Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1991, p. 1070-10740099-2240/91/041070-05$02.00/0Copyright © 1991, American Society for Microbiology
Rapid Method for Direct Extraction of DNA fromSoil and Sediments
YU-LI TSAI AND BETTY H. OLSON*
Environmental Design and Analysis, Program in Social Ecology, University of California, Irvine, California 92717
Received 10 October 1990/Accepted 23 January 1991
A rapid method for the direct extraction of DNA from soil and sediments was developed. The indigenousmicroorganisms in the soil and sediments were lysed by using lysozyme and a freeze-thaw procedure. The lysatewas extracted with sodium dodecyl sulfate and phenol-chloroform. In addition to a high recovery efficiency(>90%), the yields ofDNA were high (38 and 12 ,ug/g [wet weight] from sediments and soil, respectively). Thismethod generated minimal shearing of the extracted DNA. The crude DNA could be further purified with anElutip-d column if necessary. An additional advantage of this method is that only 1 g of sample is required,which allows for the analysis of small samples and the processing of many samples in a relatively short (7 h)period.
Isolation of bacterial DNA from natural environments hasbecome a useful tool with which to study the ecologicalfunctions of certain characterized genes that encode impor-tant metabolic pathways (25), allow tracking of geneticallyengineered organisms (8, 10), and reveal bacterial DNAdiversity (21, 22) in microbial ecosystems. The application ofDNA extraction methods to environmental samples canobviate the need for cell cultivation, since cell cultivationhas the disadvantage of obtaining only a very small propor-tion of the total microbial community. DNA extractionmethods can also help researchers to understand the occur-rence of particular bacterial genes in situ through applica-tions of nucleic acid technology.
Several techniques have been described for direct detec-tion and extraction ofDNA from aquatic environments (3, 6,13-15, 19), but there are only a few reports related toextraction of DNA from soil and sediments (8, 12, 23). Twocurrent methods used to obtain microbial DNA from soils orsediments are (i) bacterial cell extraction followed by celllysis and DNA recovery (8, 23) and (ii) direct extraction byalkaline lysis (12). The direct extraction method produces abetter DNA yield than the cell extraction method (20).However, both approaches share three disadvantages, i.e.,they (i) are lengthy and laborious, (ii) require a relativelylarge sample size (50 to 100 g), and (iii) produce low yields ofDNA.
In this study we have developed a rapid method for directextraction of DNA from soil and sediments by applying afreeze-thaw approach. This method overcomes the draw-backs of the previous techniques. The high yield from asmall sample size (1 g [wet weight]) means that the methoddescribed in this report can be used efficiently to study in situgene occurrence and to detect the distribution of geneticallyengineered microorganisms in the environment.
MATERIALS AND METHODS
Soil characterization. Subsurface soil samples from a man-ufactured gas site in Southern California (SC) and sedimentsamples from a settling pond in Oak Ridge, Tenn. (ORT),were used for DNA extraction. The SC soils were contam-
* Corresponding author.
inated with polynuclear aromatic hydrocarbons (6.5 lug/g[wet weight]) and mercury (0.07 ,ug/g), and the ORT sedi-ments were contaminated with mercury (up to 80 ,ug/g) andother heavy metals (Global Geochemistry Corp., CanogaPark, Calif.; Oak Ridge National Laboratory, Oak Ridge,Tenn.). The heterotrophic bacterial population in both sam-ples was enumerated on plate count agar (Difco, Detroit,Mich.). The indigenous mercury-resistant strains were re-covered on plate count agar amended with 25 ,ug of Hg2+ asHgCI2 per ml. Naphthalene degraders were selectively cul-tivated on a minimal medium saturated with naphthalenevapor (18). Sterile soil and sediment samples were producedby autoclaving at 121°C for 30 min. All soil and sedimentquantities are expressed as wet weight unless otherwiseindicated. The percentage of water was determined to be51% in the ORT sediments and 10% in the SC soil (2).
