Embed Size (px)
Citation preview

Vol. 33, No. 11
Quinolones and Coumarins Eliminate Chloroplasts fromEuglena gracilis
JURAJ KRAJXOVII,* LIBOR EBRINGER, AND JOZEF POLONYIInstitute of Molecular and Subcellular Biology, Comenius University, 811 07 Bratislava, Czechoslovakia
Received 24 April 1989/Accepted 16 August 1989
Quinolones and coumarins were potent eliminators of chloroplasts from Euglena graciis. There was aremarkable similarity between antichloroplastic and antibacterial activities of DNA gyrase inhibitors.Quinolones produced 100% chloroplast-free cells in concentrations which do not affect cell viability. Optimalconditions were exponential growth, continuous illumination, and neutral or slightly alkaline pH. Coumarinswere more toxic than quinolones. Among the quinolones, ofloxacin was the most potent in eliminatingchloroplasts. Among the coumarins, coumermycin A1 was the most potent. New quinolones and coumermycinA1 were able to induce the complete inability of originally green cells to form green colonies after 24 h of drugexposure, while clorobiocin and novobiocin required several days of exposure. Darkness, heat shock (42°C, 10min), or simultaneous treatment with chloramphenicol or rifampin decreased the potency of DNA gyraseinhibitors for producing chloroplast-free cells. Remarkably, in cells in which division was blocked by threedifferent methods (resting medium, hyperthermic conditions [3rC], or addition of cyclohexinmide), new
quinolones and coumermycin A1 nevertheless eliminated chloroplasts. The antichloroplastic activity of DNAgyrase inhibitors is additional data suggesting an evolutionary relationship between chloroplasts and eubacteria.
Antibacterial agents affect mitochondria and chloroplastsof eucaryotic cells. These organelles are semiautonomous(with their own DNA and protein synthesis apparatus) andappear to be of eubacterial origin (9, 10, 13, 21). Wepreviously demonstrated that chloroplasts of the flagellateEuglena gracilis are susceptible to antibacterial agents andother biologically active compounds (3, 6-10). In contrast tothe higher plants, chloroplasts in E. gracilis are not essential,thus placing this unicellular eucaryotic microorganism on theboundary between the plant and animal kingdoms. Theantichloroplastic activities of various drugs on E. gracilis aremacroscopically manifested by the permanent loss of theability to form green colonies, with cells changing fromautotrophy to heterotrophy and becoming white (bleaching).As determined by electron microscopic evaluation, bleach-ing is associated with degradation and loss of chloroplasts (3,24; J. Polonyi, L. Ebringer, J. Krajcovic, and K. Kapeller,Z. Mikroskop.-Anat. Forsch., in press).
Quinolones and coumarins are antibacterial agents thatbelong to the group of inhibitors of procaryotic topoisomer-ase II (DNA gyrase). Quinolones are a rapidly expandingclass of agents with promising properties for the treatment ofinfectious diseases (15, 16, 22, 28, 30). Nalidixic acid belongsto the group of subunit gyrA DNA gyrase antagonists(quinolones), and its effect on Euglena chloroplasts has beenextensively evaluated (5, 14, 19, 20, 23). Exposure to nali-dixic acid causes bleaching and loss of chloroplast DNA atconcentrations of drug which do not affect the viability ofcells. From the group of inhibitors of subunit gyrB DNAgyrase (coumarins), novobiocin has also been found to be a
bleaching agent (6).We now present studies evaluating the effect of additional,
more potent antagonists ofDNA gyrase on chloroplasts of E.gracilis under various experimental conditions.
* Corresponding author.
MATERIALS AND METHODS
Organism and cultivation conditions. E. gracilis (Klebs) Z(Pringsheim) was originally obtained from S. H. Hutner,Haskins Laboratories, Pace University, New York, N.Y.Cells were cultivated statically to about 106/ml in 500-mlErlenmeyer flasks containing 150 ml of a modified Cramerand Myers (CM) medium (4) supplemented with ethanol(0.5%) and adjusted to pH 7.0 (unless otherwise specified).Euglena cells were incubated at 27°C with continuous illu-mination (2,200 lx). Cells were harvested by filtration andsuspended either in growth CM medium supplemented withethanol (0.2%) or in resting medium, i.e., buffering compo-nents of CM medium (KH2PO4 and sodium citrate). The celldensity was adjusted to 105/ml with appropriate medium. Inthe case of resting conditions, cells were preincubated inresting medium for 4 days as described above.
Antibacterial agents. Fresh solutions of the following sub-stances were used: ofloxacin (Hoechst Ltd., Hounslow,United Kingdom), ciprofloxacin (Bayer Ltd., Newbury,United Kingdom), enoxacin (Parke-Davis/Warner-LambertLtd., Eastleigh, United Kingdom), norfloxacin (Krka, NovoMesto, Yugoslavia), cinoxacin (Eli Lilly & Co., Indianapo-lis, Ind.), and clorobiocin (May and Baker Ltd., Dagenham,United Kingdom). These were generous gifts from the man-ufacturers. The other agents were purchased: oxolinic acidand coumermycin A1 (Sigma Chemical Co., St. Louis, Mo.),nalidixic acid and novobiocin (Serva Feinbiochemica GmbHand Co., Heidelberg, Federal Republic of Germany), cyclo-heximide (Fluka AG, Buchs, Switzerland), rifampin (LepetitS.p.A., Milan, Italy), and chloramphenicol (Spofa, Prague,Czechoslovakia).Treatment regimens. Drug susceptibility assays were per-
formed in 10-ml test tubes containing 5 ml of E. gracilisculture. The pH values of both types of medium were 7.0. Insome experiments the broad pH scale (3.0 to 9.0) was used,adjusted with 10% H2SO4 or 1 N KOH. All the quinoloneswere dissolved in 0.1 N KOH and diluted in appropriate test
1883
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Nov. 1989, p. 1883-18890066-4804/89/111883-07$02.00/0Copyright © 1989, American Society for Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 02
Feb
ruar
y 20
22 b
y 22
1.12
7.18
.159
.
