Embed Size (px)
Citation preview

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
Quadrant - I
4.1. GLYCOGEN METABOLISM
The major site of daily glucose consumption (75%) is the brain via
aerobic pathways. Most of the remainder of is used by erythrocytes, heart muscle, and skeletal muscle. The body gets glucose either directly
from the diet or from amino acids and lactate via gluconeogenesis. Glucose got from these two primary sources either remains soluble in
the body fluids or is stored in a polymeric form, glycogen. Glycogen is considered the main storage form of glucose and is found mostly in liver
and muscle. The kidney and intestines adds minor storage sites. With up to 10% of its weight as glycogen, the liver has the maximum specific
content of any body tissue. Muscle has a much lower quantity of glycogen per unit mass of tissue, but since the total mass of muscle is
so much greater than that of liver, total glycogen stored in muscle is about twice that of liver. Glycogen storage in the liver are considered
the main buffer of blood glucose levels.
OBJECTIVES
- To give an overview of glycogen metabolism
- To understand the factors that control the glycogen metabolism by intracellular signalling
- To understand the function of the pentose phosphate pathway in
production of NADPH and ribose precursors for nucleic acid synthesis.
Summary
Glycogen is the animal storage form of branched poly(glucose). The
storage polysaccharide of animals is glycogen. All cells contain glycogen, but it is most prevalent in the liver and the muscles. Glycogen comprises of glucose molecules linked together with α(1 →4)linkages with α(1 →6)
branch points occurring every 8 to 12 residues. The purpose of the high branched structure is to have many nonreducing ends so that glucose
can be rapidly mobilized in times of metabolic needs.Glycogen
homeostasis involves the concerted regulation of the rate of glycogen synthesis (glycogenesis) and the rate of glycogen breakdown
(glycogenolysis). These two processes are reciprocally regulated such that hormones that stimulate glycogenolysis (e.g. glucagon, cortisol,
epinephrine, norepinephrine) at the same time inhibit glycogenesis. Conversely, insulin, which directs the body to store excess carbon for

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
future use, stimulates glycogenesis while concurrently inhibiting glycogenolysis.
Introduction
Glycogen metabolism is vital for several reasons.
• Glycogen stores in the liver are used to maintain a constant blood
glucose concentration. Glycogen storage is also maintained by muscles as
a reservoir of glucose for strenuous muscular activity.
• The synthesis and degradation of glycogen take place by different
metabolic pathways allowing for reciprocal regulation.
• In addition, the enzymes of glycogen metabolism are under hormonal
regulation.
The biochemical pioneers of glycogen metabolism were the Cori’s, Carl
and Gerty, a husband and wife team. They demonstrated that the
glycogen is broken down by phosphorolysis. The complete breakdown
process of glycogen breakdown is:
(glucose)n → glucose-1-phosphate + (glucose)n-1.
Processes of glycogen synthesis is:
(glucose)n-1 + UDP-glucose → (glucose)n
The glycogen breakdown and synthesis is controlled by two key enzyme
(glycogen phosphorylase and glycogen synthase) activities which are
activated/inactivated by allosteric regulation and phosphorylation /
dephosphorylation.
The storage form of glucose in most eukaryotic cells (except plants) is
glycogen, a large highly branched polysaccharide containing glucose units
joined by α-1→4 and α-1→6 glycosidic bonds. Both the liver and muscle
store glycogen and hence have the necessary anabolic and catabolic
enzymes. The degradation and synthesis of glycogen occurs in the cytosol
and the substrate for these reactions is the free ends of the branched
polymer. The large number of branch points in glycogen results in the

