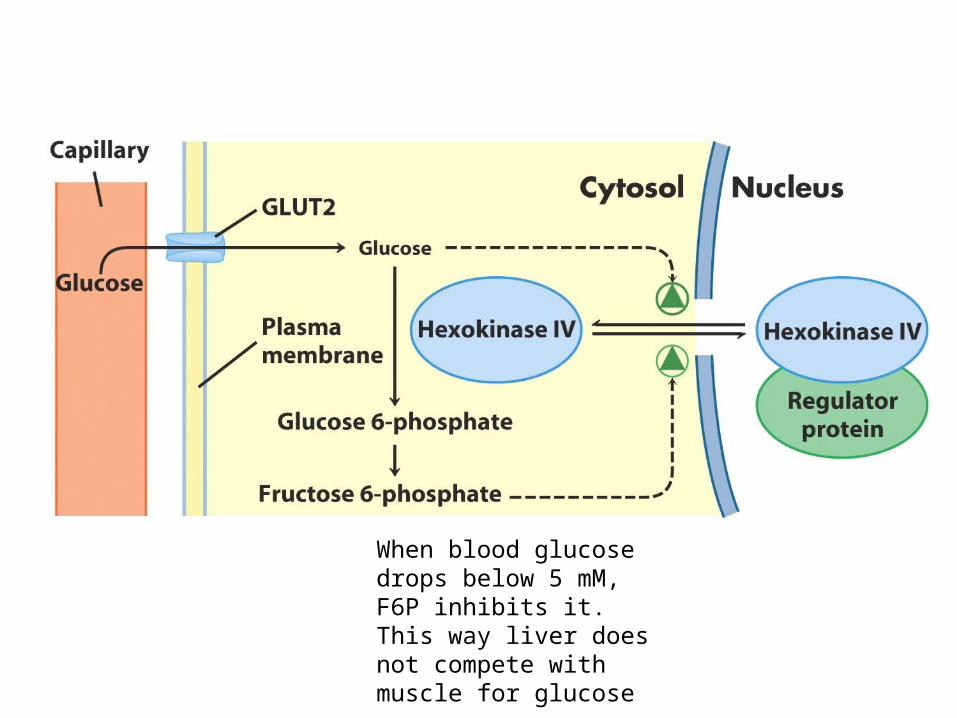
When blood glucose drops below 5 mM, F6P inhibits it. This way liver does not compete with muscle for glucose
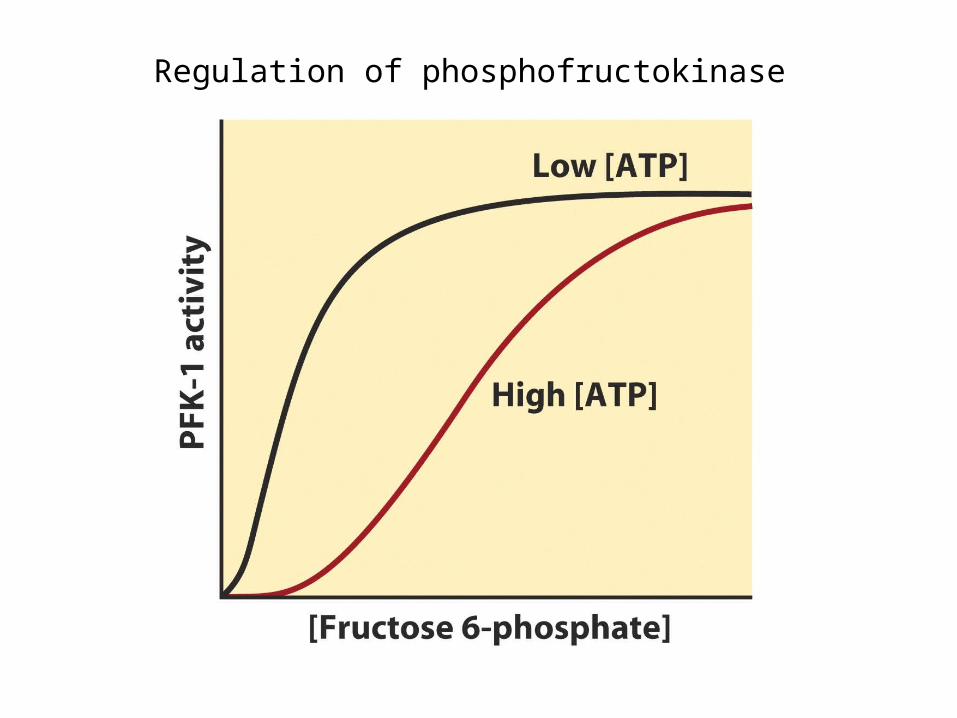
Regulation of phosphofructokinase



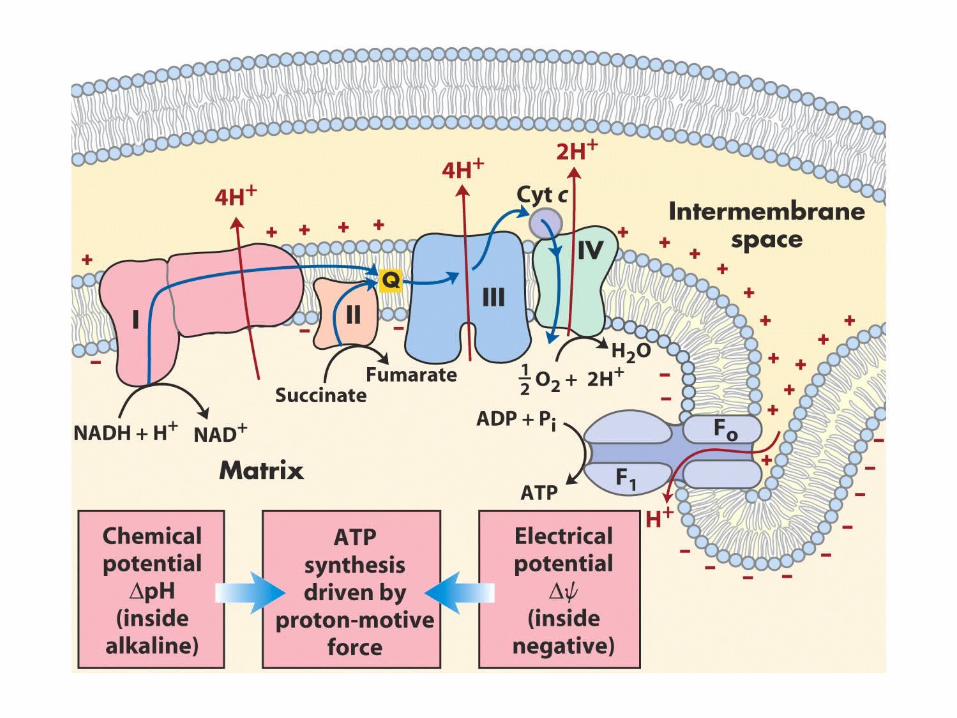
Electron transport and oxidative phosphorylation
Glucose is completely oxidized to CO2 through the enzymatic reactions of glycolysis and the citric acid cycle. The redox equation for this process is:
C6H12O6 + 6O2 ---> 6CO2 + 6H2O ΔG°’ = -2823 kJ.mol-1
Which can be represented by two half reactions:
C6H12O6 + 6H2O ---> 6CO2 + 24H+ + 24e- glucose is oxidized
6O2 + 24H+ + 24e- ---> 12H2O molecular oxygen is reduced
In living systems the electron transfer process connecting these two half reactions occurs through a multistep pathway that harnesses the liberated free energy to form ATP.
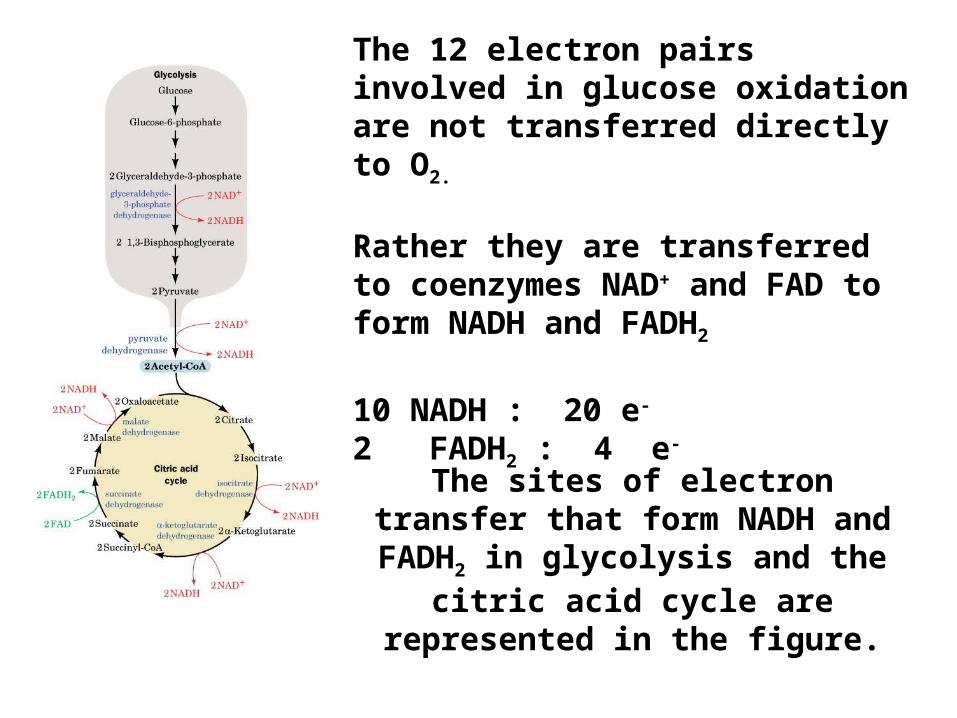
The sites of electron transfer that form NADH and FADH2 in
glycolysis and the citric acid cycle are represented in the figure.
The 12 electron pairs involved in glucose oxidation are not transferred directly to O2.
Rather they are transferred to coenzymes NAD+ and FAD to form NADH and FADH2
10 NADH : 20 e-
2 FADH2 : 4 e-

The electrons are extracted from the cofactors by reoxidation and then join the electron-transport chain, in this process, protons are expelled from the mitochondrion. The free energy stored in the resulting pH gradient drives the synthesis of ATP from ADP and Pi (inorganic phosphate) through oxidative phosphorylation.
Reoxidation of NADH ~ 3 ATPReoxidation of FADH2 ~ 2 ATP
A total of 38 ATP are produced per each molecule of glucose completely oxidized to CO2 and H2O (including the 2 ATP made in glycolysis and the 2 ATP made in the citric acid cycle)