ANTIMICROB. AGENTS CHEMOTHER.
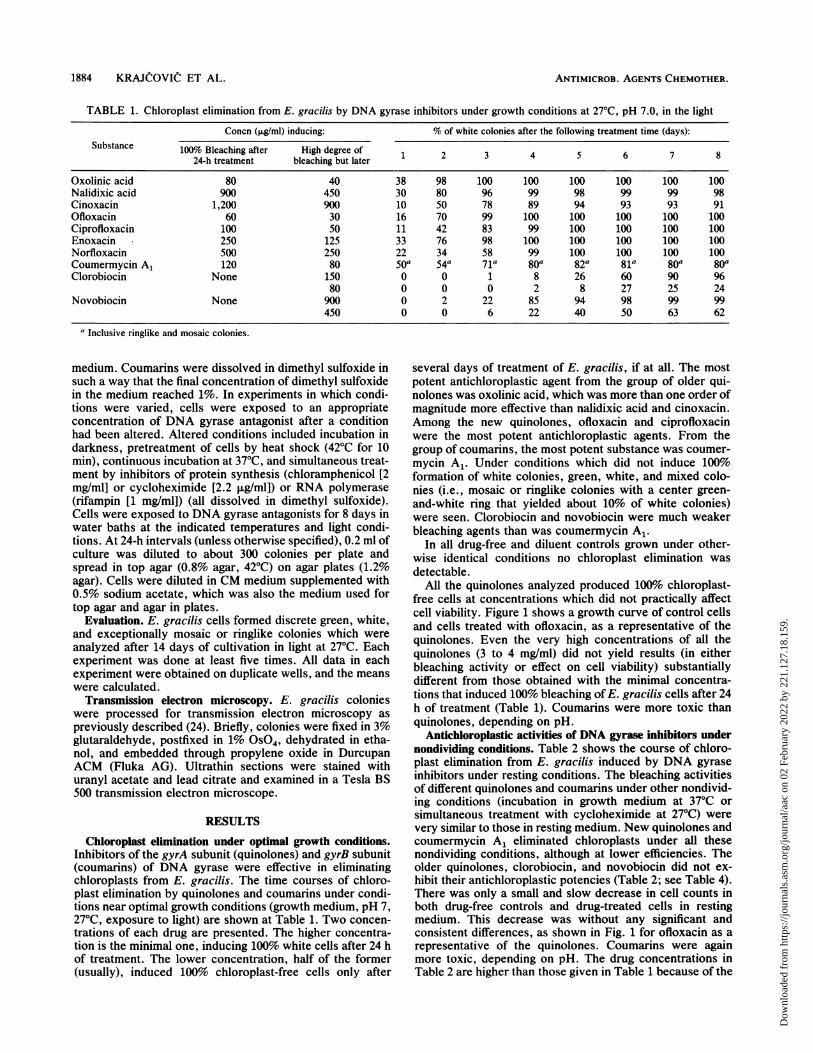
TABLE 1. Chloroplast elimination from E. gracilis by DNA gyrase inhibitors under growth conditions at 27°C, pH 7.0, in the light
Concn (,ug/ml) inducing: % of white colonies after the following treatment time (days):Substance 100% Bleaching after High degree of
24-h treatment bleaching but later 1 2 3 4 5 6 7 8
Oxolinic acid 80 40 38 98 100 100 100 100 100 100Nalidixic acid 900 450 30 80 96 99 98 99 99 98Cinoxacin 1,200 900 10 50 78 89 94 93 93 91Ofloxacin 60 30 16 70 99 100 100 100 100 100Ciprofloxacin 100 50 11 42 83 99 100 100 100 100Enoxacin 250 125 33 76 98 100 100 100 100 100Norfloxacin 500 250 22 34 58 99 100 100 100 100Coumermycin A1 120 80 50a 54a 71a 80a 82a 81a 80a 80aClorobiocin None 150 0 0 1 8 26 60 90 96
80 0 0 0 2 8 27 25 24Novobiocin None 900 0 2 22 85 94 98 99 99
450 0 0 6 22 40 50 63 62
aInclusive ringlike and mosaic colonies.
medium. Coumarins were dissolved in dimethyl sulfoxide insuch a way that the final concentration of dimethyl sulfoxidein the medium reached 1%. In experiments in which condi-tions were varied, cells were exposed to an appropriateconcentration of DNA gyrase antagonist after a conditionhad been altered. Altered conditions included incubation indarkness, pretreatment of cells by heat shock (42°C for 10min), continuous incubation at 37°C, and simultaneous treat-ment by inhibitors of protein synthesis (chloramphenicol [2mg/ml] or cycloheximide [2.2 ,ug/ml]) or RNA polymerase(rifampin [1 mg/ml]) (all dissolved in dimethyl sulfoxide).Cells were exposed to DNA gyrase antagonists for 8 days inwater baths at the indicated temperatures and light condi-tions. At 24-h intervals (unless otherwise specified), 0.2 ml ofculture was diluted to about 300 colonies per plate andspread in top agar (0.8% agar, 42°C) on agar plates (1.2%agar). Cells were diluted in CM medium supplemented with0.5% sodium acetate, which was also the medium used fortop agar and agar in plates.
Evaluation. E. gracilis cells formed discrete green, white,and exceptionally mosaic or ringlike colonies which wereanalyzed after 14 days of cultivation in light at 27°C. Eachexperiment was done at least five times. All data in eachexperiment were obtained on duplicate wells, and the meanswere calculated.
