Embed Size (px)
Citation preview

Quantitative molecular detection of Salmonella typhimurium in soiland demonstration of persistence of an active
but non-culturable population
Peter Marsh, Nathan Z. Morris, Elizabeth M.H. Wellington *Department of Biological Sciences, University of Warwick, Coventry CV4 7AL, UK
Received 5 February 1998; received in revised form 7 August 1998; accepted 20 August 1998
Abstract
Several methods were used to study survival of Salmonella typhimurium LT2 in soil. An ion exchange resin-based extractionmethod was used to concentrate biomass from soil, from which DNA was extracted in order to quantify a Salmonella-specificsequence by a quantitative polymerase chain reaction (QPCR). S. typhimurium LT2 was detected at a minimum density of 103
cells g31 in non-sterile soil, and the method proved to be specific for this organism in microcosm experiments. Non-sterile soilmicrocosms were inoculated with S. typhimurium LT2 at 107 cfu g31 dry soil and survival monitored at three matric potentials.Viable counts on XLD indicated a rapid decline in cell density over 54 days, whereas direct counts of active cells using therespiration-sensitive dye 5-cyano-2,3-ditolyl tetrazolium chloride (CTC) remained relatively constant. XLD did notunderestimate culturable cells in comparison to non-selective agar. QPCR revealed that the number of salmonella targets(H-li) remained constant up to day 13. After that time, a decrease occurred, corresponding to that of the plate counts, due to anincrease in resistance of the cells to lysis, as incorporation of a lysozyme step into the DNA extraction method allowed moreefficient DNA extraction. This resulted in a constant QPCR signal over 54 days which correlated with direct, active cell counts.QPCR showed H-li was present at levels only slightly lower than those at day 0. There was no difference in survival between thethree different moisture regimes. Direct CTC counts of S. typhimurium LT2 in the soil microcosms confirmed that intact cellswere present in a metabolising state after 54 days in non-sterile soil, indicating a significant proportion of uncultured but activecells. z 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.
Keywords: Quantitative polymerase chain reaction; Active staining; Salmonella typhimurium LT2; Soil ; Non-culturable
1. Introduction
Salmonellae, particularly Salmonella enteritidisand S. typhimurium, are a major cause of food poi-soning and reported disease incidents are dramati-cally increasing [1^3]. Viable salmonellae have beenfound in anaerobically digested sewage sludge ap-plied to farmland (P. Marsh, unpublished data).
0168-6496 / 98 / $19.00 ß 1998 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 1 6 8 - 6 4 9 6 ( 9 8 ) 0 0 0 8 3 - X
FEMSEC 969 14-12-98
* Corresponding author. Tel. : +44 (1203) 523184;Fax: +44 (1203) 523701; E-mail: [email protected]
Abbreviations: QPCR, quantitative PCR; CTC, 5-cyano-2,3-ditolyl tetrazolium chloride; FITC, fluoroscein isothiocyanate;INT, 2-(p-iodophenyl)-3-(p-nitrophenyl)-5-phenyltetrazoliumchloride; VBNC, viable but non-culturable; PVPP, polyvinyl-polypyrrolidone; TNPE, 50 mM Tris, pH 8.0, 100 mM NaCl,1% (w/v) PVPP, 10 mM EDTA; NA, nutrient agar; SDW,sterile distilled water; XLD, xylose lysine deoxycholate agar;MHC, moisture holding capacity
FEMS Microbiology Ecology 27 (1998) 351^363

An early study using viable counting on selectiveagar suggested culturable salmonellae were undetect-able within three weeks of sludge application to land[4]. Following exposure to a change in their environ-ment, Gram-negative bacteria can cease to be cultur-able on selective agar, whilst retaining apparent via-bility according to direct physiological observations.This state has been described as viable but non-cul-turable (VBNC). Certain enteric bacteria can retaintheir pathogenicity whilst VBNC [5,6]. A recentstudy showed that intact S. typhimurium cells residein non-sterile soil for long periods of time in spite ofapparent rapid die-o¡ according to viable counts onselective agar [7]. Total direct counting showed thatcells remained at a constant density, whereas cellswhich had been UV-killed prior to inoculation de-clined at a steady rate, suggesting that viable cellsmight have persisted at densities close to that ofthe initial inoculum. This has serious implicationsfor methods of detection using the viable platecount, as this remains the most economic and simplemethod of determining persistence in the environ-ment.
Certain growth conditions can facilitate survival ofbacteria. Starvation prior to inoculation allowedprolonged survival of Pseudomonas £uorescens insoil [8] whereas starvation lengthened the survivalperiod of Escherichia coli in two di¡erent soils [9].Survival of P. £uorescens was also prolonged in wetsoils, in comparison to dry conditions [10]. Salmo-nellae produce a large array of speci¢c proteins inresponse to starvation stress. This response is gov-erned by starvation inducible genes, and some of theproducts are known to aid survival during long peri-ods of nutrient-limiting conditions and also bestowresistance to other challenges such as oxidative stressfrom hydrogen peroxide [11]. These studies suggestthat many stress responses in salmonellae and otherbacteria are highly tuned adaptive response mecha-nisms to environmental changes.
The present study aimed to investigate further thepossibility that intact, active salmonellae persist insoil despite apparent loss of culturability on selectivemedia. A dual approach was taken. The ¢rst strategyinvolved quantitatively monitoring DNA by ampli¢-cation of a Salmonella-speci¢c sequence, H-li, whichencodes the phase 1 £agellar ¢lament protein [12]from soil extracts. H-li is a single copy chromosomal
gene in Salmonella species [13] and one target se-quence is therefore equivalent to one intact cell.Although not exclusive evidence of intact or viablecells, comparison of DNA quanti¢cation results withtotal direct- and direct-active staining may indicatepotentially viable Salmonella cells. The second strat-egy aimed at assessing metabolic activity of salmo-nellae in soil extracts by a combination of direct vitalstaining using the respiration-sensitive £uorochromeCTC [14] and speci¢c antibody-linked £uorescencemicroscopy [7]. Previous studies with heavy clay soilsusing the direct viable count method with nalidixicacid proved unsuccessful, probably due to adsorp-tion of nalidixic acid to clay [7]. The present studyused the combined CTC-£uorescent antibody techni-que in concert with molecular techniques to monitorsurvival of intact and metabolising S. typhimurium innatural soil over a long time period. Survival in soilat three di¡erent matric potentials (360, 36 and 32kPa) was investigated, based on the hypothesis thatin a relatively dry soil, stress exerted on cells mightcause a greater loss in culturability due to desicca-tion [15,16]. We also investigated the possibility thatcounts on selective agar may under-estimate salmo-nellae following long periods in soil compared tocounts on non-selective complex agar. This wouldindicate whether the selective medium inhibitedgrowth of sublethally injured or starved cells.