Organisms. Two strains (V55 and VNM43) isolated fromSC soil were used as seed organisms. They were identified asPseudomonas luteola and Pseudomonas putida, respec-tively, by the API Rapid NFT? kits (Analytab Products,Plainview, N.Y.). In the remainder of this paper thesestrains are referred to as P. luteola V55 and P. putidaVNM43. Both strains have the ability to degrade naphtha-lene and are resistant to mercuric ions. They were grown tolate exponential phase in 50% plate count broth (Difco) with25 pLg of Hg2+ per ml and were washed once with 10 mMphosphate buffer (pH 7.0) and resuspended in phosphatebuffer before inoculation into the soils or sediments. Theinoculated soils and sediments were shaken at 100 rpm(Orbit Shaker; Lab-Line Instrument, Inc., Melrose Park,Ill.) at 23°C for 30 min prior to DNA extraction.
Direct extraction of DNA. SC soil or ORT sediment sam-ples (1 g) were mixed with 2 ml of 120 mM sodium phosphatebuffer (pH 8.0) by shaking at 150 rpm for 15 min. The slurrywas pelleted by centrifugation at 6,000 x g for 10 min. Thepellet was washed again with phosphate buffer, resuspendedin 2 ml of lysis solution (0.15 M NaCl, 0.1 M Na2EDTA [pH8.0]) containing 15 mg of lysozyme/ml, and incubated in a37°C water bath for 2 h with agitation at 20- to 30-minintervals, and then 2 ml of 0.1 M NaCl-0.5 M Tris-HCl (pH8.0)-10% sodium dodecyl sulfate was added. Three cycles offreezing in a -70°C dry ice-ethanol bath and thawing in a65°C water bath were conducted to release DNA from the
1070
Vol. 57, No. 4
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 27
Dec
embe
r 20
21 b
y 93
.89.
201.
113.

DIRECT DNA EXTRACTION FROM SOIL AND SEDIMENTS
microbial cells in the soil or sediments. The efficiency oflysis was determined by direct cell counts with phase-contrast microscopy and by acridine orange direct cellcounts (7) with fluorescence microscopy (model BH-2;Olympus, Tokyo, Japan) and was found to be greater than95% for P. putida VNM 43 and P. luteola V55 as well as a
Bacillus sp. After the freeze-thaw cycles, 2 ml of 0.1 MTris-HCl (pH 8.0)-saturated phenol (17) was added and thesample was briefly vortexed to obtain an emulsion. Themixture was centrifuged at 6,000 x g for 10 min. A 3-mlsample of the top aqueous layer was collected and thenmixed with 1.5 ml of phenol and 1.5 ml of chloroformmixture (chloroform/isoamyl alcohol ratio, 24:1). A 2.5-mlportion of the resulting extract was further extracted with an
equal volume of chloroform mixture. Finally, nucleic acidsin the extracted aqueous phase (2 ml) were precipitated with2 ml of cold isopropanol at -20°C for 1 h or overnight. Thepellet of crude nucleic acids was obtained by centrifugationat 10,000 x g for 10 min and then vacuum dried at 23°C. Thenucleic acid pellets, which were brownish as a result of thehumic materials in both ORT and SC samples, were resus-
pended in 100 ,ul ofTE buffer (20 mM Tris-HCl, 1 mM EDTA[pH 8.0]). The RNA molecules in the crude extracts were
removed by incubation with heat-treated pancreatic RNaseA (final concentration, 0.2 jig/[ld) for 2 h at 37°C. TheRNA-free DNA was then purified with an Elutip-d column(Schleicher & Schuell, Keene, N.H.) attached to a Schlei-cher & Schuell NA010/27 (0.45-,um [pore size] celluloseacetate) prefilter. DNA was recovered from the column as
suggested by the manufacturer. The efficiency of recovery oftotal DNA from sediment and/or soil crude extracts was
determined as the percentage of DNA extracted from a
known density of P. putida VNM43 in seeded sterile samplescompared with DNA recovered from the same density ofpure culture controls. Cell numbers used in all seedingexperiments resembled heterotrophic plate counts of thesediments and soils under investigation. The yield of DNAfrom the direct extraction was expressed in micrograms ofcrude DNA obtained per gram of unseeded nonsterile sam-
ples.Gel electrophoresis. Samples of extracted DNA were ana-
lyzed on a 0.7% agarose gel containing 1 p.g of ethidiumbromide per ml. To determine the quality of extracted DNA,we analyzed undigested DNA extract and appropriate con-
trols including DNase-digested (0.5 pig/p.l) and RNase-di-gested (0.2 p.g/p.l) extracts. The concentration of DNA in thecrude extract was determined spectrophotometrically at 260nm. Restriction digestion of DNA was conducted withEcoRI (2 U/p.l), BamHI (1 U/pul), and HindIll (1 U/pul) withincubation for 5 h at 37°C, followed by gel electrophoresis.