Transmission electron microscopy. E. gracilis colonieswere processed for transmission electron microscopy aspreviously described (24). Briefly, colonies were fixed in 3%glutaraldehyde, postfixed in 1% OS04, dehydrated in etha-nol, and embedded through propylene oxide in DurcupanACM (Fluka AG). Ultrathin sections were stained withuranyl acetate and lead citrate and examined in a Tesla BS500 transmission electron microscope.
RESULTS
Chloroplast elimination under optimal growth conditions.Inhibitors of the gyrA subunit (quinolones) and gyrB subunit(coumarins) of DNA gyrase were effective in eliminatingchloroplasts from E. gracilis. The time courses of chloro-plast elimination by quinolones and coumarins under condi-tions near optimal growth conditions (growth medium, pH 7,27°C, exposure to light) are shown at Table 1. Two concen-trations of each drug are presented. The higher concentra-tion is the minimal one, inducing 100% white cells after 24 hof treatment. The lower concentration, half of the former(usually), induced 100% chloroplast-free cells only after
several days of treatment of E. gracilis, if at all. The mostpotent antichloroplastic agent from the group of older qui-nolones was oxolinic acid, which was more than one order ofmagnitude more effective than nalidixic acid and cinoxacin.Among the new quinolones, ofloxacin and ciprofloxacinwere the most potent antichloroplastic agents. From thegroup of coumarins, the most potent substance was coumer-mycin A1. Under conditions which did not induce 100%formation of white colonies, green, white, and mixed colo-nies (i.e., mosaic or ringlike colonies with a center green-and-white ring that yielded about 10% of white colonies)were seen. Clorobiocin and novobiocin were much weakerbleaching agents than was coumermycin A1.
In all drug-free and diluent controls grown under other-wise identical conditions no chloroplast elimination wasdetectable.
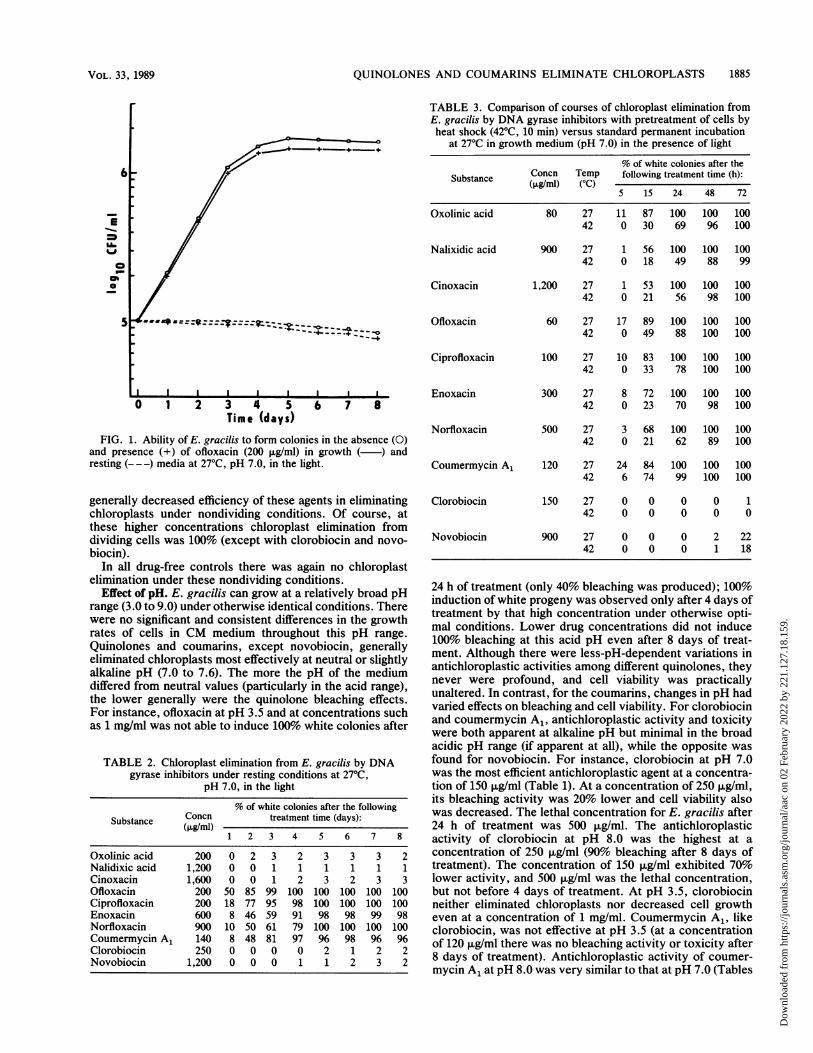
All the quinolones analyzed produced 100%o chloroplast-free cells at concentrations which did not practically affectcell viability. Figure 1 shows a growth curve of control cellsand cells treated with ofloxacin, as a representative of thequinolones. Even the very high concentrations of all thequinolones (3 to 4 mg/ml) did not yield results (in eitherbleaching activity or effect on cell viability) substantiallydifferent from those obtained with the minimal concentra-tions that induced 100% bleaching of E. gracilis cells after 24h of treatment (Table 1). Coumarins were more toxic thanquinolones, depending on pH.
Antichloroplastic activities of DNA gyrase inhibitors undernondividing conditions. Table 2 shows the course of chloro-plast elimination from E. gracilis induced by DNA gyraseinhibitors under resting conditions. The bleaching activitiesof different quinolones and coumarins under other nondivid-ing conditions (incubation in growth medium at 37°C orsimultaneous treatment with cycloheximide at 27°C) werevery similar to those in resting medium. New quinolones andcoumermycin A1 eliminated chloroplasts under all thesenondividing conditions, although at lower efficiencies. Theolder quinolones, clorobiocin, and novobiocin did not ex-hibit their antichloroplastic potencies (Table 2; see Table 4).There was only a small and slow decrease in cell counts inboth drug-free controls and drug-treated cells in restingmedium. This decrease was without any significant andconsistent differences, as shown in Fig. 1 for ofloxacin as arepresentative of the quinolones. Coumarins were againmore toxic, depending on pH. The drug concentrations inTable 2 are higher than those given in Table 1 because of the
1884 KRAJCOVIC ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 02
Feb
ruar
y 20
22 b
y 22
1.12
7.18
.159
.
