Embed Size (px)
Citation preview

Efecto de la vegetación sobre la abundancia y diversidad
de depredadores de semillas de Pinus teocote Schl. et
Cham. en hábitats contrastantes del estado de Veracruz,
México
T E S I S
QUE PARA OPTAR POR EL GRADO DE
MAESTRO EN
ECOLOGÍA FORESTAL
PRESENTA
Rafael Flores Peredo
DIRIGIDA POR
Dr. Jorge Galindo González
Xalapa, Veracruz, México Marzo de 2005

DEDICATORIA
Dedico este trabajo a mis padres Ana María del Socorro Peredo de Flores (†) y Octavio
Flores Fuentes (†), a los cuáles llamó Dios para formar parte de algo que en la actualidad me
gusta observar……el cielo, a esas dos nuevas estrellas que amo y que están conmigo en mis
triunfos y derrotas, secando mis lágrimas y gozando de mis sonrisas, a ustedes que me enseñaron
a luchar por lo que se quiere les dedico esta investigación.
Dedico el mismo a mis hermanos Ana Lourdes, Octavio y Abraham, por haber visto en
ustedes la superación de salir adelante, aún sin tener el amor de nuestros padres cuando más lo
necesitamos, y viéndolos ahora como todos unos profesionistas, estoy orgulloso de ustedes.
Dedico el mismo a María de Jesús Flores Fuentes (mi Tía Chuy), por haber llorado el
esfuerzo de querer y preocuparse por nosotros, brindarnos el cariño y el consejo de una madre
que siempre esta pendiente de lo que le pase a sus hijos, por el tiempo dedicado al vernos crecer,
por lo que eres, te dedico este pequeño escalón más de mi vida.
Dedico el mismo a mis primos Efrén López Flores y Pedro López Flores, por convivir en
parte de lo que es nuestra vida, de constituir lo que llamo yo una familia, por sus consejos, sus
risas, su tiempo, gracias.
Dedico el mismo a Joaquina Flores Fuentes (mi Tía Pina), por tu amor y cariño demostrado
para nosotros desde chicos y más cuando murió mi papa, gracias tía.
Dedico el mismo a mi novia Fabiola Carolina Espinosa Gómez por ser una linda mujer que
siempre esta en mi corazón, por tus sonrisas, tu compañía, tu cariño tan tierno, por quererme y
dejarme quererte, por ser ese lindo sueño al que recurro en mis noches, por ser como eres, te amo
Fabi.
Dedico el mismo a mis amigos, por estar conmigo cuando más los necesité por sus risas,
sus lágrimas, sus consejos y oraciones, su tiempo, su compañía, por ser parte de mi familia, a :
Alejandro Aramburu, Itzia, Selene, Claudia Ochoa, Pilar, Thania, Alonso, Emilio, Esther, René,
Enrique, Liliana, Jair, Carlos, Patricia, Humberto, Lizbeth, Vanessa, Ángel Antonio, Migue,
Magdiel, Mario, Héctor, Margarita, Eda, Helxine, Carolina, Gabriela, Guadalupe, Juan Carlos,
Amabel, Ariana, Berenice, Diana, Gloria, Yuriana, Chely, Janeth, Yadira, Juan José, Myriam,
Erika y todos aquellos que me brindaron su amistad Gracias.

AGRADECIMIENTOS
Agradezco a Dios el darme fuerzas para alcanzar mis metas, y permitirme ver en la
naturaleza lo hermoso que es este mundo.
Agradezco al M. en C. Juan Alba Landa (Director del Instituto de Genética Forestal) el
permitirme cursar la Maestría en Ecología Forestal que él coordina, misma que me brindo la
oportunidad de subir un escalón más a nivel profesional. Gracias Maestro Alba.
Agradezco al Consejo Nacional de Ciencia y Tecnología “CONACYT” el haber aceptado
la postulación de mi persona como Becario para estudios de Maestría, otorgándome el registro
175058, mismo que fomentó en mí el esfuerzo por culminar este postgrado mediante la
presentación de mi trabajo de investigación.
Agradezco al Biól. Wilfrido Márquez Ramírez (Subdirector de Planeación y Recursos
Naturales) de la Coordinación Estatal de Medio Ambiente, el apoyarme para que llevara con
éxito la culminación de este Postgrado, por su apoyo al permitirme ausentar de mis horas
laborales los viernes. Gracias.
Agradezco al Biól. Celso Hernández Aponte (Coordinador Estatal de Medio Ambiente) de
la Coordinación Estatal de Medio Ambiente, el apoyarme para que llevara con éxito la
culminación de este postgrado al ausentarme en horas laborales.
Agradezco al Dr. Jorge Galindo González, Investigador del Laboratorio de Biotecnología y
Ecología Aplicada de la Universidad Veracruzana, por haber sido mi Director y compartir
conmigo sus conocimientos, consejos y comentarios que culminaron con la presentación de este
trabajo. Gracias Doc.
Agradezco al M. en C. Álvar González Christen, Investigador del Instituto de
Investigaciones Biológicas, por permitirme cotejar mis registros con la información y
conocimiento con el que cuenta de los roedores del Estado de Veracruz.
Agradezco a la Dra. Rosa Amelia Pedraza, Investigadora del Instituto de Genética Forestal
de la Universidad Veracruzana haber compartido sus conocimientos y sugerencias, al revisar mi
trabajo.
Agradezco a cada uno de los Catedráticos en este Postgrado Juan Alba, Juan Márquez,
Rosa Amelia Pedraza, Álvar González, Lilia Mendizábal, Virginia Rebolledo, Jorge Galindo,
Lázaro Sánchez, por haber compartido con nosotros parte de sus conocimientos. Gracias.
Agradezco a mis compañeros de Generación en esta Maestría el haberme dado la
oportunidad de convivir con ellos, y hacer más amenas las clases y las salidas de campo, gracias
amigos.

Agradezco a Margarito Becerra Luna, Benjamín Becerra López (Vigilantes del Centro de
Educación y Convivencia Ambiental de San Juan del Monte) y al Biol. Helxine Fuentes Moreno
(Egresado de la Facultad de Biología de Córdoba) el haberme apoyado en mis trabajos en campo,
así como haber hecho mis estancias de investigación más agradables, gracias amigos.
Agradezco a mis compañeros y compañeras de la Dirección de Planeación y Recursos
Naturales de la Coordinación Estatal de Medio Ambiente, Lic. María de los Ángeles Carreño,
CP. María Esther Beristain, Ing. Rafael Sánchez (†), Biól. Roberto Marín Gómez, Ing. Leo
Miguel Hernández Sánchez, Biól. Ernesto Romero, Ing. Francisco Guzmán, Biól. Flora Zitácuaro
Contreras, Biól. Carlos Martínez, Biól. Miguel Ángel Portilla Morales, Biól. Ana María Gómez
Viera, el ser buenos amigos y conformar un buen equipo de trabajo.
Agradezco a todas aquellas personas que ven en la naturaleza la expresión de algo
hermoso como lo es la vida; aquellos que se preocupan por conservar lo poco que nos queda y
con sus ganas tratan de estudiarlo para generar conocimiento, mismo que permita a la sociedad
ver su importancia, por compartir las mismas ideas que yo, muchas gracias.

i CONTENIDO
INDICE DE FIGURAS Y TABLAS. iv
RESUMEN. vii
SUMMARY.
vii
i
1. INTRODUCCIÓN. 1
2. OBJETIVOS. 3
3. HIPÓTESIS. 3
4. REVISIÓN DE LITERATURA. 4
4.1. Efecto de la vegetación sobre poblaciones de aves
y roedores en algunos ecosistemas: Una revisión.
4
4.1.1. Importancia de las interacciones ecológicas
planta-animal.
4
4.1.2. Efecto de la vegetación sobre la abundancia y
riqueza de aves en un ecosistema.
7
4.1.3. Efecto de la vegetación sobre la abundancia y
riqueza de roedores en un ecosistema.
8
5. MATERIAL Y MÉTODO. 12
5.1. Área de estudio. 12
5.1.1. Localización del área de estudio. 12
5.1.2. Características físicas. 13
5.1.2.1. Clima. 13
5.1.2.2. Fisiografía. 13
5.1.2.3. Hidrología. 13
5.1.2.4. Geología y recursos minerales. 14

ii
5.1.2.5. Suelos. 14
5.1.2.6. Aptitud del suelo. 15
5.1.2.7. Características bióticas. 15
5.1.2.7.1. Comunidades vegetales. 15
5.1.2.7.2. Comunidades animales. 16
5.2. Métodos. 16
5.2.1. Obtención de semillas para el experimento. 16
5.2.2. Selección e identificación de los sitios de
muestreo.
17
5.2.3. Fase experimental. 18
5.3. Análisis Estadístico. 19
6. RESULTADOS. 21
6.1. Aves presentes en la zona de estudio. 21
6.2. Ratones y pequeños mamíferos presentes en la
zona de estudio.
21
6.3. Aves depredadoras de semillas de Pinus teocote. 21
6.4. Ratones depredadores de semillas de Pinus
teocote.
21
6.5. Diversidad y abundancia de depredadores de
semillas de Pinus teocote.
24
6.5.1. Diversidad y abundancia de aves en dos tipos de
hábitat (bosque y pastizal).
24
6.5.2. Diversidad y abundancia de pequeños mamíferos
en dos tipos de hábitat (bosque y pastizal).
24
6.6. Depredación de semillas. 24

iii
6.6.1. Depredación de semillas por aves. 24
6.6.2. Depredación de semillas por roedores. 24
6.7. Consumo total de semillas por aves y roedores en
los dos tipos de hábitat estudiados.
24
6.8. Esfuerzo de muestreo. 25
6.8.1. Curvas de acumulación de especies de aves. 25
6.8.2. Curvas de acumulación de especies de pequeños
mamíferos.
25
7. DISCUSIÓN. 27
8. CONCLUSIONES. 30
9. LITERATURA CITADA. 31
10. ANEXOS. 41
1. Artículo publicado como requisito parcial. 43
2. Listado de especies vegetales del Área Natural
Protegida de San Juan del Monte con status de riesgo
NOM-059-SEMARNAT-2001.
50
3. Listado de especies animales del Área Natural
Protegida de San Juan del Monte con status de riesgo
NOM-059-SEMARNAT-2001.
50

iv ÍNDICE DE FIGURAS Y TABLAS
Figura 1. Se muestra la probabilidad de supervivencia de las semillas bajo la
copa del árbol que las produjo. P, probabilidad de sobrevivencia de las
semillas en relación con la distancia del árbol padre; I, número de semillas
con respecto a la distancia del árbol padre; PRC, curva que marca la
probabilidad de establecimiento de un nuevo individuo (Janzen, 1970).
9
Figura 2. Ubicación geográfica del sitio de estudio, el Área Natural
Protegida de San Juan del Monte, las Vigas de Ramírez, Veracruz. 12
Figura 3. Localización de los sitios de muestreo en el Área Natural Protegida
de San Juan del Monte, las Vigas de Ramírez, Veracruz. Los puntos blancos
indican los sitios que se estudiaron, dos correspondientes a pastizal y dos a
bosque, la simbología indica la vegetación actual existente en el área.
17
Figura 4. Curvas de acumulación de especies de aves en total y por hábitat
registradas en (pastizal y bosque) en el Área Natural Protegida de San Juan
del Monte, las Vigas de Ramírez, Veracruz. Se presentan en el eje X los días
de muestreo, en el eje Y el número de especies acumuladas totales y para
cada tipo de hábitat.
25
Figura 5. Curva de acumulación de especies para los pequeños mamíferos
capturados en dos tipos de hábitat (pastizal y bosque) en el Área Natural
Protegida de San Juan del Monte, las Vigas de Ramírez, Veracruz. Se
presentan en el eje X los días de muestreo, en el eje Y el número de especies
acumuladas totales y para cada tipo de hábitat.
26
Tabla 1. Composición Geológica del Área Natural Protegida de San Juan del
Monte, las Vigas de Ramírez, Veracruz (Geissert et al., 1994). 14
Tabla 2. Tipos de suelo presentes en el Área Natural Protegida de San Juan
del Monte, las Vigas de Ramírez, Veracruz (Geissert et al., 1994). 15
Tabla 3. Lista de especies de aves identificadas en los dos tipos de hábitat
correspondientes al Área Natural protegida de San Juan del Monte, las Vigas
de Ramírez, Veracruz.
22

v
Tabla 4. Lista de pequeños mamíferos identificados en los dos tipos de
hábitat estudiados en el Área Natural Protegida de San Juan del Monte, las
Vigas de Ramírez, Veracruz.
23