Hybridization. A 1-kb fragment of the merA gene encodingmercuric reductase was labeled with [ot-32P]dCTP by randompriming (specific activity, 108 dpm/pg of DNA) and used as
a probe to detect the presence of mercury detoxificationgenes in the environmental samples. The merA fragment ofplasmid pDU1358 was kindly provided by S. Silver. DNAwas transferred from the agarose gel to membranes byvacuum blotting (11). The extracted DNA was also filteredonto membranes through a slot blot apparatus (Minifold II;Schleicher & Schuell). Nylon membranes (GeneScreen Plus;NEN Products, Boston, Mass.) and nitrocellulose mem-
branes (BA85; pore size, 0.45 pum; Schleicher & Schuell)were used as solid supports. Hybridization was carried outat 42°C in the presence of 50% formamide, followed byhigh-stringency washes as previously described (24). TheRadioanalytic Imaging System (AMBIS Systems, San Di-
A B
FIG. 1. Total DNA extracted from ORT sediments. (A) Ethid-ium bromide-stained agarose gel; (B) autoradiogram of DNA hybrid-ization signals with merA after Southern blot of the gel in panel A.Aliquots (3 ,ul) of total extracts (100 ,ul) were loaded into each well.Lanes: 1, HindlIl-cut lambda bacteriophage molecular size marker(1 p.g); 2, unseeded; 3, unseeded and digested with RNase; 4,unseeded and digested with DNase; 5, seeded; 6, seeded anddigested with RNase; 7, seeded and digested with DNase; 8, plasmidsize marker from E. coli V517.
ego, Calif.) was used to quantify the radioactivity signals ofhybridized DNA on the membranes.
RESULTS AND DISCUSSION
Figure 1A shows an ethidium bromide-stained agarose gelused to visualize the DNA extracted from ORT sedimentsafter DNase and RNase treatments. The addition of 2.5 x108 cells of P. luteola V55 to 1 g of nonsterile sedimentsresulted in a larger quantity of DNA than in the unseededsample. The efficiency of recovery was calculated to be 92%when sterile ORT sediment was seeded with a known celldensity of P. luteola V55. The sterile samples (autoclaved orgamma irradiated) did not contain indigenous DNA afterextraction. The total DNA yield from nonsterile, unseededORT sediments was determined to be 38 ,ug/g (Table 1). Thelargest DNA was greater than 23 kb, and most DNA was inthe size range of 6.5 to 23.1 kb. The DNA-shearing effect ofthe extraction procedure was less prominent than in themethod described by Ogram et al. (12), as evidenced bysmearing of smaller fragments. Because the DNA extractsbefore RNase digestion contained RNA contaminants (Fig.1A), introduction of RNase during the extraction procedurewas necessary. Impurities such as humic materials in theextracts did not affect RNase or DNase digestion, but mayaffect DNA hybridization efficiency. RNA was not observedin unseeded samples, indicating that RNA degradation or
TABLE 1. Yields and efficiency of direct DNAextraction method
Sample Heterotrophic Recovery Yieldb (pLg/gSample bacteria (CFU/g) efficiency' (%) [wet wt])
ORT sediments (8.0 + 0.7) x 107 92 ± 3 38 ± 5SC soil (4.1 +0.1) x 106 95 4 12 4
aThe recovery efficiency was calculated from the total DNA obtained froma known cell density of sterile seeded samples compared with DNA extractionfrom bacterial pure cultures.
b The yield was measured from nonsterile unseeded samples.