QUINOLONES AND COUMARINS ELIMINATE CHLOROPLASTS 1885
=
Ch0
0 1 2 3 4 5 6 7 8Time (days)
FIG. 1. Ability of E. gracilis to form colonies in the absence (O)and presence (+) of ofloxacin (200 ,ug/ml) in growth (-) andresting (- --) media at 27°C, pH 7.0, in the light.
generally decreased efficiency of these agents in eliminatingchloroplasts under nondividing conditions. Of course, atthese higher concentrations chloroplast elimination fromdividing cells was 100% (except with clorobiocin and novo-biocin).
In all drug-free controls there was again no chloroplastelimination under these nondividing conditions.
Effect of pH. E. gracilis can grow at a relatively broad pHrange (3.0 to 9.0) under otherwise identical conditions. Therewere no significant and consistent differences in the growthrates of cells in CM medium throughout this pH range.Quinolones and coumarins, except novobiocin, generallyeliminated chloroplasts most effectively at neutral or slightlyalkaline pH (7.0 to 7.6). The more the pH of the mediumdiffered from neutral values (particularly in the acid range),the lower generally were the quinolone bleaching effects.For instance, ofloxacin at pH 3.5 and at concentrations suchas 1 mg/ml was not able to induce 100% white colonies after
TABLE 2. Chloroplast elimination from E. gracilis by DNAgyrase inhibitors under resting conditions at 27°C,
pH 7.0, in the light
% of white colonies after the followingSubstance Concn treatment time (days):
(ILg/ml)1 2 3 4 5 6 7 8
Oxolinic acid 200 0 2 3 2 3 3 3 2Nalidixic acid 1,200 0 0 1 1 1 1 1 1Cinoxacin 1,600 0 0 1 2 3 2 3 3Ofloxacin 200 50 85 99 100 100 100 100 100Ciprofloxacin 200 18 77 95 98 100 100 100 100Enoxacin 600 8 46 59 91 98 98 99 98Norfloxacin 900 10 50 61 79 100 100 100 100Coumermycin A1 140 8 48 81 97 96 98 96 96Clorobiocin 250 0 0 0 0 2 1 2 2Novobiocin 1,200 0 0 0 1 1 2 3 2
TABLE 3. Comparison of courses of chloroplast elimination fromE. gracilis by DNA gyrase inhibitors with pretreatment of cells byheat shock (42°C, 10 min) versus standard permanent incubation
at 27°C in growth medium (pH 7.0) in the presence of light
% of white colonies after theSubstance Concn Temp following treatment time (h):
5 15 24 48 72
Oxolinic acid 80 27 11 87 100 100 10042 0 30 69 96 100
Nalixidic acid 900 27 1 56 100 100 10042 0 18 49 88 99
Cinoxacin 1,200 27 1 53 100 100 10042 0 21 56 98 100
Ofloxacin 60 27 17 89 100 100 10042 0 49 88 100 100
Ciprofloxacin 100 27 10 83 100 100 10042 0 33 78 100 100
Enoxacin 300 27 8 72 100 100 10042 0 23 70 98 100
Norfloxacin 500 27 3 68 100 100 10042 0 21 62 89 100
Coumermycin A1 120 27 24 84 100 100 10042 6 74 99 100 100
Clorobiocin 150 27 0 0 0 0 142 0 0 0 0 0
Novobiocin 900 27 0 0 0 2 2242 0 0 0 1 18
24 h of treatment (only 40% bleaching was produced); 100%induction of white progeny was observed only after 4 days oftreatment by that high concentration under otherwise opti-mal conditions. Lower drug concentrations did not induce100% bleaching at this acid pH even after 8 days of treat-ment. Although there were less-pH-dependent variations inantichloroplastic activities among different quinolones, theynever were profound, and cell viability was practicallyunaltered. In contrast, for the coumarins, changes in pH hadvaried effects on bleaching and cell viability. For clorobiocinand coumermycin A1, antichloroplastic activity and toxicitywere both apparent at alkaline pH but minimal in the broadacidic pH range (if apparent at all), while the opposite wasfound for novobiocin. For instance, clorobiocin at pH 7.0was the most efficient antichloroplastic agent at a concentra-tion of 150 ,ug/ml (Table 1). At a concentration of 250 ,ug/ml,its bleaching activity was 20% lower and cell viability alsowas decreased. The lethal concentration for E. gracilis after24 h of treatment was 500 jig/ml. The antichloroplasticactivity of clorobiocin at pH 8.0 was the highest at aconcentration of 250 ,ug/ml (90% bleaching after 8 days oftreatment). The concentration of 150 ,g/ml exhibited 70%lower activity, and 500 jig/ml was the lethal concentration,but not before 4 days of treatment. At pH 3.5, clorobiocinneither eliminated chloroplasts nor decreased cell growtheven at a concentration of 1 mg/ml. Coumermycin A1, likeclorobiocin, was not effective at pH 3.5 (at a concentrationof 120 ,ug/ml there was no bleaching activity or toxicity after8 days of treatment). Antichloroplastic activity of coumer-mycin A1 at pH 8.0 was very similar to that at pH 7.0 (Tables
VOL. 33, 1989
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 02
Feb
ruar
y 20
22 b
y 22
1.12
7.18
.159
.