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
generation of multiple nonreducing ends that provide a highly efficient
mechanism to quickly release and store glucose.
4.1.1.GLYCOGEN SYNTHESIS - Luis Leloir discovered the glycogen
biosynthetic pathway.
Glycogen + UDP-glucose → Glycogenn+1 + UDP
If we compare the synthetic pathway to the degradative pathway it is
clear that the glycogen biosynthesis is not merely the reversal of the
degradative pathway. The two pathways are distinct providing a
mechanism for reciprocal control.
A. UDP-glucose formation by UDP-glucose pyrophosphorylase - In the
glycogen synthesis pathway, at first, the uridine diphosphate(UDP) is
attached to glucose. The reaction is catalyzed by UDP-glucose
pyrophosphorylase.
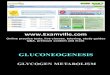
B. Glycogen synthesis by glycogen synthase
1.The glycosidic bond between glucose and UDP in UDP-glucose is
hydrolyzed.
2. The glucose unit of UDP-glucose is transferred to the C4-OH group on
one of glycogen’s non-reducing ends to form an α (1→4) glycosidic bond.
Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
C. Glycogen Branching -
Glycogen synthetase can synthesize α(1→4) linkages. Another enzyme is
required to form the α(1→6) linkages that make the branches. The
branching enzymes takes a block of seven or so residues of a nonreducing
end and transfers these seven residues to an interior site and creates an
α(1→6) linkage. The chain that contributes the seven residues must be at
least 11 residues long, and the new branch point must be at least four
residues away from pre-existing branch points. About 7 units of the non-
reducing end of α-amylose chain are removed at α(1→4) linkage, and
reattached to the C6 of other α-amylose chain by α(1→6) linkage. This
transfer is carried out by amylo-(1,4→1→6)-transglycosylase (branching
enzyme).

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
4.1.2.GLYCOGENOLYSIS
Three enzymes are required for glycogen breakdown
1. Glycogen phosphorylase

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
(Glycogen)n + Pi ↔
(glycogen)n-1 + G1P (n residues).
Glycogen phosphorylase catalyzes this reaction. Glycogen phosphorylase
catalyzes the successive phosphorolysis of glucose residues from a
nonreducing end. The phosphorylated glucose cannot diffuse out of the
cell. Glycogen phosphorylase requires a pyridoxl-5’-phosphate cofactor.
The pyridoxl-5’-phosphate cofactor is covalently bound to a lysine residue
via a Schiff base.
Glycogen phosphorylase degrades glycogen’s α(1→4) glycosidic bonds
sequentially until it gets four residues away from a α(1→6) branch point
when its action ceases and another enzyme is required to remove the
branches.This enzyme is called the debranching enzyme. This enzyme
releases a glucose unit one by one until it reaches ~ five units (limit
branch) from a branch point. There are two forms of the enzyme
phosphorylase :Phosphorylase a → E-O-PO3 and
Phosphorylase b → No phosphate attachment.

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
Reaction mechanism includes cleavage of the glycosidic bond by an acid
catalysis to form G1P. The other enzyme active site, the α-1,6-
glucosidase, cleaves the α(1→6) glycosidic linkage. After which the
glycogen debranching enzyme removes branches so that glycogen
phosphorylase can complete reaction. The glucose-1,6-bisphosphate
formed can dissociate out of the active site before transferring the C1
phosphoryl group. When this happens, the active site serine must be
rephosphorylated.
There is an enzyme called phosphoglucokinase which phosphorylates
glucose-1-phosphate to form glucose-1,6-bisphosphate which can bind to
the dephosphorylated enzyme and transfer the C1-phosphoryl group to
reactivate the enzyme and produce glucose-6-phosphate.
Phosphoglucomutase - G1P produced from the glycogen breakdown must
be converted to G6P in order to enter glycolysis or to produce glucose in
liver. Phosphoglucomutase catalyzes the conversion of G1P to G6P.
The reactions of glycogen biosynthesis are shown below.
1. Glucose-6-phosphate→ Glucose-1-phosphate ( Phosphoglucomutase)
2. Glucose-1-phosphate + UTP → UDP-Glucose + PPi (UDP-glucose
pyrophosphorylase)
3. PPi + H2O → 2Pi ( Inorganic pyrophosphatase)
4. UDP-Glucose + Glycogen UDP + Glycogen n+1 (Glycogen synthase)
5. UDP + ATP → UTP + ADP (Nucleotide diphosphokinase)
Sum: Glucose-6-phosphate + ATP + Glycogen + H2O → Glycogen n+1 +
ADP + 2Pi
90% of glycogen is phosphorylytically cleaved into glucose-1-phosphate
which is isomerized into glucose- 6-phosphate.10% are the branched
residues which are hydrolyzed into glucose which can be phosphorlated
into glucose-6-phosphate.The complete oxidation of glucose-6-phosphate
through glycolysis, the citric acid cycle and oxidative phosphorylation
yields 38 molecules of ATP. The overall efficiency of storage is 97%.
4.1.3. Regulation of glycogen metabolism