2. Materials and methods
2.1. Bacterial strain and soil
S. typhimurium LT2 was prepared for inoculationaccording to Turpin et al. [7]. Soil was taken from alocal ¢eld site (Cry¢eld, University of Warwick), andanalysis of particle size indicated a composition of(percent, dry weight) 63.6% sand, 18.4% silt, 11.7%clay and 6.2% loss on ignition. The pH of the freshsoil was 6.5^6.8 [17]. Microcosms were prepared byadding 40 g soil to 300 ml medical £at bottles.
2.2. Survival experiment
Triplicate sets of non-sterile soil microcosms wereinoculated with S. typhimurium LT2 at 1U107 cfug31 at three di¡erent matric potentials: 360, 36
FEMSEC 969 14-12-98
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363352

and 32 kPa (equivalent to 15%, 22% and 32% (v/w)soil moisture content, respectively, or 40%, 60% and90% moisture holding capacity (MHC), respectively).Microcosms were incubated at 22³C for 54 days andsampled at days 0, 3, 13, 33 and 54.
2.3. Inhibitory e¡ects of selective agar
S. typhimurium LT2 was inoculated at 1U107 cfug31 into microcosms of sterile soil with a matricpotential of 360 kPa. These were incubated at22³C for 189 days and sampled at days 0, 70 and189. Dilutions of each extract were plated on XLD(Oxoid) for the selective medium and NA (Oxoid)for the non-selective medium and incubated at37³C for 24 h.
2.4. E¡ects of desiccation on Salmonella survival insoil
The e¡ects of desiccation stress were assessed byinoculating sterile soil microcosms with sterile dis-tilled water to 360 kPa (15% (v/w) moisture), andS. typhimurium LT2 at 107 cfu g31, followed by in-cubation at room temperature for 189 days withsampling on days 0, 5, 27, 42, 70 and 189. The mi-crocosms were covered in ultra thin cellophane ¢lmto allow evaporation of water without contamina-tion. At about 0.1% (v/w) moisture after 30 days ofincubation, the moisture content of the soil micro-cosms was brought up to 15% (v/w) (360 kPa) bythe addition of an appropriate amount of sterile dis-tilled water. Sub-lethal damage was assessed by theability to grow on spread plates of XLD and NA,and on plates of TSA overlayed with XLD or NAaccording to the double layer agar technique of Mor-inìigo et al. [18]. This allowed sub-lethally injuredcells, unable to grow on spread plates, to resuscitateand form colonies. Direct CTC counts and total di-rect counts were also used as described later to esti-mate cell numbers in the soil extracts.
2.5. Extraction and concentration of cells and DNAfrom soil
An adaptation of the ion exchange-based method(Chelex extraction) of Turpin et al. [19] was used forthe extraction and concentration of Salmonella cells
from soil. Soil (10 g) was mixed with 10 ml of 2.5%polyethylene glycol 6000 and 2 g of Chelex 100 (Bio-Rad). The mixture was gently agitated on a Gri¤n£ask shaker for 2 h at 4³C. Large soil particles weresedimented by centrifugation at 177Ug for 30 s,after which the supernatant was further centrifugedat 3500Ug for 15 min. The resulting pellet was re-suspended in either 1 ml of one quarter strengthRinger's solution for plating, or 1 ml of TNPE(50 mM Tris, pH 8.0, 100 mM NaCl, 1% (w/v) poly-vinylpolypyrrolidone, 10 mM EDTA [20]) for DNAextraction. For plate counts, dilutions of the extractwere plated on appropriate agar(s) and incubated at37³C for 24 h. The pellet, resuspended in TNPE, wasadded to a 7-ml Braun bead-beating tube with 1 g of0.10^0.11-mm glass beads (Braun). TNPE was addedto ¢ll the tube completely (total volume: 7 ml), andcells were ruptured by agitation in a Braun bead-beating machine for 5 min with CO2 cooling [21].Solid matter in the mixture was pelleted by centrifu-gation at 1660Ug for 10 min. The supernatant wassaved and 0.5 volume of ice-cold 8 M potassiumacetate was added, and left to stand in ice for 15min to precipitate salt from the extract. This wascentrifuged at 12 800Ug for 30 min, after whichthe supernatant was added to 2 volumes of ice coldabsolute ethanol and 0.1 volume of 3 M sodiumacetate and incubated at 370³C for 1 h. After fur-ther centrifugation at 12 800Ug for 30 min, the pelletwas washed in ice cold 70% ethanol, centrifugedagain at 12 800Ug for 15 min, and ¢nally resus-pended in 500 Wl of TE. The soil DNA was puri¢edby partition and ion exchange chromatography [22].The ¢nal eluate was stored at 320³C.
2.6. QPCR from soil DNA
QPCR on extracts used the primers H-li 1 and H-li2 (Table 1) and conditions described by Way et al.[12]. The reaction was made quantitative by the in-clusion of 6 Wl of a 10 mg ml31 solution of bovineserum albumin (BSA; Sigma) [23], which served toinhibit humic-based inhibition of the PCR reaction[24]. The quantity of H-li sequences in soil DNAextracts was estimated by densitometry of photo-graphic negatives of ethidium bromide-stained gelsof the products and compared to a calibration curve[23]. The calibration curve was made using extracts
FEMSEC 969 14-12-98
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363 353

from soil inoculated with a known range of S. typhi-murium LT2 densities between 107 and 104 cfu g31
dry soil. When QPCR was performed on soil extractsduring experiments, control samples of extracts usedin construction of the calibration curve were alsosubjected to QPCR. PCR products ampli¢ed fromsoil DNA were con¢rmed as being H-li by semi-nested PCR [22]. A third downstream primer, H-li3 (Table 1), was designed from a 20-bp sequence inthe centre of the 173-bp H-li product. On completionof the ¢rst PCR, a second reaction mixture identicalto the ¢rst minus the BSA and including primers H-li1 and H-li 3 and 1 Wl of the ¢rst reaction mixturewas subjected to a second PCR. A 107-bp productresulting from H-li 1 and H-li 3 was ampli¢ed and allproducts were visualised on 1.6% agarose gel con-taining 0.2 Wg ml31 ethidium bromide.