NAD+ and FAD coenzymes are reduced during glucose oxidation
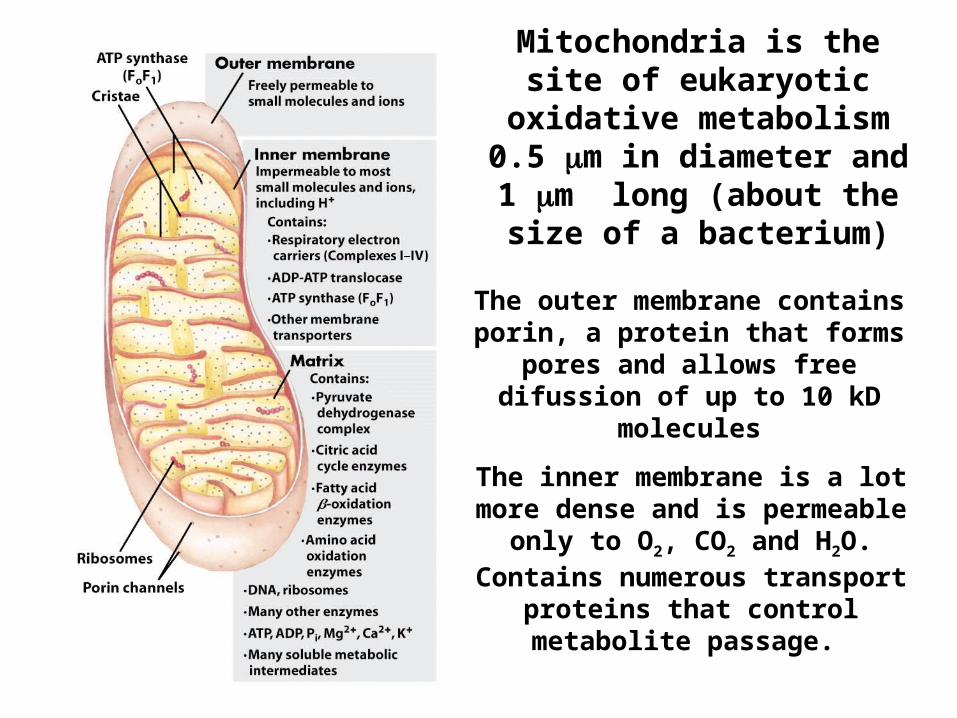
Mitochondria is the site of eukaryotic oxidative metabolism 0.5 m in
diameter and 1 m long (about the size of a
bacterium)
The outer membrane contains porin, a protein that forms pores and
allows free difussion of up to 10 kD molecules
The inner membrane is a lot more dense and is permeable only to O2, CO2 and H2O. Contains numerous
transport proteins that control metabolite passage.
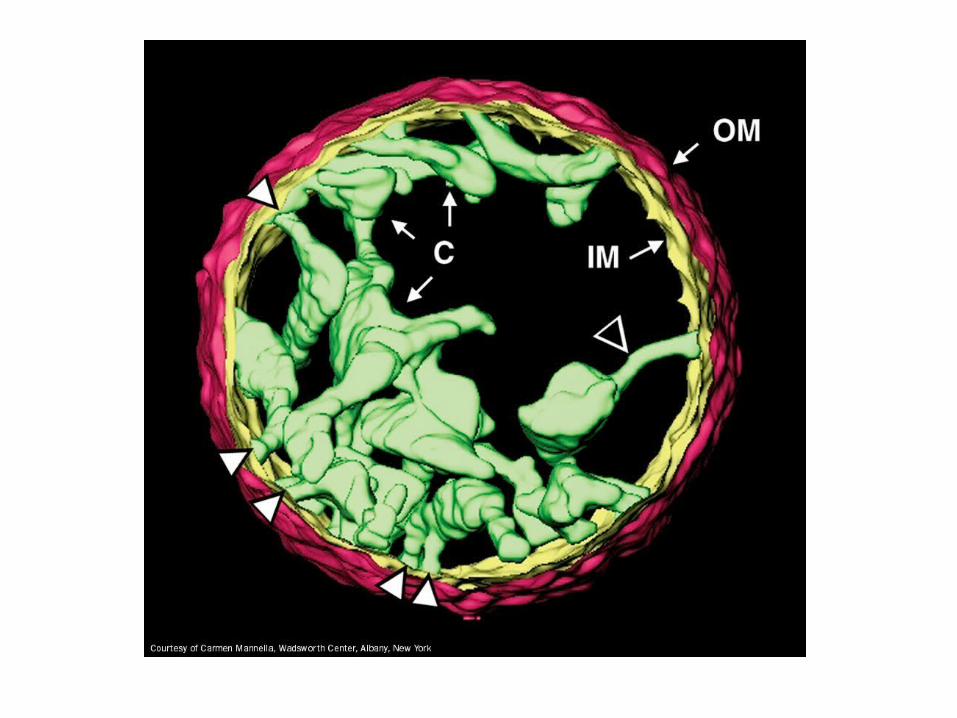
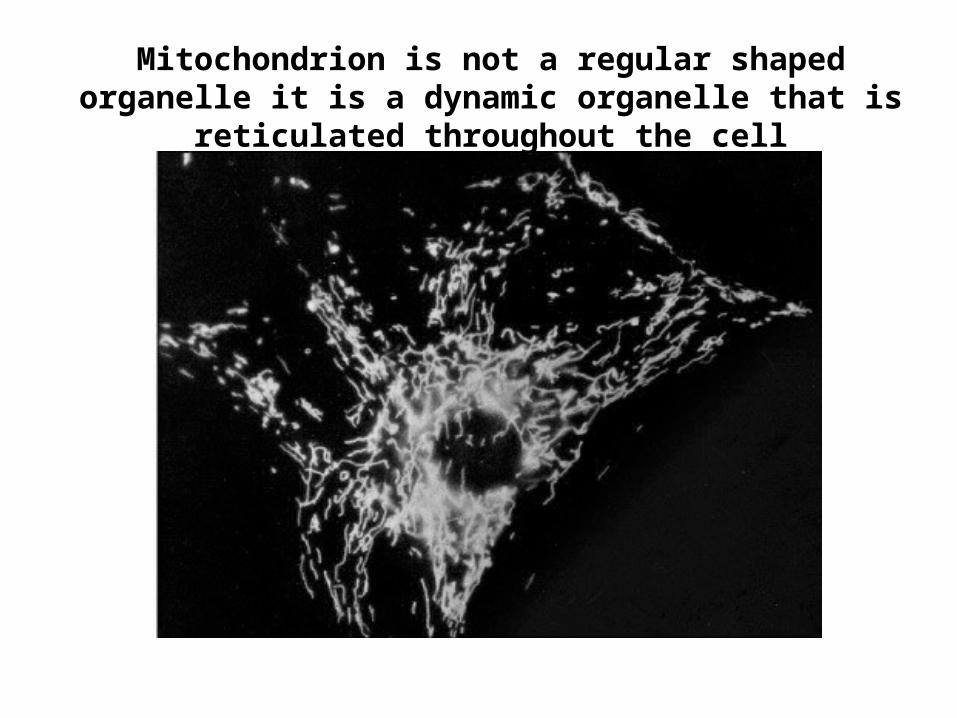
Mitochondrion is not a regular shaped organelle it is a dynamic organelle that is reticulated throughout the cell
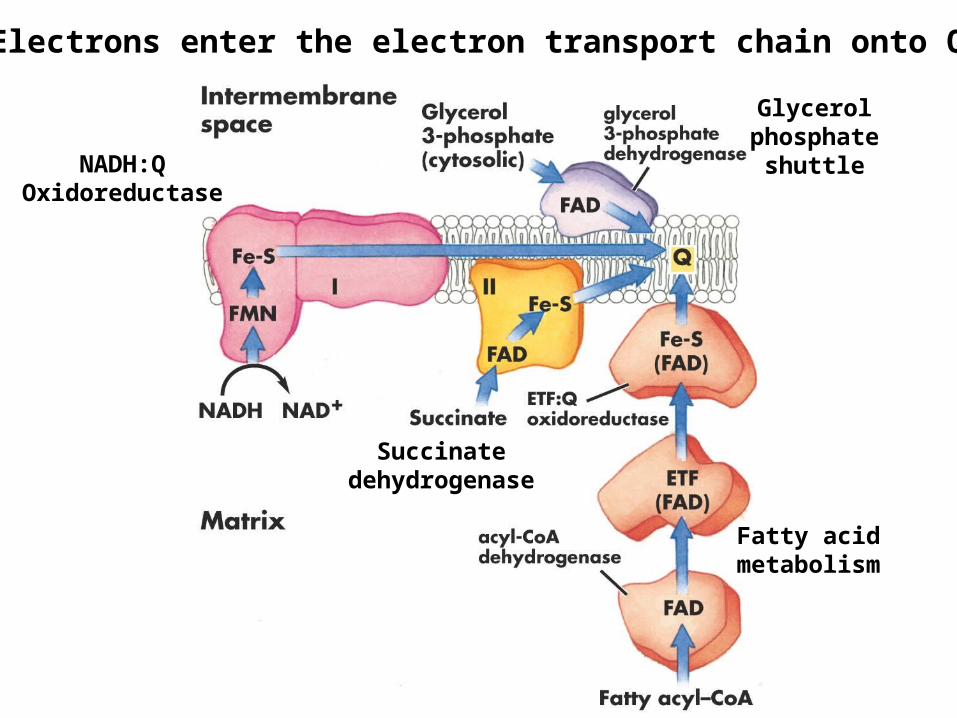
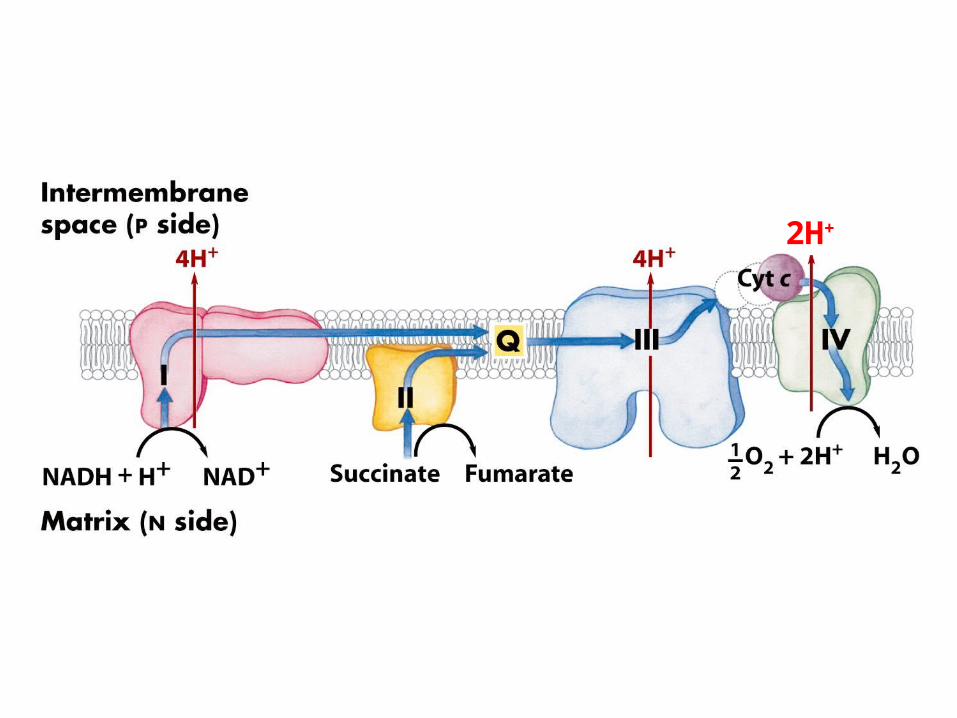
Electrons enter the electron transport chain onto Q
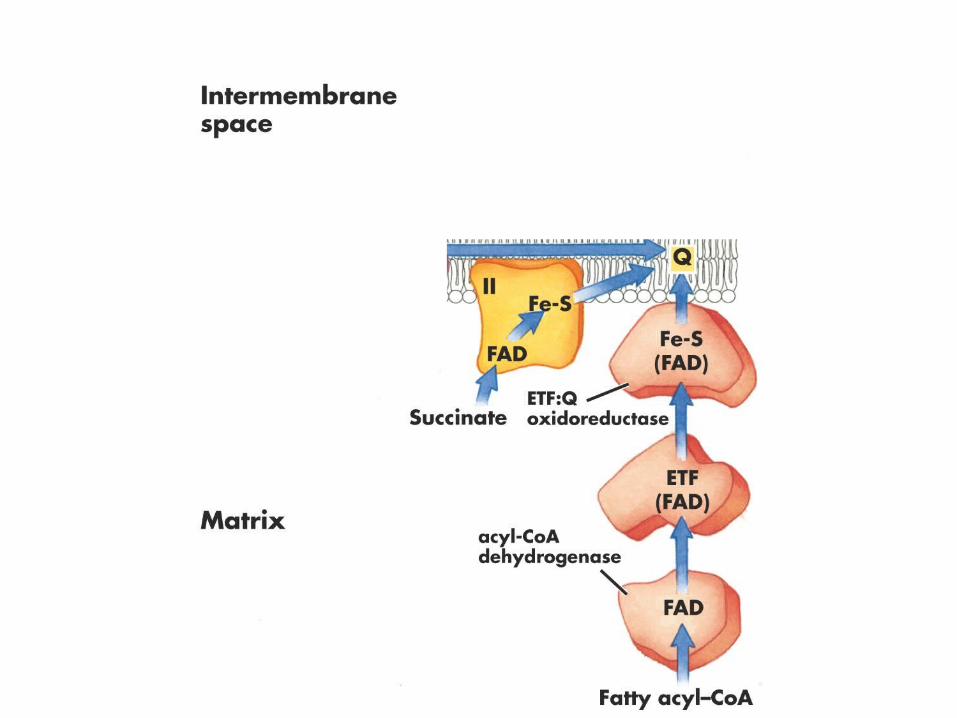
Fatty acid metabolism
Glycerol phosphate
shuttle
Succinate dehydrogenase
NADH:QOxidoreductase

Reduced state has more protons than the oxidized state!

Redox loops pumps out four protons!