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
Allosteric regulation of glycogen synthase and phosphorylase Hormonal control of glycogen metabolism
Regulatory differences between liver and muscle phosphorylase
Allosteric regulation of glycogen synthase
and phosphorylase
The allosteric regulatory effects exercised by glucose-6-phosphate, ATP
and AMP on glycogen phosphorylase and glycogen synthase make good physiological sense. Exhaustion of ATP is an excellent reason to release
glucose from the store in order to make some more. On the other hand, glucose-6-phosphate will be plentiful when glucose itself is abundant, and
therefore signals an opportunity for replenishing the glycogen stores.
2. Hormonal control of glycogen metabolism

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
. Insulin is the anabolic hormone and promotes the storage of fuels. It
activates the storage of glucose as glycogen in the liver and muscles. The conversion of glucose into triacylglycerides and the storage of
triacylglycerides in the adipose tissue is also promoted by insulin. Glucagon is the hormone that promotes the mobilization of fuels.
Glucagon acts to maintain glucose availability in the absence of dietary glucose by stimulating the release of glucose from the liver. Glucogon
stimulates glycogenolysis and gluconeogenesis. Glucagon also activates
the mobilization of fatty acids from the adipose tissue. The sites of glucagon action are principally in the liver and adipose tissue. The release
of glucagon is suppressed by insulin and glucose. The lowest levels of glucagon occur after a high carbohydrate meal. All of glucagon’s effects
are opposed by the effects of insulin. The stimulation of insulin release suppresses the release of glucagon.

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
The glycogen synthase and the phosphorylase respond in opposite ways to phosphorylation: The synthase is inactivated, whereas the
phosphorylase is activated.
3. Regulatory differences between liver and
muscle phosphorylase
Liver enzyme Muscle enzyme
Inhibition by glucose + −
Activation by Ca2+ − +
Activation by AMP even when
unphosphorylated
− +
There are regulatory differences between glycogen phosphorylase in muscle and liver. The liver enzyme inhibited by glucose. But this does not
inhibit the muscle enzyme. Ca2+ stimulates the muscle enzyme but not the liver enzyme. Recall that Ca2+ is also the trigger for muscle
contraction; the simultaneous stimulation of glycogen breakdown therefore anticipates an increased demand for ATP.
Inter organ relationships in glycogen metabolism
Liver glycogen utilization

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
Muscle glycogen utilization The Cori cycle
The two tissues that have the most significant pools of glycogen are the liver and skeletal muscle. Liver glycogen is turned over quickly;
it serves as the major reserve of blood glucose during short-term fasts. After liver glycogen is depleted, muscle glycogen can be
drawn down; this, however, requires some roundabout metabolic trickery.
Liver glycogen utilization
The liver mobilizes glucose from its glycogen store via glycogen
phosphorylase and phosphoglucomutase, which yields glucose-6-phosphate. The latter is transported to the endoplasmic reticulum, where
it is dephosphorylated. Glucose is taken back to the cytosol and released into the bloodstream.Some of the glucose will be rephosphorylated before
making it out of the cell, creating the futile cycle.However, the dominant glucose phosphorylating enzyme in the liver is glucokinase. This enzyme
has fairly low affinity for glucose; therefore, enough glucose will escape
rephosphorylation and be released into the bloodstream.
Muscle glycogen utilization

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
Muscle glycogen primarily serves the energy needs of muscle tissue
itself.During prolonged physical exercise, maximum of it is broken down to glucose-6-phosphate and then consumed via the usual pathways right
within the cells that stored it. As discussed above, this usage is facilitated
by calcium-mediated activation of glycogen phosphorylase.
Under suitable conditions, that is, prolonged fast without physical
exercise, muscle glycogen can also contribute to the replenishment of blood glucose. However, even though muscle cells have been shown to
express glucose-6-phosphatase and thus are, in principle, able to produce free glucose, they should find it difficult to release it. The reason is
muscle contains hexokinase, which has a much greater substrate affinity than glucokinase and therefore will keep the intracellular level of free
glucose well below the extracellular concentration. The net transport of glucose should therefore be directed inward at all times.