ANTIMICROB. AGENTS CHEMOTHER.
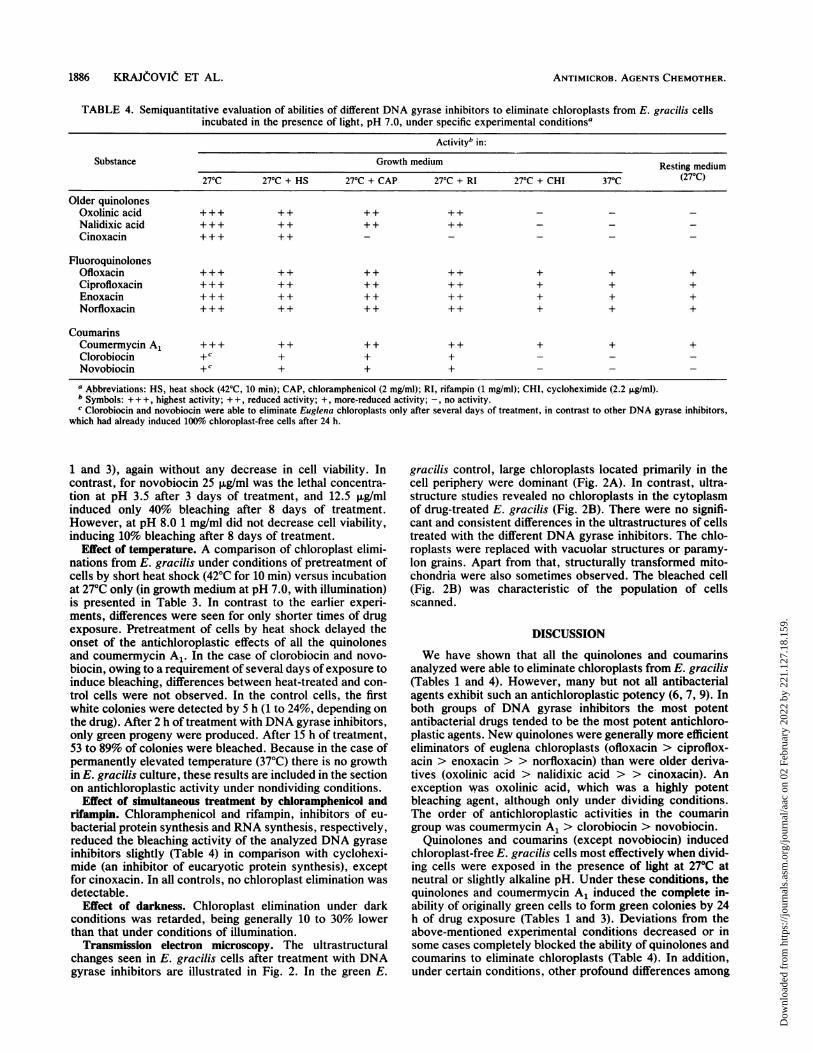
TABLE 4. Semiquantitative evaluation of abilities of different DNA gyrase inhibitors to eliminate chloroplasts from E. gracilis cellsincubated in the presence of light, pH 7.0, under specific experimental conditionsa
Activityb in:
Substance Growth medium Resting medium
270C 270C + HS 27°C + CAP 27°C + RI 27°C + CHI 370C (270C)Older quinolones
Oxolinic acid +++ + + + + ++ - - -Nalidixic acid +++ + + + + + +Cinoxacin +++ ++ - - - - -
FluoroquinolonesOfloxacin + + + + + + + + + + +Ciprofloxacin + + + + + + + + + + +Enoxacin + + + + + + + + + + +Norfloxacin + + + + + + + + + + +
CoumarinsCoumermycin A1 + ++ ++ + + +Clorobiocin +C + + +Novobiocin +C + + + -
a Abbreviations: HS, heat shock (42'C, 10 min); CAP, chloramphenicol (2 mg/ml); RI, rifampin (1 mg/ml); CHI, cycloheximide (2.2 FLg/ml).b Symbols: +++, highest activity; + +, reduced activity; +, more-reduced activity; -, no activity.c Clorobiocin and novobiocin were able to eliminate Euglena chloroplasts only after several days of treatment, in contrast to other DNA gyrase inhibitors,
which had already induced 100%o chloroplast-free cells after 24 h.
1 and 3), again without any decrease in cell viability. Incontrast, for novobiocin 25 ,ug/ml was the lethal concentra-tion at pH 3.5 after 3 days of treatment, and 12.5 ,ug/mlinduced only 40% bleaching after 8 days of treatment.However, at pH 8.0 1 mg/ml did not decrease cell viability,inducing 10% bleaching after 8 days of treatment.
Effect of temperature. A comparison of chloroplast elimi-nations from E. gracilis under conditions of pretreatment ofcells by short heat shock (42°C for 10 min) versus incubationat 27°C only (in growth medium at pH 7.0, with illumination)is presented in Table 3. In contrast to the earlier experi-ments, differences were seen for only shorter times of drugexposure. Pretreatment of cells by heat shock delayed theonset of the antichloroplastic effects of all the quinolonesand coumermycin A1. In the case of clorobiocin and novo-biocin, owing to a requirement of several days of exposure toinduce bleaching, differences between heat-treated and con-trol cells were not observed. In the control cells, the firstwhite colonies were detected by 5 h (1 to 24%, depending onthe drug). After 2 h of treatment with DNA gyrase inhibitors,only green progeny were produced. After 15 h of treatment,53 to 89% of colonies were bleached. Because in the case ofpermanently elevated temperature (37°C) there is no growthin E. gracilis culture, these results are included in the sectionon antichloroplastic activity under nondividing conditions.