The glycerophosphate shuttle mainly occurs in rapidly metabolizing tissues.
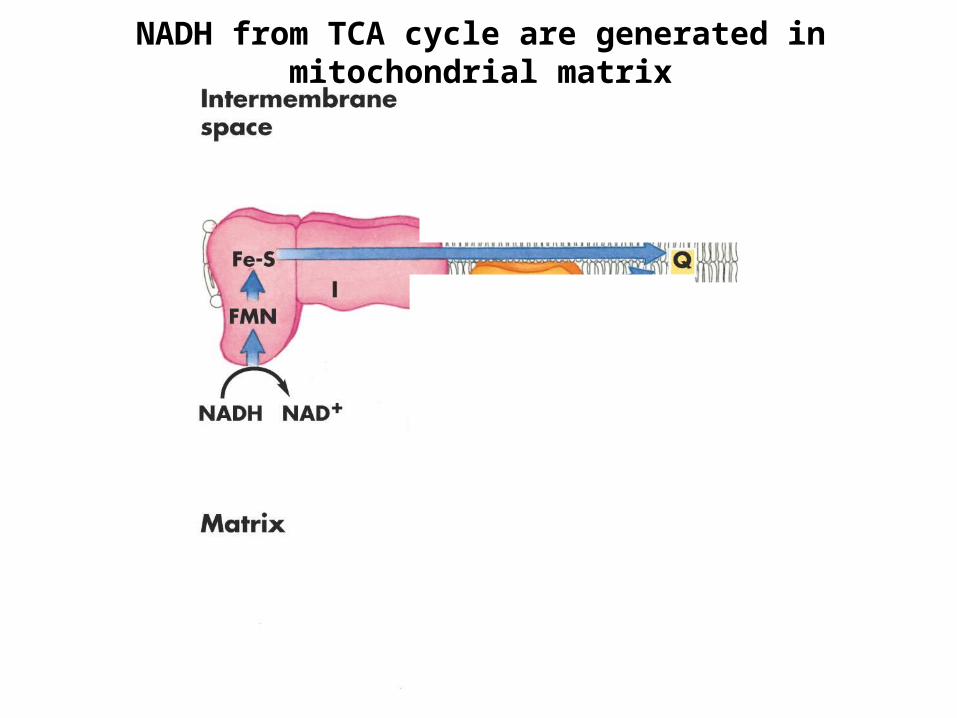
NADH from TCA cycle are generated in mitochondrial matrix
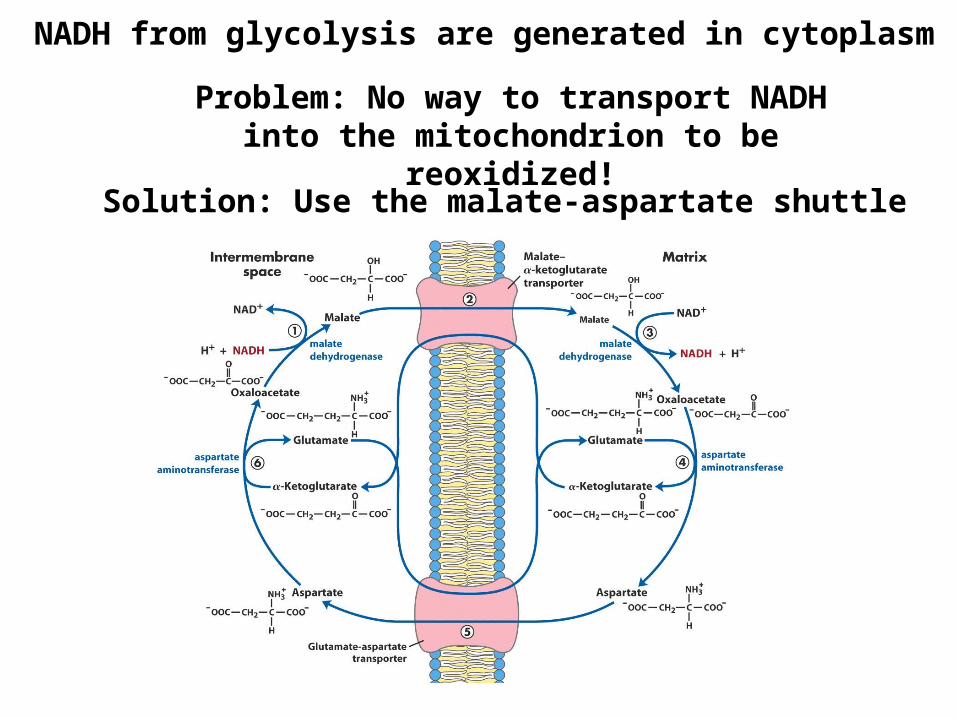
NADH from glycolysis are generated in cytoplasm
Problem: No way to transport NADH into the mitochondrion to be reoxidized!
Solution: Use the malate-aspartate shuttle
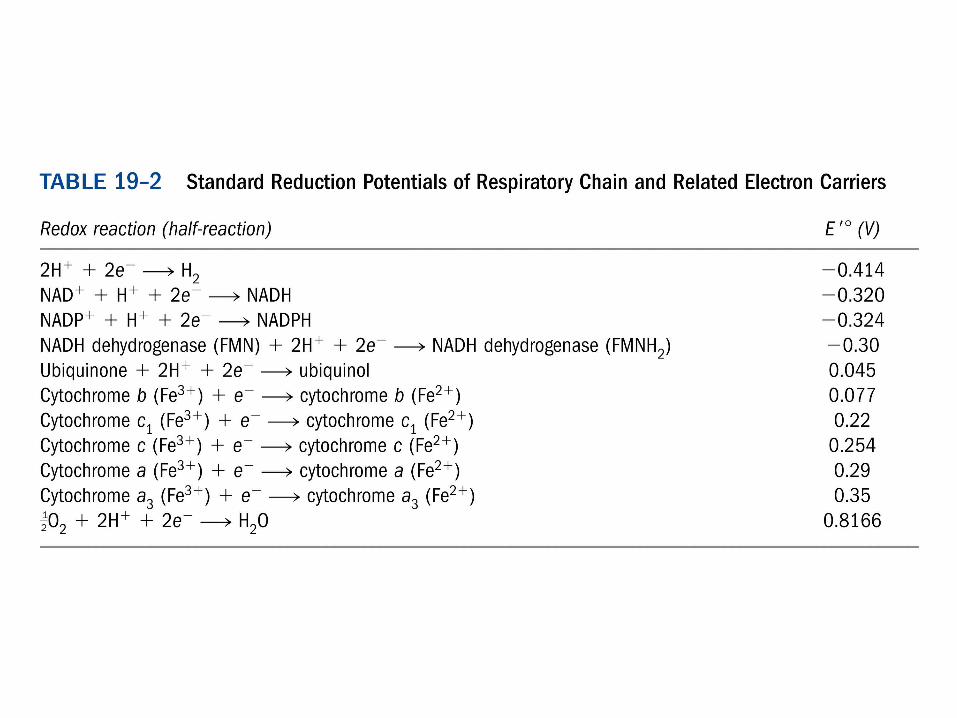
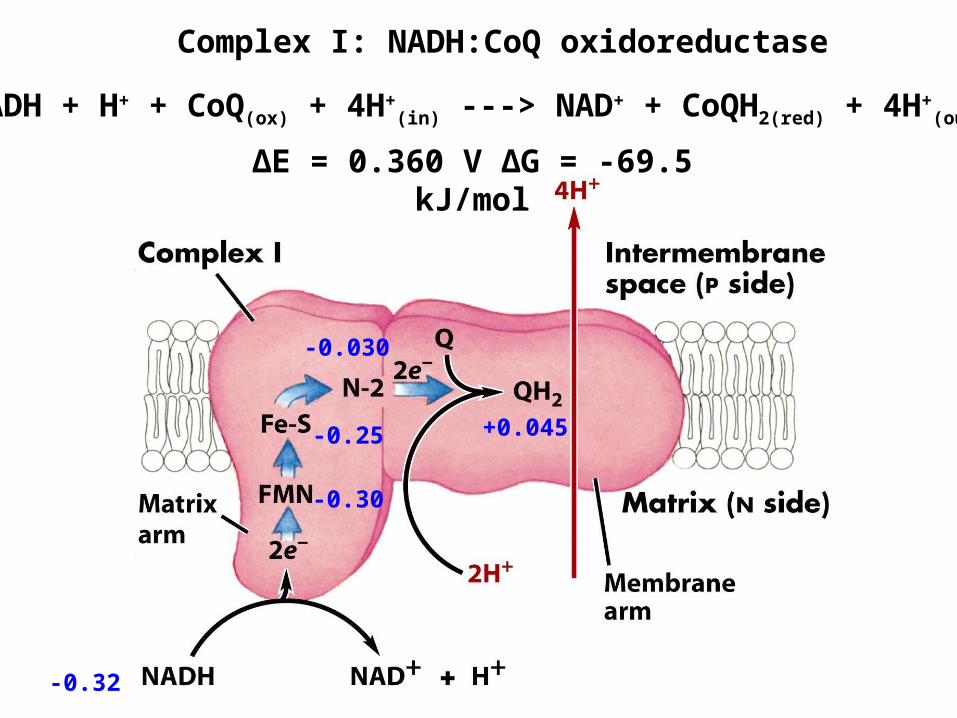
Complex I: NADH:CoQ oxidoreductase
∆E = 0.360 V ∆G = -69.5 kJ/mol
NADH + H+ + CoQ(ox) + 4H+(in) ---> NAD+ + CoQH2(red) + 4H+
(out)
-0.32
-0.30
-0.030
+0.045-0.25

Ubiquinone +0.045
-0.30
-0.32
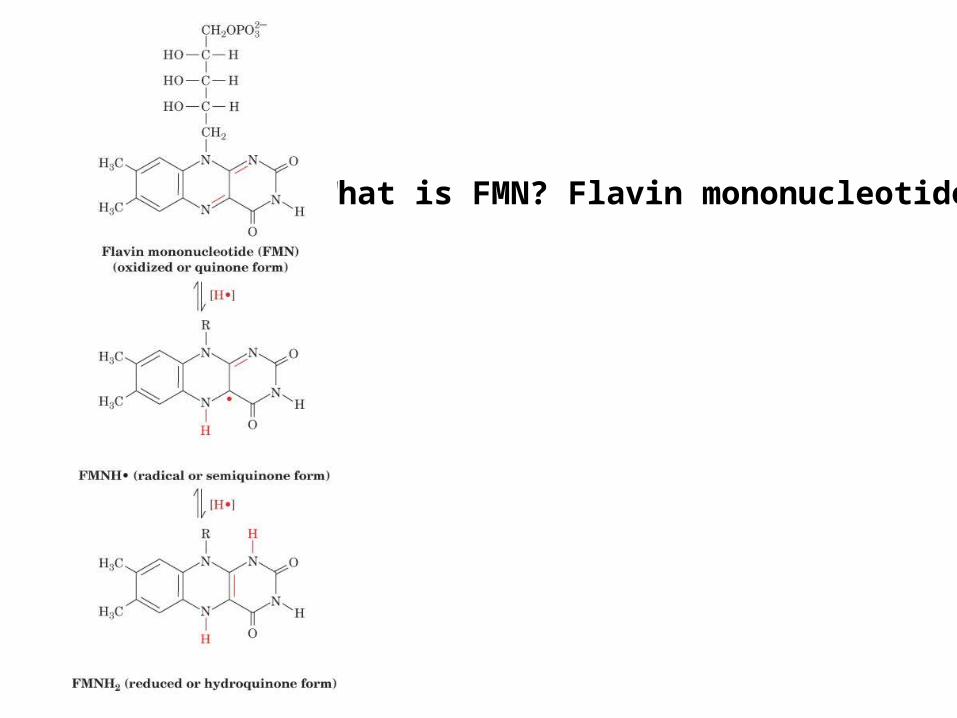
What is FMN? Flavin mononucleotide

What are FeS clusters?
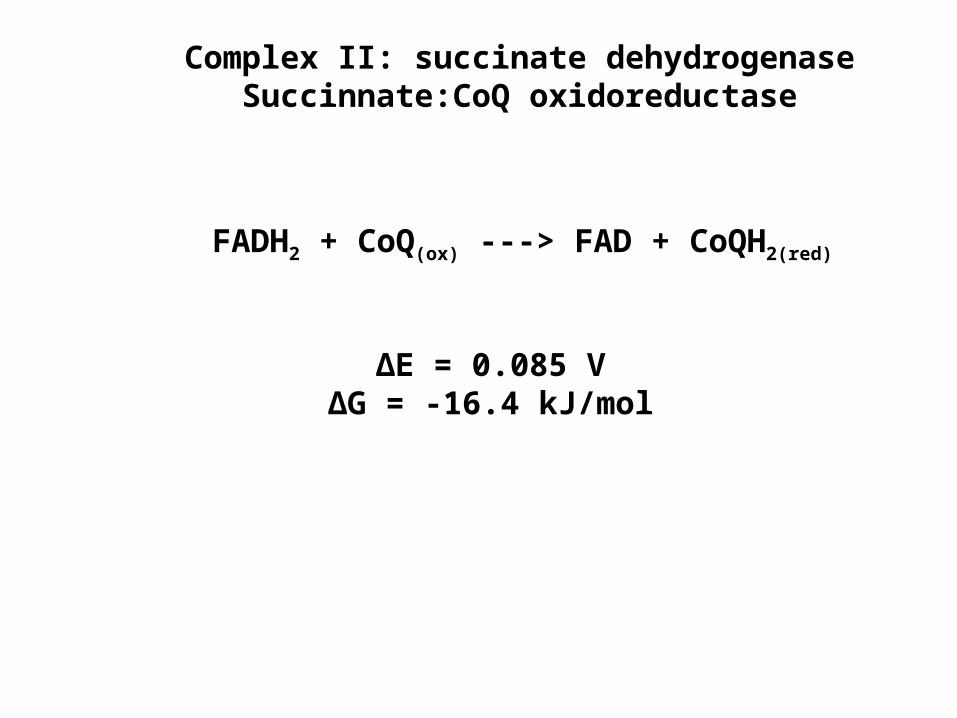
Complex II: succinate dehydrogenaseSuccinnate:CoQ oxidoreductase
FADH2 + CoQ(ox) ---> FAD + CoQH2(red)
∆E = 0.085 V∆G = -16.4 kJ/mol
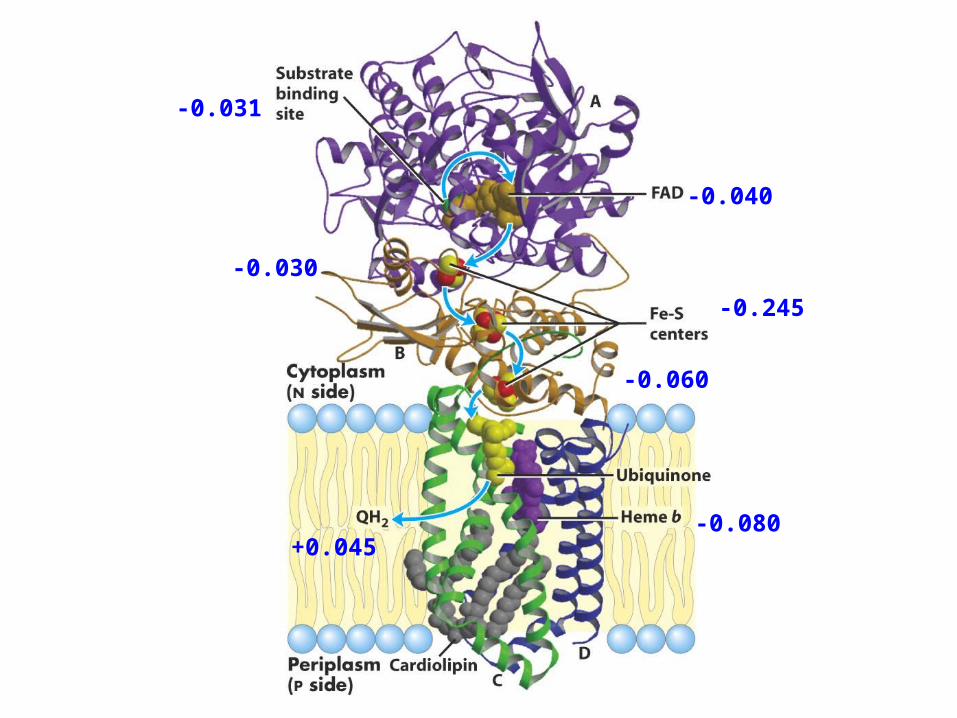
-0.031
-0.040
-0.030
-0.245
-0.060
-0.080+0.045