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
The Cori cycle
While skeletal muscle relies on oxidative metabolism most of the time,
some other tissues, especially red blood cells and lymphocytes, which together depend mostly or even exclusively on anaerobic glycolysis even
under aerobic conditions. The peripheral tissues releases lactate and it is scooped up by the liver, which converts it back to glucose through
gluconeogenesis. The process is known as the Cori cycle, named after its discoverers Carl and Gerti Cori, who worked it out as early as 1929.
Skeletal muscles produce lactate at a very much higher rate during short bouts of maximal exercise when the ATP demand exceeds the capacity for
aerobic metabolism. During intense exercise, the cardiac blood output is
diverted from the visceral organs to skeletal muscle. Thus, when ATP demand exceeds the oxygen supply of skeletal muscle, the oxygen
shortfall would be even greater in the liver, should it indeed attempt to make enough ATP for sustaining the muscle through gluconeogenesis;
and even with adequate oxygen, its capacity for producing glucose would fall far short of the muscles’ voracious appetite.
4.2.HEXOSE MONOPHOSPHATE SHUNT
The hexose monophosphate pathway(Pentose phosphate pathway)is used
to produce the NADPH required for a variety of cellular functions. This
NADPH production is controlled by the relative levels of NADP and NADPH
that alter the activity of glucose-6-phosphate dehydrogenase. This is the
control enzyme for the pathway. The ribose-5- phosphate required for

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
nucleotide and nucleic acid biosynthesisis also synthesized in the hexose
monophosphate pathway. All the enzymes after 6-phosphogluconate
dehydrogenase step in hexose monophosphate pathway are fully
reversible. These enzymes let the synthesis of ribose-5-phosphate from
the glycolytic intermediates fructose-6-phosphate and glyceraldehyde-3-
phosphate, and the conversion of ribose-5-phosphate into glycolytic
intermediates. NADPH is part of the mechanism that red blood cells use to
prevent oxygen-mediated damage to their membranes and proteins, and
to maintain haemoglobin in the oxidation state required for oxygen
transport.
4.2.1.Hexose monophosphate shunt
Use and Importance:
The hexose monophosphate pathway is used for production of NADPH
from NADP. The biosynthetic reactions such as cholesterol synthesis, fatty
acid synthesis, drug reduction requires NADPH.It also acts as a cofactor
for some non-synthetic enzymatic reactions.Furthermore, it is used for
the production of ribose for nucleotide and nucleic acid synthesis.
The hexose monophosphate pathway also permits the entry of some
carbohydrates into the glycolytic pathway (especially ribose, but also
some others), and hence acts as a connection route between different
pathways.
4.2.2.Overview of the shunt pathway
The HMP shunt pathway has oxidative and non oxidative phases. In
the oxidative phase, glucose-6-phosphate is oxidised with the generation
of 2 molecules of NADPH, and one molecule of pentose phosphate, with
the release of one molecule of CO2. In the non oxidative phase, the
pentose phosphate is transformed to intermediates of glycolysis.