vi
RESUMEN
La depredación de semillas llevada a cabo por animales es de gran importancia en varios
procesos de los ecosistemas tales como: la sucesión, regeneración, colonización, y
establecimiento de nuevos individuos y especies. Los roedores y las aves desempeñan un papel
preponderante en la depredación de semillas en bosques templados, ya que pueden determinar
cierto tipo de composición vegetal, sin embargo la fisonomía de la vegetación puede influir en la
abundancia y diversidad de especies de aves y roedores. El objetivo de este trabajo fue evaluar el
efecto del tipo de vegetación sobre la abundancia y diversidad de depredadores de semillas de
Pinus teocote en dos hábitats de la Reserva Ecológica San Juan del Monte, Mpio. de las Vigas de
Ramírez, Veracruz, México. Se localizaron 4 sitios de ca. 1 ha, que representan dos tipos de
hábitat (bosque y pastizal). En cada sitio se colocaron 5 cajas de petri plásticas (cada 200 m) con
10 semillas de P. teocote (sin ala de dispersión) en cada caja. Mediante unos binoculares se
identificaron, registraron y contaron las especies de aves que visitaron los sitios y las que
depredaron semillas. Para los roedores se colocaron 5 trampas sherman en cada sitio, éstos fueron
identificados, se conservó piel y cráneo de cada especie (macho y hembra). Los sitios se visitaron
2 veces por mes durante un año (enero-diciembre de 2003). Se determinó la abundancia y
composición de especies de las comunidades de aves y roedores, la riqueza se comparó mediante
curvas de acumulación de especies. La diversidad se estimó mediante el índice de Shanon-
Wiener, los índices de diversidad se compararon usando una prueba de t; la diferencia entre la
cantidad de semillas consumidas por aves y roedores, en bosque y pastizal, se determinó
mediante una prueba de X². Se contaron un total de 2,099 aves de 43 especies; en el bosque la
diversidad de aves (H´=1.33; 35 especies) fue mayor que en el pastizal (t = 8.41; P<0.001), 11 de
las cuales fueron exclusivas al bosque. En el pastizal, se registraron 32 especies (H´= 1.20), 7 de
las cuales fueron exclusivas al pastizal; entre los tipos de hábitats se compartieron 24 especies de
las cuales 17 fueron de hábitos granívoros, 12 depredaron semillas de P. teocote y 9 de estas
fueron las más abundantes en la zona. Se determinaron 25 nuevos registros. La abundancia de
aves no fue diferente entre los tipos de hábitat (X²=26.01, P>0.05). La depredación de semillas
por aves, fue mayor en el pastizal (X² = 23.35, P< 0.05). El total de especies de aves acumuladas
se estabilizó a los 45 días, de un total de 57 días muestreados; las 32 especies del pastizal se
obtuvieron a los 41 días, mientras que las 35 especies del bosque se obtuvieron a los 45 días. Se
capturaron un total de 80 pequeños mamíferos de 6 especies; en el bosque la diversidad fue
mayor (H´= 0.65) que en el pastizal (t = 3.84; P<0.001), se registraron 6 especies (2 musarañas
exclusivas (un nuevo registro para la zona), y 4 roedores). En el pastizal se registraron las mismas
4 especies de roedores que en el bosque, no hubo especies exclusivas (H´= 0.40), las 4 especies
de roedores depredaron semillas de P. teocote, dos de estas son las mas abundantes en la zona. La
abundancia de roedores fue diferente entre los hábitats (X² = 6.40; P> 0.05), la depredación de
semillas por roedores fue mayor en el bosque que en el pastizal (X² = 67.86, P<0.05). El total de
especies de pequeños mamíferos acumulados se estabilizó a los 53 días de un total de 57 días
muestreados, las 4 especies de pastizal se obtuvieron a los 30 días, mientras que las 6 especies de
bosque se obtuvieron a los 52 días. De los resultados se derivan las siguientes conclusiones: La
vegetación influyó sobre la diversidad de depredadores de semillas de P. teocote, el bosque fue
más diverso, la abundancia de individuos no varió entre los hábitats, en el pastizal se presentó
mayor depredación de semillas de Pinus teocote.

vii
SUMMARY
Seed predations by animals are of great importance in ecosystems processes like: the
succession, regeneration, colonization and establishment of new individuals and species. Rodents
and birds have a basic roll on seed predation in temperate forests, they can establish certain type
of vegetation composition, but also, vegetation physiognomy may influence abundance and
diversity of birds and rodents species. The objective of this work was to evaluate the effect of
vegetation on abundance and diversity of Pinus teocote seed predators at San Juan del Monte
Ecological Reserve, Mpio. of Las Vigas de Ramírez, Veracruz, Mexico. Four sites of ca. 1 ha
were located with two types of habitat (forest and grassland). Five plastic petri boxes, with 10
seeds of P. teocote (without dispersion wing) in each box, were placed in each site (separated 200
m). Using binoculars, bird species that visited study sites were counted, registered, identified, and
also annotated those that feed on seeds. Rodents were collected with 5 sherman traps placed in
each site, organisms were identified, skin and skull of each species (male and female) were
conserved. Sites were visited twice a month during a year (January-December. 2003). Birds and
rodents abundance and community composition were determined; richness was compared by
species accumulation curves; species diversity was calculated with Shanon-Wiener index,
diversity indices were compared using a t test; differences between the amount of seeds
consumed by birds and rodents, in forest and grassland, was determined by X² test. A total of
2,099 birds of 43 species were counted; in forest, bird diversity (H´=1.33) was greater than in
grassland (t = 8.41; P<0.001), registering 35 species (11 exclusive to forest); in grassland, 32
species were registered (7 exclusive) (H´= 1.20); 24 species are shared between habitats, 17 are
granivorous, 12 of them predated P. teocote seeds, and 9 are the most abundant in the zone. I
found 25 new registries. Bird abundance was not different between habitats (X²=26.01; P>0.05),
but seed predation was greater at grassland (X² = 23,35; P< 0,05). Total bird species
accumulation curve, become stabilized at 45 days, from a total of 57 sampling days; in grassland
a total of 32 species were obtained at 41 sampling days, while in forest 35 species were obtained
at 45 days. A total of 80 small mammals of 6 species were captured; diversity of small mammals
in forest (H´= 0,65) was greater than grassland (t = 3.84; P<0.001) accounting 6 species (2
shrews exclusive to forest); in grassland and forest the same 4 species of rodents were registered,
there were not exclusive species to grassland (H´ = 0,40). Seeds of P. teocote were predated by
all species of rodents, and two of these were abundant. I determine 1 new registry of shrew.
Rodent abundance was not different between habitats (X²= 6.40; P> 0.05). Seed predation by
rodents was greater in forest than in grassland (X² = 67,86, P<0.05). Total small mammal species
accumulation curve become stabilized at 53 days from a total of 57 sampling days, in grassland 4
species were obtained at 30 sampling days, while in forest 6 species were obtained at 52 days.
From results the following conclusions are derived: Vegetation influenced diversity of P. teocote
seed predators, forest was most diverse; abundance of organisms did not vary between habitats;
seed predation of was greater in grassland.


1 1. INTRODUCCIÓN
La falta de conocimientos técnicos y el deficiente manejo de los bosques es un grave
problema en México (Geréz, 1983); el aprovechamiento y la conservación forestal deben ir
íntimamente relacionados al conocimiento de los procesos ecológicos de los ecosistemas. La
diversidad biológica del país se debe estudiar, conocer y manejar de manera eficiente para
garantizar su permanencia.
La fuerte presión que se ejerce sobre los bosques templados en el territorio nacional ha
transformado la cubierta vegetal en un paisaje fragmentado, lo que ha provocado una importante
pérdida de hábitat para muchas especies, así como extinciones locales (Turner, 1996; Galindo-
González, en edición). No obstante, el conocimiento que se tiene tanto de las especies, como de
los ecosistemas en su conjunto es aún escaso. Las interacciones planta-animal son de gran
importancia en los procesos ecológicos de los ecosistemas, en particular, la depredación de
semillas puede estar relacionada con la abundancia y diversidad biológica del sitio (Ostfeld et al.,
1997; Mittelbach y Gross, 1984), al igual que los mecanismos de dispersión de semillas, entre
otros, lo que afecta la permanencia de las comunidades arbóreas, incluso en hábitats perturbados
(Connell y Slatyer, 1977; Galindo-González, 1998; Galindo-González et al., 2000).
En la actualidad la mayoría de los métodos propuestos para medir la biodiversidad de
especies están enfocados a la medición del número de especies dentro de las comunidades
conocida como diversidad (alfa), algunos de estos métodos son los siguientes: 1)Basados en la
cuantificación del número de especies presentes (riqueza específica); 2) Basados en la estructura
de la comunidad, es decir, la distribución proporcional del valor de importancia de cada especie
(abundancia relativa de los individuos), estos métodos basados en la estructura pueden a su vez
clasificarse según la dominancia o equidad de cada especie en la comunidad (Moreno, 2000).
Uno de los índices más reconocidos de diversidad que se basa principalmente en el concepto de
equidad, es el de Shannon-Wiener. Este índice expresa la uniformidad de los valores de
importancia a través de todas las especies muestreadas. Con respecto a los índices más usados y
que se basan principalmente en el concepto de dominancia, sobresale el índice de Simpson, el
cuál es un parámetro inverso al concepto de uniformidad o equidad de la comunidad ya que toma
en cuenta la representatividad de las especies con mayor importancia sin evaluar la contribución
del resto de las especies (Moreno, 2000).
La depredación y remoción de semillas llevada a cabo por animales en los ecosistemas es
de gran importancia en algunos procesos como: la colonización, sucesión, regeneración, y
establecimiento de nuevos individuos y especies (Mittelbach y Gross, 1984). Entre los
mamíferos, los roedores tienen un papel primordial en la depredación de semillas en bosques
templados (Gill y Marks, 1991; Ostfeld y Canham, 1993; Ostfeld et al., 1997). La depredación de
semillas llevada a cabo por roedores puede ser intensa y en relación con los patrones de
alimentación de otros herbívoros, puede influenciar el grado, la composición y la invasión de
árboles dentro de los ecosistemas. Los patrones de depredación de semillas llevados a cabo por
roedores, se pueden predecir basando esta predicción en las preferencias de granivoría exhibidas
por roedores en microhábitats particulares (Manson y Stiles, 1998).

2
La depredación de semillas llevada a cabo por aves también resulta de importancia en un
ecosistema ya que puede influir en los procesos de sucesión o colonización de plantas. A su vez,
puede determinar la presencia de componentes vegetales regulando el establecimiento y
crecimiento de determinada especie vegetal (Davidson, 1993).
Se ha probado que la fisonomía de la vegetación puede determinar la abundancia y
distribución de aves en un ecosistema, al estar relacionada con recursos tales como alimento,
sitios de nidificación o refugio (Rotenberry y Wiens, 1980), no obstante hace falta mayor
investigación enfocada al estudio de las interacciones planta-animal, para comprender mejor las
complejas relaciones presentes en el bosque y así, proponer alternativas para su manejo. Con base
en lo anterior, surgen las siguientes preguntas objeto del presente trabajo ¿La vegetación influye
sobre la abundancia y diversidad de depredadores de semillas de Pinus teocote en 2 tipos de
hábitat (bosque conservado y pastizal)?, ¿La diversidad y abundancia de depredadores de
semillas de Pinus teocote se relaciona con las características de cada hábitat estudiado?, ¿La
presencia de vegetación influye en el grado de depredación de semillas?, ¿La vegetación es un
elemento importante relacionado con la presencia de depredadores de semillas?, que fueron el
motor del mismo.

3
2. OBJETIVOS
Objetivo General
Analizar el efecto del tipo de vegetación sobre la abundancia y diversidad de depredadores
de semillas de Pinus teocote en dos tipos de hábitat contrastantes: un bosque y un pastizal, en la
Reserva Ecológica de San Juan del Monte.
Objetivos Particulares
Identificar a las aves y sus hábitos alimenticios, presentes en el área de estudio.
Identificar a los ratones y pequeños mamíferos así como sus hábitos alimenticios, presentes
en el área de estudio.
Identificar los principales depredadores de semillas de Pinus teocote.
Determinar la diversidad y abundancia de los depredadores de semillas en dos tipos de
hábitat (bosque y pastizal).
Comparar la tasa de depredación de semillas en dos hábitats con condiciones diferentes de
vegetación.
3. HIPÓTESIS
La vegetación es un factor fundamental que influye sobre la abundancia y diversidad de
especies de aves y roedores que depredan las semillas de Pinus teocote.

4
4. REVISION DE LITERATURA
4.1. Efecto de la vegetación sobre poblaciones de aves y roedores en algunos ecosistemas:
Una revisión
4.1.1. Importancia de las interacciones ecológicas planta-animal
Los bosques están formados por una gran variedad de especies de plantas y animales, son
sistemas dinámicos ya que en ellos los árboles de gran altura dan sombra y pueden determinar
cambios de luz, temperatura y humedad bajo sus copas, de esta manera crean microhábitats, que a
su vez, son utilizados por otras especies que requieren esas condiciones, ya sea para obtener
refugio o alimento (Roldán et al., 2001). Las hojas que caen de los árboles, constituyen una
importante fuente de nutrientes para el mismo ecosistema, estas son degradadas por
microorganismos que facilitan el desdoblamiento de nutrientes para que sean fácilmente
absorvibles. La disponibilidad de alimento en un ecosistema fomenta que existan relaciones muy
estrechas relacionadas a la ocupación de hábitats y a la distribución de los depredadores
(Bonaventura y Suárez, 1992).
Las interacciones ecológicas son muy importantes en el funcionamiento de los ecosistemas,
así como, en la supervivencia de poblaciones de flora y fauna, y están estrechamente relacionadas
con procesos evolutivos. Durante la evolución de estas interacciones, cada organismo desarrolla
diversas adaptaciones o especializaciones, que hacen aún más estrechas estas relaciones inter-
específicas (Cueto et al., 1995). En los bosques de pino se pueden observar ejemplos de
estrategias adaptativas como: las plantas del sotobosque adaptadas a vivir bajo la sombra de los
árboles, mismas que crean características microclimáticas particulares; algunos pájaros
migratorios que en cierta época del año sólo comen semillas, en otra, se alimentan de frutos e
insectos, mientras otros únicamente se alimentan de insectos que en ocasiones son dañinos para
las cortezas de los árboles. Las ardillas comen bellotas y otras semillas, los búhos depredan a los
roedores, y los roedores a las semillas, mientras que los coyotes se alimentan de conejos, ardillas
y roedores (Vander Wall, 1992). De igual manera el grado de especialización en el régimen
alimenticio, el sitio elegido para vivir, el tamaño del territorio, la elección de materiales y
ubicación de madriguera y las asociaciones con otros organismos, son el resultado de procesos de
coevolución entre individuos de diferentes especies en el espacio y tiempo, de la acción de la
selección natural que determina un equilibrio en la comunidad y del medio ambiente que
interactúa con ellos (Mills et al., 1991).
Para muchas plantas y animales, la presencia de otros seres vivos en su comunidad, en
ocasiones, es más importante para su supervivencia que los factores abióticos (luz, humedad,
temperatura, etc.), sobre todo en zonas tropicales donde estos factores son más constantes que en
zonas templadas o desérticas. Una de las causas que conduce a que se presenten interacciones
perjudiciales severas, es que el medio experimente cambios drásticos repentinos; como los
provocados por el hombre, mismos que pueden derivarse, por ejemplo, en la aparición de plagas
(Peace, 1962).