Succinate -0.031
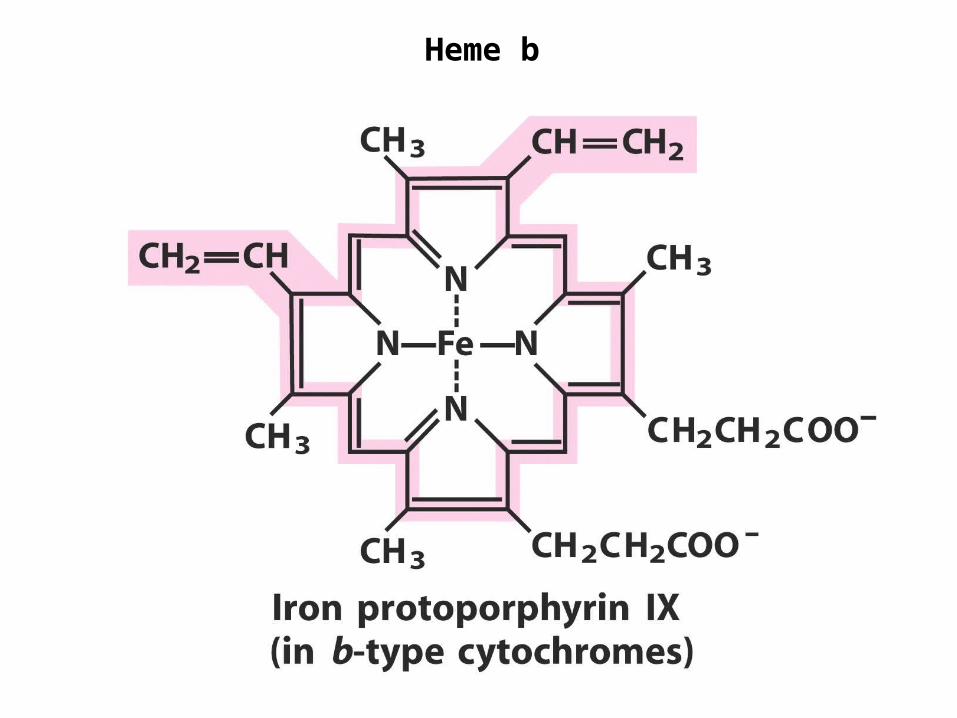
Heme b
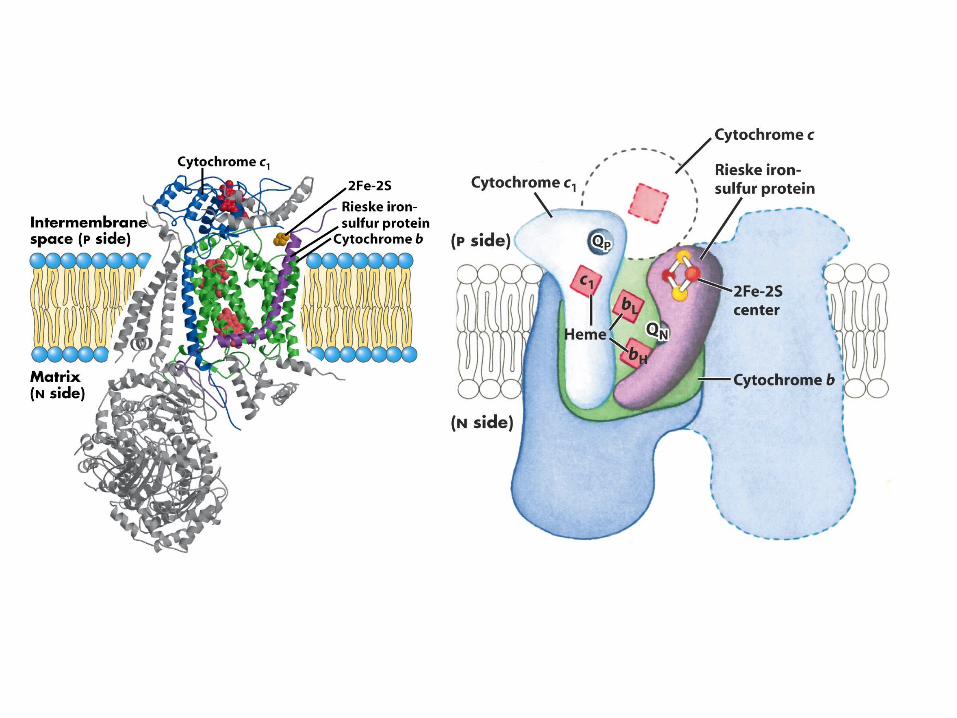
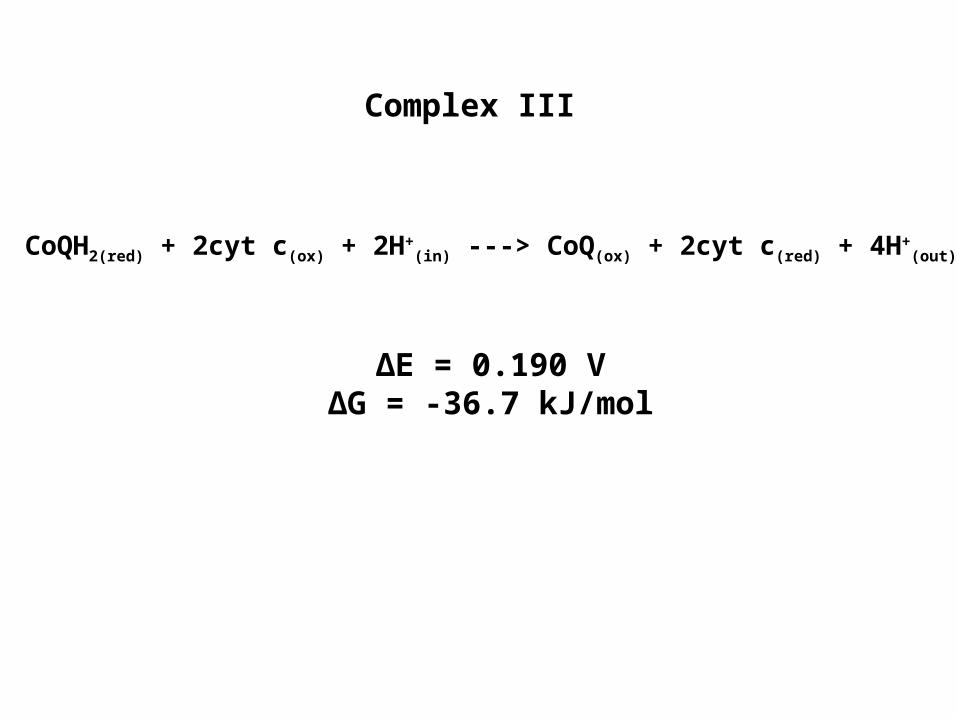
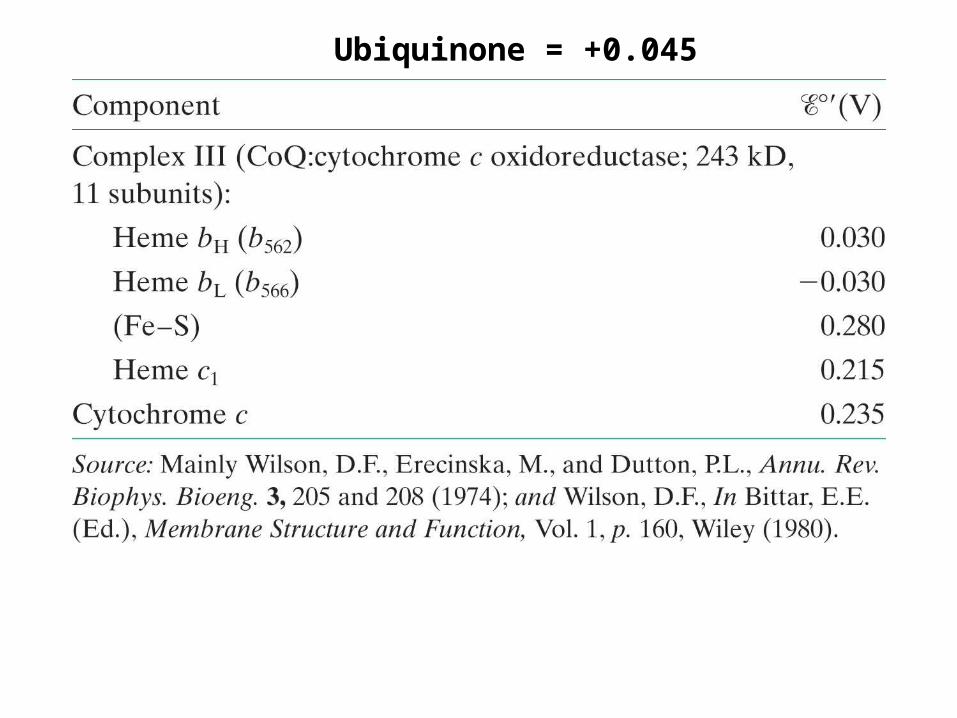
Complex III
CoQH2(red) + 2cyt c(ox) + 2H+(in) ---> CoQ(ox) + 2cyt c(red) + 4H+
(out)
∆E = 0.190 V∆G = -36.7 kJ/mol