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
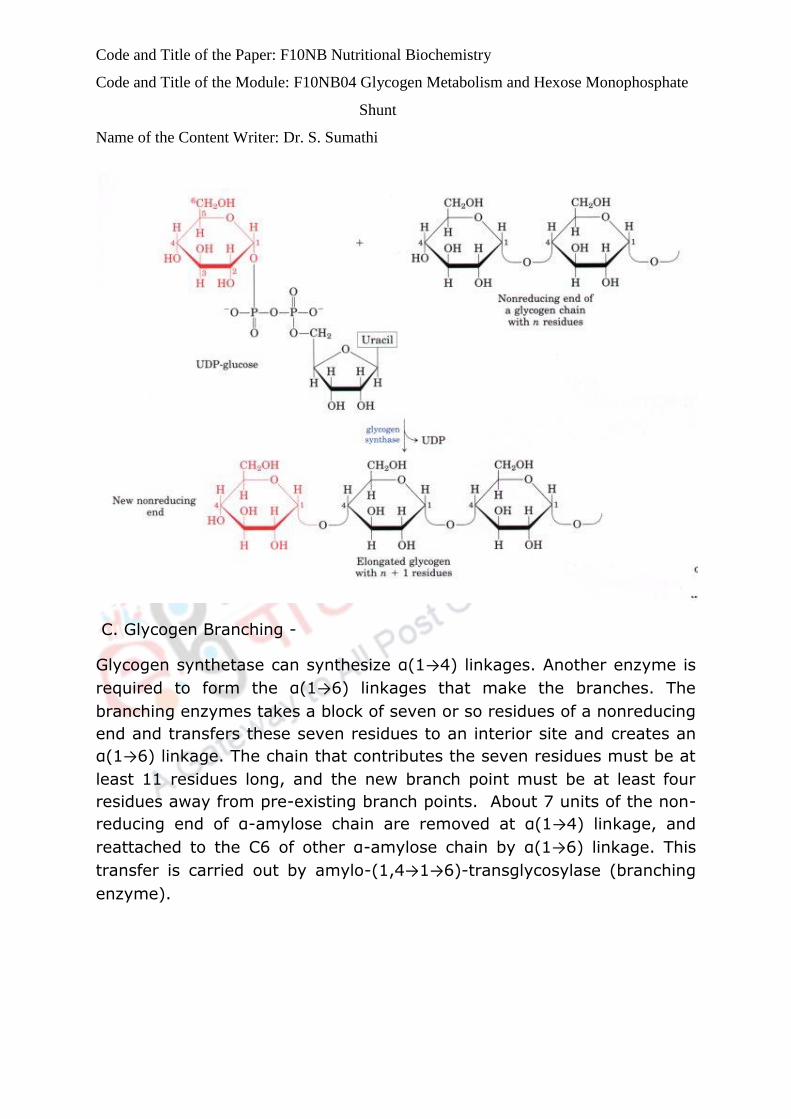
A. Oxidative phase
Step 1
Glucose-6-phosphate is oxidised by NADP+ to produce 6-phosphoglucono
lactone. In this reaction one molecule of NADPH is formed. Regulation is
effected by glucose-6- phosphate dehydrogenase enzyme.
Step 2
The lactone is hydrolysed by lactonase to form 6-phosphogluconate.
Step 3
This is an oxidative step together with decarboxylation.
The enzyme is 6- phosphogluconate dehydrogenase.
Here Ribulose 5 phosphate is formed and a second molecule ofNADPH is
generated.
Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
B. Non oxidative phase