Effect of simultaneous treatment by chloramphenicol andrifampin. Chloramphenicol and rifampin, inhibitors of eu-bacterial protein synthesis and RNA synthesis, respectively,reduced the bleaching activity of the analyzed DNA gyraseinhibitors slightly (Table 4) in comparison with cyclohexi-mide (an inhibitor of eucaryotic protein synthesis), exceptfor cinoxacin. In all controls, no chloroplast elimination wasdetectable.
Effect of darkness. Chloroplast elimination under darkconditions was retarded, being generally 10 to 30% lowerthan that under conditions of illumination.
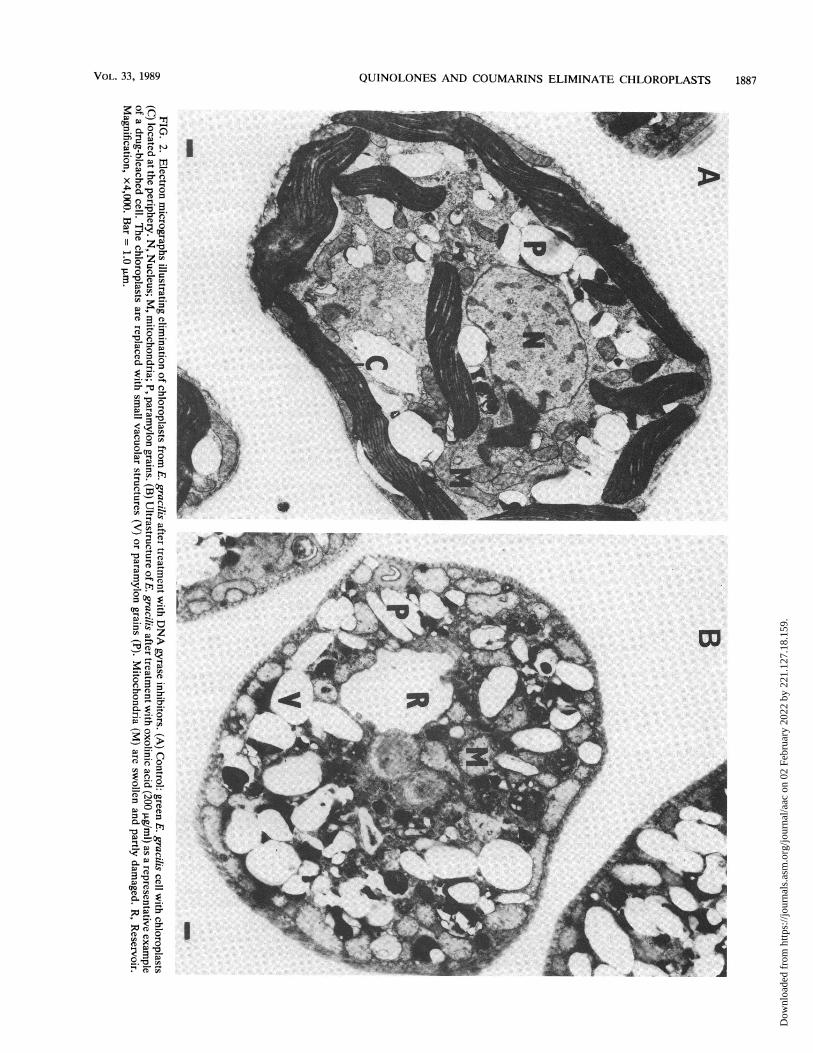
Transmission electron microscopy. The ultrastructuralchanges seen in E. gracilis cells after treatment with DNAgyrase inhibitors are illustrated in Fig. 2. In the green E.
gracilis control, large chloroplasts located primarily in thecell periphery were dominant (Fig. 2A). In contrast, ultra-structure studies revealed no chloroplasts in the cytoplasmof drug-treated E. gracilis (Fig. 2B). There were no signifi-cant and consistent differences in the ultrastructures of cellstreated with the different DNA gyrase inhibitors. The chlo-roplasts were replaced with vacuolar structures or paramy-lon grains. Apart from that, structurally transformed mito-chondria were also sometimes observed. The bleached cell(Fig. 2B) was characteristic of the population of cellsscanned.
DISCUSSION
We have shown that all the quinolones and coumarinsanalyzed were able to eliminate chloroplasts from E. gracilis(Tables 1 and 4). However, many but not all antibacterialagents exhibit such an antichloroplastic potency (6, 7, 9). Inboth groups of DNA gyrase inhibitors the most potentantibacterial drugs tended to be the most potent antichloro-plastic agents. New quinolones were generally more efficienteliminators of euglena chloroplasts (ofloxacin > ciproflox-acin > enoxacin > > norfloxacin) than were older deriva-tives (oxolinic acid > nalidixic acid > > cinoxacin). Anexception was oxolinic acid, which was a highly potentbleaching agent, although only under dividing conditions.The order of antichloroplastic activities in the coumaringroup was coumermycin A1 > clorobiocin > novobiocin.
Quinolones and coumarins (except novobiocin) inducedchloroplast-free E. gracilis cells most effectively when divid-ing cells were exposed in the presence of light at 27°C atneutral or slightly alkaline pH. Under these conditions, thequinolones and coumermycin A1 induced the complete in-ability of originally green cells to form green colonies by 24h of drug exposure (Tables 1 and 3). Deviations from theabove-mentioned experimental conditions decreased or insome cases completely blocked the ability of quinolones andcoumarins to eliminate chloroplasts (Table 4). In addition,under certain conditions, other profound differences among
1886 KRAJtOVIIC ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 02
Feb
ruar
y 20
22 b
y 22
1.12
7.18
.159
.
QUINOLONES AND COUMARINS ELIMINATE CHLOROPLASTS 1887
0-
0-t
z_
l
OX CD '4
o o z _
0e0 P00s
CD
o
_.0 0
Pz O
_. 4.