+0.045
+0.030
-0.030
+0.280+0.215
+0.235

CoQH2 + cyt c1(ox) ---> CoQ•- + cyt c1(red) + 2H+ (out)
CoQH2 + CoQ•- + cyt c1(ox) + 2H+(in) ---> CoQ + CoQH2 + cyt c1(red) + 2H+ (out)
Cycle I
Cycle II
CoQH2 + 2cyt c1(ox) + 2H+(in) ---> CoQ + 2cyt c1(red) + 4H+ (out)
Net Reaction4 protons pumped instead of 2
Ubiquinone = +0.045
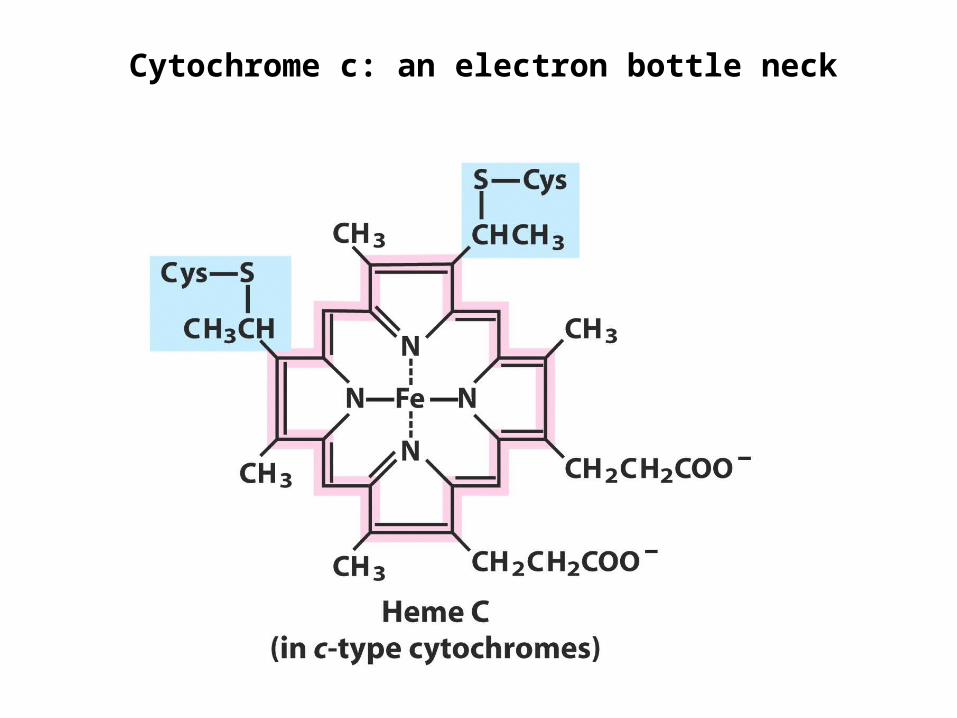
Cytochrome c: an electron bottle neck
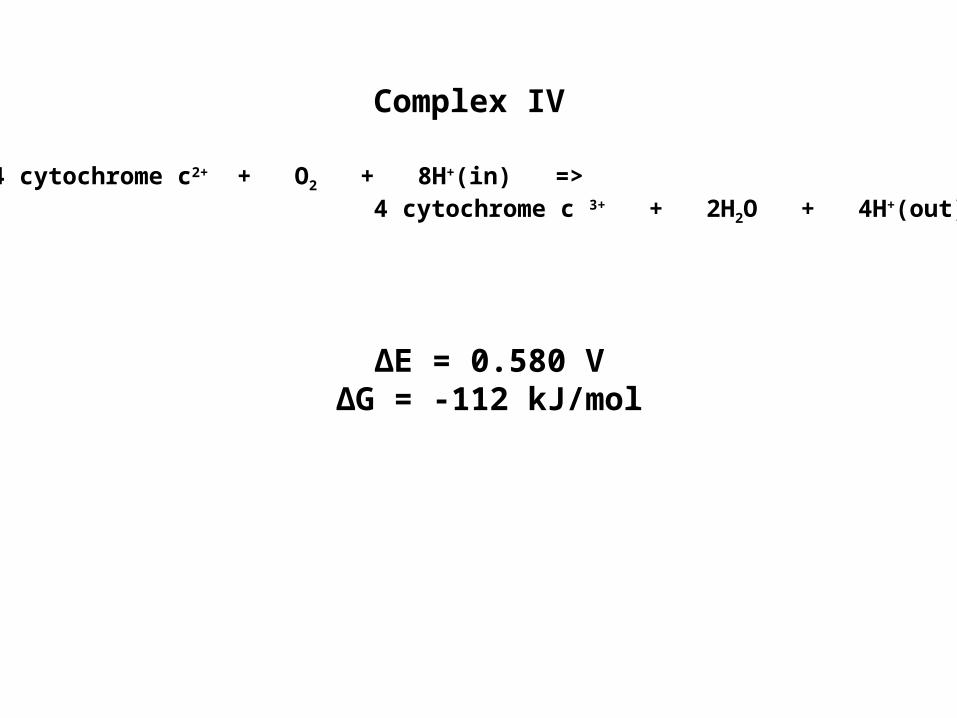
Complex IV
∆E = 0.580 V∆G = -112 kJ/mol
4 cytochrome c2+ + O2 + 8H+(in) =>4 cytochrome c 3+ + 2H2O + 4H+(out)
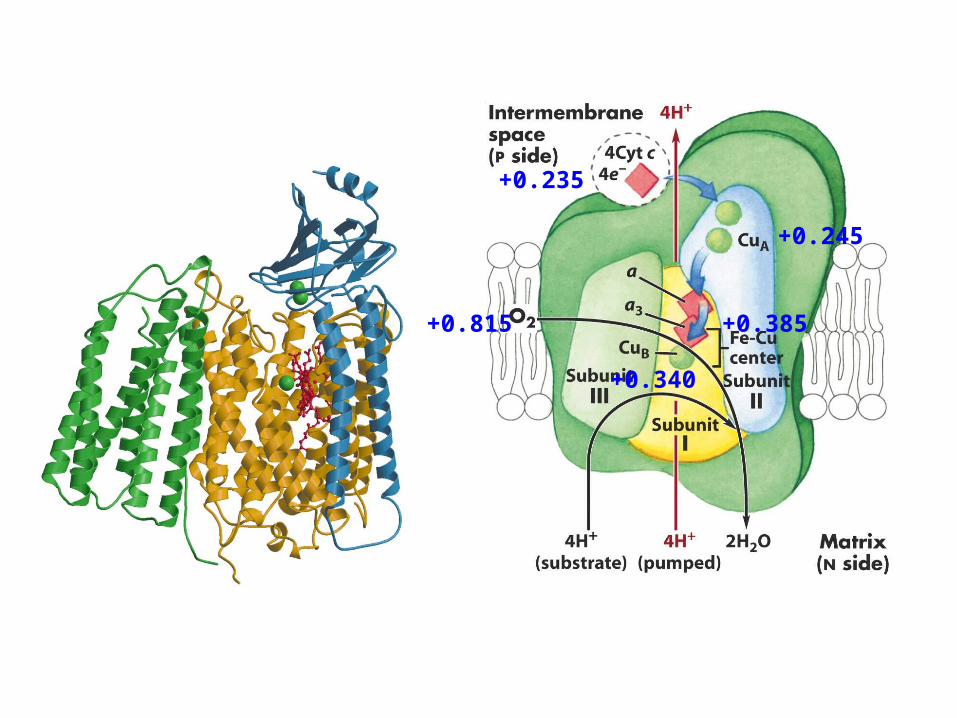
+0.235
+0.245
+0.385
+0.340
+0.815

2H+

If 2 electrons enter at complex I4 + 4 + 2 = 10 protons pumped out
If 2 electrons enter at complex II or Glycerol dehydrogenase or fatty
acid metabolism4 + 2 = 6 protons pumped out
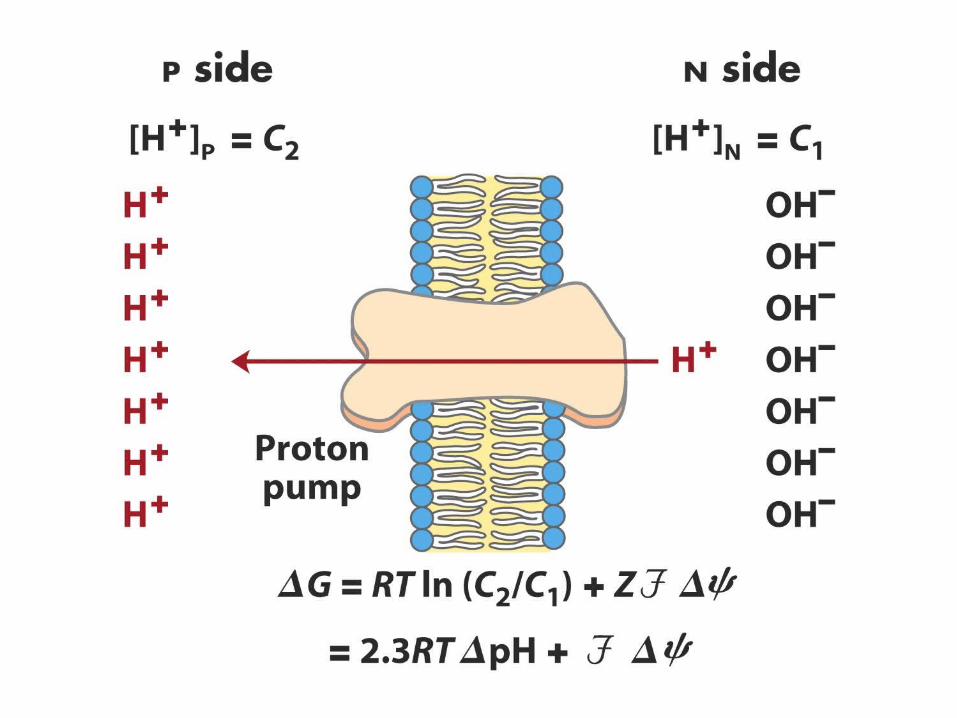
Proton concentration gradient
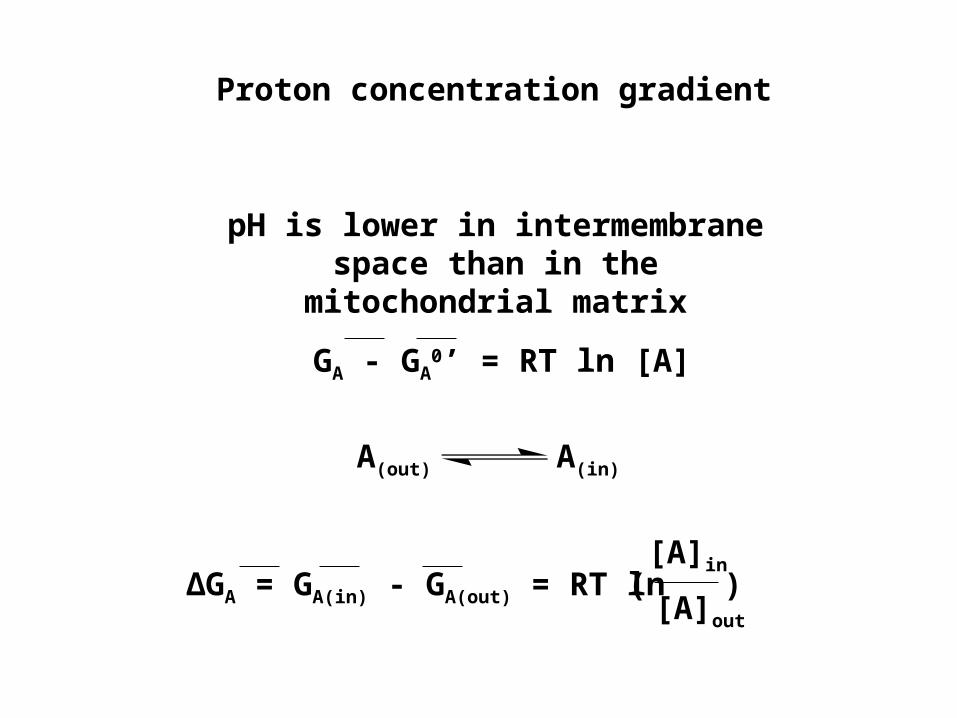
pH is lower in intermembrane space than in the mitochondrial matrix
GA - GA0’ = RT ln [A]
A(out) A(in)
∆GA = GA(in) - GA(out) = RT ln[A]in
[A]out
( )