5
Los bosques del país son sobre-explotados o deteriorados, para dedicarlos a la
agricultura y/o ganadería, provocando la destrucción de los ecosistemas, la desaparición de
especies y el desequilibrio ecológico. Algunos insectos que se alimentan de las plantas del
bosque, pueden encontrar una forma de vida en los cultivos, aunque bajo estas nuevas
condiciones puede no existir el factor biológico que controlaba la población (ejemplo aves
insectívoras), por lo que la población del insecto crecerá sin control convirtiéndose en una plaga
(Labrador y Alfieri, 2001).
Las interacciones ecológicas en la naturaleza se presentan en dos niveles: entre individuos
de la misma especie (interacciones intra-específicas), y entre individuos de diferentes especies
(inter-específicas). Estas relaciones a su vez, pueden ser benéficas o perjudiciales (también
denominadas positivas o negativas) para los organismos que interactúan, dependiendo del efecto
que tiene dicha interacción sobre los individuos, por ejemplo: un árbol que depende de cierto
organismo para ser polinizado estará en riesgo de desaparecer si el agente polinizador es
destruido; o una planta que difícilmente crece bajo la sombra de otras, puede ser favorecida si
otro organismo se alimenta de plantas de rápido crecimiento o colonizadoras, permitiéndole el
desarrollo de la primera (Begon et al., 1996). Cuando las condiciones del medio son optimas para
la vida, algunos organismos dominan y suprimen a sus asociados, pero cuando las condiciones
cambian o son desfavorables en cierta época del año, las asociadas se convierten en dominantes,
esto es común verlo en especies de aves estacionales o migratorias (Fretwell y Lucas, 1970).
Las relaciones interespecíficas se llevan a cabo en una comunidad entre individuos de
especies diferentes dentro de un ecosistema dentro de estas relaciones tenemos las siguientes
(Begon et al., 1996):
Mutualismo: Interacción entre dos especies en la que ambas se benefician en el uso de un
recurso (+/+).En el caso de los líquenes, las algas aportan el alimento, y los hongos el soporte y la
humedad, en el suelo existe un tipo de bacterias llamado Rhizobium, que se asocia e interactúa
con las raíces de algunas plantas leguminosas, como el trébol. Esta asociación tiene como
resultado la obtención de nitrógeno, capturado por las bacterias y que el trébol utiliza para
fabricar sustancias químicas. Estas sustancias permiten la supervivencia de ambos, puesto que,
tanto el trébol como la bacteria se benefician en esta interacción.
Depredación: Cuando los miembros de una población se alimentan de los de otra
población, no siempre significa la muerte de la presa (+/-). Por ejemplo un ratón que se alimenta
de alguna plántula en su totalidad, o un grillo que se alimenta de partes de una hoja o herviboría.
Comensalismo: Son relaciones que solo benefician a uno de los organismos y no perjudican
al otro (+/0). un ejemplo de ellas son las aves carroñeras que se alimentan de los restos de
animales que otros organismos cazan o el pez rémora que nada junto al tiburón y se alimenta de
los restos de comida que este deja.

6
Parasitismo: es la interdicción entre dos organismos de manera que uno de ellos se
beneficia a expensas del otro (-/+). Este tipo de interacción es muy común en animales
domésticos como el perro o el gato, que suelen ser afectados por "huéspedes" tan impopulares
como las pulgas. El beneficiado se denomina huésped o parásito y el parasitado hospedante. Se
aprecia claramente que el huésped se beneficia de la relación, ya que el hospedante, gato o perro,
le ofrece un lugar donde vivir, protección y alimento al parásito, pero se ve dañado por la
presencia del indeseable invitado. Sin embargo, el hospedante rara vez corre peligro de muerte,
pues si esto ocurriera el huésped también moriría.
Competencia: es la relación que se establece entre poblaciones que necesitan un recurso
del ambiente y que se encuentra en cantidades limitadas (-/-). Dos seres vivos pueden competir
por el mismo alimento si éste escasea en el ambiente, en esta interacción los organismos
compiten entre sí por un factor de tipo biótico. También puede establecerse la interacción de
competencia entre dos poblaciones cuando escasean factores de tipo abiótico. Así, dos plantas
podrán competir por la cantidad de agua que hay en el suelo, o bien, dos especies de aves podrán
competir por el lugar donde construir sus nidos, es decir, por el espacio de nidificación. Como
resultado de la interacción de competencia ambas especies se ven perjudicadas
Amensalismo: es la interacción que se produce cuando un organismo se ve perjudicado en
la relación y el otro no experimenta ninguna alteración es decir la relación le resulta neutra (-/0).
En algunos bosques de la selva amazónica, hay árboles de mayor tamaño que impiden la llegada
de luz solar a las hierbas que se encuentran a ras del suelo, éste es un ejemplo de amensalismo, y
se diferencia de la competencia en que las plantas de menor tamaño no afectan en nada la
supervivencia de los árboles de mayor tamaño.
Neutralismo: dos especies conviven sin afectarse mutuamente, dos poblaciones se
encuentran simultáneamente en el ambiente sin que exista relación entre ellas (0/0). En un bosque
templado habitan pájaros carpinteros y mapaches ambos se encuentran en el ambiente mas no
existe una relación entre ellos.
Como se puede ver, las relaciones planta-animal son muy importantes en los procesos
ecológicos de los ecosistemas, en el caso que nos ocupa, depredación de semillas por aves y
roedores, esta interacción se relaciona con la abundancia y diversidad biológica del sitio, con la
presencia de carnívoros y puede afectar la regeneración de una comunidad de árboles (Connell y
Slatyer, 1977).

7
4.1.2. Efecto de la vegetación sobre la abundancia y riqueza de aves en un ecosistema
La vegetación es un elemento importante para las aves en un ecosistema, ya que es un
componente primordial en la cadena alimenticia, proporcionando semillas y frutos que dan
sustento a la población (Cueto et al; 2001). Un estudio realizado en la estación de investigación
“La Selva” en Costa Rica, mostró que la eliminación de una sola especie arbórea (Casearia
corymbosa) afectó a la población de tucanes, ya que durante la época de escasez de frutos de
otras especies vegetales, los tucanes utilizaban los de C. corymbosa como sustento para su
alimentación; lo que trajo como consecuencia que la población de tucanes disminuyera, dando
como resultado una menor dispersión de las semillas de este árbol, que depende del tucán para
ello (Howe y Smallwood, 1982).
La presencia de árboles en un ecosistema favorece la biodiversidad y abundancia de
muchas aves que interactúan de diversas maneras con la vegetación. La restauración de hábitats
ha puesto de manifiesto que mientras más vegetación nativa y bien conservada exista en las
zonas, mayor será la diversidad de animales así como de endemismos (Panayotou y Ashton,
1992; Wyatt-Smith et al., 1963). A su vez, se ha visto que la heterogeneidad ambiental que
presenta un sitio y que es determinada principalmente por la vegetación es importante en la
distribución y abundancia de aves (Rotenberry, 1985). La vegetación no perturbada influye en la
distribución y abundancia de depredadores potenciales de semillas (Mittelbach y Gross, 1984),
sin embargo, si esta vegetación es perturbada se producen cambios drásticos en las poblaciones
vegetales y animales dando lugar al establecimiento de nuevas poblaciones e interacciones entre
las especies (Picket y Cadenasso, 1995). En bosques de Pinus ponderosa sujetos a tala y/o quema
en Arizona, Estados Unidos, se encontró que las perturbaciones al bosque fueron acompañadas
de un aumento sustancial en la densidad de aves granívoras, y cuando estos dos factores (tala y
fuego) se presentaron combinados, la comunidad de aves estuvo compuesta casi exclusivamente
por aves granívoras (Blake 1982).
El consumo de semillas y las relaciones de competencia entre aves granívoras, constituyen
dos tipos de interacciones de importancia en la estabilidad y diversidad de un ecosistema
(Bustamante y Marone, 2001). Las semillas dispersadas a mayor distancia de la planta madre,
tienen mayor probabilidad de sobrevivir a la depredación; mientras que semillas depositadas bajo
la copa del árbol o a poca distancia, tienen menos probabilidades de sobrevivir debido a procesos
denso-dependientes, así como a la presencia de depredadores de semillas, ataques de patógenos,
competencia entre plántulas entre otros (Janzen, 1971). Las preferencias de las aves por distintos
tipos de semillas, indica que éstas tienen un efecto sobre la composición del banco de semillas y
en última instancia, sobre la composición y abundancia de la vegetación (Cueto et al., 2001).

8
La diversidad de aves en diferentes ecosistemas depende de factores que actúan espacial y
temporalmente. En ambientes terrestres, la estructura de la vegetación determina la distribución
de aves al estar asociadas con recursos críticos tales como alimento, sitios de refugio, resguardo
contra depredadores y sitios para el anidamiento (Isacc y Martínez, 2001). En los bosques se
presentan diversos microhábitats con condiciones particulares (temperatura, humedad,
iluminación, etc.), creando un ambiente heterogéneo que favorece la presencia de ciertas especies
de aves, que requieren características específicas para sobrevivir (James y Wamer, 1982). Así, las
aves granívoras seleccionan los sitios de alimentación de acuerdo con las variaciones espaciales y
temporales del recurso alimenticio disponible, lo cual, no sólo tiene repercusiones para las aves,
sino también para las poblaciones de plantas, (Marone, 1991).
La disponibilidad de recursos alimenticios en un ecosistema se ha propuesto como uno de
los factores más importantes en la estructura y composición de las comunidades de aves
(MacArthur y Levins, 1964; Cody, 1968; Fretwell, 1972). En los bosques de pino, los incendios
tienen un efecto dual, ya sea al abrir los conos serótinos (conos que requieren de incendios para
abrir y liberar las semillas), como de estimular la producción de semillas; además, las gramíneas
presentan una gran cobertura, por consiguiente las aves granívoras son más abundantes en
relación con otros gremios alimenticios. Así, la presencia y abundancia de aves nectarívoras,
insectívoras o frugívoras, son escasas en bosques de pino, donde por lo general estos recursos son
escasos (Nocedal, 1984).
La estructura de la vegetación determina la diversidad de aves. La disponibilidad de
alimento para aves, sitios de percha y distintos tipos de sustratos de forrajeo están estrechamente
ligados con la estructura de la vegetación. En bosques templados del Valle de México, se
encontró que las aves utilizaron selectivamente distintos perfiles de estratificación del follaje,
esta ocupación varia con respecto a los meses del año, posiblemente como resultado de la
presencia de alimento o refugio (Nocedal, 1984). Las preferencias de las aves granívoras por
distintos tipos de semillas, permiten medir las consecuencias del consumo de semillas sobre los
bancos de semillas del suelo, a su vez, sobre la composición y abundancia de la vegetación
(Cueto et al., 2001).
4.1.3. Efecto de la vegetación sobre la abundancia y riqueza de roedores en un ecosistema
La vegetación es un elemento importante para las poblaciones de roedores que habitan un
ecosistema ya que proporciona semillas, frutos y raíces que les sirven de alimento, al mismo
tiempo los espacios libres entre las raíces de los árboles, debajo de los zacatonales y de las
cortezas son utilizados como madrigueras.

9
Estudios realizados en la Isla de Barro colorado en Panamá, demostraron que bajo la copa
de los árboles que producen frutos o semillas se presenta una mayor densidad de granívoros
(depredadores de semillas); un típico ejemplo de una relación denso-dependiente, es decir, que la
densidad de depredadores bajo la copa de los árboles depende de la cantidad de semillas
producida. Esta situación disminuye la probabilidad de supervivencia de una semilla que cae
cerca del árbol que la produjo. Mientras que, aquellas semillas dispersadas lejos del árbol padre
mediante el viento o por algún frugívoro, tienen mayor probabilidad de supervivencia, pues
disminuye la densidad de depredadores al aumentar la distancia de dispersión (Janzen, 1970;
Howe, 1984). Así, la probabilidad de maduración de una plántula o semilla dependen de: (1) la
cantidad de semillas producidas, (2) el tipo de agente dispersor, (3) la distancia del árbol padre, y
(4) la actividad de los depredadores de semillas y plántulas figura 1 (Janzen, 1970).
PI
PRC
Distancia del árbol padre
Número desemillas por
unidad de área
Probabilidad de supervivencia que tiene la semilla o
plántula
Figura 1. Se señala la probabilidad de supervivencia de las semillas bajo la copa del árbol que las
produjo, P probabilidad de sobrevivencia de la semillas en relación con la distancia del árbol
padre; I número de semillas con respecto a la distancia del árbol padre; PRC curva que marca la
probabilidad de establecimiento de un nuevo individuo (Janzen, 1970).
La vegetación proporciona microhábitats particulares que favorecen la presencia y
abundancia de ciertas especies que requieren características muy específicas para sobrevivir. Un
árbol puede crear características microclimáticas (temperatura, humedad, iluminación, etc.) que
favorecen la presencia de ciertos roedores, que se establecen donde hay semillas en abundancia,
cavan túneles y madrigueras que promueven la filtración del agua y la aireación del suelo; sin
embargo, aumentan las probabilidades de depredación de las semillas (Janzen ,1971; Mittelbach
y Gross 1984; Geenwood, 1985; Graves et al., 1988; Manson y Stiles, 1998). El recurso
alimenticio que proporcionan los árboles para diversos organismos, es uno de los elementos más
importantes para el mantenimiento de una población animal (Moen et al., 1993). Por lo tanto, el
decremento en una población de roedores que se alimentan específicamente de semillas de cierto
árbol puede estar relacionado al descenso en las poblaciones de otras especies de árboles que les
brindan alimento. El impacto causado por el hombre por la desforestación, resulta más dañino si
existen roedores específicos de esos sitios, pudiendo disminuir el número de individuos en una
población.