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
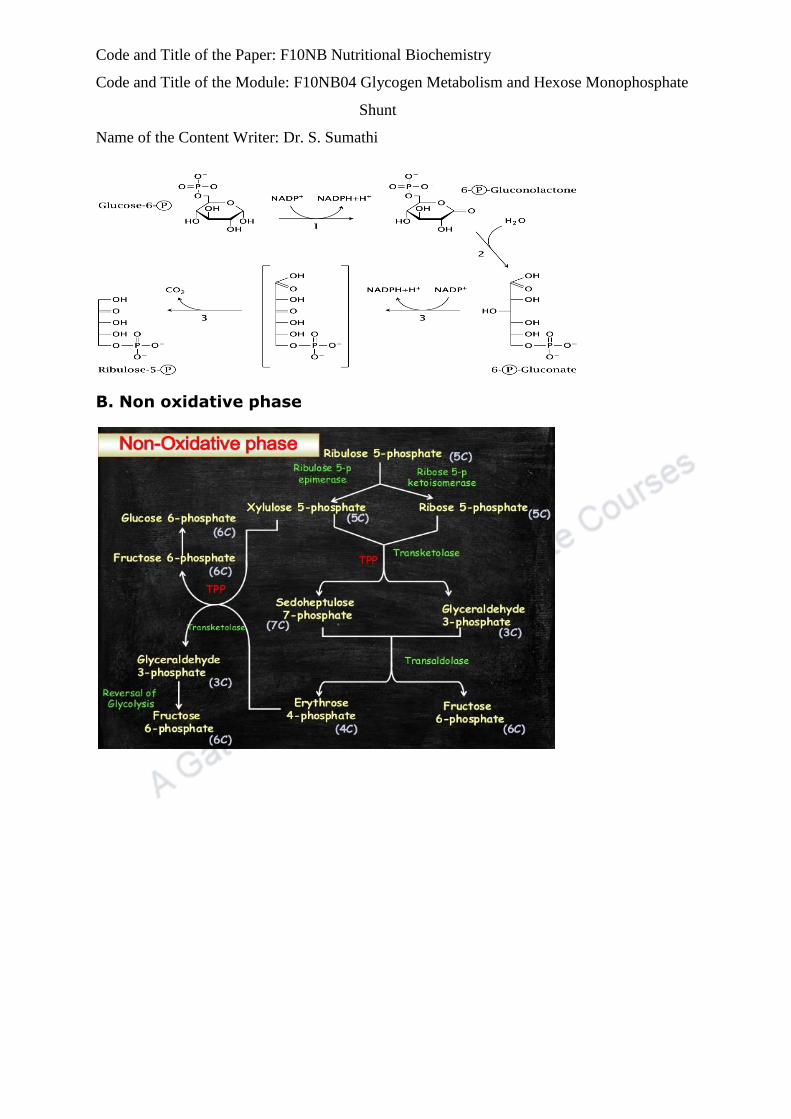
Step 4: Isomerisation The ribulose-5- phosphate is then isomerised to
ribose-5 phosphate or epimerised to Xylulose-5-phosphate.
Step 5: Transketolase reaction Transketolase is a thiamine
pyrophosphate (TPP) enzyme. This enzyme transfers two carbon unit with
keto group from Xylulose-5 phosphate to ribose-5-phosphate to a form 3
carbon (glyceraldehyde-3-phosphate) and 7 carbon (sedoheptulose-7
phosphate).
Step 6: Transaldolase reaction involves transfer of a 3 carbon unit
from sedoheptulose-7-phostate to glyceraldehyde-3-phosphate to form
fructose-6-phosphate.

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
Step 7: second transketolase reaction In one more transketolase
reaction a 2C unit is transferred from Xylulose-5- phosphate to erythrose-
4-phosphate to form fructose-6- phosphate and glyceraldehyde-3-
phosphate.
Step 8: Regeneration of glucose -6- phosphate.
Two molecules of glyceraldehyde-3-phosphate formed in step 7 are
condensed to form one fructose-6-phosphate.This fructose-6-phosphate is
then converted to glucose-6-phosphate (reversal of step 2 of glycolysis).
4.2.3.SIGNIFICANCE
1.Pentose phosphate pathway protects cells against reactive oxygen
species (ROS)which includes molecular oxygen and partially reduced,
reactive forms of oxygen. Reduction of molecular O2 in a series of
one‐electron steps yields superoxide, hydrogen peroxide, hydroxyl radical,
and water which forms the reactive oxygen species (ROS)
2. NADPH and glutathione in protects the cells against highly reactive
oxygen derivatives. The cells are protected by reduced glutathione (GSH)
by destroying hydrogen peroxide and hydroxyl free radicals. The
regeneration of GSH from its oxidized form (GS‐SG) requires the NADPH
produced in the glucose 6‐ phosphate dehydrogenase reaction.
3.Importance in RBCs: The role of red blood cells in O2 transport makes
them liable for oxidative damage by H2O. H2O2 causes both oxidation of
iron in hemoglobin to form methemoglobin and lipid peroxidation in RBCs.
The production of NADPH is the major role of HMP in red blood cells that
protect these cells from oxidative damage by providing reduced
glutathione for removal of H2O2.