∆GA = RT ln[A]in
[A]out
( ) + ZAF ∆
If the solute is charged there is another aspect of the equation: electrical potential
Membrane potential = ∆ = (in) - (out)
Free energy is a combination of chemical and electrical potential
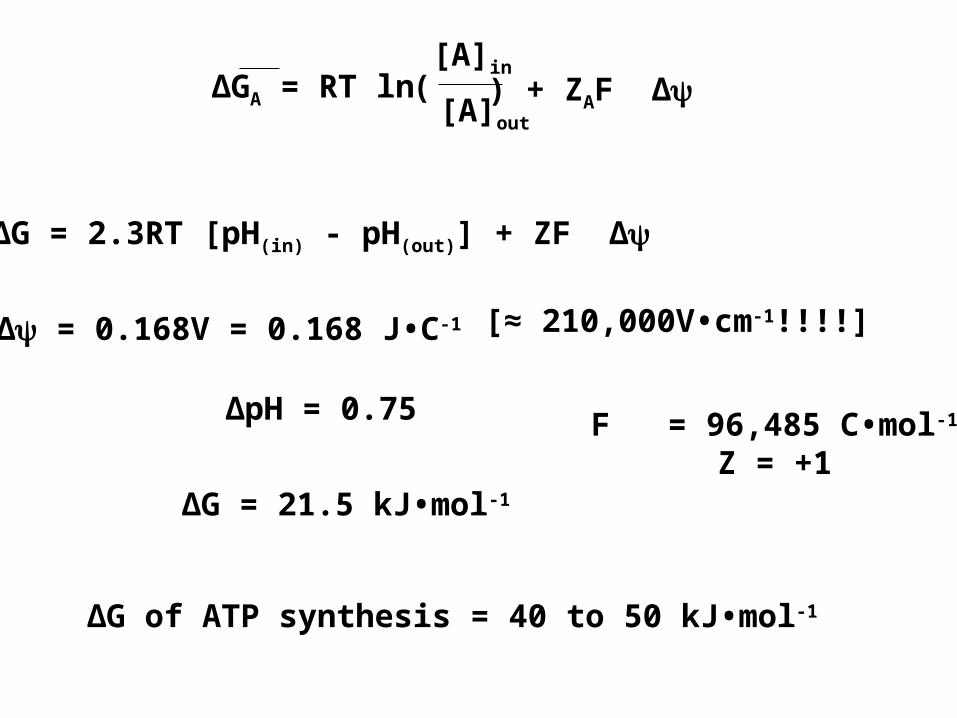
∆GA = RT ln[A]in
[A]out
( ) + ZAF ∆
∆G = 2.3RT [pH(in) - pH(out)] + ZF ∆
∆ = 0.168V = 0.168 J•C-1
∆pH = 0.75
∆G = 21.5 kJ•mol-1
F = 96,485 C•mol-1
Z = +1
∆G of ATP synthesis = 40 to 50 kJ•mol-1
[≈ 210,000V•cm-1!!!!]
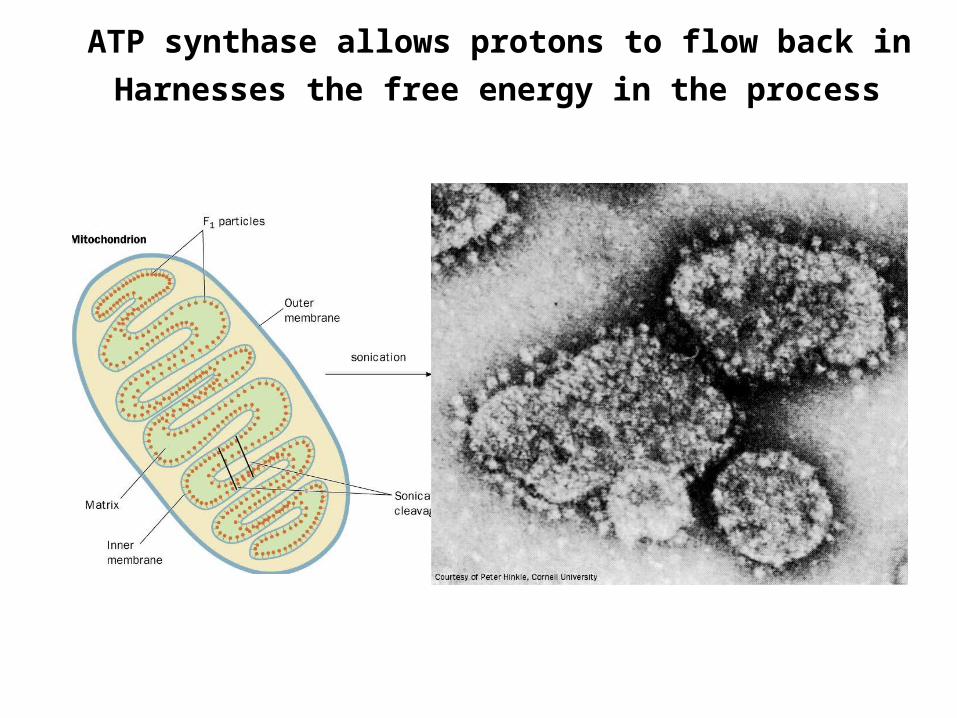
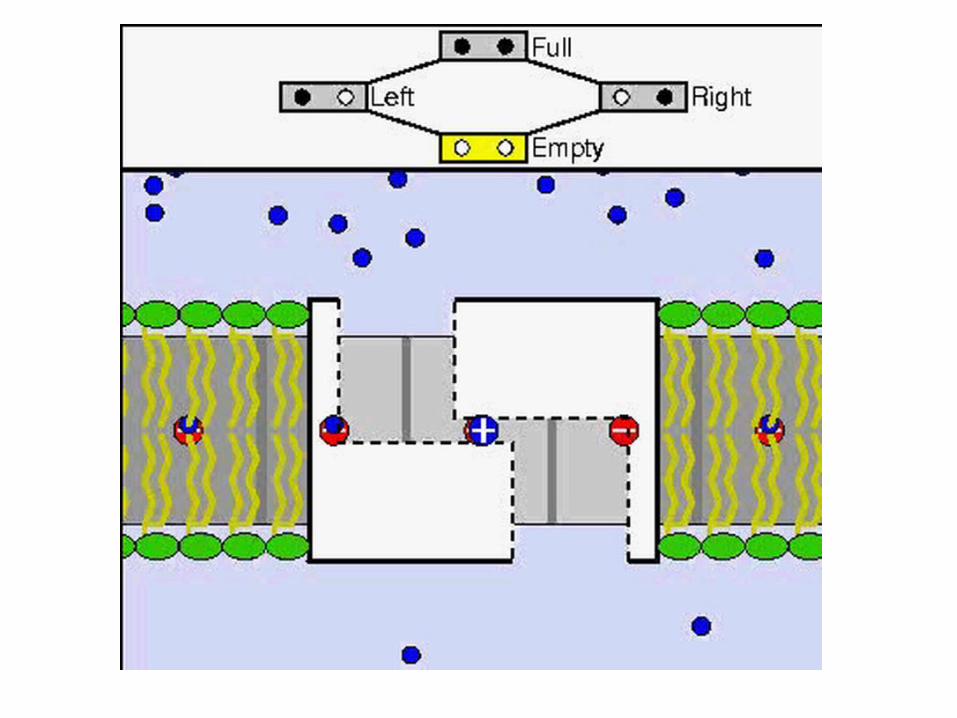
ATP synthase allows protons to flow back in
Harnesses the free energy in the process
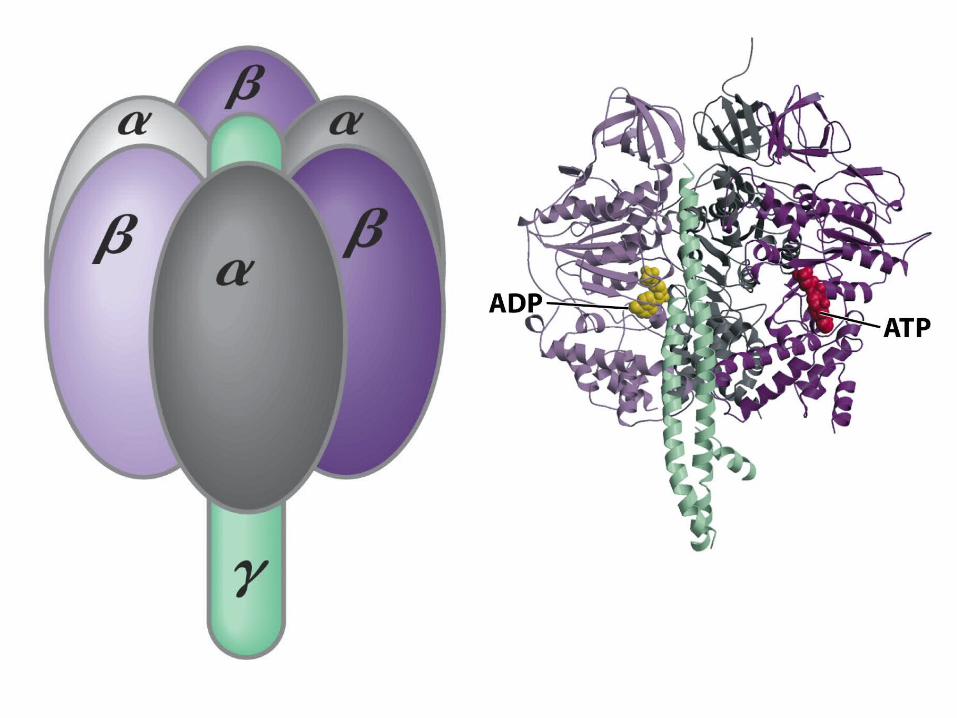

The gamma subunit: the rotor

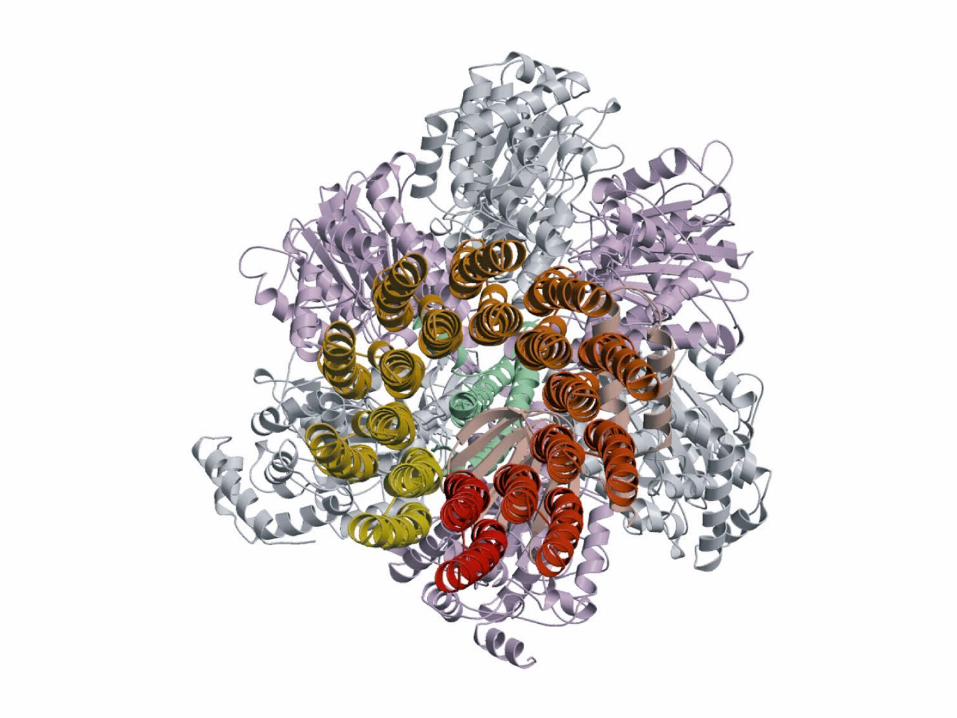
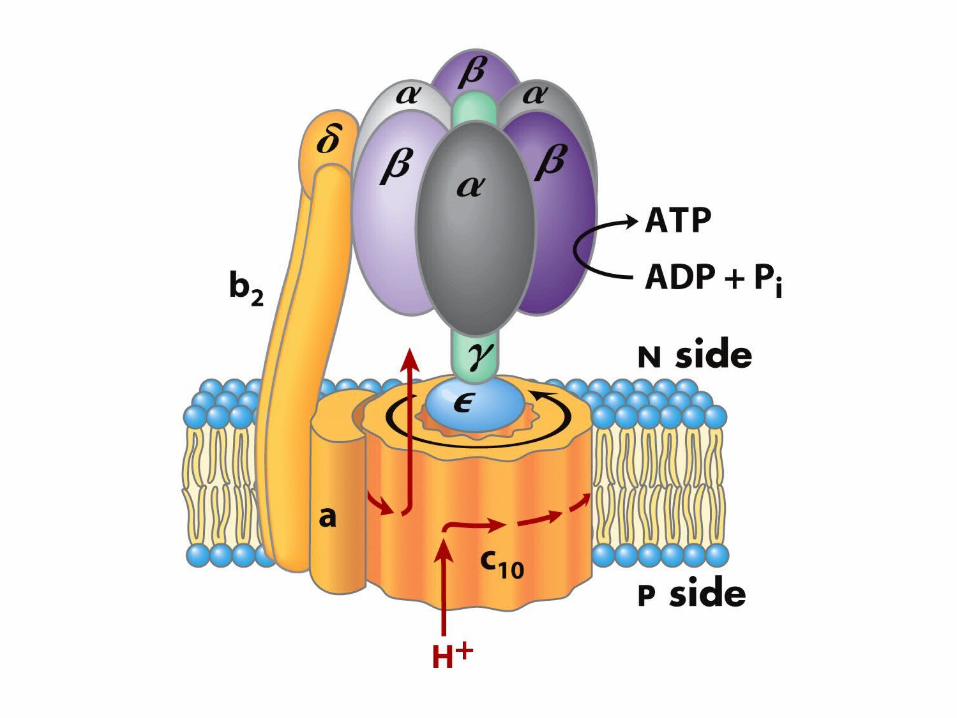
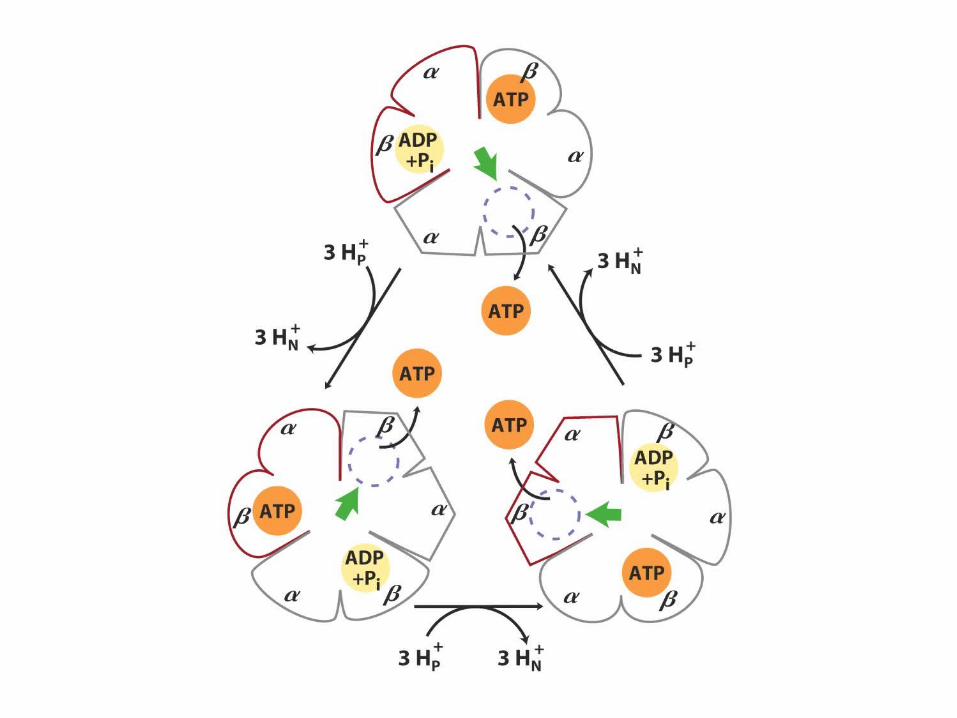