VOL. 57, 1991 1071
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 27
Dec
embe
r 20
21 b
y 93
.89.
201.
113.
1072 TSAI AND OLSON
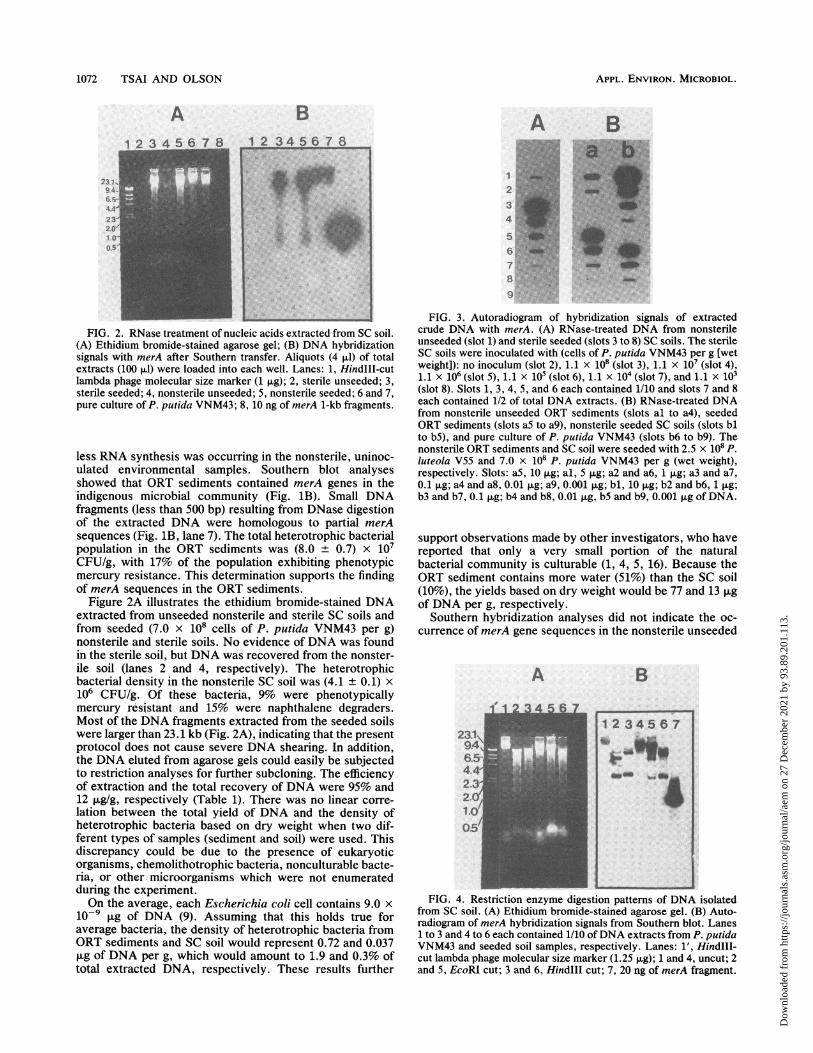
FIG. 2. RNase treatment of nucleic acids extracted from SC soil.(A) Ethidium bromide-stained agarose gel; (B) DNA hybridizationsignals with merA after Southern transfer. Aliquots (4 ,ul) of totalextracts (100 pLI) were loaded into each well. Lanes: 1, HindIII-cutlambda phage molecular size marker (1 ,ug); 2, sterile unseeded; 3,sterile seeded; 4, nonsterile unseeded; 5, nonsterile seeded; 6 and 7,pure culture of P. putida VNM43; 8, 10 ng of merA 1-kb fragments.
less RNA synthesis was occurring in the nonsterile, uninoc-ulated environmental samples. Southern blot analysesshowed that ORT sediments contained merA genes in theindigenous microbial community (Fig. 1B). Small DNAfragments (less than 500 bp) resulting from DNase digestionof the extracted DNA were homologous to partial merAsequences (Fig. 1B, lane 7). The total heterotrophic bacterialpopulation in the ORT sediments was (8.0 + 0.7) x 107CFU/g, with 17% of the population exhibiting phenotypicmercury resistance. This determination supports the findingof merA sequences in the ORT sediments.