2.7. Southern hybridisation of PCR products from soil
H-li products were hybridised to Hybond1-N+nylon membrane (Amersham) and probed withpure GT32P-labelled H-li (previously ampli¢ed frompure chromosomal S. typhimurium LT2 DNA) usinga Random Primed DNA Labelling Kit (BoehringerMannheim) according to the manufacturers' instruc-tions.
2.8. Detection limit of QPCR from non-sterile soil
Six non-sterile soil microcosms (10g) were inocu-lated with S. typhimurium LT2 at densities between107 and 103 cfu g31, plus a control inoculated withSDW, and the inoculants were allowed to soak intothe soil at room temperature for 1 h and were thensubjected to DNA extraction and QPCR.
2.9. Direct active staining
Ten g of soil and 10 ml of 1% (w/v) CaCl2 weremixed and then shaken on a Gri¤n £ask shaker atmaximum speed for 10 min. Soil from this suspen-sion was £occulated by standing for 1 h at roomtemperature and 100 Wl of the clear upper layerwas centrifuged at 12 000Ug for 5 min. The pelletwas then washed three times in phosphate bu¡eredsaline (PBS; pH 7.2), and resuspended in 100 Wl of a1.52 mg ml31 solution of CTC (Polysciences) and10 Wl of tryptone soya broth (Oxoid) [14]. Thereaction mixture was incubated in a Gallenkamporbital shaker (200 rpm) at 37³C in the dark for16 h. The cells were again washed three times inPBS and resuspended in SDW. The extended incu-bation overnight was used as slowly metabolisingcells took longer to develop £uorescence.
2.10. Counter-staining for total direct counts
Resuspended CTC-stained Salmonella (10-Wl ali-quots) were speci¢cally counter-stained with FITCusing an immuno£uorescence technique [7]. FITC-antibody labelled and CTC cells were examinedwith a Zeiss Axioskop microscope equipped withan epi£uorescent illumination system. The micro-scope was ¢tted with Zeiss ¢lter sets 09 and 15suitable for FITC and CTC, respectively, and a Ha-mamatsu CCD Camera C4742 for image capture.Image analysis on captured U400 images employedthe Improvision system using the software Bio-vision 2.2a6. Ten ¢elds per replicate sample wereenumerated.
FEMSEC 969 14-12-98
Table 1Primers for PCR ampli¢cation of H-li from Salmonella species
Oligonucleotide sequence Orientation and position onDNA sequencesa
Ampli¢cation region(bp)
Reference
H-li 1 (5P-AGCCTCGGCTACTGGTCTTG-3P) 624C643 173b Way et al. (1993)H-li 2 (5P-CCGCAGCAAGAGTCACCTCA-3P) 7776796H-li 3 (5P-GTTCCCCCCGTAACGGTAAC-3P) 7126731 107c Joys (1985)
aArrows indicate upstream (C) or downstream (6).bProduct size from primers H-li 1 and H-li 2.cProduct size from primers H-li 1 and H-li 3.
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363354
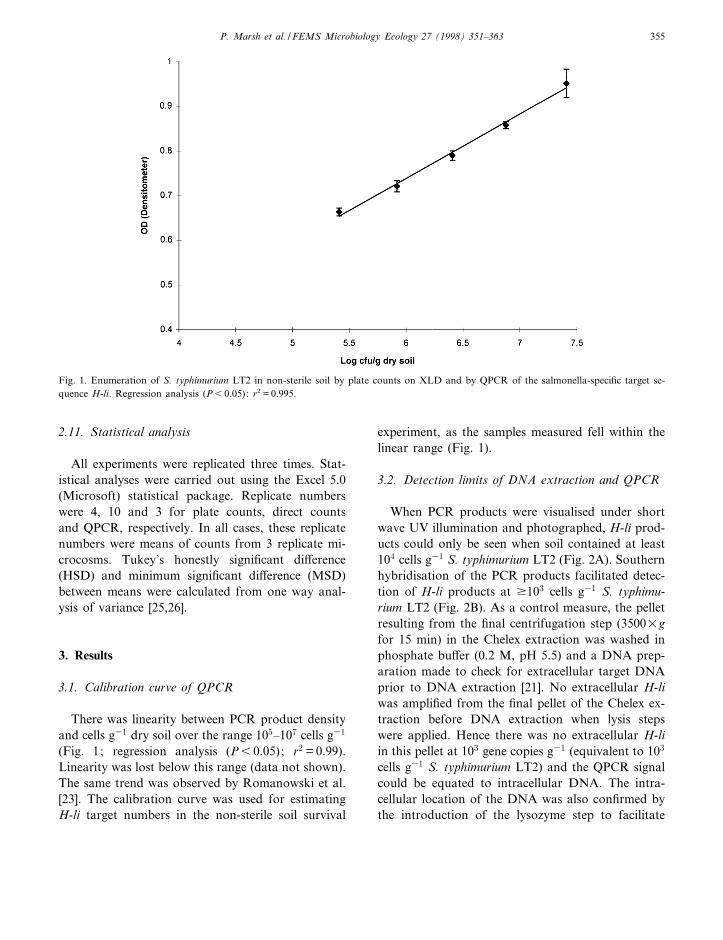
2.11. Statistical analysis
All experiments were replicated three times. Stat-istical analyses were carried out using the Excel 5.0(Microsoft) statistical package. Replicate numberswere 4, 10 and 3 for plate counts, direct countsand QPCR, respectively. In all cases, these replicatenumbers were means of counts from 3 replicate mi-crocosms. Tukey's honestly signi¢cant di¡erence(HSD) and minimum signi¢cant di¡erence (MSD)between means were calculated from one way anal-ysis of variance [25,26].