Per glucose10 NADH : 20 e-
2 FADH2 : 4 e-
If 2 electrons enter at complex I4 + 4 + 2 = 10 protons pumped out
If 2 electrons enter at complex II or Glycerol dehydrogenase4 + 2 = 6 protons pumped out
Per glucose120 protons120/3 = 40
+ 4ATP from glycolysis and
TCA
44 ATP! Why only 38?
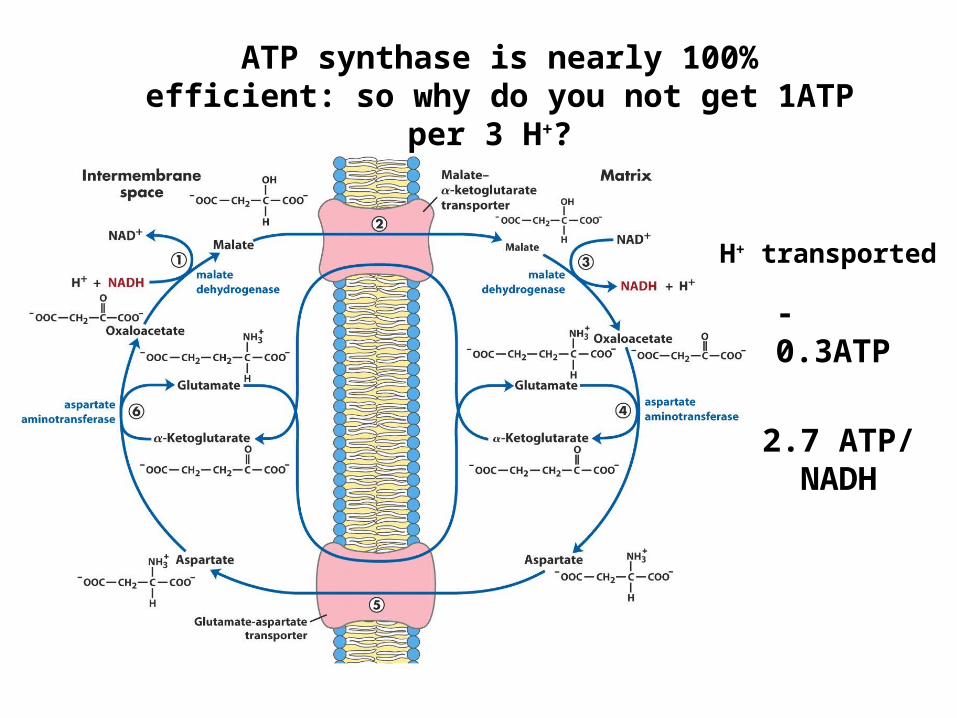
H+ transported
-0.3ATP
2.7 ATP/NADH
ATP synthase is nearly 100% efficient: so why do you not get 1ATP per 3 H+?
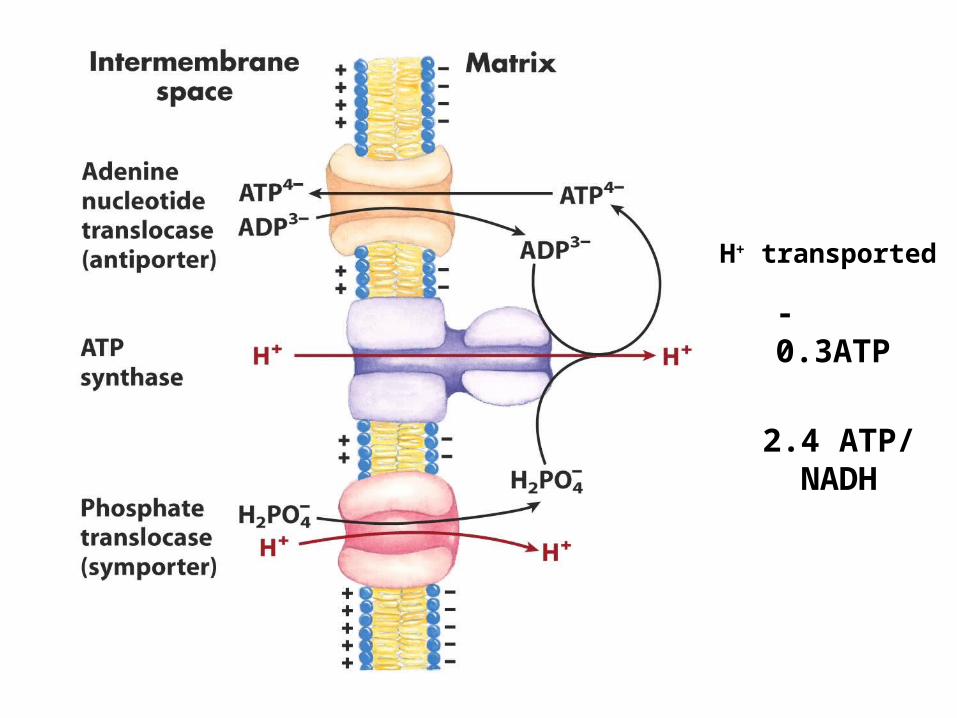
H+ transported
-0.3ATP
2.4 ATP/NADH

What about fatty acid biosynthesis, succinate dehydrogenase and glycerol
phosphate shuttle?
1.5 ATP per pair of electrons...
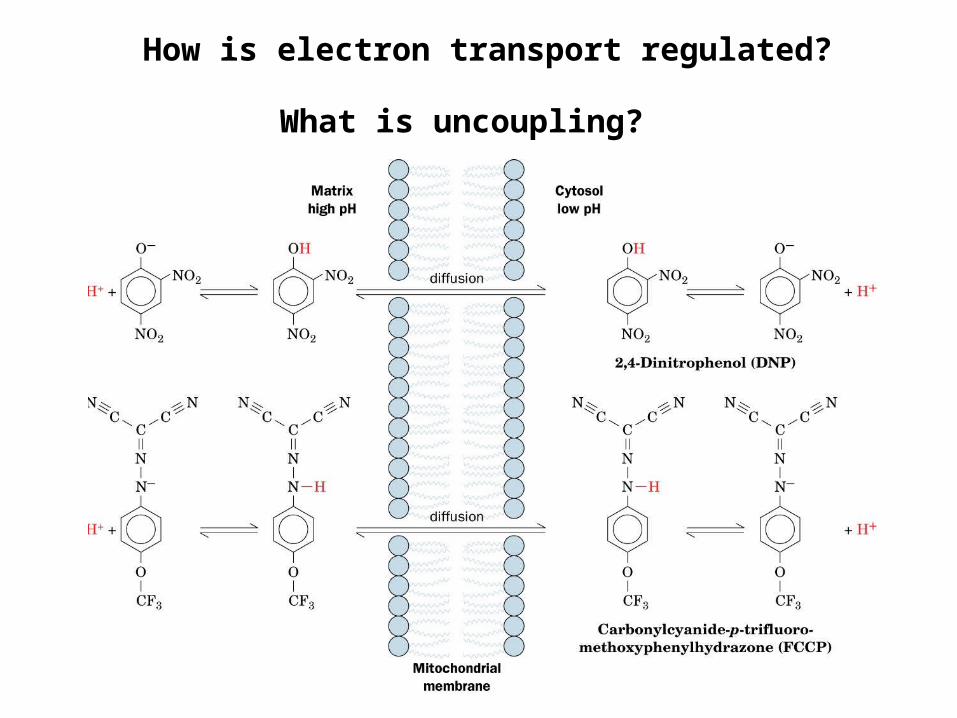
How is electron transport regulated?
What is uncoupling?
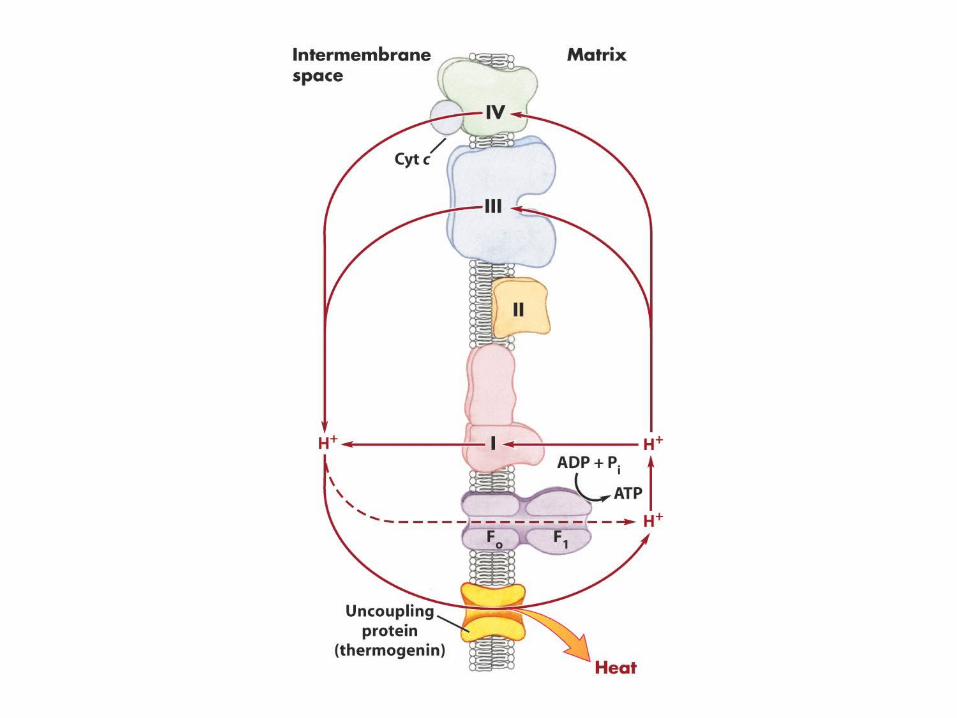
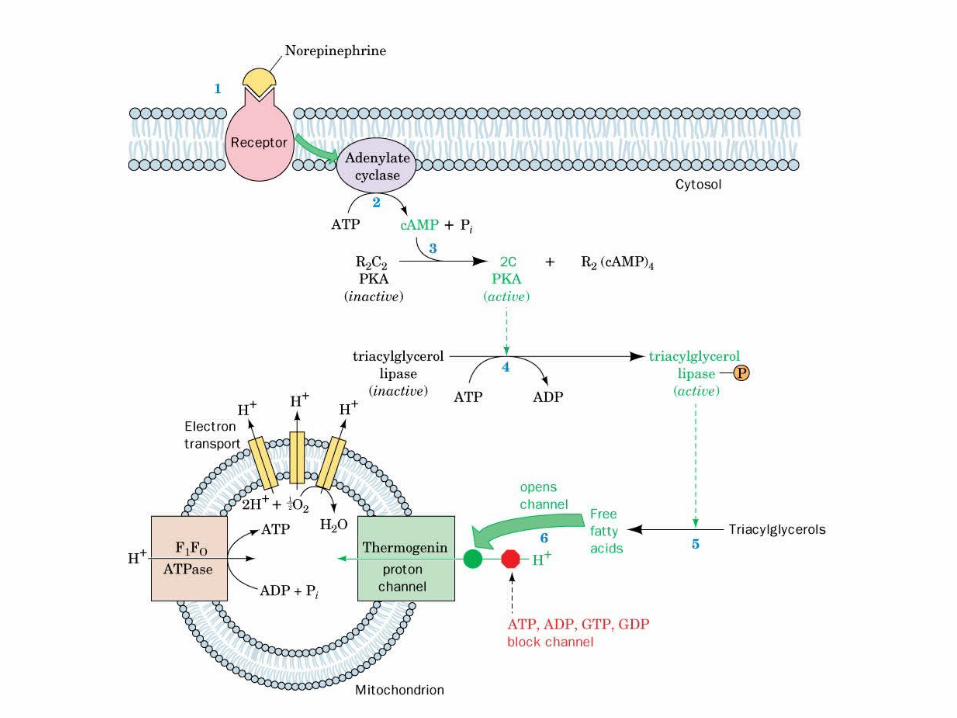
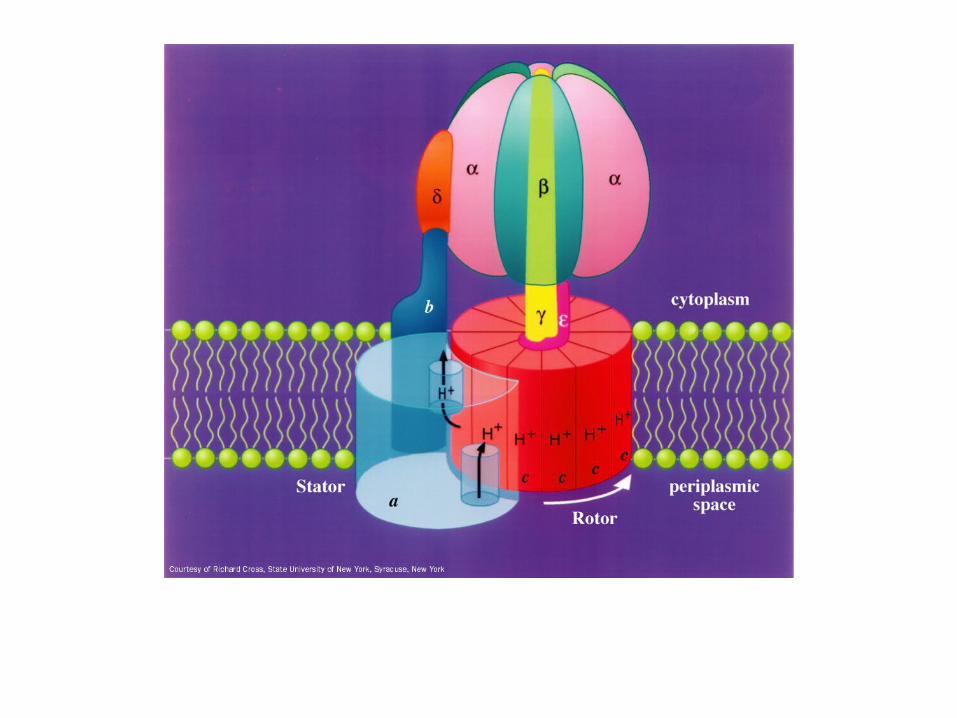
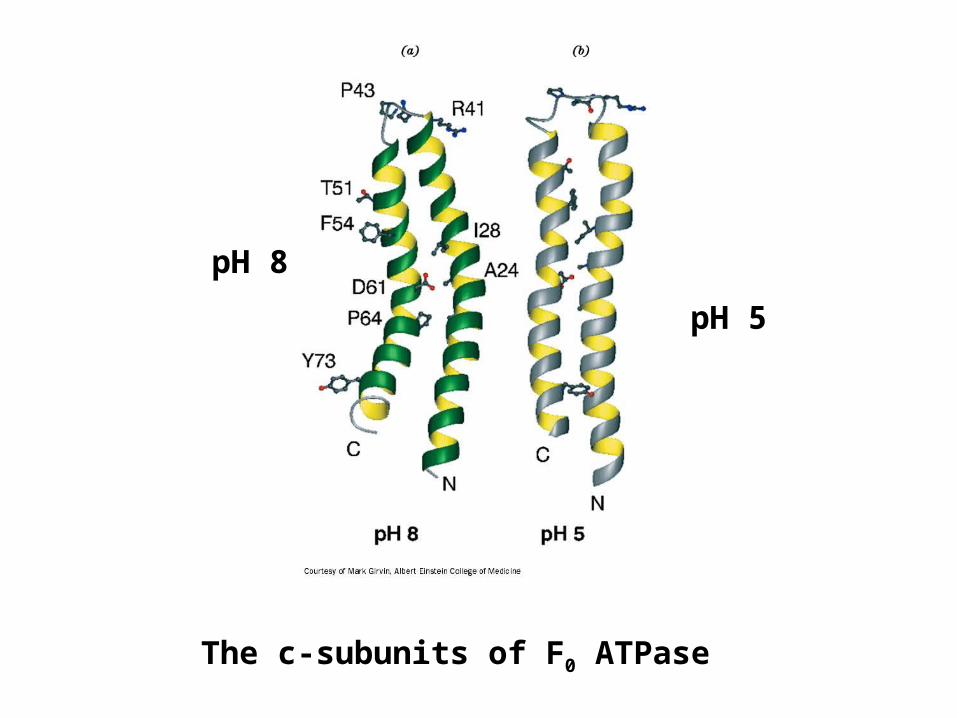
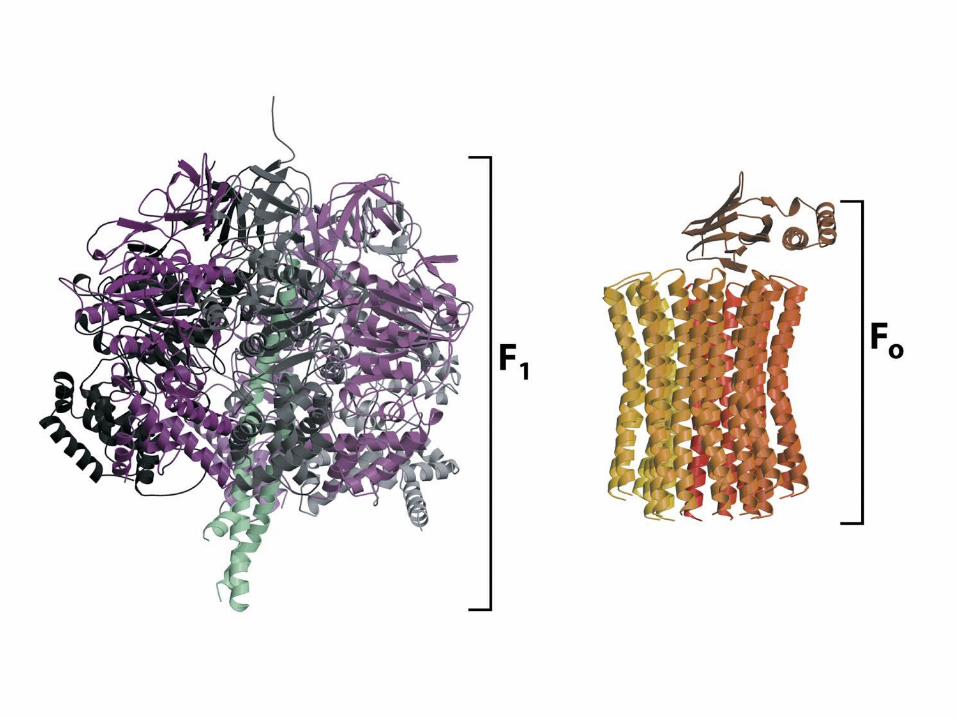
The c-subunits of F0 ATPase
pH 8
pH 5