10
La diversidad de componentes vegetales en un área, está directamente relacionada con la
estructura de la vegetación, que a su vez, favorece la creación de microclimas, y microhábitats
ocupados por una gama de organismos terrestres (Hornfeldt, 1994). Un estudio sobre el ratón de
campo Peromyscus difficilis, indicó que esta especie es mas abundante cuando el recurso
alimenticio es copioso, el cuál se vincula con su éxito reproductivo; así un hábitat que brinda
refugio, alimentación y condiciones favorables, es un lugar idóneo para la reproducción,
(Galindo-Leal y Krebs, 1998). Por esto mismo, la presencia y abundancia de recursos
alimenticios para ciertas especies puede influir sobre las interacciones sociales y la distribución
espacial de los individuos (Davies y Lundberg, 1984; Ostfeld, 1985; Rubenstein, 1982), la
interrupción de estas interacciones, podría afectar las características demográficas de una
población de roedores (Dunbar, 1985), y consecuentemente de la vegetación.
La cubierta vegetal puede ser uno de los más importantes factores que determinan la
distribución espacial de roedores y el uso del hábitat de Cricétidos (ratas y ratones de campo)
(Suárez y Bonaventura, 2001). En la parte baja del Delta de Paraná, en Argentina, la distribución
de las especies de roedores se encontró extremadamente influenciada por la estructura de la
vegetación, las ratas Akodon azarae y Oxymycterus rutilans tuvieron una preferencia marcada
por hábitats con alta cobertura herbácea (Bonaventura et al., 1991; Bonaventura et al., 1992;
Cueto et al., 1996), Scapteromys tumidus prefirió hábitats con alta cobertura de vegetación arriba
de los 50 cm de altura (Cueto et al., 1995) y Oligoryzomyz delticola se restringió a hábitats
boscosos y arbustivos (Langguth, 1963; Barlow, 1969), esta distribución responde a variables
como la disponibilidad de alimento y las condiciones del hábitat (Suárez y Bonaventura, 2001).
Sitios como las praderas cuya vegetación se restringe a herbáceas y algunos elementos
arbóreos, pueden ser utilizados por roedores como corredores de alimentación, fomentando con
esto la repoblación arbórea en zonas degradadas (Tamarin et al., 1984). La invasión de árboles
hacia otros campos puede estar determinada por la depredación de semillas y plántulas por
roedores en bosques y campos cercanos, los efectos depredatorios de los roedores sobre los
propágulos de árboles pueden ser modificados por la variación en la distribución y abundancia de
propágulos de árboles de acuerdo a épocas del año (Manson et al., 1998).
En el Este de Estados Unidos la depredación de semillas y plántulas realizada por 2
especies de roedores (Peromyscus leucopus y Microtus pennsylvanicus) reguló la invasión de
árboles a otros sitios mediante el consumo de las semillas que formaban el banco de semillas
(Mittelbach y Gross, 1984; Gill y Marks, 1991; Ostfeld y Canham, 1993; Ostfeld, et al., 1997), la
distribución y abundancia de semillas y plántulas en un bosque, puede influir en la abundancia de
pequeños mamíferos granívoros consumidores de las mismas, permitiendo con esto, tener una
idea del impacto de los roedores en la invasión de árboles hacia otros sitios (Manson et al., 1998).

11
Los campos que han sido invadidos por nuevas especies vegetales presentarán cambios en
la composición de especies animales, ya que existen animales vinculados estrechamente con
especies vegetales ya sea en cuestión de refugio o de alimentación (McDonnell y Stiles, 1983;
Myster y Pickett, 1993). La falta de cubierta vegetal en determinado sitio, puede significar un
factor de riesgo para especies de roedores, los árboles brindan protección contra depredadores,
por consiguiente, el riesgo de ser depredado puede ser un elemento importante en la decisión de
alimentarse en un hábitat o microhábitat en particular (Manson y Stiles, 1998). Mientras mayor
cantidad de microhábitats proporcione determinado tipo de vegetación, la presencia de especies
relacionadas a esos microhábitats aumentará (Janzen, 1970).
Cuando un área que cuenta con vegetación conservada es perturbada, las hembras de
roedores son más susceptibles a los cambios presentados por esta perturbación (cambios
climáticos y sobre su hábitat), y aún más, cuando están en actividad reproductiva; ya que si
escasea el alimento, la madre no proporcionará alimento a sus crías, perdiendo toda la camada
(Anderson, 1989). En estas circunstancias, un roedor macho puede recorrer mayor distancia en
busca de alimento, que una hembra gestante, que necesita mayor energía para desplazarse. La
distancia que un macho recorre, puede ser un factor de riesgo de depredación; los machos
enfocan su actividad en dos aspectos: a) mayor consumo de semillas, b) diversificación de su
alimentación; al no haber una buena cantidad de semillas para alimentarse, se suplementa con
semillas de otras especies vegetales, favoreciendo el establecimiento de nuevos árboles, arbustos
y herbáceas, actuando como control biológico de las especies vegetales de las que se alimenta.
La estructura de la vegetación y los componentes que la forman se relaciona con la
presencia de microhábitats específicos a ciertas especies, por esto mismo la selección de un
hábitat por un granívoro, se reflejará en sus estrategias de sobrevivencia y de reproducción. Un
organismo que habita un tipo de hábitat específico y que produce poca descendencia, es más
propenso a extinguirse localmente si aumentan las perturbaciones en su ambiente, que un
organismo adaptado a una amplia gama de hábitats con descendencia numerosa, aunque sea
mayormente depredado por carnívoros tendrá una estructura demográfica bien conformada y su
población será más estable (Pulliam y Danielson, 1991).

12
5. MATERIAL Y MÉTODO
5.1. Área de Estudio
El sitio de estudio correspondió al área natural protegida de San Juan del Monte, dicha
zona esta conformada con bosques de segundo crecimiento, debido a los aprovechamientos
forestales llevados a cabo en el sitio. El estudio aborda la depredación de semillas de Pinus
teocote ya que esta especie es la dominante del bosque en el sitio (68% de cobertura), ocupando
un total de 412.47 ha de 609 ha que comprenden el área natural protegida (Gobierno del Estado
de Veracruz y Coordinación Estatal de Medio Ambiente, 2002).
5.1.1. Localización del Área de Estudio
San Juan del Monte se ubica en la vertiente norte del Volcán Cofre de Perote (figura 2),
entre las coordenadas geográficas 19º 39´ 00´´ y 19º 35´ 00´´ norte y 97º 05´ 00´´ y 97º 07´ 30´´
oeste, dentro del Municipio de Las Vigas de Ramírez Cano, Veracruz (Gobierno del Estado de
Veracruz y Coordinación Estatal de Medio Ambiente, 2002).
Figura 2. Ubicación Geográfica del sitio de estudio, el Área Natural Protegida de San Juan del
Monte, las Vigas de Ramírez, Veracruz.

13 5.1.2. Características Físicas
5.1.2.1. Clima
El clima es templado subhúmedo C (W1), corresponde al más seco de los templados,
presenta lluvias en verano con una precipitación entre 43.2 mm y 55 mm, y porcentaje de lluvia
invernal entre 5 mm y 10.2 mm (Soto y Angulo, 1990).
Se presentan nublados frecuentes, ya que la humedad generada en el Golfo de México
golpea la zona. Los inviernos son muy fríos con largos periodos de neblinas. En los primeros
meses del año las heladas son muy frecuentes, el verano es fresco con lluvias abundantes y
noches despejadas (Soto y Angulo, op. cit.).
5.1.2.2. Fisiografía
El Área Natural Protegida de San Juan del Monte se ubica en la provincia fisiográfica del
eje neovolcánico y en la subprovincia de los lagos y volcanes del Anáhuac. La región se
caracteriza por constar de sierras volcánicas, alternadas con llanuras y vasos lacustres. Dentro de
este tipo de zonas se encuentra el cofre de Perote, que es una masa montañosa de origen
volcánico ya erosionada, constituida por rocas lávicas andesíticas que marcan las características
típicas del Cofre (Geissert et al., 1994).
En San Juan del Monte predominan los lomeríos con orientación Norte, Noroeste. El 32%
del predio presenta pendientes suaves de entre 1 y 6º; el 58% con pendientes moderadas en un
rango de 6 a 22º y el 10% restante presenta pendientes mayores a 22º pero sin rebasar los 39º
(INEGI-ORSTORN, 1991). En promedio presenta una altitud de 1327 msnm, el punto más alto
es la Cueva del Obispo a 3100 msnm, y el más bajo del área corresponde a la parte norte con
2470 msnm en el paraje denominado el “Tostón”.
5.1.2.3. Hidrología
San Juan del Monte se encuentra ubicado en las regiones hidrológicas 27 y 28, sobre la
cuenca del Río Bobos y la del Río Actopan, ambos ríos escurren hacia el Golfo de México
(Geissert et al., 1994). Dentro del Área Natural no se aprecian escurrimientos importantes, la
mayoría son intermitentes, todos los que desembocan hacia el río Bobos, se infiltran antes de
llegar a un curso permanente.
En el Cerro de Las Lajas y la comunidad de “El Paraíso”, existe una red de arroyos
intermitentes que se juntan en el arroyo San Juan, que atraviesa el Área Natural Protegida.
Existen 3 manantiales cuyo gasto mayor es en temporada de lluvias, el manantial Palo de Rayo,
es permanente (Gobierno del Estado de Veracruz y Coordinación Estatal de Medio Ambiente,
2002).

14
5.1.2.4. Geología y recursos minerales
El área esta comprendida dentro de la Sierra Volcánica Perote-Orizaba en el eje
neovolcánico transmexicano, el paisaje volcánico esta parcialmente modificado por emisiones
lávicas como es el caso del lugar denominado El Volcancillo, situado 2 km al este del Área
Natural Protegida de San Juan del Monte.
Los perfiles geológicos se sitúan en tres edades: Mesozóico, Oligomioceno y Cenozóico
Cuaternario. La geología del predio se manifiesta por materiales de origen extrusivo, en su mayor
parte brechas volcánicas, tobas intermedias y andesitas, hacia el lado noroeste existe una franja
compuesta de basalto (Geissert et al., 1994) tabla 1.
Tabla 1. Composición geológica del Área Natural Protegida de San Juan del Monte, las Vigas de
Ramírez, Veracruz (Geissert et al., 1994).
LITOLOGÍA PORCENTAJE
Andecita y Dacita 46.6
Basalto 44.19
Flujo de cenizas 6.37
Cono de escroias de caída
(tezontle) 1.97
Flujo de cenizas y bloqueos 0.86
5.1.2.5. Suelos
Los suelos más comunes son el andosol húmico y el andosol ócrico, de origen volcánico,
los cuales frecuentemente tienen un horizonte superficial oscuro rico en materia orgánica. Deben
sus propiedades específicas a la presencia de abundantes minerales amorfos, algunos se presentan
claros en la parte superficial, principalmente en las partes más planas de la zona y negros hacia
las barrancas. Son muy fértiles pero muy frágiles, suelen ser esponjosos o muy sueltos, tienen la
capacidad de almacenar y regular grandes volúmenes de agua, estos suelos se presentan en el
56% del predio (Geissert y Meza, 1996; INEGI-ORSTOM, 1991; Ortiz, 1994)
En el 44% de la superficie se presenta el litosol, suelos limitados en profundidad por un
estrato duro, continuo y coherente, dentro de los 10 cm superiores; son someros frágiles y de
textura media, están ubicados principalmente en la parte oeste del predio, la capa dura que los
define está conformada por tezontle (materiales piroclásticos medianamente consolidados con
alto contenido en hierro, lo que les confiere el color rojizo) (Geissert et al., 1994) tabla 2.

15
Tabla 2. Tipos de suelo presentes en el Área Natural Protegida de San Juan del Monte, las Vigas
de Ramírez, Veracruz (Geissert et al, 1994).
TIPO DE SUELO PORCENTAJE DEL PREDIO
Andosol húmico moderadamente
profundo 46.6
Litosol ándico y andosol ócrico 44.2
Andosol húmico y andosol ócrico
dístrico 4.83
Andosol húmico poco profundo 1.97
Andosol ócrico distrito 1.54
Andosol ócrico dístrico y andosol
húmico poco profundo 0.86
100
5.1.2.6. Aptitud del suelo
Debido a sus características físicas y climáticas, los suelos tienen vocación forestal, no
soportan actividades agrícolas y pecuarias de alto rendimiento. La presencia de litosoles en parte
del predio disminuye la capacidad de crecimiento del bosque y por esta razón es difícil plantear
un aprovechamiento forestal de alta productividad en términos de rentabilidad económica
(Gobierno del Estado de Veracruz y Coordinación Estatal de Medio Ambiente, 2002).
5.1.2.7. Características bióticas
La vegetación de la zona es el bosque de coníferas, los pinos se encuentran ubicados en
complejos arbóreos bien definidos, la mayoría de los pinos constituyen elementos dominantes y
codominantes en la vegetación actual del área. Los pinos tienen preferencia por suelos volcánicos
y la mayor parte de las coníferas toleran mejor los suelos ácidos. Los bosques de pinos con un
estado de sucesión avanzada generalmente constituyen comunidades arbóreas con pocas especies
dominantes (Almeida, 1997).
5.1.2.7.1. Comunidades vegetales
El bosque de pino es el componente principal de la vegetación de San Juan del Monte
donde existen 220 hectáreas afectadas por un incendio en el año de 1993. Se han reportado un
total de 447 especies de flora susceptible a ser encontrada en este sitio de las siguientes familias:
Bromeliaceae, Lauraceae y Pinaceae. Las principales especies encontradas en la zona son Pinus
patula, P. pseudoestrobus, y P. teocote, aunque existan también individuos de Cupressus
benthamii alrededor del antiguo aserradero.

16
El área presenta rodales puros de Pinus teocote y rodales mixtos con P. patula asociado
con P. pseudoestrobus, Quercus ssp., Alnus jorullensis y Arbutus xalapensis, mismos que se
distribuyen de acuerdo al tipo de suelo; P. teocote soporta suelos delgados y poco fértiles como el
litosol, P. patula asociado con P. pseudoestrobus, Quercus ssp., Alnus jorullensis y Arbutus
xalapensis, se encuentran en suelos profundos. En el estrato inferior predominan arbustos como:
Bacharis conferta, Pertnettia ciliata, Acciena elongata y Buddleia sp. En el estrato herbáceo
predomina Alchemilla pectinata, Archibaccharis androgyna, Pteridium aquilinum dentro de las
gramíneas Muhlembergia macroura, Brachypodium mexicanum y Triniochloa stipoides dentro de
las hierbas Piptochaetium pirescens (Berninger y Junikka, 1994; Narave-Flores, 1985). Se
encuentran especies registradas en la NOM 059-SEMARNAT-2001 (anexo 2).
5.1.2.7.2. Comunidades animales
La diversidad biológica presente en el Área Natural Protegida de San Juan del Monte es
producto de variaciones en la topografía y el clima de la zona, lo que crea un mosaico de
condiciones ambientales y microambientales muy particulares. Se han reportado para la zona 55
especies de mamíferos, 23 de aves, 27 de reptiles y 6 de anfibios, no se cuenta con información
sobre invertebrados (Gobierno del Estado y Coordinación Estatal de Medio Ambiente, 2002). Se
encuentran especies registradas en la NOM 059-SEMARNAT-2001 (anexo 3).
El bosque de coníferas en México ocupa el tercer lugar en número de vertebrados terrestres
endémicos para Mesoamérica, el segundo lugar en especies endémicas de México y el cuarto
lugar en especies de distribución restringida (especies que han sido registradas para un sólo tipo
de vegetación), por lo tanto son más susceptibles a cambios en su hábitat (Flores-Villela y Geréz,
1994).
5.2. Métodos
5.2.1. Obtención de semillas para el experimento
A través de recorridos en el Área Natural Protegida de San Juan del Monte, se localizaron
árboles de Pinus teocote con presencia de conos en sus ramas; durante los meses de noviembre-
diciembre, temporada de producción de semilla (Patiño et al., 1983), se colectaron 60 conos
maduros, los cuáles se colocaron en una tela de peyón dispuesta en cajas de cartón y sometidas a la
acción de la luz solar, para agilizar la apertura de los conos y la obtención de semillas (Arriaga et
al.,1994). Una vez abiertos los conos, fueron sacudidos sobre la tela de peyón para separar las
semillas. Se contaron y almacenaron en frascos de vidrio esterilizados (INIFAP, 1994).
1.