3. Results
3.1. Calibration curve of QPCR
There was linearity between PCR product densityand cells g31 dry soil over the range 105^107 cells g31
(Fig. 1; regression analysis (P6 0.05); r2 = 0.99).Linearity was lost below this range (data not shown).The same trend was observed by Romanowski et al.[23]. The calibration curve was used for estimatingH-li target numbers in the non-sterile soil survival
experiment, as the samples measured fell within thelinear range (Fig. 1).
3.2. Detection limits of DNA extraction and QPCR
When PCR products were visualised under shortwave UV illumination and photographed, H-li prod-ucts could only be seen when soil contained at least104 cells g31 S. typhimurium LT2 (Fig. 2A). Southernhybridisation of the PCR products facilitated detec-tion of H-li products at v103 cells g31 S. typhimu-rium LT2 (Fig. 2B). As a control measure, the pelletresulting from the ¢nal centrifugation step (3500Ugfor 15 min) in the Chelex extraction was washed inphosphate bu¡er (0.2 M, pH 5.5) and a DNA prep-aration made to check for extracellular target DNAprior to DNA extraction [21]. No extracellular H-liwas ampli¢ed from the ¢nal pellet of the Chelex ex-traction before DNA extraction when lysis stepswere applied. Hence there was no extracellular H-liin this pellet at 103 gene copies g31 (equivalent to 103
cells g31 S. typhimurium LT2) and the QPCR signalcould be equated to intracellular DNA. The intra-cellular location of the DNA was also con¢rmed bythe introduction of the lysozyme step to facilitate
FEMSEC 969 14-12-98
Fig. 1. Enumeration of S. typhimurium LT2 in non-sterile soil by plate counts on XLD and by QPCR of the salmonella-speci¢c target se-quence H-li. Regression analysis (P6 0.05): r2 = 0.995.
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363 355

lysis of the cells surviving in soil (see DNA quanti-¢cation).
3.3. Selective e¡ects of isolation media
Sterile microcosms were incubated longer thannon-sterile microcosms to allow the S. typhimuriumLT2 to decline to a count on XLD approximate tothat in the non-sterile soil survival experiment after54 days. Sterile soil was necessary in this experiment
so that S typhimurium LT2 counts on NA could becompared with counts on XLD without interferencefrom indigenous soil micro£ora which would growon NA. XLD is recommended as a sensitive selectivemedium for isolation of salmonellae from environ-mental samples [18]. S. typhimurium LT2 counts de-clined from 107 to 104^105 cfu g31 over 189 daysaccording to plate counts on XLD and NA (Table2). There was no signi¢cant di¡erence (P6 0.05) be-tween counts on XLD and NA at days 0, 70 or 189(Table 2). This was only con¢rmed in extracts fromsoil at a matric potential of 360 kPa.
3.4. E¡ects of desiccation on Salmonella survival insoil
Following inoculation, there was a 10-fold in-crease in cfu (estimated both by spread plates andoverlay plates), followed by a decline in numbers upto the re-wetting point at day 30 (Fig. 3A). Thispopulation increase and subsequent decrease was re-
FEMSEC 969 14-12-98
Fig. 2. Soil DNA extracted from non-sterile soil inoculated with S. typhimurium LT2 and PCR ampli¢ed with primers H-li 1 and H-li 2(Table 1). A: Soil DNA and PCR ampli¢cation. Lanes 1^6: Soil DNA extracts. Soil inoculated with LT2 at cells g31 dry soil as follows:1: 107 ; 2: 106 ; 3 : 105 ; 4: 104 ; 5: 103 ; 6 : SDW; 7: 1-kb size marker. Lanes 8^15: PCR ampli¢ed H-li products from soil extracts shownin lanes 1^6. Lanes 8^13 correspond to lanes 1^6, respectively. Lane 14: Positive control (LT2 chromosomal DNA as target) ; lane 15:negative control (no DNA); lane 16: 1-kb size marker. B: Southern hybridisation of products ampli¢ed from soil to con¢rm the detectionlimit of 103 cells g31. Lanes 1^8: PCR ampli¢ed H-li from soil extracts as in A hybridised with GT32P-labelled pure H-li. Lanes 1^8 cor-respond to lanes 8^15 in A. Size of H-li is marked in bp.
Table 2Comparison of counts of S. typhimurium LT2 on XLD and NAfollowing extraction from sterile soil
Days in soil Log cfu g31
dry soilHSD(P6 0.05)
XLD NA
0 7.46 7.26 0.0570 6.27 6.59 0.98
189 4.33 5.09 0.94
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363356
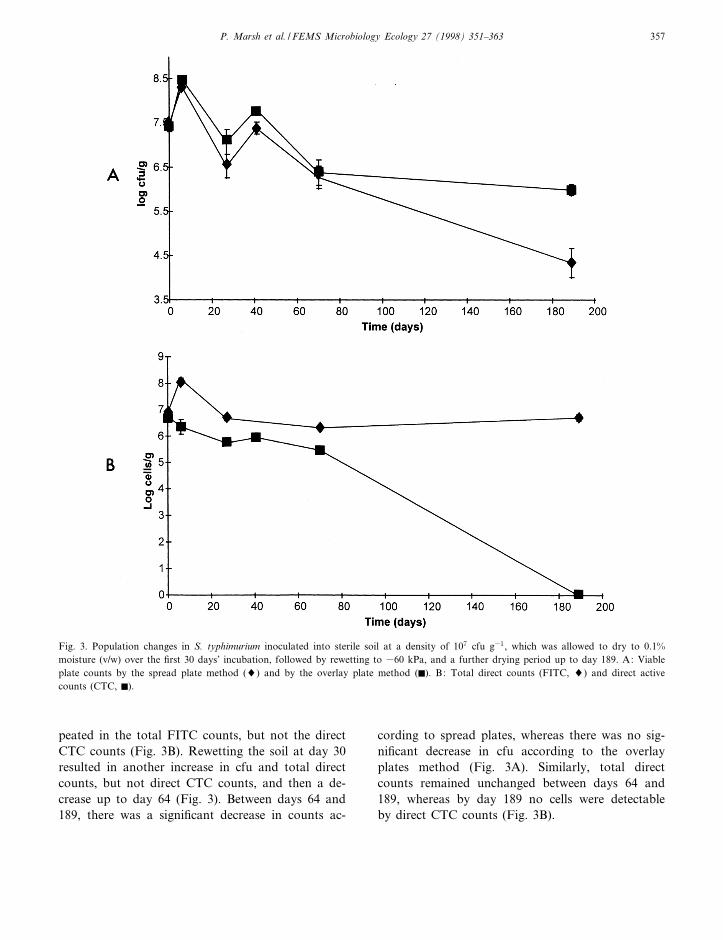
peated in the total FITC counts, but not the directCTC counts (Fig. 3B). Rewetting the soil at day 30resulted in another increase in cfu and total directcounts, but not direct CTC counts, and then a de-crease up to day 64 (Fig. 3). Between days 64 and189, there was a signi¢cant decrease in counts ac-
cording to spread plates, whereas there was no sig-ni¢cant decrease in cfu according to the overlayplates method (Fig. 3A). Similarly, total directcounts remained unchanged between days 64 and189, whereas by day 189 no cells were detectableby direct CTC counts (Fig. 3B).