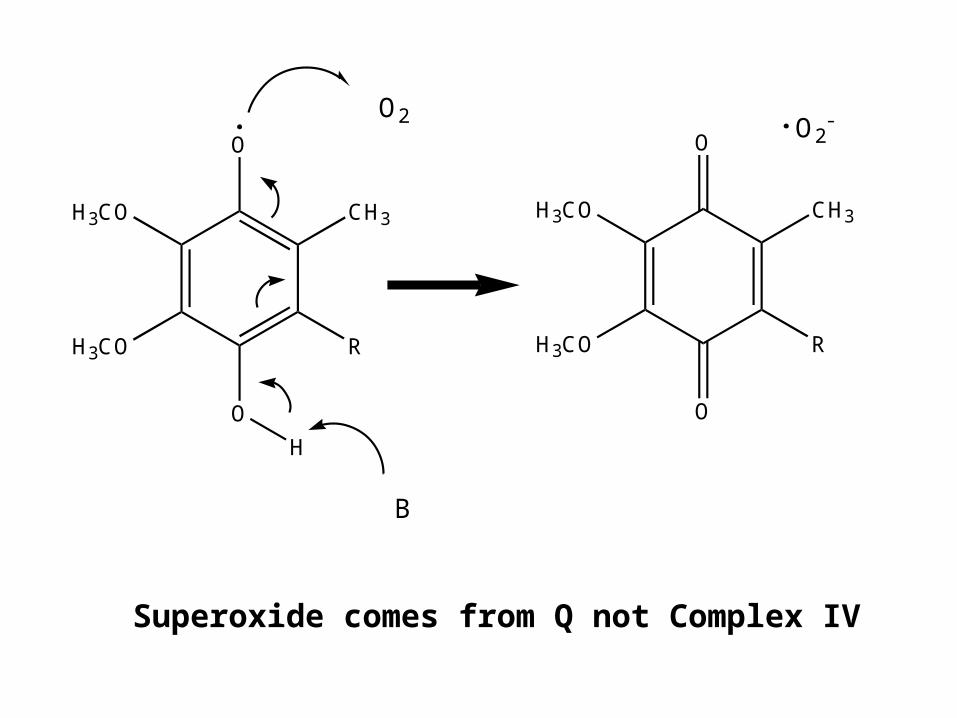
Electrons sometimes leak out of the chain onto molecular oxygen. As much 5% leak
out onto oxygen
Incompletely reduced oxygen is toxic
Reactive oxygen species
Superoxide •O2-
Peroxide O22-
Hydroxyl radical •OH
Where does this occur?
O
O
H3CO
H3CO
CH3
R
O2
O
O
H3CO
H3CO
CH3
R
H
B
O2-
Superoxide comes from Q not Complex IV
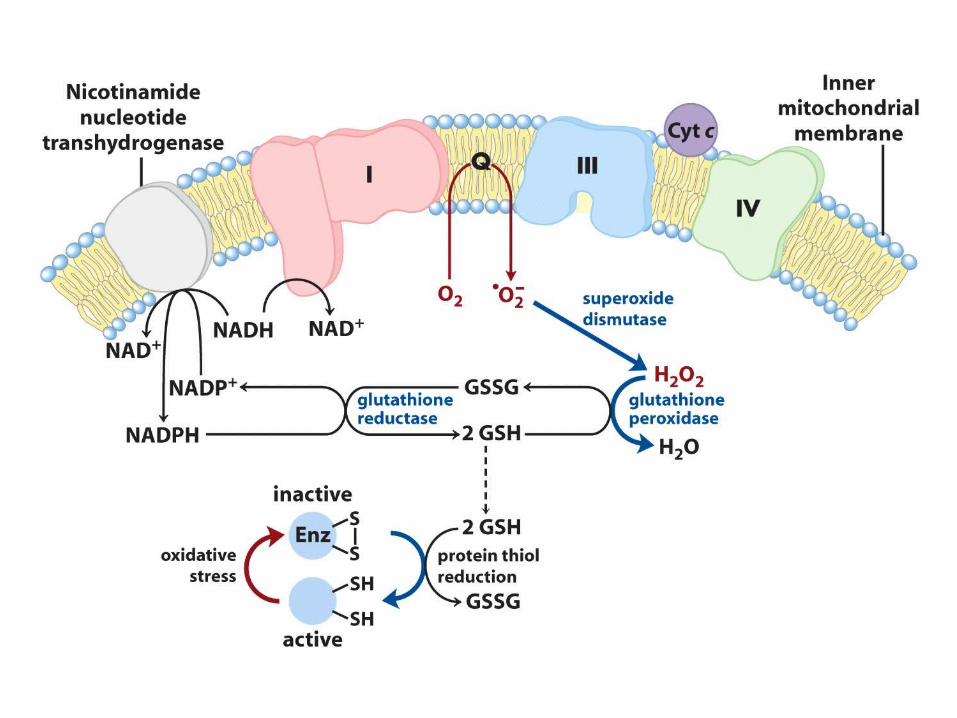

Why does this happen?
The ox-tox hypothesis
Oxygen is soluble in membranes and it can oxidize lipids, albeit slowly. Too much
oxygen is toxic.
Mitochondria detoxify oxygen by reducing it to water. This is how they
were beneficial

What about superoxide and peroxide?
Maybe these are allowed to leak out as signal molecules that apprise the cell of the energetic state….
Maybe the generation of superoxide, which is not soluble in membrane is a way to
detoxifying oxygen……
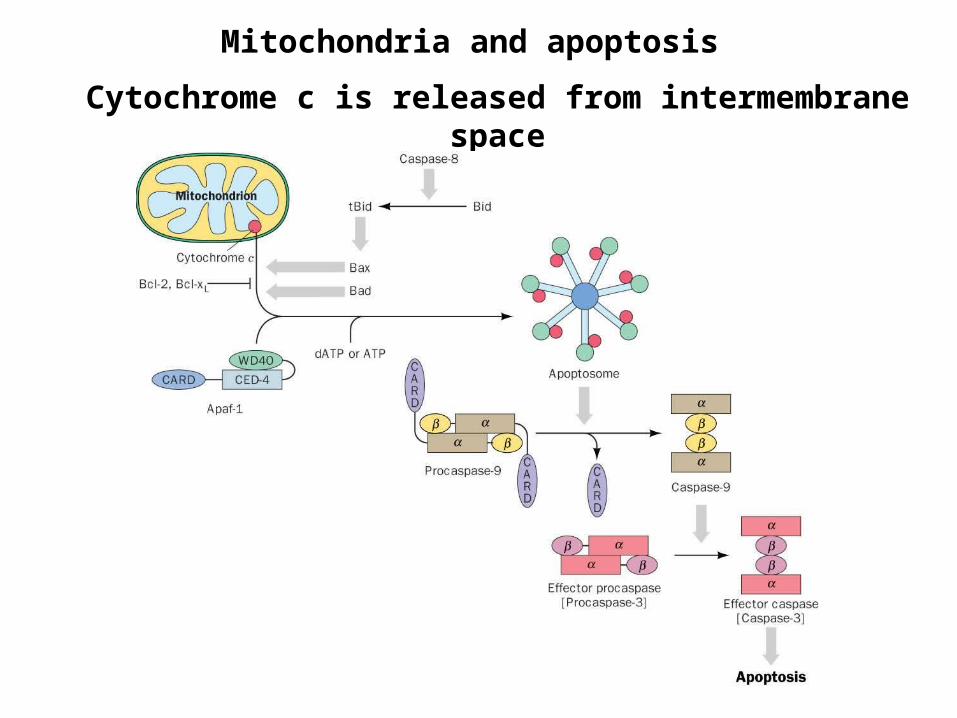
Mitochondria and apoptosis
Cytochrome c is released from intermembrane space












![Inhibitors of Pyruvate Carboxylase - Bentham Open · Inhibitors of Pyruvate Carboxylase The Open Enzyme Inhibition Journal, 2010, Volume 3 9 tumors in rats [18], demonstrating that](https://img.pdfslide.net/doc/110x75/5d4ebe6088c99358578b489f/inhibitors-of-pyruvate-carboxylase-bentham-open-inhibitors-of-pyruvate-carboxylase.jpg)