CD
-t5o
CD CD$
0~-
CD
0 0
0
0. k
0 -t0
CD
0B-.
0)
CD5
<0Q
CD°
CD
O CL _
CD (A C
CD 0_
0 _
I
VOL. 33, 1989
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 02
Feb
ruar
y 20
22 b
y 22
1.12
7.18
.159
.

ANTIMICROB. AGENTS CHEMOTHER.
DNA gyrase inhibitors were observed. Under nondividingconditions induced by resting medium (Table 2), hyperther-mic conditions (37°C), or cycloheximide (Table 4), only newquinolones and coumermycin A1 exhibited antichloroplasticactivity, although this activity was somewhat less than withgrowing cells. That these agents are able to eliminate chlo-roplasts from both growing and nongrowing cells suggeststhe presence of more than one mechanism of chloroplastantagonism, as has been suggested for killing of bacteria byciprofloxacin and ofloxacin (16, 26, 28, 31) and even norflox-acin (27) based on similar types of studies. These findings arealso remarkable in that previously evaluated antibioticseliminated E. gracilis chloroplasts only from growing cells(6-9). In contrast, mutagens and carcinogens eliminatedchloroplasts from either growing or nongrowing cells (3, 8,9). We recently identified differences between DNA gyraseinhibitors and classical mutagens in their action on E.gracilis chloroplasts. Mutagens eliminate Euglena chloro-plasts more rapidly (in minutes to hours) and only at near-toxic concentrations. Nonmutagenic antibacterial agents re-quire longer periods (1 to 4 days) for induction of 100%bleaching; however, they exhibit this antichloroplastic activ-ity in concentrations which do not affect cell viability. Inaddition, heat shock and hyperthermia stimulate the bleach-ing activity of some mutagens (3) while antagonizing thiseffect of quinolones and coumermycin A1 (Tables 3 and 4).Exposure to coumermycin A1 results in growth of not only
green and white colonies but also mosaic or ringlike colo-nies. Such mixed colonies are only sometimes induced byexposure to other DNA gyrase antagonists but are com-monly seen after exposure to mutagens and carcinogens (3).From an evolutionary viewpoint, the correlation between
antibacterial and antichloroplastic activities of DNA gyraseantagonists represents additional data suggestive of a closerelationship between eubacteria and chloroplasts (9, 10, 13,21). In agreement with this possibility are the observationsthat inhibitors of eucaryotic topoisomerase II (etoposide,teniposide, adriamycin, and mitoxantrone) do not eliminatechloroplasts from E. gracilis (data not shown).The putative targets for DNA gyrase antagonists in chlo-
roplasts are likely similar to those found in eubacteria. DNAgyrase activity was recently also detected in chloroplasts(18, 29). The apparent molecular weights and dual bands ofthe putative chloroplast topoisomerase II suggest that theprotein may be similar to a bacterial gyrase type II enzymewith two subunits (25). The amount of enzyme per chloro-plast is highest at the time when chloroplast replication ismaximal.The facultative nature of chloroplasts in E. gracilis cells
makes this flagellate an object for the study of antibacterialagents in vivo. It appears to be suggestive for an evaluationof their antibacterial potencies in vivo (2, 11). In this contextthe close resemblance of the orders of antichloroplasticactivities of quinolones and their activities against Chla-mydia trachomatis (17) are also very interesting becausethese pathogens are obligate intracellular parasites and chlo-roplasts are supposed to be eubacterial endosymbionts ofplant cells. On the other hand, the study of the action ofantibacterial chemotherapeutic agents on E. gracilis couldhelp identify some side or latent effects of analyzed sub-stances on the eucaryotic cell itself. DNA gyrase inhibitorsmay also antagonize eucaryotic cells, including human cells(12), possibly by inhibition of a DNA gyrase-like enzyme inmitochondria (1). Recently, we demonstrated that quinolo-nes and coumermycin A1 also affect the mitochondria of E.gracilis (Pol6nyi et al., in press).
In conclusion, the quinolones and coumarins are potenteliminators of chloroplasts from E. gracilis. The study oftheir antichloroplastic activities under a broad spectrum ofexperimental conditions has revealed several remarkabledifferences among DNA gyrase inhibitors. We point outagain that the flagellate E. gracilis, with its facultativechloroplasts, could be a sensitive model for an evaluation ofthe activities of various antibacterial agents.
ACKNOWLEDGMENTS
We thank John S. Wolfson for critical reading of the manuscript.The skillful technical assistance of M. Barnasova is gratefullyacknowledged. We express our gratitude for the donations ofofloxacin, ciprofloxacin, enoxacin, norfloxacin, cinoxacin, and clo-robiocin to Hoechst Ltd., Bayer Ltd., Parke-Davis/Warner-Lam-bert Ltd., Krka, Eli Lilly & Co., and May and Baker Ltd.,respectively.
LITERATURE CITED1. Castora, F. J., M. Lazarus, and D. Kunes. 1985. The presence of
two mitochondrial DNA topoisomerases in human acute leuke-mia cells. Biochem. Biophys. Res. Commun. 130:854-866.
2. Celmer, W. D., and L. Ebringer. 1%7. Effects of certain 0-acylderivatives of oleandomycin and erythromycin on chloroplastsof Euglena gracilis. J. Protozool. 14:263-267.
3. ChreAo, O., J. Krajkovil, L. Ebringer, and J. Pol6nyi. 1988.Effect of heat shock on the mutagenicity of mutagens andcarcinogens in Euglena gracilis. Teratog. Carcinog. Mutagen.8:161-168.
4. Cramer, M., and J. Myers. 1952. Growth and photosyntheticcharacteristic of Euglena gracilis. Arch. Microbiol. 17:384-402.