FEMSEC 969 14-12-98
Fig. 3. Population changes in S. typhimurium inoculated into sterile soil at a density of 107 cfu g31, which was allowed to dry to 0.1%moisture (v/w) over the ¢rst 30 days' incubation, followed by rewetting to 360 kPa, and a further drying period up to day 189. A: Viableplate counts by the spread plate method (8) and by the overlay plate method (F). B: Total direct counts (FITC, 8) and direct activecounts (CTC, F).
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363 357
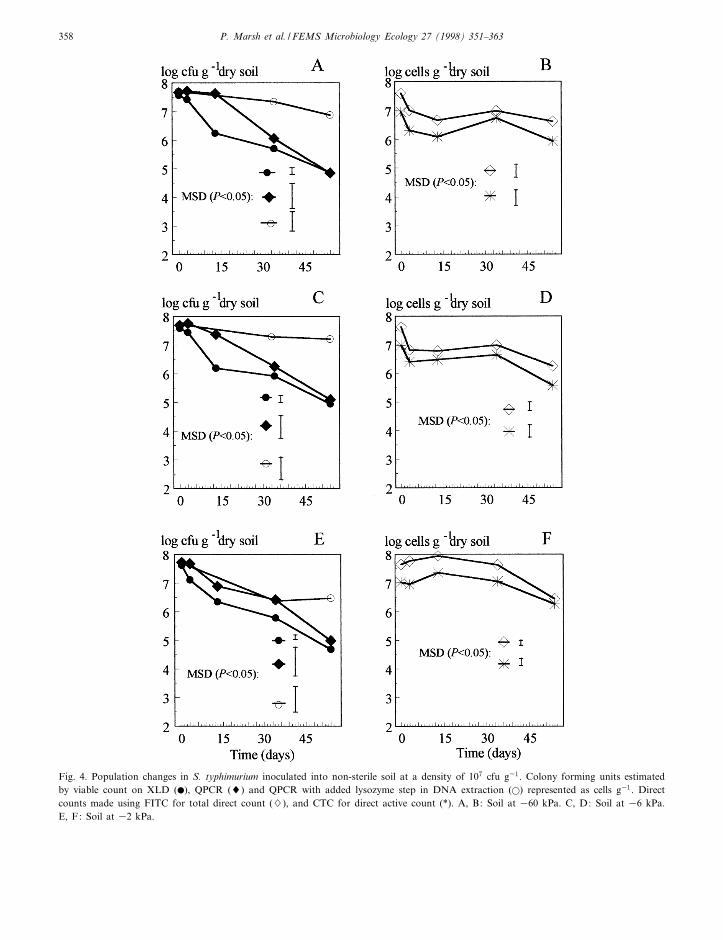
FEMSEC 969 14-12-98
Fig. 4. Population changes in S. typhimurium inoculated into non-sterile soil at a density of 107 cfu g31. Colony forming units estimatedby viable count on XLD (b), QPCR (8) and QPCR with added lysozyme step in DNA extraction (a) represented as cells g31. Directcounts made using FITC for total direct count (7), and CTC for direct active count (*). A, B: Soil at 360 kPa. C, D: Soil at 36 kPa.E, F: Soil at 32 kPa.
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363358

3.5. Detection of metabolising S. typhimurium
In the three di¡erent moisture regimes, the platecounts on XLD declined rapidly between days 0 and13, after which the rate of decline was less rapid (Fig.4A, C and E). In contrast, the total direct counts inthe two drier soils fell slightly between days 0 and 3,and were then relatively unchanged at between about107 and 106 cells g31 throughout the remaining 54-day incubation period (Fig. 4B and D). The totaldirect counts in soil at 32 kPa rose between days 0and 13, then declined gradually for the remainder ofincubation (Fig. 4F). Direct counts of CTC-stainedcells followed the same trend as the total directcounts in all three water regimes, but at about oneorder of magnitude lower than the total direct countsof FITC-stained cells for 54 days (Fig. 4B, D and F).This trend was constant throughout the experiment,and may be explained by a low staining e¤ciency ofthe CTC method. Trends in culturable viable countsdid not di¡er between the three moisture regimes.Cells became visibly smaller with time. RespiringFITC-stained Salmonella could be speci¢cally di¡er-entiated and counted using the Improvision imageanalysis equipment by highlighting and enumeratingcells in a known ¢eld size from an image capturedfrom a view under ¢lter 09 (FITC-stained cells). Thehighlighted areas could then be stored, and appliedto the same ¢eld from a view under ¢lter 15 (CTC-stained cells).
3.6. DNA quanti¢cation
DNA extraction by purely physical means (beadbeating) gave results from QPCR suggesting that theDNA quantity followed similar trends to the platecounts (Fig. 4A, C and E). The QPCR-bead beatingestimations did, however, appear to show steady celldensities up to day 13 in 32 kPa soil (Fig. 4A),whereas the day 13 QPCR-bead beating estimationsfrom soil at 36 and 32 kPa were progressively lower(Fig. 4C and E). After day 13, all QPCR-bead beat-ing estimations followed the trend of the platecounts. It was postulated that this e¡ect might bedue to increased resistance to lysis due to long-term residence of the cells in soil and decrease insize. To overcome this problem, a lysis step was in-troduced to the DNA extraction prior to bead-beat-
ing. This involved an additional lysozyme stepwhereby the pellet, following centrifugation(3500Ug for 15 min) was resuspended in 1 ml of20 mg ml31 lysozyme in 50 mM Tris-HCl (pH 8.0),0.25 M sucrose [27,28] and incubated at 37³C for 30min. The mixture was then transferred to the bead-beating tube and made up with TNPE, and extrac-tion proceeded as described in Section 2. With themodi¢ed method, estimations of H-li were higher inall three moisture regimes and at 360 and 36 kPathere were no signi¢cant declines in cfu g31 (Fig. 4Aand C). The cfu g31 in 32 kPa soil declined by oneorder of magnitude (Fig. 4E). All PCR products am-pli¢ed from soil extracts were con¢rmed as being H-li according to semi-nested PCR using primers H-li 1and H-li 3 (Fig. 5).