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
4.Chemically NAD and NADP are very similar molecules. These two
molecules are structurally closely identical (the only difference is the
phosphate on the 2´-position of the adenosine ribose of NADP instead of
the free hydroxyl at this position in NAD). In catabolic processes (such as
the TCA cycle), NAD is used to accept electrons, whereas NADPH is
primarily used to donate electrons for synthetic reactions. NADPH can
thus act as a strong driving force for otherwise unfavorable reactions
because of both the concentration differential and the fact that NADPH
contains more energy than NADP. Variety of electron donation reactions
uses NADPH. In some cell types, NADPH is required to maintain normal
functioning in addition to its involvement in synthetic processes. As an
example, red blood cells, that perform very few synthetic reactions, yet
require significant amounts of NADPH. NADPH plays two critical roles in
red blood cells. These two roles are related to the oxygen-transport
function of the red blood cell. The first role is related to glutathione:
erythrocytes require NADPH to maintain their levels of reduced
glutathione.
5. Oxygen is toxic and without reduced glutathione, peroxides that are
spontaneously formed from molecular oxygen would oxidize the lipid
components of the red blood cell membranes. Furthermore, peroxides
have a tendency to damage hemoglobin, causing precipitation of the
protein. Insoluble aggregates of hemoglobin have severely impaired
oxygen carrying capacity, and insoluble protein aggregates also tend to
be inflexible enough to prevent the normal deformations of the red blood
cell. Two enzymes are essential to deal with the peroxides -Glutathione
peroxidase converts the peroxide to an alcohol using glutathione.
Glutathione reductase then uses NADPH to regenerate the reduced
glutathione. The second role of NADPH in red blood cells is associated to
hemoglobin. Oxygen tends to oxidize the hemoglobin iron from +2 to the
more stable +3 oxidation state (resulting in methemoglobin). This is
problematic: the +3 state of heme iron binds oxygen very poorly. NADPH
is used to supply reducing equivalents to methemoglobin reductase, the
enzyme which returns the haemoglobin to the +2 oxidation state.
6.Glucose-6-phosphate dehydrogenase deficiency:Glucose-6-phosphate
dehydrogenase deficiency is a very common genetic abnormality that is
found in 5-10% of the global population. The effect of the deficiency is a

Code and Title of the Paper: F10NB Nutritional Biochemistry
Code and Title of the Module: F10NB04 Glycogen Metabolism and Hexose Monophosphate
Shunt
Name of the Content Writer: Dr. S. Sumathi
decreased ability to form NADPH in red blood cells. Malaria parasites live
inside red blood cells. The normal lifespan of red blood cells are about 120
days. The lifespan of the red blood cell is shortened by a number of
mutations (glucose-6-phosphate dehydrogenase deficiency, sickle cell
anaemia, and the thalassemias), causing death of the malaria parasites
that have not reached maturity. In glucose-6-phosphate dehydrogenase
deficiency, the most common mutations seem to decrease the half-life of
the enzyme that is effective in reducing the life-span of the cell since red
blood cells lack the ability to synthesize new proteins. The erythrocytes in
individuals with glucose-6-phosphate dehydrogenase deficiency are
asymptomatic unless challenged with oxidants; oxidant exposure,
however, results in hemolyticanemia (a reduced number of red blood cells
due to lysis of the cells).
4.2.4.Regulation of pentose phosphate pathway
• The entry of glucose 6‐phosphate into the pentose phosphate pathway
is controlled by the cellular concentration of NADPH. NADPH is a strong
inhibitor of glucose 6‐ phosphate dehydrogenase. As NADPH is used in
various pathways, inhibition is relieved, and the enzyme is accelerated to
produce more NADPH.The synthesis of glucose 6‐phosphate
dehydrogenase is induced by the increased insulin/glucagon ratio after a
high carbohydrate.