Figure 2A illustrates the ethidium bromide-stained DNAextracted from unseeded nonsterile and sterile SC soils andfrom seeded (7.0 x 108 cells of P. putida VNM43 per g)nonsterile and sterile soils. No evidence of DNA was foundin the sterile soil, but DNA was recovered from the nonster-ile soil (lanes 2 and 4, respectively). The heterotrophicbacterial density in the nonsterile SC soil was (4.1 + 0.1) x106 CFU/g. Of these bacteria, 9% were phenotypicallymercury resistant and 15% were naphthalene degraders.Most of the DNA fragments extracted from the seeded soilswere larger than 23.1 kb (Fig. 2A), indicating that the presentprotocol does not cause severe DNA shearing. In addition,the DNA eluted from agarose gels could easily be subjectedto restriction analyses for further subcloning. The efficiencyof extraction and the total recovery of DNA were 95% and12 ,ug/g, respectively (Table 1). There was no linear corre-lation between the total yield of DNA and the density ofheterotrophic bacteria based on dry weight when two dif-ferent types of samples (sediment and soil) were used. Thisdiscrepancy could be due to the presence of eukaryoticorganisms, chemolithotrophic bacteria, nonculturable bacte-ria, or other microorganisms which were not enumeratedduring the experiment.On the average, each Escherichia coli cell contains 9.0 x
io-9 ,ug of DNA (9). Assuming that this holds true foraverage bacteria, the density of heterotrophic bacteria fromORT sediments and SC soil would represent 0.72 and 0.037,ug of DNA per g, which would amount to 1.9 and 0.3% oftotal extracted DNA, respectively. These results further
FIG. 3. Autoradiogram of hybridization signals of extractedcrude DNA with merA. (A) RNase-treated DNA from nonsterileunseeded (slot 1) and sterile seeded (slots 3 to 8) SC soils. The sterileSC soils were inoculated with (cells of P. putida VNM43 per g [wetweight]): no inoculum (slot 2), 1.1 x 108 (slot 3), 1.1 x 107 (slot 4),1.1 x 106 (slot 5), 1.1 x 105 (slot 6), 1.1 X 104 (slot 7), and 1.1 X 103(slot 8). Slots 1, 3, 4, 5, and 6 each contained 1/10 and slots 7 and 8each contained 1/2 of total DNA extracts. (B) RNase-treated DNAfrom nonsterile unseeded ORT sediments (slots al to a4), seededORT sediments (slots aS to a9), nonsterile seeded SC soils (slots blto b5), and pure culture of P. putida VNM43 (slots b6 to b9). Thenonsterile ORT sediments and SC soil were seeded with 2.5 x 108 p.luteola V55 and 7.0 x 108 P. putida VNM43 per g (wet weight),respectively. Slots: aS, 10 Rg; al, 5 jig; a2 and a6, 1 ,ug; a3 and a7,0.1 ,ug; a4 and a8, 0.01 ,ug; a9, 0.001 ,ug; bl, 10 ,ug; b2 and b6, 1 ,ug;b3 and b7, 0.1 ,ug; b4 and b8, 0.01 pug, b5 and b9, 0.001 jig of DNA.
support observations made by other investigators, who havereported that only a very small portion of the naturalbacterial community is culturable (1, 4, 5, 16). Because theORT sediment contains more water (51%) than the SC soil(10%), the yields based on dry weight would be 77 and 13 ,ugof DNA per g, respectively.