17
5.2.2. Selección e identificación de los sitios de muestreo
Dentro del Área Natural Protegida de San Juan del Monte se seleccionaron 4 sitios de 1 ha
cada uno, dos cubiertos de pastizales o zacatonales y los otros dos cubiertos con bosque (figura
3). Los pastizales fueron sitios abiertos cubiertos principalmente de macoyos de Mublembergia
macroura y Brachypodyum mexicanum la distancia entre los macoyos de zacate oscilaba entre
los 10 a 15 cm aunque en algunos macoyos de gran tamaño los espacios entre estos eran mínimos
o no perceptibles creando una cobertura densa, al ser sitios abiertos, en el día manejaban las
temperaturas mas altas y por la noche presentaban temperaturas muy bajas. Los cubiertos por
bosque fueron sitios que presentaron arbolado principalmente de Pinus teocote y árboles de Alnus
jorullensis, en su estrato herbáceo contaban con la presencia de helechos Pteridium aquilinum y
hierbas como Alchemilla pectinata a su vez de estar cubiertos por hoja de pino (ocochal) por el
día eran sitios frescos con humedad misma que se generaba por la presencia de árboles y por la
noche eran sitios fríos, no obstante de que su topografía no era plana como en los pastizales.
Figura 3. Localización de los sitios de muestreo en el Área Natural Protegida de San Juan del
Monte, las Vogas de Ramírez, Veracruz. Los puntos blancos indican los sitios que se estudiaron,
dos correspondientes a pastizal y dos a bosque, la simbología indica la vegetación actual
existente en el área.

18
Los sitios se marcaron con una placa de PVC rotulada, que indicó el siguiente texto acorde
al sitio de muestreo: Universidad Veracruzana, Instituto de Genética Forestal, Investigación sobre
interacciones ecológicas, Pastizal, Superficie de muestreo 1 ha, de la misma manera para el
Bosque.
5.2.3. Fase experimental
Con el objetivo de identificar a las aves y sus hábitos alimenticios presentes en la zona de
estudio se realizaron observaciones directas de aves depredando semillas de Pinus teocote, el área
se visitó cuatro veces al mes durante un año (enero-diciembre de 2003). Durante las visitas
mensuales, se dedicaron dos días a los sitios de bosque y dos días a los pastizales. Se registraron,
contaron e identificaron las aves (Peterson y Chalif, 1989; Howell y Webb, 2001) que visitaron y
se percharon en los sitios de muestreo. Las observaciones se realizaron mediante unos
binoculares (UNIC 10 x 50) en cuatro periodos diarios de observación (6-8 am, 10 a 12 am, 2 a 4
pm y 6 a 8 pm), a una distancia de 200 m (Bernis, 1966).
Con el objetivo de identificar a los ratones y pequeños mamíferos de la zona de estudio,
se colocaron 10 trampas sherman (5 en cada sitio) durante 57 noches a lo largo del año
mencionado. Los especimenes capturados se identificaron a nivel de especie (Hall y Dalquest,
1963), se les tomaron medidas morfométricas (largo de la cola, longitud de la oreja, largo de la
pata, longitud total, sexo, si fue macho condición testicular) y se liberaron. Se conservó un macho
y una hembra de cada especie (piel y cráneo).
Para identificar a los principales depredadores de semillas de Pinus teocote, durante el día
se colocaron 5 cajas de petri plásticas cada 200 m en cada uno de los 4 sitios de observación.
Cada caja contenía 10 semillas de P. teocote, a las que se les retiró el ala de dispersión, con el
objeto de que el viento no las removiera (Manson y Stiles, 1998; Bernis, 1966; Patiño et
al.,1983). Mediante el uso de los binoculares se identificaron las aves que consumieron las
semillas, durante los periodos de observación mencionados. Cada dos horas se contaron las
semillas y se repusieron las depredadas. Igualmente durante las noches de muestreo se colocaron
las cajas de petri con semillas de P. teocote cerca de las trampas sherman, cuando se colectó
algún ratón se asumió que las semillas depredadas de la caja petri cercana a la trampa fueron
depredadas por ese ratón. En ningún momento durante el día o la noche se observó algún otro
depredador (hormigas, escarabajos, etc.) de las semillas colocadas en las cajas de petri.

19 5.3. Análisis Estadístico.
Se determinó la abundancia y la composición de la comunidad de roedores y aves
tomando en cuenta su orden, familia, especie, gremio alimenticio y para el caso de aves, estatus
(residente o migratorio). La variabilidad del número de aves y roedores entre sitios, se evaluó
mediante una prueba de X² (Zar, 1999).
La riqueza de especies en cada tipo de hábitat se comparó mediante curvas de
acumulación de especies, que predicen el número máximo de especies (la asíntota) presentes en
cierta área. De acuerdo al número de especies acumuladas durante el periodo de muestreo, se
asume que la probabilidad de añadir una nueva especie disminuye linealmente con el tamaño
de la lista, donde finalmente todas las especies serán registradas (Soberón y Llorente, 1993). Se
ajusto la función mediante una regresión no lineal usando el algoritmo quasi-newton del
paquete estadístico Statística (Stat Soft, 1998).
Para determinar si existía diferencia entre el consumo total de semillas llevado a cabo
por aves y roedores en los dos tipos de hábitat estudiados, se realizó una prueba de X² (Zar, op.
cit.).
La diferencia entre el consumo de semillas en cada área, así como la comparación de la
abundancia de aves y roedores se realizó mediante una prueba de X² (Zar, op. cit).
Para determinar el efecto de la vegetación sobre la abundancia de depredadores de
semillas de Pinus teocote (aves y roedores) se realizaron análisis de varianza (Zar, op. cit.).
Para conocer si existe diferencia entre la presencia de vegetación y el número de especies
presentes, así como la vegetación en relación al grado de depredación de semillas por aves y
roedores, se utilizó la siguiente formula (Moreno, 2000) pastizal (H1) y bosque (H2):
FiLog²Fi–(FiLogFi)²/n
S²H2=
n²
Donde S² es igual a la varianza, H1 es igual al Pastizal, H2 es igual al Bosque, Fi Log Fi
es igual al Logaritmo de organismos observados de una especie por el número de organismos
observados de esa especie, Fi log ² Fi es igual al logaritmo de organismos observados de una
especie elevado al cuadrado por el número de organismos observados de esa especie, n² es
igual al número total de organismos observados al cuadrado.

20
Se comparó la diversidad y abundancia de depredadores de semillas de cada tipo de
hábitat mediante el uso del índice de diversidad de Shannon-Wiener (Dirzo, 1990; Moreno,
2000); para la determinación del índice de Shannon se utilizó la siguiente fórmula para cada
sitio de muestreo:
Donde n log n es igual al logaritmo del total de organismos observados por el total de
organismos observados, ΣFi log Fi es igual a la sumatoria del logaritmo del número de
organismos observados de todas las especies por el numero de organismos observados de
todas las especies, y n es el numero de organismos observados de todas las especies para cada
área de estudio. La prueba de diversidad de Shannon-Wiener se aplicó para cada sitio, para
determinar si la vegetación influye en el número de especies presentes. La significancia se
evaluó con una prueba de t (Zar, 1999):
t = H´1 – H´2 / SH´1 – H´2
Donde t es igual a H´1 (índice de Shannon del pastizal) – H´2 (índice de Shannon del
bosque), entre la varianza del índice de Shannon del pastizal (H´1) menos el índice de
Shannon del bosque (H´2).
La varianza del índice de Shannon del pastizal menos el índice de Shannon del bosque,
es igual a la raíz cuadrada de la varianza del índice de Shannon del pastizal, mas la varianza
del índice de Shannon del bosque.
Los grados de libertad que permiten ver la significancia en la prueba t se obtuvieron con la
siguiente formula (Zar, 1999):
La elaboración de la base de datos, así como las tablas y gráficos de las especies
observadas y colectadas en el sitio se realizó mediante el paquete STATISTIC, en su modulo
de tables y banners y Excel (Statsoft, 1998, Excel Office XP, 2001).

21
6. RESULTADOS
6.1. Aves presentes en la zona de estudio
Se registraron un total de 2099 aves de 43 especies, pertenecientes a 12 familias, agrupadas
en 3 órdenes: Passeriformes, Piciformes y Caprimulgiformes. El 95% del total de aves registradas
perteneció al orden Passeriformes con 9 familias y 39 especies (tabla 3). El 57% de las especies
(24 especies) incluyen semillas en su dieta: 20 de estas especies (48%) son de hábitos granívoro-
insectívoro, 3 especies (7%) son granívoro-insectívoro-frugívoro, y 1 especie (2%) es granívoro-
nectarívoro (Ramírez Albores y Ramírez Cedillo, 2002; Howell y Webb, 2001).
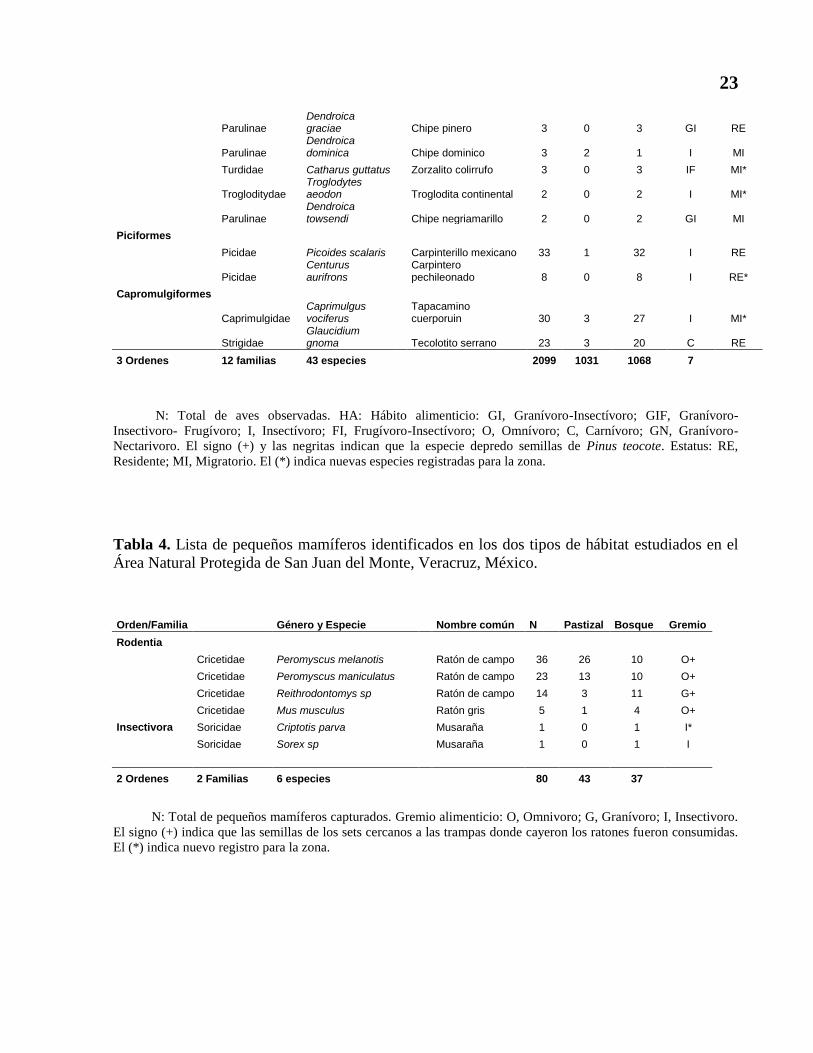
6.2. Ratones y pequeños mamíferos presentes en la zona de estudio
Se capturaron un total de 80 pequeños mamíferos de seis especies, de los cuales 78 fueron
roedores y dos musarañas (insectívoras), pertenecientes a dos familias, agrupadas en dos órdenes:
Rodentia e Insectívora (tabla 4).
6.3. Aves depredadoras de semillas de Pinus teocote.
De las 24 especies de aves granívoras registradas en la zona (tabla 3), el 50% de las
especies depredaron semillas de Pinus teocote y nueve de éstas fueron las más abundantes en las
zonas de estudio. En el pastizal representan el 36% (755.6) de la abundancia total (2099 = 100%),
mientras que en el bosque el 20% (419.8) (tabla 3).
6.4. Ratones depredadores de semillas de Pinus teocote.
Se colectaron un total de 78 ratones pertenecientes a cuatro especies (tabla 4). Las cuatro
especies de roedores pertenecen al gremio alimenticio omnívoro e incluyen en su alimentación
semillas de Pino (Cevallos-González y Galindo-Leal, 1984). Se determinó que estas cuatro
especies depredaron semillas de Pinus teocote, ya que las cajas de petri que contenían estas
semillas y estaban cercanas a las trampas no contaban con éstas, y presentaban excretas de
ratones. De las cuatro especies de roedores capturados, dos fueron las más abundantes en las
zonas de estudio, en el pastizal representan el 90.7% de la abundancia total y en el bosque
representan el 57.1% (tabla 4), sin incluir a las musarañas.