4. Discussion
Sewage sludge is applied in large quantities toagricultural land in the UK. This poses a potentialrisk to human health as there is a probability that
FEMSEC 969 14-12-98
Fig. 5. Semi-nested PCR ampli¢cation products using primers H-li 1 and H-li 3 (Table 1) in the presence of PCR-ampli¢ed H-lifrom extracts of the 54-day microcosm experiment. The semi-nested product can be seen at 107 bp, whereas a residual 173-bpproduct from the QPCR (with lysozyme step) ampli¢cation runcan often be seen. Lanes 1^3, 4^6, and 7^9 represent triplicatesamples from days 0, 34 and 54, respectively, of the soil micro-cosms at 360 kPa. Lane 10: Positive control (LT2 chromosomalDNA as target) ; lane 11: negative control (no DNA negativecontrol from QPCR); lane 12: 1-kb size marker. Sizes aremarked in bp.
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363 359

viable human pathogens in the sludge may endure,survive and return to the human food chain viacrops and livestock farmed on sludge-fertilisedland. Enteric bacteria have evolved survival strat-egies to adapt to long periods of time outside thehost-animal-body following egestion of waste, main-taining pathogenicity when ingested by a new host.This work explored the possibility that S. typhimu-rium persists and survives in soil.
Plate counting on XLD did not underestimate S.typhimurium LT2 in comparison to counts using thenon-selective NA. Hence estimations of culturablesalmonellae from soil microcosms using XLD didnot exclude sublethally injured cells. XLD was foundto be e¤cient for enumerating salmonellae in envi-ronmental samples such as seawater microcosms andsoil [7,18,19]. S. typhimurium cells exposed to sub-lethal oxidative stress showed equal e¤ciency ofplating on both XLD and NA (N.Z. Morris and P.Marsh, unpublished data).
The DNA extraction method appeared to be com-parable to other methods in its e¤ciency, i.e. ena-bling detection of PCR targets at v103 cells g31 [29].The detection limit for the viable count by ion ex-change resin-based extraction was 102 cfu g31 (datanot shown), the greater sensitivity possibly arisingfrom the number and nature of clean-up steps re-quired for DNA extraction.
In all of the control microcosms where non-sterilesoil was inoculated with SDW alone, no Salmonella-speci¢c products were ampli¢ed. All the QPCRproducts were con¢rmed by semi-nested PCR prov-ing that H-li ampli¢ed from non-sterile soil inocu-lated with S. typhimurium LT2 was being ampli¢edfrom the DNA of that strain alone.
Desiccation of the soil containing S. typhimuriumcells resulted in apparent loss of activity according todirect CTC counts, whereas viable counts by thedouble layer plate technique suggested that a similarpopulation density of viable cells was present in thesterile soil after 189 days. The double layer methodallowed recovery of sublethally injured cells after 80days. This observation alludes to the survival of thecells in a sessile state for prolonged periods, and anability to resuscitate under the appropriate condi-tions. The decline in CTC-stained cells may be dueto changes in permeability following a signi¢cantreduction in size, and indicated that the CTC-stain-
ing was not always a reliable method for the detec-tion of viable cells.
The use of three di¡erent moisture regimes al-lowed observation of survival under di¡ering matricpotentials. Warwick Cry¢eld soil at 360 kPa waswell drained, contained air-spaces between aggre-gates and was suitable for growth of ¢lamentousbacteria such as streptomycetes which are less de-pendent on water ¢lms in soil [30]. Soil at 36 kPais close to typical ¢eld capacity in the UK, whereaswater in soil at 32 kPa was readily availablealthough the soil was not water-logged. There wasno apparent di¡erence between moisture regimes forthe survival of S. typhimurium LT2 as shown by thesimilar rates of decline in the active counts for thethree treatments in addition to similar trends ob-served for the direct counts (FITC) which remainedbetween 107 and 106 cells g31 throughout the 54days. The CTC counts indicated that a large propor-tion of the cells remained active for 54 days, in con-trast to the declining counts on XLD over this peri-od. Active Salmonella cells in soil samples couldtherefore be detected and enumerated using a combi-nation of CTC and FITC counter-staining. Thistechnique was greatly improved by incorporationof the Improvision image analysis system with Bio-vision 2.26a. The method could be applied to enu-meration of other species in soil where antibody-linked £uorochromes are available, or the enumera-tion of indigenous soil bacteria using CTC and coun-ter-staining with non-speci¢c £uorochromes such asDAPI or acridine orange. Although the cells werenot assessed for their potential to elongate in thepresence of substrate with naladixic acid to preventdivision [31], only metabolising cells stained posi-tively with CTC and hence may be capable of growthor pathogenesis. The Kogure et al. [31] method failedwith this soil due to adsorption of the nalidixic acidto clay [7].
Substrate response/naladixic acid and the CTCmethods for direct active counts of Flavobacteriumstrain P25 in soil showed very similar trends duringa survival study [32]. The extraction method for di-rect staining had several advantages over that of Pyleet al. [33], which involved ¢ltration of samplethrough membrane ¢lters so as to concentrate andimmobilise cells. Dense environmental samples suchas sewage sludge and soil may easily block mem-
FEMSEC 969 14-12-98
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363360

brane ¢lters, and cells which have entered a dwarfstate following long periods in soil may be smallenough (6 0.07 Wm3) to pass through most mem-brane ¢lters [34].