Southern hybridization analyses did not indicate the oc-currence of merA gene sequences in the nonsterile unseeded
A B
FIG. 4. Restriction enzyme digestion patterns of DNA isolatedfrom SC soil. (A) Ethidium bromide-stained agarose gel. (B) Auto-radiogram of merA hybridization signals from Southern blot. Lanes1 to 3 and 4 to 6 each contained 1/10 ofDNA extracts from P. putidaVNM43 and seeded soil samples, respectively. Lanes: 1' Hindlll-cut lambda phage molecular size marker (1.25 p.g); 1 and 4, uncut; 2and 5, EcoRI cut; 3 and 6, Hindlll cut; 7, 20 ng of merA fragment.
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 27
Dec
embe
r 20
21 b
y 93
.89.
201.
113.

DIRECT DNA EXTRACTION FROM SOIL AND SEDIMENTS
FIG. 5. Restriction enzyme digestion patterns of DNA extracted from ORT sediments. (A) Ethidium bromide-stained agarose gel. (B)Autoradiogram ofmerA hybridization signals from Southern blot of gel in panel A. Lanes 2 to 4 and 5 to 7 each contained 1/30 ofDNA extractsfrom unseeded and seeded samples, respectively. Lanes 8 to 10 and 11 to 13 contained 1/20 of Elutip-d column-purified DNA from unseededand seeded samples, respectively. Lanes: 1, HindIll-cut lambda phage molecular weight marker with ladder of 23.1, 9.4, 6.5, 4.4, 2.3, and2.0 kb; 2 and 8, unseeded and uncut; 3 and 9, unseeded and EcoRI cut; 4 and 10, unseeded and BamHI cut; 5 and 11, seeded and uncut; 6and 12, seeded and EcoRI cut; 7 and 13, seeded and BamHI cut.
SC soil (Fig. 2B, lane 4), but the direct application of DNAto membranes by the slot blot hybridization techniqueshowed that crude DNA extracted from the same soil didcontain merA genes (Fig. 3A, slot 1). The discrepancybetween these two observations was due to incompletetransfer of DNA from the gel to the filters during Southernblotting. Therefore, the existence of mercury-resistant bac-terial strains such as P. putida VNM43 and P. luteola V55 inthe polynuclear aromatic hydrocarbon-contaminated SCsoils was further supported by the gene probe data. TheDNA extracts from seeded sterile (Fig. 2B, lane 3) andnonsterile (lane 5) soils revealed smaller DNA fragments(between 1 and 20 kb) containing homologous merA se-quences. This shearing is probably due to the grinding effectof the sand particles in SC soil during the extraction proce-dure because pure-culture controls (lane 6 and 7) did notexhibit this effect. The two soils differ in cation exchangecapacity and particle size distribution, with SC soils havinglow cation exchange capacity (9 mEq/100 g) and high sandconcentration (18.4%), and ORT sediments having highcation exchange capacity and low sand concentration (Uni-
versity of California, Riverside, Department of Soil andEnvironmental Sciences; Oak Ridge National Laboratory).
Figure 3A shows the merA hybridization signals from thecrude DNA extracted from sterile SC soil seeded withdifferent quantities of P. putida VNM43. Signals were de-tected in a 50% aliquot of total DNA extract when as few as1.1 x 104 cells per g (wet weight) were inoculated into soil,indicating that the sensitivity was 5.5 x 103 cells per g. Acomparative sensitivity of 4.3 x 104 cells per g (dry weight)was reported for the cell extraction method (8). Figure 3Billustrates hybridization signals of putative merA genes ex-tracted from nonsterile unseeded and seeded ORT sedimentsand nonsterile seeded SC soils. As the extracted DNA wasnormalized at 1 ,ug, DNA from seeded samples (Fig. 3B,slots a6 and b2) demonstrated higher merA hybridizationsignals (7,049 and 24,477 cpm) than that from unseededsamples (800 cpm; slot a2). As would be expected, the signalfor the SC soils inoculated with a higher cell density (slot b2)had a higher-intensity signal of merA sequences than didORT sediments inoculated with a lower cell density (slot a6).The pure-culture DNA control showed the highest signal
VOL. 57, 1991 1073
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 27
Dec
embe
r 20
21 b
y 93
.89.