22
Tabla 3. Lista de especies de aves identificadas en las dos tipos de hábitat
correspondientes al Área Natural Protegida de San Juan del Monte, Veracruz, México.
Orden/Familia Género y Especie Nombre común N Pastizal Bosque Hábito Estatus
Passeriformes
Emberizinae Junco phaeonotus Junco Ojilumbre 285 154 131 GI+ RE
Turdinae Turdus migratorius Zorzal pechirojo 146 58 88 GIF+
RE
Parulinae Vermivora peregrina Chipe peregrino 137 77 60 GI+
MI
Emberizinae Junco hyemalis Junco ojioscuro 123 73 50 GI+ MI*
Fringillidae Carpodacus mexicanus Gorrión Doméstico 118 64 54 Gi+ RE
Emberizinae Spizella passerina Gorrión cejiblanco 114 104 10 GI+ RE*
Parulinae Wilsonia pusilla Chipe coroninegro 99 99 0 GI+ MI
Fringillidae Carduelis Pinus Jilguero pinero rayado 95 2 93 GN RE
Emberizinae Poocetes gramíneus Gorrión zapatero 93 86 7 GI+ RE*
Turdidae Turdus grayi Primavera 89 54 35 GI+ RE
Corvidae Cyanocitta stelleri Chara copetona 79 2 77 O RE
Sylvidae Regulus calendula Reyezuelo sencillo 77 77 0 GI MI*
Parulinae Dendroica pinus Chipe nororiental 66 1 65 GI MI*
Corvidae Aphelocoma ultramarina Chara pechigris 65 6 59 O MI*
Emberizinae Aimophila ruficeps Gorrión bigotudo 48 48 0 GI RE*
Parulinae Dendroica occidentalis Chipe negriamarillo 37 1 36 GI+ MI
Emberizinae Sicalis luteola Gorrión americano 34 1 33 GI RE
Fringillidae Carduelis psaltria Jilguero dorsioscuro 33 0 33 GI RE*
Tyrannidae Contopus pertinax Contopus Jose María 30 0 30 I MI*
Aegithalidae Psaltriparus minimus Sastrecito 28 0 28 GIF RE*
Tyrannidae Mitrephanes phaeocercus Mosquerito copetón 22 0 22 I RE*
Ptilogonatidae Ptilogonys cinereus Capulinero gris 22 3 19 GIF MI*
Tyrannidae Contopus borealis Contopus de chaleco 21 21 0 I MI*
Parulinae Peucedramus taeniatus Peeucedramo 21 20 1 GI+ MI
Tyrannidae Myarchus tuberculifer Papamoscas copetón 16 14 2 I RE*
Parulinae Dendroica discolor Chipe Gaalán 15 0 15 GI MI*
Emberizinae Spizella wortheni Gorrión altiplanero 15 15 0 GI MI*
Parulinae Myoborus miniatus Pavito alioscuro 15 0 15 FI MI*
Tyrannidae Empidonax difficilis Empidonax difícil 12 11 1 I MI*
Emberizinae Melospiza lincolnii Gorrión de lincoln 11 9 2 GI+ MI
Tyrannidae Empidonax affinis Empidonax afin 10 10 0 I RE*
Parulinae Dendroica coronata Chipe grupidorado 7 7 0 I MI*
Turdinae Sialia sialis Azulejo gorjicanelo 5 1 4 FI MI
Tyrannidae Empidonax fulvifrons Empidonax canelo 4 4 0 I MI*
23
Parulinae Dendroica graciae Chipe pinero 3 0 3 GI RE
Parulinae Dendroica dominica Chipe dominico 3 2 1 I MI
Turdidae Catharus guttatus Zorzalito colirrufo 3 0 3 IF MI*
Trogloditydae Troglodytes aeodon Troglodita continental 2 0 2 I MI*
Parulinae Dendroica towsendi Chipe negriamarillo 2 0 2 GI MI
Piciformes
Picidae Picoides scalaris Carpinterillo mexicano 33 1 32 I RE
Picidae Centurus aurifrons
Carpintero pechileonado 8 0 8 I RE*
Capromulgiformes
Caprimulgidae Caprimulgus vociferus
Tapacamino cuerporuin 30 3 27 I MI*
Strigidae Glaucidium gnoma Tecolotito serrano 23 3 20 C RE
3 Ordenes 12 familias 43 especies 2099 1031 1068 7
N: Total de aves observadas. HA: Hábito alimenticio: GI, Granívoro-Insectívoro; GIF, Granívoro-
Insectivoro- Frugívoro; I, Insectívoro; FI, Frugívoro-Insectívoro; O, Omnívoro; C, Carnívoro; GN, Granívoro-
Nectarivoro. El signo (+) y las negritas indican que la especie depredo semillas de Pinus teocote. Estatus: RE,
Residente; MI, Migratorio. El (*) indica nuevas especies registradas para la zona.
Tabla 4. Lista de pequeños mamíferos identificados en los dos tipos de hábitat estudiados en el
Área Natural Protegida de San Juan del Monte, Veracruz, México.
Orden/Familia Género y Especie Nombre común N Pastizal Bosque Gremio
Rodentia
Cricetidae Peromyscus melanotis Ratón de campo 36 26 10 O+
Cricetidae Peromyscus maniculatus Ratón de campo 23 13 10 O+
Cricetidae Reithrodontomys sp Ratón de campo 14 3 11 G+
Cricetidae Mus musculus Ratón gris 5 1 4 O+
Insectivora Soricidae Criptotis parva Musaraña 1 0 1 I*
Soricidae Sorex sp Musaraña 1 0 1 I
2 Ordenes 2 Familias 6 especies 80 43 37
N: Total de pequeños mamíferos capturados. Gremio alimenticio: O, Omnivoro; G, Granívoro; I, Insectivoro.
El signo (+) indica que las semillas de los sets cercanos a las trampas donde cayeron los ratones fueron consumidas.
El (*) indica nuevo registro para la zona.

24
6.5. Diversidad y abundancia de depredadores de semillas de Pinus teocote
6.5.1. Diversidad y Abundancia de aves en dos tipos de hábitat (bosque y pastizal)
La diversidad de especies de aves fue significativamente diferente entre los dos tipos de
hábitat (P<0.05). En el bosque, la diversidad (H´= 1.33) fue mayor que en el pastizal (t = 8.41;
P<0.001), donde se registraron 35 especies, de las cuales 11 (26%) fueron exclusivas; mientras
que en el pastizal, se registraron 32 especies, 7 (17%) de las cuales fueron exclusivas (H´=1.20).
Entre los tipos de hábitat se comparten 24 especies (57%). Se registraron 25 nuevas especies para
la zona (Tabla 3). La abundancia de aves no presentó diferencias significativas entre los tipos de
hábitat (X2
= 26.01, P>0.05). En el bosque se registraron un total de 1,068 individuos, mientras
que en el pastizal fueron 1,031 (tabla 3).
6.5.2. Diversidad y Abundancia de pequeños mamíferos en dos tipos de hábitat (bosque y
pastizal)
La diversidad de especies de pequeños mamíferos presentó diferencias entre los dos tipos
de hábitat (P< 0.05). En el bosque, la diversidad fue mayor (6 especies) (H´= 0.65) que en el
pastizal (H´= 0.40) (t = 3.84; P<0.001), las dos especies de musarañas representan el 33.33% del
total de especies capturadas y fueron exclusivas del bosque, mientras que el pastizal se
registraron las mismas 4 especies de roedores que en el bosque, no hubo especies exclusivas a
este tipo de hábitat. Entre los hábitats se comparten 4 especies (66.66%), fue registrada una nueva
especie de musaraña para la zona (tabla 4). La abundancia de roedores no presentó diferencias
significativas entre los tipos de hábitat, (X2
= 6.40 P>0.05). En el bosque se capturaron un total
de 35 individuos, mientras que en el pastizal fueron 43 (tabla 4).
6.6. Depredación de semillas
6.6.1. Depredación de semillas por aves
La depredación de semillas llevada a cabo por aves fue significativamente mayor en el
pastizal (733 semillas) que en el bosque (320) (X²= 23.35; P<0.05).
6.6.2. Depredación de semillas por roedores
La depredación de semillas llevada a cabo por roedores fue significativamente mayor en
el bosque (1100 semillas) que en el pastizal (969) (X²= 67.86; P<0.05).
6.7. Consumo total de semillas por aves y roedores en los dos tipos de hábitat estudiados
El consumo total de semillas llevado a cabo por aves y roedores fue significativamente
mayor en el pastizal (1,702 semillas consumidas) mientras que en el bosque fueron (1,402
semillas consumidas) (X²=145.995; P<0.001).

25
6.8. Esfuerzo de muestreo
6.8.1. Curvas de acumulación de especies de aves
El total de especies acumuladas (43 especies) se estabilizó a los 45 días, de un total de 57
días de muestreo (456 horas de observación). En el pastizal, el número máximo de especies (32
especies) se obtuvo a los 41 días, mientras que en el bosque (35 especies) fue a los 45 días (figura
4). La función de acumulación de especies predice un máximo de 43 especies, así, el muestreo
proporciona una muy buena estimación de la riqueza de especies de aves en la zona.
TOTALES
Pastizal
Bosque
Dias muestreados
Núm
ero
de e
specie
s a
cum
ula
das
-5
5
15
25
35
45
55
-5 5 15 25 35 45 55 65
Figura 4. Curvas de acumulación de especies de aves en total y por hábitat registradas en
(pastizal y bosque) en el Área Natural Protegida de San Juan del Monte, las Vigas de Ramírez,
Veracruz. Se presentan en el eje X los días de muestreo, en el eje Y el número de especies
acumuladas totales y para cada hábitat.
6.8.2. Curvas de acumulación de especies de pequeños mamíferos
El total de especies de pequeños mamíferos acumulados (6) en ambos hábitats se
estabilizó a los 53 días de muestreo de un total de 57 días. En el pastizal, el número máximo de
especies se obtuvo a los 30 días, mientras que en el bosque fue a los 52 días (figura. 5). En el
bosque se colectaron un total de 35 individuos, mientras que en el pastizal fue de 43.

26
TOTALES
Pastizal
Bosque
Dias muestreados
Especie
s a
cum
ula
das
0
1
2
3
4
5
6
7
-5 5 15 25 35 45 55 65
Figura. 5 Curva de acumulación de especies de pequeños mamíferos capturados en dos tipos de
hábitat (pastizal y bosque) en el Área Natural Protegida de San Juan del Monte, las Vigas de
Ramírez, Veracruz. Se presentan en el eje X los días de muestreo, en el eje Y el número de
especies acumuladas totales y para cada hábitat.

27 7. DISCUSIÓN
Nuestros datos muestran que la vegetación es un factor importante relacionado con la
diversidad de aves y pequeños mamíferos, en el bosque ambos grupos fueron significativamente
más diversos. La disponibilidad de recursos alimenticios en un ecosistema se ha propuesto como
uno de los factores más importantes en la conformación de las comunidades de aves (Carrascal,
2003; Fretwell, 1972). La disponibilidad de alimento para aves, sitios de percha y distintos tipos
de estratos de forrajeo están estrechamente ligados con la estructura de la vegetación. Las aves
utilizan selectivamente los perfiles del follaje de acuerdo a la época del año, sin embargo, las
diferentes formas presentes en la vegetación incrementan la diversidad de aves (Carrascal, 2004;
Nocedal, 1984). Para el caso de roedores, la vegetación es un factor denso-dependiente, ya que
muchos roedores se distribuyen en sitios donde se encuentra mayor disponibilidad de alimento
(Janzen, 1970), el ratón de campo Psammomys obesus en el desierto de Israel, construye
madrigueras cerca de las riveras densamente cubiertas por los arbustos de las especies Atriplex
halimus y Anabis articulata que le proporcionan refugio y alimento (Tchabovsky y Krasnov,
2002), si la vegetación es perturbada, la dieta de semillas de algunos roedores puede ser
sustituida por otro tipo de dieta (Castellarini et al., 2003). La cubierta vegetal puede considerarse
como uno de los más importantes recursos que determinan el uso de hábitat por roedores (Suárez
y Bonaventura, 2001). La vegetación en pastizales permite que aquellas semillas que logran
“escapar” de la depredación por roedores, o son transportadas por el viento, restablezcan zonas
degradadas o formen un banco de semillas. Para los consumidores de semillas, la disponibilidad
de alimento, así como su comportamiento para disminuir el riesgo a ser depredados por un
carnívoro, puede influir en su distribución y por consiguiente sobre los depredadores de semillas
en sitios con cobertura arbolada. El roedor Gerbillus dasyurus en el desierto de Israel establece
relaciones muy estrechas entre individuos de su misma especie, lo que le permiten aumentar su
sobrevivencia ante depredadores, mientras más organismos se desplacen juntos, mayor confusión
se genera al depredador (Gromov et al., 2001); un roedor que consume semillas bajo el dosel de
un árbol tiene menor riesgo a ser depredado, que uno en un campo abierto donde puede ser presa
fácil de aguilillas, lechuzas, coyotes entre algunos carnívoros (Moore et al., 2003; Manson y
Stiles, 1998; Suárez y Bonaventura, 2001).
La diversidad de aves depredadoras de semillas de Pinus teocote estuvo relacionada con
los tipos y características de los hábitats estudiados (bosque y pastizal). Encontramos especies
exclusivas en cada tipo de hábitat, ya que presentan ciertas características microclimáticas,
edáficas y biológicas particulares, que ciertas especies de aves requieren para completar sus
ciclos biológicos (Carrascal, 2003). En un sitio heterogéneo los hábitats presentes cuentan con
distintos factores bióticos y abióticos que influyen en la presencia física de organismos (Bush et
al., 2001). La diversidad de aves puede incrementar o disminuir de acuerdo con características
especificas de clima, sitios de percha, alimentación, tipo de vegetación presentes en determinada
zona (Blendinger y Ojeda, 2001; Carrascal, 2004).