Quantitative PCR has been used in several studiesto estimate copy number and cell number in environ-mental samples [23,35]. The QPCR estimation ofcells showed a clear di¡erence between the DNAextraction methods with and without the lysozymestep, indicating changes in physiological state of thecells in soil. Without the lysozyme step, the cellsbecame increasingly di¤cult to lyse as shown bythe decreasing QPCR signal. This decline followedsimilar rates in all soil moisture regimes. However,once the lysozyme step was introduced, almost con-stant QPCR signals were obtained throughout the 54days indicating e¤cient lysis in the two drier soils.The QPCR signal from 32 kPa soil still declinedwith the lysozyme step by an order of magnitudeduring the experiment. The higher direct counts inthe 32 kPa soil compared to the other moistureregimes may be related to di¡erences in extractione¤ciencies with varying soil moistures. Cells in thedrier soils may have become bound to soil aggregatesand were not separated during the CaCl2 extractionfor direct counts, whereas in the wettest soil theamount of water present may have prevented closebinding of cells to soil particles giving higher directcounts.
The resistance to lysis may indicate that the cellshave entered a dwarf state [34], in which cells becameresistant to lysis following long periods of inactivityin nutrient-limited environments. There was a closecorrelation between the rates of decline detected byQPCR (without the lysozyme step) and viable countson XLD. This correlation may have been due to thesame alterations in cell wall composition, resulting inboth resistance to lysis and reduced culturability onselective agar. Use of the two QPCR extractionmethods may therefore be used in concert to evalu-ate cell state in soil. Weichart and Kjelleberg [36]found that VBNC Vibrio vulni¢cus cells were resist-ant to lysis by sonication as evidenced by the pres-ence of intact cells. In this study we have clearlydemonstrated a population of non-culturable but ac-tive S. typhimurium cells in soil with increased resist-ance to lysis as shown by release of DNA.
The number of active CTC-stained Salmonella
cells in non-sterile soil was approximately one orderof magnitude lower than the initial inocula after 54days incubation, despite a far greater loss of cultur-ability on selective agar. Therefore populations insoil may remain pathogenic, although it has beenreported that virulence potential of S. typhimuriumdecreased following starvation in seawater which ap-peared attributable to hydrophobicity alterations onthe cell surface [37]. It is interesting to note that S.typhimurium LT2 has an altered rpoS allele [38].However, despite poor stress tolerance in vitro itcould survive as well as the wild type in vivo. Ourstudies have shown that S. typhimurium LT2 canalso survive in the soil environment.
The loss of culturability resulting from residence insoils has been reported in a number of bacterialgroups including indigenous bacteria such as P. £uo-rescens [39,40] and Enterobacter cloacae [15]. Furtherwork is needed to establish the pathogenic status ofthe Salmonella cells detected in soil and determine ifthe population consists of non-culturable cells capa-ble of resuscitation under speci¢c growth conditions.
Acknowledgments
This research work was funded by BBSRC Grant88/FQS02672. We also acknowledge the help of theWilliam Edwards Charity for Postgraduate Study(N.Z. Morris, M.Sc.).
References
[1] Baird-Parker, A.C. (1991) Foodborne salmonellosis. In:Foodborne Illness (Waites, W.M. and Arbuthnott, J.P.,Eds.), pp. 53^61, Edward Arnold, London.
[2] Hinton, M., and Bale, J.A. (1994) Is Salmonella enteritidisPT4 a superbug? Food Res. Int. 27, 233^235.
[3] Roberts, J.A. and Sockett, P.N. (1994) The socioeconomicimpact of human Salmonella enteritidis infection. Int. J.Food Microbiol. 21, 117^129.
[4] Watson, D. (1985) Potential risks to human and animal healtharising from land disposal of sewage sludge. J. Appl. Bacter-iol. Symp. Suppl., 95S^103S.
[5] Xu, S.-H., Roberts, N., Singleton, F.L., Attwell, R.W.,Grimes, D.J. and Colwell, R.R. (1982) Survival and viabilityon nonculturable Escherichia coli and Vibrio cholerae in theestuarine and marine environment. Microb. Ecol. 8, 313^323.
FEMSEC 969 14-12-98
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363 361

[6] Byrd, J.J., Xu, S.-H. and Colwell, R.R. (1991) Viable butnonculturable bacteria in drinking water. Appl. Environ. Mi-crobiol. 57, 875^878.
[7] Turpin, P.E., Maycroft, K.A., Rowlands, C.L. and Welling-ton, E.M.H. (1993) Viable but non-culturable salmonellas insoil. J. Appl. Bacteriol. 74, 421^427.
[8] van Elsas, J.D., Wolters, A.C., Clegg, C.D., Lappinscott,H.M. and Anderson, N. (1994) Fitness of genetically modi¢edPseudomonas £uorescens in competition for soil and root col-onization. FEMS Microbiol. Ecol. 13, 259^272.
[9] Sjogren, R.E. (1994) Prolonged survival of an environmentalEscherichia coli in laboratory soil microcosms. Water Air SoilPollut. 75, 389^403.
[10] Meikle, A., Aminhanjani, S., Glover, L.A., Killham, K. andProsser, J.I. (1995) Matric potential and the survival and ac-tivity of a Pseudomonas £uorescens inoculum in soil. Soil Biol.Biochem. 27, 881^892.
[11] Foster, J.W. and Spector, M.P. (1995) How Salmonella sur-vive against the odds. Annu. Rev. Microbiol. 49, 145^174.
[12] Way, J.S., Josephson, K.L., Pillai, S.D., Abbaszadegan, M.,Gerba, C.P. and Pepper, I.L. (1993) Speci¢c detection of Sal-monella spp. by multiplex polymerase chain reaction. Appl.Environ. Microbiol. 59, 1473^1479.
[13] Joys, T.M. (1985) The covalent structures of the phase 1 £ag-ellar ¢lament protein of Salmonella typhimurium and its com-parison with other £agellins. J. Biol. Chem. 260, 15758^15761.
[14] Rodriguez, G.G., Phipps, D., Ishiguro, K. and Ridgway, H.F.(1992) Use of a £uorescent redox probe for direct visualizationof actively respiring bacteria. Appl. Environ. Microbiol. 58,1801^1808.
[15] Pederson, J.C. and Jacobsen, C.S. (1993) Fate of Enterobactercloacae JP120 and Alcaligenes eutrophus AE0106(PR0101) insoil during water-stress-e¡ects on culturability and viability.Appl. Environ. Microbiol. 59, 1560^1564.