201.
113.

1074 TSAI AND OLSON
(28,648 cpm) in 1 ,ug of DNA extract. The radioactivitycounts for hybridized DNA were lower from soil and sedi-ment samples than from pure cultures, indicating that DNAhad absorbed into the soil particles or that more nonhomol-ogous merA sequences were present in the former samples.The detection limit of homologous merA DNA was 10 ng oftotal extracted DNA from both P. putida VNM43 pureculture and P. putida VNM43-seeded SC soils (Fig. 3B). ThemerA hybridization signals were found in 1 ,ug of total DNAextracted from nonsterile unseeded ORT sediments. Incomparison, Ogram et al. (12) required 1.2 ,ug of total DNAto detect nif nitrogen fixation genes from sediments otherthan those used in the present study. In the present studythere is no indication that humic materials in the crudeextract affect DNA-DNA hybridization (Fig. 3).
Figure 4 shows the restriction patterns of DNA extractsfrom a pure bacterial culture and from a seeded SC soil. Theunpurified DNA extract was clearly digested by Hindlll butonly slowly digested by EcoRI. The application of restrictionenzyme analyses to directly extracted DNA may allow moreprecise studies of in situ gene rearrangements or amplifica-tion. The impurities in ORT sediments did affect the restric-tion enzyme digestion (Fig. 5). EcoRI and BamHI did notdigest DNA extracts from either unseeded or seeded sam-ples within the 5-h incubation period. The DNA yieldsuffered a 40% loss during the Elutip-d column purificationstep, and the purified DNA (A260/A280 = 1.8) was cut byEcoRI but not by BamHI. Therefore, for studying in situgene transfer the purification step might be needed depend-ing on the soil type, but it is not necessary for determinationof gene occurrence.
In conclusion, this rapid method for the direct extractionof DNA needs only 1 g of soil or sediments and can detectthe presence of a target gene, such as merA, from a minimumof 5,000 bacterial cells. It is simple and can be completed in7 h. In addition, because of the small sample size (1 g), alarge number of soil or sediment samples can be analyzed atone time. Finally, the high yield in combination with the highquality of DNA will enable microbial ecologists to studyDNA from environmental samples in a more detailed fashionby molecular biology techniques.
ACKNOWLEDGMENTS
This study was funded by grant 8000-25 provided by the ElectricPower Research Institute.We are grateful to Marie Park for developing the autoradiograms.
We also thank 0. A. Ogunseitan, P. A. Rochelle, and C. C. Tebbefor valuable discussions and suggestions and R. Turner for provisionand analysis of ORT sediments.
REFERENCES1. Alexander, M. 1977. Introduction to soil microbiology, 2nd. ed.
John Wiley & Sons, Inc., New York.2. American Public Health Association. 1989. Standard methods for
the examination of water and wastewater, 17th ed. AmericanPublic Health Association, Washington, D.C.
3. DeFlaun, M. F., J. H. Paul, and D. Davis. 1986. Simplifiedmethod for dissolved DNA determination in aquatic environ-ments. Appl. Environ. Microbiol. 52:654-659.
4. Ferguson, R. L., E. N. Buckley, and A. V. Palumbo. 1984.Response of marine bacterioplankton to differential filtrationand confinement. Appl. Environ. Microbiol. 47:49-55.
5. Francisco, D. E., R. A. Mah, and A. C. Rabin. 1973. Acridine
orange epifluorescent technique for counting bacteria in naturalwaters. Trans. Am. Microsc. Soc. 92:416-421.
6. Fuhrman, J. A., D. E. Comeau, A. Hagstrom, and A. M. Chan.1988. Extraction from natural planktonic microorganisms ofDNA suitable for molecular biological studies. Appl. Environ.Microbiol. 54:1426-1429.