28
En cambio la diversidad de roedores no dependió de las características de los hábitats
estudiados, las cuatro especies registradas se presentaron en ambos hábitats, aunque si se
registraron dos especies de musarañas exclusivas para el bosque. Los diferentes ambientes que
presenta un sitio, y la fisonomía de la vegetación influyen fuertemente sobre la abundancia y
distribución de animales; los bosques presentan diversos microhábitats con condiciones
particulares de temperatura, iluminación, humedad, etc., creando características ambientales que
favorecen la presencia de ciertas especies (James y Warner, 1982; Rotenberry, 1985; Rotenberry
y Wiens, 1980; Stancampiano y Schnella, 2003), si estos hábitats son dañados, fragmentados o
reducidos, las poblaciones de animales asociados, pueden disminuir drásticamente (Ojeda et al.,
2002).
A diferencia de la diversidad, la abundancia de aves y roedores no se relacionó con las
características de los hábitats estudiados, aunque ambos grupos fueron similarmente abundantes
en el bosque y pastizal, se presentaron especies exclusivas en cada tipo de hábitat, ya que cada
hábitat proporciona alimento, refugio, sitios de percha y madrigueras para diversas especies de
aves y roedores. La diversidad de hábitats proporciona diversidad de fauna (aves y pequeños
mamíferos).
El tipo de vegetación influyó sobre la cantidad de semillas depredadas por aves y roedores.
Las aves depredaron mayor número de semillas en el pastizal; la mayoría de aves registradas en
este hábitat fueron passeriformes granívoros, en el pastizal se presentan mayor número de
especies de gramíneas, muchas de las cuales se establecen después de algún tipo de perturbación,
como el desmonte y los incendios forestales (Ericson et al., 2003; Geréz, 1983), situación que se
ha presentado en el sitio de estudio frecuentemente. Mientras que los roedores, depredaron mayor
cantidad de semillas en el bosque; la presencia de árboles productores de semillas genera una
denso-dependencia hacia el sitio donde se encuentra el alimento, en este caso bajo la copa de los
árboles (Janzen, 1970); En el sudeste de Brasil, por ejemplo, la disponibilidad de alimento y
precipitación fueron dos factores importantes para el establecimiento y crecimiento poblacional
de los roedores Nectomys squamipes y Oryzomys russatus (Bergallo y Magnusson, 2002).
El tipo de vegetación es un factor importante relacionado con la presencia de aves y
roedores depredadores de semillas. El mayor consumo de semillas llevado a cabo por aves en el
pastizal, permite que se genere un mayor banco de semillas en el bosque, que en condiciones
óptimas, pueden germinar hasta establecerse como plántulas. Los roedores, que se alimentan en
mayor parte de semillas depositadas bajo los árboles, permiten que semillas depositadas en el
pastizal formen un mayor banco de semillas mismas que germinarán en condiciones favorables.
El consumo de semillas llevado a cabo por ambos grupos (aves y roedores) regula los bancos de
semillas en los dos tipos de hábitat, lo cual se relaciona con el establecimiento y colonización de
especies vegetales hacia otros sitios, los animales se encuentran estrechamente relacionados con
los procesos de regeneración exitosa de árboles en zonas degradadas (Harold, 2001).

29
Sin embargo, en total el mayor consumo de semillas llevado a cabo por aves y roedores en
los sitios estudiados se registró en el pastizal, esto permite la formación de un mayor banco de
semillas en el bosque, en “espera” de que se presenten las condiciones ideales para su
germinación. Esta situación también puede explicar, en parte, porqué los pastizales en las áreas
de bosque templado se mantienen y no son invadidos por árboles. La depredación de semillas y
plántulas realizada por dos especies de roedores (Peromyscus leucopus y Microtus
pensylvanicus) reguló la invasión de árboles a otros sitios mediante el consumo de las semillas
que formaban el banco de semillas en bosques del Este de Estados Unidos (Gill y Marks, 1991;
Ostfeld y Canham, 1993). En los sitios estudiados, los roedores depredaron menor número de
semillas en el pastizal, las semillas (dispersadas por viento) son menos abundantes en sitios
lejanos a los árboles productores, por lo que a pesar de la menor depredación, ésta puede ser de
mayor impacto sobre la tasa de reclutamiento de especies arbóreas en un pastizal. Si los
depredadores de semillas desaparecieran, la dinámica de los pastizales y bosques sin duda
cambiaría.
Las aves y roedores granívoros presentan patrones selectivos de forrajeo enfocado al
consumo de semillas en cada tipo de hábitat, relacionados con la composición de la vegetación y
el banco de semillas en el suelo del bosque, las diferencias en la composición de especies de las
comunidades de aves y roedores, y el grado de depredación de semillas, están relacionados con el
tipo de hábitat. Cuando la depredación de semillas de árboles y arbustos disminuye, las
probabilidades de regeneración natural se incrementan.
Pinus teocote es la especie dominante en el bosque del área natural protegida de San Juan
del monte ocupando el 68% de cobertura forestal, 412.47 ha de 609 ha que comprenden el área
natural protegida, esta especie ah sido presa de los incendios forestales registrados en la zona, no
obstante muchos árboles han sobrevivido y por medio de sus semillas cuando estas caen al suelo
se está propiciando una regeneración natural de esta especie, a su vez de que dichos árboles son
usados como refugio por una amplia gama de organismos.

30
8. CONCLUSIONES
En el bosque la diversidad de especies de aves fue mayor, además presentó el mayor
número de especies exclusivas.
En el bosque la diversidad de especies de pequeños mamíferos fue mayor, donde se
registraron dos especies exclusivas. En el pastizal no se presentaron especies exclusivas.
Las características de cada hábitat no influyen sobre la abundancia de aves y roedores, el
bosque y el pastizal fueron similares en abundancia, ambos hábitats cuentan con recursos para
brindar alimento y refugio a aves y roedores, sin embargo en el pastizal se depredaron
significativamente más semillas, por aves y roedores, que en el bosque.
La diversidad de aves se relacionó con las características de los hábitats, presentando
especies exclusivas para cada hábitat (bosque y pastizal).
La presencia de Pinus teocote en la mayor parte del territorio es un factor de importancia ya
que brinda de recursos alimenticios a las comunidades de aves y roedores, no obstante las
perturbaciones a las que ah sido objeto la vegetación de pino han favorecido al establecimiento
de gramíneas y con esto a la presencia de organismos granívoros.
Las preferencias de los granívoros por alimentarse de ciertos tipos de semillas es la pauta
para proponer algunas hipótesis sobre las consecuencias del consumo de semillas sobre los
bancos de semillas del suelo, a su vez sobre la composición y abundancia de la vegetación.

31
9. LITERATURA CITADA
ALMEIDA, M.E. 1997. Recomposición de la producción y del mercado de la madera en el Cofre
de Perote. Tésis de Maestría en Desarrollo Rural Regional. Universidad de Chapingo.
Estado de México. 117 p.
ANDERSON, P.K. 1989. Dispersal in rodents: a resident fitness hypothesis. Special Publication.
The American Society of Mammalogist 9:1-139.
ARRIAGA, V.; CERVANTES, V. y VARGAS-MENA, A. 1994. Manual de Reforestación con
Especies Nativas: Colecta y Preservación de Semillas, Propagación y Manejo de Plantas.
SEDESOL / INE – Facultad de Ciencias UNAM. México, D.F. 52 p.
BARLOW, J.C. 1969. Observations on the biology of rodents in Uruguay. Life Scientific 75: 1-
59.
BEGON, M.; HARPER, J.L. y TOWSEND, C.P. 1996. Ecology: Individuals, populations and
communities. Segunda edición. Sinauer Associates. Suderland, Massachusetts. 1068 p.
BERGALLO, H.G. y MAGNUSSON, W.E. 2002. Effects of weather and food availability on the
condition and growth of two species of rodents in Southeastern Brazil. Mammalia 66: 17-
31
BERNINGER, K. y JUNIKKA, L. 1994. Biodiversidad en los Bosques de Coníferas en la Región
del Cofre de Perote, Veracruz. Estudio técnico No 5, Acuerdo de cooperación en materia
forestal México-Finlandia. Helsinki, Finlandia. 32 p.
BERNIS, F. 1966. Migración en aves. Tratado teórico y práctico. SEO. Madrid. 238-242.
BONAVENTURA, S.M.; PIANTANIDA, M.J. y GURINI, L. 1991. Habitat selection in
population of cricetine rodents in the region Delta (Argentina). Mammalia 55: 339-354.
BONAVENTURA, S.M., PIANTANIDA, M.J y GURINI, L. 1992. Selección de hábitat en
roedores cricétidos del Delta Bnaerense. Acta Científica Venezolana 43: 373-380.
BONAVENTURA, S.M. y SUÁREZ, O.V. 1992. The relationship between food availability,
sapace use and territoriality in Akodon azarae. Mammalia 56:407-416.

32
BLAKE, J.G. 1982. Influence of the fire and logging on non-breeding bird communities of
ponderosa pine forest. Ecology 55: 828-837.
BLENDINGER, P.G. y OJEDA, R.A. 2001. Seed supply as a limiting factor for granivorous bird
assemblages in the Monte Desert, Argentina. Austral Ecology 26: 413-422.
BUSTAMANTE, R. y MARONE, L. 2001. Granivoría en ambientes áridos y semiáridos
patrones y procesos a escala local y regional. Memorias del primer encuentro Binacional de
Ecología. Asociación Argentina de Ecología y Sociedad Chilena de Ecología.
BUSH, M.; MIÑO, M.H.; DADON, J.R. y HODARA, K. 2001. Hábitat selection by Akodon
azarae and Calomys laucha (Rodentia, Muridae) in pampean agroecosystems. Mammalia
65:29-48.
CARRASCAL, L.M. 2003. Patrones de preferencias de hábitat, valencia ecológica y densidad de
paseriformes. Atlas de las Aves Reproductoras de España. Dirección General de la
Conservación de la Naturaleza-Sociedad Española de Ornitología, Madrid Pp 718-721.
CARRASCAL, L.M. 2004. Distribución y abundancia de las aves en la Península Ibérica. Una
aproximación biogeográfica y macroecológica. La Ornitología hoy. Homenaje al Profesor
Francisco Bernis. Editorial Complutense. Universidad Complutense de Madrid, Madrid. 48
p.
CASTELLARINI, F.; DELLAFIORE, C. y POLOP, J. 2003. Feeding habits of small mammals
in agroecosystems of Central Argentina. Mammalian Biology 68: 91-101.
CEBALLOS-GONZÁLEZ, G. y GALINDO-LEAL, C. 1984. Mamíferos silvestres de la Cuenca
de México. Editorial Limusa. 292 p.
CODY, M.L. 1968. On the methods of resource division in grassland bird communities.
American Naturalist 102: 107-147.
CONNELL, J.H. y SLATYER, R.O. 1977. Mechanisms of succesión in natural comunities and
their role in community stability and organization. American Naturalist 111:1119-1144.
CUETO, G.R.; BILENCA, D.N. y KRAVETZ, F.O. 1995. Interspecific social relationships in
three murid rodent species of central Argentina, alter fasting and unlimited food. Behaviour
132: 811-820.
CUETO, V.R.; CAGNONI, M.C. y PIANTANIDA, M.J. 1995. Hábitat use of Scapteromys
tumidus (Rodentia:Cricetidae) in the Delta of Paraná River, Argentina. Mammalia 59: 24-
34.

33
CUETO, V.R.; CAGNONI, M.C. y PIANTANIDA, M.J. 1996. Efecto de la cobertura vegetal
sobre el uso de hábitat del ratón de campo Akodon azarae. Acta científica Venezolana 47:
111-116.
CUETO, V.R.; MARONE, L. y LÓPEZ DE CASENAVE, J. 2001. Seed preferences by birds
effects of the design of feeding – preference experiments. Journal of Avian Biology 32:
275-278.
DAVIES, N.B. y LUNDBERG, A. 1984. Food distribution and a variable mating system in the
dunnock Prunella modularis . The Journal of Animal Ecology 53: 895-912.
DAVIDSON, W.D. 1993. The effects of hervibory and granivory on terrestrial plant succession.
Oikos 68: 25-35.
DIRZO, R. 1990. La biodiversidad como crisis ecológica actual ¿Qué sabemos?, Ciencias (No
especial) 4:48:55.
DUNBAR, R.I.M. 1985. Population consequences of social structure Pp 507-519. In Behavioural
Ecology. Ecological consequences of adaptive behaviour (R.M. Sibly and R.H. Smith,
eds).Blackwell Scientific Publications.
ERICSON, P.G.P.; IRESTEDT, M. y JOHANSSON, U.S. 2003. Evolution, biogeography, and
patterns of diversification in passerine birds. Journal of Avian Biology. 34: 3-15.
EXCEL OFFICE XP. 2001. Copyrigth. Microsoft Corporation.United States of America.
FLORES-VILLELA, O. y GERÉZ, P. 1994. Biodiversidad y conservación en México. Segunda
Edición. Universidad Autónoma de México. México, D.F. 439 p.
FRETWELL, S.D. y LUCAS, H.L. 1970. On the territorial behavior and other factors influencing
habitat distribution in birds. I. Theoretical development. Acta Biotheoretica 19: 16-36.
FRETWELL, S.D. 1972. Population in seasonal environment. Monographs in Population
Biology 5. Princeton Univ. Press. Princeton.217 p.
GALINDO-LEAL, C. y KREBS, C.J. 1998. Effects of food abundance on individuals and
populations of the rock mouse (Peromyscus difficilis). Journal of Mammalogy 79:1131-
1142.
GALINDO-GONZÁLEZ, J. (en edición). Efectos de la fragmentación del paisaje sobre las
poblaciones de mamíferos, el caso de los murciélagos de Los Tuxtlas Veracruz. En: G.
Sánchez Rojas y A. E. Rojas Martínez (editores). Tópicos en Sistemática, Biogeografía,
Ecología y Conservación de Mamíferos. Universidad Autónoma del Estado de Hidalgo,
México.

34
GALINDO-GONZÁLEZ, J. 1998. Dispersión de semillas por murciélagos: su importancia en la
conservación y regeneración del bosque tropical. Acta Zoológica Mexicana (nueva serie)
73: 57 – 74.
GALINDO-GONZÁLEZ, J.; GUEVARA, S. y SOSA, V.J. 2000. Bat-and bird-generated seed
rains at isolated trees in pastures in tropical rain forest. Conservation Biology 14: 1693 –
1703.
GEISSERT, F.P.; DUBROEUCK, D.; CAMPOS, A. y MEZA, P.E. 1994. Carta de Paisajes
Geomorfoedafológicos de la Región Volcánica Cofre de Perote Edo de Veracruz, México.
Escala 1:75,000. Instituto de Ecología AC y ORSTOM Xalapa, Veracruz.
GEISSERT, K.D. y MEZA, P.E. 1996. Propiedades físicas e hídricas de los Andosoles del Cofre
de Perote, Veracruz. Instituto de Ecología AC. Xalapa, Veracruz.
GERÉZ, F.P. 1983. Crónica del Uso de los recursos naturales en la cuenca de Perote, Libres en
Goldberg, A.D. (Compilador), El cofre de Perote, investigaciones ecológicas en un área
conflictiva. Cuadernos de divulgación. 9:11-16. Inireb, Xalapa. (Compilador), El Cofre de
Perote, investigaciones ecológicas en un área conflictiva. Cuadernos de divulgación 9:11-
16 INIREB. Xalapa.
GILL, D.S. y MARKS, P.L. 1991. Tree and shurb seedling colonization of old fields in central
New York. Ecology. Monografy 61:183-205.
GOBIERNO DEL ESTADO DE VERACRUZ. y COORDINACIÓN ESTATAL DE MEDIO
AMBIENTE. 2002. Programa de Manejo de San Juan del Monte. Vol.9. Editora de
Gobierno del Estado de Veracruz. 85 p.
GRAVES, S.; MALDONADO, J. y WOLF, J.O. 1988. Use of ground and arboreal microhábitats
by Peromyscus leucopus and Peromyscus maniculatus. Canadian Journal of Zoology
66:277-278.
GREENWOOD, J.S.D. 1985. Frequency dependent selection by seed predators. Oikos 44:195-
210.
GROMOV, V.S.; KRASNOV, B.R. y SHENBROT, G.I. 2001. Behavioural correlates of spatial
distribution in Wagner´s gerbil Gerbillus dasyurus (Rodentia, Gerbillinae). Mammalia 65:
111-120.
HALL, R.E. y DALQUEST, W.W. 1963. The mammals of Veracruz. University of Kansas
Publications. Museum of Natural History 14:165-362.