[16] Teixeira, P., Castro, H. and Kirby, R. (1995) Spray drying asa method for preparing concentrated cultures of Lactobacillusbulgaricus. J. Appl. Bacteriol. 78, 456^462.
[17] Wellington, E.M.H., Cresswell, N. and Saunders, V.A. (1990)Growth and survival of streptomycete inoculants and extentof plasmid transfer in sterile and nonsterile soil. Appl. Envi-ron. Microbiol. 56, 1413^1419.
[18] Morinìigo, M.A., Martinez-Manzarares, E., Munìoz, A., Cor-nax, R., Romero, P. and Borrego, J.J. (1989) Evaluation ofdi¡erent plating media used in the isolation of salmonellasfrom environmental samples. J. Appl. Bacteriol. 66, 353^360.
[19] Turpin, P.E., Maycroft, K.A., Rowlands, C.L. and Welling-ton, E.M.H. (1993) An ion-exchange based extraction methodfor the detection of salmonellas in soil. J. Appl. Bacteriol. 74,181^190.
[20] Curtis, T.P., Mara, D.D. and Silva, S.A. (1992) In£uence ofpH, oxygen, and humic substances on ability of sunlight todamage faecal coliforms in waste stabilization pond water.Appl. Environ. Microbiol. 58, 1335^1343.
[21] Cresswell, N., Saunders, V.A. and Wellington, E.M.H. (1991)Detection and quanti¢cation of Streptomyces violaceolatus
plasmid DNA from soil. Lett. Appl. Microbiol. 13, 193^197.
[22] Straub, S.T., Pepper, I.L., Abbaszadegan, M. and Gerba, C.P.(1994) A method to detect enteroviruses in sewage sludge-amended soil using the PCR. Appl. Environ. Microbiol. 60,1014^1017.
[23] Romanowski, G., Lorenz, M.G. and Wackernagel, W. (1993)Use of polymerase chain reaction and electroporation of Es-cherichia coli to monitor the persistence of extracellular plas-mid DNA introduced into natural soils. Appl. Environ. Mi-crobiol. 59, 3438^3446.
[24] Kreader, C.A. (1996) Relief of ampli¢cation inhibition inPCR with bovine serum-albumin or T4 gene-32 protein.Appl. Environ. Microbiol. 62, 1102^1106.
[25] Peterson, R.G. (1985) Design and analysis of experiments. In:Statistics : Textbooks and Monographs, Vol. 66, pp. 72^111.Marcel Decker, New York.
[26] Fry, J.C. (1989) Analysis of variance and regression in aquaticbacteriology. Binary 1, 83^88.
[27] Johnson, J.L. (1991) Isolation and puri¢cation of nucleicacids. In: Nucleic Acid Techniques in Bacterial Systematics(Stackebrandt,E. and Goodfellow, M., Eds.), pp. 1^19. Wiley,Chichester.
[28] Moreè, M.I., Herrick, J.B., Silva, M.C., Ghiorse, W.C. andMadsen, E.L. (1994) Quantitative cell lysis of indigenous mi-croorganisms and rapid extraction of microbial DNA fromsediment. Appl. Environ. Microbiol. 60, 1572^1580.
[29] Smalla, K., Cresswell, N., Mendonca-Hagler, L.C., Wolters,A. and van Elsas, J.D. (1993) Rapid DNA extraction protocolfrom soil for polymerase chain reaction-mediated ampli¢ca-tion. J. Appl. Bacteriol. 74, 78^85.
[30] Herron, P.R. and Wellington, E.M.H. (1990) New method forextraction of streptomycete spores from soil and applicationto the study of lysogeny in sterile amended and nonsterile soil.Appl. Environ. Microbiol. 56, 1406^1412.
[31] Kogure, K., Simidu, U. and Taga, N. (1979) A tentative directmicroscopic method for counting living marine bacteria. Can.J. Microbiol. 25, 415^420.
[32] Heijnen, C.E., Page, S. and van Elsas, J.D. (1995) Metabolicactivity of Flavobacterium strain P25 during starvation andafter introduction into bulk soil and the rhizosphere of wheat.FEMS Microbiol. Ecol. 18, 129^138.
[33] Pyle, B.H., Broadway, S.C. and McFeters, G.A. (1995) Arapid, direct detection method for enumerating respiring en-terohaemorragic Escherichia coli 0157:H7 in water. Appl. En-viron. Microbiol. 61, 2641^2619.
[34] Bakken, L.R. and Olsen, R.A. (1989) DNA-content of soilbacteria of di¡erent cell size. Soil Biol. Biochem. 21, 789^793.
[35] Rosado, A.S., Seldin, L., Wolters, A.C. and van Elsas, J.D.(1996) Quantitative 16S rDNA-targeted polymerase chain re-action and oligonucleotide hybridization for the detection ofPaenibacillus azoto¢xans in soil and the wheat rhizosphere.FEMS Microbiol. Ecol. 19, 153^164.
[36] Weichart, D. and Kjelleberg, S. (1996) Stress resistance andrecovery potential of culturable and viable but nonculturablecells of Vibrio vulni¢cus. Microbiology 142, 845^853.
[37] Galdiereo, E., Donnarumma, G., de Martino, L., Marcatili,
FEMSEC 969 14-12-98
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363362

A., de l'Ero, G.C. and Merone, A. (1994) E¡ect of low-nu-trient seawater on morphology, chemical composition, andvirulence of S. typhimurium. Arch. Microbiol. 162, 41^47.
[38] Wilmes-Riensenberg, M.R., Foster, J.W. and Curtiss, R.(1997) An altered rpoS allele contributes to the avirulence ofSalmonella typhimurium LT2. Infect. Immun. 65, 203^210.
[39] Winding, A., Binnerup, S.J. and SÖrensen, J. (1994) Viability
of indigenous soil bacteria assayed by respiratory activity andgrowth. Appl. Environ. Microbiol. 60, 2869^2875.
[40] van Overbeek, L.S., Eberl, L., Givskov, M., Molin, S. and vanElsas, J.D. (1995) Survival of, and induced stress resistance in,carbon-starved Pseudomonas £uorescens cells residing in soil.Appl. Environ. Microbiol. 61, 4202^4208.
FEMSEC 969 14-12-98
P. Marsh et al. / FEMS Microbiology Ecology 27 (1998) 351^363 363





