7. Hobbie, J. E., R. J. Daley, and S. Jasper. 1977. Use ofNuclepore filters for counting bacteria by fluorescence micros-copy microscopy. Appl. Environ. Microbiol. 33:1225-1228.
8. Holben, W. E., J. K. Jansson, B. K. Chelm, and J. M. Tiedje.1988. DNA probe method for the detection of specific microor-ganisms in the soil bacterial community. Appl. Environ. Micro-biol. 54:703-711.
9. Ingraham, J. L., 0. Maaloe, and F. C. Neidhardt. 1983. Growthof the bacterial cell, p. 1-48. Sinauer Associates, Inc., Sunder-land, Mass.
10. Jansson, J. K., W. E. Holben, and J. M. Tiedje. 1989. Detectionin soil of a deletion in an engineered DNA sequence by usingDNA probes. Appl. Environ. Microbiol. 55:3022-3025.
11. Medveczky, P., C.-W. Chang, C. Oste, and C. Mulder. 1987.Rapid vacuum driven transfer of DNA and RNA from gels tosolid supports. BioTechniques 5:242-246.
12. Ogram, A., G. S. Sayler, and T. Barkay. 1987. The extractionand purification of microbial DNA from sediments. J. Microb.Methods 7:57-66.
13. Paul, J. H., and D. J. Carlson. 1984. Genetic material in themarine environment: implications for bacterial DNA. Limnol.Oceanogr. 29:1091-1097.
14. Paul, J. H., W. H. Jeffrey, and M. DeFlaun. 1985. ParticulateDNA in subtropical oceanic and estuarine planktonic environ-ments. Mar. Biol. 90:95-101.
15. Paul, J. H., and B. Myers. 1982. Fluorometric determination ofDNA in aquatic microorganisms by use of Hoechst 33258. Appl.Environ. Microbiol. 43:1393-1399.
16. Roszak, D. B., and R. R. Colwell. 1987. Survival strategies ofbacteria in the natural environment. Microbiol. Rev. 51:365-379.
17. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
18. Sayler, G. S., M. S. Shields, E. T. Tedford, A. Breen, S. W.Hooper, K. M. Sirotkin, and J. W. Davis. 1985. Application ofDNA-DNA colony hybridization to the detection of catabolicgenotypes in environmental samples. Appl. Environ. Microbiol.49:1295-1303.
19. Somerville, C. C., I. T. Knight, W. L. Straube, and R. R.Colwell. 1989. Simple, rapid method for direct isolation ofnucleic acids from aquatic environments. Appl. Environ. Mi-crobiol. 55:548-554.
20. Steffan, R. J., J. Goks0yr, A. K. Bej, and R. M. Atlas. 1988.Recovery of DNA from soils and sediments. Appl. Environ.Microbiol. 54:2908-2915.
21. Torsvik, V., J. Goks0yr, and F. L. Daae. 1990. High diversity inDNA of soil bacteria. Appl. Environ. Microbiol. 56:782-787.
22. Torsvik, V., K. Salte, R. S0rheim, and J. Goks0yr. 1990.Comparison of phenotypic diversity and DNA heterogeneity ina population of soil bacteria. Appl. Environ. Microbiol. 56:776-781.
23. Torsvik, V. L. 1980. Isolation of bacterial DNA from soil. SoilBiol. Biochem. 12:15-21.
24. Tsai, Y.-L., and B. H. Olson. 1990. Effects of Hg2+, CH3-Hg',and temperature on the expression of mercury resistance genesin environmental bacteria. Appl. Environ. Microbiol. 56:3266-3272.
25. Walia, S., A. Khan, and N. Rosenthal. 1990. Construction andapplications of DNA probes for detection of polychlorinatedbiphenyl-degrading genotypes in toxic organic contaminatedsoil environments. Appl. Environ. Microbiol. 56:254-259.
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 27
Dec
embe
r 20
21 b
y 93
.89.
201.
113.





