35
HAROLD, A.C. 2001. Remoción y Germinación de semillas de Dipteryx panamensis y Carapa
guianensis en bosques fragmentados de Sarapiquí, Costa Rica. Revista Forestal
Centroamericana 34: 42-46.
HORNFELDT, B. 1994. Delayed density dependence as a determinant of vole cycles. Ecology
75:791-806.
HOWELL, S. y WEBB, S. 2001. A Guide to the birds of México and Northern Central América.
Published by University Oxford Press. 851 p.
HOWE, H.F. y SMALLWOOD, J. 1982. Ecology of seed dispersal. Annual Review Ecology
Systematics 13:201-228.
HOWE, H.F. 1984. Implications of seed dispersal by animals for tropical reserve management.
Annual Review Ecology Systematics 30:261-281.
INEGI-ORSTOM. 1991. Cuaderno de Información Básica. Región Cofre de Perote. Instituto
Nacional de Estadística, Geografía e Informática. Instituto Francés de Investigación
Científica para el Desarrollo en Cooperación. Aguascalientes,Ags. México. Anexo
Cartográfico 30 mapas. 58 p.
INSTITUTO DE INVESTIGACIONES FORESTALES. y AGROPECUARIAS. 1994. Semillas
Forestales. División Forestal. Publicación Especial No. 2. 137 p.
ISAAC, J.P. y MARTÍNEZ, M.M. 2001. Estacionalidad y relaciones con la estructura del hábitat
de las comunidades de aves de pastizales de paja colorada manejadas con fuego en la
provincia de Buenos Aires, Argentina. Ornitología Neotropical. Departamento de Biología.
Universidad Nacional de Mar de Plata.
JAMES, F.C. y WAMER, N.O. 1982. Relationships between temperate forest bird communities
and vegetation structure. Ecology 63: 159-171.
JANZEN, D.H. 1971. Seed predation by animals. Annual Review Ecology Systematics 2: 465-
492.
JANZEN, D.H. 1970. Herbivores and the number of tree species in tropical forests. The
American Naturalist 104: 501-528.
LABRADOR, M.J. y ALFIERI, M.A. 2001. Agroecología y Desarrollo. Editorial Mundi-Prensa.
566 pps.

36
LANGGUTH, A. 1963. Las especies Uruguayas del género Oryzomyz (Rodentia:Cricetidae).
Community Zoology Museum History Natural Montevideo 7: 1-19.
MAC ARTHUR, R.H. y LEVINS, R. 1964. Competition, habitat selection, and character
displacement in a patchy environment. Proceedings of the National Academy of Sciences,
USA 51: 1207-1210.
MACDONNELL, M.J. y STILES, E.W. 1983. The estructural complexity of old field vegetation
and the recuitment of bird-dispersed plant species. Oecología 56:109-114.
MANSON, H.R. y STILES, E.W. 1998. Links between microhábitat preferences and seed
predation by small mammals in old fields. Oikos 82:37-50.
MANSON, H.R.; OSTFELD, R.S. y CANHAM, C.D. 1998. The effects of tree seed and seedling
density on predation rates by rodents in old fields. Ecoscience 5:183-190.
MARONE, L. 1991. Habitat features affecting bird species distribution in the Monte desert,
Argentina. Ecología Austral. P 23-28.
MILLS, J.N.; ELLIS, B.A.; MCKEE, K.T.; MAIZTEGUI, J.I. y CHILDS, J.E. 1991. Habitat
associations and relative densities of rodent populations in cultivated areas of central
Argentina. Journal of Mammalogy 72: 470-479.
MITTELBACH, G.G. y GROSS, K.L. 1984. Experimental studies of seed predation in old fields.
Oecología 65:7-13.
MOEN, J.; GARDFJELL, H.; OKSANEN, L.; ERICSON, L. y EKERHOLM, P. 1993. Grazing
by food limited microtine rodents on a productive experimental plant community: does the
“green desert” exist?. Oikos 68: 401-413.
MOORE, N.P.; ASKEW, N.L. y BISHOP, J.D. 2003. Small mammals in new farm woodlands.
Mammal Review 33: 101-104.
MORENO, C.E. 2000. Manual de Métodos para medir la Biodiversidad. Universidad
Veracruzana. Xalapa, Veracruz, México. 49 p.
MYSTER, R.W. y PICKETT, S.T.A. 1993. Effects of litter, distance, density, and vegetation
patch type on postdispersal tree and seed predation in old fields. Oikos 66:381-388.
NOCEDAL, J. 1984. Estructura y utilización del follaje de las comunidades de pájaros en
bosques templados del Valle de México. Acta Zoológica Mexicana. (Nueva serie) 6: 1-4.

37
NARAVE-FLORES, H. 1985. “La vegetación del Cofre de Perote, Veracruz, México”. Instituto
Nacional de Investigaciones sobre Recursos Bióticos. Xalapa,Veracruz. Biótica 1:35-64.
OJEDA, R.A.; BORGHI, C.E. y VIRGILIO, G.R. 2002. Diversidad y Conservación de los
mamíferos de Argentina: Un Panorama Sintético. en "Diversidad y Conservación de los
Mamíferos Neotropicales" (Ceballos, G. y J. Simonetti, Eds). CONABIO, Mexico, D.F. Pp
23-63
ORTIZ, S.C. 1994. Introducción a la leyenda del mapa mundial de suelos FAO/UNESCO,
versión 1988. Cuaderno de edafología 20. Instituto de Recursos naturales, Programa de
Edafología, Colegio de Postgraduados en Ciencias Agrícolas, Montecillos, Edo de México.
México. 40 p.
OSTFELD, R.S. 1985. Limiting resources and territoriality in microtine rodents. The American
Naturalist 126: 1-15.
OSTFELD, R.S. y CANHAM, C.D. 1993. Effects of meadow vole population density on tree
seedling survival in old fields. Ecology 74:1792-1801.
OSTFELD, R.S.; MANSON, R.H. y CANHAM, C.D. 1997. Effects of rodents on tree invasion
of old fields. Ecology 78:1531-1542.
PANAYOTOU, T y ASHTON, P.S. 1992. International Non-Timber Forest Product Issues.
Journal of Sustainable Forestry. 13: 5-23.
PATIÑO, F.; DE LA GARZA, P.; VILLA GOMEZ, Y.; TALAVERA, L. y CAMACHO, F.
1983. Guía para la Recolección y Manejo de Semillas de Especies Forestales. Boletín
divulgativo No. 63. INIF, México, D.F. 181 p.
PEACE, T.R. 1962. Pathology of trees and shrubs. Clarendon Press. Oxford. New Jersey. 753p.
PETERSON, R.T. y CHALIF, E.L. 1989. Aves de México, Guía de campo. Editorial Diana,
México D.F. 443 pp.
PICKETT, S.T.A. y CADENASSO, M.L. 1995. Landscape ecology, spatial heterogeneity in
ecological systems. Science 269:331-334.
PULLIAM, H.R. y DANIELSON, B.J. 1991. Sources, Sinks, and hábitat selection. The
American Naturalist 137: 650-666.
RAMÍREZ-ALBORES, J. y RAMÍREZ-CEDILLO, G. 2002. Avifauna de la Región oriente de la
Sierra de Huautla-Morelos, México. Anales del Instituto de Biología, Universidad Nacional
Autónoma de México. Serie Zoológica 73: 91-111.

38
ROLDÁN, M.J.; CAMPOS, G.; DE PABLO, C.L. y DE AGAR, P.M. 2001. Interacciones
ecológicas en el paisaje: una aproximación basada en el estudio de mosaicos. 1ª Jornadas
Ibéricas de Ecología del Paisaje. Bragança (Portugal). P 2-8.
ROTENBERRY, J.T. y WIENS, J.A. 1980. Habitat structure patchiness, and avian communities
in North American Steppe vegetation a multivariate analysis. Ecology 61:1228-1250.
ROTENBERRY, J.T. 1985. The role of hábitat in avian community composition: physiognomy
or floristic. Oecología 67: 213-217.
RUBENSTEIN, D.I. 1982. Individual variation and competition in the everglades pygmy sunfish.
The Journal of Animal Ecology 50: 337-350.
SOBERÓN, J. y LLORENTE, J. 1993. The use of species acumulation functions for the
prediction of species richness. Conservation Biology 7:480-488.
SOTO, E.M. y ANGULO, R. 1990. Estudio climático de la región del Cofre y Valle de Perote.
Instituto de Ecología, A.C. Publicación 28. Xalapa, Veracruz, México. 103 p.
STANCAMPIANO, A.J. y SCHNELLA, D. 2003. Microhabitat affinities of small mammals in
Southwestern Oklahoma. Journal of Mammalogy 85: 948-958.
STATSOFT, INC. 1998. Statistica for Wndows. Tulsa Oklahoma.
SUÁREZ, O.V. y BONAVENTURA, S.M. 2001. Habitat use a diet sympatric species of rodents
of the low Paraná Delta, Argentina. Mammalia 65: 167-176.
TAMARIN, R.H.; REICH, L.M. y MOYER, C.M. 1984. Meadow vole cycles within fences.
Canadian Journal of Zoology 62:1796-1802.
TCHABOVSKY, A.V. y KRASNOV, R. 2002. Spatial distribution of Psammomys obesus
(Rodentia, Gerbillinae) in relation to vegetation in the Negev Desert of Israel. Mammalia
66: 361-368.
TURNER, M.I. 1996. Species loss in fragments of tropical rain forest: a review of the evidence.
Journal Applied Ecology 33:200-209.
VANDER WALL, S.B. 1992. The role of animals in dispersing a “wind-dispersed” pine.
Ecology 73:614.621.

39
WYATT-SMITH, J.; PANTON, W.P. y MITCHELL, B.A. 1963. Manual of malayan silviculture
for island forest (malayan forest records num 23) Yau Seng Press. Kuala Lumpur. “citado
por Matthews, 1989”. 32-46 p.
ZAR, J.H. 1999. Biostatistical Analysis. 4th Edition. Prentice-Hall, Inc., Upper Saddle River, NJ.
931 p.

40

41
10. ANEXOS

42

43

44

45

46

47

48

49

50
ANEXO 2
Especies vegetales del Área Natural Protegida de San Juan del Monte con status de riesgo
NOM 059-SEMARNAT-2001
Familia Genero Especie Estatus de riesgo
Bromeliaceae Tillandsia imperialis A no endémica
Lauraceae Litsea glaucescens P no endémica
Pinaceae Abies hickelii P endémica
Cupressaceae Cupressus lusitanica Pr no endémica
Malvaceae Phymosia rosea Pr no endémica
ANEXO 3
Especies animales del Área Natural Protegida de San Juan del Monte
con status de riesgo NOM 059-SEMARNAT-2001
Aves
Familia Especie Nombre común Estatus
Fringillidae Junco phaeonotus Junco Ojilumbre Pr endémica
Strigidae Glaucidium gnoma Tecolotito serrano Pr endémica
Trochilidae Athis heloisa Colibri enano zumbador A no endémica
Mamíferos
Familia Especie Nombre comun Estatus de riesgo
Soricidae Sorex macrodon Musaraña Pr endémica
Soricidae Sorex saussurei Musaraña Pr endémica
Sciuridae Spermophilus perotensis Ardillón A endémica
Sciuridae Sciurus oculatus Ardilla Pr endémica
Sciuridae Glaucomys volans Ardilla voladora A no endémica
Cricetidae Peromyscus maniculatus Ratón A endémica
Cricetidae Peromyscus boylii Ratón A endémica

51
Anfibios
Familia Especie Nombre comun Estatus de riesgo
Leptodactylidae Eleuterodactylus decoratus Rana Pr endémica
Pletodontidae Thorius sp Ajolote Pr endémica
Pletodontidae Chropterotriton lavae Tlaconete Pr endémica
Pletodontidae Chropterotriton chiropterus Tlaconete Pr endémica
P.- En peligro de extinción PE.- En peligro de extinción
Pr.- Bajo protección especial A.- Amenazada
Reptiles
Familia Especie Nombre comun Estatus de riesgo
Phrynosomatidae Sceloporus grammicus Lagartija verde Pr no endémica
Phrynosomatidae Phrynosoma orbiculare Camaleón cornudo A endémica
Anguidae Abronia graminea Escorpion Pr endémica
Anguidae Barisia imbricata Escorpion Pr endémica
Colubridae Lampropeltis triangulum Falso coralillo A no endémica
Colubridae Trimorphodon bisculatus Pr no endémica
Dipsadidae Geophis multitorques Pr endémica
Dipsadidae Rhadinea forbesi Pr endémica
Dipsadidae Thamnophis chrysocephalus Culebra de agua A endémica
Dipsadidae Thamnophis cyrtopsis Culebra de agua A no endémica
Dipsadidae Thamnophis eques Culebra de agua A no endémica
Dipsadidae Thamnophis proximus Culebra de agua A no endémica
Dipsadidae Thamnophis scalaris Culebra de agua A endémica
Dipsadidae Thamnophis sumichrasti Culebra de agua A endémica
Crotalidae Crotalus intermedius Vibora de cascabel A endémica
Crotalidae Crotalus scutulatus Vibora de cascabel Pr no endémica
Crotalidae Sistrurus ravus Cascabel sorda Pr endémica

52





