5. Ebringer, L. 1970. The action of nalidixic acid on Euglenaplastids. J. Gen. Microbiol. 61:141-144.
6. Ebringer, L. 1972. Are plastids derived from prokaryotic micro-organisms? Action of antibiotics on chloroplasts of Euglenagracilis. J. Gen. Microbiol. 71:35-52.
7. Ebringer, L. 1978. Effects of drugs on chloroplasts. Prog. Mol.Subcell. Biol. 6:271-354.
8. Ebringer, L., A. Jurfiek, J. Konitek, M. Koniakov*, N. La-hitova, and S. Trubakik. 1976. Mutagenic action of nitrofuranson Euglena gracilis and Mycobacterium phlei. Antimicrob.Agents Chemother. 9:682-689.
9. Ebringer, L., and J. KrajEovik. 1986. Are chloroplasts andmitochondria the remnants of prokaryotic endosymbionts? Fo-ha Microbiol. 31:228-254.
10. Ebringer, L., and J. Krajkovik. 1987. Prokaryotic character ofchloroplasts and mitochondria-the present knowledge. FoliaMicrobiol. 32:244-282.
11. Fasulo, M., M. Bassi, and G. Dafl'Olio. 1981. Effects of eryth-romycin on Euglena gracilis as a model for testing the toxicityof antibacterial drugs. Drug Res. 31:1869-1872.
12. Forsgren, A., S. F. Schlossman, and T. F. Tedder. 1987. 4-Quinolone drugs affect cell cycle progression and function ofhuman lymphocytes in vitro. Antimicrob. Agents Chemother.31:768-773.
13. Gray, M. W. 1988. Organelle origins and ribosomal RNA.Biochem. Cell Biol. 66:325-348.
14. Hashimoto, H., and Murakami. 1982. Chloroplast replicationand loss of chloroplast DNA induced by nalidixic acid inEuglena gracilis. Cell Struct. Funct. 7:111-120.
15. Hooper, D. C., and J. S. Wolfson. 1985. The fluoroquinolones:pharmacology, clinical uses, and toxicities in humans. Antimi-crob. Agents Chemother. 28:716-721.
16. Hooper, D. C., J. S. Wolfson, E. Y. Ng, and M. N. Swartz. 1987.Mechanisms of action of and resistance to ciprofloxacin. Am. J.Med. 82(Suppl. 4A):12-20.
17. Kmetz, M. E., W. T. Slater, Jr., and J. Schwartz. 1987. In vitroantichlamydial activity of antimicrobials and quinolones using arapid microtiter screening test system. J. Microbiol. Methods6:247-251.
18. Lam, E., and N. H. Chua. 1987. Chloroplast DNA gyrase and in
1888 KRMCOVIC ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 02
Feb
ruar
y 20
22 b
y 22
1.12
7.18
.159
.

QUINOLONES AND COUMARINS ELIMINATE CHLOROPLASTS 1889
vitro regulation of transcription by template topology and no-
vobiocin. Plant Mol. Biol. 8:415-424.19. Lyman, H. 1967. Specific inhibition of chloroplast replication in
Euglena gracilis by nalidixic acid. J. Cell Biol. 35:726-730.20. Lyman, H., A. S. Jupp, and I. Larrinua. 1975. Action of
nalidixic acid on chloroplast replication in Euglena gracilis.Plant Physiol. 55:390-392.
21. Margulis, L. 1981. Symbiosis in cell evolution. W. H. Freeman,San Francisco.
22. Neu, H. C. 1987. Ciprofloxacin: an overview and prospectiveappraisal. Am. J. Med. 82(Suppl. 4A):395-404.
23. Pienkos, P., A. Walfield, and C. L. Hershberger. 1974. Effect ofnalidixic acid on Euglena gracilis: induced loss of chloroplastdeoxyribonucleic acid. Arch. Biochem. Biophys. 165:548-553.
24. Pol6nyi, J., L. Ebringer, J. Krajeovki, and K. Kapeller. 1987.Ultrastructure of green and AF-2 nitrofuran-bleached mutantsof Euglena gracilis. I. Volumes of chloroplasts, mitochondriaand paramylon granules. Biologia (Bratislava) 42:217-229.
25. Pyke, K. A., J. Marrison, and R. M. Leech. 1989. Evidence fora type II topoisomerase in wheat chloroplasts. FEBS Lett.242:305-308.
26. Ratcliffe, N. T., and J. T. Smith. 1984. Ciprofloxacin andofloxacin exhibit a rifampicin-resistant bactericidal mechanismnot detectable in other 4-quinolone antibacterial agents. J.Pharm. Pharmacol. 36:59P.
27. Ratcliffe, N. T., and J. T. Smith. 1985. Norfloxacin has a novelbactericidal mechanism unrelated to that of other 4-quinolones.J. Pharm. Pharmacol. 37:92P.
28. Smith, J. T. 1984. Awakening the slumbering potential of the4-quinolone antibacterials. Pharm. J. 233:299-305.
29. Thompson, R. J., and G. Mosig. 1985. An ATP-dependentsupercoiling topoisomerase of Chlamydomonas reinhardtii af-fects accumulation of specific chloroplast transcripts. NucleicAcids Res. 13:873-891.
30. Wolfson, J. S., and D. C. Hooper. 1985. The fluoroquinolones:structures, mechanisms of action and resistance, and spectra ofactivity in vitro. Antimicrob. Agents Chemother. 28:581-586.
31. Zeiler, H.-J. 1985. Evaluation of the in vitro bactericidal actionof ciprofloxacin on cells of Escherichia coli in the logarithmicand stationary phases of growth. Antimicrob. Agents Chemo-ther. 28:524-527.
VOL. 33, 1989
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 02
Feb
ruar
y 20
22 b
y 22
1.12
7.18
.159
.





























